physical and transcription map in the region 14q24.3: identification of six novel transcripts
TRANSCRIPT

GENOMICS 43, 130–140 (1997)ARTICLE NO. GE974786
Physical and Transcription Map in the Region 14q24.3:Identification of Six Novel Transcripts
Anne-Francoise Roux,*,1 Johanna M. Rommens,* Lynn Read,*,†Alessandra M. V. Duncan,‡ and Diane W. Cox*,†,2
*Research Institute, The Hospital for Sick Children, Toronto, Ontario M5G 1X8, and Department of Molecular Geneticsand Microbiology, University of Toronto, Toronto, Ontario M5S 1A1; †Department of Medical Genetics,
University of Alberta, Edmonton, Alberta T6G 2S2; and ‡Department of Pathology, Queen’s University,and Kingston General Hospital, Kingston, Ontario K7L 2V7, Canada
Received October 24, 1996; accepted April 25, 1997
mon approaches consist of exon trapping (Duyck etThe region of chromosome 14q24 has been of particu- al., 1990; Buckler et al., 1991) and cDNA selection
lar interest as it is known to contain one of the early- (Parimoo et al., 1991; Lovett et al., 1991; Rommens etonset Alzheimer disease genes (AD3). Other genes of al., 1994), used singly or in combination (Couch et al.,medical interest, such as arrhythmogenic right ven- 1996; Heiss et al., 1996).tricular cardiomyopathy, have been mapped to this re- The region we have studied is directly adjacent andgion by linkage analysis or chromosome rearrange- centromeric to the Alzheimer (AD3) region on 14q24.3.ments. We have focused on the region of a balanced This region has been partially described as a yeast arti-translocation (2;14)(p25;q24). Members of a family ficial chromosome (YAC) contig during the search forwith this translocation all have anterior polar cata-
the AD3 gene (Cruts et al., 1995; Clark et al., 1995).racts, suggesting the presence of a gene involved inYAC contig and sequence-tagged sites (STS) informa-lens integrity at the vicinity of the breakpoint. Thetion is also accessible from the public databaseschromosome 14 breakpoint has been defined betweenCEPH–Genethon (Chumakov et al., 1995) and the Hu-the short tandem repeats D14S289 and D14S277, a re-man Genome Center of the Whitehead Institute (Hud-gion of overlap for yeast artificial chromosomes (YACs)son et al., 1995). The region was initially chosen for888b2 and 934d4. We have extended the study of thea study of a family presenting an apparent balancedregion to 2 Mb on chromosome 14 and present a physi-
cal map of this region, including several sequence- translocation (2;14)(p25;q24) that has been inheritedtagged sites. New probes were generated using several through two generations in four individuals, cosegreg-end clones and inter-Alu PCR fragments from YACs. ating with an anterior polar cataract (Moross et al.,cDNA selection was used to identify transcribed se- 1984). Therefore, this translocation may be associatedquences. Mapping and alignment of 17 nonoverlapping with the alteration of a gene involved in early develop-cDNAs completed by sequence and expression pattern ment of the lens, leading to an anterior polar cataract.analysis suggested that a minimum of eight putative Anterior polar cataracts represent 3–14% of congeni-transcription units is present in this region: six of tal cataracts. Cataracts are heterogeneous in theirthese units correspond to five new genes and one mem-
transmission, in their location within the lens, and inber of a new gene family. q 1997 Academic Presstheir association with eye-related or other disorders.Several animal models have enabled identification ofmutations in various crystallin genes, the products ofINTRODUCTIONwhich represent 90% of the proteins of the lens (re-viewed in Hejtmancik et al., 1995). In humans severalFine physical mapping, together with the identifi-cataract loci have been identified, by linkage analysis,cation of transcription units, is the crucial step toon chromosomes 1, 2, 16, and 17 (Hejtmancik et al.,understanding the organization of the human ge-
nome. Several techniques have proven to be efficient 1995; Padma et al., 1995; Berry et al., 1996). Geneticfor retrieving transcribed sequences. The most com- defects resulting in different types of cataracts have
been described. These include mutations in the galac-tokinase gene (Stambolian et al., 1995), the activation1 Present address: Institut fur Humangenetik und Anthropologie,
Breisacher str. 33, D-79106 Freiburg, Germany. of gE-crystallin pseudogene (Brakenhoff et al., 1994),2 To whom correspondence should be addressed at Department of and some mutations in the PAX6 gene that also lead toMedical Genetics, 670 Heritage Medical Research Centre, University
aniridia and anophthalmia (Glaser et al., 1994). Genesof Alberta, Edmonton, Alberta T6G 2S2. Telephone: (403) 492-0874.Fax: (403) 492-1998. involved in anterior polar cataracts have not yet been
1300888-7543/97 $25.00Copyright q 1997 by Academic PressAll rights of reproduction in any form reserved.
AID GENO 4786 / 6r3a$$$$$1 06-18-97 18:25:01 gnma

MAP OF SIX NOVEL TRANSCRIPTS IN 14q24.3 131
Pulsed-field gel electrophoresis. Agar plugs containing high mo-identified in humans. Anterior polar cataracts are alsolecular weight DNA were rinsed in water for 1
2 h at 47C, transferredheterogeneous, and two of them that are associatedto 72 ml of digestion buffer with 1 mg of BSA and 20–30 U of restric-with specific translocations have been described: one tion enzymes, and incubated at the temperature recommended by
involving chromosomes 2 and 14 (Moross et al., 1984) the manufacturer for 4 h to overnight. Reactions were stopped byadding 3 ml of 500 mM EDTA, and plugs were stored at 47C untiland the second, chromosomes 3 and 18 (Rubin et al.,electrophoresis. Electrophoresis was performed in a CHEF DR II1994). One linkage study in a four-generation familyelectrophoresis system (Bio-Rad) using 1% agarose gels in 0.51 TBE.has identified a locus on 17p (Berry et al., 1996).Conditions used were according to the size range of separation re-We extended the study of the family with the (2;14) quired and were as follows when not specified otherwise: 200 V, 18 h,
translocation to identify the breakpoint and the poten- 5–50 s, and 167C. DNA was transferred onto Hybond N/ membrane(Amersham) using the manufacturer’s protocol.tially interrupted gene. We generated a physical and
The sizes of the PACs were estimated by PFGE following NotItranscription map of the region of chromosome 14 en-digestion (to release the insert). Conditions of electrophoresis werecompassing the breakpoint and spanning 2 Mb. We2–20 s, 200 V, and 20 h, 167C in 1% agarose gel.
have defined the translocation breakpoint between theProbes. PCR products from STSs and ESTs, and inserts gener-
short tandem repeats (STRs) D14S289 and D14S277 ated from inter-Alu PCR clones, end clones, and cDNA clones, werefrom an affected member of the family. We have gener- radiolabeled using a random priming kit (Pharmacia). STSs
D14S625 and D14S571 were preannealed with sheared human pla-ated new STSs and probes to build the fine physicalcental DNA before hybridization. Hybridizations were performed atmap and determined the correct order of some STSsleast overnight in Church–Gilbert buffer (Church and Gilbert, 1984)previously mapped. Using direct cDNA selection on at 657C. Washes were as follows: one rinse in 21 SSC at room temper-
YAC 888b2, we have retrieved cDNAs from the region ature, one wash in 21 SSC, 0.1% SDS at 587C for 20 min, and oneand identified eight putative transcription units. We wash in 0.21 SSC, 0.1% SDS at 587C for 20 min. After autoradiogra-
phy, filters were stripped and then were exposed before use.have strong evidence from sequence- and tissue-specificcDNA selection, construction, and screening of the mini-cDNA li-expression analysis that six of them correspond to five
brary. The procedure has been described (Rommens et al., 1994).new genes and one member of a new gene family.A pool of primary cDNA libraries of eight tissues, including liver,fetal brain, adult brain, skeletal muscle, retina, testis, lung, andkidney, and of the cell line Caco-2 was hybridized simultaneously.MATERIALS AND METHODSThe cDNAs retrieved from the selection were amplified by PCR andsubcloned using the TA cloning kit (Invitrogen).
