possible impacts of zooplankton grazing on dimethylsulfide production in the antarctic ocean
TRANSCRIPT

Possible impacts of zooplankton grazing ondimethylsulfide production in the Antarctic Ocean1
Nobue Kasamatsu, So Kawaguchi, Shuichi Watanabe, Tsuneo Odate,and Mitsuo Fukuchi
Abstract: The role of zooplankton grazing on dimethylsulfide (DMS) and dissolved dimethylsulfoniopropionate(DMSPd) production was investigated in the Antarctic Ocean in January and February 2002. Dominant herbivorousmacrozooplankton of this region, the Antarctic krill Euphausia superba and the tunicate Salpa thompsoni, were used inshipboard incubation experiments. The concentration of DMSPd + DMS increased in the water during incubation withkrill. The production rate was 2.96 ± 2.78 nmol DMSPd + DMS·krill–1·h–1 (mean ± SD). In addition, the DMSPd +DMS production rate was linearly related to the ingestion rate of krill (r2 = 0.664, p ≤ 0.01). Addition of salps tonatural surface water, however, did not change the DMSPd + DMS concentrations. During the experiments, bothanimals fed on phytoplankton cells. The fecal pellets of krill contained broken phytoplankton cells, whereas those ofsalps contained unbroken cells. These results suggest that sloppy feeding by krill is a more likely mechanism for pro-ducing DMS and DMSPd than the direct ingestion of phytoplankton cells by salps. The decrease of DMS concentra-tions in the upper 200 m of the water column from January to February may be explained, in part, by changes in thecomposition of the macrozooplankton community.
Résumé : En janvier et février 2002, nous avons étudié le rôle du broutage du zooplancton sur la production de sul-fure de diméthyle et de diméthylsulfoniopropionate dissous (DMSPd) dans l’océan Antarctique. Nous avons aussi utiliséle krill antarctique Euphausia superba et le tunicier Salpa thompsoni, les espèces dominantes du macrozooplancton dela région, dans des expériences d’incubation à bord. Les concentrations de DMSPd + DMS augmentent dans l’eau du-rant l’incubation avec le krill. Le taux de production est de 2,96 ± 2,78 nmol DMSPd + DMS·krill–1·h–1 (moyenne ±écart type). De plus, le taux de production de DMSPd + DMS est en corrélation linéaire avec le taux d’ingestion dukrill (r2 = 0,664, p ≤ 0,01). En revanche, l’addition de salpes à de l’eau naturelle de surface ne change pas la concen-tration de DMSPd + DMS. Durant l’expérience, les deux espèces d’animaux se nourrissent de cellules du phytoplanc-ton. Les boulettes fécales du krill contiennent des cellules de phytoplancton brisées, alors que celles des salpescontiennent des cellules intactes. Ces résultats indiquent que l’alimentation brouillonne du krill est un mécanisme plusprobable de production de DMS et de DMSPd que l’ingestion directe des cellules du phytoplancton par les salpes. Ledéclin des concentrations de DMS dans les 200 m supérieurs de la colonne d’eau en janvier et en février peuts’expliquer, en partie, par les changements dans la communauté du macrozooplancton.
[Traduit par la Rédaction] Kasamatsu et al. 743
Introduction
Dimethylsulfide (DMS) is the most abundant form of vol-atile sulfur in the ocean. According to the CLAW hypothesis(Charlson et al. 1987), marine DMS production may act as aclimate regulatory mechanism via a feedback loop involvingsurface ocean temperature, phytoplankton, DMS, sulfate aero-sol formation, and cloud albedo, although the existence ofthis feedback has not been proven. Recently, attention has
been focused on assessing sinks and sources of DMS in theoceans and on the processes controlling the cycling of DMSin the marine environment. The different processes involvedin marine DMS emission were reviewed by Liss et al. (1997).DMS is produced by the enzymatic cleavage of dimethyl-sulfoniopropionate (DMSP), an organic compound synthe-sized by many phytoplankton species that may serve as anosmolyte and cryoprotectant (Karsten et al. 1996; Kirst1996). Since DMS is produced in seawater by the break-
Can. J. Fish. Aquat. Sci. 61: 736–743 (2004) doi: 10.1139/F04-072 © 2004 NRC Canada
736
Received 24 March 2003. Accepted 2 March 2004. Published on the NRC Research Press Web site at http://cjfas.nrc.ca on23 June 2004.J17407
N. Kasamatsu.2 The Graduate University for Advanced Studies, 173-8515, 9-10 Kaga 1-chome, Itabashi-ku, Tokyo, Japan.S. Kawaguchi.3 National Research Institute of Far Seas Fisheries, 424-8633, 5-7-1 Orido, Shimizu, Japan.S. Watanabe. Japan Marine Science and Technology Center, 237-0061, 2-15 Natsushimacho, Yokosuka, Japan.T. Odate and M. Fukuchi. The Graduate University for Advanced Studies, 173-8515, 9-10 Kaga 1-chome, Itabashi-ku, Tokyo,Japan, and National Institute of Polar Research, 173-8515, 9-10 Kaga 1-chome, Itabashi-ku, Tokyo, Japan.
1This paper is part of the proceedings of the Third International Symposium on Biological and Environmental Chemistry ofDMS(P) and Related Compounds, held in Rimouski (Québec), 26–28 September 2002.
2Corresponding author (e-mail: [email protected]).3Present address: Australian Antarctic Division, Channel Highway, Kingston Tasmania 7050, Australia.

down of its precursor, DMSP, one of the key steps towardsbetter knowledge of DMS cycling in the ocean is to under-stand the rates of and controls on production and fate ofDMSP. The conversion of DMSP into DMS may be influ-enced by bacterial activities (Kiene and Linn 2000), phyto-planktonic enzymes (Niki et al. 2000), and zooplanktongrazing (Dacey and Wakeham 1986; Cantin et al. 1996;Wolfe et al. 2000). However, there is still a need for aproper evaluation of the role of zooplankton on DMS pro-duction in natural systems.
