participation of y89 and y97 in the … participation of y89 and y97 in the conjugating activity of...
TRANSCRIPT

A r t i c l e
PARTICIPATION OF Y89 AND Y97IN THE CONJUGATING ACTIVITYOF Drosophila melanogasterGLUTATHIONE S-TRANSFERASED3 (DmGSTD3)Kithalakshmi Vignesvaran and Zazali AliasInstitute of Biological Sciences, Faculty of Science, University of Malaya,Kuala Lumpur, Malaysia
Drosophila melanogaster glutathione S-transferase D3 (DmGSTD3) has ashorter amino acid sequence as compared to other GSTs known in the fruitflies. This is due to the 15 amino acid N-terminal truncation in whichnormally active amino acid residue is located. The work has made use ofhomology modeling to visualize the arrangement of amino acid side chainsin the glutathione (GSH) substrate cavity. The identified amino acids werethen replaced with amino acids without functional groups in the sidechains and the mutants were analyzed kinetically. Homology modelingrevealed that the side chains of Y89 and Y97 were shown facing toward thesubstrate cavity proposing their possible role in catalyzing the conjugation.Y97A and Y89A GSH gave large changes in Km (twofold increase), Vmax
(fivefold reduction), and Kcat/Km values for GSH suggesting theirsignificant role in the conjugation reaction. The replacement at eitherpositions has not affected the affinity of the enzyme toward1-chloro-2,4-dinitrobenzene as no significant change in values of Kmax wasobserved. The replacement, however, had significantly reduced the catalyticefficiency of both mutants with (Kcat/Km)GSH and (Kcat/Km)CDNB of eight-and twofold reduction. The recombinant DmGSTD3 has shown no activitytoward 1,2-dichloro-4-nitrobenzene, 2,4-hexadienal, 2,4-heptadienal,p-nitrobenzyl chloride, ethacrynic acid, and sulfobromophthalein.
Grant sponsor: University of Malaya Postgraduate Research Fund; Grant number: PPP: PV091/2011A; Grantsponsor: Ministry of Higher Education under the Fundamental Research Grant; Grant number: FRGS: FP052-2014A.Correspondence to: Zazali Alias, Institute of Biological Sciences, Faculty of Science, University of Malaya, KualaLumpur 50603, Malaysia. E-mail: [email protected]
ARCHIVES OF INSECT BIOCHEMISTRY AND PHYSIOLOGY, Vol. 92, No. 3, 210–221 (2016)Published online in Wiley Online Library (wileyonlinelibrary.com).C© 2016 Wiley Periodicals, Inc. DOI: 10.1002/arch.21332

Y89 and Y97 in the Conjugating Activity of DmGSTD3 � 211
Therefore, it was evident that DmGSTD3 has made use of distal aminoacids Y97 and Y89 for GSH conjugation. C© 2016 Wiley Periodicals, Inc.
Keywords: detoxification; glutathione S-transferase; site-directed mutagenesis;Drosophila melanogaster
INTRODUCTION
The glutathione S-transferases (GSTs, EC 2.5.1.18) consist of an enzyme family with mul-tiple functionalities. GSTs catalyze the nucleophilic attack by the thiol group of reducedglutathione (GSH) on a wide range of electrophilic substrates. GSTs play importantroles in the development of resistance to a variety of exogenous xenobiotics, such aschemotherapeutic drugs (Hayes and Pulford, 1995), chemical carcinogens (Coles andKetterer, 1990), herbicides (Edwards et al., 2000), and insecticides (Clark, 1989; Yu, 1996;Enayati et al., 2005). GSTs are cytosolic, microsomal, or mitochondrial enzymes. Thecytosolic proteins have extensive ligand-binding properties in addition to their catalyticrole in detoxification. GSTs catalyze the conjugation of activated xenobiotics to reducedGSH (Vontas et al., 2001) GSTs have emerged as promising therapeutic targets due totheir isoenzyme-specific overexpression in different tumors and may play a role in theetiology of other diseases (Travensolo et al., 2008). GST can also bind to hydrophobiccompounds, which are not substrates associated with sequestration, storage and trans-portation of drugs, hormones and other metabolites (Nair and Choi, 2011). GSTs alsohave GSH peroxidase activity under oxidative stress and are implicated in physiologicalroles such as signal transduction, cell proliferation, differentiation, and apoptosis; for ex-ample, GST plays a role in regulation of the c-Jun N- terminal kinase signal transductionpathway (Udomsinprasert et al., 2005).
