phytoplankton community ecology: principles applied in san
TRANSCRIPT

MARINE ECOLOGY PROGRESS SERIESMar Ecol Prog Ser
Vol. 285: 11–28, 2005 Published January 19
INTRODUCTION
The challenge of deciphering the rules of phyto-plankton community assembly remains a central prob-lem of aquatic ecology. Phytoplankton photosynthesisfixes up to 50 Gt of carbon per year, contributing nearlyhalf of global primary production (Falkowski et al.1998). The biogeochemical consequences and fate ofthat production are determined by the way in which itis packaged into species having distinct biochemicalcontents and cell sizes spanning 6 orders of magnitude.Diatom production is a net sink for CO2, whereas coc-colithophorid production is a net source, so communitycomposition at this level determines the direction of
ocean–atmosphere CO2 exchange, with correspondinginfluence on the climate system (Harrison 2000).Micron-sized picoplankton are not directly accessibleto most metazoan consumers (Tamigneaux et al. 1995),so their energy content is routed through the microbialloop, with large respiratory losses. Some algal taxasynthesize biochemicals that are essential dietary com-ponents for animal consumers (Müller-Navarra et al.2004), while others produce potent toxins to disruptanimal growth or reproduction (Cembella 2003). Thepathways and efficiencies of energy transfer from pri-mary producers to aquatic food webs, including thosesustaining upper trophic levels, are therefore deter-mined by phytoplankton community composition.
© Inter-Research 2005 · www.int-res.com*Email: [email protected]
Phytoplankton community ecology: principlesapplied in San Francisco Bay
James E. Cloern1,*, Richard Dufford2
1US Geological Survey, MS496, 345 Middlefield Road, Menlo Park, California 94025, USA21845 Orchard Place, Fort Collins, Colorado 80521, USA
ABSTRACT: In his seminal 1961 paper ‘The paradox of the plankton’ Am Nat 95:137–147, G. E.Hutchinson asked why many species of phytoplankton can coexist while competing for a small num-ber of limiting resources in an unstructured habitat. Hutchinson anticipated the resolution of hisparadox, recognizing that communities are organized by processes beyond resource competitionincluding species interactions, habitat variability and dispersal. Since 1961 we have made funda-mental discoveries that have revolutionized our conceptual understanding of pelagic ecology, includ-ing (1) habitat heterogeneity at all scales relevant to plankton population dynamics, (2) communityshifts in response to global climate cycles, (3) fast and selective predation as a powerful top–downforce to shape phytoplankton communities, (4) turbulent mixing as a physical process that selectsspecies on the basis of their size and form, (5) mixotrophy that allows some algal species to taporganic nutrient pools and function at multiple trophic levels, (6) taxon-specific life cycles includingalternating vegetative and resting stages, and (7) the pelagic as an open system where communitiesare continually reshaped by species immigration. Here we synthesize these discoveries to show howthey validate and amplify Hutchinson’s hypothesis that phytoplankton communities are assembledby many processes. Our synthesis is built around observations of phytoplankton species compositionfrom a decade of study in San Francisco Bay, used as a case study to illustrate the contemporary prin-ciples of phytoplankton community ecology. We apply these principles to address 2 central questions:(1) What processes assemble phytoplankton communities? (2) How does phytoplankton communitycomposition influence ecosystem functions such as production in pelagic and benthic food webs?
KEY WORDS: Phytoplankton · Community ecology · Ecosystem functions · Aquatic ecosystems ·Food webs · Biological diversity · Paradox of the plankton
Resale or republication not permitted without written consent of the publisher

Mar Ecol Prog Ser 285: 11–28, 2005
In his seminal paper ‘The paradox of the plankton’,Hutchinson (1961) asked how ‘is it possible for a numberof species to coexist in a relatively isotropic or unstruc-tured environment all competing for the same sorts ofmaterials’? The paradox was coexistence of many spe-cies when the competitive exclusion principle predicts‘that one species alone would out compete all the othersso that in a final equilibrium situation the assemblagewould reduce to a population of a single species.’ Theparadox was grounded on assumptions that: (1) phyto-plankton communities are assembled on the basis ofdifferential growth rates among species, determined bythe availability of inorganic nutrients, (2) species inter-act through competition for mineral nutrients, and (3)pelagic habitats are homogeneous closed systems. In the4 decades since, our concepts of pelagic ecology havebeen revolutionized by fundamental new discoveriesthat resolve the paradox, largely through confirmation ofmechanisms anticipated by Hutchinson (1961): speciesimmigration, biological interactions, and habitat vari-ability that precludes community equilibrium.
Here, we synthesize those discoveries and their con-tribution to the revolutionized view of pelagic ecologysince Hutchinson’s era. We build this synthesis withexamples of species occurrence and distributional pat-terns from a decade-long survey of phytoplanktoncommunities in San Francisco Bay. We use this casestudy as a foundation to synthesize important dis-coveries since Hutchinson’s paper to address 2 funda-mental ecological questions: (1) What are the patternsand processes of phytoplankton community assemblyin open ecosystems such as estuaries and oceaniccurrent systems? and (2) How does phytoplanktoncommunity composition influence ecosystem functionssuch as food supply to consumers and energy transferto pelagic and benthic food webs?
MATERIALS AND METHODS
San Francisco Bay: the case study. San FranciscoBay, the estuary between California’s 2 largest riversand the Pacific Ocean, is a site of long-term ecologicalresearch designed to reveal patterns and processes ofvariability in a coastal ecosystem (Cloern 1996). Estu-aries are open systems strongly influenced by riverinflows, mixing with the coastal ocean, and exchangesacross the sediment–water interface that drive physi-cal and biogeochemical variability. Estuaries are tur-bulent advective systems in which the pelagic andits biota are dispersive and transient. Their pelagichabitats are time-varying mixtures of seawater andfreshwater, situated within spatial gradients of salinity,temperature, nutrients, turbidity and heterotroph com-munities along a river–ocean continuum. Long-term
observations across the habitat gradients of San Fran-cisco Bay provide an appropriate data set for illustrat-ing principles that have emerged from the discovery ofprocesses beyond resource competition that shapephytoplankton communities.
Methods. For a decade (1992 to 2001), we sampledseasonally along the salinity gradients of San FranciscoBay between the tidal freshwater Sacramento River,marine-influenced Central Bay, and brackish-marineSouth San Francisco Bay (Fig. 1). We measured salin-ity, temperature, irradiance, phytoplankton biomass aschlorophyll a, suspended sediment and dissolved in-organic nutrient (N, P, Si) concentrations to capturevariability associated with annual cycles of river flowand biomass and activity of herbivores (Cloern 1996).Phytoplankton samples were collected within the 6sub-regimes of the river-estuary system (Fig. 1), identi-fied from tree-based regressions of chlorophyll dis-tributions (Jassby et al. 1997). These 6 primary stationswere sampled during spring 1992 and then roughlybimonthly each year from 1993 through 2001 (seeFig. 2E). Additional (ca. weekly) samples were collectedin South Bay when the biomass fluctuated aroundneap-spring periodicity of tidal mixing.
Vertical profiles of temperature, salinity and photo-synthetically active radiance (Iz) were measured with aSeabird Electronics CTD and LiCor 192 quantum sen-sor. The light-attenuation coefficient k was calculatedas the slope of linear regression of ln(Iz) againstdepth z. Vertical distributions of suspended particulatematter (SPM) and chlorophyll a were measured with aD&A Instruments optical backscatter sensor and Sea-Tech fluorometer calibrated with discrete measuresof SPM and chlorophyll a in near-surface and near-bottom water samples. Chlorophyll a samples werefiltered onto Gelman A/E glass-fiber filters and frozen.Dried filters were ground in 90% acetone, extracted ina freezer for 24 h, and the absorbance of centrifugedextracts measured with a Hewlett Packard 8452Adiode-array spectrophotometer. Chlorophyll a concen-tration was calculated using Lorenzen’s (1967) equa-tions with corrections for phaeopigments. SPM wasdetermined gravimetrically using samples collectedonto pre-weighed Nuclepore (0.4 µm pore size) filters.From 1992 through 2001, we measured 4118 verticalprofiles along the river–estuary transect. We present(Fig. 2) only measurements at 1 m depth, correspond-ing to the depth of phytoplankton sampling.
Nutrient samples were filtered through 0.4 µm Nucle-pore filters and frozen until analyzed. Concentrationsof dissolved inorganic nitrogen (DIN = NH4
+ + NO3– +
NO2–), reactive phosphorus and silica were mea-
sured using modifications of standard colorimetricmethods (Hager & Schemel 1997) with a TechniconAutoAnalyzer II. We computed indices of nutrient
12

