p27kip1 signaling- transcriptional and post-translational regulation
DESCRIPTION
p27 siganllingTRANSCRIPT

S
p
SZa
b
Ac
d
e
f
g
a
ARRAA
KpMSKCP
ptFaampPwespS
e
M
h1
The International Journal of Biochemistry & Cell Biology 68 (2015) 9–14
Contents lists available at ScienceDirect
The International Journal of Biochemistry& Cell Biology
jo ur nal home page: www.elsev ier .com/ locate /b ioce l
ignalling networks in focus
27Kip1 signaling: Transcriptional and post-translational regulation
u Su Thae Hnita, Chanlu Xiea, Mu Yaob, Jeff Holstc,d, Alan Bensoussane, Paul De Souzaf,hong Lig,∗∗, Qihan Donga,b,f,∗
School of Science and Health, University of Western Sydney, AustraliaCentral Clinical School and Charles Perkins Centre, The University of Sydney and Department of Endocrinology, Royal Prince Alfred Hospital, Sydney,ustraliaOrigins of Cancer Program, Centenary Institute, Camperdown, NSW, AustraliaSydney Medical School, The University of Sydney, Sydney, NSW, AustraliaNational Institute of Complementary Medicine, University of Western Sydney, AustraliaSchool of Medicine, University of Western Sydney, AustraliaDongzhimen Hospital, Beijing University of Chinese Medicine, Beijing, China
r t i c l e i n f o
rticle history:eceived 17 April 2015eceived in revised form 8 August 2015ccepted 10 August 2015vailable online 14 August 2015
eywords:27Kip1
YC
a b s t r a c t
p27Kip1 is an inhibitor of a broad spectrum of cyclin-dependent kinases (CDKs), and the loss of a singlep27Kip1 allele is thereby sufficient to increase tumor incidence via CDK-mediated cell cycle entry. As such,down-regulation of p27Kip1 protein levels, in particular nuclear expressed p27Kip1, is implicated in bothdisease progression and poor prognosis in a variety of cancers. p27Kip1 expression is positively regulatedby the transcription factor MENIN, and inhibited by oncogenic transcription factors MYC and PIM. How-ever, regulation of p27Kip1 protein expression and function is predominantly through post-translationalmodifications that alter both the cellular localization and the extent of E3 ubiquitin ligase-mediateddegradation. Phosphorylation of p27Kip1 at Thr187 and Ser10 is a prerequisite for its degradation via the E3
10
KP2PCRM1IRH2ubiquitin ligases SKP2 (nuclear) and KPC (cytoplasmic), respectively. Additionally, Ser phosphorylatedp27Kip1 is predominantly localized in the cytoplasm due to the nuclear export protein CRM1. Another E3ubiquitin ligase, PIRH2, degrades p27Kip1 in both the cytoplasm and nucleus independent of phosphory-lation state. As such, inhibition of cell cycle entry and progression in a variety of cancers may be achievedwith therapies designed to correct p27Kip1 localization and/or block its degradation.
Abbreviations: ABL, abelson murine leukemia viral oncogene homolog; AKT,rotein kinase B; Ala, alanine; AMPK, 5′ adenosine monophosphate-activated pro-ein kinase; ATP, adenosine triphosphate; E2F, elongation 2 transcription factor;OXO3a, forkhead box O3a; JAK2, janus kinase 2; kDa, kilodalton; KIS, kinase inter-cting stathmin; LYN, lck/yes-related novel protein tyrosine kinase; MAT1, menage
trois 1-CDK-activating kinase assembly factor; MAX, myc-associated factor X;iR, microRNA; MYC, v-myc avian myelocytomatosis viral oncogene homolog;
21Cip1, CDK-interacting protein 1; p27Kip1, cyclin dependent kinase inhibitor 1B;IM, proto-oncogene serine/threonine-protein kinase; PIRH2, p53-induced proteinith a RING-H2 domain; RB, retinoblastoma; RING finger domain, really inter-
sting new gene finger domain; SCF, Skp1-Cullin-Fox complex; Ser, serine; SGK,erum/glucocorticoid-regulated kinase; siRNA, small interference RNA; SKP2, s-hase kinase-associated protein 2; SRC, proto-oncogene tyrosine-protein kinaseRC; TGF�, transforming growth factor �; Thr, threonine; Tyr, tyrosine.∗ Corresponding author at: School of Science and Health, The University of West-rn Sydney, Penrith South, NSW 2751, Australia.∗ ∗ Corresponding author at: Dongzhimen Hospital, Beijing University of Chinese
edicine, 5 Haiyuncang Hutong, Dongcheng, Beijing, China.E-mail addresses: [email protected] (Z. Li), [email protected] (Q. Dong).
ttp://dx.doi.org/10.1016/j.biocel.2015.08.005357-2725/© 2015 Elsevier Ltd. All rights reserved.
© 2015 Elsevier Ltd. All rights reserved.
1. Introduction
In 1994, a 27 kDa protein was found to inhibit the activity ofthe cyclin-dependent kinase 2 (CDK2) following treatment of minklung epithelial cells with TGF� (Polyak et al., 1994). This pro-tein was named CDK inhibitor 1B or p27. As a member of kinaseinhibitor protein (KIP) family, p27 was subsequently named asp27Kip1 and was thought to control cell cycle progression in G1phase (Polyak et al., 1994). While this has been confirmed by othergroups (Grimmler et al., 2007; O’Hagan et al., 2000; Sheaff et al.,1997), p27Kip1 can also bind and inhibit cyclin-CDK complexesbeyond the G1 phase of cell cycle (James et al., 2008).
p27Kip1 is haploinsufficient, with the loss of a single p27Kip1
allele being sufficient to increase tumor incidence (Gao et al., 2004).Decreased levels of p27Kip1 protein, in particular nuclear expressedp27Kip1, is implicated in disease progression and poor prognosisin lung, head and neck, colon, breast, ovary and prostate cancer
(Chu et al., 2008). p27Kip1-null mice show multiorgan hyperplasia,retinal dysplasia, increased body size and tumor formation (Feroet al., 1996; Gao et al., 2004), suggesting an indispensable function
1 l of Bio
oso
2
eptce
2
tbetspHtriocR
2
ecplpattpponiebcs
2
CeapeiDmuy
0 S.S.T. Hnit et al. / The International Journa
f p27Kip1. Hence, understanding the regulation of p27Kip1 expres-ion via both transcriptional and post-translational mechanisms isf clinical importance.
