overexpression of a tobacco j-domain protein enhances drought tolerance in transgenic arabidopsis
TRANSCRIPT

lable at ScienceDirect
Plant Physiology and Biochemistry 83 (2014) 100e106
Contents lists avai
Plant Physiology and Biochemistry
journal homepage: www.elsevier .com/locate/plaphy
Research article
Overexpression of a tobacco J-domain protein enhances droughttolerance in transgenic Arabidopsis
Zongliang Xia a, *, 1, Xiaoquan Zhang a, 1, Junqi Li a, Xinhong Su b, Jianjun Liu c
a Henan Agricultural University, Zhengzhou 450002, PR Chinab Henan Tobacco Company, Zhengzhou 450008, PR Chinac Zhengzhou Branch, Henan Tobacco Company, Zhengzhou 450001, PR China
a r t i c l e i n f o
Article history:Received 6 April 2014Accepted 27 July 2014Available online 6 August 2014
Keywords:DroughtGene expressionJ-domain proteinTobacco
Abbreviations: DnaJ, J-protein; Hsp40, 40 kDa heHistidine, proline, aspartate motif; MS, Murashige anPCR.* Corresponding author. College of Life Science, H
Zhengzhou 450002, PR China. Tel.: þ86 371 6357967E-mail address: [email protected] (Z. Xia).
1 Contributed equally.
http://dx.doi.org/10.1016/j.plaphy.2014.07.0230981-9428/© 2014 Elsevier Masson SAS. All rights re
a b s t r a c t
DnaJ proteins constitute a DnaJ/Hsp40 family and are important regulators involved in diverse cellularfunctions. To date, the molecular mechanisms of DnaJ proteins involved in response to drought stress inplants are largely unknown. In this study, a putative DnaJ ortholog from Nicotiana tabacum (NtDnaJ1),which encodes a putative type-I J-protein, was isolated. The transcript levels of NtDnaJ1 were higher inaerial tissues and were markedly up-regulated by drought stress. Over-expression of NtDnaJ1 in Arabi-dopsis plants enhanced their tolerance to osmotic or drought stress. Quantitative determination of H2O2
accumulation has shown that H2O2 content increased in wild-type and transgenic seedlings under os-motic stress, but was significantly lower in both transgenic lines compared with the wild-type.Expression analysis of stress-responsive genes in NtDnaJ1-transgenic Arabidopsis revealed that therewas significantly increased expression of genes involved in the ABA-dependent signaling pathway(AtRD20, AtRD22 and AtAREB2) and antioxidant genes (AtSOD1, AtSOD2, and AtCAT1). Collectively, thesedata demonstrate that NtDnaJ1 could be involved in drought stress response and its over-expressionenhances drought tolerance possibly through regulating expression of stress-responsive genes. Thisstudy may facilitate our understandings of the biological roles of DnaJ protein-mediated abiotic stress inhigher plants and accelerate genetic improvement of crop plants tolerant to environmental stresses.
© 2014 Elsevier Masson SAS. All rights reserved.
1. Introduction
J-domain proteins (also called DnaJ proteins) constitute a DnaJ/Hsp40 family and are conserved co-chaperones for HSP70s (Caplanet al., 1993; Silver andWay,1993; Qiu et al., 2006). DnaJ proteins areinvolved in a variety of essential cellular processes including pro-tein folding, assembly, translocation, degradation, stabilization andrefolding (Wang et al., 2004; Mayer and Bukau, 2005; Craig et al.,2006; Rajan and D’Silva, 2009). Besides their coechaperone activ-ity, DnaJ proteins function as protein disulfide isomerases to cata-lyze protein disulfide formation, reduction, and isomerization (deCrouy-Chanel et al., 1995).
at shock protein; HPD motif,d skoog; qPCR, Quantitative
enan Agricultural University,6; fax: þ86 371 63555790.
served.
DnaJ was originally identified in Escherichia coli as a 41 kDa heatshock protein (Goffin and Georgopoulos, 1998). Subsequently,members of the J-protein family were found to function as mo-lecular chaperones by binding Hsp70 to stimulate ATP hydrolysis,and stabilizing the Hsp70 interaction with substrate proteins(Szyperski et al., 1994; Szabo et al., 1996; Cheetham and Caplan,1998). The J domain is a highly conserved ~70 amino acid a-heli-cal region in DnaJ proteins (Hennessy et al., 2005). Plant J-domainproteins have been classified into four types (I, II, III, and IV) basedon the presence of other conserved domains (Rajan and D’Silva,2009; Miernyk, 2001; Walsh et al., 2004). Traditional type-I J-domain proteins contain four domains including a J domain, a Gly/Phe (G/F) domain, a CXXCXGXG zinc-finger domain and a lessconserved C-terminal domain. Type-II J proteins lack the zinc-finger domain, whereas type-III J- proteins contain only the Jdomain. Type-IV J proteins have been recently described and clas-sified as ‘J-like proteins’, with significant sequence and structuralsimilarities with the J domain, but they lack the HPD motif (Walshet al., 2004). The well-characterized Arabidopsis genome is knownto harbor 120 predicted J-domain proteins (Rajan and D’Silva,2009).