Cloern & Dufford: Phytoplankton community ecology
limitation as measured concentrations normalized byhalf-saturation constants for growth (K), assuming thatKN = 0.7 µM, KP = 0.1 µM, KSi = 1.8 µM (Chapra 1997).We computed an index of light limitation as meanwater-column irradiance [= E/(kH), where H is waterdepth and E is daily surface irradiance], scaled by acharacteristic irradiance at which growth rate is half themaximum (KI = 2.4 mol quanta m–2 d–1; Cloern 1999).
Phytoplankton samples were preserved with acidLugol’s solution, and 2 to 50 ml aliquots were settled inchambers for 6 to 24 h, and then counted and identifiedusing a phase-contrast inverted microscope (Utermöhl1958). Entire aliquots were examined at 125× magnifi-cation and all cells >30 µm diameter were enumerated.Cells smaller than 30 µm were counted at 1250× magni-fication; at least 100 cells of the most numerous taxonwere counted using the strip-count method (APHA1989). Diatoms and dinoflagellates were identified afterthe cell contents were cleared in 30% hydrogen per-oxide and mounted in Hyrax Mounting Medium. Cellvolumes were estimated for dominant taxa by measuring50 to 100 cells (Hillebrand et al. 1999) and applying thegeometric formulas of Wetzel & Likens (1991). The bio-mass of each taxon was computed as the product ofabundance (cells ml–1) and cell biovolume (µm3 ml–1).
From mean cell dimensions we computed the size ofeach species as equivalent spherical diameter (ESD).
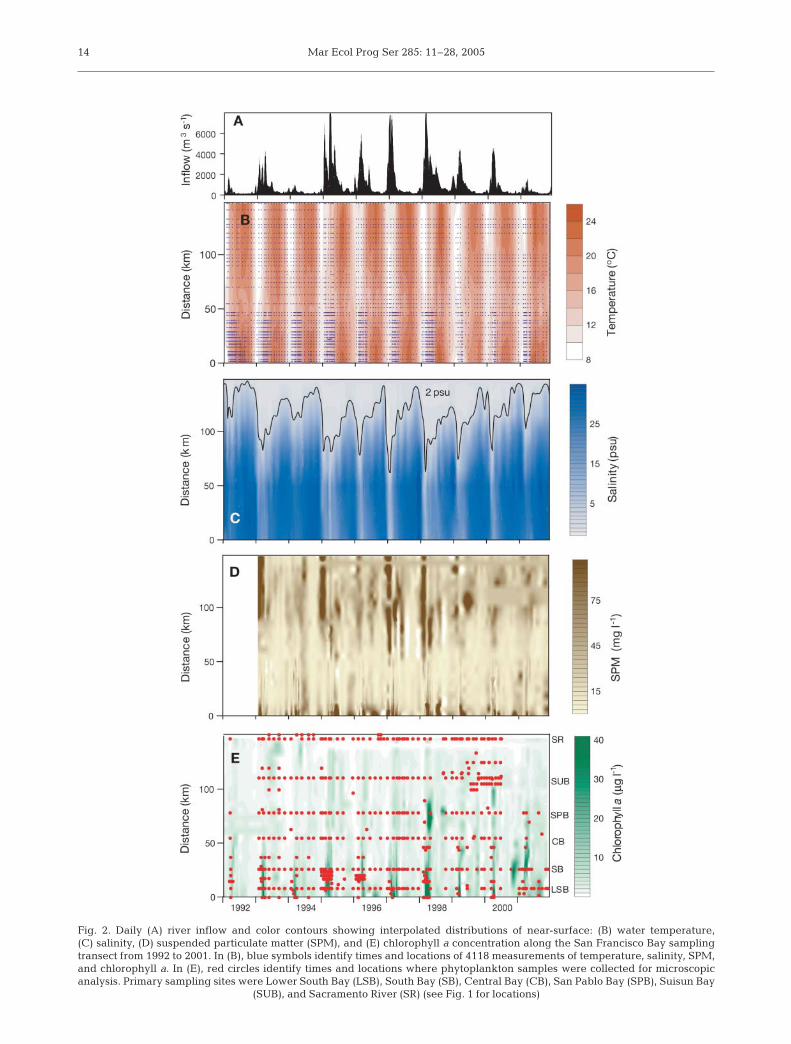
San Francisco Bay as a habitat mosaic. Phytoplank-ton communities were sampled across a mosaic ofphysical–chemical habitats, where the variability ofsalinity (0.05 to 32.2 psu), temperature (8.7 to 22.7°C),turbidity (SPM from 1 to 556 mg l–1) and nutrient con-centrations (dissolved inorganic nitrogen, DIN, from0.05 to 221 µM) reflected a broad range of environ-mental factors that regulate phytoplankton growth(Fig. 2). Hydrologic conditions ranged from seasonallow riverine inflow of ~100 m3 s–1 during summer andautumn to exceptional flood events (>9000 m3 s–1,Fig. 2A). The salinity distribution responded with rapidestuary-wide displacements of seawater and its plank-ton community during periods of high inflow followedby progressive salinity intrusion during the low flow insummer and autumn (Fig. 2C). Spatial distributions ofSPM (Fig. 2D) created large river–ocean gradients inturbidity and light limitation of phytoplankton growth.Most samples had chlorophyll a concentrations <3 µgl–1. Exceptions occurred as blooms, mostly in SouthBay (Fig. 2E), when chlorophyll a exceeded 20 µg l–1
and reached maxima >100 µg l–1. DIN, Si and P usu-ally exceeded concentrations that limit phytoplankton
13
Fig. 1. Sampling locations in SanFrancisco Bay (�). Vertical profilesof chlorophyll a, suspended partic-ulate matter, irradiance, tempera-ture, and salinity were measured atall stations. Phytoplankton and nu-trient samples were collected at theprimary stations distributed among6 subregions along the salinity gra-dient. Data are available at http://sfbay.wr.usgs.gov/access/wqdata
Mar Ecol Prog Ser 285: 11–28, 200514
Fig. 2. Daily (A) river inflow and color contours showing interpolated distributions of near-surface: (B) water temperature, (C) salinity, (D) suspended particulate matter (SPM), and (E) chlorophyll a concentration along the San Francisco Bay samplingtransect from 1992 to 2001. In (B), blue symbols identify times and locations of 4118 measurements of temperature, salinity, SPM,and chlorophyll a. In (E), red circles identify times and locations where phytoplankton samples were collected for microscopicanalysis. Primary sampling sites were Lower South Bay (LSB), South Bay (SB), Central Bay (CB), San Pablo Bay (SPB), Suisun Bay
(SUB), and Sacramento River (SR) (see Fig. 1 for locations)

Cloern & Dufford: Phytoplankton community ecology
growth. Scaled concentrations of DIN (N' = DIN/KN)indicate that phytoplankton growth was potentiallylimited by N (N' <1) in only 4% of samples (Fig. 3). Siwas at rate-limiting concentrations for diatoms (Si' <1)in only 1% of samples, and P limitation was never indi-cated. Irradiance profiles indicated light limitation ofgrowth in 74% of samples (Fig. 3D), confirming thatphytoplankton growth is predominately limited by thelight resource in this enriched estuary (Cloern 1999).
Phytoplankton community of San Francisco Bay. Wedetected 500 distinct phytoplankton taxa, of which 396could be identified to species. Accurate enumeration andidentification of prokaryotes and some small eukaryoticalgae is not possible by light microscopy, and some frag-ile taxa (e.g. the raphidophyte Heterosigma akashiwo)can only be detected in live samples. Therefore, ouranalysis of size and taxonomic diversity is constrained tothose forms that were well-preserved and identifiableby light microscopy. Of these taxa, cell volume rangedfrom 1 to 3 µm3 for the smallest taxa (Nannochloropsissp., Chromulina mikroplankton, Merismopedia spp.,Aphanocapsa spp.) to 4–6 × 105 µm3 for the largest(Coscinodiscus centralis var. pacifica, Thalassiosiraanguste-lineata, Coscinodiscus oculus-iridis, Noctilucascintillans). We compiled mean cell size for the 81 mostimportant species (Table 1), a community of cosmo-politan phytoplankton commonly observed in temperateestuaries and coastal waters globally.
The phytoplankton community was dominated by asmall number of species: the top 10 contributed 77% tocumulative biomass in the 599 samples, and the top100 species contributed >99% (Fig. 4A). Diatoms (divi-sion Bacillariophyta) contributed 81% to cumulativebiomass in all samples (Fig. 4B). Dinoflagellates (Pyrro-phyta) and cryptophytes contributed 11 and 5%,respectively, to cumulative biomass. Other divisions(Chlorophyta, Cyanophyta, Chrysophyta, Eugleno-phyta) were minor components of overall biomass,although their contributions were important in someindividual samples. We probed the San Francisco Baydata set to extract patterns illustrating principles ofphytoplankton community assembly that have emergedfrom discoveries since Hutchinson’s era.
RESULTS AND DISCUSSION
Eight principles of phytoplankton community assembly
(1) Cell size is determined by nutrient supply andselective grazing: Major conceptual breakthroughs ofpelagic ecology have come from discoveries of themicrobial loop (Pomeroy 1974, Azam et al. 1983) andthe distinction between new and regenerated produc-
15
A
B
C
0 3 6 9I'
0
20
40
60
Occ
urr
en
ces
0 30 60 90P'
0
20
Occ
urr
en
ces
0 30 60 90Si'
0
8
16
Occ
urr
en
ces
0 30 60 90N'
0
20
Occ
urr
en
ces
D
N Limitation 4% < 1
P Limitation 0% < 1
Si Limitation 1% < 1
Light Limitation 74% < 1
Fig. 3. Frequency distributions (number of occurrences) of nu-trient (N, Si, P) concentrations and water-column irradiance(I), measured in San Francisco Bay from 1992 to 2001. Mea-surements were normalized to half-saturation constantsdefining resource limitation of phytoplankton growth. Note
difference in x-axis scale between (A–C) and (D)

