protein synthetic requirements for the outgrowth of trophoblast cells from mouse blastocysts
TRANSCRIPT

THE JOURNAL OF EXPERIMENTAL ZOOLOGY 224:401-408 (1982)
Protein Synthetic Requirements for the Outgrowth of Trophoblast Cells From Mouse Blastocysts
EMILY J. BLAKE, JOKL SCHINDLER, AND MICHAEL I. SHERMAN Roehe Institute of Molecular Biology, Nutley, New Jersey 07110 (J.S., M.I.S.) and Deprtment of Human. Genetics and Dmelopment, Colum.bia University College of Physicians and Surgeons, New York, New York 10052 (E,.J.B,, M . I.S.)
ABSTRACT Mouse blastocysts cultured under optimal conditions attach to the surface of the culture dish and subsequently give rise to outgrowths of trophoblast cells. The migration of trophoblast cells appears to be analogous to their behavior during the invasive phase of implantation in utero. In these studies, we have attempted to determine the time of synthesis and nature of the products required for trophoblast outgrowth. Fourth-day blastocysts were cul- tured in two different media: cNCTC, a nutrient medium supplemented with fetal calf serum, and PCMF, a simple preimplantation medium lacking amino acids and serum. Trophoblast outgrowth occurs in the former, but not in the latter, medium. Most blastocysts will give rise to outgrowths following as little as 12 hr of exposure to cNCTC at the appropriate time, even if they are subsequently placed in PCMF. Outgrowth fails to occur if blastocysts are treated with cycloheximide during the interval of exposure to cNCTC, whereas treat- ment with the antimetabolite after the period of culture in cNCTC does not block outgrowth. These observations are consistent with the view that protein(s) essential for trophoblast outgrowth are synthesized many hours prior to the actual event.
Successful implantation of the mammalian embryo requires that both the mother and the conceptus undergo a series of specific, ordered changes in a coordinated fashion. Disruption of the endocrinological balance in the uterus can prevent implantation and interrupt the developmental program of the blastocyst. In mice, implantation delay occurs naturally when a pregnant female suckles a previous litter or it can be induced by ovariectomy; in both instances, implantation can be achieved, and the developmental program of the concep- tus restored, when the hormonal insufficiency is rectified (Whitten, '55; Yoshinaga and Ad- ams, '66). During implantation delay, the de- velopmental program of the blastocyst appears to be disrupted by virtue of a generalized suppression of metabolic rates (Menke and McLaren, '70; Weitlauf, '74) and biosynthetic activities (McLaren, '68; Weitlauf and Green- wald, '68; Sherman and Barlow, '72; Weitlauf, '73; Chavez and Van Blerkom, '79; Van Bler- kom et al., '79). Although some early experi- ments suggested that hormone administration reverses implantation delay through a direct effect upon the blastocyst (Psychoyos, '67;
Prasad et al., '68; Smith, '68; Smith and Smith, '71), it now appears more likely that hormones act indirectly by altering the uterine milieu, thereby permitting restoration of normal met- abolic processes in the conceptus (see Sher- man and Wudl, '76; Van Blerkom et al., '79).
During implantation in the mouse, tropho- blast cells attach to, and subsequently invade,. the uterine wall. Under appropriate culture conditions, trophoblast cells will behave in an apparently analogous way (see Sherman and Wudl, '76): They initially become adherent to the substratum and then migrate out along it. These events can be blocked by placing blas- tocysts in suboptimal culture medium, e.g., in the absence of serum factors (Gwatkin, '66; Rizzino and Sherman, '79; Sellens and Sher- man, '80), amino acids (Gwatkin, '66; Spindle and Pedersen, '73; Naeslund, '79; Sellens and Sherman, '80; Sherman and Matthaei, '80), or
Joel Schindler is now at the Department of Anatomy. University
Address reprint requests t o Michael I. Sherman, Roche Institute of Cincinnati Medical Center, Cincinnati. OH 45267.
of Molecular Biology, Nutley, NJ 07110.
0022-104W82/2243-0401$02 50 'c, 1982 ALAN R. LISS. INC

402 E.J. BLAKE, J. SCHINDLER, AND M. 1. SHERMAN
glucose (Wordinger and Brinster, '76; Naes- lund, '79; Van Blerkom et al., '79). Sellens and Sherman ('80) provided evidence that blasto- cysts cultured in these suboptimal media re- semble those blocked from implanting in utero in several respects. Preliminary studies sug- gested that trophoblast outgrowth was blocked in suboptimal culture medium be- cause blastocysts could not enter a critical biosynthetic phase (Sherman and Matthaei, '80). The purpose of the present study was to define this biosynthetic period further and to learn the nature of the biosynthetic products that are critical for trophoblast outgrowth.
