phylogeny of red palm weevil
TRANSCRIPT

Full Terms & Conditions of access and use can be found athttp://www.tandfonline.com/action/journalInformation?journalCode=toin20
Download by: [King Saud University] Date: 25 October 2015, At: 00:36
Oriental Insects
ISSN: 0030-5316 (Print) 2157-8745 (Online) Journal homepage: http://www.tandfonline.com/loi/toin20
Phylogeny of red palm weevil (Rhynchophorusferrugineus) based on ITS1 and ITS2
Monther T. Sadder, Polana S. P. V. Vidyasagar, Saleh A. Aldosari, MahmoudM. Abdel-Azim & Abdullah A. Al-Doss
To cite this article: Monther T. Sadder, Polana S. P. V. Vidyasagar, Saleh A. Aldosari, MahmoudM. Abdel-Azim & Abdullah A. Al-Doss (2015): Phylogeny of red palm weevil (Rhynchophorusferrugineus) based on ITS1 and ITS2, Oriental Insects, DOI: 10.1080/00305316.2015.1081639
To link to this article: http://dx.doi.org/10.1080/00305316.2015.1081639
Published online: 19 Oct 2015.
Submit your article to this journal
Article views: 4
View related articles
View Crossmark data

Oriental insects, 2015http://dx.doi.org/10.1080/00305316.2015.1081639
Phylogeny of red palm weevil (Rhynchophorus ferrugineus) based on ITS1 and ITS2
Monther T. Saddera,b, Polana S. P. V. Vidyasagarc, Saleh A. Aldosaric, Mahmoud M. Abdel-Azimc and Abdullah A. Al-Dossb
aFaculty of agriculture, Department of Horticulture and crop science, the University of Jordan, amman, Jordan; bDepartment of Plant Production, college of Food and agricultural sciences, King saud University, riyadh, saudi arabia; cDepartment of Plant Protection, college of Food and agricultural sciences, chair of Date Palm research, King saud University, riyadh, saudi arabia
Introduction
Red palm weevil (RPW), Rhynchophorus ferrugineus (Olivier) (Curculionidae: Coleoptera), was reported as a pest of coconut palm in India for more than one hundred years ago (Lefroy 1906; Gosh 1912). Recently, it became the major destructive insect of date palms (Abraham et al. 1998; Gómez and Ferry 1999). RPW is polyphagous and attacks both commercial palms and several ornamental palms as well. The invasive pest is very difficult to detect in early stages of infesta-tion because of its cryptic nature (Nirula 1956). Several control methods have been practiced to control PRW such as chemical control with pesticides, mechanical control and cultural practices (Vidyasagar and Bhat 1991).
ABSTRACTRed palm weevil (Rhynchophorus ferrugineus Olivier) populations were collected from several regions in Saudi Arabia. Insects were graded based on different patterns of pronotum markings. The entire ITS1-5.8S-ITS2 region was cloned and sequenced for both R. ferrugineus and its related species R. vulneratus (Panzer). The novel ITS1 sequence form Rhynchophorus was found to be unique in the current Genbank database. Discrimination power of ITS1 region was shown to be much higher than ITS2 region. Penetrance of different pronotum markings varied from one region to another. The pronotum-based clustering diverged from that revealed by ribosomal sequence. Several Indels and nucleotide substitutions were detected along the ITS1-5.8S-ITS2 region between R. ferrugineus and R. vulneratus. The data support a two-species classification rather than considering them colour morphs of the same species.
© 2015 taylor & Francis
KEYWORDS5.8s; its1; its2; Rhynchophorus ferrugineus; R. vulneratus
ARTICLE HISTORYreceived 21 October 2013 accepted 6 august 2015
CONTACT Monther t. sadder [email protected]
Dow
nloa
ded
by [
Kin
g Sa
ud U
nive
rsity
] at
00:
36 2
5 O
ctob
er 2
015

2 M. T. SADDeR eT Al.
The insect has been spreading steadily westward since the mid 1980s. It reached the eastern region of Kingdom of Saudi Arabia (KSA) in 1987 (Al-Abdulmohsin 1987) and spread to many other areas in the country (Abraham et al. 1998). A total of about 25 million date palms are in risk of infestation with RPW in KSA (Ministry of Agriculture 2010). It was first recorded in northern United Arab Emirates (UAE) in 1985. Afterwards, it has covered the entire country and neighbouring Oman (El-Ezaby et al. 1998). In Iran, it was discovered in southern regions in 1990 (Faghih 1996) and in Egypt at the end of 1992 (Cox 1993). In 1994, it had been captured in Spain (Barranco et al. 1996) and subsequently spread to other EU countries (Italy, Portugal, France, Greece and Cyprus). RPW was found in Jordan in 1999 (Kehat 1999), in Japan in 2003 (Abe et al. 2009) and in USA in 2010 (USDA-APHIS-PPQ 2010). Therefore, it is considered a universal and key pest of all palm tree species and new innovative methods should be utilised in combating this menace.