Preparation of DNA. DNA from the patient with the transloca-Identification of PAC and cosmid clones. The PAC library, de-tion (EM1504) was prepared from a transformed lymphoblast cell
scribed elsewhere (Ioannou et al., 1994), was screened with sevenline. High molecular weight DNA used for pulsed-field gel electropho-clones. Southern blots containing EcoRI digests of the PACs wereresis (PFGE) was prepared as described (Scherer and Tsui, 1991).probed with the various landmarks (shown in Table 1 and in Fig. 3).PAC and cosmid DNAs were generated by Quiagen miniprep proto-
Human chromosome 14-specific cosmid library provided by thecols for routine Southern blot analysis.Human Genome Center (Los Alamos) was screened with 1-46, 1-
YAC and cosmid analysis. YAC clones were obtained from the 4, 1-58, 1-69, and 1-106. No cosmids could be retrieved with 1-4CEPH library and were chosen according to their STS content. The and 1-58.sizes of the YACs were obtained from CEPH or determined by PFGE.
Northern blot analysis. A human multiple tissue Northern blotInter-Alu PCR was performed on YACs 804g11, 924c7, 938e10, andfrom Clontech containing 2-mg samples of poly(A)/ RNA was succes-888b2 using the conditions and oligonucleotides described (Bull etsively probed with each cDNA. Hybridization and stripping of theal., 1993). The fragments were subcloned with the TA cloning kitblot was done as recommended by the manufacturer. Consecutive(Invitrogen). End clones were generated from both arms of YACswashes were performed with 21 SSC at room temperature, with 21804g11, 2h10, 972g10, 888b2, and 962d8 using HincII/RsaI or EcoRVSSC, 01% SDS at 587C for 15 min, and finally with 0.21 SSC, 0.1%digestion followed by inverse PCR and subcloning with the TA clon-SDS, at 587C for 15 min.ing kit (Triglia et al., 1988; Ochmann et al., 1988). End clones were
Sequencing. Automated fluorescence sequencing was performedgenerated from cosmids (1-69) 8 and 11 using inverse PCR as de-with an ABI 377 Prism sequencer. Sequences were subjected toscribed (Byth et al., 1994).alignment searches through GenBank and dbEST databases usingPublicly available STSs, STRs, and expressed-sequence tagsBLAST programs from the National Center for Biotechnology Infor-(ESTs) (accessible on the World Wide Web) were used to confirm themation (Altschul et al., 1990) and GRAIL1a (Uberbacher and Mural,YAC contig. PCR conditions for D14S289 and D14S258 have been1991) and were completed by using GCG sequence analysis softwaredescribed (Gyapay et al., 1994). D14S102E was a gift from M. Crutspackage.(Cruts et al., 1995). D14S571, D14S625, D14S858, and D14S841,
initially generated by the Whitehead Institute/MIT Center for Ge-nome Research (MIT), were obtained from Research Genetics. PCR
RESULTS AND DISCUSSIONconditions for STS 2h10-L (D14S1418) were 947C for 45 s, 587C for45 s, and 727C for 30 s for 35 cycles with a final MgCl2 concentrationof 1.5 mM. Generation of the Physical Map
In situ hybridization. Mapping of the YACs and PACs was per-Genomic Contigformed by fluorescence in situ hybridization (FISH) to normal human
lymphocyte and patient EM1504 lymphoblast chromosome spreadsThe YAC contig was generated with CEPH mega-counterstained with DAPI. The YAC and PAC probes were labeled
YACs obtained from the CEPH–Genethon and MITeither by nick-translation (Bionick Labeling System) with biotin-14–dATP or by oligolabeling (Boehringer Mannheim DIG DNA labeling databases. The initial landmarks used to select YACskit). Fluorescent hybridization signals were captured by a CCD cam- on the basis of reported STS content had been pre-era connected to a Cytovision image analysis system (Applied Im- viously shown to flank the translocation breakpoint inaging). Hybridization signals were merged using image analysis soft-
patient EM1504 by analysis of the two derivative chro-ware for pseudocolored blue (DAPI) and yellow (FITC) stains andoverlaid electronically. mosomes obtained by flow sorting (D. W. Cox et al.,
AID GENO 4786 / 6r3a$$$$$1 06-18-97 18:25:01 gnma
ROUX ET AL.132
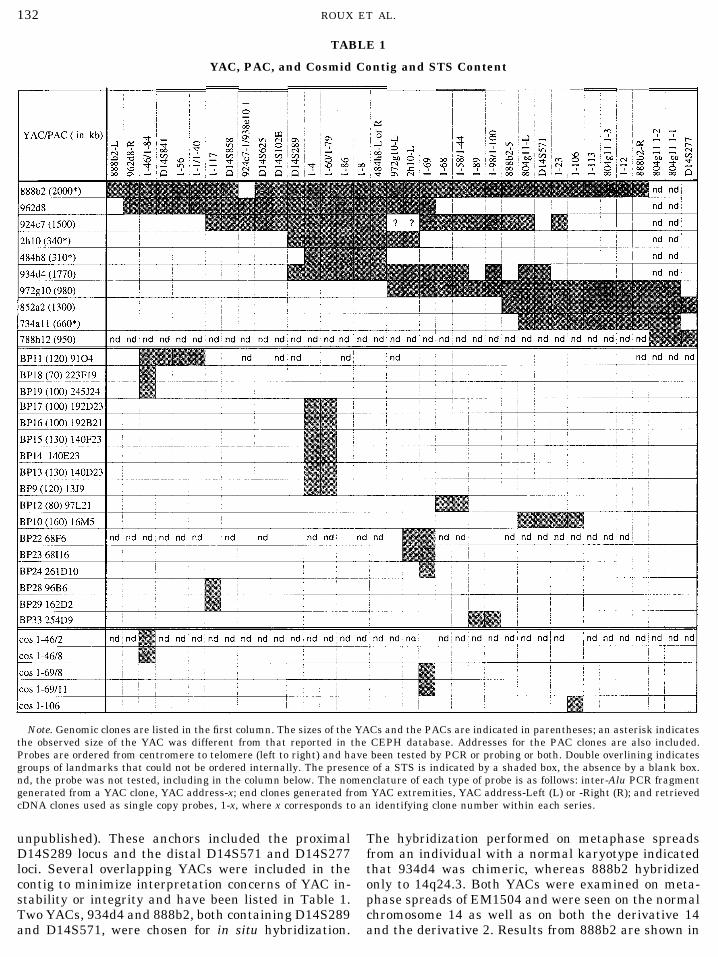
TABLE 1
YAC, PAC, and Cosmid Contig and STS Content
Note. Genomic clones are listed in the first column. The sizes of the YACs and the PACs are indicated in parentheses; an asterisk indicatesthe observed size of the YAC was different from that reported in the CEPH database. Addresses for the PAC clones are also included.Probes are ordered from centromere to telomere (left to right) and have been tested by PCR or probing or both. Double overlining indicatesgroups of landmarks that could not be ordered internally. The presence of a STS is indicated by a shaded box, the absence by a blank box.nd, the probe was not tested, including in the column below. The nomenclature of each type of probe is as follows: inter-Alu PCR fragmentgenerated from a YAC clone, YAC address-x; end clones generated from YAC extremities, YAC address-Left (L) or -Right (R); and retrievedcDNA clones used as single copy probes, 1-x, where x corresponds to an identifying clone number within each series.