MATERIALS AND METHODS Materials
NCTC-109 medium and fetal calf serum (heat-inactivated at 56°C for 20 min) were pur- chased from Microbiological Associates (Be- thesda, MD). Antibiotics were purchased from Gibco (Grand Island, NY). Bovine serum al- bumin (BSA; PentexlMiles, Elkhart, IN) and fetuin (Calbiochem, La Jolla, CA) were solu- bilized and dialyzed extensively as described by Sellens and Sherman ('80). Cycloheximide (Calbiochem) was dissolved in distilled water at 1 mglml and filter sterilized. Fetuin and cycloheximide stock solutions were stored at -2O"C, thawed once and maintained at 4°C during use for a maximum of 1 (cycloheximide) or 2 (fetuin) weeks. Plastic tissue culture dishes (35 mm) were purchased from Falcon Plastics (Oxnard, CA). [3"S]Methionine ( - 1000 Cilmmol) and unlabled methionine were pur- chased from Amersham (Arlington Heights, IL) and Calbiochem, respectively. Eppendorf centrifuge tubes were obtained from Fisher. Scientific (Pittsburgh, PA). Hydrofluor (Na- tional Diagnostics, Somerville, NJ), NCS (Amersham), and LSC (Yorktown Research, Hackensack, NJ) were used for scintillation spectrometry, as indicated.
Culture media cNCTC is NCTC-109 medium supplemented
with penicillin (100 ulml), streptomycin (100 pglml), kanamycin (100 ulml), and 10% heat- inactivated fetal calf serum. PCM is preim- plantation culture medium formulated as de- scribed by Goldstein et al. ('75) except that the BSA was dialyzed prior to use (Sellens and Sherman, '80). PCM supplemented with 0.05% dialyzed fetuin is referred to as PCMF.
Embryos SWRlJ female mice (Jackson Laboratory,
Bar Harbor, ME) were superovulated (Run-
ner and Palm, '53) and mated with SJLlJ males (Jackson Laboratory). The day of observation of the sperm plug was considered the first day of pregnancy. Blastocysts were collected on the afternoon of the fourth day of pregnancy. All embryos were obtained by flushing uter- ine horns with phosphate-buffered saline [so- lution A of Dulbecco and Vogt ('54)]. Embryos were washed in PCM and then transferred for culture into either PCMF or cNCTC.
Cultur-e conditions Embryos were cultured in groups of 20 to
35 in 3 ml of PCMF or cNCTC medium, as indicated, in 35-mm tissue culture-grade plas- tic dishes and maintained at 37OC in a humid- ified atmosphere of 5% CO, in air. Embryos were scored as positive for outgrowth when at least one trophoblast cell and its nucleus could be seen flattened out on the culture dish surface.
Radioactive labehzg of embryos Labeling of embryos with [35S]methionine
was carried out in 1.5-ml Eppendorf centri- fuge tubes in 30-50 p1 of radioactive medium. The [35S]methionine concentration ranged from 0.25 to 0.35 pCilpl in PCMF and from 1.0 to 1.5 pCi/pl in cNCTC for incubations of 16- 24 hr. Different concentration ranges were used in the two media because of differences in protein synthetic rates and because of the presence of unlabeled methionine in cNCTC but not PCMF. Under these conditions, blas- tocysts cultured in PCMF or cNCTC incorpo- rated approximately 24,000 and 13,000 c p d embryo, respectively, into trichloroacetic acid- insoluble material (see below). Following the labeling periods, the embryos were washed twice with 1 ml PCM lacking sodium pyru- vate, sodium lactate and BSA but supple- mented with 0.02 mglml cold L-methionine, and stored frozen at -70°C.
Determination of trichloroacetic acid-soluble and -insoluble radioactivity
To determine the total uptake of [Y3]methio- nine by embryos, 0.5 ml of a 1.0 mglml BSA solution was added to the embryo samples. Following three rounds of freezing and thaw- ing, ice-cold trichloroacetic acid (TCA) was added to yield a final concentration of 10%. The samples were placed on ice for 30 min and centrifuged, and then supernates were re- moved and saved. Pellets were resuspended in hot TCA, placed in a boiling water bath for 15 min, centrifuged, and the supernates pooled with those from the 10% cold TCA extraction.
CONTROL OF TROPHOBLAST OUTGROWTH 403
Hydrofluor was added to the pooled super- nates, and samples were counted to determine TCA-soluble counts. The precipitated material was digested for 1 hr at 37OC with NCS, transferred to scintillation vials, and counted with LSC to determine the extent of incorpo- ration into TCA-insoluble material.