PCR-based DNA markers have been successfully used for estimating genetic diversity and studying population genetics on a wide range of geographic scale. These techniques vary in complicity, reliability and information generating capacity. Among others, randomly amplified polymorphic DNA (RAPD) was extensively used for genetic diversity studies in different insect species. It was used to identify necrophagous insects (Benecke 1998), to distinguish species and strains of rice weevil (Hidayat et al. 1996), gypsy moth (Garner and Slavicek 1996) and Indianmeal moth (Dowdy and MeGaughey 1996). RAPD was also used to study introgression in budworm species (Deverno et al. 1998), genetic structuring and gene flow among populations of boll weevils in South America (Scataglini et al. 2000), gene flow among curculionid weevils (Scataglini et al. 2000; Vandewoestijne and Baguette 2002; Ayres et al. 2003) and RPW genetic diversity (Hallett et al. 2004; Gadelhak and Enan 2005). However, RAPD technique is not suitable for phylogenetic analysis and has several drawbacks (Landry and Lapointe 1996; Bardakci 2001). A phylogeny can be achieved utilising sequence data, e.g. cytochrome oxidase subunit 1 (COI) (El-Mergawy, Faure, et al. 2011) and internal transcribed spacer (ITS) (El-Mergawy, Al Ajlan, et al. 2011).
Although the biology of RPW has been studied extensively, the population genetics and biodiversity is still elusive. The aim of this study was to elucidate the diversity of RPW from different countries based on pronotum markings and ribosomal sequence.
Materials and methods
Insect samples
For collection and preservation of R. ferrugineus (Olivier) adult samples, 300 ml glass jars were filled with 95% ethanol. Jars were securely packed in a spill-proof box. Insects were collected under the jurisdiction of the Ministry of Agriculture from seven different areas in Saudi Arabia (Table 1). From each location, fresh and healthy adults (20 males and females each) were selected and preserved.
Dow
nloa
ded
by [
Kin
g Sa
ud U
nive
rsity
] at
00:
36 2
5 O
ctob
er 2
015
ORIenTAl InSecTS 3
Collection glass jars were labelled (governorate, location, sex, number and name of collector or authorised person) and sent to the laboratory. In total, 280 insect samples were procured and kept in refrigerator for further use.
Table 1. Palm weevil samples utilised in this study along with their origins.
*Wattanapongsiri (1966).
# country, region Host palm GPS coordinates Species existence (years)
1 Ksa, Qatif Date palm 26°33′18.21″n R. ferrugineus ~2549°59′10.45″e
2 Ksa, al ahsa Date palm 25°23′5.07″n R. ferrugineus ~2249°36′50.66″e
3 Ksa, Kharj Date palm 24°13′0.88″n R. ferrugineus ~1747°15′43.42″e
4 Ksa, Diriyah Date palm 24°44′47.05″n R. ferrugineus ~1846°33′25.85″e
5 Ksa, Wadi al Dawasir Date palm 20°28′4.79″n R. ferrugineus ~2144°51′32.48″e
6 Ksa, Mecca Date palm 21°25′6.11″n R. ferrugineus ~ 1939°46′40.67″e
7 Ksa, najran Date palm 17°27′48.29″n R. ferrugineus ~1844°6′10.41″e
8 Uae, abu Dhabi Date palm na R. ferrugineus ~249 spain, catalonia canary island date palm na R. ferrugineus ~1610 italy, sicily canary island date palm na R. ferrugineus ~1411 indonesia coconut na R. vulneratus ~100*
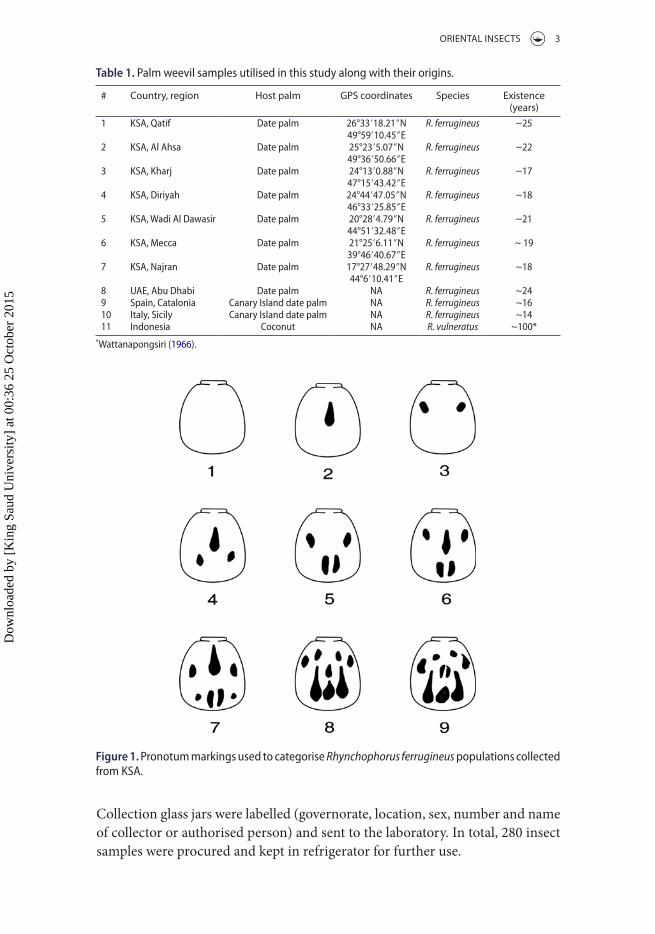
Figure 1. Pronotum markings used to categorise Rhynchophorus ferrugineus populations collected from Ksa.
