phosphorylation of either crh or hpr mediates binding of ccpa to the bacillus subtilis xyn cre and...
TRANSCRIPT

Article No. jmbi.1998.2492 available online at http://www.idealibrary.com on J. Mol. Biol. (1999) 286, 307±314
COMMUNICATION
Phosphorylation of Either Crh or HPr MediatesBinding of CcpA to the Bacillus subtilis xyn cre andCatabolite Repression of the xyn Operon
1 2 3
Anne Galinier , Josef Deutscher * and Isabelle Martin-Verstraete1Institut de Biologie et Chimiedes ProteÂines, CNRS UPR 412F-69367, Lyon Cedex 07France2Laboratoire de GeÂneÂtique desMicroorganismes, INRA-CNRSERS 567, F-78850Thiverval-Grignon, France3Unite de BiochimieMicrobienne, CNRS URA1300, Institut PasteurF-75724, Paris, France
*Corresponding author
15 in histidine-containing prote
E-mail address of the [email protected]
Present address: I. Martin-VerstrReÂgulation de l'Expression GeÂneÂtiqInstitut Pasteur, F-75724 Paris, Fran
Abbreviations used: CCR, carbonrepression; HPr, histidine-containinCrh, catabolite repression HPr; cre,response element; FBP, fructose 1,6PEP, phosphenol pyruvate; PTS, PEphosphotransferase system; EI, enz
0022-2836/99/070307±08 $30.00/0
Carbon catabolite repression (CCR) of several Bacillus subtilis catabolicgenes is mediated by ATP-dependent phosphorylation of Ser46 of the his-tidine-containing protein (HPr), a phosphocarrier protein of the phos-phoenolpyruvate (PEP): sugar phosphotransferase system. A recentlydiscovered HPr-like protein of B. subtilis, Crh, cannot be phosphorylatedby PEP and enzyme I but becomes phosphorylated at Ser46 by the ATP-dependent, metabolite-activated HPr kinase. Genetic data suggested thatCrh is also implicated in CCR. We here demonstrate that in a ptsH1 crh1mutant, in which Ser46 of both HPr and Crh is replaced with an alanylresidue, expression of the b-xylosidase-encoding xynB gene was comple-tely relieved from CCR. No effect on CCR could be observed in strainscarrying the crh1 allele, suggesting that under the experimental con-ditions P-Ser-HPr can substitute for P-Ser-Crh in CCR. By contrast, aptsH1 mutant was slightly relieved from CCR of xynB, indicating thatP-Ser-Crh can substitute only partly for P-Ser-HPr. Mapping experimentsallowed us to identify the xyn promoter and a catabolite responsiveelement (cre) located 229 bp downstream of the transcription start point.Using DNase I footprinting experiments, we could demonstrate that simi-lar to P-Ser-HPr, P-Ser-Crh stimulates binding of CcpA to the xyn cre.Fructose 1,6-bisphosphate was found to strongly enhance binding of theP-Ser-HPr/CcpA and P-Ser-Crh/CcpA complexes to the xyn cre, but hadno effect on binding of CcpA alone.
# 1999 Academic Press
Keywords: PEP:carbohydrate phosphotransferase system; cataboliterepression HPr; carbon catabolite repression; DNase I footprinting; xylan
metabolismThe bacterial phosphoenolpyruvate (PEP):sugarphosphotransferase system (PTS) catalyses thetransport and concomitant phosphorylation ofcarbohydrates via a protein phosphorylation chainincluding PEP-dependent phosphorylation of His-
in (HPr) by enzyme
ing author:
aete, Unite deue, CNRS URA 1129,ce.catabolite
g protein;catabolite-bisphosphate;P:carbohydrate
yme I.
I (EI). In Gram-positive bacteria, the phosphorylcarrier protein HPr is the central regulatory proteinmodulating the activity of enzymes, antitermina-tors and transcriptional activators controllinginduction and carbon catabolite repression (CCR)of numerous catabolic genes (Saier, 1996;Deutscher et al., 1997). HPr of Gram-positive bac-teria can be phosphorylated at a regulatory serineresidue (Ser46) by ATP and the HPr kinase(Deutscher & Saier, 1983; Galinier et al., 1998;Reizer et al., 1998) in addition to phosphorylationat the catalytic His15 by PEP and EI (Gassner et al.,1977). Seryl-phosphorylated HPr (P-Ser-HPr) playsa role in CCR (Deutscher et al., 1994) and inducerexclusion (Ye et al., 1994).
A new Bacillus subtilis gene encoding the HPr-
like protein Crh (catabolite repression HPr) which# 1999 Academic Press

308 Crh- and HPr-Dependent Catabolite Repression
is composed of 85 amino acid residues was discov-ered within the B. subtilis genome sequencing pro-ject (Galinier et al., 1997). Crh exhibits 45 %sequence identity when compared to HPr, but theactive-site His15 of HPr is replaced with glutaminein Crh. Crh is therefore not phosphorylated by PEPand EI, but is phosphorylated by ATP and the HPrkinase at Ser46 (Galinier et al., 1997, 1998). ATP-dependent phosphorylation of both HPr and Crh isstimulated by fructose 1,6-bisphosphate (FBP). Inmutants in which Ser46 of HPr had been replacedwith alanine (ptsH1 mutation), which prevents theATP-dependent phosphorylation of HPr, severalenzymes were found to be insensitive to CCR(Deutscher et al., 1994). By contrast, the enzymesb-xylosidase, inositol dehydrogenase and levanasewere not, or only partly, relieved from CCR in theptsH1 mutant and only the additional disruption ofthe crh gene caused complete relief from CCR(Galinier et al., 1997), suggesting that both P-Ser-HPr and seryl-phosphorylated Crh (P-Ser-Crh) areimplicated in CCR of these three enzymes.