RESULTS Conditions that allow blastocysts to form
trophoblast outgrowths Blastocysts cultured in cNCTC hatch from
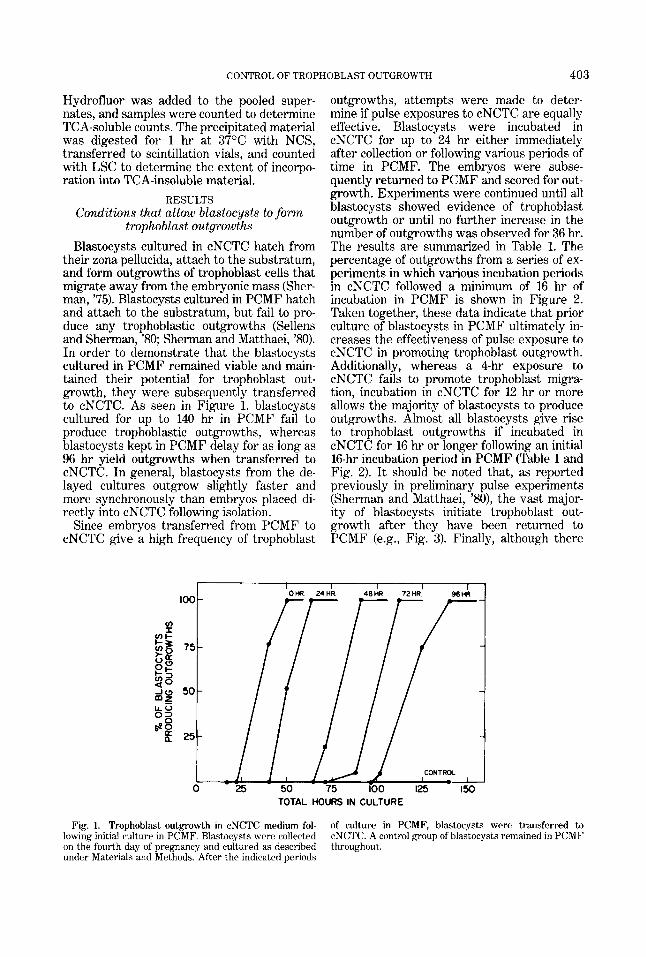
their zona pellucida, attach to the substratum, and form outgrowths of trophoblast cells that migrate away from the embryonic mass (Sher- man, '75). Blastocysts cultured in PCMF hatch and attach to the substratum, but fail to pro- duce any trophoblastic outgrowths (Sellens and Sherman, '80; Sherman and Matthaei, '80). In order to demonstrate that the blastocysts cultured in PCMF remained viable and main- tained their potential for trophoblast out- growth, they were subsequently transferred to cNCTC. As seen in Figure 1, blastocysts cultured for up to 140 hr in PCMF fail to produce trophoblastic outgrowths, whereas blastocysts kept in PCMF delay for as long as 96 hr yield outgrowths when transferred to cNCTC. In general, blastocysts from the de- layed cultures outgrow slightly faster and more synchronously than embryos placed di- rectly into cNCTC following isolation.
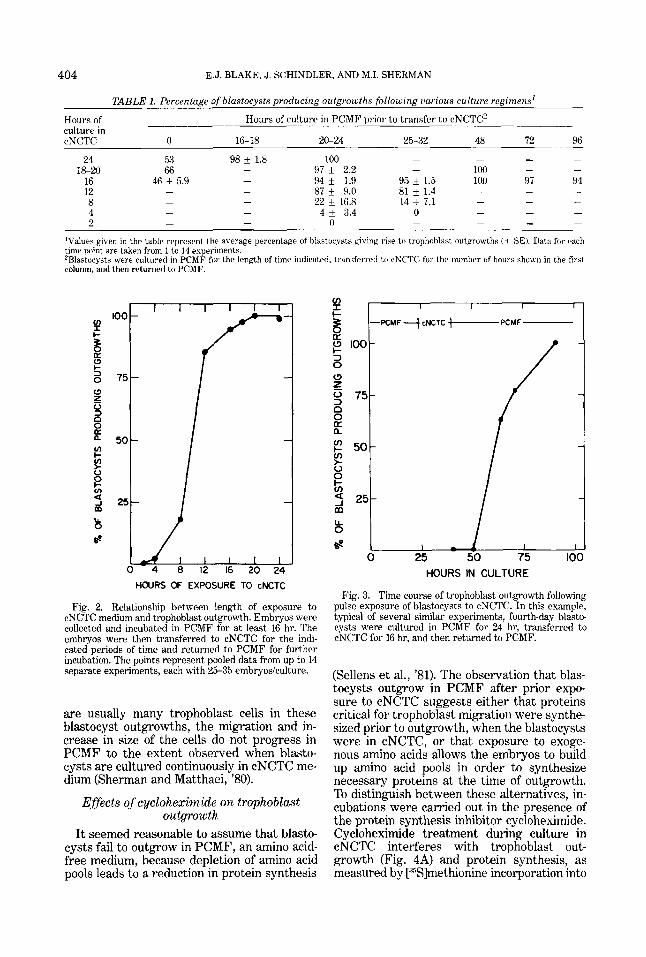
Since embryos transferred from PCMF to cNCTC give a high frequency of trophoblast
outgrowths, attempts were made to deter- mine if pulse exposures to cNCTC are equally effective. Blastocysts were incubated in cNCTC for up to 24 hr either immediately after collection or following various periods of time in PCMF. The embiyos were subse- quently returned to PCMF and scored for out- growth, Experiments were continued until all blastocysts showed evidence of trophoblast outgrowth or until no further increase in the number of outgrowths was observed for 36 hr. The results are summarized in Table 1. The percentage of outgrowths from a series of ex- periments in which various incubation periods in cNCTC followed a minimum of 16 hr of incubation in PCMF is shown in Figure 2. Taken together, these data indicate that prior culture of blastocysts in PCMF ultimately in- creases the effectiveness of pulse exposure to cNCTC in promoting trophoblast outgrowth. Additionally, whereas a 4-hr exposure to cNCTC fails to promote trophoblast migra- tion, incubation in cNCTC for 12 hr or more allows the majority of blastocysts to produce outgrowths. Almost all blastocysts give rise to trophoblast outgrowths if incubated in cNCTC for 16 hr or longer following an initial 16-hr incubation period in PCMF (Table 1 and Fig. 2). I t should be noted that, as reported previously in preliminary pulse experiments (Sherman and Matthaei, '80), the vast major- ity of blastocysts initiate trophoblast out- growth after they have been returned to PCMF (e.g., Fig. 3). Finally, although there