Dow
nloa
ded
by [
Kin
g Sa
ud U
nive
rsity
] at
00:
36 2
5 O
ctob
er 2
015

4 M. T. SADDeR eT Al.
Adult insects were studies in a careful and non-destructive way to record the dark colour markings on the pronotum. Depending on the number of dots or marks present on the pronotum, nine categories were prepared (Figure 1). Examined insects were grouped into these nine categories and their percentage in the population was calculated. Data were subjected to hierarchical cluster analysis based on Euclidean distance using SPSS software version 20.
Molecular analysis
A total of 20 insects, preserved in absolute ethanol, were used for the molecular analysis (Table 1). Two R. ferrugineus insects (male and female) were used for each KSA collection region. In addition, two R. ferrugineus samples were used from UAE, a field sample and laboratory-reared one and two European samples one from Italy and another from Spain. Furthermore, two R. vulneratus (Panzer) insects (male and female) from Indonesia served as outgroup species.
Samples were washed thoroughly with water and ground with sand using mor-tar and pestle. Total gDNA was isolated using Wizard genomic DNA purification kit (Promega, USA) with some modifications. Each ground insect was incubated overnight in rotary incubator at 55 °C in insect lysis buffer (500 μl nuclei lysis solution, 120 μl EDTA (pH 8.0), 17.5 μl of 20 mg/ml proteinase K (Sigma, USA)). Isolated DNA was quantified with gel electrophoresis and spectrophotometer. Samples were diluted accordingly and stored at −20 °C.
DNA was amplified using thermocycler machine with heated lid (ABI, USA). The PCR programme includes 3 min at 95 °C, 35 amplification cycles (94 °C for 30 s; 50 °C for 30 s; 72 °C for 1 min) and an extra extension cycle for 10 min at 72 °C. PCR reactions were carried out in 20 μl volumes using DNA polymerase master mix (Promega, USA), 0.4 μM primer (IDT-DNA, Belgium) and 10 ng of gDNA. PCR primers were designed to amplify ribosomal DNA regions; a primer pair for 12S ribosomal DNA (forward: 5′-TAGTAGTAGCTATGTTCTTG-3′; reverse: 5′-GAGTGTAAAGTTGTATTTCC-3′) and another pair for 18S(partial)-ITS1–5.8S-ITS2–28S(partial) region (forward: 5′-AACCTGCGGAAGGATCATTA-3′; reverse: 5′-AGTCTCACCTGCTCTGAGG-3′).
PCR amplicons were run using 1% agarose gel electrophoresis for 90 min at 100 volt in 1x TAE buffer (40 mM Tris-acetate, 1 mM EDTA, pH 8.0). Hyperladder VI (Bioline, USA) was used as DNA marker. Gels were stained with SYBR Gold (Life Technologies, USA) and bands were excised out with scalpel under safe blue light. DNA was recovered from gel pieces using Wizard Gel and PCR Clean-Up kit (Promega, USA). Fragments were cloned into pGEM vector (Promega, USA) and transformed with heat shock into E. coli DH5α cells (Stratagene, USA). Competent E. coli cells were chemically prepared with CaCl2 (Sambrook et al. 1989). Putative colonies with recombinant plasmids were selected over LB plates containing ampi-cillin, IPTG and X-Gal (Sambrook et al. 1989). Plasmids were isolated from over-night cultures from selected clones using plasmid Miniprep kit (Promega, USA)
Dow
nloa
ded
by [
Kin
g Sa
ud U
nive
rsity
] at
00:
36 2
5 O
ctob
er 2
015

ORIenTAl InSecTS 5
and bi-directionally sequenced with Sanger method (ABI, USA). The sequences were deposited in Genbank with accession numbers (KC954631 to KC954642).
ITS2 sequences (621 bp) for Rhynchophorus spp. and 5.8S ribosomal sequences (163 bp) from related insects were retrieved from Genbank (NCBI 2013). Sequences were aligned using ClustalW multiple alignment function available in BioEdit (Hall 1999). Alignments were bootstrapped 100 times using SEQBOOT func-tion available in Phylip (Felsenstein 1989) and subjected to maximum likelihood method using the DNAML function (Felsenstein 1989; Felsenstein and Churchill 1996). The transition/transversion ratio was two and a global rearrangement was selected, which causes each possible group to be removed and re-added after the last species is added to the tree. Extended majority rule consensus tree was gener-ated using CONSENSE function and plotted with TreeView software (Page 1996).
Results
Pronotum markings
The penetrance of pronotum markings in adult RPW varied for studied regions. Insects without any marking or with one spot (marking 1) were recorded only in Mecca with 16.67% and 3.33%, respectively. On the other hand, insects with multiple spots (marking 9) were prominent in Wadi Al Dawasir region (30%). Four different markings were recorded for insects from Qatif region with 22.22, 16.67, 22.22 and 38.89% for markings 3, 5, 6 and 8, respectively.
Pronotum markings-based cluster analysis revealed two major clusters (Figure 2). The first cluster grouped insects form Najran, Kharj, Mecca and Wadi Al Dawasir. The second cluster grouped insects from Diriyah, Al Ahsa and Qatif regions.