There are total of 41 GST proteins as retrieved from the Drosophila melanogaster genomedatabase. In the fruit fly, there are theta, delta, epsilon, omega, zeta, and sigma classesof GSTs. The polymorphic expression of the GSTs may suggest the variation of biologicalfunctions of the isoforms. Biochemical characterization of the delta class (DmGSTD1 toDmGSTD10), an epsilon class (DmGSTE1), and sigma class (DmGSTS1) GSTs has beeninvestigated (Sawicki et al., 2003). Further preliminary characterization of the full set ofthe GSTs was performed as described by Saisawang et al. (2012). In the study, the GSTswere categorized into theta (DmGSTT1-DmGSTT4), delta (DmGSTD1-DmGSTD11), ep-silon (DmGSTE1-GSTE14), omega (GSTO1-DmGSTO4), sigma (DmGSTS1), and zeta(DmGSTZ1-DmGSTZ2) classes.
Drosophila melanogaster glutathione S-transferase D3 (DmGSTD3) lacks more than 15amino acid residues at N terminus as observed in Figure 1. It was initially proposedthat DmGSTD3 was a pseudogene of D. melanogaster (Toung et al., 1993) on the basisof “suboptimal Kozak context” of the translation–initiation codon, which may preventits synthesis and a 15-amino acid N-terminal truncation that might cause the proteinto be inactive even if synthesized. On the contrary, GSTD3 mRNA was demonstratedto be translated and expressed in bacterial cells but it was cautioned that might notmean efficient translation of the gene in Drosophila in vivo (Sawicki et al., 2003). Itwas proposed that its expression was restricted to a preadult stage (Toung et al., 1993),having demonstrated the presence of GSTD3 transcript in a third instar larvae. GSTD3 wasidentified in adult flies by sulfobromophthalein (BSP)/GSH-agarose and GSH-agarose-based affinity matrices (Alias and Clark, 2007).
Archives of Insect Biochemistry and Physiology

212 � Archives of Insect Biochemistry and Physiology, July 2016
Figure 1. Multiple sequence alignment of members of delta class GSTs of D. melanogaster. All of the sequenceswere retrieved from Flybase (http://flybase.org). Y89 and Y97 of DmGSTD3 are as marked in box.
For a GST-catalyzed conjugation, the enzymatic reaction involves initial ionizationof GSH sulfhydryl group to yield a nucleophilic thiolate anion (GS-). This is generallyachieved when tyrosyl or seryl residues (located in the N-terminal of the sequence),which act as general bases, abstract hydrogen from GSH. This lowers the pKa of thethiol of GSH (Armstrong 1997). The GSTD3 is however shorter by 15 amino acids at theN-terminus compared to the other D-class GSTs (Sawicki et al., 2003). The absence ofN-terminal sequence means the absence of highly conserved catalytic tyrosine or serineresidues (Fig. 1). The purified recombinant GSTD3 was active toward 4-hydroxynonenal(Sawicki et al., 2003). Possible interaction of other amino acid residues that resultedin ionization of sulfyldryl group could have made the GST functional. Based on thisjudgment, GSTD3 could have adopted an “electron-sharing network” within the G-site assuggested to occur in AdGSTD3-3 (Winayanuwattikun and Ketterman, 2005). Our interestwas to investigate if there was a possible role of any far distant amino acid residues in thecatalysis. Thus, the study was directed to observe whether this amino acid residue/s makesany contribution to the catalytic mechanism of GSTD3 using bioinformatics analysis andsite-directed mutagenesis.
MATERIALS AND METHODS
Chemicals
Unless otherwise stipulated, chemicals employed were of the highest grade obtainable.Buffer components were purchased from Sigma-Aldrich.
Sample Preparation
Drosophila melanogaster that were used in this study were obtained from the GeneticsDepartment of University of Malaya. Adult fruit flies were bred and maintained at roomtemperature on standard oats agar medium. Newly emerged adult flies were continuouslysubcultured into new medium every week.