unpublished). These anchors included the proximal The hybridization performed on metaphase spreadsfrom an individual with a normal karyotype indicatedD14S289 locus and the distal D14S571 and D14S277
loci. Several overlapping YACs were included in the that 934d4 was chimeric, whereas 888b2 hybridizedonly to 14q24.3. Both YACs were examined on meta-contig to minimize interpretation concerns of YAC in-
stability or integrity and have been listed in Table 1. phase spreads of EM1504 and were seen on the normalchromosome 14 as well as on both the derivative 14Two YACs, 934d4 and 888b2, both containing D14S289
and D14S571, were chosen for in situ hybridization. and the derivative 2. Results from 888b2 are shown in
AID GENO 4786 / 6r3a$$$$$1 06-18-97 18:25:01 gnma
MAP OF SIX NOVEL TRANSCRIPTS IN 14q24.3 133
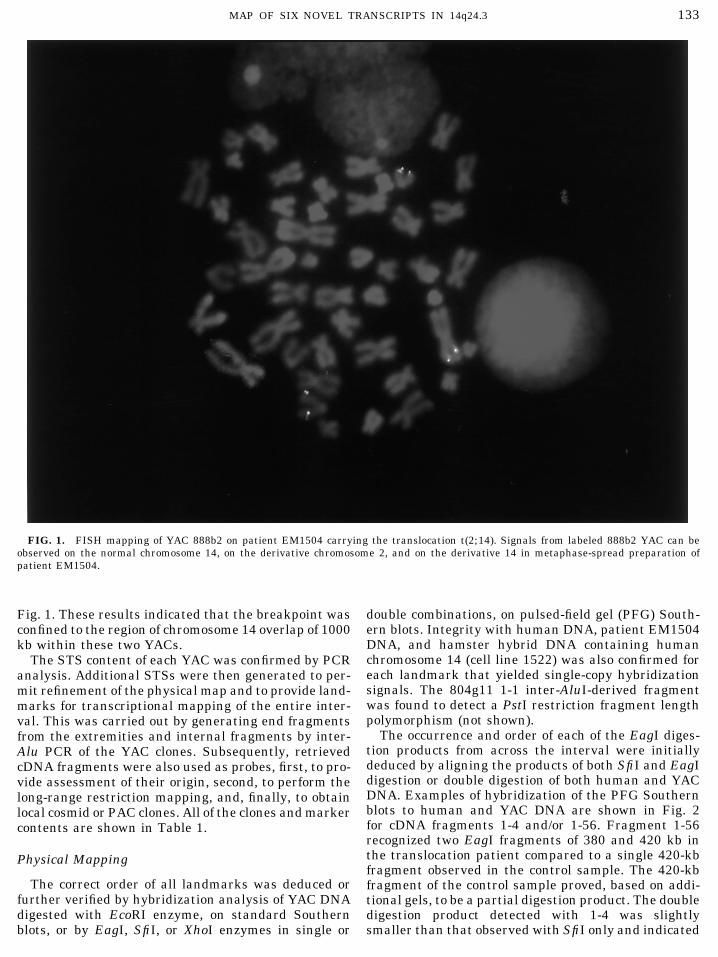
FIG. 1. FISH mapping of YAC 888b2 on patient EM1504 carrying the translocation t(2;14). Signals from labeled 888b2 YAC can beobserved on the normal chromosome 14, on the derivative chromosome 2, and on the derivative 14 in metaphase-spread preparation ofpatient EM1504.
Fig. 1. These results indicated that the breakpoint was double combinations, on pulsed-field gel (PFG) South-ern blots. Integrity with human DNA, patient EM1504confined to the region of chromosome 14 overlap of 1000DNA, and hamster hybrid DNA containing humankb within these two YACs.chromosome 14 (cell line 1522) was also confirmed forThe STS content of each YAC was confirmed by PCReach landmark that yielded single-copy hybridizationanalysis. Additional STSs were then generated to per-signals. The 804g11 1-1 inter-AluI-derived fragmentmit refinement of the physical map and to provide land-was found to detect a PstI restriction fragment lengthmarks for transcriptional mapping of the entire inter-polymorphism (not shown).val. This was carried out by generating end fragments
The occurrence and order of each of the EagI diges-from the extremities and internal fragments by inter-tion products from across the interval were initiallyAlu PCR of the YAC clones. Subsequently, retrieveddeduced by aligning the products of both SfiI and EagIcDNA fragments were also used as probes, first, to pro-digestion or double digestion of both human and YACvide assessment of their origin, second, to perform theDNA. Examples of hybridization of the PFG Southernlong-range restriction mapping, and, finally, to obtainblots to human and YAC DNA are shown in Fig. 2local cosmid or PAC clones. All of the clones and markerfor cDNA fragments 1-4 and/or 1-56. Fragment 1-56contents are shown in Table 1.recognized two EagI fragments of 380 and 420 kb inthe translocation patient compared to a single 420-kbPhysical Mappingfragment observed in the control sample. The 420-kb
The correct order of all landmarks was deduced or fragment of the control sample proved, based on addi-further verified by hybridization analysis of YAC DNA tional gels, to be a partial digestion product. The doubledigested with EcoRI enzyme, on standard Southern digestion product detected with 1-4 was slightly
smaller than that observed with SfiI only and indicatedblots, or by EagI, SfiI, or XhoI enzymes in single or
AID GENO 4786 / 6r3a$$$$$1 06-18-97 18:25:01 gnma
ROUX ET AL.134
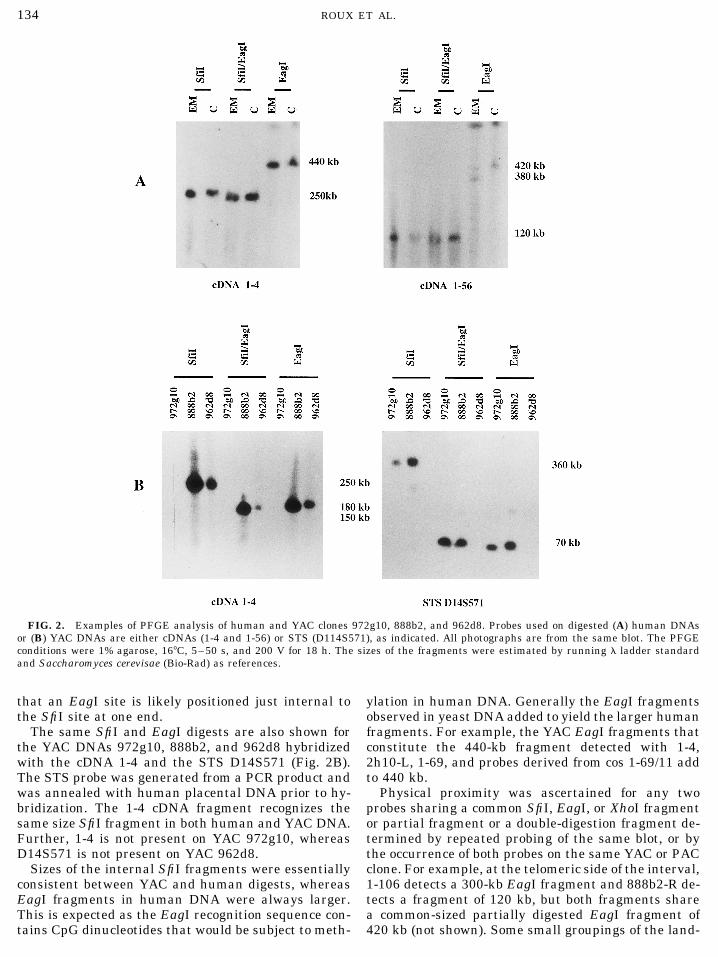
FIG. 2. Examples of PFGE analysis of human and YAC clones 972g10, 888b2, and 962d8. Probes used on digested (A) human DNAsor (B) YAC DNAs are either cDNAs (1-4 and 1-56) or STS (D114S571), as indicated. All photographs are from the same blot. The PFGEconditions were 1% agarose, 167C, 5–50 s, and 200 V for 18 h. The sizes of the fragments were estimated by running l ladder standardand Saccharomyces cerevisae (Bio-Rad) as references.
that an EagI site is likely positioned just internal to ylation in human DNA. Generally the EagI fragmentsobserved in yeast DNA added to yield the larger humanthe SfiI site at one end.
The same SfiI and EagI digests are also shown for fragments. For example, the YAC EagI fragments thatconstitute the 440-kb fragment detected with 1-4,the YAC DNAs 972g10, 888b2, and 962d8 hybridized
with the cDNA 1-4 and the STS D14S571 (Fig. 2B). 2h10-L, 1-69, and probes derived from cos 1-69/11 addto 440 kb.The STS probe was generated from a PCR product and
was annealed with human placental DNA prior to hy- Physical proximity was ascertained for any twoprobes sharing a common SfiI, EagI, or XhoI fragmentbridization. The 1-4 cDNA fragment recognizes the
same size SfiI fragment in both human and YAC DNA. or partial fragment or a double-digestion fragment de-termined by repeated probing of the same blot, or byFurther, 1-4 is not present on YAC 972g10, whereas
D14S571 is not present on YAC 962d8. the occurrence of both probes on the same YAC or PACclone. For example, at the telomeric side of the interval,Sizes of the internal SfiI fragments were essentially
consistent between YAC and human digests, whereas 1-106 detects a 300-kb EagI fragment and 888b2-R de-tects a fragment of 120 kb, but both fragments shareEagI fragments in human DNA were always larger.