TOTAL HOURS IN CULTURE
Fig. 1. Trophoblast outgrowth in cNCTC medium fol- lowing initial culture in PCMF. Blastocysts were collected on the fourth day of pregnancy and cultured as described under Materials and Methods. After the indicated periods
of culture in PCMF, blastocysts were transferred to cNCTC. A control group of blastocysts remained in PCMF throughout.
404 E.J. BLAKE, J. SCHINDLER, AND M.I. SHERMAN
v) I
[L
3 0 (3
loo-
z 0 75- 3 0
2 n
5 0 - > V
E 3 25- m
6." b
TABLE 1. Percentage of blastocysts producing outgrowths following iiarious culture regimens'
Hours of culture in cNCTC 0 16-18 20-24 25-32 48 72 96
24 53 98 f 1.8 100 - 97 * 2.2 - 100 - -
16 46 f 5.9 - 94 f 1.9 95 f 1.5 100 97 94 12 - - 87 i 9.0 81 k 1.4
- - 22 i 16.8 14 +_ 7.1 - - - 8 4 - - 4 * 3.4 0 2
Hours of culture in PCMF prior to transfer to cNCTC'
- - - -
18-20 66
- - -
- - - - - - - - - 0
'Values given in the table represent the average percentage of blastocysts giving rise to trophohlast outgrowths ( 5 SE). Data for each time point are taken from 1 to 14 experiments. 'Blastocysts were cultured i n PCMF for the length of time indicated. transferred tr, cNCTC for the nirmber of hnurs shown in the first column, and then returned to PCMF.
I I I I
--PtMFfcNCTC I-f'CMF
I 0 25 50 75 to
1 1 1 1 1 1 100
L 24
HOURS OF EXPOSURE TO cNCTC
Fig. 2. Relationship between length of exposure to cNCTC medium and trophoblast outgrowth. Embryos were collected and incubated in PCMF for a t least 16 hr. The embryos were then transferred to cNCTC for the indi- cated periods of time and returned to PCMF for further incubation. The points represent pooled data from up to 14 separate experiments, each with 25-35 embryosiculture.
are usually many trophoblast cells in these blastocyst outgrowths, the migration and in- crease in size of the cells do not progress in PCMF to the extent observed when blasto- cysts are cultured continuously in cNCTC me- dium (Sherman and Matthaei, '80).