Phylogeny of RPW
Repetitive 12S ribosomal DNA was used to screen isolated gDNA for PCR. 12S ribosomal DNA was successfully amplified in all samples. Likewise, the second designed primer pair covering the entire region spanning ITS1–5.8S-ITS2 was
Figure 2. Dendrogram clustering R. ferrugineus populations collected from Ksa based on pronotum markings.
Dow
nloa
ded
by [
Kin
g Sa
ud U
nive
rsity
] at
00:
36 2
5 O
ctob
er 2
015
6 M. T. SADDeR eT Al.
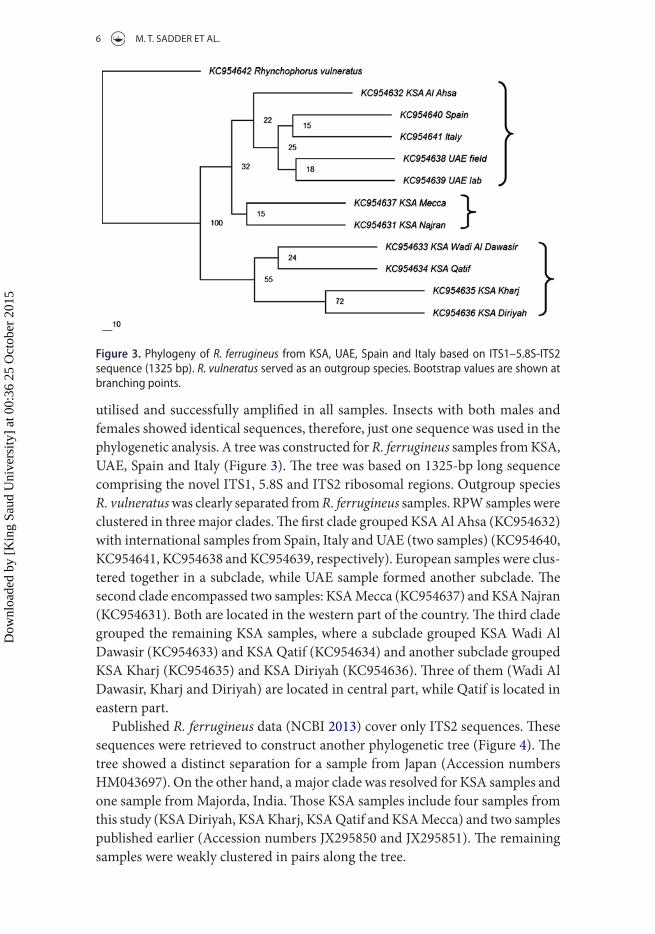
utilised and successfully amplified in all samples. Insects with both males and females showed identical sequences, therefore, just one sequence was used in the phylogenetic analysis. A tree was constructed for R. ferrugineus samples from KSA, UAE, Spain and Italy (Figure 3). The tree was based on 1325-bp long sequence comprising the novel ITS1, 5.8S and ITS2 ribosomal regions. Outgroup species R. vulneratus was clearly separated from R. ferrugineus samples. RPW samples were clustered in three major clades. The first clade grouped KSA Al Ahsa (KC954632) with international samples from Spain, Italy and UAE (two samples) (KC954640, KC954641, KC954638 and KC954639, respectively). European samples were clus-tered together in a subclade, while UAE sample formed another subclade. The second clade encompassed two samples: KSA Mecca (KC954637) and KSA Najran (KC954631). Both are located in the western part of the country. The third clade grouped the remaining KSA samples, where a subclade grouped KSA Wadi Al Dawasir (KC954633) and KSA Qatif (KC954634) and another subclade grouped KSA Kharj (KC954635) and KSA Diriyah (KC954636). Three of them (Wadi Al Dawasir, Kharj and Diriyah) are located in central part, while Qatif is located in eastern part.
Published R. ferrugineus data (NCBI 2013) cover only ITS2 sequences. These sequences were retrieved to construct another phylogenetic tree (Figure 4). The tree showed a distinct separation for a sample from Japan (Accession numbers HM043697). On the other hand, a major clade was resolved for KSA samples and one sample from Majorda, India. Those KSA samples include four samples from this study (KSA Diriyah, KSA Kharj, KSA Qatif and KSA Mecca) and two samples published earlier (Accession numbers JX295850 and JX295851). The remaining samples were weakly clustered in pairs along the tree.
Figure 3. Phylogeny of R. ferrugineus from Ksa, Uae, spain and italy based on its1–5.8s-its2 sequence (1325 bp). R. vulneratus served as an outgroup species. Bootstrap values are shown at branching points.
Dow
nloa
ded
by [
Kin
g Sa
ud U
nive
rsity
] at
00:
36 2
5 O
ctob
er 2
015

ORIenTAl InSecTS 7
Rhynchophorus spp
An additional phylogenetic investigation was applied to reveal the relationship among Rhynchophorus spp. Available ITS2 sequences for related species R. phoe-nicis, R. bilineatus and R. palmarum were retrieved from Genbank (NCBI 2013). The phylogenetic tree clearly separated the outgroup species Sitophilus zeamais (Figure 5). The tree revealed a consequent evolution of Rhynchophorus spp., where R. palmarum was branched out first, followed by R. phoenicis and then R. ferrugineus. On the other hand, the two species R. bilineatus and R. vulneratus clustered in one clade with good bootstrap value (63%). Moreover, the novel ITS1 sequence supports the speciation of palm weevils into two species: R. ferrugineus and R. vulneratus. The alignment of the consensus sequence between R. ferrugi-neus and R. vulneratus resolved eleven Insertion–Deletion (Indel) polymorphisms
Figure 4. Phylogeny of R. ferrugineus from this study (accession numbers Kc955631–42) and published data (ncBi 2013) based on its2 sequence (621 bp). Bootstrap values are shown at branching points.