Dacey and Wakeham (1986) found that one third of thephytoplankton DMSP ingested by copepods was released inthe seawater as DMS. Daly and DiTullio (1996) showed thatDMS concentrations increased when krill were added to bot-tles filled with seawater during shipboard experiments. Theinterpretation of these results is, however, limited, as zoo-plankton ingestion rates were not measured during these ex-periments.
Zooplankton grazing experiments conducted during DMSstudies were mainly done with copepods and krill, which areboth known as sloppy feeders (Dacey and Wakeham 1986;Daly and DiTullio 1996; Tang et al. 2000a). Macro-zooplankton have various grazing mechanisms in nature. Tounderstand the impact of zooplankton on DMS production,we need to assess the effect of the different grazing mecha-nisms on DMS production. The Antarctic krill Euphausiasuperba and the tunicate Salpa thompsoni are both known asdominant macrozooplankton groups in the Antarctic Ocean.Although both krill and salps are filter feeders (e.g.,Harbison et al. 1986), they have different grazing mecha-nisms. Krill are known as sloppy feeders, because they breakalgal cells with their mouthparts (Strom et al. 1997), whereassalps filter particles on their mucous nets and swallow themdirectly into their guts (Alldredge and Madin 1982). Break-age of the algal cells through sloppy feeding may result ingreater dissolved DMSP (DMSPd) production than doesgrazing by direct ingestion of cells without breakage(Kawaguchi 2001). Because of their different grazing mech-anisms and their importance as consumers of primary pro-ducers in the Antarctic Ocean, we believe that it is importantto compare the contribution of krill and salps to DMS pro-duction in this ocean system. The objective of this study wasto determine the influence of macrozooplankton grazing onDMS dynamics in the Antarctic Ocean during the summer-time.
Materials and methods
Water samplingSampling was carried out in the Australian sector of the
Antarctic Ocean (approximately 65°S, 140°E) during cruiseKH01-3 of the R/V Hakuho-Maru (University of Tokyo) inJanuary 2002 and during the 43rd Japanese Antarctic Re-search Expedition Marine Science Cruise of the R/VTangaroa in February 2002 (Table 1). Water samples werecollected at 10 or 11 depths from 2 to 200 m with a rosettesampler equipped with 12-L Niskin bottles and a conductivity–temperature–depth (CTD) probe (SeaBird SBE911 plus). Inaddition, surface seawater was taken with a plastic bucket.On some occasions, seawater pumped by the ship from adepth of approximately 5 m was used for culture experi-
ments (Table 2). These samples were used for the determi-nation of DMS, DMSPd, particulate DMSP (DMSPp), andchlorophyll a (Chl a). For DMS(P) analyses, water sampleswere transferred with 100-mL glass syringes.
Incubation experimentsTo determine the influence of macrozooplankton grazing
on DMSP and DMS production, shipboard incubation exper-iments were conducted during the February cruise using thetwo dominant macrozooplankton species, the Antarctic krillE. superba and the tunicate S. thompsoni (Table 2). Krill andsalp individuals were sampled by vertical tows with aNORPAC net at a hauling speed of 0.5 m·s–1. Salps werehandled as gently as possible to avoid any damage to the an-imals. The animals were acclimated for several days underlaboratory conditions, and only those in good condition wereused for the experiments. This time span also ensured the re-generation of their mucous nets even if they were acciden-tally damaged during the sampling. Except for experiment 3,krill were collected before the February cruise and kept alivein the laboratory until performing the experiments. Approxi-mate body length of krill and salps was 35 and 40 mm, re-spectively. All animals were kept in Whatman GF/F (nominalpore size of 0.7 µm) filtered seawater for 24 h before the be-ginning of the experiments to ensure that the gut contents ofthe animals were cleared out.
For the incubation experiments, water was collected at thesea surface or at the depth of the Chl a maximum, as de-scribed above, or pumped by the ship from a depth of ap-proximately 5 m (Table 2). The water was transferred into a75-L plastic bucket through 330-µm mesh, which removedmacro- and meso-zooplankton. Initial concentrations ofDMS, DMSPd, DMSPp, Chl a, and bacteria of the seawaterin this 75-L bucket were determined in triplicate. The sea-water was then immediately subdivided into six narrow-mouthed 10-L polycarbonate bottles as gently as possible.One animal each was placed in three of these bottles. Theremaining three bottles without animals were incubated ascontrols. All of the labware was previously cleaned with di-luted HCl.
All bottles were incubated, with a little headspace, for 6–24 h in a darkened roller incubator installed in a low-temperature room (approximately 0.5 °C). At the end of theincubation, subsamples were collected in each bottle for thedetermination of DMS(P), Chl a, and bacterial abundance.The appearance of fecal pellets in the incubation bottles waschecked every 3–6 h. Fecal pellets were removed from thetreatment bottles when they were present. They were ob-served and photographed under a microscope. Clearance rateof Chl a by macrozooplankton was calculated using theequation suggested by Frost (1972).
Sulfur determinationsDMS and DMSP concentrations were measured with a
modified purge and trap apparatus as described by Watanabeet al. (1987) and Uzuka et al. (1996). For DMS measure-ment, an aliquot of 4–14 mL of seawater was filtered on aWhatman GF/F filter (47 mm) and the filtrate was intro-duced into a glass purge chamber. DMS concentration wasdetermined on a gas chromatograph equipped with a flamephotometric detector (Shimadzu GC-14B). For total DMSP
© 2004 NRC Canada
Kasamatsu et al. 737
(DMSPt) (DMSPt = DMSPd + DMSPp) + DMS measure-ment, an aliquot of 4–14 mL of seawater sample was pouredinto a 30-mL serum bottle containing 4 mL of 6 molNaOH·L–1. The alkali treatment permits the cleavage ofDMSPt into gaseous DMS. For DMSPd + DMS measure-ment, an aliquot of 4–14 mL of seawater was filteredthrough a Whatman GF/F filter and the filtrate was pouredinto a serum bottle containing NaOH. Serum bottles werestored at 4 °C for at least 24 h to complete the cleavage be-fore analyses. For DMSP analyses, the serum bottles werepurged with pure nitrogen gas. DMSPd concentration wascomputed as the difference between DMSPd + DMS andDMS. DMSPp concentration was calculated as the differencebetween DMSPt + DMS and DMSPd + DMS. The analyticalerror for each measurement was 11%.