Effects of cycloheximide on trophoblast outgrowth
It seemed reasonable to assume that blasto- cysts fail to outgrow in PCMF, an amino acid- free medium, because depletion of amino acid pools leads to a reduction in protein synthesis
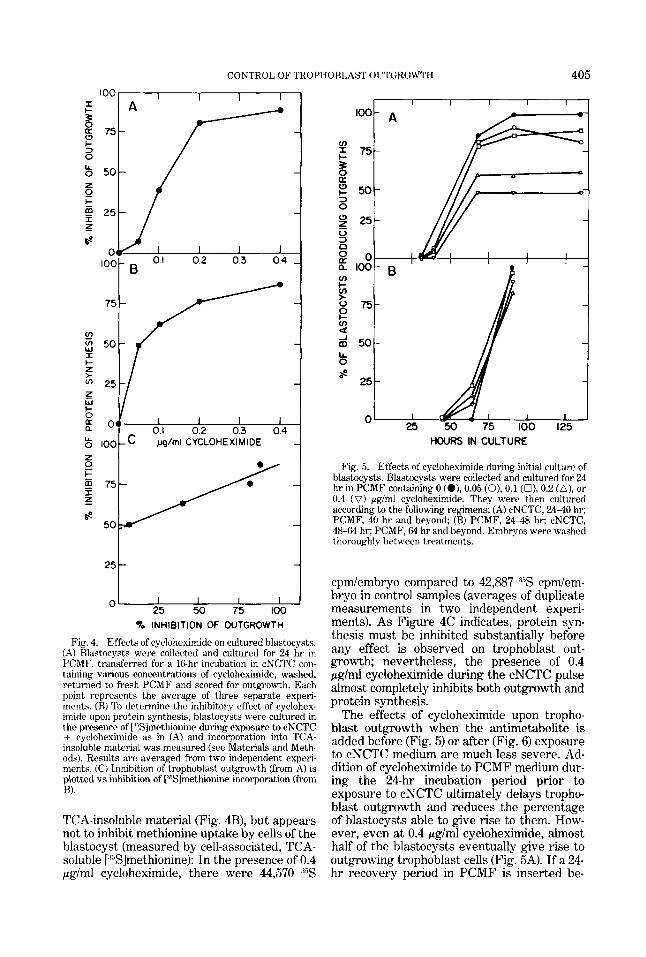
(Sellens et al., '81). The observation that blas- tocysts outgrow in PCMF after prior expo- sure to cNCTC suggests either that proteins critical for trophoblast migration were synthe- sized prior to outgrowth, when the blastocysts were in cNCTC, or that exposure to exoge- nous amino acids allows the embryos to build up amino acid pools in order to synthesize necessary proteins at the time of outgrowth. To distinguish between these alternatives, in- cubations were carried out in the presence of the protein synthesis inhibitor cycloheximide. Cycloheximide treatment during culture in cNCTC interferes with trophoblast out- growth (Fig. 4A) and protein synthesis, as measured by [:%fmethionine incorporation into
CONTROL OF TROPHOBLAST OUTGROWTH 405
100-
v)
I- = 75-
a F! 50- 3 0 * 25- z 0 3 0
HIS-, v) c v) > 0 75- 0
U d 50-
z
!i
B 25
100 I I I I
A
-
i5 50 75 100 125 0 I 0.2 0.3 0.4
25 t O' 215 ;o 715 Id0
% INHIBITION OF OUTGROWTH
Fig. 4. Effects of cycloheximide on cultured blastocysts. (A) Rlastocysts were collected and cultured for 24 hr in PCMF, transferred for a 16-hr incubation in cNCTC con- taining various concentrations of cycloheximide, washed, returned to fresh PCMF and scored for outgrowth. Each point represents the average of three separate experi- ments. (B) To determine the inhibitory effect of cyclohex- imide upon protein synthesis, blastocysts were cultured in the presence of [ "Slmethionine during exposure to cNCTC
cycloheximide as in (A) and incorporation into TCA- insoluble material was measured (see Materials and Meth- ods). Results are averaged from two independent experi- ments. (C) Inhibition of trophoblast outgrowth (from A) is plotted vs inhibition of [''Slrnethionine incorporation (from B).
TCA-insoluble material (Fig. 4B), but appears not to inhibit methionine uptake by cells of the blastocyst (measured by cell-associated, TCA- soluble [:%]methionine): In the presence of 0.4 pg/ml cycloheximide, there were 44,570 ,35S
I I I I I I
HOURS IN CULTURE
Fig. 5. Effects of cycloheximide during initial culture of blastocysts. Blastocysts were collected and cultured for 24 hr in PCMF containing 0 (O), 0.05 (0). 0.1 (O), 0.2 (A), or 0.4 ( V ) pg/ml cycloheximide. They were then cultured according to the following regimens: (A) cNCTC, 24-40 hr; PCMF, 40 hr and beyond; (B) PCMF, 24-48 hr; cNCTC, 48-64 hr; PCMF, 64 hr and beyond. Embryos were washed thoroughly between treatments.
cpmlembryo compared to 42,887 d5S cpdem- bryo in control samples (averages of duplicate measurements in two independent experi- ments). As Figure 4C indicates, protein syn- thesis must be inhibited substantially before any effect is observed on trophoblast out- growth; nevertheless, the presence of 0.4 p g l d cycloheximide during the cNCTC pulse almost completely inhibits both outgrowth and protein synthesis.