Mar Ecol Prog Ser 285: 11–28, 200516
Table 1. Common phytoplankton taxa in San Francisco Bay, listing those species occurring in >10 (of 599) samples and contribut-ing >0.01% of cumulative biomass as biovolume contained in all samples. Mean cell size as equivalent spherical diameter (ESD).
n = no. of occurrences
Taxon Division Biomass (%) ESD (µm) n
Thalassiosira rotula Meunier Bacillariophyta 20.970 28 140Chaetoceros socialis Lauder Bacillariophyta 11.980 12 48Skeletonema costatum (Greville) Cleve Bacillariophyta 9.51 8 277Ditylum brightwellii (T. West) Grunow Bacillariophyta 7.44 85 103Gymnodinium sanguineum Hirasaka Pyrrophyta 7.40 61 41Coscinodiscus oculus-iridis Ehrenberg Bacillariophyta 6.30 101 66Thalassiosira hendeyi Hasle & Fryxell Bacillariophyta 4.85 51 203Thalassiosira punctigera (Castracane) Hasle Bacillariophyta 3.31 72 27Plagioselmis prolonga var. nordica Novarino, Lucas & Morrall Cryptophyta 2.65 5 495Coscinodiscus curvatulus Grunow Bacillariophyta 2.15 73 49Mesodinium rubrum Lohmann Holotrich ciliate 2.03 30 190Teleaulax amphioxeia (Conrad) Hill Cryptophyta 2.02 9 375Chaetoceros debilis Cleve Bacillariophyta 1.84 11 31Eucampia zodiacus Ehrenberg Bacillariophyta 1.80 30 29Coscinodiscus radiatus Ehrenberg Bacillariophyta 1.77 53 66Thalassiosira eccentrica (Ehrenberg) Cleve Bacillariophyta 1.48 55 136Protoperidinium sp. Pyrrophyta 1.45 37 21Thalassiosira decipiens (Grunow) Jorgensen Bacillariophyta 0.90 20 136Coscinodiscus centralis var. pacifica Gran & Angst Bacillariophyta 0.72 103 23Rhizosolenia setigera Brightwell Bacillariophyta 0.49 29 102Noctiluca scintillans (Macartney) Kofoid & Swezy Pyrrophyta 0.46 98 36Nitzschia bilobata W. Smith Bacillariophyta 0.44 35 33Cyclotella atomus Hustedt Bacillariophyta 0.43 7 304Coscinodiscus jonesianus (Greville) Ostenfeld Bacillariophyta 0.33 68 17Pyramimonas orientalis Butcher Chlorophyta 0.31 5 175Rhodomonas marina (Dangeard) Lemmermann Cryptophyta 0.26 10 116Protoperidinium depressum (Bailey) Balech Pyrrophyta 0.23 60 41Heterocapsa triquetra (Ehrenberg) Stein Pyrrophyta 0.23 18 138Protoperidinium claudicans (Paulsen) Balech Pyrrophyta 0.22 26 52Cyclotella choctawhatcheeana Prasad Bacillariophyta 0.22 12 74Alexandrium tamarense (Lebour) Balech Pyrrophyta 0.21 23 109Nannochloropsis sp. Chrysophyta 0.20 2 424Thalassiosira nodulolineata (Hendey) Hasle & Fryxell Bacillariophyta 0.18 49 40Chlorella salina Butcher Chlorophyta 0.17 3 97Chaetoceros wighamii Brightwell Bacillariophyta 0.17 9 43Eutreptia lanowii Steur Euglenophyta 0.16 11 167Prorocentrum minimum (Pavillard) Schiller Pyrrophyta 0.16 14 116Aulacoseira lirata (Ehrenberg) Ross Bacillariophyta 0.15 12 12Rhizosolenia styliformis Brightwell Bacillariophyta 0.13 48 22Entomoneis paludosa (W. Smith) Reimer Bacillariophyta 0.12 28 30Odontella mobiliensis (Bailey) Grunow Bacillariophyta 0.11 71 17Gyrosigma balticum (Ehrenberg) Rabenhorst Bacillariophyta 0.11 53 36Pleurosigma strigosum W. Smith Bacillariophyta 0.09 32 61Leptocylindrus minimus Gran Bacillariophyta 0.09 11 36Gyrodinium spirale (Bergh) Kofoid & Swezy Pyrrophyta 0.09 51 21Thalassiosira visurgis Hustedt Bacillariophyta 0.08 21 50Oxytoxum milneri Murray & Whitting Pyrrophyta 0.08 28 75Lithodesmium undulatum Ehrenberg Bacillariophyta 0.08 69 14Gyrosigma fasciola (Ehrenberg) Griffith & Henfrey Bacillariophyta 0.08 32 91Cyclotella striata (Kuetzing) Grunow Bacillariophyta 0.08 13 63Heterocapsa rotundata (Lohmann) Hansen Pyrrophyta 0.07 11 94Pyramimonas disomata Butcher Chlorophyta 0.07 14 112Chaetoceros subtilis Cleve Bacillariophyta 0.07 7 57Chaetoceros gracilis Schutt Bacillariophyta 0.05 7 23Asterionellopsis glacialis (Castracane) Round Bacillariophyta 0.05 20 33Guinardia delicatula (Cleve) Hasle Bacillariophyta 0.05 21 100Chlorella marina Butcher Chlorophyta 0.05 5 86Paralia sulcata (Ehrenberg) Cleve Bacillariophyta 0.05 13 166Actinoptychus senarius Ehrenberg Bacillariophyta 0.05 58 39

Cloern & Dufford: Phytoplankton community ecology
tion (Dugdale & Goering 1967). We know now thatphytoplankton biomass and production are dominatedby micron-sized picoplankton in oligotrophic lakes andmuch of the ocean. Small size (high surface:volumeratio) provides a competitive advantage in nutrient-impoverished regenerating systems (Chisholm 1992),but this advantage disappears in new-production sys-tems when physical processes inject nutrients into the
euphotic zone and promote selective growth of largecells that escape predation by fast-responding micro-heterotrophs (Kiørboe 1993). As predicted from thisprinciple, phytoplankton biomass in the nutrient-richSan Francisco Bay is dominated by large taxa: cells<8 µm contributed only 4% of community biomass, butcells >30 µm contributed 40% (Fig. 4B). This is con-sistent with an earlier study demonstrating that ultra-
17
Table 1 (continued)
Taxon Division Biomass (%) ESD (µm) n
Pleurosigma normanii Ralfs Bacillariophyta 0.04 47 17Pseudo-nitzschia seriata (Cleve) Peragallo Bacillariophyta 0.04 15 34Cyclotella meneghiniana Kuetzing Bacillariophyta 0.04 13 57Gyrosigma acuminatum (Kuetzing) Rabenhorst Bacillariophyta 0.04 42 24Melosira varians Agardh Bacillariophyta 0.03 22 26Nitzschia pusilla Grunow Bacillariophyta 0.03 7 69Alexandrium catenella (Whedon & Kofoid) Balech Pyrrophyta 0.03 26 22Ceratium furca (Ehrenberg) Claparede & Lachmann Pyrrophyta 0.03 41 11Thalassiosira nordenskioeldii Cleve Bacillariophyta 0.03 16 40Gyrosigma macrum (W. Smith) Griffith & Henfrey Bacillariophyta 0.03 31 71Stephanodiscus hantzschii Grunow Bacillariophyta 0.03 11 11Prorocentrum micans Ehrenberg Pyrrophyta 0.02 30 29Scrippsiella hangoei (Schiller) Larsen Pyrrophyta 0.02 32 18Prorocentrum gracile Schutt Pyrrophyta 0.02 20 17Aulacoseira granulata (Ehrenberg) Simonsen Bacillariophyta 0.02 14 32Synechocystis salina Wislouch Cyanophyta 0.02 2 61Aulacoseira italica (Ehrenberg) Simonsen Bacillariophyta 0.02 13 18Protoperidinium bipes (Paulsen) Balech Pyrrophyta 0.02 22 43Nitzschia closterium (Ehrenberg) W. Smith Bacillariophyta 0.02 7 122Nitzschia longissima (Brebisson) Ralfs Bacillariophyta 0.02 11 143Thalassionema nitzschioides (Grunow) Mereschkowsky Bacillariophyta 0.02 13 47Cryptomonas ovata Ehrenberg Cryptophyta 0.02 15 27
10 10020 40 60 808640
0.2
0.4
0.6
0.8
1
Diatoms 81%
Dinoflagellates 11%
Cryptophytes 5%
Mesodinium 2%
B
1 10 1000
0.2
0.4
0.6
0.8
1
0.77
0.99
A
Fig. 4. (A) Cumulative distribution of total phytoplankton biomass in 599 samples versus number of species included, ranked bybiomass; 10 most important species contributed 77% of cumulative biomass, and 100 species accounted for > 99% of cumulativebiomass. (B) Cumulative frequency distribution of phytoplankton biomass versus cell size (ESD, equivalent spherical diameter);top inset shows relative contributions to total biomass by diatoms, dinoflagellates, cryptophytes and phototrophic ciliate Meso-dinium rubrum. Photomicrograph inset shows a sample collected during a spring diatom bloom dominated by Thalassiosira
punctigera having mean diameter ~90 µm

Mar Ecol Prog Ser 285: 11–28, 2005
plankton (cells passing a 5 µm filter) contributed only3 to 28% to chlorophyll biomass and annual primaryproduction in different regions of San Francisco Bay(Cole et al. 1986). Much of the large-cell productionoccurred during blooms, when biomass was dominatedby Thalassiosira rotula, T. hendeyi, T. punctigera,Chaetoceros socialis, C. debilis, Skeletonema costa-tum, Ditylum brightwellii, Coscinodiscus oculus-iridis,C. curvatulus, C. radiatus and Eucampia zodiacus(Table 1). Analyses by epifluorescence microscopy(Ning et al. 2000) confirmed that picoplankton (primar-ily cyanobacteria Synechococcus spp.) contributedabout 15% to community biomass during nonbloomconditions and only 2% during blooms. Phytoplanktondynamics in upwelling systems and many estuariesare characterized by boom–bust cycles of large cellsagainst a background of stable picoplankton biomass(Cullen et al. 2002). The damped biomass fluctuationsof small cells result from tight coupling between theirproduction and consumption by fast-responding pro-tistans (Murrell & Hollibaugh 1998). As a consequenceof fast grazing in the microbial loop and laggedresponses of metazoan consumers (Malone 1992),nutrient enrichment amplifies large-cell production(Duarte et al. 2000).
(2) Diatoms respond rapidly to nutrient pulses: Thephytoplankton community of San Francisco Bay isoverwhelmingly dominated by diatoms (Fig. 4B), aresult of dominance during blooms when their cell-division rate exceeds loss rates from grazing, sedimen-tation, transport and mortality. Large diatoms are keyspecies in this estuary just as they are in the oceanfollowing physically-driven events such as upwellingevents that inject nutrients into the euphotic zone(Goldman & McGillicuddy 2003). The bloom mecha-nism in nutrient-rich San Francisco Bay is a releasefrom strong light limitation by other physical pro-cesses, including runoff-induced salinity stratification(Cloern 1996) or increased euphotic-zone depth assediment suspension is damped during weak neaptides (May et al. 2003).
Community studies in San Francisco Bay and in up-welling systems support the empirically-based princi-ple that diatoms respond rapidly to episodic high-lighthigh-nutrient conditions. Surprisingly, there is no con-sensus explanation of why diatoms best exploit theseopportunities for fast population growth. One view isfrom the bottom up: diatoms divide faster than othertaxa, either because they have inherently high growthrates (Smayda 1997), accelerated N assimilation undernitrate-rich conditions (Dugdale & Wilkerson 1992),high growth efficiency at low light (Goldman & Mc-Gillicuddy 2003), or because they can utilize bicarbon-ate during blooms when CO2 limits photosynthesis ofother algae (Hobson 1988). Other explanations work
from the top down: diatoms succeed because theirsilica cell wall resists the cracking force of feedingstructures such as copepod mandibles (Hamm et al.2003) and attack by small predatory flagellates orpathogens (Smetacek 1995). Large diatoms can regu-late their buoyancy (Waite et al. 1992) to suppress sink-ing, and this is important in shallow environments suchas San Francisco Bay, where most phytoplankton pro-duction is consumed by benthic suspension feedersand avoidance of the benthic boundary layer is a key topopulation survival.
(3) Pelagic habitats select phytoplankton species onthe basis of their form and function: Hutchinson’sparadox came from the premise that phytoplanktonspecies are functional equivalents competing for finiteresources in an ‘isotropic’ fluid medium. However,Margalef (1978) later recognized the ecological signif-icance of variability among species in their form (size,density) and function (motility, behavior) as adapta-tions to an ‘unstable and turbulent environment’.Margalef’s Mandala is now a central paradigm ofphytoplankton community ecology, expressing theprinciple that phytoplankton life forms are adaptationsto specific pelagic habitat types defined along gradi-ents of turbulence intensity and nutrient concentra-tions (Cullen et al. 2002). As an alternative to theresource-competition perspective, Margalef (1978)postulated that the ‘combination of sedimentation withturbulence’ shapes communities as the pelagic variesbetween extremes of a fertile-turbulent state (pro-moting diatom growth) and exhausted-stratified state(promoting growth of flagellates and dinoflagellatesthat migrate vertically to exploit nutrient gradients;Smayda & Reynolds 2001). This framework provides abasis for exploring the diverse patterns of speciesoccurrence in estuaries. One pattern in San FranciscoBay is restricted seasonal occurrence, exemplified bythe marine diatoms (Fig. 5A) that dominate springblooms. A second pattern, exemplified by Mesodiniumrubrum and some dinoflagellates (Fig. 5B), is nonsea-sonal population growth in response to short-termevents, such as sunny calm weather that establishes anephemeral thin upper layer within which motile spe-cies accumulate (Cloern et al. 1994). Gyrosigma fasci-ola illustrates a different nonseasonal pattern (Fig. 5C)driven by wind events and periodic tidal stresses thatsuspend benthic forms into the plankton.
Margalef’s (1978) conceptual framework also pro-vides a foundation for exploring the diversity ofcommunity dynamics among coastal ecosystems. Thetextbook depiction of phytoplankton seasonality attemperate latitudes is dominance by diatoms in springand dinoflagellates in summer, but this idealized sea-sonal succession does not occur in San Francisco Bayor in the Bay of Brest (perennial diatom dominance:
18