. Functions of p27Kip1
The p27Kip1 gene is located on chromosome 12p13 (Pietenpolt al., 1995). The CDK-inhibitory domain resides in the N-terminalortion of p27Kip1 and is sufficient to arrest cells at G0/G1. The C-erminal portion of p27Kip1 is less conserved in the KIP family andontains a nuclear localization signal (Polyak et al., 1994; Russot al., 1996).
.1. Inhibitor of cyclin E/A-CDK2
The role of p27Kip1 in blocking cyclin E/A-CDK2 is well charac-erized. p27Kip1 interacts with cyclin A through a conserved cyclininding domain, and CDK2 via the kinase binding region (Russot al., 1996). p27Kip1 inhibits cyclin E/A-CDK2 activity by three dis-inct mechanisms: (i) blocking ATP access to CDK (ii) preventingubstrate access to the CDK, and (iii) preventing the activatinghosphorylation of CDK by the CDK-activating kinase (i.e. cyclin/CDK7/MAT1). As a result, the cyclin E/A-CDK2 complex is unable
o phosphorylate RB, causing a pause in the transcription of genesequired for progression to S phase. Furthermore, this results innhibition of other S-phase promoting activities such as formationf the origin recognition complex for DNA replication through theyclin E/A-CDK2 complex (Aleem et al., 2005; James et al., 2008;ay et al., 2009; Russo et al., 1996).
.2. The mode of action on cyclin D-CDK4/6
The relationship of p27Kip1 and cyclin D-CDK4/6 is complex (Rayt al., 2009). p27Kip1-bound cyclin D-CDK4/6 complexes remainatalytically active in proliferating keratinocytes, epithelial or lym-hoid cells (James et al., 2008). In quiescent epithelial and resting
ymphoid cells, however, p27Kip1-bound cyclin D-CDK4/6 com-lexes are inactive. It is believed that the effect of p27Kip1 on thectivity of cyclin D-CDK4/6 is dependent on the growth state ofhe cells (James et al., 2008; Ray et al., 2009). In cycling cells,he p27Kip1-cyclin D-CDK4/6 complex is active due to the phos-horylation at Tyr74, Tyr88 and/or Tyr89 (Ray et al., 2009). Thehosphorylation of p27Kip1 pushes its tail out of the catalytic cleftf CDK4/6, exposing the CDK4/6 active site to ATP and substrate. Inon-cycling cells, Tyr88 and Tyr89 are not phosphorylated, result-
ng in occupation of catalytic cleft of the CDK by p27Kip1 tail (Rayt al., 2009), i.e., the bound and inhibitory mode of p27Kip1. Bylocking cyclin D-CDK4/6 complexes from phosphorylating RB, andonsequent repression of the transcription activity of E2F, the tran-cription of the genes required for G1-S transition is halted.
.3. Inhibitor of cyclin B-CDK1
Studies in mice have demonstrated that knockout of CDK2, CDK3,DK4 and CDK6 are not lethal (Adhikari et al., 2012; Santamariat al., 2007). Mouse oocytes lacking these interphase CDKs are stillble to proliferate, revealing that CDK1 can replace these inter-hase CDKs and drive cell division (Adhikari et al., 2012; Santamariat al., 2007). In CDK2 knockout mice, p27Kip1 was found to bind andnhibit CDK1 activity in the thymus and spleen (Aleem et al., 2005).
eletion of p27Kip1 significantly upregulated CDK1 activity and pro-oted G1-S transition (Aleem et al., 2005). The exact mechanismnderlying p27Kip1 regulation of G1-S transition through CDK1 iset to be determined.
chemistry & Cell Biology 68 (2015) 9–14
Apart from controlling proliferation, p27Kip1 has also beenshown to have a role in differentiation, affecting terminal differen-tiation of neuronal stem cells (Bahrami et al., 2005) and erythroiddifferentiation of a multipotent human leukemia cell line (Munoz-Alonso et al., 2005). Furthermore, cytoplasmic p27kip1 promotescell migration and metastasis in HepG2 hepatocellular carcinomacells (Denicourt and Dowdy, 2004; McAllister et al., 2003).
3. Transcriptional and post-translational regulation ofp27Kip1 by MYC, FOXO, PIM and MENIN
MYC is an oncogenic transcription factor of thehelix–loop–helix/leucine zipper family. The human MYC genefamily is composed of MYC, MYCN and MYCL1, and all threeMYC gene products have similar effects on cell cycle progression(Bretones et al., 2015). MYC-MAX heterodimer binds to E-boxes inthe regulatory regions of its targeted genes and recruits transcrip-tional coactivators or repressors. Transient overexpression of MYCin Jurkat T cells and breast cancer cells increased binding of theMYC homology box II to the p27Kip1 promoter, directly repressingp27Kip1 gene transcription (Yang et al., 2001). By comparison,Forkhead box O proteins (FOXO4, FOXO3a, and FOXO1a) aretranscription factors that can induce p27Kip1 gene expression,leading to cell cycle arrest at G0/G1 (Fig. 1). Interestingly, MYCcan also suppress p27Kip1 transcription through an interactionbetween the MYC homology box II domain and the DNA-bindingportion of FOXO3a in mouse fibroblasts, breast cancer cells, JurkatT cells and human embryonic kidney cells (Bretones et al., 2015;Chandramohan et al., 2008).