The trans-acting protein CcpA, a member of theLacI-GalR family of repressors (Weickert & Adhya,1992), is a key regulator of CCR in B. subtilis. Itmediates CCR by binding to the cis-active operatorsequence cre (catabolite response element: Weickert& Chambliss, 1990; Henkin, 1996). An interactionof P-Ser-HPr with CcpA has been demonstratedin vitro (Deutscher et al., 1995; Jones et al., 1997)and the P-Ser-HPr/CcpA complex binds speci®-cally to the cre of the gnt and xyl operons (Fujitaet al., 1995; GoÈsseringer et al., 1997) and of theamyE gene (Kim et al., 1998). It was possible that P-Ser-Crh exerts its effect on CCR also via CcpA,since for certain operons a ccpA mutant and aptsH1 crh double mutant exhibited the same relieffrom CCR.
To demonstrate the presumed interactionbetween CcpA and P-Ser-Crh and binding of thecomplex to cre sequences, we carried out exper-iments with the promoter region of the b-xylosi-dase-encoding gene (Hastrup, 1988), since in aptsH1 mutant b-xylosidase activity was stronglyrepressed by glucose, whereas it was completelyrelieved from CCR in a ptsH1 crh double mutant(Galinier et al., 1997). Together with an extracellu-lar b-xylanase and intracellular D-xylose isomeraseand xylulokinase, b-xylosidase is involved in xylanutilization. Due to the lack of a xylose-speci®c per-mease, B. subtilis cannot grow on xylose as the solecarbon source (Lindner et al., 1994), but slowxylose uptake seems to be catalysed by the AraEprotein (Krispin & Allmansberger, 1998). However,B. subtilis can transport xylosides produced fromxylan degradation that are intracellularly hydro-lysed to form xylose. The ynaJ gene and the adja-cent xynB gene (Kunst et al., 1997) were thought toform an operon and to encode a xyloside transpor-ter and b-xylosidase. To con®rm that the B. subtilisxynB gene codes for the enzyme b-xylosidase, weconstructed a xynB mutant strain by integrating
plasmid pRC24 containing a PCR-ampli®edinternal xynB fragment fused to the Escherichia colilacZ gene into the xynB locus. The resulting inte-grant QB7146 carrying a disrupted xynB geneexhibited a xylose-inducible b-xylosidase activityof only six units per milligram of protein comparedto 2790 units/mg protein measured for the corre-sponding wild-type strain. In the absence of xylose,similar low b-xylosidase activities were determinedfor the wild-type strain (31 units/mg protein) andfor QB7146 (four units/mg protein). These resultscon®rm the fact that xynB encodes the enzymeb-xylosidase.
To localise the xyn promoter, expression studieswere carried out with QB7146, described aboveand with strain QB7144, which carries a transcrip-tional ynaJ0-lacZ fusion inserted at the amyE locus.This ynaJ0-lacZ fusion contains fragment A (ÿ279,�380; Figure 1) of the ynaJ gene. Since the syn-thesis of b-xylosidase has been reported to be indu-cible by xylose and repressed by glucose (Galinieret al., 1997), bacteria were grown in CSK mediumor in CSK medium containing either 0.2 % (w/v)xylose or 0.2 % xylose and 1 % (w/v) glucose.Expression of both lacZ fusions was low in theabsence of xylose but was increased 66-fold forynaJ0-lacZ and 40-fold for xynB0-lacZ when 0.2 %xylose was added to the growth medium (Table 1).The addition of 1 % glucose strongly repressed theexpression of both transcriptional fusions. Theobserved coregulation of ynaJ and xynB expressionfurther supported the assumption that these twogenes form an operon. The location of the promo-ter was con®ned by using transcriptional ynaJ0-lacZfusions that carried varying 50 deletions comparedto the ynaJ0-lacZ fusion with fragment A describedabove. These fusions were integrated as singlecopies at the amyE locus of B. subtilis 168. Thestrain containing the fusion with fragment B (ÿ49,�380; Figure 1) remained inducible by xylose,whereas the strain containing the fusion with frag-ment C (ÿ2, �380) could no longer be induced(Table 1). According to this result, the promotermust be located between the 50 ends of fragmentsB and C, i.e. between ÿ49 and ÿ2 (Figure 1).