This is expected as the EagI recognition sequence con- a common-sized partially digested EagI fragment of420 kb (not shown). Some small groupings of the land-tains CpG dinucleotides that would be subject to meth-
AID GENO 4786 / 6r3a$$$$$1 06-18-97 18:25:01 gnma

MAP OF SIX NOVEL TRANSCRIPTS IN 14q24.3 135
marks could not be ordered because of the limits of isolated with the 1-69 cDNA, detected the 440-kb EagIfragment and the 80-kb SfiI fragment in human DNA.resolution or because STRs did not provide suitable
probes even with extensive preannealing with human The 80-kb fragment was also seen in the DNA of theYACs, but in the patient EM an extra SfiI fragment ofplacental DNA. They are indicated in common inter-
vals as listed in Table 1. 160 kb was seen, suggesting that it was a junctionfragment. This was confirmed by FISH mapping; theThe YAC 888b2 was unstable, but included a very
large human fragment that was not chimeric as judged PAC clones BP11 and BP13 and the clones BP12 andBP10 are positioned centromeric and telomeric, respec-by any of our analyses, including in situ hybridization.
Propagation of individually grown colonies following tively, while the clone BP24 was found to contain thebreakpoint (data not shown).streaking on a minimal medium selection agar plate
revealed YACs of varying size upon ethidium bromide The comparison of results obtained from our physicalmap with those recently published by others (Cruts etstaining. The largest clones appeared to be greater
than 2.0 Mb. This was consistent with the size of the al., 1995) indicates some differences. First, in additionto the new probes we have generated, two additionalregion as measured from the EagI digestions of human
DNA and is markedly different from the 820 kb initially STSs, D14S841 and D14S858 (Hudson et al., 1995),have been included. The STS contents show discrepanc-reported by the CEPH–Genethon Database. The
largest 2.0-Mb YAC preparation was used for the selec- ies probably due to the instability of the megaYACs,as in our hands, YACs 924c7 and 888b2 do contain thetion experiments. The different-sized derivative 888b2
YACs also revealed a discrepancy with D14S625, which markers D14S102E and D14S625. The probe D14S102Ethat was previously described as an EST (Cruts et al.,was present on some, but absent on others, including
the large 2.0-Mb YAC. The deletion including D14S625 1995) that mapped to the interval was not accuratelyplaced on the long-range map as any attempt to use(and 924c7-1) is not large, since the adjacent SfiI and
EagI sites that were detected in other YACs and human the PCR product as a hybridization probe failed to yieldspecific signals. The ordering of the STSs on the mapDNA appear consistent with each other and with
888b2. Thus, we included D14S625 (and 924c7-1) in cen–D14S841–D14S858–D14S625–tel is in order op-posite to that previously reported (Hudson et al., 1995).the physical map.
The YAC end fragments could be precisely localized The confidence of the presented order is high, becausethe results from the mapping of the probes to EagI,in several cases. The end clone 2h10-L recognizes an
80-kb EagI fragment in 888b2 and 962d8, but the SfiI, and XhoI digestions together with the YAC cloneends support our interpretation.smaller EagI end fragment of 40 kb in 972g10. Also, 1-
23 detects 360-kb SfiI fragments with 852a2, 972g10, The distances between anchor STSs of the publicmaps vary from 140 kb (D14S841, D14S858) to at leastand 888b2, but detects ends of 250 and 150 kb of YACs
734a11 and 804g11, respectively (not shown). 1.1 Mb (D14S625, D14S571). A new STS was generatedbetween D14S625 and D14S571 from the left extremityAn estimate of the size of the region under study was
determined by adding the sizes of the EagI digestion of YAC 2h10. The STS 2h10-L (D14S1418) is Ç250 kbtelomeric of D14S625. The PCR product generated withproducts of human DNA detected with probes from
across the region, which indicate a total of 2.2 Mb. the oligonucleotides with sequences (1) 5*-TAT TTCTTG GTC CCT GGC TTG and (2) 5*-ATT TGA AGTDespite the number of probes, there were limitations
to the map at two positions; one of them is represented GCC TGC CTG AA (GDB: 3942317 and 3942615) is100 bp long. As the recently reported STS D14S1205by diagonal slashes (//) on Fig. 3. Fragments of common
size were not observed between D14S625 and any of with a PCR product size of 117 bp had been generatedfrom the same extremity of YAC 2h10, it is likely to bethe probes of the cluster around 1-4 or between the
cDNA fragments 1-69 and 1-58 in either YAC or human similar (Clark et al., 1995).DNA. To obtain the size estimate across these two posi-
Generation of the Transcription Maptions, EagI fragments of the human map were assumedto be adjacent. Additional internal fragments, if pres- Characterization of the Retrieved cDNA Fragmentsent, would be expected to be very small based on theoverall size and the summation of fragments in the cDNA selection was used to retrieve transcribed se-
quences of the isolated 2-Mb YAC 888b2. Of a total ofYAC clones. Rearrangements within the YACs are nota major concern as more than one YAC was used to 300 clones, 50% were found to contain repetitive se-
quences and were eliminated. Of the remaining clones,confirm the map. Although there is good correlationbetween the human map and the overall size of the 32 did not yield discrete signals on human or YAC DNA
or contained low-copy repeats. Clones were verified toYAC contig, there may be some additional EagI andSfiI sites on the YACs that were not detected with the originate from the region of interest by hybridization
to YAC DNA, to human DNA, and to human1 hamsterexisting probes (Fig. 3).The long-range mapping results revealed the posi- hybrid DNA containing only human chromosome 14.
The clones were also hybridized to each other to iden-tion of the translocation breakpoint to within 80 kb.The end fragment and the EcoRI restriction fragments tify overlapping fragments. A total of 17 nonoverlap-
ping clones were selected and positioned on the physi-of the chromosome 14-specific cosmid, cos11, which was
AID GENO 4786 / 6r3a$$$$$1 06-18-97 18:25:01 gnma
ROUX ET AL.136
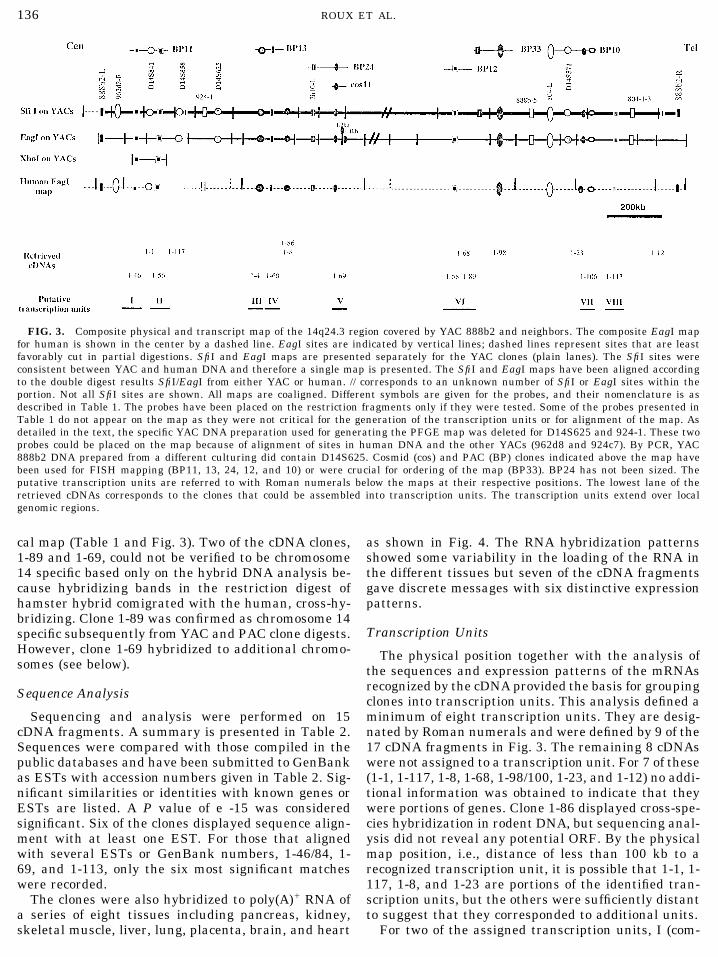
FIG. 3. Composite physical and transcript map of the 14q24.3 region covered by YAC 888b2 and neighbors. The composite EagI mapfor human is shown in the center by a dashed line. EagI sites are indicated by vertical lines; dashed lines represent sites that are leastfavorably cut in partial digestions. SfiI and EagI maps are presented separately for the YAC clones (plain lanes). The SfiI sites wereconsistent between YAC and human DNA and therefore a single map is presented. The SfiI and EagI maps have been aligned accordingto the double digest results SfiI/EagI from either YAC or human. // corresponds to an unknown number of SfiI or EagI sites within theportion. Not all SfiI sites are shown. All maps are coaligned. Different symbols are given for the probes, and their nomenclature is asdescribed in Table 1. The probes have been placed on the restriction fragments only if they were tested. Some of the probes presented inTable 1 do not appear on the map as they were not critical for the generation of the transcription units or for alignment of the map. Asdetailed in the text, the specific YAC DNA preparation used for generating the PFGE map was deleted for D14S625 and 924-1. These twoprobes could be placed on the map because of alignment of sites in human DNA and the other YACs (962d8 and 924c7). By PCR, YAC888b2 DNA prepared from a different culturing did contain D14S625. Cosmid (cos) and PAC (BP) clones indicated above the map havebeen used for FISH mapping (BP11, 13, 24, 12, and 10) or were crucial for ordering of the map (BP33). BP24 has not been sized. Theputative transcription units are referred to with Roman numerals below the maps at their respective positions. The lowest lane of theretrieved cDNAs corresponds to the clones that could be assembled into transcription units. The transcription units extend over localgenomic regions.