Archives of Insect Biochemistry and Physiology

Y89 and Y97 in the Conjugating Activity of DmGSTD3 � 213
Expression Plasmids and Site-Directed Mutagenesis
The total genomic DNA was extracted from the adult fruit flies by using the Qia-gen Genomic Extraction Kit (Hilden, Germany) and was used to amplify the targetgene, DmGSTD3. The DmGSTD3 gene of D. melanogaster was amplified using a ther-mal cycler with specific primers that produced a fragment of around 600 bp. For-ward primer was designed to introduce an NdeI restriction site of which the initia-tion codon, ATG, was included. Reverse primer spanned all or part of the stop codon(double underline) and was designed to introduce EcoRI restriction site (boldface);Forward DmGSTD3: 5′ CGCTCCGTTCATATGGTGGGCAAG; Reverse DmGSTD3: 5′
TCGAATTCAATGAATTATTTAGCAGCATTCTG.The amplified DNA was then purified and ligated into the plasmid pGEM-T Easy
(Promega, Madison, WI). The ligation reaction product was then transformed into Nov-aBlue Competent Cells (Escherichia coli JM109) (Novagen; Merck Millipore, Nottingham,United Kingdom) on Luria–Bertani (LB) agar plate containing ampicillin (100 mg/ml)and isopropyl β-D-1-thiogalactopyranoside (IPTG)-Xgal. Through the blue-white screen-ing method, the transformants were chosen and the pGEMT-D3 plasmid was extractedusing innuPREP Plasmid Rapid Kit (Analytik Jena, Jena, Germany). The recombinantplasmid was further confirmed by restriction site digestion and was sequenced. TheDNA encoding the GST gene was then subcloned into an expression vector pET-30a(+) (Novagen, Merck Millipore). The pET-D3 plasmid was then transformed into E. coliBL21 (DE3) pLysS (Novagen, Merck Millipore) strain on LB medium with kanamycin (30mg/ml). The QuikChange Lightning Site-Directed Mutagenesis Kit (Agilent, Santa Clara,CA) was used to make point mutations. Two complimentary oligonucleotides primers con-taining the desired mutation (Tyr to Ala), flanked by unmodified nucleotide sequence,were synthesized according to the primer design guidelines from the manufacturer’smanual:
Y89A: 5′ tgatatggcgctgatggctccgaccctggcgaac-3′
Y89A antisense: 5′-gttcgccagggtcggagccatcagcgccatatca-3′
Y97A: 5′-atccgaccctggcgaactattatgctaaagcgtttaccacc-3′,Y97A antisense: 5′-ggtggtaaacgctttagcataatagttcgccagggtcggat-3′
Protein Expression and Purification
A single colony of the transformed E. coli BL21 pET-D3 was inoculated into 5 ml LBmedium containing kanamycin (30 μg/μl) and incubated on a shaking incubator at37°C for 16 h. Then, 500 μl of the overnight culture was diluted into a fresh 500 ml LBmedium containing kanamycin. The culture was grown at 37°C until optical density at600 nm reached 0.6. The culture was then induced with 0.4 mM IPTG, and the incubationwas continued for an additional 4 h in the same conditions. After incubation, the culturewas then harvested by centrifugation at 7,000 × g for 10 min at 4°C. The pelleted cellswere resuspended in 2X Cellytic B buffer (Sigma-Aldrich) and mixed thoroughly with avortex before incubating it on a shaker for 15 min. The lysate was then centrifuged at16,000 × g for 60 min at 4°C and the supernatant was used for purification.
Chromatography was carried out using an AKTA Purifier FPLC (GE Healthcare,Little Chalfont, United Kingdom) equipped with UNICORN software Version 5.1 anda fraction collector (Frac900) was employed for greater automation of the purificationprocess. The protein was purified using anion exchange column (5 ml HiTrapTM QHP)
Archives of Insect Biochemistry and Physiology

214 � Archives of Insect Biochemistry and Physiology, July 2016
(GE Healthcare) (Sawicki et al., 2003) preequilibrated with sodium phosphate buffer(pH 8.0). The sample was applied using a 5-ml syringe and the flow rate was fixed at0.3 ml/min. Once sample had been applied, the preequilibrated column was washed withsodium phosphate buffer (pH 8.0) to remove the nonspecific binding proteins. Gradientelution was performed in order to identify the NaCl concentration that gives the mostpurified fraction. Elution of the bound enzyme with 0.5 M NaCl solution gave the highestactivities and a homogenously prepared band as judged from SDS-PAGE analysis. Thecollected fractions were then measured for activity and the active fractions were pooled,desalted, and freeze-dried for further analysis.
Substrate Specificities and Kinetic Characterization
Enzymatic assays with 1-chloro-2,4-dinitrobenzene (CDNB), ethacrynic acid, BSP,p-nitrobenzyl chloride, trans-4-phenyl-3-butene-2-one (Habig et al., 1974), and 1,2-dichloro-4-nitrobenzene (Motoyama and Dauterman, 1977) were performed. Abilityto conjugate trans-2-octenal, hexadienal, and trans,trans-2,4-heptadienal was performed(Brophy et al., 1989). All assays were determined spectrophotometrically of which re-quired concentration of substrates and GSH was mixed in the presence of enzymes in afinal 3 ml reaction volume. The progression of the enzymatic catalysis was monitored for3 min of with the rate of the conjugated product formation was measured. In all instances,the true activities were calculated by deducting the activities due to the spontaneous con-jugation reaction (formation of conjugated product in the absence of enzyme).