In addition, primer-extension experiments wereperformed according to Sambrook et al. (1989) inorder to localize the start point of the presumedynaJ xynB transcript. Primer IM118 (50-CCCCTTCCATCCATATTAGTAAGATCTTATC-30) corresponding to positions �84 to �55 (seeFigure 1) was hybridized with 8 mg of RNAextracted from the wild-type strain 168 aftergrowth in CSK medium (Martin-Vestraete et al.,1990) or in CSK medium containing either 0.2 %xylose or 0.2 % xylose plus 1 % glucose (data notshown). The obtained position of the transcriptionstart point is indicated with an asterisk in Figure 1.A sequence representing a potential promoter (50-TTGAAA-30 separated by 17 bp from the sequence50- TAATAT-30) is present upstream from the tran-scriptional start point within the region ÿ49 to ÿ2determined by the promoter mapping described
above. Primer extension was observed only with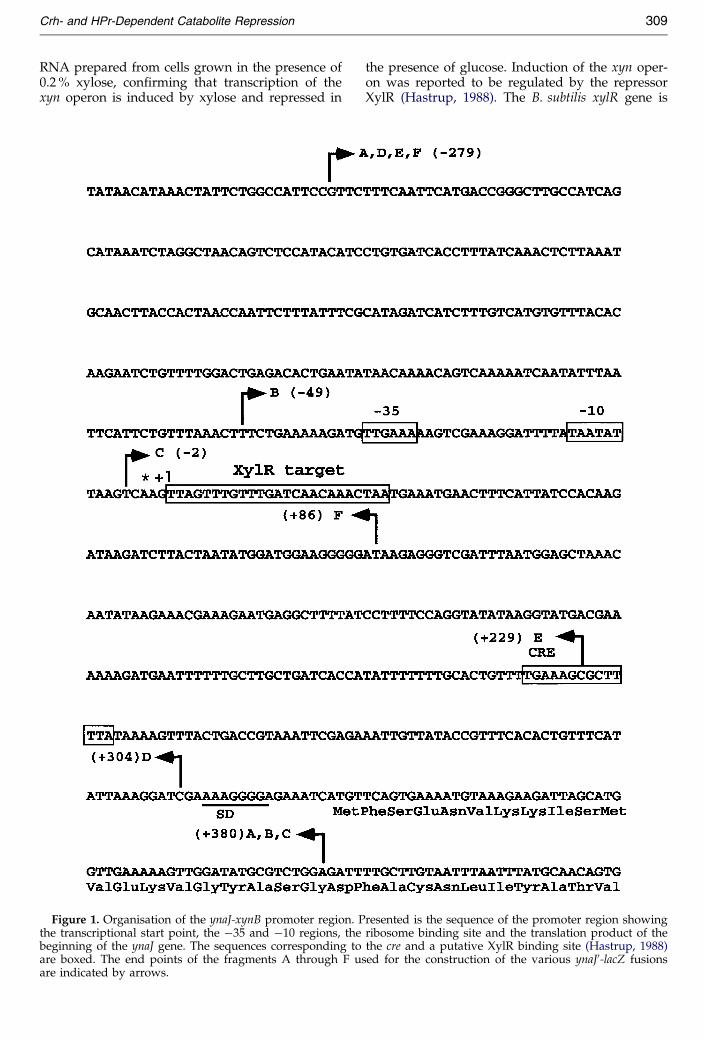
Crh- and HPr-Dependent Catabolite Repression 309
RNA prepared from cells grown in the presence of0.2 % xylose, con®rming that transcription of the
xyn operon is induced by xylose and repressed inFigure 1. Organisation of the ynaJ-xynB promoter region. Pthe transcriptional start point, the ÿ35 and ÿ10 regions, thebeginning of the ynaJ gene. The sequences corresponding toare boxed. The end points of the fragments A through F usare indicated by arrows.
the presence of glucose. Induction of the xyn oper-on was reported to be regulated by the repressor
XylR (Hastrup, 1988). The B. subtilis xylR gene isresented is the sequence of the promoter region showingribosome binding site and the translation product of thethe cre and a putative XylR binding site (Hastrup, 1988)ed for the construction of the various ynaJ0-lacZ fusions

Table 1. Regulation of the expression of a xynB0-lacZ fusion and different ynaJ0-lacZ fusions carrying fragments Athrough F (Figure 1)
b-Galactosidase activity (Units/mg of protein)b
Fusionsa Plasmid CSK CSK Xyl CSK Xyl Glu
pA ynaJ0-lacZ� pRC21 9 598 5xynB0-lacZ� pRC24 6 238 3.5A. 50 DeletionspB ynaJ0-lacZ� pRC25 9 555 4.5pC ynaJ0-lacZ� pRC26 4 3.5 0.5B. 30 DeletionspDynaJ0-lacZ� pRC28 25 1990 13pEynaJ0-lacZ� pRC22 6 380 65pFynaJ0-lacZ� pRC27 5 215 47
a Transcriptional fusions with varying fragments of ynaJ and lacZ integrated in amyE by double crossing-over were constructedusing the vector pAC6 (StuÈ lke et al., 1997). pAC6 carries the pC194 chloramphenicol resistance gene and a lacZ gene without a pro-moter located between two fragments of the B. subtilis amyE gene. PCR fragments corresponding to fragments A through D and F(Figure 1) and carrying an EcoRI and a BamHI restriction site at the 50 and 30 ends, respectively, were inserted into pAC6. FragmentE was obtained by cutting PCR product A with EcoRI and Eco47III, and inserting the resulting fragment into pAC6 cut with EcoRIand SmaI. The xynB0-lacZ transcriptional fusion was integrated in the chromosomal xyn operon by single crossing-over using plasmidpRC24, a derivative of pDIA5307 (Calogero et al., 1994), containing a SmaI/SphI internal xynB fragment fused to lacZ.
b Speci®c activities of b-galactosidase were determined in extracts prepared from exponentially growing cells (A600 � 0.7-1). Themean values of at least three independent experiments are presented. Cells were grown in CSK medium which is C minimal med-ium supplemented with potassium succinate (6 g/l) and potassium glutamate (8 g/l) (Martin-Verstraete et al., 1990) or on CSK med-ium containing 0.2 % xylose or 0.2 % xylose plus 1 % glucose. The method of Miller was used for the determination ofb-galactosidase activity.