cal map (Table 1 and Fig. 3). Two of the cDNA clones, as shown in Fig. 4. The RNA hybridization patternsshowed some variability in the loading of the RNA in1-89 and 1-69, could not be verified to be chromosome
14 specific based only on the hybrid DNA analysis be- the different tissues but seven of the cDNA fragmentsgave discrete messages with six distinctive expressioncause hybridizing bands in the restriction digest of
hamster hybrid comigrated with the human, cross-hy- patterns.bridizing. Clone 1-89 was confirmed as chromosome 14
Transcription Unitsspecific subsequently from YAC and PAC clone digests.However, clone 1-69 hybridized to additional chromo- The physical position together with the analysis ofsomes (see below). the sequences and expression patterns of the mRNAs
recognized by the cDNA provided the basis for groupingSequence Analysis clones into transcription units. This analysis defined aminimum of eight transcription units. They are desig-Sequencing and analysis were performed on 15
cDNA fragments. A summary is presented in Table 2. nated by Roman numerals and were defined by 9 of the17 cDNA fragments in Fig. 3. The remaining 8 cDNAsSequences were compared with those compiled in the
public databases and have been submitted to GenBank were not assigned to a transcription unit. For 7 of these(1-1, 1-117, 1-8, 1-68, 1-98/100, 1-23, and 1-12) no addi-as ESTs with accession numbers given in Table 2. Sig-
nificant similarities or identities with known genes or tional information was obtained to indicate that theywere portions of genes. Clone 1-86 displayed cross-spe-ESTs are listed. A P value of e -15 was considered
significant. Six of the clones displayed sequence align- cies hybridization in rodent DNA, but sequencing anal-ysis did not reveal any potential ORF. By the physicalment with at least one EST. For those that aligned
with several ESTs or GenBank numbers, 1-46/84, 1- map position, i.e., distance of less than 100 kb to arecognized transcription unit, it is possible that 1-1, 1-69, and 1-113, only the six most significant matches
were recorded. 117, 1-8, and 1-23 are portions of the identified tran-scription units, but the others were sufficiently distantThe clones were also hybridized to poly(A)/ RNA of
a series of eight tissues including pancreas, kidney, to suggest that they corresponded to additional units.For two of the assigned transcription units, I (com-skeletal muscle, liver, lung, placenta, brain, and heart
AID GENO 4786 / 6r3a$$$$$1 06-18-97 18:25:01 gnma
MAP OF SIX NOVEL TRANSCRIPTS IN 14q24.3 137
TABLE 2
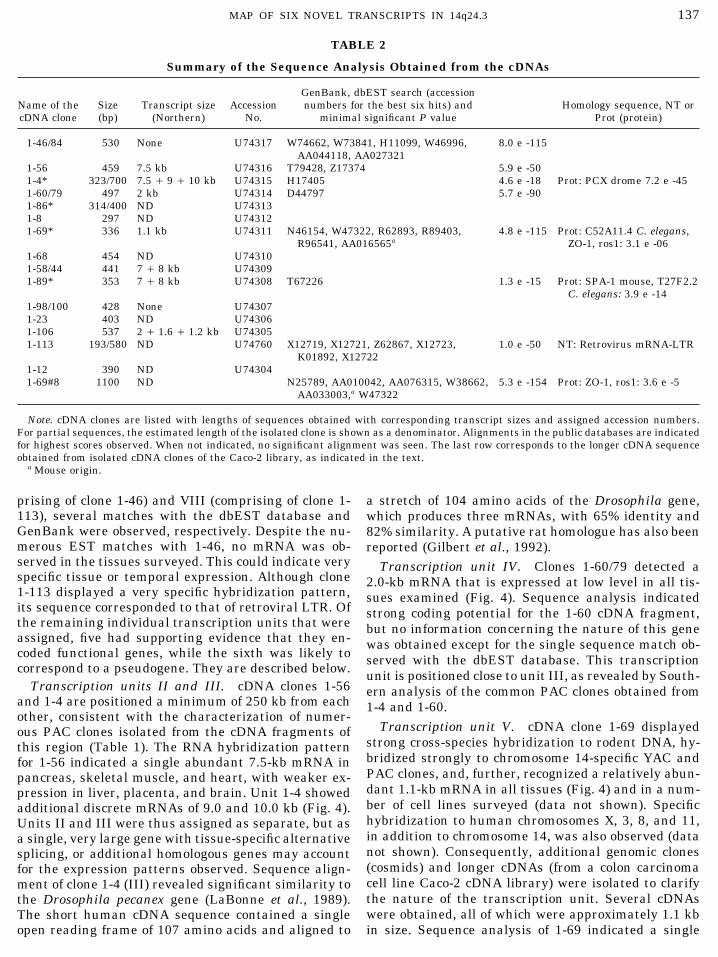
Summary of the Sequence Analysis Obtained from the cDNAs
GenBank, dbEST search (accessionName of the Size Transcript size Accession numbers for the best six hits) and Homology sequence, NT orcDNA clone (bp) (Northern) No. minimal significant P value Prot (protein)
1-46/84 530 None U74317 W74662, W73841, H11099, W46996, 8.0 e -115AA044118, AA027321
1-56 459 7.5 kb U74316 T79428, Z17374 5.9 e -501-4* 323/700 7.5 / 9 / 10 kb U74315 H17405 4.6 e -18 Prot: PCX drome 7.2 e -451-60/79 497 2 kb U74314 D44797 5.7 e -901-86* 314/400 ND U743131-8 297 ND U743121-69* 336 1.1 kb U74311 N46154, W47322, R62893, R89403, 4.8 e -115 Prot: C52A11.4 C. elegans,
R96541, AA016565a ZO-1, ros1: 3.1 e -061-68 454 ND U743101-58/44 441 7 / 8 kb U743091-89* 353 7 / 8 kb U74308 T67226 1.3 e -15 Prot: SPA-1 mouse, T27F2.2
C. elegans: 3.9 e -141-98/100 428 None U743071-23 403 ND U743061-106 537 2 / 1.6 / 1.2 kb U743051-113 193/580 ND U74760 X12719, X12721, Z62867, X12723, 1.0 e -50 NT: Retrovirus mRNA-LTR
K01892, X127221-12 390 ND U743041-69#8 1100 ND N25789, AA010042, AA076315, W38662, 5.3 e -154 Prot: ZO-1, ros1: 3.6 e -5
AA033003,a W47322
Note. cDNA clones are listed with lengths of sequences obtained with corresponding transcript sizes and assigned accession numbers.For partial sequences, the estimated length of the isolated clone is shown as a denominator. Alignments in the public databases are indicatedfor highest scores observed. When not indicated, no significant alignment was seen. The last row corresponds to the longer cDNA sequenceobtained from isolated cDNA clones of the Caco-2 library, as indicated in the text.
a Mouse origin.