Dow
nloa
ded
by [
Kin
g Sa
ud U
nive
rsity
] at
00:
36 2
5 O
ctob
er 2
015

8 M. T. SADDeR eT Al.
and sixteen nucleotide substitutions in the ITS1 region (Table 2), while one Indel and eleven nucleotide substitutions were detected in ITS2 region (Table 2).
Complete 5.8S ribosomal sequence of the genus Rhynchophorus was further utilised to get a prospective of palm weevils among related insects. The 5.8S sequence of 19 different insects were retrieved and utilised to construct a phy-logenetic tree with Rhynchophorus (Figure 6). The tree was resolved into three major clades, while Cryptolaemus montrouzieri did not cluster with any clade. First clade clustered the genera Campoletis, Anselmella, Mallada, Dasymutilla, Anthonomus and Sitophilus. Second clade grouped the genera Bracon, Leiophron, Tenebrio, Trichogramma and Solenopsis. And finally, Rhynchophorus was clustered in a third clade with Altica litigata and Lysathia ludoviciana.
Figure 5. Phylogeny of Rhynchophorus species based on its2 sequence (621 bp). R. ferrugineus (Kc954636 Ksa Diriyah) and R. vulneratus (Kc954642) are from this study. R. phoenicis (HM043703), R. bilineatus (HM043701) and R. palmarum (HM043699) are from Genbank (ncBi 2013). Sitophilus zeamais (aF276518) served as an outgroup species. Bootstrap values are shown at branching points.
Table 2. indels and nucleotide substitutions detected in its1 and its2 regions sequenced from both R. ferrugineus and R. vulneratus.
Base Region R. ferrugineus R. vulneratus Base Region R. ferrugineus R. vulneratus
30 its1 t 636 its1 a c31 its1 t 672 its1 t44 its1 t 673 its1 c45 its1 a 674 its1 G46 its1 t 690 its1 G c47 its1 a 695 its1 G t93 its1 t c 704 its1 G a120 its1 G a 998 its2 c G212 its1 a G 917 its2 t a228 its1 G a 919 its2 t c233 its1 t 942 its2 t c234 its1 a 983 its2 a G309 its1 G c 1032 its2 c t318 its1 a t 1072 its2 a ∙347 its1 c t 1081 its2 c t369 its1 a G 1177 its2 t c489 its1 c G 1200 its2 a G507 its1 a c 1252 its2 G a508 its1 c t 1264 its2 t a516 its1 t G
Dow
nloa
ded
by [
Kin
g Sa
ud U
nive
rsity
] at
00:
36 2
5 O
ctob
er 2
015

ORIenTAl InSecTS 9
Discussion
Ministry of Agriculture in KSA has implemented various control programmes to combat RPW. A pioneer IPM project for RPW was carried out in 1992 (Abraham and Vidyasagar 1992). The project included pheromone trapping techniques and pest management procedures. However, RPWs are still spreading in KSA and worldwide. Our data revealed a detectable genetic diversity among investigated samples. Such diversity could help the insect to escape management schemes and combating plans.
Collected populations of R. ferrugineus demonstrated a wide spectrum of pronotum markings (Figure 1). In a comprehensive survey, 24 different pat-terns of pronotum markings were recorded for R. ferrugineus (Wattanapongsiri 1966), only two of which overlapped with ours (Figure 1). It is worth noting that Wattanapongsiri’s survey (1966) was conducted when the insects were still restricted to Southeast Asia (India, Philippines, Vietnam, Thailand, Taiwan and Burma). Accordingly, it is important to understand the nature behind changes in pronotum markings along the spread and distribution of the insect.
We investigated the potential use of these markings for diversity studies. The pronotum markings-based tree (Figure 2) was compared with the phylogenetic tree (Figure 3). Clustering data correlates for some populations but contradicts for others. Similar conclusions were found in earlier reports utilising different genetic data. Marvaldi et al. (2002) studied phylogeny of weevils based on combined 18S rDNA and morphological data. They suggested that diversification in weevils
Figure 6. Phylogeny of Rhynchophorus (this study) and related insects based on 5.8s sequence (163 bp). accession numbers are followed by scientific names (ncBi 2013). Bootstrap values are shown at branching points.
Dow
nloa
ded
by [
Kin
g Sa
ud U
nive
rsity
] at
00:
36 2
5 O
ctob
er 2
015

10 M. T. SADDeR eT Al.
was accompanied by niche shifts in host–plant associations and larval habits. Pronounced conservatism was evident in larval feeding habits, particularly in the host tissue consumed. On the other hand, a genetic relationship between three phenotypic different forms of RPW from one location of KSA Al Ahsa (Al-Hassa) region was investigated using six RAPD primers (Al-Ayied et al. 2006). Although weevils were collected from the same geographical region, it was suggested that banding profile acquired from black and brown coloured morphs were genetically closer compared to the brown spotted morphs. Intra genetic variation was lower in black than brown and brown spotted morphs. The authors explained this genetic variation by either mutated non-spotted/spotted morphs or a different weevil race.