310 Crh- and HPr-Dependent Catabolite Repression
located just downstream of ynaJ and xynB, but istranscribed in the opposite direction (Kunst et al.,1997). The xylR gene is followed by the xylABoperon, with xylA encoding xylose isomerase andxylB encoding xylulose kinase. Induction of thexylAB operon has been demonstrated to be regu-lated by XylR and an operator site has beenidenti®ed: TTAGTTTGTTTGGGCAACAAACTAA(Kreuzer et al., 1989). A putative XylR binding site(Hastrup, 1988) identical with the xylAB operatorsite (with the exception of the three G located inthe middle) is situated between positions �4 and�28 (boxed in Figure 1) and might be involved inmodulating ynaJ and xynB expression in responseto the presence of intracellular xylose formed byxyloside degradation.
CCR in Gram-positive bacteria requires anothercis-active site (cre) usually located in front or at thebeginning of an operon (Weickert & Chambliss,1990). To localize the cre regulating CCR of theB. subtilis xyn operon, a series of transcriptionalfusions was constructed. For this purpose, frag-ment A (ÿ279, �380) and fragments carrying vary-ing 30 deletions of A were fused to the lacZ genelacking its own promoter. These fusions wereinserted as single copies into the amyE locus andb-galactosidase activities were measured aftergrowth of the resulting strains in CSK medium orin CSK medium containing either 0.2 % xylose or0.2 % xylose plus 1 % glucose (Table 1). While thefusions with fragments A (ÿ279, �380) and D(ÿ279, �304) were 120-fold repressed by glucose,the fusions with fragments E (ÿ279, �229) and F(ÿ279, �86) were repressed only ®ve- to sixfold.According to this result, the cre implicated in CCRof the xyn operon must be located between the30 ends of fragments E and D, i.e. between posi-
tions �229 and �304. A cre-like sequenceTGAAAGCGCTTTTA extending from position�224 to �237 (downstream from the transcrip-tional start site) was indeed found in this region.
Galinier et al. (1997) had suggested that phos-phorylation of both HPr and Crh at Ser46 might benecessary for CCR of the B. subtilis b-xylosidase-encoding xynB gene. To study the proposedrequirement of Crh phosphorylation for CCR ofxynB, a chromosomal crh1 mutant (encodingCrhS46A) was constructed. Plasmid pRC33, apHT315 derivative (Arantes & Lereclus, 1991) con-taining the crh1 allele and yvcN, the gene locateddownstream from crh, disrupted by insertion of akanamycin resistance cassette at the unique BstBIsite, was used to replace the wild-type crh gene bythe crh1 allele by cotransformation with the anti-biotic resistance cassette. By using this technique, acrh1 (QB7157) and a ptsH1 crh1 double mutant(QB7156) carrying the ynaJ0-lacZ fusion with frag-ment A (Figure 1) were constructed. The presenceof the crh1 allele in these strains was con®rmed bysequencing appropriate PCR products obtainedwith chromosomal DNA of these strains. Therepressive effect of glucose on the expression of theynaJ0-lacZ fusion in the crh1 and the ptsH1 crh1strains was compared to that obtained for a ccpAand a ptsH1 strain. Strong glucose repression wasobserved for a ptsH1 or a crh1 mutant strain (40-fold and 100-fold repression, respectively, com-pared to 110-fold repression for the wild-typestrain). By contrast, in the ptsH1 crh1 mutantQB7156, expression of the ynaJ0-lacZ fusion wasrelieved from glucose repression to an extent simi-lar to that observed for the ccpA mutant QB7147(1050 and 1200 units under inducing conditions,650 and 700 units under repressing conditions,respectively). In summary, these results show that
(i) CCR of the B. subtilis ynaJ gene is mediated via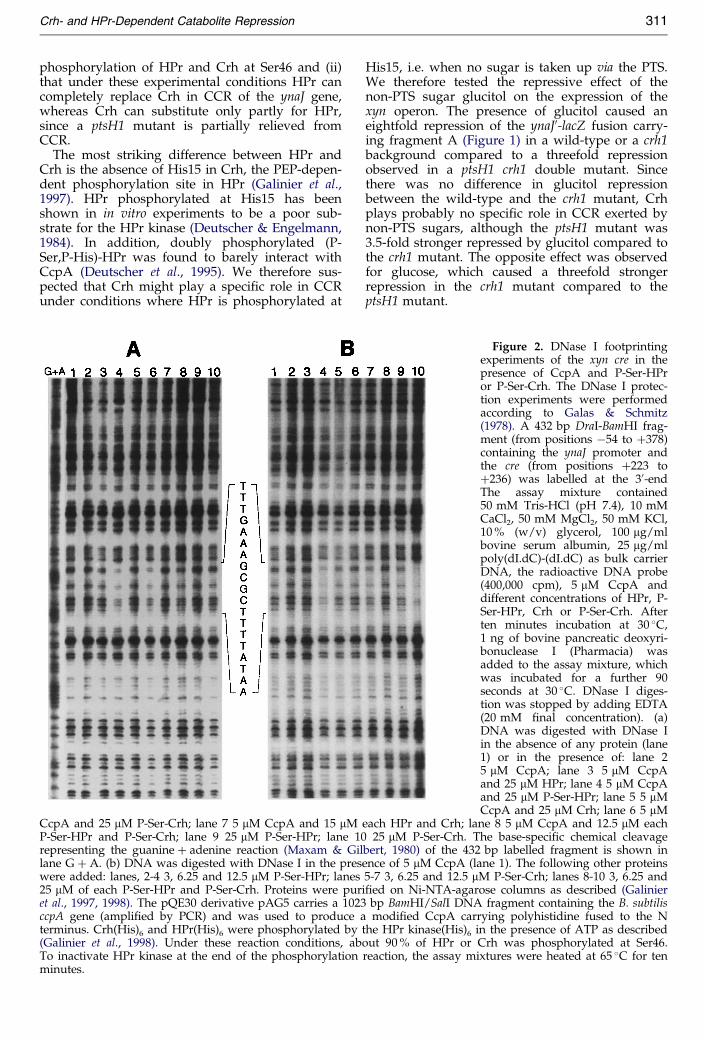
Crh- and HPr-Dependent Catabolite Repression 311
phosphorylation of HPr and Crh at Ser46 and (ii)that under these experimental conditions HPr cancompletely replace Crh in CCR of the ynaJ gene,whereas Crh can substitute only partly for HPr,since a ptsH1 mutant is partially relieved fromCCR.