Pigment analysisSamples for pigment analysis (Chl a and pheopigments)
were filtered on Whatman GF/F filters. Pigments were ex-tracted in dimethylformamid (Suzuki and Ishimaru 1990) for24 h at approximately –80 °C. Concentrations of pigmentswere determined onboard the ship (Parsons et al. 1984) us-ing a model 10AU Turner Designs fluorometer.
Bacterial abundanceSamples for the determination of bacterial abundance
were fixed with 0.5% glutaraldehyde and kept refrigeratedfor later processing. At a shore laboratory, cells were stainedwith 4′,6-diamidino-2-phenylindole and counted by epi-fluorescence microscopy (Kirchman et al. 1982).
Statistical methodsStatistical analyses were conducted to evaluate the possi-
ble impacts of zooplankton on DMS(P) production, Chl aconcentration, and bacterial abundance in the experimentalseawater during the incubation. Because the values in thecontrol bottles changed during the incubation periods as wellas in the treatment bottles, differences of the final values be-tween treatment bottles and control bottles (mean of three
replicates for each experiment) were compiled for both krilland salps and applied to the tests. Because we were uncer-tain about the normality of the distribution of the values, weused both parametric (one-tailed t test) and nonparametrictests (the sign test) to assess the significance of their differ-ences. Nonparametric tests are less powerful than parametrictest but require fewer assumptions. Therefore, the results ofthe nonparametric tests will be more conservative (Zar 1996).To evaluate the relationship between the zooplankton graz-ing activity and DMS(P) production, a linear regression wasfitted. Statistical analyses were conducted using Statview(version 5.0) software (SAS Institute Inc., Cary, N.C.).
Results
Overview of the survey siteIn January 2002, the northern edge of the sea ice cover
was observed near 65.5°S. In February, sea ice cover had re-treated more than 93 km further south, near the coast of theAntarctic continent. The sampling station was thus free ofice in February. The surface mixed layer depth (defined asthe depth where potential density is 0.1 greater than surfacevalues) at the sampling station increased from 5 m in Janu-ary to 50 m in February. An extensive phytoplankton bloomcomposed mainly of diatoms (M. Miki, University of Tokyo,Tokyo 113-8657, Japan, personal communication, e-mail:[email protected]) was observed at the iceedge in January. Chl a concentrations integrated from 0 to200 m were 412 mg·m–2 in January and 87 mg·m–2 in Febru-ary (Table 1). The dominant macrozooplankton communitychanged from krill in January (J. Nishikawa, Ocean Re-search Institute, University of Tokyo, Tokyo 164-8639, Ja-pan, personal communication, e-mail: [email protected])to salps in February (S. Kawaguchi, personal observation).
Concentrations of DMS and related compoundsIn January, DMS and DMSPd concentrations integrated from
0 to 200 m were very high, reaching 1.45 and 2.18 mmol·m–2,respectively (Table 1). In February, these concentrations de-
© 2004 NRC Canada
738 Can. J. Fish. Aquat. Sci. Vol. 61, 2004
Sample location
DateLatitude(S)
Longitude(E)
DMS(mmol·m–2)
DMSPd
(mmol·m–2)DMSPp
(mmol·m–2)Chl a(mg·m–2)
Dominantzooplankton
10 Jan. 2002 65°41′ 140°08′ 1.45 2.18 1.93 411.78 Krill18 Feb. 2002 65°26′ 139°51′ 0.54 0.75 3.36 86.99 Salp
Table 1. Dominant zooplankton and integrated values for DMS, DMSPd, DMSPp, and Chl a in the water column(depth = 0–200 m).
Details of seawater used
Experiment Latitude (S) DepthExperimentalanimal
Initial Chl a(µg·L–1)
Initial DMSPd+DMS(nmol·L–1)
1 66°26′ Chl a maximum (5 m) Krill 4.20 4.832 64°00′ Chl a maximum (16 m) Krill 0.83 6.203 65°26′ Surface Krill 2.25 6.854 65°26′ Surface Krill 1.38 3.815 64°00′ Pumped (5 m) Salp 0.14 4.916 64°45′ Pumped (5 m) Salp 0.34 10.54
Table 2. Details of shipboard incubation experiments.
creased to 0.54 and 0.75 mmol·m–2, respectively. In contrastwith DMS and DMSPd, DMSPp concentration increased from1.93 mmol·m–2 in January to 3.36 mmol·m–2 in February (Ta-ble 1).