For the determination of kinetic parameters, initial velocity data was obtained byvarying the concentration of both substrates (CDNB and GSH). The kinetic measurementassay for CDNB and GSH was carried out independently; varying CDNB concentrations(0.4–1 mM) at fixed GSH (1 mM) or varying GSH concentrations (0.4–1 mM) at fixedCDNB. All assays were performed using a Jasco V-630 spectrophotometer equipped withtemperature controller, and kinetic data were evaluated by nonlinear regression analysiswith Michaelis–Menten equation (υ = Vmax × [S]/Km + [S]) using SigmaPlot 12.0 graphand analysis software. The catalytic constant (Kcat) was calculated using the equationKcat = Vmax/[E], where [E] is the total enzyme concentration.
Protein Quantification (Bradford Assay)
Protein concentration was determined using Coomassie Brilliant Blue G-250, and bovineserum albumin was used as standard (Bradford, 1976).
SDS Polyacrylamide Gel Electrophoresis
Electrophoresis was then run in 12% polyacrylamide gel in the presence of 0.1% SDSat 150 V. Gels were stained with Colloidal Coomassie Blue G-250 (Neuhoff et al., 1988).Stained gels were scanned with Image Scanner III (GE Healthcare) and visualized andanalyzed with Image Master Software.
Homology Modeling and Bioinformatic analysis
Homology modeling was performed to construct three-dimensional structure of theDmGSTD3 using the experimentally determined structure of a related family mem-ber as the template. The comparative protein structure modeling was performed with
Archives of Insect Biochemistry and Physiology

Y89 and Y97 in the Conjugating Activity of DmGSTD3 � 215
the help of SWISS-MODEL workspace (Arnold et al., 2006) as an integrated web-basedmodeling expert system, and the model was viewed in Swiss-PdbViewer 4.10 (Guex et al.,2009). On the basis of a sequence alignment between the target protein and the templatestructure (GST 1-1), a three-dimensional model for the target protein was generated.SWISS-MODEL workspace derived the restraints automatically from related known struc-tures (template) present in the database. All of the Delta class fruit flies GSTs wereretrieved from Flybase (http://flybase.org). Multiple-sequence alignment was performedusing CLUSTALW programme with default parameterization.
RESULTS AND DISCUSSION
Bioinformatic Analysis
An electron-sharing network occurs when negatively and positively charged residues in-teract electrostatically and are further stabilized by a network of hydrogen bonding. It wasshown that such interactions resulted in the glutamyl α-carboxylate of GSH functioning asthe catalytic base, accepting the hydrogen from the sulfyldryl group of GSH (Winderstenet al., 1996; Gustafsson et al., 2001). In AdGSTD3-3 (Winayanuwattikun and Ketterman,2005), Ser65 and Arg66 were shown to form hydrogen bonds to the GSH glutamyl α-carboxylate. The Arg66 residue was further stabilized by Asp100. The Ser65 interacts withThr158 via a water molecule. Thus, glutamyl α-carboxylate is essential to the catalyticeffect of the GSTs (Adang et al., 1988a, 1988b, 1989, 1990, 1991).
A homology modeling of GSTD3 three-dimensional structure was processed using theSWISS-model programme (Fig. 2). The N-terminal domain of most GSTs adopts the β-α-β-α-β-β-α structural motif. The C-terminal domain has all α-helices. The β-β-α structuralmotif of the domain is the most conserved among classes of GSTs (Wan et al., 2008).As analyzed from the GSTD3 SWISS-model structure shown in Figure 2, the N-terminaldomain consists of α-α-β-β-α. Figure 2 shows the presence of Y89 and Y97 on the C-terminal α-helix that point toward GSH. The same polar nature of the H-site was reportedin DmGSTS1 (residues Y153 and Y208), Lucilia cuprina LcGSTT1 (Y105 and Y113) (Wilceand Parker, 1994), and Anopheles dirus AdGST1-4 (Y111 and Y119) (Oakley et al., 1999).The residues in these proteins are all orientated similarly.
It was also observed that several amino acid residues are orientated so as to interactwith the GSH molecule in the G-site by hydrogen bonding (Fig. 3). In the model, tenta-tively the side chains of Ser49 and Arg50 interact with the GSH glutamyl α-carboxylate,while side chains of His34 and Ser35 interact with the glycine terminal carboxylate group.It was also observed that Glu48 could interact with the glutamyl amino group. In theGSTD3, it was seen that Arg50 interacts through hydrogen bonding to the glutamyl car-boxylate, and perhaps offers a role equivalent to Arg45 and Arg15 of DmGSTS1 andhGSTA4-4, respectively. It was proposed that not only glutamyl carboxylate serves as basefor thiol ionization but also may modulate the affinity for the electrophilic compoundvia indirect interactions with the C-terminal α-helix (Windersten et al., 1991). The par-ticipation of C- terminal α-helix in the binding of both GSH and electrophilic substrateis a characteristic of all α-class GSTs (Allardyce et al., 1995; Bruns et al., 1999). Struc-tural change in the C-terminus of the protein influences the binding of substrates. Thus,from this argument it was evident that in the absence of the N-terminal catalytic serineor tyrosine, GSTD3 can adopt a glutamyl carboxyl stabilization strategy to facilitate theionization of the thiol group by influencing the mode of binding of the GSH.