Evaluation of genetic diversity based on morphological traits does not usually provide accurate estimates of genetic differences as they are highly influenced by environmental factors. This can also be influenced by genetic diversity between insect species and even within populations of the same species (Apostol et al. 1993; Stevens and Wall 1995). Nonetheless, some pronotum markings were found to be more frequent than others in certain RPW populations. Therefore, it is highly recommended to investigate both the penetrance and expressivity of pronotum markings under controlled laboratory conditions. This can be achieved by utilising planned mating systems for insects with various pronotum markings.
Limited genetic diversity studies have been published for the RPW (Salama and Saker 2002; Gadelhak and Enan 2005; Al-Ayied et al. 2006). From Egypt, three morphologically different forms of RPWs collected from infested date palm fields were subjected to nine RAPD primers (Salama and Saker 2002). PCR amplification products indicated that three different RAPD patterns align with three different forms of weevils. They concluded that the detected genetic variations could be assigned to the generation of possible mutants or they may belong to different races. Seven other populations of the RPW insects collected from different loca-tions from UAE were analysed using six RAPD 10-mers (Gadelhak and Enan 2005). A 51.4% of the generated bands were polymorphic. The populations were clustered into three groups with genetic variability ranging from 38 to 94% and good correlation between genetic and geographical distances among investigated populations. However, RAPD markers generate non-specific amplifications, which is a major drawback when applied for genetic diversity studies. DNA from RPW gut microorganism (Khiyami and Alyamani 2008) can also be amplified. These microorganisms vary from one isolate to another (Ibrahim et al. 2011) and from one season to another (Jia et al. 2013).
Phylogenetic data of R. ferrugineus were limited to ITS2 region as this was the only published sequence (El-Mergawy, Al Ajlan, et al. 2011; NCBI 2013). El-Mergawy, Faure, et al. (2011) could not detect ITS2 polymorphism among different RPW samples from thirteen international locations. The limited discrim-ination power of ITS2 resulted in consolidation of some clades (Figure 4), which could be resolved after incorporating ITS1 sequence (Figure 3). This scenario emphasises the polymorphic features embedded in the novel ITS1 sequence for
Dow
nloa
ded
by [
Kin
g Sa
ud U
nive
rsity
] at
00:
36 2
5 O
ctob
er 2
015

ORIenTAl InSecTS 11
R. ferrugineus. This sequence is so unique that no single blastn hit could be detected in the Genbank (NCBI 2013).
There are reports of about ten species of Rhynchophorus distributed in various geographical regions of the world. Among them, R. ferrugineus and R. vulneratus are sympatric in nature. Based on morphological and molecular studies, it was suggested to consider them two morphs of one species (Hallett et al. 2004). Our findings disagree with this classification for several reasonable facts. The most comprehensive description of the genus Rhynchophorus was published by Wattanapongsiri (1966), where several morphological features were described to distinguish between the two species R. ferrugineus and R. vulneratus. On the other hand, supporting data for considering them as synonyms (Hallett et al. 2004) was based on two molecular tools. The first was RAPD technique, which cannot be utilised for phylogenetic analysis, while the second was COI, which has some inherent limitations. In some cases, COI was found to have little reso-lution at the species level (Derycke et al. 2010), while in other cases, the nuclear copies of COI can lead to overestimation of the taxonomic diversity (Song et al. 2008). Nonetheless, our conclusion is supported by recent work based on COI (Rugman-Jones et al. 2013). Moreover, detected Indels and nucleotide substi-tutions based on both novel ITS1 and ITS2 sequences (Table 2) revealed an undisputed evidence of molecular diversification between R. ferrugineus and R. vulneratus. Finally, the phylogenetic analysis encountering available sequences of related Rhynchophorus spp. revealed a striking genetic proximity between R. vulneratus and R. bilineatus rather than between R. vulneratus and R. ferrug-ineus (Figure 5).
Acknowledgements
The authors would like to thank the Ministry of Agriculture in KSA for facilitating the collec-tion of insect samples and our collaborators for kindly supplying the International samples.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
Abe F, Hata K, Sone K. 2009. Life history of the red palm weevil, Rhynchophorus ferrugineus (Coleoptera: Dryophtoridae), in Southern Japan. Florida Entomologist. 92:421–425.
Abraham VA, Al Shuaibi MA, Faleiro JR, Abuzuhairah RA, Vidyasagar PSPV. 1998. An integrated management approach for the red palm weevil, Rhynchophorus ferrugineus Oliv., a key pest of date palm in the Middle East. Sultan Qabus University Journal for Scientific Research, Agricultural Sciences. 3:77–84.
Abraham VA, Vidyasagar PSPV. 1992. Strategy for the control of red palm weevil of date palm in the Kingdom of Saudi Arabia. Part II. Consultancy report submitted to the Ministry of Agriculture and Water, Riyadh, Kingdom of Saudi Arabia, 32 pp.