Krill experimentsDuring most experiments, krill ingested phytoplankton
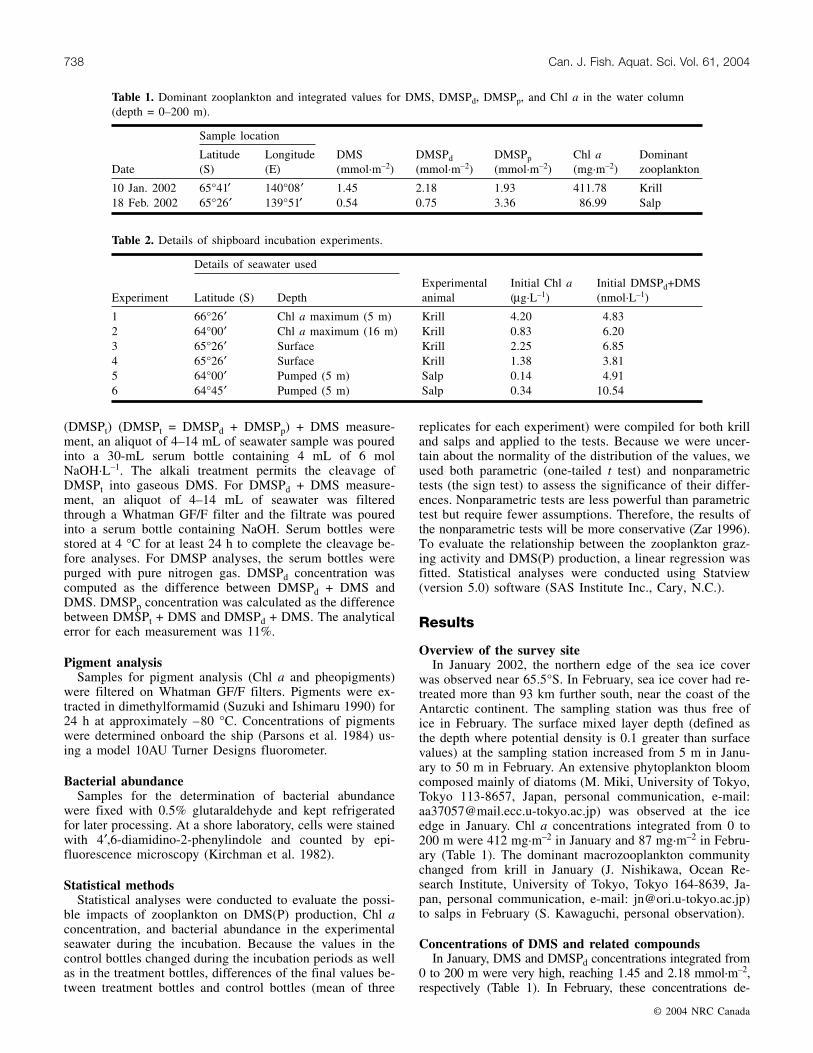
pigments. Chl a concentrations in treatment bottles were sig-nificantly lower than those of final control bottles (one-tailedt test, t = –2.713, p ≤ 0.05, n = 12; sign test, p ≤ 0.01, n =12) (Fig. 1a). For all experiments, the clearance rate was262 ± 239 mL·individual–1·h–1 (mean ± SD) and the inges-tion rate ranged from 0.00 to 0.48 µg Chl a·individual–1·h–1
(Table 3). DMSPd + DMS concentrations in treatment bot-tles were significantly higher than in the final control bottles(one-tailed t test, t = 2.937, p ≤ 0.05, n = 12; sign test,
p ≤ 0.01, n = 12) (Fig. 1b). Production rates of DMS andDMSPd + DMS were 2.91 ± 2.75 and 2.96 ± 2.78 nmol·individual–1·h–1, respectively (Table 4). Mean bacterialabundance under the different experimental conditionsranged from 1.1 to 1.4 × 105 cells·mL–1 (Table 5). There wasa statistical significant difference in bacterial abundances be-tween treatment bottles and final control (one-tailed t test,t = 3.559, p ≤ 0.01, n = 12; sign test, p ≤ 0.05, n = 12).
Salp experimentsInitial Chl a concentrations in salp grazing experiments
were lower than those in the krill experiments (Table 2). Sig-nificant statistical differences between Chl a concentrationsin treatment bottles and final control bottles were observed(one-tailed t test, t = –2.936, p ≤ 0.05, n = 6; sign test, p ≤0.05, n = 6). During the experiments with salps, clearancerates ranged from 5.3 to 1048 mL·individual–1·h–1, while in-gestion rates were 0.06 ± 0.06 µg Chl a·individual–1·h–1
(mean ± SD) (Table 3). However, changes in DMSPd + DMSconcentrations during the incubations were not statisticallysignificant (one-tailed t test, t = 1.071, p = 0.333, n = 6; signtest, p = 0.688, n = 6) (Fig. 1b). Mean bacterial abundanceunder the different experimental conditions ranged from 0.8to 1.0 × 105 cells·mL–1 (Table 5). There was no significantdifference in bacterial abundance between the treatment bot-tles and final control bottles (one-tailed t test, t = –1.777, p =0.136, n = 6; sign test, p = 0.219, n = 6).
© 2004 NRC Canada
Kasamatsu et al. 739
Fig. 1. Differences in (a) Chl a concentrations and (b) DMSPd +DMS concentrations between treatment bottles and final controlbottles (treatment minus final control) with the incubation time.The broken line indicates where the value of treatment bottlesequals that of final control bottles. Positive values indicate thatvalues of treatment bottles are higher than those of final controlbottles.
Animal Mean Max. Min. SD n
Clearance rate (mL·individual–1·h–1)Krill 262.26 717.58 0.00 238.64 12Salp 336.51 1047.96 5.34 400.42 6Ingestion rate (µg Chl a·individual–1·h–1)Krill 0.25 0.48 0.00 0.17 12Salp 0.06 0.17 0.00 0.06 6
Note: Max., maximum; Min., minimum; n, the number of individuals.
Table 3. Clearance and ingestion rates of zooplankton in ship-board incubation experiments.
Animal Mean Max. Min. SD n
DMS production rates (nmol�individual–1·h–1)Krill 2.91 7.26 –0.70 2.75 12Salp na na na na 6DMSPd + DMS production rates (nmol·individual–1·h–1)Krill 2.96 9.86 –0.60 2.78 12Salp na na na na 6
Note: Max., maximum; Min., minimum; n, the number of individuals;na, not applicable.
Table 4. DMS and DMSPd + DMS production rates in shipboardincubation experiments.
Bacteria count (× 105 cells·mL–1) (mean ±SD)
Animal Initial control Final control Treatment
Krill 1.1±0.7 1.2±0.7 1.4±0.7Salp 1.0±0.5 0.9±0.2 0.8±0.2
Table 5. Bacterial abundance in incubation experiments.