Archives of Insect Biochemistry and Physiology
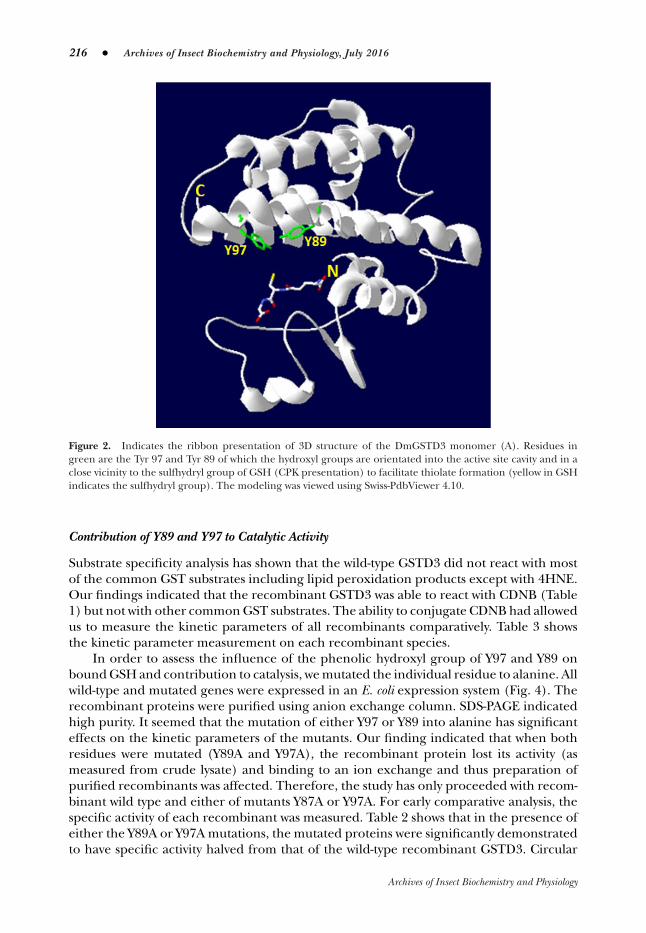
216 � Archives of Insect Biochemistry and Physiology, July 2016
Figure 2. Indicates the ribbon presentation of 3D structure of the DmGSTD3 monomer (A). Residues ingreen are the Tyr 97 and Tyr 89 of which the hydroxyl groups are orientated into the active site cavity and in aclose vicinity to the sulfhydryl group of GSH (CPK presentation) to facilitate thiolate formation (yellow in GSHindicates the sulfhydryl group). The modeling was viewed using Swiss-PdbViewer 4.10.
Contribution of Y89 and Y97 to Catalytic Activity
Substrate specificity analysis has shown that the wild-type GSTD3 did not react with mostof the common GST substrates including lipid peroxidation products except with 4HNE.Our findings indicated that the recombinant GSTD3 was able to react with CDNB (Table1) but not with other common GST substrates. The ability to conjugate CDNB had allowedus to measure the kinetic parameters of all recombinants comparatively. Table 3 showsthe kinetic parameter measurement on each recombinant species.
In order to assess the influence of the phenolic hydroxyl group of Y97 and Y89 onbound GSH and contribution to catalysis, we mutated the individual residue to alanine. Allwild-type and mutated genes were expressed in an E. coli expression system (Fig. 4). Therecombinant proteins were purified using anion exchange column. SDS-PAGE indicatedhigh purity. It seemed that the mutation of either Y97 or Y89 into alanine has significanteffects on the kinetic parameters of the mutants. Our finding indicated that when bothresidues were mutated (Y89A and Y97A), the recombinant protein lost its activity (asmeasured from crude lysate) and binding to an ion exchange and thus preparation ofpurified recombinants was affected. Therefore, the study has only proceeded with recom-binant wild type and either of mutants Y87A or Y97A. For early comparative analysis, thespecific activity of each recombinant was measured. Table 2 shows that in the presence ofeither the Y89A or Y97A mutations, the mutated proteins were significantly demonstratedto have specific activity halved from that of the wild-type recombinant GSTD3. Circular
Archives of Insect Biochemistry and Physiology

Y89 and Y97 in the Conjugating Activity of DmGSTD3 � 217
Figure 3. Putative hydrogen bonding interaction of the side chains of several amino acid residues with glu-tathione in the active site of DmGSTD3. Possible hydrogen bonding (shown in dotted line) of glutamyl car-boxylate (GSH in blue) with side chains of E48, S49, and R50. It also indicates possible hydrogen bonding ofside chains of H34 and S35 with the glycyl carboxylate. It also indicates the distance of possible hydrogen bondformation between hydroxyl groups of Y89 and Y87 to the sulfhydryl group of GSH. The side chains of theamino acids are in CPK presentation. The modeling was generated using Swiss-PdbViewer 4.10.