Another mechanism by which MYC regulates p27Kip1 expressionis through transcriptional regulation of other genes and microR-NAs. MYC transcriptionally regulates expression of miR-221 andmiR-222, which have been shown to target p27Kip1 mRNA inglioblastoma cell lines (le Sage et al., 2007; Pineau et al., 2010).MYC also regulates expression of E3 ubiquitin ligase componentsSKP2 and CUL1, leading to p27Kip1 protein degradation (Bretoneset al., 2011; O’Hagan et al., 2000). Other MYC target genes, such asE2F (E2F1, E2F2 and E2F3) transcription factors, may also play a rolein regulating p27Kip1, by inducing cyclin E expression (Muller et al.,1997; Pérez-Roger et al., 1997) (Fig. 1).
The serine/threonine kinase PIM family members (PIM1, PIM2and PIM3) have been shown to directly phosphorylate p27Kip1 atThr157 and Thr198, resulting in cytoplasmic localization (Morishitaet al., 2008). Furthermore, PIM1S (PIM1 short isoform) can phos-phorylate and inactivate FOXO3a at Thr32 and Ser253 and FOXO1a atThr24, Ser256 and Ser319, thereby repressing FOXO-mediated p27Kip1
gene transcription in human embryonic kidney cells (Morishitaet al., 2008). MENIN is encoded by the MEN1 gene, and its mutationis responsible for multiple endocrine neoplasia type 1. MENIN bindsto the p27Kip1 gene promoter and activates p27Kip1 gene expressionby modulating histone methyltransferase activity (Chu et al., 2008;Karnik et al., 2005) (Fig. 1).
4. Phosphorylation of p27Kip1 at Thr187 promotesSCFSKP2-dependent degradation in the nucleus
Despite these clear transcriptional mechanisms, regulation ofp27Kip1 is mainly via post-translational modifications. The proteinlevel of p27Kip1 is controlled by ubiquitin-dependent proteolysis,which requires phosphorylation of p27Kip1 (Chu et al., 2008). Thephosphorylation of p27Kip1 at Thr187 by cyclin E-CDK2 is recognized
by the F-box protein SKP2, a subunit of the SCF E3 ubiquitin lig-ase (SCFSKP2), and thus triggers ubiquitin-dependent degradationin the nucleus (Fig. 2A). The importance of Thr187 phosphoryla-tion is demonstrated by introduction of the Ala187 mutation, which
S.S.T. Hnit et al. / The International Journal of Biochemistry & Cell Biology 68 (2015) 9–14 11
Fig. 1. Transcriptional and post-translational regulation of p27Kip1 by MYC, PIM and MENIN. MYC represses the transcription of p27Kip1 gene (CDKN1B) directly or indirectlyv nd mi Kip1
t regulap xpresb
lmtcc1EeSgpTubt
ia(soo(o
5K
icc2st
ia inactivating FOXO3a-mediated CDKN1B transcription. MYC induces miR-221 aranscription directly or indirectly through E2F transcription factors. MYC also up27Kip1 protein by SCFSKP2-mediated ubiquitination. PIM suppresses p27Kip1 gene eind to the p27Kip1 promoter and induce p27Kip1 gene expression.
eads to resistance to degradation of p27Kip1 and G1 arrest inouse fibroblasts (Sheaff et al., 1997). Similar mutation of Thr187
o Gly187 also increased p27Kip1 stability in the extracts of HeLaells (Tsvetkov et al., 1999). As p27Kip1 binds the cyclin E/A-CDK2omplex, p27Kip1 is also a substrate of the complex (Sheaff et al.,997). The progressive decrease of p27Kip1 in G1 permits cyclin/A-CDK2 to promote the G1-S transition in HeLa cells (Tsvetkovt al., 1999). Despite clear evidence that degradation of p27Kip1 byCFSKP2 occurs during S phase (Tsvetkov et al., 1999), there are sug-estions that SCFSKP2 may also degrade p27Kip1 in other cell cyclehases (Chu et al., 2008; Hattori et al., 2007; Kamura et al., 2004).here is also an accessory protein, cyclin-dependent kinases reg-latory subunit 1 (CKS1), which appears to be necessary to bridgeetween p27Kip1 and SCFSKP2 as determined by the 3D structure ofhe ternary complex (Hao et al., 2005).
There is also evidence to support SCFSKP2-mediated proteolysisndependent of Thr187 phosphorylation, working together with
subunit of the damaged-DNA binding protein (DDB1), CUL4ACullin 4A) and the COP9 signalosome (Bondar et al., 2006).iRNA-mediated knockdown of DDB1, CUL4A, and CSN1 (a subunitf the signalosome) causes an accumulation of p27Kip1, whereasverexpression of DDB1 reduces the level of p27Kip1 in HeLa cellsBondar et al., 2006). Further studies are needed to verify this formf p27Kip1 regulation.