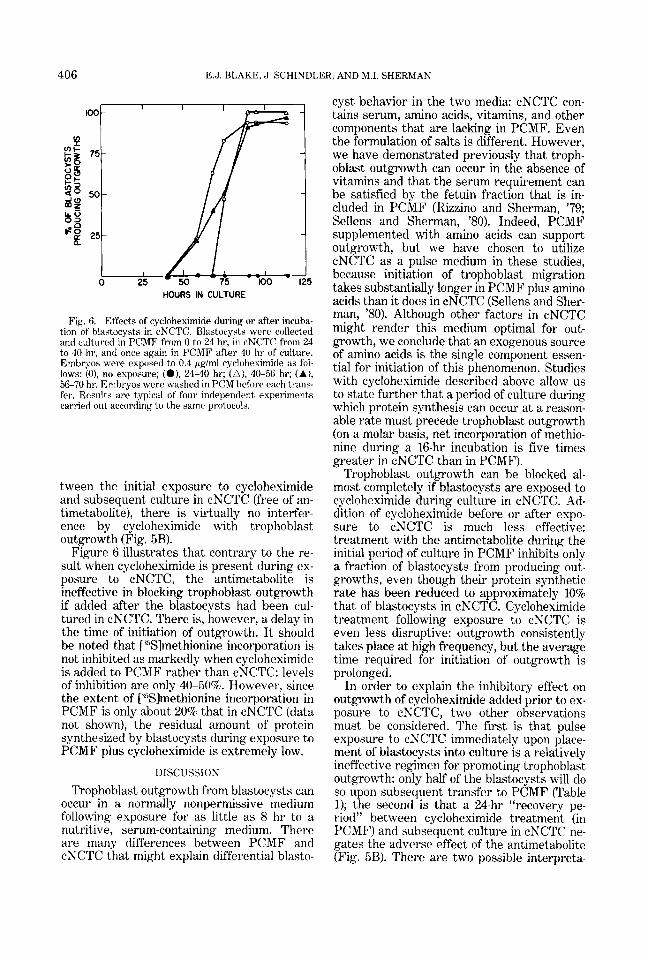
The effects of cycloheximide upon tropho- blast outgrowth when the antimetabolite is added before (Fig. 5) or after (Fig. 6) exposure to cNCTC medium are much less severe. Ad- dition of cycloheximide to PCMF medium dur- ing the 24-hr incubation period prior to exposure to cNCTC ultimately delays tropho- blast outgrowth and reduces the percentage of blastocysts able to give rise to them. How- ever, even at 0.4 pglml cycloheximide, almost half of the blastocysts eventually give rise to outgrowing trophoblast cells (Fig. 5A). If a 24- hr recovery period in PCMF is inserted be-
406 E.J. BLAKE, J. SCHINDL ,ER, AND M.I. SHERMAN
cyst behavior in the two media: cNCTC con- tains serum, amino acids, vitamins, and other components that are lacking in PCMF. Even the formulation of salts is different. However, we have demonstrated previously that troph- oblast outgrowth can occur in the absence of vitamins and that the serum requirement can be satisfied by the fetuin fraction that is in- cluded in PCMF (Rizzino and Sherman, ’79; Sellens and Sherman, ’80). Indeed, PCMF supplemented with amino acids can support outgrowth, but we have chosen to utilize cNCTC as a pulse medium in these studies, because initiation of trophoblast migration takes substantially longer in PCMF plus amino acids than it does in cNCTC (Sellens and Sher- man, ’80). Although other factors in cNCTC might render this medium optimal for out- growth, we conclude that an exogenous source of amino acids is the single component essen- tial for initiation of this phenomenon. Studies with cycloheximide described above allow us to state further that a period of culture during which protein synthesis can occur at a reason- able rate must precede trophoblast outgrowth (on a molar basis, net incorporation of methio- nine during a 16-hr incubation is five times greater in cNCTC than in PCMF).
Trophoblast outgrowth can be blocked al- most completely if blastocysts are exposed to cycloheximide during culture in cNCTC. Ad- dition of cycloheximide before or after expo- sure to cNCTC is much less effective: treatment with the antimetabolite during the initial period of culture in PCMF inhibits only a fraction of blastocysts from producing out- growths, even though their protein synthetic rate has been reduced to approximately 10% that of blastocysts in cNCTC. Cycloheximide treatment following exposure to cNCTC is even less disruptive: outgrowth consistently takes place at high frequency, but the average time required for initiation of outgrowth is prolonged.
In order to explain the inhibitory effect on outgrowth of cycloheximide added prior to ex- posure to cNCTC, two other observations must be considered. The first is that pulse exposure to cNCTC immediately upon place- ment of blastocysts into culture is a relatively ineffective regimen for promoting trophoblast outgrowth: only half of the blastocysts will do so upon subsequent transfer to PCMF (Table 1); the second is that a 24-hr “recovery pe- riod” between cycloheximide treatment (in PCMF) and subsequent culture in cNCTC ne- gates the adverse effect of the antimetabolite (Fig. 5B). There are two possible interpreta-
5 HOURS IN CULTURE
Fig. 6. Effects of cycloheximide during or after incuba- tion of blastocysts in cNCTC. Bbastocysts were collected and cultured in PCMF from 0 to 24 hr, in cNCTC from 24 to 40 hr, and Once again in PCMF after 40 hr of culture. Embryos were exposed to 0.4 pgiml cycloheximide as fol- lows: (O), no elposure; (0). 24-40 hr; (A) , 40-56 hr; (A), 56-70 hr. Emhryos were washed in PCM before each trans- fer. Results are typical of four independent experiments carried out according to the same protocols.
tween the initial exposure to cycloheximide and subsequent culture in cNCTC (free of an- timetabolite), there is virtually no interfer- ence by cycloheximide with trophoblast outgrowth (Fig. 5B).