Table 1. Substrate Specificities of DmGSTD3 Against Selected Substrates
Substrate Specific activity (μmol/min/mg)
1-Chloro-2,4-dinitrobenzene 7.38 ± 0.411,2-Dichloro-4-nitrobenzene ND*trans-2-Hexenal NDtrans,trans 2,4-Hexadienal NDtrans-2-Octenal NDtrans-4-Phenyl-butene-2-one NDtrans,trans-Hepta-2,4-dienal NDEthacryanic acid NDp-Nitrobenzyl chloride ND
ND*, not detectable.
Table 2. Specific Activities of the Purified Recombinant DmGSTD3 Wild Type and Mutants (Y89 and Y97)When Acted Against CDNB
Sample Specific activity (μmol/min/mg)
Wild type 7.38 ± 0.41Y89A 3.51 ± 0.09Y97A 3.01 ± 0.15
Archives of Insect Biochemistry and Physiology

218 � Archives of Insect Biochemistry and Physiology, July 2016
Table 3. Kinetic Parameters of the DmGSTD3 WT, Y89, and Y97 With CDNB as Hydrophobic Substrate
Vmax Km Kcat Kcat/Km
CDNB GSH CDNB GSH CDNB GSH CDNB GSH
Wt 25.72 ± 4.11 27.46 ± 5.31 2.77 ± 0.49 2.66 ± 0.87 1.29 ± 0.21 1.38 ± 0.37 1.38 ± 0.37 0.519Y89A 13.15 ± 2.49 5.48 ± 0.91 2.60 ± 0.63 4.28 ± 0.90 0.64 ± 0.12 0.27 ± 0.09 0.247 0.063Y97A 10.87 ± 2.09 7.24 ± 1.01 2.32 ± 0.89 5.56 ± 0.71 0.52 ± 0.09 0.35 ± 0.05 0.226 0.063
Figure 4. SDS-PAGE of (A) crude protein extracted from the E. coli BL21 cloned with wild-type DmGSTD3and mutated proteins. (B) Purified recombinant wild-type and mutated GSTD3.(A) Arrow indicates the overexpression of recombinant GSTD3 proteins of wild-type and mutated clones afterIPTG induction. Lane 1 protein marker (Benchmark; Invitrogen); Lane 2 crude lysate of DmGSTD3 wildtype; Lane 3 crude lysate of DmGSTD3-Y89A; Lane 4 crude lysate of DmGSTD3-Y97A; Lane 5 crude lysate ofDmGSTD3-Y89A and Y97A. Gel was stained with Coomassie blue.(B) The purified recombinant proteins (wild type, Y89A, and Y97A) appeared single band at MW approximately25 kDa. Gel was stained with Coomassie blue.
dichroism spectroscopy reconfirmed that there was no conformational change due to thesite-directed mutagenesis (data not shown) and any resulted catalytic reaction was merelyaffected from the mutated amino acid. Mutation of both residues (Y89A and Y97A) hasresulted in loss of activity (as there was no activity detected in crude sample) and capabilityin binding to the ion exchange chromatography. This indicates that the presence of bothor either residues have essential contribution to the GSH binding in the G-site of theenzyme Y97 and Y89 are located in the helix 4 and their hydroxyl groups are pointinginto the G-site where the GSH is located. Our assumption was that the homology modelindicates a hydrogen bonding interaction between the groups and the sulfhydryl group of
Archives of Insect Biochemistry and Physiology

Y89 and Y97 in the Conjugating Activity of DmGSTD3 � 219
the GSH and this could stabilize the GSH moiety. Data in Table 3 indicate that both Y89Aand Y97A proteins have equal Km
GSH values. The values increased twofold as compared tothat of the wild-type protein, suggesting the reduction of affinity of both mutants towardGSH.
The mutated residues however did not affect the affinity of the proteins towardCDNB. Due to changes in observed Km
GSH, the maximal velocity of the rate of reactionvery much affected. Significant reduction (fivefold) of Vmax
GSH was observed in Y89A(5.48 ± 0.91 μmol/min/mg) and Y97A (7.24 ± 1.01 μmol/min/mg) when GSHwas varied as compared to wild-type protein (27.46 ± 5.31 μmol/min/mg). Thisimplies that both tyrosines at position 97 and 89 play important roles in stabiliza-tion of the GSH in the G-site. Interestingly, the effect of replacement of each Tyrposition has not changed the Kmax value, implying that both residues have not af-fected the binding of cosubstrate, CDNB. The Kca
GSH and KcaCDNB for both mu-
tants have been appreciably impaired as compared to the wild type. An increasein Km
GSH and reduction of KcatGSH have affected the catalytic efficiency of both
mutants, where the KcatGSH/Km
GSH for both Y97A (0.063 mM−1sec−1) and Y89A(0.063 mM−1sec−1) was reduced eightfold.