. Phosphorylation of p27Kip1 at Ser10 fostersPC-dependent degradation in the cytoplasm
By analyzing crude extracts of SCFSKP2 knockout cells, the SKP2ndependent E3 ubiquitin ligases Kip1 ubiquitylation-promotingomplex (KPC), containing KPC1 and KPC2, were found in the
ytoplasmic fraction of the rabbit reticulocyte sample (Chu et al.,008; Kamura et al., 2004). Through binding to CDK interactingite of p27Kip1 (Kotoshiba et al., 2005), KPC1 and KPC2 mediatehe cytoplasmic p27Kip1 ubiquitination in G1-S phase in a Ser10R-222, which can silence p27 mRNA. MYC also induces cyclin E gene (CCNE1)tes SKP2 transcription and the increased SKP2 levels contribute to degradation ofsion by inactivating FOXO1a and FOXO3a by phosphorylation. MENIN can directly
phosphorylation-dependent manner (Fig. 2B). Overexpression ofboth KPC1 and KPC2 stimulated the degradation of p27Kip1 whilethe ubiquitination of p27Kip1 was reduced with a loss-of-functionmutation in KPC1 in mouse fibroblasts. Moreover, the treatment ofNIH 3T3 cells with KPC1 RNAi induced a delay in p27Kip1 degrada-tion, with a similar effect observed in mouse embryonic fibroblastsresulting in a decrease of p27Kip1 degradation in G0-G1 transition(Kamura et al., 2004). This study demonstrated that the action ofKPC regulates p27Kip1 degradation in G0 to S phase (Kamura et al.,2004) and therefore further studies are required to examine itsaction on G2 and M phase in the presence of active SKP2.
6. Phosphorylation at Ser10 stimulates nuclear export ofp27Kip1 by CRM1
As an inhibitor of DNA duplication and cell division, p27Kip1
exerts its anti-proliferative action inside the nucleus. Chromosomeregion maintenance 1 (CRM1) is a carrier protein for nuclear export(Wang et al., 2013), which mediates the cytoplasmic translocationof p27Kip1 through binding to its CDK interacting site (Chu et al.,2008). In early G1, phosphorylation of p27Kip1 at Ser10 by kinasesincluding AKT, CDK5 and KIS facilitates its binding to CRM1 andsubsequent transport from the nucleus to the cytoplasm (Fig. 2B).Mutation of Ser10 to Ala10 reduces p27Kip1 nuclear export, and ledto accumulation of p27Kip1 in the nucleus of mouse fibroblasts evenafter serum stimulation (Ishida et al., 2002). In CRM1 siRNA treatedovarian cancer cells, a reduction of CRM1 protein levels and coinci-dent accumulation of Ser10 phosphorylated p27Kip1 in the nuclearfraction were observed (Wang et al., 2013). These data suggestthat CRM1 plays an essential role in the subcellular localization
of p27Kip1 and subsequently in cell cycle regulation. Since Ser10phosphorylation promotes KPC-dependent degradation in thecytoplasm, it is possible that KPC-mediated degradation of p27Kip1
in G1 phase requires CRM1 to export p27Kip1 from the nucleus.
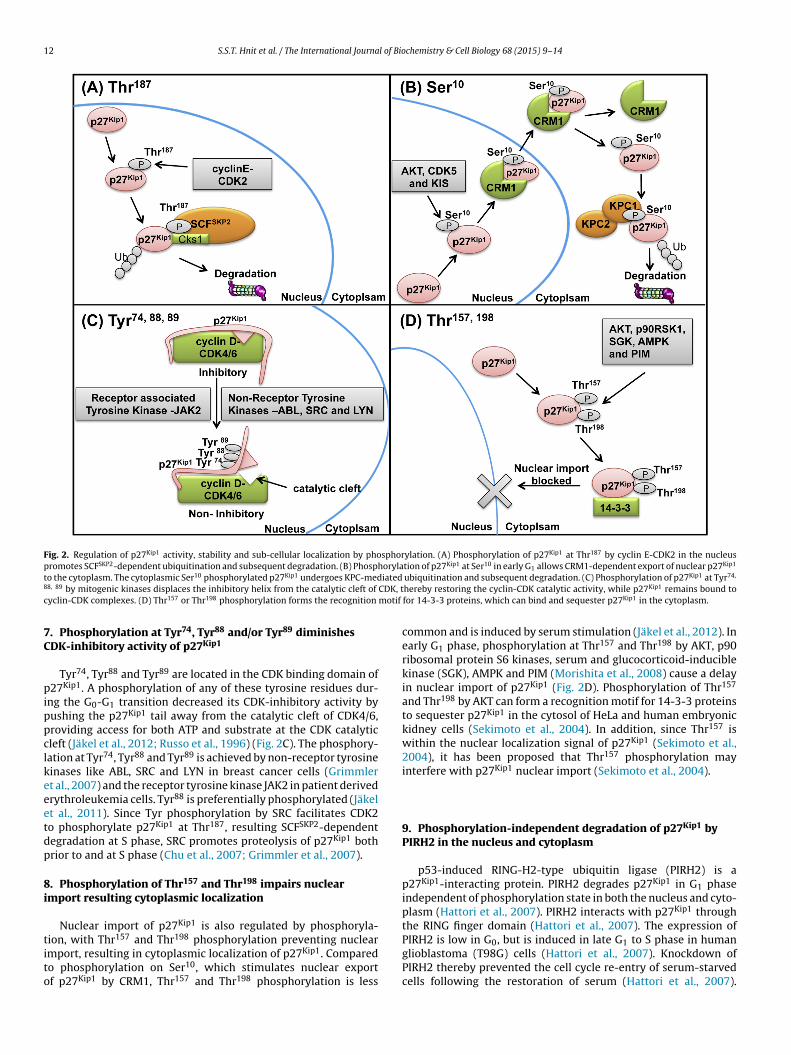
12 S.S.T. Hnit et al. / The International Journal of Biochemistry & Cell Biology 68 (2015) 9–14
Fig. 2. Regulation of p27Kip1 activity, stability and sub-cellular localization by phosphorylation. (A) Phosphorylation of p27Kip1 at Thr187 by cyclin E-CDK2 in the nucleuspromotes SCFSKP2-dependent ubiquitination and subsequent degradation. (B) Phosphorylation of p27Kip1 at Ser10 in early G1 allows CRM1-dependent export of nuclear p27Kip1
t iated8 CDK, tc motif
7C
pippclkeeetdp
8i
tito
o the cytoplasm. The cytoplasmic Ser10 phosphorylated p27Kip1 undergoes KPC-med8, 89 by mitogenic kinases displaces the inhibitory helix from the catalytic cleft of
yclin-CDK complexes. (D) Thr157 or Thr198 phosphorylation forms the recognition
. Phosphorylation at Tyr74, Tyr88 and/or Tyr89 diminishesDK-inhibitory activity of p27Kip1
Tyr74, Tyr88 and Tyr89 are located in the CDK binding domain of27Kip1. A phosphorylation of any of these tyrosine residues dur-
ng the G0-G1 transition decreased its CDK-inhibitory activity byushing the p27Kip1 tail away from the catalytic cleft of CDK4/6,roviding access for both ATP and substrate at the CDK catalyticleft (Jäkel et al., 2012; Russo et al., 1996) (Fig. 2C). The phosphory-ation at Tyr74, Tyr88 and Tyr89 is achieved by non-receptor tyrosineinases like ABL, SRC and LYN in breast cancer cells (Grimmlert al., 2007) and the receptor tyrosine kinase JAK2 in patient derivedrythroleukemia cells. Tyr88 is preferentially phosphorylated (Jäkelt al., 2011). Since Tyr phosphorylation by SRC facilitates CDK2o phosphorylate p27Kip1 at Thr187, resulting SCFSKP2-dependentegradation at S phase, SRC promotes proteolysis of p27Kip1 bothrior to and at S phase (Chu et al., 2007; Grimmler et al., 2007).