Cloern & Dufford: Phytoplankton community ecology 19
0
10
20
30S
alin
ity (
psu
)
A Coscinodiscus oculus-iridis
D Plagioselmis prolonga
B Alexandrium catenella
0
10
20
30
Sa
linity
(p
su)
C Gyrosigma fasciola
0
10
20
30
Sa
linity
(p
su)
E Skeletonema potamos F Prorocentrum gracile
Spring bloom
J F M A M J J A S O N D J F M A M J J A S O N D
1992 1994 1996 1998 2000
0
10
20
30
Sa
linity
(p
su)
G Coscinodiscus marginatus
1992 1994 1996 1998 2000
H Thalassiosira nodulolineata
Fig. 5. (A–F) Six different patterns of phytoplankton seasonal occurrence and distribution along the estuarine salinity gradient ofSan Francisco Bay; size of data points is proportional to log(biomass) of individual species in each sample; small dots: samplesfrom which the species was absent. (A) Dominant during spring blooms; (B) episodic occurrences of planktonic species;(C) episodic occurrences of benthic species in plankton; (D) persistent and cosmopolitan distribution; (E) input of freshwaterspecies by river inflow; (F) input of coastal ocean species. (G, H) Changes during the decade of observation, showing coastal
species that disappeared after 1996 (G) and appeared after 1997 (H)
Mar Ecol Prog Ser 285: 11–28, 200520
Anabaena affinis Synedra ulna Cyclotella pseudostelligera Chlorella vulgaris Cryptomonas ovata Fragilaria crotonensis Oscillatoria agardhii Melosira lineata Aulacoseira granulata Aphanizomenon flos-aquae Merismopedia tenuissima Skeletonema potamos Scenedesmus acuminatus Bacillaria paxillifer Euglena proxima Stephanodiscus hantzschii Oscillatoria lutea Monoraphidium convolutum Nitzschia sigma Aulacoseira lirata Gyrosigma acuminatum Cyclotella meneghiniana Cyclotella striata Cyclotella choctawhatcheeana Cyclotella atomus Plagioselmis prolongaNannochloropsis sp.Gyrosigma macrum Teleaulax amphioxeia Skeletonema costatum Thalassiosira decipiens Chaetoceros subtilis Coscinodiscus radiatus Prorocentrum aporum Gyrosigma fasciola Chaetoceros wighamii Thalassiosira nordenskioeldii Pyramimonas disomata Mesodinium rubrum Oxytoxum milneriAlexandrium tamarense Prorocentrum minimum Thalassiosira rotula Coscinodiscus oculus-iridis Scrippsiella hangoei Heterocapsa triquetra Chaetoceros didymus Ditylum brightwellii Heterocapsa rotundata Eutreptia lanowii Gonyaulax spinifera Chaetoceros socialis Paralia sulcata Nitzschia closterium Alexandrium minutum Thalassiosira nodulolineata Chaetoceros decipiens Protoperidinium bipes Navicula distans Rhizosolenia setigera Alexandrium catenella Noctiluca scintillansThalassionema nitzschioides Chaetoceros debilis Prorocentrum micans Asterionellopsis glacialis Protoperidinium pellucidum Gymnodinium sanguineum Leptocylindrus danicus Protoperidinium brevipes Gyrodinium spirale Prorocentrum gracile Dinophysis acuminataRhizosolenia hebetata Thalassiosira punctigera Detonula confervacea Pyramimonas parkeae Biddulphia pulchella Ceratium furca Thalassionema frauenfeldii
0 10 20 30
Salinity (psu)
8 12 16 20
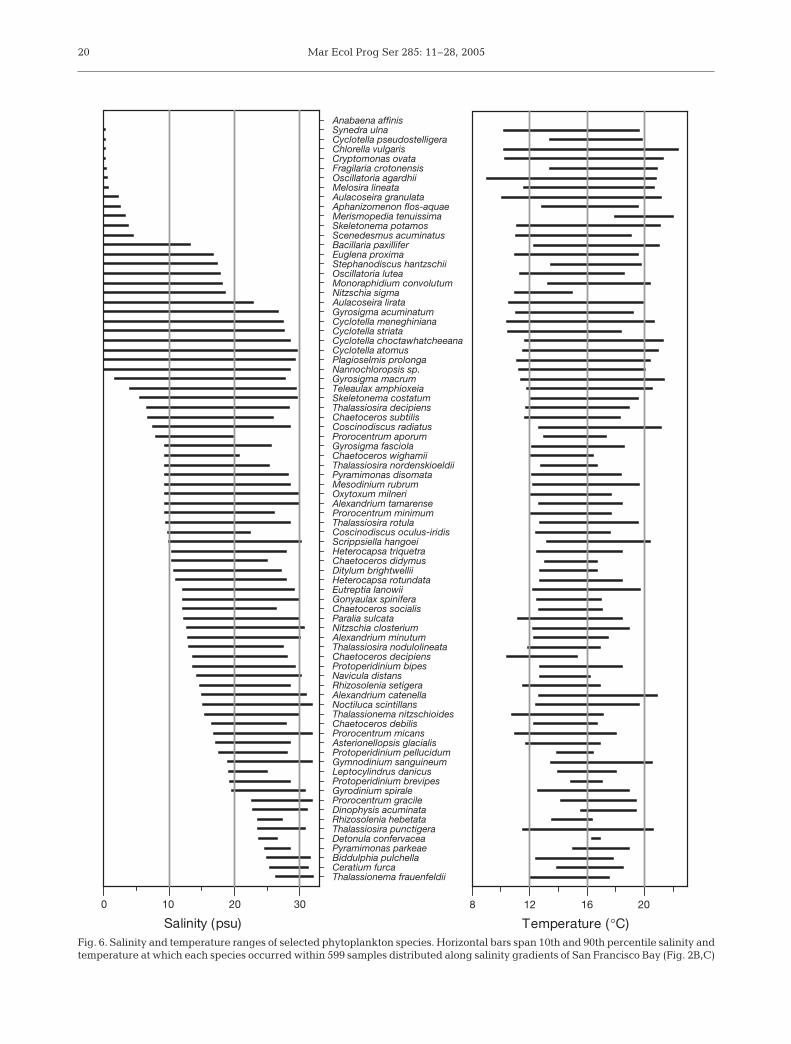
Temperature (°C)Fig. 6. Salinity and temperature ranges of selected phytoplankton species. Horizontal bars span 10th and 90th percentile salinity andtemperature at which each species occurred within 599 samples distributed along salinity gradients of San Francisco Bay (Fig. 2B,C)