prising of clone 1-46) and VIII (comprising of clone 1- a stretch of 104 amino acids of the Drosophila gene,113), several matches with the dbEST database and which produces three mRNAs, with 65% identity andGenBank were observed, respectively. Despite the nu- 82% similarity. A putative rat homologue has also beenmerous EST matches with 1-46, no mRNA was ob- reported (Gilbert et al., 1992).served in the tissues surveyed. This could indicate very Transcription unit IV. Clones 1-60/79 detected aspecific tissue or temporal expression. Although clone 2.0-kb mRNA that is expressed at low level in all tis-1-113 displayed a very specific hybridization pattern, sues examined (Fig. 4). Sequence analysis indicatedits sequence corresponded to that of retroviral LTR. Of strong coding potential for the 1-60 cDNA fragment,the remaining individual transcription units that were but no information concerning the nature of this geneassigned, five had supporting evidence that they en- was obtained except for the single sequence match ob-coded functional genes, while the sixth was likely to served with the dbEST database. This transcriptioncorrespond to a pseudogene. They are described below. unit is positioned close to unit III, as revealed by South-
Transcription units II and III. cDNA clones 1-56 ern analysis of the common PAC clones obtained fromand 1-4 are positioned a minimum of 250 kb from each 1-4 and 1-60.other, consistent with the characterization of numer-
Transcription unit V. cDNA clone 1-69 displayedous PAC clones isolated from the cDNA fragments ofstrong cross-species hybridization to rodent DNA, hy-this region (Table 1). The RNA hybridization patternbridized strongly to chromosome 14-specific YAC andfor 1-56 indicated a single abundant 7.5-kb mRNA inPAC clones, and, further, recognized a relatively abun-pancreas, skeletal muscle, and heart, with weaker ex-dant 1.1-kb mRNA in all tissues (Fig. 4) and in a num-pression in liver, placenta, and brain. Unit 1-4 showedber of cell lines surveyed (data not shown). Specificadditional discrete mRNAs of 9.0 and 10.0 kb (Fig. 4).hybridization to human chromosomes X, 3, 8, and 11,Units II and III were thus assigned as separate, but asin addition to chromosome 14, was also observed (dataa single, very large gene with tissue-specific alternativenot shown). Consequently, additional genomic clonessplicing, or additional homologous genes may account(cosmids) and longer cDNAs (from a colon carcinomafor the expression patterns observed. Sequence align-cell line Caco-2 cDNA library) were isolated to clarifyment of clone 1-4 (III) revealed significant similarity tothe nature of the transcription unit. Several cDNAsthe Drosophila pecanex gene (LaBonne et al., 1989).were obtained, all of which were approximately 1.1 kbThe short human cDNA sequence contained a single
open reading frame of 107 amino acids and aligned to in size. Sequence analysis of 1-69 indicated a single
AID GENO 4786 / 6r3a$$$$$1 06-18-97 18:25:01 gnma
ROUX ET AL.138
ple, the ZO-1 gene produces a 7.8-kb mRNA and en-codes a large multidomain protein that is a componentof tight junctions. Tight junctions form the intercellularbarrier between epithelial cells, separate tissue com-partments, and maintain cellular polarity (Willott etal., 1993). The small size of the mRNA precludes theisolated cDNAs as corresponding to a ZO-1 homologue,but the shared domain, whose function is not yetknown, is clearly present. Both cosmid clones cos8 andcos11, isolated with 1-69, were confirmed by end-frag-ment analysis to originate from chromosome 14. Thesequence corresponding to the ORF region of cos11 re-vealed at least one insertion and one deletion leadingto stop codons, indicating that the copy of this sequencefrom chromosome 14 was likely to correspond to a pseu-dogene.
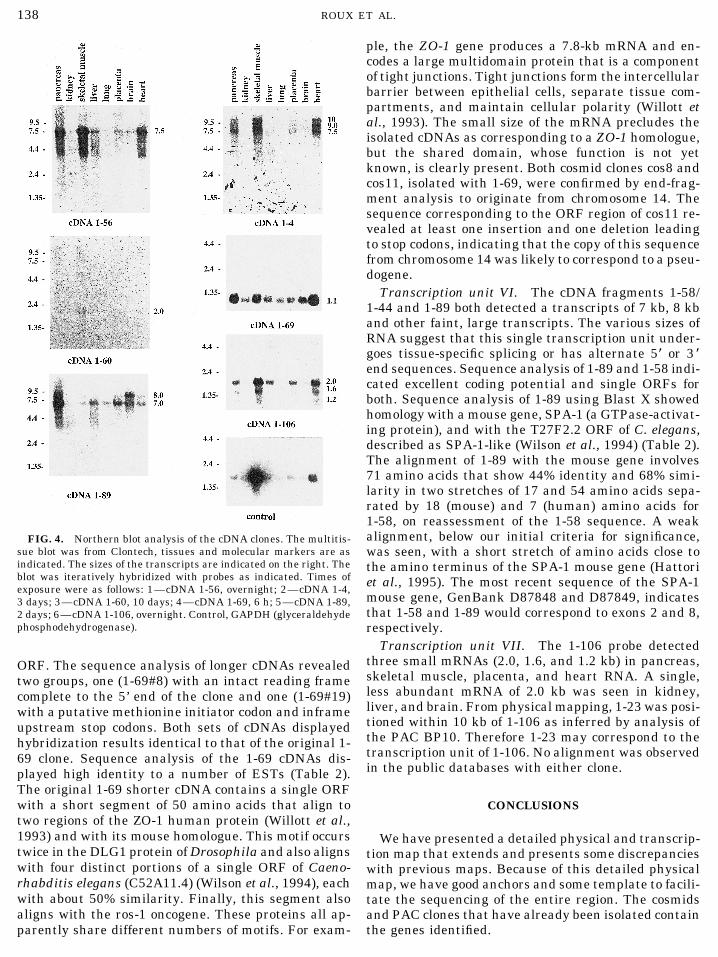
Transcription unit VI. The cDNA fragments 1-58/1-44 and 1-89 both detected a transcripts of 7 kb, 8 kband other faint, large transcripts. The various sizes ofRNA suggest that this single transcription unit under-goes tissue-specific splicing or has alternate 5* or 3 *end sequences. Sequence analysis of 1-89 and 1-58 indi-cated excellent coding potential and single ORFs forboth. Sequence analysis of 1-89 using Blast X showedhomology with a mouse gene, SPA-1 (a GTPase-activat-ing protein), and with the T27F2.2 ORF of C. elegans,described as SPA-1-like (Wilson et al., 1994) (Table 2).The alignment of 1-89 with the mouse gene involves71 amino acids that show 44% identity and 68% simi-larity in two stretches of 17 and 54 amino acids sepa-rated by 18 (mouse) and 7 (human) amino acids for1-58, on reassessment of the 1-58 sequence. A weakalignment, below our initial criteria for significance,FIG. 4. Northern blot analysis of the cDNA clones. The multitis-
sue blot was from Clontech, tissues and molecular markers are as was seen, with a short stretch of amino acids close toindicated. The sizes of the transcripts are indicated on the right. The the amino terminus of the SPA-1 mouse gene (Hattoriblot was iteratively hybridized with probes as indicated. Times of et al., 1995). The most recent sequence of the SPA-1exposure were as follows: 1—cDNA 1-56, overnight; 2—cDNA 1-4,
mouse gene, GenBank D87848 and D87849, indicates3 days; 3—cDNA 1-60, 10 days; 4—cDNA 1-69, 6 h; 5—cDNA 1-89,that 1-58 and 1-89 would correspond to exons 2 and 8,2 days; 6—cDNA 1-106, overnight. Control, GAPDH (glyceraldehyde
phosphodehydrogenase). respectively.Transcription unit VII. The 1-106 probe detected
three small mRNAs (2.0, 1.6, and 1.2 kb) in pancreas,ORF. The sequence analysis of longer cDNAs revealedskeletal muscle, placenta, and heart RNA. A single,two groups, one (1-69#8) with an intact reading frameless abundant mRNA of 2.0 kb was seen in kidney,complete to the 5’ end of the clone and one (1-69#19)liver, and brain. From physical mapping, 1-23 was posi-with a putative methionine initiator codon and inframetioned within 10 kb of 1-106 as inferred by analysis ofupstream stop codons. Both sets of cDNAs displayedthe PAC BP10. Therefore 1-23 may correspond to thehybridization results identical to that of the original 1-transcription unit of 1-106. No alignment was observed69 clone. Sequence analysis of the 1-69 cDNAs dis-in the public databases with either clone.played high identity to a number of ESTs (Table 2).