. Phosphorylation of Thr157 and Thr198 impairs nuclearmport resulting cytoplasmic localization
Nuclear import of p27Kip1 is also regulated by phosphoryla-
ion, with Thr157 and Thr198 phosphorylation preventing nuclearmport, resulting in cytoplasmic localization of p27Kip1. Comparedo phosphorylation on Ser10, which stimulates nuclear exportf p27Kip1 by CRM1, Thr157 and Thr198 phosphorylation is lessubiquitination and subsequent degradation. (C) Phosphorylation of p27Kip1 at Tyr74,
hereby restoring the cyclin-CDK catalytic activity, while p27Kip1 remains bound tofor 14-3-3 proteins, which can bind and sequester p27Kip1 in the cytoplasm.
common and is induced by serum stimulation (Jäkel et al., 2012). Inearly G1 phase, phosphorylation at Thr157 and Thr198 by AKT, p90ribosomal protein S6 kinases, serum and glucocorticoid-induciblekinase (SGK), AMPK and PIM (Morishita et al., 2008) cause a delayin nuclear import of p27Kip1 (Fig. 2D). Phosphorylation of Thr157
and Thr198 by AKT can form a recognition motif for 14-3-3 proteinsto sequester p27Kip1 in the cytosol of HeLa and human embryonickidney cells (Sekimoto et al., 2004). In addition, since Thr157 iswithin the nuclear localization signal of p27Kip1 (Sekimoto et al.,2004), it has been proposed that Thr157 phosphorylation mayinterfere with p27Kip1 nuclear import (Sekimoto et al., 2004).
9. Phosphorylation-independent degradation of p27Kip1 byPIRH2 in the nucleus and cytoplasm
p53-induced RING-H2-type ubiquitin ligase (PIRH2) is ap27Kip1-interacting protein. PIRH2 degrades p27Kip1 in G1 phaseindependent of phosphorylation state in both the nucleus and cyto-plasm (Hattori et al., 2007). PIRH2 interacts with p27Kip1 throughthe RING finger domain (Hattori et al., 2007). The expression of
PIRH2 is low in G0, but is induced in late G1 to S phase in humanglioblastoma (T98G) cells (Hattori et al., 2007). Knockdown ofPIRH2 thereby prevented the cell cycle re-entry of serum-starvedcells following the restoration of serum (Hattori et al., 2007).
l of Bio
Hd
1
ceuaiosdp2pHimrPop2(tcMegws
cssdsac2rCv
R
A
A
B
B
B
B
S.S.T. Hnit et al. / The International Journa
ence, PIRH2 is another negative regulator of p27Kip1 via ubiquitin-ependent proteasomal degradation.
0. Associated pathologies and therapeutic implication
Low protein levels or cytoplasmic expression of p27Kip1 is asso-iated with cancer aggressiveness and poor clinical prognosis (Chut al., 2008). The main cause of low p27Kip1 expression in cancer isnlikely via mutations or epigenetic silencing, but rather throughberrant p27Kip1 degradation, repressed p27Kip1 expression, andncreased cytoplasmic localization (Chu et al., 2008). High levelsf SKP2, KPC and PIRH2 together with low levels of p27Kip1 werehown in many human cancers, suggesting that accelerated degra-ation of p27Kip1 by E3 complexes is involved in carcinogenesis androgression (Bretones et al., 2011; Chu et al., 2008; Huang et al.,011; Kamura et al., 2004). In the presence of MYC, low levels of27Kip1 result in tumor formation in animals (Bretones et al., 2011).igh level of MYC also inhibit FOXO-mediated p27Kip1 expression
n many tumor cells (Chandramohan et al., 2008). miRNA-221 andiRNA-222 are frequently upregulated in tumor tissues, and cor-
elate with low levels of p27Kip1 in cancers (le Sage et al., 2007;ineau et al., 2010). Finally, cytoplasmic localization of p27Kip1 alsoccurs in cancers (Sekimoto et al., 2004). Since the nuclear export of27Kip1 is regulated by SRC (Chu et al., 2007) and CRM1 (Wang et al.,013), while the cytoplasmic-nuclear shuttling is blocked by PIMMorishita et al., 2008) and 14-3-3 proteins (Sekimoto et al., 2004),he increased levels of SRC, CRM1 and PIM in many cancers couldontribute to p27Kip1 cytoplasmic localization (Chu et al., 2007;orishita et al., 2008; Wang et al., 2013). Indeed, CRM1-positive
xpression in ovarian cancer was associated with higher tumorrade and lymph node metastases (Wang et al., 2013). Patientsith high Ser10 phosphorylated p27Kip1 had significantly shorter
urvival time (Wang et al., 2013).Since stability of p27Kip1 protein is a function of site spe-
ific phosphorylation, a strategy targeting the key phosphorylationites responsible for ubiquitination-mediated p27Kip1 degradationhould be considered. Benzoquinone ansamycin 17-allylamino gel-anamycin (17-AAG) is a proteasome inhibitor that is able totabilize p27Kip1 by depletion of Cks1, the bridge between SKP2nd p27Kip1 (Khattar et al., 2014). Treatment of colon carcinomaells with 17-AAG resulted in G1 cell cycle arrest (Khattar et al.,014). Restoring p27Kip1 levels by influencing its gene expressionegulators and rectifying p27Kip1 mislocalization via targeting SRC,RM1 and PIM, may also provide new avenue for treatment of aariety of cancers.