Cloern & Dufford: Phytoplankton community ecology
Del Amo et al. 1997), Neuse River Estuary (dinoflagel-late dominance in winter: Mallin & Paerl 1994), Gulfof Naples (small diatoms in summer: Ribera d’Alcalàet al. 2004), Orbetello Lagoon (cryptophytes in winterand spring: Nuccio et al. 2003), or Norwegian fjords(coccolithophorids in summer: Smayda 1980). Diatomscontribute over 80% to phytoplankton biomass in SanFrancisco Bay, but dinoflagellates contribute over 85%to biomass in the Patapsco River estuary (Sellner etal. 2001), and biomass is evenly distributed amongdiatoms, dinoflagellates, cryptophytes, chlorophytes andcyanobacteria in the Neuse River Estuary (Pinckneyet al. 1998). Margalef’s Mandala suggests the (as yetuntested) hypothesis that this large community vari-ability between ecosystems is a consequence of differ-ences in their input of exogenous energy and the per-sistence of vertical structure in their pelagic habitats.
Margalef’s (1978) conceptual framework was builtfrom the principle that specialized life forms haveevolved as adaptations to physical habitat variability,but some species are clearly generalists. The pico-cyanobacteria and some small eukaryotes (Nanno-chloropsis sp., Teleaulax amphioxeia, Plagioselmisprolonga) are persistent and ubiquitous across largehabitat gradients in San Francisco Bay (Fig. 5D),implying resilience to variability in turbulent mixing,optical properties, salinity and temperature. Somemarine species (Thalassiosira frauenfeldii, Ceratiumfurca, Pyramimonas parkeae) are confined to near-coastal domains of San Francisco Bay (Fig. 6) and somefreshwater species (Anabaena affinis, Synedra ulna,Cryptomonas ovata) are confined to the tidal river, butmany others occur across broad salinity and tem-perature ranges (Fig. 6). These generalist species areapparently not constrained to occupying fixed niches,so their seasonal occurrences and timing of peak bio-mass can vary markedly from year to year (Karentz &Smayda 1984). One mode of resilience to environmen-tal variability is the occurrence within morphospeciesof genetically distinct strains having variable life histo-ries (French & Hargraves 1986, Montresor et al. 2003),growth rates (Rynearson & Armbrust 2000), and toler-ance to salinity and temperature variability (Krawiec1982).
(4) Pelagic communities are shaped by speciesinteractions across trophic levels: The mean doublingtime of phytoplankton in San Francisco Bay is onlyabout 4 d (Cloern et al. 1985) but, except duringblooms, biomass does not build up (Fig. 2E) becausemost phytoplankton primary production is consumedat a comparable rate. Many metazoan and protistanconsumers are selective in their herbivory, exerting atop–down control on phytoplankton community com-position. Based on this principle, Verity & Smetacek(1996) suggested that the traditional emphasis on
resource-driven community regulation has been self-limiting, and they proposed a revised conceptualframework ‘that predation and resource availabilityact through morphologies and life history strategiesof organisms to structure pelagic ecosystems’. Thecalanoid copepod Acartia sp. adapts its diet withselectivity in San Francisco Bay for large flagellatesand diatoms (Coscinodiscus spp., Thalassiosira spp.),the chain-forming diatom Skeletonema costatum, anddinoflagellates (Rollwagen-Bollens & Penry 2003).The microheterotrophs ingest small forms, with veryfast (up to 2 d–1) consumption of picocyanobacteria(Murrell & Hollibaugh 1998).
The phytoplankton have evolved a battery of taxon-specific chemical, morphological and behavioral de-fenses against predation. For example, some dino-flagellates (Gymnodinium catenatum, Dinophysis acu-minata, Alexandrium spp.) synthesize metabolites thatinhibit feeding and growth of copepods (Carlsson et al.1995, Teegarden 1999, Calbet et al. 2002). Some algaesynthesize multiple compounds which inhibit growthof bacteria, other phytoplankton, protozoan and meta-zoan grazers (Fistarol et al. 2003) to structure plank-tonic communities from both the top down and bottomup. At least 20 marine phytoplankton species are in-fected with host-specific viruses (Zingone et al. 1999),suggesting that pathogens play an important role inregulating phytoplankton biomass and species compo-sition (Short & Suttle 2003). We have just begun toexplore the population ecology of algal pathogens andchemically-based species interactions, but these forcesof community assembly are just as powerful as re-source competition and habitat selection (Zingone &Wyatt 2004). Many of these biological interactionswere unknown in Hutchinson’s era, so the 1961 con-ceptual framework was missing key processes nowviewed as central regulators of pelagic communitystructure.
(5) Phytoplankton species have mixed nutritionalmodes: Hutchinson (1961) and his contemporaries knewthe phytoplankton as obligate phototrophs competing fora finite number of resources. The presumption of strictphototrophy was shattered by discoveries (e.g. Bird &Kalff 1986, Estep et al. 1986) that some phototrophic fla-gellates ingest bacteria, and dinoflagellates feed onother algae with ‘zipper-like ‘mouths’ for swallowingfood, peduncles or feeding tubes for ‘sucking- up’ wholeprey or their cytoplasmic contents, and feeding veils forencasing and externally digesting large prey’ (Coats2002). Mixotrophic algae are key components of phyto-plankton communities including the 2 prominent non-diatom groups in San Francisco Bay: dinoflagellates(mixotrophic species of Dinophysis, Prorocentrum andAlexandrium) and cryptophytes (species of Plagioselmis,Teleaulax, Rhodomonas and Cryptomonas).
21

Mar Ecol Prog Ser 285: 11–28, 2005
The discovery of mixotrophy was important becauseit revealed that phytoplankton can tap the nutrientpools contained in microbial biomass, relaxing theconstraint of strict growth regulation by the supply ofinorganic nutrients. Bacterial grazing by algal flagel-lates is a mechanism for acquiring particulate organicP to sustain blooms even at growth-limiting concentra-tions of dissolved inorganic P (Nygaard & Tobiesen1993). Although we know very little about the ener-getics of mixotrophs, their capacity to acquire energyheterotrophically also provides a mechanism to survivein habitats such as turbid estuaries where photosyn-thesis is strongly light-limited. The ubiquity and per-sistence of cryptophytes (Fig. 5D) may reflect theirability to ingest bacteria as a heterotrophic supplementto light-limited phototrophy. The discovery of mixo-trophy revealed that phytoplankton species are not allfunctional equivalents occupying one trophic level.Even for the primary producers, species do not ‘clearlyaggregate into discrete, homogeneous trophic levels’(Polis & Strong 1996).
(6) Phytoplankton species have variable life histo-ries: Since Hutchinson’s era we have discovered thatmany algal species have different life forms, includingplanktonic vegetative and benthic or suspended rest-ing stages (e.g. Dale 2001). Life cycles were not explic-itly included in conceptual models of phytoplanktoncommunity ecology until Smetacek (1985) postulatedthat mass sedimentation of bloom diatoms is a life his-tory adaptation to nutrient depletion of surface waters,a survival strategy creating a stock of resting cells toseed population growth when seasonal mixing bringsnutrient-rich bottom waters back to the surface. Manyneritic species produce benthic resting stages, and thesediments of coastal ecosystems contain a reservoir ofdiatom spores or resting cells and dinoflagellate cyststhat provide a ready inoculum to seed blooms. Produc-tion of benthic stages is also a mechanism to retaina species’ genome within strongly advective systemssuch as estuaries, and this retentive function mayexplain why 92% of the biomass in San Francisco Baycomes from the 2 algal divisions (diatoms and dinofla-gellates) in which spore or cyst production is common.
The cues and periodicity of life-form alternation varyamong species, providing another mechanism of bio-geographic diversity beyond physical-habitat selec-tion. The dinoflagellate Alexandrium tamarense pro-duces cysts that require prolonged dormancy beforegermination, with a ‘univoltine’ life cycle of 1 annualalternation between pelagic vegetative and benthicresting stages (Montresor et al. 1998). Scrippsiellatrochoidea has a ‘multivoltine’ life strategy, producingresting stages that require short periods of dormancyand repeated cycles between life forms within a year.Annual cycles of excystment are regulated by endo-
genous rhythms (A. tamarense, Anderson & Keafer1987), or triggered by exogenous cues. Spore stages ofcommon diatoms in San Francisco Bay (Chaetocerossocialis, C. debilis, C. decipiens, Thalassiosira norden-skioeldii, Rhizosolenia setigera) germinate only whenthe photoperiod reaches 13 h (Eilertsen et al. 1995), amechanism of providing viable inocula to seed springblooms that is not related to water column stability ornutrient availability.
(7) Pelagic ecosystems are open: We now conceivethe pelagic as an open system in which immigrationand dispersal sustain community diversity. The phyto-plankton metapopulation of San Francisco Bay in-cludes all taxa in the linked river–estuary–coastalocean and pelagic–benthic systems (Table 1). Theopenness of aquatic ecosystems is central to the di-versity paradox because theoretical models show thatmany species can coexist when immigration is in-cluded as a process of community assembly (Hubbell2001). Many key phytoplankton species in San Fran-cisco Bay are the same taxa that develop blooms in theadjacent coastal upwelling system, suggesting thatphytoplankton diversity inside the estuary is influ-enced by exchanges with the coastal Pacific Ocean.Some dinoflagellates and flagellates (Ceratium spp.,Alexandrium catenella, Prorocentrum micans, P. gracile,Dinophysis acuminata, Heterosigma akashiwo) occuronly within the high-salinity zone of rapid oceanicexchange (Fig. 5F), suggesting these are allochthonousspecies produced in coastal waters and transportedinto the bay by surface currents or tidal dispersion.Allochthonous freshwater taxa, such as Skeletonemapotamos (Fig. 5E), are delivered to the estuary bypulses of river flow. Phytoplankton communities insidethe estuary are influenced by exchanges across a thirdinterface, between the sediments and overlying water.Many diatoms sampled in the plankton are benthic(Entomoneis paludosa, Gyrosigma balticum, G. fasci-ola, G. acuminatum, Pleurosigma strigosum, P. nor-manii, Nitzschia pusilla, N. [Cylindrotheca] closterium,N. sigma) or meroplanktonic forms (Thalassiosira de-cipiens, Cyclotella striata, C. meneghiniana, Paraliasulcata, Actinoptychus senarius) common in surficialsediments. Many freshwater taxa (Melosira varians,M. moniliformis, Bacillaria paxillifer, Achnanthes spp.)are common periphyton components in the tributaryrivers, so plankton species richness is augmented bysuspension of epipelic-epiphytic microalgae as an im-migration process that was recognized by Hutchinson(1961).
(8) Communities respond to large-scale climaticperiodicity: In Hutchinson’s era we were unaware oflarge-scale climatic fluctuations such as ENSO eventsor longer-period climatic cycles such as the PacificDecadal Oscillation (PDO) and their influence on
22