Figure 6 illustrates that contrary to the re- sult when cycloheximide is present during ex- posure to cNCTC, the antimetabolite is ineffective in blocking trophoblast outgrowth if added after the blastocysts had been cul- tured in cNCTC. There is, however, a delay in the time of initiation of outgrowth. I t should be noted that [ “Slmethionine incorporation is not inhibited as markedly when cycloheximide is added to PCMF rather than cNCTC: levels of inhibition are only 40-50%. However, since the extent of [ Y3lmethionine incorporation in PCMF is only about 20% that in cNCTC (data not shown), the residual amount of protein synthesized by blastocysts during exposure to PCMF plus cycloheximide is extremely low.
DISCUSSION
Trophoblast outgrowth from blastocysts can occur in a normally nonpermissive medium following exposure for as little as 8 hr to a nutritive, serum-containing medium. There are many differences between PCMF and cNCTC that might explain differential blasto-

CONTROL OF TROPHOBLAST OUTGROWTH 407
tions of these results: the first, which we be- lieve most reasonable, is that many fourth-day blastocysts at the time of collection are not capable of producing adequate amounts of out- growth-related proteins, even in cNCTC, and that prerequisite developmental processes re- quiring protein synthesis must occur. A sec- ond explanation, which we cannot yet eliminate, is that fourth-day blastocysts are normally capable of producing outgrowth-re- lated proteins in vivo, but that the trauma of being placed in culture precludes their doing so; cycloheximide treatment during the initial culture period exacerbates the effect, whereas recovery incubations in PCMF prior to expo- sure to cNCTC overcome it.
The observation that cycloheximide has a minimal effect upon trophoblast outgrowth when added after exposure to cNCTC sug- gests that the critical time for synthesis of outgrowth-related proteins is several hours prior to the event per se (for example, Fig. 3 illustrates that about 30 hr elapsed between the midpoint of the pulse exposure to cNCTC and the time at which half the blastocysts initiated outgrowth). It would appear that there is also a substantial hiatus between the time at which outgrowth-related proteins are translated and the time at which their mRNA had been transcribed, since exposure to a-amanitin at any time after collection of blas- tocysts on the fourth day fails to influence either the frequency or time of initiation of outgrowth (Schindler and Sherman, '81; Sher- man e t al., '81). Although the critical period for production of outgrowth-related proteins appears to be during the cNCTC pulse, pro- tein synthesis (even if a t reduced rates) follow- ing transfer to PCMF appears to be beneficial for outgrowth, since cycloheximide treatment during this period delays initiation of out- growth. Furthermore, the extent of tropho- blast outgrowth in PCMF following exposure to cNCTC is rarely equal to that observed when blastocysts are maintained in cNCTC. Whether this means that outgrowth-related proteins must be continually replenished or that other types of outgrowth-promoting pro- teins are made after return of blastocysts to PCMF remains to be determined.
Since levels of protein synthesis in PCMF are much lower than those in cNCTC, failure to outgrow in the former medium could be due to either qualitative or quantitative differ- ences between the proteins synthesized by blastocysts in the two media. In preliminary studies we have attempted to distinguish be- tween these two possibilities by incubating
cells with [."S]methionine in cNCTC or PCMF during the synthetic period critical for out- growth and analyzing the products by two- dimensional gel electrophoresis (O'Farrell, '75). Analyses of these gels did not reveal ob- vious qualitative differences (unpublished ob- servations). It is possible, therefore, that the synthesis of outgrowth-related proteins can take place in PCMF at the appropriate devel- opmental time, but that outgrowth is pre- cluded because insufficient quantities of these proteins are produced.
ACKNOWLEDGMENTS
We are grateful to Ms. Perla Pine for assist- ance. We also thank Drs. J. Monahan and C. Coleclough for comments on the manuscript.
LITERATURE CITED
Chavez, D.J., and J. Van Rlerkom (1979) Persistence of embryonic RNA synthesis during facultative delayed implantation in the mouse. Dev. Riol., 7039-49.
Dulbecco, R., and M. Vogt (19c54) Plaque formation and isolation of pure lines with poliomyelitis viruses. J. Exp. Med., 99:167-199.
Goldstein, C.S., A.I. Spindle and R.A. Pedersen (1975) X- ray sensitivity of the prcimplantation mouse embryo in vitro. Radiat. Res., 6&27f-287.
Gwatkin, R.R.L. (1966) Defined media and development of mammalian eggs in vitro. Ann. N.Y. Acad. Sci.. 159.79- 90.
McLaren, A. (1968) A study of blastocysts during delay and siibsequent implantation in lactating mice. J. Entlo- crinol., 42453-464.