The original 1-69 shorter cDNA contains a single ORFwith a short segment of 50 amino acids that align to CONCLUSIONStwo regions of the ZO-1 human protein (Willott et al.,1993) and with its mouse homologue. This motif occurs We have presented a detailed physical and transcrip-twice in the DLG1 protein of Drosophila and also aligns tion map that extends and presents some discrepancieswith four distinct portions of a single ORF of Caeno- with previous maps. Because of this detailed physicalrhabditis elegans (C52A11.4) (Wilson et al., 1994), each map, we have good anchors and some template to facili-with about 50% similarity. Finally, this segment also tate the sequencing of the entire region. The cosmidsaligns with the ros-1 oncogene. These proteins all ap- and PAC clones that have already been isolated contain
the genes identified.parently share different numbers of motifs. For exam-
AID GENO 4786 / 6r3a$$$$$1 06-18-97 18:25:01 gnma

MAP OF SIX NOVEL TRANSCRIPTS IN 14q24.3 139
S. S., and Shiels, A. (1996). A locus for autosomal dominant ante-FISH analysis initially indicated the position of therior polar cataract on chromosome 17p. Hum. Mol. Genet. 5: 415–translocation breakpoint in patient EM1504. It is also419.likely that the extra SfiI fragment of 160 kb present in
Brakenhoff, R. H., Henskens, H. A. M., van Rossum, M. W. P. C.,the patient and recognized by cDNA 1-69 corresponds Lubsen, N. H., and Schoenmakers, J. G. G. (1994). Activation ofto the junction fragment. From 10 different enzyme the gE-crystallin pseudogene in the human hereditary Coppock-
like cataract. Hum. Mol. Genet. 3: 279–283.digestions on a hybridization blot, no junction fragmentcould be found using 1-69, cos 11-L, 1.1kb-Eco, or 1.2kb- Buckler, A. J., Chang, S. L., Graw, S. L., Brook, J. D., Haber, D. A.,
Sharp, P. A., and Housman, D. E. (1991). Exon amplification: AEco as probes. This suggests we have not localized astrategy to isolate mammalian genes based on RNA splicing. Proc.gene at the breakpoint or that the crucial gene occursNatl. Acad. Sci. USA 88: 4005–4009.on chromosome 2. Although the cDNA selection was
Bull, P. C., Barwell, J. A., Hannah, H. T.-L., Paulter, S. E., Higgins,carried out with a series of tissues (including retina M. J., Lalande, M., and Cox, D. W. (1993). Isolation of new probesand fetal brain), not all genes may have been retrieved in the region of the Wilson disease locus, 13q14.2–q14.3. Cyto-or insufficient numbers of clones may have been ana- genet. Cell Genet. 64: 12–17.lyzed. It therefore appears necessary to clone the Byth, B. C., Thomas, G. R., Hofland, N., and Cox, D. W. (1994). Appli-
cation of inverse PCR to isolation of end probes from cosmids.breakpoint and sequence the corresponding regionsNucleic Acids Res. 22: 1766–1767.from chromosomes 2 and 14. The anterior polar cata-
Chumakov, I. L., Rigault, P., Le Gal, I., Bellane-Chantelot, C., Bil-ract is possibly not involved in the translocation, andlault, A., Guillou, S., Soularue, P., Guasconi, G., Poullier, E., Gros,the association observed in the affected family could beI., Belova, M., Sanbucy, J.-L., Susini, L., Gervy, P., Glibert, F.,
spurious (1 chance in 16). One other possibility is that Deaufils, S., Bui, H., Massart, C., De Tand, M.-F., Dukasz, F.,the affected gene does not occur at the breakpoint and Lecoulant, S., Ougen, P., Perrot, V., Bertrand, S., Codani, J.-J.,
Caterina, D., Georges, I., Lacroix, B., Lucotte, G., Sahbatou, M.,altered longer range cis-acting factors affect either itsSchmit, C., Sangouard, M., Tubacher, E., Dib, C., Faure, S., Fi-regulation or transcription as a result of a change ofzames, C., Gyapay, G., Millasseau, P., Nguyen, S., Muselet, D.,state of the local chromatin. Therefore, any gene sur- Vignal, A., Morissette, J., Menninger, J., Lieman, J., Desai, T.,
rounding cDNA 1-69 could be considered a potential Banks, A., Bray-Ward, P., Ward, D., Hudson, T., Gerety, S., Foote,candidate (such as 1-60/1-79 and 1-58/1-89). Further S., Stein, L., Page, D. C., Lander, E. S., Weissenbach, J., Le Paslier,
D., and Cohen, D. (1995). A YAC contig map of the human genome.experiments will address this possibility and study theNature 377: 155–297.developmental expression of these genes in lens.
Church, G. M., and Gilbert, W. (1984). Genomic sequencing. Proc.Several transcription units remain tentative asNatl. Acad. Sci. USA 81: 1991–1995.mRNA could not be detected with the Northern blots.
Clark, R. F., Cruts, M., Korenblat, K. M., He, C., Talbot, C., VanThis may be due to low or no expression in the tissues orBroeckoven, C., and Goate, A. M. (1995). A yeast artificial chromo-
in the stages of their development examined. Possibly, some contig from human chromosome 14q24 spanning the Alzhei-these cDNAs are intronic and do not correspond to ma- mer’s disease locus AD3. Hum. Mol. Genet. 4: 1347–1354.ture mRNA (Couch et al., 1996). Arrhythmogenic right Couch, F. J., Rommens, J. M., Neuhausen, S. L., Belanger, C., Du-
mont, M., Abel, K., Bell, R., Berry, S., Bogden, R., Cannon-Albright,ventricular cardiomopathy has been localized in theL., Farid, L., Frye, C., Hattier, T., Janecki, T., Jiang, P., Kherer,14q23–q24 region (Rampazzo et al., 1994). TranscriptR., LeBlanc, J.-F., McArthur-Morrison, J., McSweeney, D., Miki,units II and III, expressed in heart, might be consid-Y., Peng, Y., Samson, C., Schoeder, M., Snyder, S. C., Stringfellow,
ered candidate genes for this disease. M., Stroup, C., Swedlund, B., Swensen, J., Teng, D., Thakur, S.,No genes were localized to this region before the pres- Tran, T., Tranchant, M., Welver-Feldaus, J., Wong, A. K. C., Shik-
zuya, H., Labrie, F., Skolnick, M. H., Goldgar, D. E., Kamb, A.,ent study. We suggest that all the putative genes iden-Weber, B. L., Tavtigian, S. V., and Simard, J. (1996). Generationtified correspond to new sequences and at least one ofof an integrated transcription map of the BRCA2 region on chromo-them appears to be a new gene family member. some 13q12–q13. Genomics 36: 86–99.
Cruts, M., Backhovens, H., Theuns, J., Clark, R. F., Le Paslier, D.,ACKNOWLEDGMENTS Weissenbach, J., Goate, A. M., Martin, J.-J., and Van Broeckhoven,
C. (1995). Genetic and physical characterization of the early-onsetAlzheimer’s disease AD3 locus on chromosome 14q24.3. Hum. Mol.We are grateful to Stephan Gasca and Gail Billingsley for theirGenet. 4: 1355–1364.constant support and critical comments. We thank Naomi Kautz for
technical help. We acknowledge with thanks the support of the Ge- Duyck, G. M., Kim, S. M., Myers, R. M., and Cox, D. R. (1990). Exonnetic Diseases Section of the Network of Centers of Excellence for trapping: A genetic screen to identify candidate transcribed se-sequencing (Sylvia Soder) and for FISH mapping (Linda Anderson). quences in cloned mammalian genomic DNA. Proc. Natl. Acad.PAC screening was carried out by Jack Huizenza in the facilities of Sci. USA 87: 8995–8999.the Canadian Genome Analysis & Technology Program. This work Gilbert, T. L., Hadelman, B. A., Mulvihill, E., and O’Hara, P. J.is supported by a grant from the Medical Research Council (MRC) (1992). A mammalian homologue of a transcript from the Drosoph-of Canada (MT5426) and the Centers of Excellence. J.M.R. is a ila pecanex locus. J. Neurogenet. 8: 181–187.Scholar of the MRC.