Relationship between ingestion rates and productionrates
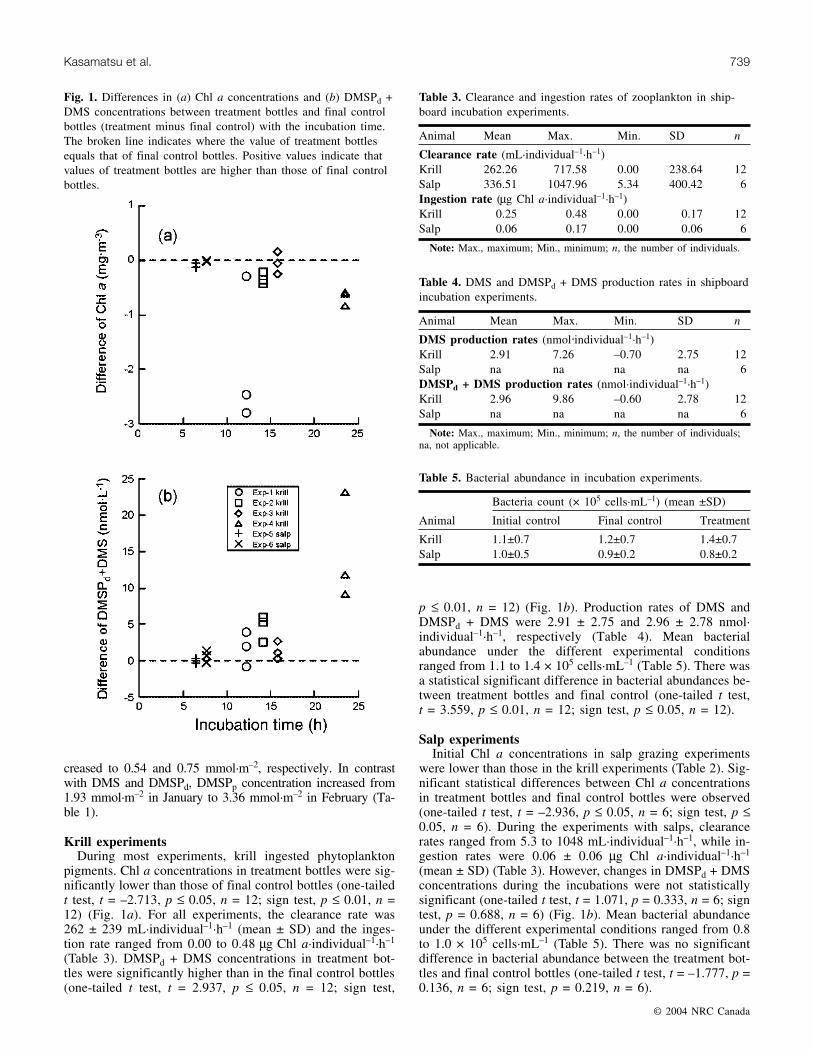
Ingestion rate of krill was linearly related to DMSPd +DMS production rate (linear regression, r2 = 0.664, p ≤ 0.01,n = 12) (Fig. 2). In one case, the DMSPd + DMS productionrate of krill was calculated as a negative value. This mayhave been caused by the poor physiological condition of thekrill.
Discussion
During this study, there were substantial changes in theposition of the ice edge, in the concentrations of biogenicsulfur compounds and Chl a, and in the species compositionof the macrozooplankton population. In January, the south-ernmost sampling station was located near the ice edge andthe macrozooplankton community was dominated by krill.The integrated DMS value measured that month in the upper200 m of the water column was relatively high (1.45 mmol·m–2) near the ice edge where krill were the dominant macro-zooplankton. DMS concentration at the sea surface was31 nmol·L–1, a value 15 times higher than those measured inmost oceans areas (Aumont et al. 2002). In February, whenthe sea ice had retreated further south, the integrated DMSvalue (0–200 m) decreased to one third of that in January.DMS concentrations within the mixed layer were stable atapproximately 3 nmol·L–1, a value closer to the mean oce-anic DMS concentration. The integrated DMSPd value inFebruary also decreased to one third of the January value. Incontrast with DMS and DMSPd, the integrated DMSPp valuein February showed about a twofold increase from that inJanuary. The krill was replaced by salps during this interval.Phytoplankton biomass, estimated by Chl a concentrations,dramatically decreased from 412 mg·m–2 in January to87 mg·m–2 in February, although the DMSPp integrated value
increased over the 2 months. This may be due to a change inthe composition of the algal assemblage and (or) growthphase of the dominant phytoplankton species between Janu-ary and February.
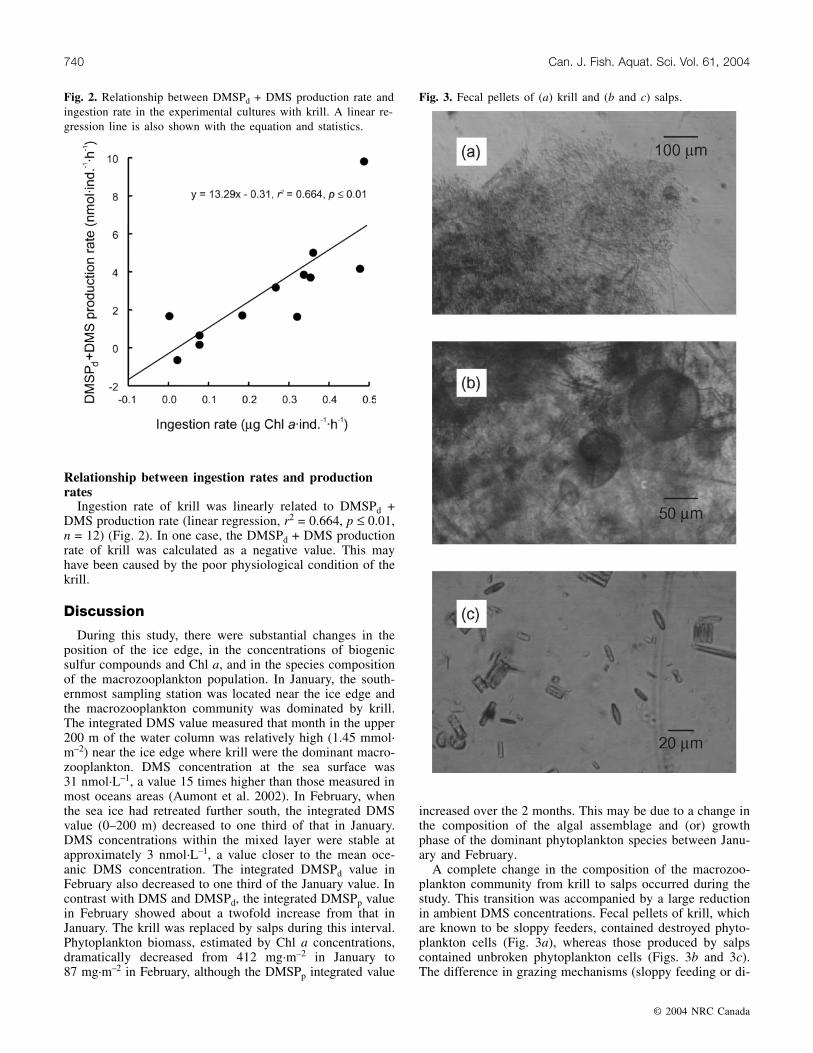
A complete change in the composition of the macrozoo-plankton community from krill to salps occurred during thestudy. This transition was accompanied by a large reductionin ambient DMS concentrations. Fecal pellets of krill, whichare known to be sloppy feeders, contained destroyed phyto-plankton cells (Fig. 3a), whereas those produced by salpscontained unbroken phytoplankton cells (Figs. 3b and 3c).The difference in grazing mechanisms (sloppy feeding or di-
© 2004 NRC Canada
740 Can. J. Fish. Aquat. Sci. Vol. 61, 2004
Fig. 2. Relationship between DMSPd + DMS production rate andingestion rate in the experimental cultures with krill. A linear re-gression line is also shown with the equation and statistics.