Dow
nloa
ded
by [
Kin
g Sa
ud U
nive
rsity
] at
00:
36 2
5 O
ctob
er 2
015

12 M. T. SADDeR eT Al.
Al-Abdulmohsin AM. 1987. First record of red date palm weevil in Saudi Arabia. Arab World Agriculture. 3:15–16.
Al-Ayied HY, Alswailem AM, Shair O, Al Jabr AM. 2006. Evaluation of phylogenetic relationship between three phenotypically different forms of red date palm weevil Rhynchophorus ferrugineus Oliv. using PCR-based RAPD technique. Archives of Phytopathology and Plant Protection. 39:303–309.
Apostol BL, Black WC, Miller BR, Reiter P, Beaty BJ. 1993. Estimation of the number of full sibling families at an oviposition site using RAPD–PCR markers: applications to the mosquito Aedes aegypti. Theoretical and Applied Genetics. 86:991–1000.
Ayres CFJ, Melo-santos MAV, Solé-Cava AM, Furtado AF. 2003. Genetic differentiation of Aedes aegypti (Diptera: Culicidae), the major dengue vector in Brazil. Journal of Medical Entomology. 40:430–435.
Bardakci F. 2001. Random amplified polymorphic DNA (RAPD) markers. Turkish Journal of Biology. 25:185–196.
Barranco P, De La Peña J, Cabello T. 1996. The red palm weevil, Rhynchophorus ferrugineus (Olivier), new pest in Europe. (Coleoptera, Curculionidae). Phytoma España. 76:36–40.
Benecke M. 1998. Random amplified polymorphic DNA (RAPD) typing of necrophageous insects (Diptera, Coleoptera) in criminal forensic studies: validation and use in practice. Forensic Science International. 98:157–168.
Cox ML. 1993. Red palm weevil, Rhynchophorus ferrugineus in Egypt. FAO Plant Protection Bulletin. 41:30–31.
Derycke S, Vanaverbeke J, Rigaux A, Backeljau T, Moens T. 2010. Exploring the use of cytochrome oxidase c subunit 1 (COI) for DNA barcoding of free-living marine nematodes. PLoS ONE. 5(10):e13716.
Deverno LL, Smith GA, Harrison KJ. 1998. Randomly amplified polymorphic DNA evidence of introgression in two closely related sympatric species of coniferophagous Choristoneura (Lepidoptera: Tortricidae) in Atlantic Canada. Annals of the Entomological Society of America. 91:248–259.
Dowdy AK, McGaughey WH. 1996. Using random amplified polymorphic DNA to differentiate strains of the Indian meal moth (Lepidoptera: Pyralidae). Environmental Entomology. 25:396–400.
El-Ezaby FA, Khalifa O, El Assal A. 1998. Integrated pest management for the control of red palm weevil Rhynchophorus ferrugineus Oliv. in The United Arab Emirates, Eastern Region, Al Ain. Proceedings of the First International Conference on Date Palms; Al-Ain, UAE, 269–281 pp.
El-Mergawy RAAM, Al Ajlan AM, Abdalla N, Vassiliou V, Capdevielle-Dulac C, Kontodimas DC, Silvain JF, Nasr MI. 2011. Preliminary study on geographical variation of cytochrome b gene and ITS2-rDNA among populations of Rhynchophorus ferrugineus. Journal of Agricultural Science and Technology B. 1:189–197.
El-Mergawy RAAM, Faure N, Nasr MI, Avand-Faghih A, Rochat D, Silvain J-F. 2011. Mitochondrial genetic variation and invasion history of red palm weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae), in Middle-East and Mediterranean basin. International Journal of Agriculture and Biology. 13:631–637.
Faghih AA. 1996. The biology of red palm weevil, Rhynchophorus ferrugineus Oliv. (Coleopter, Curculionidae) in Savaran region (Sistan province, Iran). Applied Entomological Phytopathology. 63:16–86.
Felsenstein J. 1989. PHYLIP – Phylogeny inference package (version 3.2). Cladistics. 5:164–166.Felsenstein J, Churchill GA. 1996. A hidden Markov model approach to variation among sites
in rate of evolution. Molecular Biology and Evolution. 13:93–104.
Dow
nloa
ded
by [
Kin
g Sa
ud U
nive
rsity
] at
00:
36 2
5 O
ctob
er 2
015

ORIenTAl InSecTS 13
Gadelhak GG, Enan MR. 2005. Genetic diversity among populations of red palm weevil, Rhynchophorus ferrugineus Olivier (Coleoptera: Curculionidae), determined by random amplified polymorphic DNA polymerase chain reaction (RAPD-PCR). International Journal of Agriculture and Biology. 7:395–399.
Garner KJ, Slavicek JM. 1996. Identification and characterization of a RAPD-PCR marker for distinguishing Asian and North American gypsy moths. Insect Molecular Biology. 5:81–91.
Gómez VS, Ferry M. 1999. Attempts at biological control of date-palm pests recently found in Spain. In: Canard M, Beyssatarnaouty V, editors. Proceedings of the First Regional Symposium for Applied Biological Control in Mediterranean Countries; 25–29 October 1998; Cairo. Toulouse, France: Imprimerie Sacco, 121–125 pp.
Gosh CC. 1912. The rhinoceros beetle and red or palm weevil. Mem. Department of Agriculture, India II.