Glaser, T., Jepeal, L., Edwards, J. G., Young, S. R., Favor, J., andMaas, R. L. (1994). PAX6 gene dosage effect in a family with con-
REFERENCES genital cataracts, aniridia, anophthalmia and central nervous sys-tem defects. Nature Genet. 7: 463–470.
Altschul, S., Gish, W., Miller, W., Myers, E., and Lipman, D. J. Gyapay, G., Morissette, J., Vignal, A., Did, C., Fizames, C., Millas-(1990). Basic local alignment search tool. J. Mol. Biol. 215: 403– seau, P., Marc, S., Bernadi, G., Lathrop, M., and Weissenbach,410. J. (1994). The 1993–1994 Genethon human genetic linkage map.
Nature Genet. 7: 246–339.Berry, V., Ionides, A. C. W., Moore, A. T., Plant, C., Battacharya,
AID GENO 4786 / 6r3a$$$$$1 06-18-97 18:25:01 gnma

ROUX ET AL.140
Hattori, M., Tsukamoto, N., Nur-E-kamal, M. S. A., Rubinfeld, B., Parimoo, S., Patanjali, S. R., Shukla, H., Chaplin, D. D., and Weiss-man, S. M. (1991). cDNA selection: Efficient PCR approach for theIwai, K., Kubota, H., Maruta, H., and Minato, N. (1995). Molecular
cloning of a novel mitogen-inducible nuclear protein with a Ran selection of cDNAs encoded in large chromosomal DNA fragments.Proc. Natl. Acad. Sci. USA 88: 9623–9627.GTPase-activating domain that affects cell cycle progression. Mol.
Cell. Biol. 15: 552–560. Rampazzo, A., Nava, A., Danieli, G. A., Buja, G., Dallento, L., Fasoli,Heiss, N. S., Rogner, U. C., Kiochis, P., Korn, B., and Poustka, A. G., Scognamiglio, R., Corrado, D., and Thiene, G. (1994). The gene
(1996). Transcriptional mapping in a 700kb region around DXS52 for arrhythmogenic right ventricular cardiomyopathy maps tolocus in Xq28: Isolation of six novel transcripts and a novel ATPase chromosome 14q23–q24. Hum. Mol. Genet. 3: 959–962.isoform (hPMCA5). Genome Res. 6: 478–491. Rommens, J. M., Mar, L., McArthur, J., Tsui, L.-C., and Scherer,
Hejtmancik, J. F., Kaiser, M. I., and Piatigorsky, J. (1995). Molecular S. W. (1994). Towards a transcriptional map of the q21–q22 regionbiology and inherited disorders of the eye lens. In ‘‘The Metabolic of chromosome 7. In ‘‘Identification of Transcribed Sequences’’ (U.and Molecular Bases of Inherited Disease’’ (C. R. Scriver, A. L. Hochgeschwender and K. Gardiner, Eds.), pp. 65–79, Plenum,Beaudet, W. S. Sly, and D. Valle, Eds.), pp. 4325–4349, McGraw– New York.Hill, New York. Rubin, S. E., Nelson, L. B., and Pletcher, B. A. (1994). Anterior polar
Hudson, T. J., Stein, L. D., Gerety, S. S., Ma, J., Castle, A. B., Silva, cataract in two sisters with an unbalanced 3;18 chromosomalJ., Slonim, D. K., Baptista, R., Kruglyak, L., Xu, S.-H., Hu, X., translocation. Am. J. Ophthalmol. 117: 512–515.Colbert, A. M. E., Rosenberg, C., Reeve-Daly, M. P., Rozen, S., Hui,
Scherer, S., and Tsui, L.-C. (1991). Cloning and analysis of largeL., Wu, X., Vestergaard, C., Wilson, K. M., Bae, J. S., Maitra, S.,molecules. In ‘‘Advanced Techniques in Chromosome Research’’Ganiatsas, S., Evans, C. A., DeAngelis, M. M., Ingalls, K. A., Nahf,(K. W. Adolph, Ed.), pp. 33–72, Dekker, New York/Basel/HongR. W., Horton, L. T., Anderson, M. O., Collymore, A. J., Ye, W.,Kong.Kouyoumjian, V., Zemsteva, I. S., Tam, J., Devine, R., Courtney,
Stambolian, D., Ai, Y., Sidjanin, D., Nesburn, K., Sathe, G., Rosen-D. F., Turner Renaud, M., Nguyen, H., O’Connor, T. J., Fizames,berg, M., and Bergsma, D. J. (1995). Colning of the galactokinaseC., Faure, S., Gyapay, G., Dib, C., Morissette, J., Orlin, J. B., Bir-cDNA and identification of mutations in two families with cata-ren, B. W., Goodman, N., Weissenbach, J., Hawkins, T. L., Foote,racts. Nature Genet. 10: 307–312.S., Page, D. C., and Lander, E. S (1995). An STS-based map of the
human genome. Science 270: 1945–1954. Triglia, T., Peterson, M. G., and Kemp, D. J. (1988). A procedurefor in vitro amplification of DNA segments that lie outside theIoannou, P. A., Amemiya, C. T., Garnes, J., Kroisel, P. M., Shizuya,boundaries of known sequences. Nucleic Acids Res. 16: 8186.H., Chen, C., Batzer, M. A., and de Jong, P. J. (1994). A new bacte-
riophage P1-derived vector for the propagation of large human Uberbacher, E. C., and Mural, R. J. (1991). Locating protein codingDNA fragments. Nature Genet. 6: 84–89. regions in human DNA sequences by a multiple sensor–neural
LaBonne, S. G., Sunitha, I., and Mahowald, A. P. (1989). Molecular network approach. Proc. Natl. Acad. Sci. USA 88: 11261–11265.genetics of pecanex, a maternal-effect neurogenic locus of Drosoph- Willott, E., Balsad, M. S., Fanning, A. S., Jameson, B., Van Itallie,ila melanogaster that potentially encodes a large transmembrane C., and Anderson, J. M. (1993). The tight junction protein ZO-1 isprotein. Dev. Biol. 136: 1–16. homologous to the Drosophila discs-large tumor suppressor protein
Lovett, M., Kere, J., and Hinton, L. M. (1991). Direct selection: A of septate junctions. Proc. Natl. Acad. Sci. USA 90: 7834–7838.method for the isolation of cDNAs encoded by large genomic region. Wilson, R., Ainscough, R., Anderson, K., Baynes, C., Berks, M., Bon-Proc. Natl. Acad. Sci. USA 88: 9628–9632. field, J., Burton, J., Connell, M., Copsey, T., Cooper, J., Coulson,
Moross, T., Vaithilingam, S. S., Styles, S., and Gardner, H. A. (1984). A., Craxton, M., Dear, S., Du, Z., Durbin, R., Favello, A., Fulton,Autosomal dominant anterior polar cataracts associated with a L., Garner, A., Green, P., Hawkins, T., Hillier, L., Jier, M., John-familial 2;14 translocation. J. Med. Genet. 21: 52–53. ston, L., Jones, M., Kershaw, J., Kirsten, J., Laister, N., Latreille,
Ochmann, H., Gerber, A. S., and Hartl, D. L. (1988). Genetic applica- P., Lightning, J., Lloyd, C., McMurray, A., Mortimore, B., O’Cal-tions of an inverse polymerase chain reaction. Genetics 120: 621– laghan, M., Parson, J., Percy, C., Rifken, L., Roopra, A., Saunders,623. D., Sulston, J., Thierry-Mieg, J., Thomas, K., Vaudin, M.,
Vaughan, K., Waterson, R., Watson, A., Weinstock, L., Wilkinson-Padma, T., Ayyagari, R., Murty, J. S., Basti, S., Fletcher, T., Rao,G. N., Kaiser-Kupfer, M., and Hejtmancik, J. F. (1995). Autosomal Sproat, J., and Wohldman, P. (1994). 2.2Mb of contiguous nucleo-
tide sequence from chromosome III of C.elegans. Nature 368: 32–dominant zonular cataract with sutural opacities localized to chro-mosome 17q11–2. Am. J. Hum. Genet. 57: 840–845. 38.
AID GENO 4786 / 6r3a$$$$$1 06-18-97 18:25:01 gnma