Fig. 3. Fecal pellets of (a) krill and (b and c) salps.

rect ingestion) may lead to differences in the magnitude ofDMS and DMSPd release from the ingested phytoplanktoncells. Therefore, among the many possible factors influenc-ing DMS concentrations, we focused on the contribution ofthe various grazing impacts.
The ingestion rate of krill was linearly related to theDMSPd + DMS production rate (r2 = 0.664, p ≤ 0.01, n =12). This indicates that krill grazing on phytoplankton is di-rectly linked to the increasing DMSPd + DMS concentration.The difference in bacterial abundance between treatmentbottles and final control bottles was significant. If the bacte-rial abundance has increased in the treatment bottles, it mayalso have affected the DMSPd + DMS production in krill ex-periments. However, the linear relationship found betweeningestion rates of krill and DMSPd + DMS production ratesupports the assumption that DMSPd + DMS production hasbeen stimulated by krill grazing during the experiments.
The rate of DMSPd + DMS production by krill grazing inour study was 2.96 nmol·krill–1·h–1 and the DMS productionrate was 2.91 nmol·krill–1·h–1. DMS production rate throughkrill grazing reported by Daly and DiTullio (1996) ac-counted for only 5% of the value obtained in our work. Dalyand DiTullio (1996) noted that their values may have beenunderestimated because of confinement effects. Furthermore,in their work, the initial Chl a concentration was low(0.31 mg·m–3) compared with that of our krill incubation ex-periments. The containers that we used were relatively large(10 L) and were rotated during the experiment so that thephytoplankton were always suspended in the incubation bot-tles. Moreover, most of the animals used during our incuba-tions were well adapted to the tanks and consequently lessstressed than wild animals recently captured. All of our ex-perimental conditions should have produced values with lessexperimental stress, which may be closer to the in situ value.
In the case of salps, clearance rates varied among individ-uals. Huntley et al. (1989) conducted shipboard incubationexperiments and obtained maximum clearance rates of376 mL·individual–1·h–1 for S. thompsoni in the AntarcticPeninsula region. Pakhomov et al. (2002) reported that theclearance rate of S. thompsoni is a function of body length.According to their regression, the clearance rate of salps of40 mm length should be 1491 mL·individual–1·h–1. In ourstudy, clearance rates of salps ranged from 5.3 to 1048 mL·individual–1·h–1 (337 mL·individual–1·h–1 on average). Ourclearance rates are slightly smaller than those of Pakhomov etal. (2002), but our maximum rates are only marginallyhigher than those of Huntley et al. (1989). Pakhomov et al.(2002) also reported that the changes in ingestion rates as afunction of salp length are best represented by a power func-tion. This equation gives an ingestion rate of 0.6 µg pigment·individual–1·h–1 for a 40-mm animal. In our experiment,the ingestion rates of salps ranged between 0.0 and 0.2 µgChl a · individual–1·h–1. These low values result mainly fromthe low Chl a concentrations present in our incubation ves-sels and not from the clearance performance. Hence, we be-lieve that our incubation experiments on salps areappropriate for grazing experiments.
Our results indicate that krill grazing on phytoplanktonwas an important mechanism for producing DMSPd andDMS. On the other hand, salps had no detectable effect on
the production of this biogenic gas. The difference in graz-ing mechanisms (sloppy feeding by krill or direct ingestionby salps) may have led to differences in the magnitude ofDMS release from the ingested phytoplankton. What was theimportance of zooplankton grazing on DMS and DMSPdconcentrations in the field? In January, near the study site(65°S), krill abundance obtained by RMT net (0–800 m) was64 individuals·1000 m–3 (S. Toczko, Ocean Research Insti-tute, University of Tokyo, Tokyo 164-8639, Japan, personalcommunication, e-mail: [email protected]). DMSPd +DMS production rates by the krill in January were calcu-lated to be 0.2 nmol·m–3·h–1 (based on a per individual rateof 2.96 nmol·individual–1·h–1). Turnover time expressed asambient DMSPd + DMS integrated value (0–200 m) dividedby the production rate by the krill assemblage in Januarywas estimated to be 4 days. These values are comparablewith the microbial turnover time of <1–3 days calculated inthe Labrador Sea (Wolfe et al. 2000). Furthermore, if we as-sume an interannual variation range of 4.8–511 individuals·1000 m–3 (Loeb et al. 1997) at our study site, this means thatDMSPd + DMS production rate by krill may vary inter-annually between 0.01 and 1.51 nmol·m–3·h–1. There are ob-viously many possible explanations for the rapid change inDMS concentrations measured between January and Febru-ary, including the advection of a different water mass andchanges in phytoplankton assemblage and (or) microbial ac-tivity (Liss et al. 1997; Simó 2001). Our results neverthelesssuggest that the changes in macrozooplankton dominanceand their corresponding impact on DMS production were atleast partly responsible for the variations in DMS concentra-tions.