CONCLUSIONS
In conclusion, we have demonstrated that the N-terminally truncated DmGSTD3 is afunctional protein of which its conjugation reaction was affected by tyrosine residues inpositions 97 and 89. The preliminary homology modeling study indicated the positioningof the tyrosine into the active cavity of which the hydroxyl groups could have presumablycontributed the stabilization of the thiolate formation (through hydrogen bonding) in theGSH molecule. The absence of the hydroxyl group on each position has greatly affectedthe velocity of reaction, affinity, and the catalytic efficiency of the enzyme toward GSH.The replacement has not contributed any significant change in the enzyme affinity towardCDNB. However, a lesser degree of change was also observed on the maximal velocity andthe catalytic efficiency toward CDNB as the substrate. The study again demonstrated theimportance of GSH stabilization in the conjugation reaction catalyzed by GSTs.
ACKNOWLEDGMENT
The authors are also thankful to Dr. A. G. Clark (Victoria University of Wellington) forhis critical reading of the manuscript.
Conflict of Interest
There is no conflict of interests that could be perceived as prejudicing the impartiality ofthe research reported.
LITERATURE CITED
Adang AE, Duindam AJ, Brussee J, Mulder GJ, van der Gen A. 1988a. Synthesis and nucleophilic re-activity of a series of glutathione analogues, modified at the gamma- glutamyl moiety. BiochemJ 255:715–720.
Archives of Insect Biochemistry and Physiology

220 � Archives of Insect Biochemistry and Physiology, July 2016
Adang AE, Brussee J, Meyer DJ, Coles B, Ketterer B, van der Gen A, Mulder GJ. 1988b. Substratespecificity of rat liver glutathione S-transferase isoenzymes for a series of glutathione analogues,modified at the gamma-glutamyl moiety. Biochem J 255:721–724.
Adang AE, Meyer DJ, Brussee J, van der Gen A, Ketterer B, Mulder GJ. 1989. Interaction of ratglutathione S-transferases 7-7 and 8-8 with gamma-glutamyl or glycyl-modified glutathioneanalogues. Biochem J 264:759–764.
Adang AE, Brussee J, van der Gen A, Mulder GJ. 1990. The glutathione-binding site in glutathioneS-transferases. Investigation of the cysteinyl, glycyl and gamma-glutamyldomains. Biochem J269:47–54.
Adang AE, Brussee J, van der Gen A, Mulder GJ. 1991. Inhibition of rat liver glutathione S transferaseisoenzymes by peptides stabilized against degradation by gamma-glutamyl transpeptidase. J BiolChem 266:830–836.
Alias Z, Clark AG. 2007. Studies on the gluthathione S-transferase proteome of adult Drosophilamelanogaster: responsiveness to chemical challenge. Proteomics 7:3618–3628.
Allardyce CS, McDonagh PD, Lian LY, Wolf CR, Roberts CKG. 1995. The role of tyrosine-9 and theterminal helix in the catalytic mechanism of Alpha-class glutathione S-transferases. Biochem J343:525–531.
Armstrong RN. 1997. Structure, catalytic mechanism and evolution of the glutathione transferases.Chem Res Toxicol 10:2–18.
Arnold K, Bordoli L, Kopp J, Schwede T. 2006. The SWISS-MODEL workspace: a web-based envi-ronment for protein structure homology modelling. Bioinformatics 22:195–201.
Bradford M. 1976. A rapid and sensitive method for the quantitation of microgram quantities ofprotein utilising the principle of protein-dye binding. Anal Biochem 72:248–254.
Brophy PM, Southa C, Barrett J. 1989. Glutathione transferases in the tapeworm Moniezia expansa.Biochem J 262:939–946.
Bruns CM, Hubatsch I, Ridderstrom M, Mannervik B, Tainer JA. 1999. Human glutathione trans-ferase A4-4 crystal structures and mutagenesis reveal the basis of high catalytic efficiency withtoxic lipid peroxidation products. J Mol Biol 288:427–439.
Clark AG. 1989. The comparative enzymology of the glutathione S-transferases from non-vertebrateorganisms. Comp Biochem Physiol 92:419–446.
Coles B, Ketterer B. 1990. The role of glutathione and glutathione S-transferases in chemicalcarcinogenesis. Biochem Mol Biol 25:47–70.