Menke, T.M., and A. Mc1,aren (1970) Carbon dioxide pro- duction by mouse blastocysts during lactational delay of implantation or after ovariectomy. J. Endocrinol., 4 7237- 294.
Naeslund, G. (1979) The effect of glucose-, arginine- and leucine-deprivation on mouse blastocyst outgrowth in vitro. IJps. J . Med. Sci., 84.9-20.
O'Famell, P.H. (1975) High resolution, two-dimensional electrophoresis of proteins. J. Bid. Chem., 2t50:4007-4021.
Prasad, M.R.N., C.M.S. Ilass and S. Mohla (1968) Action of oestrogen on the blastocyst and uterus in delayed implantation. An autoradiographic study (rat). J . Reprod. Fertil . , f6.?R- 104.
Psychoyos, A. (1967) Mechanismes tle la nidation. Arch. Anat. Microsc. Morphol. Exp., 56:616-631.
Rizzino, A., and M.I. Sherman (1979) Development and differentiation of mouse blastocysts i n serum-free me- dium. Exp. Cell Res., 121t221-233.
Runner, M.N., and J. Palm (1953) Transplantation antl sur- vival of unfertilized ova of the mouse in relation to post ovulatory age. J . Exp. Zool., IX303-316.
Schindler, J., and M.I. Sherman (1981) Effects of a-amani- tin on programming of mouse blastocyst development. Dev. Biol., 84:332-340.
Sellens, M.H., and M.I. Sherman (1980) Effects of culture conditions on the developmental programme of mouse hlastocysts. J. Embryol. Exp. Morphnl., 56:l-22.
Sellens, M.H., S. Stein and M.I. Sherman (1981) Protein and free amino acid content in preimplantation mouse embryos antl in hlastocysts under various culture condi- tions. J. Reprod. Fertil., 61.307-315.
Sherman, M.I. (1975) Long term culture of cells derived from mouse blastocysts. Differentiation 3:61-67.

408 E.J. BLAKE, J. SCHINDLER, AND M.I. SHERMAN
Sherman, M.I., and P.W. Barlow (1972) Deoxyribonucleic acid content in delayed mouse hlastocysts. J. Reprod. Fertil., 29:123-126.
Sherman, M.I., and K.I. Matthaei (1980) Factors involved in implantation-related events in vitro. Prog. Reprod. Biol. 79:43-53.
Sherman, M.I., and L.R. Wudl(1976) The implanting mouse bkastocyst. In: The Cell Surface in Animal Embryogene- sis and Development. G. Poste and G.L. Nicolson, eds. North-Holland, Amsterdam, pp. 81-125.
Sherman, M.I., M.H. Sellens, S.B. Atienza-Samols, A.C. Pai and J. Schindler (1981) Relationship between the programs for implantation and trophoblast differentia- tion. In: Cellular and Molecular Aspects of Implantation. S.R. Glasser and D.W. Bullock, eds. Plenum Press, New York, pp. 75-89.
Smith, D.M. (1968) The effect on implantation of treating cultured mouse blastocysts with oestrogen in vitro and the uptake of 'H-oestradiol by blastocysts. J. Endocri- nol., /1:17-29.
Smith, D.M., and A.E. Smith (1971) Uptake and incorpo- ration of amino acids by cultured mouse embryos: Estro- gen stimulation. Biol. Reprod., 4.66-73.
Spindle, A.I., and R.A. Pedersen (1973) Hatching, attach-
ing and outgrowth of mouse bldstocysts in vitro: Fixed nitrogen requirements. J. Exp. Zool., 286:305-318.
Van Blerkom, J., D.J. Chavez, and H. Bell (1979) Molecular and cellular aspects of facultative delayed implantation in the mouse. In: The Maternal Regulation of Pregnancy, CIBA Symposium, Vol. 64. B. Heap, ed. North-Holland, Amsterdam, pp. 141-163.
Weitlauf, H.M. (1973) In vitro uptake and incorporation of amino acids by bkastocysts from intact and ovariectom- ized mice. J. Exp. Zool., 183,303-308.
Weitlauf, H.M. (1974) Effect of actinomycin D on protein synthesis by delayed implanting mouse embryos in vitro.
Weitlauf, H.M., and G.S. Greenwald (1968) Survival of blastocysts in the uteri of ovariectomized mice. J. Re- prod. Fertil., 17:515-520.
Whitten, W.K. (1955) Endocrine studies on delayed implan- tation in lactating mice. J. Endocrinol., 13:l-6.
Wordinger, R.J., and R.L. Brinster (1976) Influence of re- duced glucose levels on the in vitro hatching, attachment and trophoblast outgrowth of the mouse blastocyst. Dev. Biol., 53,294-296.
Yoshinaga, K., and C.E. Adams (1966) Delayed implanta- tion in the spayed, progesterone-treated adult mouse. J. Reprod. Fertil., 22t593-595.
J. EXP. ZOO^., I89t197-202.