Bacteria attached to fecal pellets and damaged phyto-plankton cells are known to play an important role in thetransformation of DMSP and DMS (Scarratt et al. 2000).The effect of attached bacteria should have been negligibleduring our incubations, since fecal pellets were removed.However, attached bacteria may have been more abundantand (or) active on the fecal pellets from the krill than onthose from the salps, which contained mostly intact cells. Itis also important to consider the metabolism of DMSP bythe zooplankters themselves. Some copepod species can ac-cumulate a considerable amount of DMSP in their body, apool that may represent more than 70% of DMSPp in ambi-ent seawater (Tang et al. 1999, 2000b). It is assumed thatzooplankton grazing affects zooplankton body DMSP con-tent, thereby changing the partitioning of DMSP between theparticulate and dissolved phases with a potential effect onDMS production (Tang 2000; Tang et al. 2000a, 2000b).
There is considerable discussion regarding the existenceof competition between salps and krill in the Antarctic Ocean(e.g., Loeb et al. 1997; Kawaguchi et al. 1998). Nicol et al.(2000) showed clear separation of habitat by water massesbetween krill and salps. According to Pakhomov et al.(2002), krill and salps are adapted to different environmentalconditions. Whether or not they are in competition, theabundance of the two species exhibits considerable intra-and interannual variation in space and time (Loeb et al.1997; Kawaguchi et al. 1998). We believe that this importantphenomenon has a strong influence in DMS distribution.Our results clearly indicate the potential importance of zoo-
© 2004 NRC Canada
Kasamatsu et al. 741

plankton community structure and feeding mechanisms onDMS distributions in the Antarctic Ocean.
Moreover, if seawater temperature continues to increase,such as has already been observed in the Antarctic Peninsularegion (e.g., Rott et al. 1996), there is a possibility that salpsmay spread into the high Antarctic regions with strong im-plications for the Antarctic biogeochemistry. The negativeeffect that this shift in zooplankton dominance may have onDMS net production may, however, be compensated (or ex-acerbated) by parallel changes in the abundance of DMSP-rich phytoplankton species and (or) bacterial DMS yields. Afull appreciation of our results would thus require a betterknowledge of the overall effect of these climate variationson the whole plankton community and associated net DMSproduction.
Acknowledgments
We wish to acknowledge support from Shizuo Tsunogaiand Shinichiro Noriki, Hokkaido University. We would liketo thank Captain Andrew Leachman and the crew of the R/VTangaroa for their marvelous hospitality and efforts in help-ing us to carry out our sampling program. Thanks are ex-tended to Makoto Terazaki and Hiroshi Ogawa for providingspace aboard the R/V Hakuho-Maru. The captain and crewof the R/V Hakuho-Maru facilitated work in the AntarcticOcean. We are grateful to Stephen Nicol of Australian Ant-arctic Division (AAD) for providing us with krill. We thankJun Nishikawa, Sean Toczko, and Meguru Miki of the Uni-versity of Tokyo for providing us with useful comments onzooplankton and phytoplankton abundance. We wish tothank Steve Candy of the AAD for his helpful advice on ourstatistical treatments. We would also like to thank GrahamHosie of the AAD, Michael Scarratt of Maurice LamontagneInstitute, Michel Gosselin and his colleague of Université duQuébec à Rimouski for useful comments on the contents andsuggestions on the English. The manuscript was consider-ably improved by critical and constructive comments madeby the two anonymous referees and the Editors, GarySprules and Maurice Levasseur. This study was a part of theactivities of JARE-43 financially supported by the Ministryof Education, Culture, Sports, Science and Technology,Grant-in-Aid for Special Purpose 11800009.
References
Alldredge, A.L., and Madin, L.P. 1982. Pelagic tunicates: uniqueherbivores in the marine plankton. BioScience, 32: 655–663.
Aumont, O., Belviso, S., and Monfray, P. 2002. Dimethylsulfonio-propionate (DMSP) and dimethylsulfide (DMS) sea surface dis-tributions simulated from a global three-dimensional oceancarbon cycle model. J. Geophys. Res. 107(C4).
Cantin, G., Levasseur, M., Gosselin, M., and Michaud, S. 1996.Role of zooplankton in the mesoscale distribution of surfacedimethylsulfide concentrations in the Gulf of St. Lawrence,Canada. Mar. Ecol. Prog. Ser. 141: 103–117.
Charlson, R.J., Lovelock, J.E., Andreae, M.O., and Warren, S.G.1987. Oceanic phytoplankton, atmospheric sulphur, cloud albedoand climate. Nature (Lond.), 326: 655–661.
Dacey, J.W.H., and Wakeham, S.G. 1986. Oceanic dimethylsulfide:production during zooplankton grazing on phytoplankton. Sci-ence (Wash., D.C.), 233: 1314–1316.
Daly, K.L., and DiTullio, G.R. 1996. Particulate dimethylsulfonio-propionate removal and dimethylsulfide production by zooplank-ton in the Southern Ocean. In Biological and environmentalchemistry of DMSP and related sulfonium compounds. Edited byR.P. Kiene, P.T. Visscher, M.D. Keller, and G.O. Kirst. PlenumPress, New York. pp. 223–238.
Frost, B.W. 1972. Effects of size and concentration of food parti-cles on the feeding behavior of the marine planktonic copepodCalanus pacificus. Limnol. Oceanogr. 17: 805–815.
Harbison, G.R., McAlister, V.L., and Gilmer, R.W. 1986. The re-sponse of the salp, Pegea confoederata, to high levels of particu-late material: starvation in the midst of plenty. Limnol. Oceanogr.31: 371–382.
Huntley, M.E., Sykes, P.F., and Marin, V. 1989. Biometry andtrophodynamics of Salpa thompsoni foxton (Tunicata: Thaliacea)near the Antarctic Peninsula in austral summer, 1983–1984. Po-lar Biol. 10: 59–70.