The most striking difference between HPr andCrh is the absence of His15 in Crh, the PEP-depen-dent phosphorylation site in HPr (Galinier et al.,1997). HPr phosphorylated at His15 has beenshown in in vitro experiments to be a poor sub-strate for the HPr kinase (Deutscher & Engelmann,1984). In addition, doubly phosphorylated (P-Ser,P-His)-HPr was found to barely interact withCcpA (Deutscher et al., 1995). We therefore sus-pected that Crh might play a speci®c role in CCR
under conditions where HPr is phosphorylated atCcpA and 25 mM P-Ser-Crh; lane 7 5 mM CcpA and 15 mMP-Ser-HPr and P-Ser-Crh; lane 9 25 mM P-Ser-HPr; lane 1representing the guanine � adenine reaction (Maxam & Gilane G � A. (b) DNA was digested with DNase I in the preswere added: lanes, 2-4 3, 6.25 and 12.5 mM P-Ser-HPr; lanes25 mM of each P-Ser-HPr and P-Ser-Crh. Proteins were puret al., 1997, 1998). The pQE30 derivative pAG5 carries a 102ccpA gene (ampli®ed by PCR) and was used to produceterminus. Crh(His)6 and HPr(His)6 were phosphorylated by(Galinier et al., 1998). Under these reaction conditions, abTo inactivate HPr kinase at the end of the phosphorylationminutes.
His15, i.e. when no sugar is taken up via the PTS.We therefore tested the repressive effect of thenon-PTS sugar glucitol on the expression of thexyn operon. The presence of glucitol caused aneightfold repression of the ynaJ0-lacZ fusion carry-ing fragment A (Figure 1) in a wild-type or a crh1background compared to a threefold repressionobserved in a ptsH1 crh1 double mutant. Sincethere was no difference in glucitol repressionbetween the wild-type and the crh1 mutant, Crhplays probably no speci®c role in CCR exerted bynon-PTS sugars, although the ptsH1 mutant was3.5-fold stronger repressed by glucitol compared tothe crh1 mutant. The opposite effect was observedfor glucose, which caused a threefold strongerrepression in the crh1 mutant compared to the
ptsH1 mutant.Figure 2. DNase I footprintingexperiments of the xyn cre in thepresence of CcpA and P-Ser-HPror P-Ser-Crh. The DNase I protec-tion experiments were performedaccording to Galas & Schmitz(1978). A 432 bp DraI-BamHI frag-ment (from positions ÿ54 to �378)containing the ynaJ promoter andthe cre (from positions �223 to�236) was labelled at the 30-endThe assay mixture contained50 mM Tris-HCl (pH 7.4), 10 mMCaCl2, 50 mM MgCl2, 50 mM KCl,10 % (w/v) glycerol, 100 mg/mlbovine serum albumin, 25 mg/mlpoly(dI.dC)-(dI.dC) as bulk carrierDNA, the radioactive DNA probe(400,000 cpm), 5 mM CcpA anddifferent concentrations of HPr, P-Ser-HPr, Crh or P-Ser-Crh. Afterten minutes incubation at 30 �C,1 ng of bovine pancreatic deoxyri-bonuclease I (Pharmacia) wasadded to the assay mixture, whichwas incubated for a further 90seconds at 30 �C. DNase I diges-tion was stopped by adding EDTA(20 mM ®nal concentration). (a)DNA was digested with DNase Iin the absence of any protein (lane1) or in the presence of: lane 25 mM CcpA; lane 3 5 mM CcpAand 25 mM HPr; lane 4 5 mM CcpAand 25 mM P-Ser-HPr; lane 5 5 mMCcpA and 25 mM Crh; lane 6 5 mM
each HPr and Crh; lane 8 5 mM CcpA and 12.5 mM each0 25 mM P-Ser-Crh. The base-speci®c chemical cleavagelbert, 1980) of the 432 bp labelled fragment is shown inence of 5 mM CcpA (lane 1). The following other proteins5-7 3, 6.25 and 12.5 mM P-Ser-Crh; lanes 8-10 3, 6.25 andi®ed on Ni-NTA-agarose columns as described (Galinier3 bp BamHI/SalI DNA fragment containing the B. subtilisa modi®ed CcpA carrying polyhistidine fused to the Nthe HPr kinase(His)6 in the presence of ATP as describedout 90 % of HPr or Crh was phosphorylated at Ser46.reaction, the assay mixtures were heated at 65 �C for ten

Figure 3. DNase I footprinting of the xyn cre withCcpA and P-Ser-HPr or P-Ser-Crh in the presence ofFBP. DNase I protection experiments were carried outas described for Figure 2. DNA was digested withDNase I in the presence of 10 mM FBP and 5 mM CcpA(lane 1). The following other proteins were added: lanes2 and 3 3 and 6.25 mM P-Ser-HPr; lanes 4 and 5 3 and6.25 mM P-Ser-Crh; lanes 6 and 7 3 and 6.25 mM eachP-Ser-HPr and P-Ser-Crh; lane 8 15 mM HPr; lane 915 mM Crh; lane 10 15 mM each HPr and Crh. The base-speci®c chemical cleavage representing the guanine -� adenine reaction (Maxam & Gilbert, 1980) of the432 bp labelled fragment is shown in lane G � A.