In Vidyasagar PSPV. 1998. A Brief Report on Red Palm Weevil Research in India. Accessed 15 Sep 2012. http://www.redpalmweevil.com/newlook/RPWReport/India.htm
Hall TA. 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series. 41:95–98.
Hallett RH, Crespi BJ, Borden JH. 2004. Synonymy of Rhynchophorus ferrugineus (Olivier), 1790 and R. vulneratus (Panzer), 1798 (Coleoptera, Curculionidae, Rhynchophorinae). Journal of Natural History. 38:2863–2882.
Hidayat P, Phillips TW, Ffrench-Constant RH. 1996. Molecular and morphological characters discriminate Sitophilus oryzae and S. zeamais (Coleoptera: Curculionidae) and confirm reproductive isolation. Annals of the Entomological Society of America. 89:645–652.
Ibrahim AMA, Sewify GH, Moursy EB, Hassan JM. 2011. Microbial profile from gut of red palm weevil Rhynchophorus ferrugineus Olivier (Curculionidae: Coleoptera). Egyptian Journal of Biotechnology. 37:9.
Jia S, Zhang X, Zhang G, Yin A, Zhang S, Li F, Wang L, Zhao D, Yun Q, Tala Wang J, et al. 2013. Seasonally variable intestinal metagenomes of the red palm weevil (Rhynchophorus ferrugineus). Environmental Entomology. 15(11):3020–2029.
Kehat M. 1999. Threat to date palms in Israel, Jordan and the Palestinian authority by the red palm weevil, Rhynchophorus ferrugineus. Phytoparasitica. 27:107–108.
Khiyami M, Alyamani E. 2008. Aerobic and facultative anaerobic bacteria from gut of red palm weevil (Rhynchophorus ferrugineus). African Journal of Biotechnology. 7:1432–1437.
Landry P-A, Lapointe F-J. 1996. RAPD problems in phylogenetics. Zoologica Scripta. 25:283–290.
Lefroy HM. 1906. The more important insects injurious to Indian agriculture. Calcutta: Govt. Press.
Marvaldi AE, Sequeira AS, O’Brien CW, Farrell BD. 2002. Molecular and morphological phylogenetics of weevils (Coleoptera, Curculionoidea): do niche shifts accompany diversification? Systematic Biology. 51:761–785.
Ministry of Agriculture. 2010. Available from: http://www.moa.gov.sa/.NCBI. 2013. Available from: http://www.ncbi.nlm.nih.gov/.Nirula KK. 1956. Investigation on the pest of coconut palm. Part IV. Rhynchophorus ferrugineus
F. Indian Coconut Journal. 9:229–247.Page RDM. 1996. TREEVIEW: an application to display phylogenetic trees on personal
computers. Computer Applications in the Biosciences. 12:357–358.Rugman-Jones PF, Hoddle CD, Hoddle MS, Stouthamer R. 2013. The lesser of two weevils:
molecular-genetics of pest palm weevil populations confirm Rhynchophorus vulneratus (Panzer 1798) as a valid species distinct from R. ferrugineus (Olivier 1790), and reveal the global extent of both. PLoS ONE. 8(10):e78379.
Dow
nloa
ded
by [
Kin
g Sa
ud U
nive
rsity
] at
00:
36 2
5 O
ctob
er 2
015

14 M. T. SADDeR eT Al.
Salama HS, Saker MM. 2002. DNA fingerprints of three different forms of the red palm weevil collected from Egyptian date palm orchards. Archives of Phytopathology and Plant Protection. 35:299–306.
Sambrook J, Fritsch EF, Maniatis T. 1989. Molecular cloning: a laboratory manual. Cold Spring Harbor: Cold Spring Harbor Laboratory Press.
Scataglini MA, Confalonieri VA, Lanteri AA. 2000. Dispersal of the cotton boll weevil (Coleoptera: Curculionidae) in South America: evidence of RAPD analysis. Genetica. 108:127–136.
Song H, Buhay JE, Whiting MF, Crandall KA. 2008. Many species in one: DNA barcoding overestimates the number of species when nuclear mitochondrial pseudogenes are coamplified. Proceedings of the National Academy of Sciences. 105(36):13486–13491.
Stevens J, Wall R. 1995. The use of random amplified polymorphic DNA (RAPD) analysis for studies of genetic variation in populations of the blowfly Lucilia sericata (Diptera: Calliphoridae) in southern England. Bulletin of Entomological Research. 85:549–555.
USDA-APHIS-PPQ. 2010. First U.S. detection of red palm weevil, Rhychophorus ferrugineus, in California. Letter to State Plant Regulatory Officials.
Vandewoestijne S, Baguette M. 2002. The genetic structure of endangered populations in the cranberry fritillary, Boloria aquilonaris (Lepidoptera, Nymphalidae): RAPDs vs allozymes. Heredity. 89:439–445.
Vidyasagar PSPV, Bhat SK. 1991. Pest management in coconut gardens. Journal of Plantation Crops. 19:163–182.
Wattanapongsiri A. 1966. A revision to the genera Rhynchophorus and Dynamis (Coleoptera: Curculionidae). Department of Agriculture Science Bulletin. (Bangkok). 1:1–328.
Dow
nloa
ded
by [
Kin
g Sa
ud U
nive
rsity
] at
00:
36 2
5 O
ctob
er 2
015