Karsten, U., Kuck, K., Vogt, C., and Kirst, G.O. 1996. Dimethyl-sufoniopropionate production in phototrophic organisms and itsphysiological function as a cryoprotectant. In Biological envi-ronmental chemistry of DMSP and related sulfonium com-pounds. Edited by R.P. Kiene, P.T. Visscher, M.D. Keller, andG.O. Kirst. Plenum Press, New York. pp. 143–153.
Kawaguchi, S. 2001. Recent topics on Antarctic krill research.Kaiyo Monthly Spec. Issue, 27: 147–155. [In Japanese.]
Kawaguchi, S., Ichii, T., Naganobu, M., and de la Mare, W.K.1998. Do krill and salps compete? Contrary evidence from thekrill fisheries. CCAMLR Sci. 5: 205–216.
Kiene, R.P., and Linn, L.J. 2000. The fate of dissolved dimethyl-sulfoniopropionate (DMSP) in seawater: tracer studies using 35S-DMSP. Geochim. Cosmochim. Acta, 64: 2797–2810.
Kirchman, D.J., Sigda, J., Kapuscinski, R., and Mitchell, R. 1982.Statistical analysis of the direct count method for enumeratingbacteria. Appl. Environ. Microbiol. 44: 376–382.
Kirst, G.O. 1996. Osmotic adjustment in phytoplankton and macro-algae. In Biological environmental chemistry of DMSP and relatedsulfonium compounds. Edited by R.P. Kiene, P.T. Visscher, M.D.Keller, and G.O. Kirst. Plenum Press, New York. pp. 121–129.
Liss, P.S., Hatton, A.D., Malin, G., Nighingale, P.D., and Turner,S.M. 1997. Marine sulphur emissions. Philos. Trans. R. Soc.Lond. B Biol. Sci. 352: 159–169.
Loeb, V., Siegel, V., Holm-Hansen, O., Hewitt, R., Fraser, W.,Trivelpiece, W., and Trivelpiece, S. 1997. Effects of sea-ice ex-tent and krill or salp dominance on the Antarctic food web. Na-ture (Lond.), 387: 897–900.
Nicol, S., Pauly, T., Bindoff, N.L., Wright, S., Thiele, D., Hosie,G.W., Strutton, P.G., and Woehler, E. 2000. Ocean circulationoff east Antarctica affects ecosystem structure and sea-ice ex-tent. Nature (Lond.), 406: 504–507.
Niki, T., Kunugi, M., and Otsuki, A. 2000. DMSP-lyase activity infive marine phytoplankton species: its potential importance inDMS production. Mar. Biol. 136: 759–764.
Pakhomov, E.A., Froneman, P.W., and Perissinotto, R. 2002. Salp/krill interactions in the Southern Ocean: spatial segregation and im-plications for the carbon flux. Deep Sea Res. II, 49: 1881–1907.
Parsons, T.R., Maita, Y., and Lalli, C.M. 1984. A manual of chemi-cal and biological methods for seawater analysis. PergamonPress, Toronto, Ont.
Rott, H., Skvarca, P., and Nagler, T. 1996. Rapid collapse of north-ern Larsen Ice Shelf, Antarctica. Science (Wash., D.C.), 271:788–792.
Scarratt, M., Cantin, G., Levasseur, M., and Michaud, S. 2000. Partic-ulate size-fractionated kinetics of DMS production: where doesDMSP cleavage occur at the microscale? J. Sea Res. 43: 245–252.
© 2004 NRC Canada
742 Can. J. Fish. Aquat. Sci. Vol. 61, 2004

Simó, R. 2001. Production of atmospheric sulfur by oceanic plank-ton: biological, ecological and evolutionary links. Trends Ecol.Evol. 16: 287–294.
Strom, S.L., Benner, R., Ziegler, S., and Dagg, M.J. 1997. Plank-tonic grazers are a potentially important source of marine dis-solved organic carbon. Limnol. Oceanogr. 42: 1364–1374.
Suzuki, R., and Ishimaru, T. 1990. An improved method for the de-termination of phytoplankton chlorophyll using N,N-dimethyl-formamide. J. Oceanogr. Soc. Jpn. 46: 190–194.
Tang, K.W. 2000. Dynamics of dimethylsulfoniopropionate (DMSP)in a migratory grazer: a laboratory simulation study. J. Exp. Mar.Biol. Ecol. 243: 283–293.
Tang, K.W., Dam, H.G., Visscher, P.T., and Fenn, T.D. 1999.Dimethylsulfoniopropionate (DMSP) in marine copepods and itsrelation with diets and salinity. Mar. Ecol. Prog. Ser. 179: 71–79.
Tang, K.W., Fenn, T.D., Visscher, P.T., and Dam, H.G. 2000a. Reg-ulation of body dimethylsulfoniopropionate (DMSP) content by
the copepod Temora longicornis: a test of four mechanisms.Mar. Biol. 136: 749–757.
Tang, K.W., Rogers, D.R., Dam, H.G., and Visscher, P.T. 2000b.Seasonal distribution of DMSP among seston, dissolved matterand zooplankton along a transect in the Long Island Sound estu-ary. Mar. Ecol. Prog. Ser. 206: 1–11.
Uzuka, N., Watanabe, S., and Tsunogai, S. 1996. Dimethylsulfidein coastal zone of the east China Sea. J. Oceanogr. 52: 313–321.
Watanabe, S., Demura, K., and Tsunogai, S. 1987. Determinationof volatile organic sulfur compound concentrations in seawater.Bull. Fac. Fish. Hokkaido Univ. 38: 286–292.
Wolfe, G.B., Levasseur, M., Cantin, G., and Michaud, S. 2000.DMSP and DMS dynamics and microzooplankton grazing in theLabrador Sea: application of the dilution technique. Deep SeaRes. I, 47: 2243–2264.
Zar, J.H. 1996. Biostatistical analysis. 3rd ed. Prentice-Hall,Englewod Cliffs, N.J.
© 2004 NRC Canada
Kasamatsu et al. 743