312 Crh- and HPr-Dependent Catabolite Repression
P-Ser-HPr has been demonstrated to interactin vitro with CcpA (Deutscher et al., 1995; Joneset al., 1997) and the resulting protein complexbinds speci®cally to the cre sequence of the gluco-nate and xylose operons (Fujita et al., 1995;GoÈsseringer et al., 1997) and of the amyE gene (Kimet al., 1998). To test whether P-Ser-Crh can alsointeract with CcpA and allow speci®c binding ofCcpA to the xyn cre, we performed DNase I foot-printing experiments. A 432 bp PCR fragment con-taining the xyn promoter (from positions ÿ54 to�378; Figure 1) was 30-end-labelled with[a-32P]dATP. The results of the DNase I footprint-ing experiments are presented in Figure 2. Inter-action of CcpA (5 mM) with P-Ser-HPr (25 mM) orP-Ser-Crh (25 mM) causes speci®c recognition ofthe xyn cre (Figure 2(a), lanes 4 and 6). This speci®crecognition was not observed when CcpA, CcpAand HPr, CcpA and Crh, P-Ser-HPr, or P-Ser-Crhwere present in the assay mixture (Figure 2(a),lanes 2, 3, 5, 9 and 10, respectively). The regionprotected in the experiments with CcpA and P-Ser-HPr or P-Ser-Crh (TTTGAAAGCGCTTTTATAA)overlaps the cre- like sequence (in bold letters)identi®ed between positions �224 and �237(Figure 1). Under the experimental conditions, noadditional speci®c protection could be detected.When both proteins, P-Ser-HPr (12.5 mM) and P-Ser-Crh (12.5 mM), were added together to a CcpA-containing assay mixture (Figure 2(a), lane 8), aclearly weaker protection was observed as com-pared to that obtained with only 25 mM P-Ser-HPror only 25 mM P-Ser-Crh (Figure 2(a), lanes 4 and6), indicating that there is no synergetic effect ofP-Ser-HPr and P-Ser-Crh on binding of CcpA tothe xyn cre. To study the effect of each protein inmore detail, footprinting experiments were carriedout in the presence of CcpA and various concen-trations of P-Ser-HPr, P-Ser-Crh, and a mixture ofboth (Figure 2(b)). A protection of the xyn cresequence was obtained in the presence of 12.5 mMP-Ser-HPr (Figure 2(b), lane 4), whereas a clearlyweaker protection was obtained in the presence of12.5 mM P-Ser-Crh (Figure 2(b), lane 7). These datasuggest that P-Ser-HPr is slightly more ef®cientthan P-Ser-Crh in stimulating binding of CcpA tothe xyn cre sequence. When P-Ser-HPr and P-Ser-Crh were added together at a ®nal concentration of3 or 6.25 mM each (i.e. 6 or 12.5 mM total phos-phorylated proteins, respectively) no signi®cantprotection was observed (Figure 2(b), lanes 8 and9, respectively). This result con®rmed that there isno synergetic effect of P-Ser-HPr and P-Ser-Crh onbinding of CcpA to the xyn cre. In the presence of25 mM P-Ser-HPr and 25 mM P-Ser-Crh a signi®-cant protection of the xyn cre sequence wasobserved (Figure 2(b), lane 10). However, toobserve a protective effect, we had to use fairlyhigh concentrations of P-Ser-HPr and/or P-Ser-Crh, suggesting that other components might berequired to further stimulate binding of the P-Ser-HPr/CcpA or P-Ser-Crh/CcpA complex to the xyn
cre.Since Deutscher et al. (1995) had found that thepresence of FBP enhances the speci®c interaction ofCcpA from Bacillus megaterium with B. subtilisP-Ser-HPr , it was possible that FBP could in¯u-ence the binding of the P-Ser-HPr/CcpA complexto the cre. We therefore carried out footprintingexperiments in the presence of FBP (Figure 3).Addition of 10 mM FBP enhanced the binding ofCcpA to the xyn cre in the presence of 6.25 mMP-Ser-HPr (Figure 3, lane 3) or 6.25 mM P-Ser-Crh(Figure 3, lane 5). A clear protection of the xyn crewas observed in the presence of 3 mM P-Ser-HPr,whereas no protection was detected with 3 mM
P-Ser-Crh (Figure 3, lanes 2 and 4, respectively).