Cloern & Dufford: Phytoplankton community ecology
pelagic community dynamics. Our perception in thatera was that phytoplankton population dynamicsfollow recurrent patterns of species occurrence eachyear. Sustained observations and examination ofmicrofossils in the sedimentary record (e.g. Dale 2001)have since revealed that communities and seasonalpatterns change, sometimes dramatically, and thatthese shifts are coherent with annual variability andinterdecadal trends of climate change (McGowan etal. 1998). During our decade of observation in SanFrancisco Bay, some phytoplankton taxa (Prorocen-trum aporum, Coscinodiscus marginatus, Protoperi-dinium depressum, Eucampia zodiacus) apparentlydisappeared after 1996 (Fig. 5G) while others (Pro-toperidinum bipes, Pseudo-nitzschia delicatissima,Scrippsiella trochoidea, Thalassiosira nodulolineata)first appeared, and have occurred yearly, since1997–1998 (Fig. 5H). The general timing of thesechanges is coherent with a regime shift in the PDO andlarge-scale changes in ocean temperature, regionalwind patterns, and biological communities across thePacific basin (Chavez et al. 2003).
Manifestations of this shift included strengthening ofequatorward winds and surface currents, increasedabundances of cold-water species of copepods anddecreased abundances of warm-water species, andaltered distributions and abundance of euphausids inthe Eastern Pacific Boundary Current system after1997–1998 (Swartzman & Hickey 2003). Simultaneousspecies appearances and disappearances in San Fran-cisco Bay suggest that climate-driven changes incoastal plankton communities can propagate to inducecommunity change within estuaries. Estuarine com-munities evolve continuously, driven by climatic trends(Chiba & Saino 2002, McQuoid & Nordberg 2003,Zingone & Wyatt 2004), but also global species trans-locations (Hallegraeff & Bolch 1992) and disturbancessuch as eutrophication (Anderson et al. 2002).
Ecological significance of phytoplankton functionaldiversity
Phytoplankton photosynthesis is the primary energysupply to metazoan food webs of San Francisco Bay(Jassby et al. 1993, Sobczak et al. 2002) and other estu-aries. The efficiency of energy transfer from phyto-plankton to consumers and ultimate production atupper trophic levels vary with algal species composi-tion: diatom-dominated marine upwelling systems sus-tain 50 times more fish biomass per unit of phyto-plankton biomass than cyanobacteria-dominated lakes(Brett & Müller-Navarra 1997). The nutritional qualityof algae varies with its elemental composition andcontent of highly unsaturated fatty acids (HUFA), par-
ticularly eicosapentaenoic acid (EPA) and docosa-hexaenoic acid (DHA). Growth and fecundity of in-vertebrates are strongly correlated with the HUFAcontent of their food (Brett & Müller-Navarra 1997),and HUFA-rich diets are essential for development ofinvertebrate and vertebrate larvae (Olsen 1999). The17 top ranked species, contributing 89% to phyto-plankton biomass in San Francisco Bay, comprise 12diatoms, 2 dinoflagellates, 2 cryptophytes, and Meso-dinium rubrum. Marine dinoflagellates are rich inDHA, diatoms are rich in EPA (Olsen 1999), and cryp-tophytes have high contents of both (Ravet et al. 2003),so the phytoplankton in San Francisco Bay are of highnutritional quality. This suggests the hypothesis thattrophic efficiency leading to production at uppertrophic levels is also high. Estuaries are rearing habi-tats for shrimp, crabs, bivalves, demersal and pelagicfishes, and this nursery function includes algal supplyof both energy and essential biochemicals. The highnutritional quality of diatoms and dinoflagellates maycontribute to the generally higher efficiency of fishproduction in estuarine–marine regions compared tofreshwater lakes (Nixon 1988).
This traditional view of efficient diatom-based foodwebs has been challenged since discoveries that cope-pods often select other food items (Kleppel 1993) andhave lower fecundity when fed diatom versus non-diatom diets. This ‘diatom–copepod paradox’ (Ban etal. 1997) is the conflicting high nutritional qualityyet potential toxicity of diatoms to consumers. Somediatoms, including key species in San Francisco Bay(Thalassiosira rotula, Skeletonema costatum, Chaeto-ceros debilis, Ditylum brightwellii) synthesize alde-hydes that block mitosis and arrest development ofinvertebrate eggs (D’Ippolito et al. 2002). Althoughthere is uncertainty about the impairment of con-sumers in natural environments (Colin & Dam 2002),the prominence of these diatoms in San Francisco Baysuggests that secondary production might be disruptedby episodic blooms of key species that synthesizecytotoxic biochemicals.
Pathways and efficiencies of energy transfer withinpelagic food webs vary also with phytoplankton cellsize and nutritional mode. Size is important becausemany metazoan consumers, such as calanoid cope-pods, cannot capture small particles, including thenutritionally-rich nanoflagellates (Fenchel 1988). Webinned key phytoplankton species of San FranciscoBay into 2 size classes (using 15 µm as the criterionseparating small and large algae; Rollwagen Bollens& Penry 2003), and 3 nutritional modes (Fig. 7) distin-guishing strict phototrophs and heterotrophic dino-flagellates from mixotrophs capable of heterotrophicnutrition through osmotrophy (Dinophysis spp., Prym-nesiophytes) and ingestion of bacterial-sized particles
23

Mar Ecol Prog Ser 285: 11–28, 2005
(cryptophytes, Heterosigma spp., Prorocentrum spp.)or small phototrophs (Gymnodinium spp., Oxyrrhis spp.).
Taxa within each bin fulfill distinct functions, in-cluding a diverse array of feeding linkages thatcumulatively set the energy supply to metazoans. Weillustrate 5 trophic pathways (Fig. 7) leading fromphytoplankton primary production to production ofnorthern anchovy Engraulis mordax, the most abun-dant fish species in San Francisco Bay. Anchovy larvaefeed on large dinoflagellates including Gymnodinium(Akashiwo sanguineum), so they can function as pri-mary consumers (Trophic Level 2) when they utilizethis pathway exclusively (Fig. 7A). Other pathwayslead to anchovy consumption of mesozooplankton suchas copepods, and the most direct is through cope-pod predation on phototrophs, with selection for thepreferred species Mesodinium rubrum, flagellates,Skeletonema costatum, Thalassiosira spp., and Coscino-discus spp. (Rollwagen-Bollens & Penry 2003). Thispathway (Fig. 7B) places anchovies at Trophic Level 3,and it dominates during the spring bloom when large-cell primary production is high enough to support
maximum rates of copepod feeding and reproduction.Heterotrophic dinoflagellates such as Protoperidiniumspp. compete with copepods for large algal cells (Olli &Heiskanen 1999), and themselves are consumed bycopepods. The heterotrophic dinoflagellates introducean additional trophic linkage between phototrophsand mesozooplankton (Fig. 7C), moving anchoviesto Trophic Level 4. Although the energy contained insmall algal cells is not directly accessible to adult cope-pods, the mixotrophic algae function as intermediaryconsumers to repackage a fraction of this energy intoaccessible forms. The most direct pathway (Fig. 7D)links picocyanobacteria (Synechococcus spp.) to smallmixotrophs (Plagioselmis spp., Teleaulax spp.), thenlarger mixotrophs (Gymnodinium spp., Dinophysis spp.)and copepods, placing anchovies at Trophic Level 5.Longer pathways (Fig. 7E) to copepods, from bacteriato mixotrophs such as Prorocentrum spp. and then toheterotrophic dinoflagellates such as Polykrikos spp.and Noctiluca spp., place anchovies at Trophic Level 6.Assuming that the growth efficiency of protistans is0.25 and that of metazoans is 0.1, potential anchovy
24
Fig. 7. Phytoplankton classification by size (small cells <15 µm) and nutritional mode that influence pathways through which en-ergy and essential biochemicals are supplied to benthic metazoans (e.g. bivalve mollusks) and pelagic metazoans (e.g. copepodsand anchovies). Trophic Pathway A is an efficient direct link from large-cell phototrophs to larval fishes. Other pathways routeenergy through mesozooplankton: (B) directly from large-cell phototrophs, (C) from heterotrophic algae feeding on large-cellphototrophs, or (D) from mixotrophs or (E) microheterotrophs deriving energy fixed by small-cell phototrophs and routed through
the microbial loop. Some images redrawn from Tomas (1993, 1996)

Cloern & Dufford: Phytoplankton community ecology
production can range from 10% of primary productionwhen based solely on consumption of phototrophicdinoflagellates to only 0.016% of primary productionwhen based on the longest pathways (Fig. 7E). Neitherof these idealized pathways operates to the exclusionof others, but this comparison illustrates how phyto-plankton size and nutritional mode influence the eco-logical efficiency of food webs supporting pelagic fishproduction.
Phytoplankton size structure also influences energyallocation between the competing benthic and pelagicfood webs because of differences in size-selectivefeeding between benthic suspension feeders (e.g.bivalve mollusks) and crustacean zooplankton (e.g.calanoid copepods). Ciliary feeding provides bivalvesaccess to a broad size spectrum of food particles,including the bacterial-sized picoplankton and small-est eukaryotes (Wetz et al. 2002) that are inaccessibleto many pelagic crustaceans. Trophic linkages fromsmall-cell primary producers (species of Synecho-coccus, Nannochloropsis, Pyramimonas, Plagioselmis,Cyclotella) to benthic metazoans are direct and effi-cient because no intermediary steps of transformation(and associated respiratory loss) are required. Thestable biomass of picoplankton and small cryptophytesprovides a ubiquitous food resource to benthic inverte-brates, whereas the size-selective copepods are moredependent upon the episodic bursts of large-cell pro-duction during blooms. Copepods utilize trophic path-ways that are less efficient and less reliable than thoseof bivalves, and this distinction partly explains whybenthic metabolism often exceeds pelagic metabolismin shallow marine systems (Caffrey et al. 1998), benthicmacrofaunal biomass exceeds mesozooplankton bio-mass (Baretta et al. 1988), and alien bivalves success-fully compete with zooplankton for the phytoplanktonfood resource, leading to dramatic declines of zooplank-ton abundance following invasions by alien bivalves(Orsi & Mecum 1996).
Resolution of Hutchinson’s paradox and a remainingbig challenge
The plankton diversity problem was originally con-ceived as one of low dimensionality, whereby com-munities were presumably assembled by competitionoperating along a few resource gradients. Now weconceive the problem as hyperdimensional, wherebycommunities are assembled by selective forces operat-ing on variation in algal size, motility, behavior, lifecycles, biochemical specializations, nutritional mode,chemical and physiological tolerances, and dispersalprocesses. Diversity is promoted by variability alongeach of these dimensions. Four decades of discovery
and conceptual-model revisions have resulted in amore complete and mechanistic understanding of whyphytoplankton diversity exceeds that predicted fromequilibrium resource-competition theory.
Resolution of Hutchinson’s (1961) plankton diversityparadox has come from progress in understandingphytoplankton community assembly at the level ofform, function and growth habitat: small versus large,motile versus nonmotile, allochthonous versus auto-chthonous, benthic versus pelagic taxa. This has notyielded comparable advances in our knowledge ofcommunity assembly at the level of species. Marineecologists understand, at least generally, why diatomsdominate in turbulent nutrient-rich habitats, but notwhy Thalassiosira rotula is the Rank 1 species in SanFrancisco Bay or why toxic species are present but donot form harmful blooms in this estuary, as they do inothers. The rules of phytoplankton community assem-bly at the species level remain elusive, with 1 generalprinciple, that ‘the ultimate influences of species com-position are precedent and stochasticity‘ (Reynoldset al. 2000). The underlying processes will not be re-vealed until we fill in details of the matrix of key speciesand their repertoires of attributes on which the selec-tive processes of community assembly operate, includ-ing intraspecies genetic diversity (Zingone & Wyatt2004). We know surprisingly little about the life cycles,behavior, energetics, biochemical pathways, resourcerequirements, susceptibility to pathogens, genetic vari-ability and even growth rates of many key species suchas the ubiquitous and ecologically important crypto-phytes. Our knowledge base is therefore insufficientfor constructing reliable numerical models of phyto-plankton population dynamics at the species level, inspite of our recognition that the functions provided bythe phytoplankton vary among species (Fig. 7).
Resolution of the species assembly puzzle demandsinvestment in a mode of scientific investigation ground-ed in autecology, organism interactions and life historythat is perceived as unfundable or outmoded (Dale2001, Smetacek et al. 2002). Until this investment ismade, our capacity to understand the mechanisms ofphytoplankton species dynamics, their ecological andbiogeochemical significance, and their variable pat-terns between ecosystems will remain stifled (Verity &Smetacek 1996, Reynolds 1998). Given the imperativeof forecasting ecological responses across the fast-changing landscapes of the Earth system (Clark et al.2001), species-level studies are required to build thefoundation for constructing models of pelagic commu-nity responses under plausible scenarios of environ-mental change through nutrient enrichment, introduc-tions of alien species, hydrologic manipulations andglobal warming. Resolution of Hutchinson’s paradoxis a source of intellectual satisfaction derived from
25