eferences
dhikari, D., Zheng, W., Shen, Y., Gorre, N., Ning, Y., Halet, G., Kaldis, P., Liu, K.,2012. Cdk1, but not Cdk2, is the sole Cdk that is essential and sufficient to driveresumption of meiosis in mouse oocytes. Hum. Mol. Genet. 21 (11),2476–2484, http://dx.doi.org/10.1093/hmg/dds061.
leem, E., Kiyokawa, H., Kaldis, P., 2005. Cdc2-cyclin E complexes regulate the G1/Sphase transition’. Nat. Cell Biol. 7 (8), 831–836, http://dx.doi.org/10.1038/ncb1284.
ahrami, A.R., Matin, M.M., Andrews PW, 2005. The CDK, inhibitor p27 enhancesneural differentiation in pluripotent NTERA2 human EC cells but does notpermit differentiation of 2102Ep nullipotent human EC cells. Mech. Dev. 122(9), 1034–1042, http://dx.doi.org/10.1016/j.mod.2005.04.011.
ondar, T., Kalinina, A., Khair, L., Kopanja, D., Nag, A., Bagchi, S., Raychaudhuri, P.,2006. Cul4A and DDB1 associate with Skp2 to target p27Kip1 for proteolysisinvolving the COP9 signalosome. Mol. Cell. Biol. 26 (7), 2531–2539, http://dx.doi.org/10.1128/mcb.26.7.2531-2539.2006.
retones, G., Acosta, J.C., Caraballo, J.M., Ferrandiz, N., Gomez-Casares, M.T.,Albajar, M., Blanco, R., Ruiz, P., Hung, W.C., Albero, M.P., Perez-Roger, I., Leon, J.,2011. SKP2 oncogene is a direct MYC target gene and MYC down-regulates
p27(KIP1) through SKP2 in human leukemia cells. J. Biol. Chem. 286 (11),9815–9825, http://dx.doi.org/10.1074/jbc.m110.165977.retones, G., Delgado, M.D., León, J., 2015. Myc and cell cycle control. BBA: GeneRegul. Mech. 1849 (5), 506–516, http://dx.doi.org/10.1016/j.bbagrm.2014.03.013.
chemistry & Cell Biology 68 (2015) 9–14 13
Chandramohan, V., Mineva, N.D., Burke, B., Jeay, S., Wu, M., Shen, J., Yang, W., Hann,S.R., Sonenshein, G.E., 2008. c-Myc represses FOXO3a-mediated transcriptionof the gene encoding the p27Kip1 cyclin dependent kinase inhibitor’. J. Cell.Biochem. 104 (6), 2091–2106, http://dx.doi.org/10.1002/jcb.21765.
Chu, I., Sun, J., Arnaout, A., Kahn, H., Hanna, W., Narod, S., Sun, P., Tan, C.K., Hengst,L., Slingerland, J., 2007. p27 phosphorylation by Src regulates inhibition ofcyclin E-Cdk2. Cell 128 (2), 281–294, http://dx.doi.org/10.1016/j.cell.2006.11.049.
Chu, I.M., Hengst, L., Slingerland, J.M., 2008. The Cdk inhibitor p27 in humancancer: prognostic potential and relevance to anticancer therapy. Nat. Rev.Cancer 8 (4), 253–267, http://dx.doi.org/10.1038/nrc2347.
Denicourt, C., Dowdy, S.F., 2004. Cip/Kip proteins: more than just CDKs inhibitors.Genes Dev. 18 (8), 851–855, http://dx.doi.org/10.1101/gad.1205304.
Fero, M.L., Rivkin, M., Tasch, M., Porter, P., Carow, C.E., Firpo, E., Polyak, K., Tsai, L.H.,Broudy, V., Perlmutter, R.M., Kaushansky, K., Roberts, J.M., 1996. A syndrome ofmultiorgan hyperplasia with features of gigantism, tumorigenesis, and femalesterility in p27(Kip1)-deficient mice. Cell 85 (5), 733–744, http://dx.doi.org/10.1016/s0092-8674(00)81239-8.
Gao, H., Ouyang, X., Banach-Petrosky, W., Borowsky, A.D., Lin, Y., Kim, M., Lee, H.,Shih, W.-J., Cardiff, R.D., Shen, M.M., 2004. A critical role for p27kip1 genedosage in a mouse model of prostate carcinogenesis. Proc. Natl. Acad. Sci. U. S.A. 101 (49), 17204–17209, http://dx.doi.org/10.1073/pnas.0407693101.