Crh- and HPr-Dependent Catabolite Repression 313
These results suggest that, in the presence of FBP,P-Ser-HPr is more ef®cient than P-Ser-Crh in sti-mulating binding of CcpA to the the xyn cre. Fur-thermore, FBP does not induce a synergy beetweenP-Ser-HPr and P-Ser-Crh. The protective effect inthe presence of 3 or 6.25 mM P-Ser-HPr (Figure 3,lanes 2 and 3, respectively) was not markedlyincreased by the addition of an equivalent amountof P-Ser-Crh (Figure 3, lanes 6 and 7). Binding ofCcpA to the xyn cre in the absence of other proteinsor in the presence of HPr, Crh, or HPr and Crhwas not affected by FBP (Figure 3, lanes 1 and 8 to10, respectively). No additional speci®cally pro-tected region could be observed in the presence ofFBP.
In contrast to the B. subtilis gnt and mtl operons,which were found to be almost completely relievedfrom CCR in ptsH1 mutants (Deutscher et al.,1994), the lev, iol and xyn operons were onlyslightly relieved from CCR (Galinier et al., 1997).The results presented above suggest that CCR ofthe latter operons is mediated by the concertedaction of P-Ser-HPr and P-Ser-Crh. Similar to accpA mutant, a chromosomal ptsH1 crh1 doublemutant, in which Ser46 of both HPr and Crh wasreplaced with an alanyl residue, exhibited no CCRof the xyn operon. Like P-Ser-HPr, P-Ser-Crh seemsto exert its effect on CCR by interacting withCcpA. Both seryl-phosphorylated proteins stimu-lated binding of CcpA to the xyn cre identi®ed infront of the coding sequence between positions�224 and �237 (Figure 1), although the effect ofP-Ser-Crh on CcpA binding seemed to be weakerthan the effect of P-Ser-HPr. Interestingly, FBPenhanced binding of the P-Ser-HPr/CcpA andP-Ser-Crh/CcpA complexes to the xyn cre similaras had been reported for the amyE cre (Kim et al.,1998). FBP seems therefore to play a dual role inCCR of the xyn operon: high levels of intracellularFBP seem (i) to increase the amount of P-Ser-HPrand P-Ser-Crh by speci®cally stimulating the HPrkinase (Galinier et al., 1998); and (ii) to enhancebinding of the P-Ser-HPr/CcpA and P-Ser-Crh/CcpA complexes to the xyn cre.
Acknowledgements
This research was supported by the MinisteÁre del'Education Nationale de la Recherche et de la Technolo-gie, the Centre National de la Recherche Scienti®que, theInstitut National de la Recherche Agronomique, theInstitut Pasteur, the Universite de Lyon and the Univer-site Paris 7. We are grateful to G. Rapoport, in whoselaboratory part of this work was carried out, for continu-ous encouragement and critical reading of the manu-script. We are thankful to C. van Herrewege, A. Boschand C. Dugast for their expert assistance in preparation
of the manuscript.References
Arantes, O. & Lereclus, D. (1991). Construction of clon-ing vectors for Bacillus thuringiensis. Gene, 108, 115-119.
Calogero, S., Gardan, R., Glaser, P., Schweizer, J.,Rapoport, G. & DeÂbarbouilleÂ, M. (1994). RocR, anovel regulatory protein controlling arginine utiliz-ation in Bacillus subtilis, belongs to the NtrC/NifAfamily of transcriptional activators. J. Bacteriol. 176,1234-1241.
Deutscher, J. & Engelmann, R. (1984). Puri®cation andcharacterization of an ATP-dependent proteinkinase from Streptococcus faecalis. FEMS Microbiol.Letters, 23, 157-162.
Deutscher, J., Fischer, C., Charrier, V., Galinier, A.,Lindner, C., Darbon, E. & Dossonnet, V. (1997).Regulation of carbon metabolism in Gram-positivebacteria by protein phosphorylation. Folia Microbiol.42, 171-178.
Deutscher, J., KuÈ ster, E., Bergstedt, U., Charrier, V. &Hillen, W. (1995). Protein kinase-dependant HPr/CcpA interaction links glycolytic activity to carboncatabolite repression in Gram-positive bacteria. Mol.Microbiol. 15, 1049-1053.
Deutscher, J., Reizer, J., Fischer, C., Galinier, A., Saier,M. H., Jr & Steinmetz, M. (1994). Loss of proteinkinase-catalyzed phosphorylation of HPr, a phos-pho-carrier of the phosphostransferase system, bymutation of the ptsH gene confers catabolite repres-sion resistance to several catabolic genes of Bacillussubtilis. J. Bacteriol. 176, 3336-3344.
Deutscher, J. & Saier, M. H., Jr (1983). ATP-dependentprotein kinase-catalyzed phosphorylation of a serylresidue in HPr, a phosphate carrier protein of thephosphotransferase system in Streptococcus pyogenes.Proc. Natl Acad. Sci. USA, 80, 6790-6794.
Fujita, Y., Miwa, Y., Galinier, A. & Deutscher, J. (1995).Speci®c recognition of the Bacillus subtilis gnt cis-act-ing catabolite-responsive element by a protein com-plex formed between CcpA and seryl-phosphorylated HPr. Mol. Microbiol. 17, 953-960.
Galas, D. G. & Schmitz, A. (1978). DNAse footprinting:a simple method for detection of protein-DNAbinding speci®city. Nucl. Acids Res. 5, 3157-3170.
Galinier, A., Haiech, J., Kilhoffer, M.-C., Jaquinod, M.,StuÈ lke, J., Deutscher, J. & Martin-Verstraete, I.(1997). The Bacillus subtilis crh gene encodes a HPr-like protein involved in carbon catabolite repres-sion. Proc. Natl Acad. Sci. USA, 94, 8439-8444.