Edwards R, Dixon DP, Walbot V. 2000. Plant glutathione S-transferases: enzymes with multiplefunctions in sickness and health. Trends Plant Sci 5:193–198.
Enayati AA, Ranson H, Hemingway J. 2005. Insect glutathione transferases and insecticides resis-tance. Insect Mol Biol 14:3–8.
Guex N, Peitsch MC, Schwede T. 2009. Automated comparative protein structure modeling withSWISS-MODEL and Swiss-PdbViewer: a historical perspective. Electrophoresis 30:S162–S173.
Gustafsson A, Petterson PL, Grehn L, Jemth P, Mannervik B. 2001. Role of Glutamyl α- carboxylateof the substrate glutathione in the catalytic mechanism of human glutathione transferase A1-1.Biochemistry 40:15835–15845.
Habig WH, Pabst MJ, Jakoby WB. 1974. Glutathione S-transferases: the first enzymatic step inmercapturic acid formation. J Biol Chem 249:7130–7139.
Hayes JD, Pulford DJ. 1995. The glutathione S-transferase supergene family: regulation of GST andthe contribution of the isoenzymes to cancer chemoprotection and drug resistance. Crit RevBiochem Mol Biol 30:445–600.
Motoyama N, Dauterman WC. 1977. Purification and properties of housefly glutathione S trans-ferase. Insect Biochem 7:361–269.
Archives of Insect Biochemistry and Physiology

Y89 and Y97 in the Conjugating Activity of DmGSTD3 � 221
Neuhoff V, Arold N, Taube D, Ehrhardt W. 1988. Improved staining of proteins in polyacrylamidegels including isoelectric focussing gels with clear background at nanogram sensitivity usingCoomassie Brilliant Blue G-250 and R-250. Electrophoresis 9:255–262.
Oakley AJ, Bello ML, Nuccetelli M, Mazzetti AP, Parker MW. 1999. The ligandin (non substrate)binding site of human Pi class glutathione transferase is located in the electrophilic bindingsite (H-site). J Mol Biol 291:913–926.
Nair PMG, Choi J. 2011. Identification, characterization and expression profiles of Chironomusriparius glutathione S-transferase (GST) genes in response to cadmium and silver nanoparticlesexposure. Aqua Toxicol 101:550–560.
Sawicki R, Singh SP, Mondal AK, Benes H, Zimniak P. 2003. Cloning, expression and biochem-ical characterization of one Epsilon-class (GST-3) and ten Delta-class (GST-1) glutathioneS-transferases from Drosophila melanogaster, and identification of additional nine members ofthe Epsilon class. Biochem J 370:661–669.
Saisawang C, Wongsantichon J, Ketterman AJ. 2012. A preliminary characterisation of the cytosolicglutathione transferase proteome from Drosophila melanogaster. Biochem J 442:181–190.
Toung Y-PS, Hsieh T-S, Tu C-PD. 1993. The glutathione S-transferase D genes: a divergently organ-ised, intronless gene family in Drosophila melanogaster. J Biol Chem 268:9737–9746.
Travensolo RF, Garcia W, Muniz JRC, Caruso CS, Lemos EGM, Carrilho E, Araoujo APU.2008. Cloning, expression, purification and characterization of recombinant gluthathione-s-transferase from Xylella fastidious. Protein Expr Purif 59:153–160.
Udomsinprasert R, Pongjaroenkit S, Wongsantichon J, Oakley AJ, Prapantadhara L-A, Wilce MCJ,Ketterman AJ. 2005. Identification, characterization and structure of a new Delta class glu-tathione transferase isoenzyme. Biochem J 388:763–771.
Vontas JG, Small GJ, Hemingway J. 2001. Glutathione S-transferases as antioxidant defence agentsconfer pyrethroid resistance in Nilaparvata lugens. Biochem J 357:65–72.
Wan Q, Whang I, Lee J. 2008. Molecular characterization of mu class glutathione-s-transferase fromdisk abalone (Haliotis discus), a potential biomarker of endocrine-disrupting chemicals. CompBiochem Physiol 150(2):187–199.
Wilce MCJ, Parker MW. 1994. Structure and function of glutathione S-transferses. Biochem BiophysActa 205:1–18.
Winayanuwattikun P, Ketterman AJ. 2005. An electron-sharing network involved in the catalyticmechanism is functionally conserved in different glutathione transferases classes. J Biol Chem280:31776–31782.
Windersten M, Bjornestedt R, Mannervik B. 1996. Involvement of the carbonyl of glutathione inthe catalytic mechanism of human glutathione transferase A1-1. Biochemistry 35:7731–7742.
Yu SJ. 1996. Insect glutathione S-transferases. Zool Stud 35:9–19.
Archives of Insect Biochemistry and Physiology