Grimmler, M., Wang, Y., Mund, T., Cilensek, Z., Keidel, E.M., Waddell, M.B., Jakel, H.,Kullmann, M., Kriwacki, R.W., Hengst, L., 2007. Cdk-inhibitory activity andstability of p27Kip1 are directly regulated by oncogenic tyrosine kinases. Cell128 (2), 269–280, http://dx.doi.org/10.1016/j.cell.2006.11.047.
Hao, B., Zheng, N., Schulman, B.A., Wu, G., Miller, J.J., Pagano, M., Pavletich, N.P.,2005. Structural basis of the Cks1-dependent recognition of p27(Kip1) by theSCF(Skp2) ubiquitin ligase. Mol. Cell 20 (1), 9–19, http://dx.doi.org/10.1016/j.molcel.2005.09.003.
Hattori, T., Isobe, T., Abe, K., Kikuchi, H., Kitagawa, K., Oda, T., Uchida, C., Kitagawa,M., 2007. Pirh2 promotes ubiquitin-dependent degradation of thecyclin-dependent kinase inhibitor p27Kip1. Cancer Res. 67 (22), 10789–10795,http://dx.doi.org/10.1158/0008-5472.can-07-2033.
Huang, X., Qian, X., Cheng, C., He, S., Sun, L., Ke, Q., Zhang, L., Pan, X., He, F., Wang,Q., 2011. Expression of Pirh2, a p27 Kip1 ubiquitin ligase, in hepatocellularcarcinoma: correlation with p27 Kip1 and cell proliferation. Hum. Pathol. 42(4), 507–515, http://dx.doi.org/10.1016/j.humpath.2010.04.021.
Ishida, N., Hara, T., Kamura, T., Yoshida, M., Nakayama, K., Nakayama, K.I., 2002.Phosphorylation of p27Kip1 on serine 10 is required for its binding to CRM1and nuclear export. J. Biol. Chem. 277 (17), 14355–14358, http://dx.doi.org/10.1074/jbc.c100762200.
Jäkel, H., Peschel, I., Kunze, C., Weinl, C., Hengst, L., 2012. Regulation of p27Kip1 bymitogen-induced tyrosine phosphorylation. Cell Cycle 11 (10), 1910–1917,http://dx.doi.org/10.4161/cc.19957.
Jäkel, H., Weinl, C., Hengst, L., 2011. Phosphorylation of p27Kip1 by JAK2 directlylinks cytokine receptor signaling to cell cycle control. Oncogene 30 (32),3502–3512, http://dx.doi.org/10.1038/onc.2011.68.
James, M.K., Ray, A., Leznova, D., Blain, S.W., 2008. Differential modification ofp27Kip1 controls its cyclin D-cdk4 inhibitory activity. Mol. Cell. Biol. 28 (1),498–510, http://dx.doi.org/10.1128/mcb.02171-06.
Kamura, T., Hara, T., Matsumoto, M., Ishida, N., Okumura, F., Hatakeyama, S.,Yoshida, M., Nakayama, K., Nakayama, K.I., 2004. Cytoplasmic ubiquitin ligaseKPC regulates proteolysis of p27(Kip1) at G1 phase. Nat. Cell Biol. 6 (12),1229–1235, http://dx.doi.org/10.1038/ncb1194.
Karnik, S.K., Hughes, C.M., Gu, X., Rozenblatt-Rosen, O., McLean, G.W., Xiong, Y.,Meyerson, M., Kim, S.K., 2005. Menin regulates pancreatic islet growth bypromoting histone methylation and expression of genes encoding p27Kip1 andp18INK4c. Proc. Natl. Acad. Sci. U. S. A. 102 (41), 14659–14664, http://dx.doi.org/10.1073/pnas.0503484102.
Khattar, V., Fried, J., Xu, B., Thottassery, J.V., 2014. Cks1 proteasomal degradation isinduced by inhibiting Hsp90-mediated chaperoning in cancer cells. CancerChemother. Pharmacol., 1–10, http://dx.doi.org/10.1007/s00280-014-2666-7.
Kotoshiba, S., Kamura, T., Hara, T., Ishida, N., Nakayama, K.I., 2005. Moleculardissection of the interaction between p27 and Kip1 ubiquitylation-promotingcomplex, the ubiquitin ligase that regulates proteolysis of p27 in G1 phase. J.Biol. Chem. 280 (18), 17694–17700, http://dx.doi.org/10.1074/jbc.m500866200.
le Sage, C., Nagel, R., Egan, D.A., Schrier, M., Mesman, E., Mangiola, A., Anile, C.,Maira, G., Mercatelli, N., Ciafre, S.A., Farace, M.G., Agami, R., 2007. Regulation ofthe p27(Kip1) tumor suppressor by miR-221 and miR-222 promotes cancercell proliferation. EMBO J. 26 (15), 3699–3708, http://dx.doi.org/10.1038/sj.emboj.7601790.
McAllister, S.S., Becker-Hapak, M., Pintucci, G., Pagano, M., Dowdy, S.F., 2003. Novelp27kip1 C-terminal scatter domain mediates Rac-dependent cell migrationindependent of cell cycle arrest functions. Mol. Cell. Biol. 23 (1), 216–228,http://dx.doi.org/10.1128/mcb.23.1.216-228.2003.
Morishita, D., Katayama, R., Sekimizu, K., Tsuruo, T., Fujita, N., 2008. Pim kinasespromote cell cycle progression by phosphorylating and down-regulatingp27Kip1 at the transcriptional and posttranscriptional levels. Cancer Res. 68
(13), 5076–5085, http://dx.doi.org/10.1158/0008-5472.can-08-0634.Muller, D., Bouchard, C., Rudolph, B., Steiner, P., Stuckmann, I., Saffrich, R., Ansorge,W., Huttner, W., Eilers, M., 1997. Cdk2-dependent phosphorylation of p27facilitates its Myc-induced release from cyclin E/cdk2 complexes. Oncogene 15(21), 2561–2576, http://dx.doi.org/10.1038/sj.onc.1201440.