Galinier, A., Kravanja, M., Engelmann, R.,Hengstenberg, W., Kilhoffer, M. C., Deutscher, J. &Haiech, J. (1998). New protein kinase and proteinphosphatase families mediate signal transduction inbacterial catabolite repression. Proc. Natl Acad. Sci.USA, 95, 1823-1828.
Gassner, M., Stehlik, D., Schrecker, O., Hengstenberg,W., Maurer, W. & RuÈ terjans, H. (1977). The phos-phoenolpyruvate-dependent phosphotransferase sys-tem of Staphylococcus aureus 2. 1H and 31P nuclear-magnetic-resonance studies on the phosphocarrierprotein HPr, phosphohistidines and phosphorylatedHPr. Eur. J. Biochem. 75, 287-296.
GoÈsseringer, R., KuÈ ster, E., Galinier, A., Deutscher, J. &Hillen, W. (1997). Cooperative and non-cooperativeDNA binding modes of catabolite control proteinCcpA from Bacillus megaterium result from sensing
two different signals. J. Mol. Biol. 266, 665-676.
3484-3492.
314 Crh- and HPr-Dependent Catabolite Repression
Hastrup, S. (1988). Analysis of the Bacillus subtilis xyloseregulon. In Genetic and Biotechnology of Bacilli(Ganesan, A. T. & Hoch, J. A., eds), vol. 2, pp. 79-83, Academic Press, New York.
Henkin, T. M. (1996). The role of CcpA transcriptionalregulator in carbon metabolism in Bacillus subtilis.FEMS Microbiol. Letters, 135, 9-15.
Jones, E. J., Dossonnet, V., KuÈ ster, E., Hillen, W.,Deutscher, J. & Klevit, R. (1997). Binding of the cat-abolite repressor protein CcpA to its DNA target isregulated by phosphorylation of its corepressorHPr. J. Biol. Chem. 272, 26530-26535.
Kim, J.-H., Voskuil, M.-I. & Chambliss, G. H. (1998).NADP, corepressor for the Bacillus catabolite con-trol protein CcpA. Proc. Natl Acad. Sci. USA, 95,9590-9595.
Kreuzer, P., GaÈrtner, D., Allmansberger, R. & Hillen, W.(1989). Identi®cation and sequence analysis of theBacillus subtilis W23 xylR gene and xyl operator.J. Bacteriol. 171, 3840-3845.
Krispin, O. & Allmansberger, R. (1998). The Bacillus sub-tilis AraE protein displays a broad substrate speci-®city for several different sugars. J. Bacteriol. 180,3250-3252.
Kunst, F., Ogasawara, N., Moszer, I., Albertini, A. M.,Alloni, G., Azevedo, V., Bertero, M. G., Bessieres,P., Bolotin, A., Borchert, S., Borriss, R., BoursieÁr, L.,Brans, A., Braun, M., Brigriell, S. C., et al. (1997).The complete genome sequence of the Gram-posi-tive bacterium Bacillus subtilis. Nature, 390, 249-256.
Lindner, C., StuÈ lke, J. & Hecker, M. (1994). Regulationof xylanolytic enzymes in Bacillus subtilis. Micro-biology, 140, 753-757.
Martin-Verstraete, I., DeÂbarbouilleÂ, M., Klier, A. &Rapoport, G. (1990). Levanase operon of Bacillussubtilis includes a fructose-speci®c phosphotransfer-
(R
ase system regulating the expression of the operon.J. Mol. Biol. 214, 657-671.
Maxam, A. M. & Gilbert, W. (1980). Sequencing end-labelled DNA with base-speci®c chemical cleavages.Methods Enzymol. 65, 499-560.
Miller, J. H. (1972). Experiments in Molecular Genetics, pp.352-355, Cold Spring Harbor Laboratory Press,Plainview, NY.
Reizer, J., Hoischen, C., Titgemeyer, F., Rivolta, C.,Rabus, R., StuÈ lke, J., Karamata, D., Saier, M. H., Jr& Hillen, W. (1998). A novel protein kinase thatcontrols carbon catabolite repression in bacteria.Mol. Microbiol. 27, 1157-1169.
Saier, M. H., Jr (1996). Cyclic AMP-independent catabo-lite repression in bacteria. FEMS Microbiol. Letters,138, 97-103.
Sambrook, J., Fritsch, E. F. & Maniatis, T. (1989). Molecu-lar Cloning: A Laboratory Manual, 2nd edit., ColdSpring Harbor Laboratory Press, Plainview, NY.
StuÈ lke, J., Martin-Verstraete, I., Zagorec, M., Rose, M.,Klier, A. & Rapoport, G. (1997). Induction of theBacillus subtilis ptsGHI operon by glucose is con-trolled by a novel antiterminator, GlcT. Mol. Micro-biol. 25, 65-78.
Weickert, M. J. & Adhya, S. (1992). A family of bacterialregulators homologous to Gal and Lac repressors.J. Biol. Chem. 267, 15869-15874.
Weickert, M. J. & Chambliss, G. H. (1990). Site-directedmutagenesis of a catabolite repression operatorsequence in Bacillus subtilis. Proc. Natl Acad. Sci.USA, 87, 6238-6242.
Ye, J. J., Neal, J. W., Cui, X., Reizer, J. & Saier, M. H., Jr(1994). Regulation of the glucose:H� symporter bymetabolite-activated ATP-dependent phosphoryl-ation of HPr in Lactobacillus brevis. J. Bacteriol. 176,
eceived 4 Nove
Edited by I. B. Holland
mber 1998; accepted 16 December 1998)