Mar Ecol Prog Ser 285: 11–28, 2005
progress in identifying the processes that assemblecommunities at one level. The remaining big challengeis to understand how these processes interact to as-semble phytoplankton communities at the next, moreformidable, level of species.
Acknowledgements. We are indebted to our friends and col-leagues who have sustained a multi-decade program ofresearch and observation in San Francisco Bay: A. E. Alpine,B. E. Cole, S. W. Hager, T. S. Schraga, C. Burns Lopez, B.Richards, S. Conard, A. Arnsberg, J. Edmunds, S. Wienke,and J. Baylosis. A. Zingone and W. Sobczak provided insight-ful comments that greatly improved this paper. This projectwas supported by the US Geological Survey Toxic Sub-stance Hydrology Program and Place Based Program, and theRegional Monitoring Program for San Francisco Bay.
LITERATURE CITED
APHA (1989) Standard methods for the examination of waterand wastewater, 17th edn. American Public Health Asso-ciation, Washington, DC
Anderson DM, Keafer BA (1987) An endogenous annual clockin the toxic marine dinoflagellate Gonyaulax tamarensis.Nature 325:616–617
Anderson DM, Glibert PM, Burkholder JM (2002) Harmfulalgal blooms and eutrophication: nutrient sources, com-position and consequences. Estuaries 25:704–726
Azam F, Fenchel T, Field JG, Gray JS, Meyer-Reil LA,Thingstad F (1983) The ecological role of water-columnmicrobes in the sea. Mar Ecol Prog Ser 10:257–263
Ban S, Burns C, Castel J, Chaudron Y and 19 others (1997)The paradox of diatom–copepod interactions. Mar EcolProg Ser 157:287–293
Baretta J, Ruardiz P, de Wolf P (eds) (1988) Tidal flat estuaries.Simulation and analysis of the Ems Estuary. Springer-Verlag, Berlin
Bird DF, Kalff J (1986) Bacterial grazing by planktonic lakealgae. Science 231:493–495
Brett MT, Müller-Navarra DC (1997) The role of highly unsat-urated fatty acids in aquatic foodweb processes. FreshwBiol 38:483–499
Caffrey JM, Cloern JE, Grenz C (1998) Changes in produc-tion and respiration during a spring phytoplankton bloomin San Francisco Bay, California, USA: implications for netecosystem metabolism. Mar Ecol Prog Ser 172:1–12
Calbet A, Broglio E, Saiz E, Alcaraz M (2002) Low grazingimpact of mesozooplankton on the microbial communitiesof the Alboran Sea: a possible case of inhibitory effects bythe toxic dinoflagellate Gymnodinium catenatum. AquatMicrob Ecol 26:235–246
Carlsson P, Granéli E, Maestrini SY (1995) Copepod grazingon a phytoplankton community containing the toxicdinoflagellate Dinophysis acuminata. J Plankton Res 17:1925–1938
Cembella AD (2003) Chemical ecology of eukaryotic micro-algae in marine ecosystems. Phycologia 42:420–447
Chapra SC (1997) Surface water-quality modeling. McGraw-Hill, New York
Chavez FP, Ryan J, Lluch-Cota SE, Ñiquen MC (2003) Fromanchovies to sardines and back: multidecadal change inthe Pacific Ocean. Science 299:217–221
Chiba S, Saino T (2002) Interdecadal change in the upperwater column environment and spring diatom community
structure in the Japan Sea: an early summer hypothesis.Mar Ecol Prog Ser 231:23–35
Chisholm SW (1992) Phytoplankton size. In: Falkowski PG,Woodhead AD (eds) Primary productivity and biogeo-chemical cycles in the sea. Plenum Press, New York,p 213–237
Clark JS, Carpenter SR, Barber M, Collins S and 13 others(2001) Ecological forecasts: an emerging imperative.Science 293:657–660
Cloern JE (1996) Phytoplankton bloom dynamics in coastalecosystems: a review with some general lessons from sus-tained investigation of San Francisco Bay, California. RevGeophys 34:127–168
Cloern JE (1999) The relative importance of light and nutrientlimitation of phytoplankton growth:a simple index of coastalecosystem sensitivity to nutrient enrichment. Aquat Ecol33:3–15
Cloern JE, Cole BE, Wong RLJ, Alpine AE (1985) Temporaldynamics of estuarine phytoplankton: a case study of SanFrancisco Bay. Hydrobiologia 129:153–176
Cloern JE, Cole BE, Hager SW (1994) Notes on a Mesodiniumrubrum red tide in South San Francisco Bay (USA).J Plankton Res 16:1269–1276
Coats DW (2002) Dinoflagellate life-cycle complexities. J Phycol38:417–419
Cole BE, Cloern JE, Alpine AE (1986) Biomass and produc-tivity of three phytoplankton size classes in San FranciscoBay. Estuaries 9:117–126
Colin SP, Dam HG (2002) Testing for toxic effects of preyon zooplankton using sole versus mixed diets. LimnolOceanogr 47:1430–1437
Cullen JJ, Franks PS, Karl DM, Longhusrt A (2002) Physicalinfluences on marine ecosystem dynamics. In: RobinsonAR, McCarthy JJ, Rothschild BJ (eds) The sea, Vol 12.John Wiley & Sons, New York, p 297–336
Dale B (2001) The sedimentary record of dinoflagellate cysts:looking back into the future of phytoplankton blooms.Sci Mar 65 (Suppl. 2):257–272
Del Amo Y, Quéquiner B, Tréguer P, Breton H, Lampert L(1997) Impacts of high-nitrate freshwater inputs on macro-tidal ecosystems. II. Specific role of the silicic acid pump inthe year-round dominance of diatoms in the Bay of Brest(France). Mar Ecol Prog Ser 161:225–237
D’Ippolito G, Romano G, Iadicicco O, Miralto A, Ianora A,Cimino G, Fontana A (2002) New birth-control aldehydesfrom marine diatom Skeletonema costatum: characteriza-tion and biogenesis. Tetrahedron Lett 43:6133–6136
Duarte CM, Agustí S, Agawin NSR (2000) Response of aMediterranean phytoplankton community to increasednutrient inputs: a mesocosm experiment. Mar Ecol ProgSer 195:61–70
Dugdale RC, Goering JJ (1967) Uptake of new and regener-ated forms of nitrogen in primary productivity. LimnolOceanogr 12:196–206
Dugdale R, Wilkerson F (1992) Nutrient limitation of new pro-duction in the sea. In: Falkowski PG, Woodhead AD (eds)primary productivity and biogeochemical cycles in thesea. Plenum Press, New York, p 107–122
Eilertsen HC, Sandberg S, Tøllefsen H (1995) Photoperiodiccontrol of diatom spore growth: a theory to explain the onsetof phytoplankton blooms. Mar Ecol Prog Ser 116:303–307
Estep KW, Davis PG, Keller MD, Sieburth JMcN (1986) Howimportant are oceanic algal nanoflagellates in bacteri-vory? Limnol Oceanogr 31:646–649
Falkowski PG, Barber RT, Smetacek V (1998) Biogeochemicalcontrols and feedbacks on ocean primary production.Science 281:200–206
26