1 l of Bio
M
O
P
P
P
P
R
4 S.S.T. Hnit et al. / The International Journa
unoz-Alonso, M.J., Acosta, J.C., Richard, C., Delgado, M.D., Sedivy, J., Leon, J., 2005.p21Cip1 and p27Kip1 induce distinct cell cycle effects and differentiationprograms in myeloid leukemia cells. J. Biol. Chem. 280 (18), 18120–18129,http://dx.doi.org/10.1074/jbc.m500758200.
’Hagan, R.C., Ohh, M., David, G., de Alboran, I.M., Alt, F.W., Kaelin Jr., W.G.,DePinho, R.A., 2000. Myc-enhanced expression of Cul1 promotesubiquitin-dependent proteolysis and cell cycle progression. Genes Dev. 14(17), 2185–2191, http://dx.doi.org/10.1101/gad.827200.
érez-Roger, I., Solomon, D., Sewing, A., Land, H., 1997. Myc activation of cyclinE/Cdk2 kinase involves induction of cyclin E gene transcription and inhibitionof p27 (Kip1) binding to newly formed complexes. Oncogene 14 (20),2373–2381, http://dx.doi.org/10.1038/sj.onc.1201197.
ietenpol, J.A., Bohlander, S.K., Sato, Y., Papadopoulos, N., Liu, B., Friedman, C.,Trask, B.J., Roberts, J.M., Kinzler, K.W., Rowley, J.D., et al., 1995. Assignment ofthe human p27Kip1 gene to 12p13 and its analysis in leukemias. Cancer Res.55 (6), 1206–1210, http://www.ncbi.nlm.nih.gov/pubmed/7882309
ineau, P., Volinia, S., McJunkin, K., Marchio, A., Battiston, C., Terris, B., Mazzaferro,V., Lowe, S.W., Croce, C.M., Dejean A, 2010. miR-221 overexpressioncontributes to liver tumorigenesis. Proc. Natl. Acad. Sci. U. S. A. 107 (1),264–269, http://dx.doi.org/10.1073/pnas.0907904107.
olyak, K., Kato, J.Y., Solomon, M.J., Sherr, C.J., Massague, J., Roberts, J.M., Koff, A.,1994. p27Kip1, a cyclin-Cdk inhibitor, links transforming growth factor-beta
and contact inhibition to cell cycle arrest. Genes Dev. 8 (1), 9–22, http://dx.doi.org/10.1101/gad.8.1.9.ay, A., James, M.K., Larochelle, S., Fisher, R.P., Blain, S.W., 2009. p27Kip1 inhibitscyclin D-cyclin-dependent kinase 4 by two independent modes. Mol. Cell. Biol.29 (4), 986–999, http://dx.doi.org/10.1128/mcb.00898-08.
chemistry & Cell Biology 68 (2015) 9–14
Russo, A.A., Jeffrey, P.D., Patten, A.K., Massagué, J., Pavletich, N.P., 1996. Crystalstructure of the p27Kip1 cyclin-dependent-kinase inhibitor bound to thecyclin A-Cdk2 complex. Nature 382 (6589), 325–331, http://dx.doi.org/10.1038/382325a0.
Santamaria, D., Barriere, C., Cerqueira, A., Hunt, S., Tardy, C., Newton, K., Caceres,J.F., Dubus, P., Malumbres, M., Barbacid, M., 2007. Cdk1 is sufficient to drive themammalian cell cycle. Nature 448 (7155), 811–815, http://dx.doi.org/10.1038/nature06046.
Sekimoto, T., Fukumoto, M., Yoneda, Y., 2004. 14-3-3 suppresses the nuclearlocalization of threonine 157-phosphorylated p27Kip1. EMBO J. 23 (9),1934–1942, http://dx.doi.org/10.1038/sj.emboj.7600198.
Sheaff, R.J., Groudine, M., Gordon, M., Roberts, J.M., Clurman, B.E., 1997. CyclinE-CDK2 is a regulator of p27Kip1. Genes Dev. 11 (11), 1464–1478, http://dx.doi.org/10.1101/gad.11.11.1464.
Tsvetkov, L.M., Yeh, K.-H., Lee, S.-J., Sun, H., Zhang, H., 1999. p27 Kip1ubiquitination and degradation is regulated by the SCF Skp2 complex throughphosphorylated Thr187 in p27. Curr. Biol. 9 (12), 661–664, http://dx.doi.org/10.1016/s0960-9822(99)80290-5.
Wang, Y., Wang, Y., Xiang, J., Ji, F., Deng, Y., Tang, C., Yang, S., Xi, Q., Liu, R., Di, W.,2013. Knockdown of CRM1 inhibits the nuclear export of p27(Kip1)phosphorylated at serine 10 and plays a role in the pathogenesis of epithelialovarian cancer. Cancer Lett. 343 (1), 6–13, http://dx.doi.org/10.1016/j.canlet.2013.09.002.
Yang, W., Shen, J., Wu, M., Arsura, M., FitzGerald, M., Suldan, Z., Kim, D.W.,Hofmann, C.S., Pianetti, S., Romieu-Mourez, R., Freedman, L.P., Soneshein, G.E.,2001. Repression of transcription of the p 27 Kip 1 cyclin-dependent kinaseinhibitor gene by c-Myc. Oncogene 20 (14), 1688–1702, http://dx.doi.org/10.1038/sj.onc.1204245.