Cloern & Dufford: Phytoplankton community ecology
Fenchel T (1988) Marine plankton food chains. Annu Rev EcolSyst 19:19–38
Fistarol GO, Legrand C, Granéli E (2003) Allelopathic effectof Prymnesium parvum on a natural plankton community.Mar Ecol Prog Ser 255:115–125
French FW III, Hargraves PE (1986) Population dynamics ofthe spore-forming diatom Leptocylindricus danicus inNarragansett Bay, Rhode Island. J Phycol 22:411–420
Goldman JC, McGillicuddy DJ Jr (2003) Effect of largemarine diatoms growing at low light on episodic newproduction. Limnol Oceanogr 48:1176–1182
Hager SW, Schemel LE (1997) Dissolved nutrient data for theSan Francisco Bay estuary, California, January throughNovember 1995. Open-File Report 97-359, US GeologicalSurvey, Menlo Park, CA
Hallegraeff GM, Bolch CJ (1992) Transport of diatom anddinoflagellate resting spores in ships’ ballast water: im-plications for plankton biogeography and aquaculture.J Plankton Res 14:1067–1084
Hamm CE, Merkel R, Springer O, Jurkojc P, Maier C, PrechtelK, Smetacek V (2003) Architecture and material prop-erties of diatom shells provide effective mechanicalprotection. Nature 421:841–843
Harrison K (2000) Role of increased marine silica input onpaleo-pCO2 levels. Paleoceanography 15:292–298
Hillebrand H, Durselen C, Kirschtel D, Pollingher U, Zohary T(1999) Biovolume calculation for pelagic and benthicmicroalgae. J Phycol 35:403–424
Hobson LA (1988) Paradox of the phytoplankton—anoverview. Biol Oceanogr 6:493–504
Hubbell SP (2001) The unified neutral theory of biodiversityand biogeography. Princeton University Press, Princeton
Hutchinson GE (1961) The paradox of the plankton. Am Nat95:137–147
Jassby AD, Cloern JE, Powell TM (1993) Organic carbonsources and sinks in San Francisco Bay: variabilityinduced by river flow. Mar Ecol Prog Ser 95:39–54
Jassby AD, Cole BE, Cloern JE (1997) The design of samplingtransects for characterizing water quality in estuaries.Estuar Coast Shelf Sci 45:285–302
Karentz D, Smayda TJ (1984) Temperature and seasonal oc-currence patterns of 30 dominant phytoplankton speciesin Narragansett Bay over a 22-year period (1959–1980).Mar Ecol Prog Ser 18:277–293
Kiørboe T (1993) Turbulence, phytoplankton cell size, and thestructure of pelagic food webs. Adv Mar Biol 29:1–72
Kleppel GS (1993) On the diets of calanoid copepods. MarEcol Prog Ser 99:183–195
Krawiec RW (1982) Autecology and clonal variability of themarine centric diatom Thalassiosira rotula (Bacillario-phyceae) in response to light, temperature and salinity.Mar Biol 69:79–89
Lorenzen CJ (1967) Determination of chlorophyll and phaeo-pigments: spectrophotometric equations. Limnol Oceanogr12:343–346
Mallin MA, Paerl HW (1994) Planktonic trophic transfer in anestuary: seasonal, diel, and community structure effects.Ecology 75:2168–2184
Malone TC (1992) Effects of water column processes ondissolved oxygen, nutrients, phytoplankton and zoo-plankton. In: Smith DE, Leffler M, Mackiernan G (eds)Oxygen dynamics in the Chesapeake Bay. A synthesis ofrecent research. Maryland Sea Grant, College Park, MD,p 61–112
Margalef R (1978) Life-forms of phytoplankton as survivalalternatives in an unstable environment. Oceanol Acta1:493–509
May CL, Koseff JR, Lucas LV, Cloern JE, Schoellhamer DH(2003) Effects of spatial and temporal variability ofturbidity on phytoplankton blooms. Mar Ecol Prog Ser254:111–128
McGowan JA, DR Cayan, Dorman LM (1998) Climate–oceanvariability and ecosystem response in the northeastPacific. Science 281:210–217
McQuoid MR, Nordberg K (2003) Environmental influence onthe diatom and silicoflagellate assemblages in Koljö Fjord(Sweden) over the last two centuries. Estuaries 26:927–937
Montresor M, Zingone A, Sarno D (1998) Dinoflagellate cystproduction at a coastal Mediterranean site. J Plankton Res20:2291–2312
Montresor M, Sgrosso S, Procaccini G, Kooistra WHC (2003)Intraspecific diversity in Scrippsiella trochoidea (Dino-phyceae): evidence for cryptic species. Phycologia 42:56–70
Müller-Navarra DC, Brett MT, Park S, Chandra S, BallantyneAP, Zorita E, Goldman CR (2004) Unsaturated fatty acidcontent in seston and tropho-dynamic coupling in lakes.Nature 427:69–71
Murrell MC, Hollibaugh JT (1998) Microzooplankton grazingin northern San Francisco Bay measured by the dilutionmethod. Aquat Microb Ecol 15:53–63
Ning X, Cloern JE, Cole BE (2000) Spatial and temporalvariability of picocyanobacteria Synechococcus sp. in SanFrancisco Bay. Limnol Oceanogr 45:695–702
Nixon SW (1988) Physical energy inputs and the comparativeecology of lake and marine ecosystems. Limnol Oceanogr33:1005–1025
Nuccio C, Melillo C, Massi L, Innamorati M (2003) Phyto-plankton abundance, community structure and diversityin the eutrophicated Orbetello Lagoon (Tuscany) from1995 to 2001. Oceanol Acta 26:15–25
Nygaard K, Tobiesen A (1993) Bacterivory in algae: a survivalstrategy during nutrient limitation. Limnol Oceanogr 38:273–279
Olli K, Heiskanen AS (1999) Seasonal stages of phytoplank-ton community structure and sinking losses in the Gulf ofRiga. J Mar Syst 23:165–184
Olsen Y (1999) Lipids and essential fatty acids in aquatic foodwebs: what can freshwater ecologists learn from mari-culture? In: Arts MT, Wainman BC (eds) Lipids in fresh-water ecosystems. Springer-Verlag, New York, p 161–202
Orsi JJ, Mecum WL (1996) Food limitation as the probablecause of a long-term decline in the abundance of Neo-mysis mercedis the opossum shrimp in the Sacramento–San Joaquin Estuary. In: Hollibaugh JT (ed) San FranciscoBay: the ecosystem. Pacific Division, American Asso-ciation for the Advancement of Science, San Francisco,p 375–401
Pinckney JL, Paerl HW, Harrington MB, Howe KE (1998)Annual cycles of phytoplankton community-structureand bloom dynamics in the Neuse River Estuary, NorthCarolina. Mar Biol 131:371–381
Polis GA, Strong DR (1996) Food web complexity and com-munity dynamics. Am Nat 147:813–846
Pomeroy LR (1974) The ocean’s food web: a changing para-digm. BioScience 24:542–544
Ravet JL, Brett MT, Müller-Navarra DC (2003) A test of therole of polyunsaturated fatty acids in phytoplankton foodquality for Daphnia using liposome supplementation.Limnol Oceanogr 48:1938–1947
Reynolds CS (1998) What factors influence the species com-position of phytoplankton in lakes of different trophicstatus? Hydrobiologia 369/370:11–26
Reynolds CS, Dokulil M, Padisák J (2000) Understanding the
27

Mar Ecol Prog Ser 285: 11–28, 2005
assembly of phytoplankton in relation to the trophic spec-trum: where are we now? Hydrobiologia 424:147–152
Ribera d’Alcalà M, Conversano F, Corato F, Lincandro P and9 others (2004) Seasonal patterns in plankton communitiesin a pluriennal time series at a coastal Mediterranean site(Gulf of Naples): an attempt to discern recurrences andtrends. Sci Mar 68(Suppl. 1):65–83
Rollwagen Bollens GC, Penry DL (2003) Feeding dynamics ofAcartia spp. copepods in a large, temperate estuary (SanFrancisco Bay, CA). Mar Ecol Prog Ser 257:139–158
Rynearson TA, Armbrust EV (2000) DNA fingerprintingreveals extensive genetic diversity in a field population ofthe centric diatom Ditylum brightwellii. Limnol Oceanogr45:1329–1340
Sellner KG, Sellner SG, Lacouture RV, Magnien RE (2001)Excessive nutrients select for dinoflagellates in the strati-fied Patapsco River estuary: Margalef reigns. Mar EcolProg Ser 220:93–102
Short SM, Suttle CA (2003) Temporal dynamics of naturalcommunities of marine algal viruses and eukaryotes.Aquat Microb Ecol 32:107–119
Smayda TJ (1997) Harmful algal blooms: their ecophysiologyand general relevance to phytoplankton blooms in the sea.Limnol Oceanogr 42:1137–1153
Smayda TJ (1980) Phytoplankton species succession. In:Morris I (ed) The physiological ecology of phytoplankton.University of California Press, Berkeley, p 493–570
Smayda TJ, Reynolds CS (2001) Community assembly inmarine phytoplankton: application of recent models toharmful dinoflagellate blooms. J Plankton Res 23:447–461
Smetacek V (1985) Role of sinking in diatom life-history cycles:ecological, evolutionary and geological significance. MarBiol 84:239–251
Smetacek V (1995) Diatoms and the ocean carbon cycle.Protist 150:25–32
Smetacek V, Montresor M, Verity P (2002) Marine produc-tivity: footprints of the past and steps to the future. In:Williams PJ LeB, Thomas DN, Reyonolds CS (eds) Phyto-plankton productivity: carbon assimilation in marine andfreshwater ecosystems. Iowa State University Press, Ames,p 350–369
Sobczak WV, Cloern JE, Jassby AD, Müller-Solger AB (2002)Bioavailability of organic matter in a highly disturbed
estuary: the role of detrital and algal resources. Proc NatlAcad Sci USA 99:8101–8105
Swartzman G, Hickey B (2003) Evidence for a regime shiftafter the 1997–1998 El Niño, based on 1995, 1998, and2001 acoustic surveys in the Pacific Eastern BoundaryCurrent. Estuaries 26:1032–1043
Tamigneaux E, Vazquez E, Mingelbier M, Klein B, LegendreL (1995) Environmental control of phytoplankton assem-blages in nearshore marine waters, with special em-phasis on phototrophic ultraplankton. J Plankton Res 17:1421–1447
Teegarden GJ (1999) Copepod grazing selection and particlediscrimination on the basis of PSP toxin content. Mar EcolProg Ser 181:163–176
Tomas CR (ed) (1993) Marine phytoplankton. A guide to thenaked flagellates and coccolithophorids. Academic Press,San Diego
Tomas CR (ed) (1996) Identifying marine diatoms and dino-flagellates. Academic Press, San Diego
Utermöhl H (1958) Zur Vervollkommnung der quantitativenPhytoplankton-Methodik. Mitt Int Verh Theor AngewLimnol 9:1-38
Verity PG, Smetacek V (1996) Organism life cycles, preda-tion, and the structure of marine pelagic ecosystems. MarEcol Prog Ser 130:277–293
Waite AM, Thompson PA, Harrison PJ (1992) Does energycontrol the sinking rates of marine diatoms? LimnolOceanogr 37:468–477
Wetz MS, Lewitus AJ, Koepfler ET, Hayes KC (2002) Impactof the Eastern oyster Crassostrea virginica on micro-bial community structure in a salt marsh estuary. AquatMicrob Ecol 28:87–97
Wetzel RG, Likens GE (1991) Limnological analyses, 2nd edn.Springer-Verlag, New York
Zingone A, Wyatt T (2004) Harmful algal blooms: keys to theunderstanding of phytoplankton ecology. In: RobinsonAR, Brink KH (eds) The global coastal ocean: multi-scaleinterdisciplinary processes. The sea, Vol 13. HarvardUniversity Press, Cambridge, MA, p 867–926
Zingone A, Sarno D, Forlani G (1999) Seasonal dynamics inthe abundance of Micromonas pusilla (Prasinophyceae)and its viruses in the Gulf of Naples (Mediterranean Sea).J Plankton Res 21:2143–2159
28
Editorial responsibility: Otto Kinne (Editor-in-Chief), Oldendorf/Luhe, Germany
Submitted: May 25, 2004; Accepted: September 14, 2004Proofs received from author(s): January 3, 2005