Z. Xia et al. / Plant Physiology and Biochemistry 83 (2014) 100e106 101
Increasing evidence has shown that J-domain proteins playimportant roles in growth and development (Kneissl et al., 2009;Yang et al., 2009; Shen et al., 2011; Bekh-Ochir et al., 2013), dis-ease resistance (Bekh-Ochir et al., 2013; Liu andWhitham, 2013; Duet al., 2013), and abiotic stress responses (Yang et al., 2010; Zhouet al., 2012). For example, the Arabidopsis J-domain protein OWL1is involved in the regulation of germination, cotyledon opening,hypocotyl elongation, and perception of very low light influences(Kneissl et al., 2009). TMS1, a DnaJ proteinwith disulfide isomeraseactivity, is required for thermotolerance of pollen tubes in Arabi-dopsis, possibly by functioning as a co-molecular chaperone (Yanget al., 2009). The Arabidopsis Type-I J-domain protein J3 mediatesthe integration of flowering signals during the floral transition andregulates the plasma membrane Hþ-ATPase through interactingwith the PKS5 kinase under high salinity conditions (Yang et al.,2009; Shen et al., 2011). The J-protein AtDjB1 facilitates thermo-tolerance by protecting cells against heat-induced oxidative dam-age in Arabidopsis (Zhou et al., 2012). Recently, it has been reportedthat a type-III J domain protein GmHSP40.1 causes HR-like celldeath in tobacco (Liu and Whitham, 2013). More interestingly, atobacco type-I J-domain protein NbMIP1 has been shown to berequired for both tobacco mosaic virus infection and plant innateimmunity through functioning as co-chaperones (Du et al., 2013).
In spite of progress made in understanding function of DnaJproteins, few reports have been concerned the role of J-domainproteins in drought stress in plants (Wang et al., 2014). Droughtstress often adversely affects plant growth and productivity, thus itis still a serious problem in agriculture worldwide. Tobacco is animportant crop as well as amodel plant system, and its productivityis vulnerable to drought. To identify genes important in droughtstress response in tobacco, we previously identified mRNAs up-regulated by drought stress through microarray analysis (unpub-lished data). One highly induced mRNA encoding a J-domain pro-tein (NtDnaJ1) was characterized in detail. We further characterizedthe putative NtDnaJ1 in transgenic Arabidopsis to investigatedrought tolerance and possible function mechanisms.
2. Materials and methods
2.1. Plant materials and stress treatment
Tobacco (Nicotiana tabacum cv. Xanthi) was used throughoutthis study. Arabidopsis thaliana ecotype Col-0 was used for genetransformation. Plants were grown in a growth room as describedpreviously (Xia et al., 2012a). Drought stress in four-week-oldplants was realized by replacing the water with 15% PEG 6000and leaves were sampled at 0, 6, 12, 24, or 48 h for expressionanalysis as described by us (Xia et al., 2013).
2.2. Cloning of NtDnaJ1 and sequence analysis
The drought-induced EST encoding a putative DnaJ/Hsp40 pro-tein was used to do BLAST (http://www.ncbi.nlm.nih.gov/) andmRNA sequences containing such an EST were downloaded forgene prediction. The gene is highly homologous to DnaJ/Hsp40family member, and thus is named NtDnaJ1. Two primers NtDnaJ1-F and NtDnaJ1-R (Table S1) were designed for amplifying the openreading frame (ORF) of NtDnaJ1. The 1257 bp PCR product wasverified by sequencing.
The primary structural analysis was performed using Inter-ProScan (http://www.ebi.ac.uk/InterProScan). The alignment of thededuced protein sequences and phylogenetic tree analyses weredone by DNASTAR and MEGA 5.1, respectively, using standard pa-rameters (Tamura et al., 2011).
2.3. Real-time PCR analysis
Real-time PCR was used to determine the expression patterns ofNtDnaJ1 in different organs and under drought condition. The qRT-PCR was performed in triplicate with an IQ5 light cycler system(Bio-Rad) using SYBR Premix ExTaq II (Takara, Japan) with gene-specific primers NtDnaJ1-F1 and NtDnaJ1-R1 (Table S1), whichproduces a 195-bp product. The tobacco NtActin transcript wasused as an internal control to quantify the relative transcript levelsas described by us previously (Xia et al., 2013). The relative level ofgene expression was detected using the 2�DDC
T method (Livak andSchmittgen, 2001).
To examine the relative expression of NtDnaJ1 in transgenicArabidopsis plants, the AtActin2 transcript (gene-specific primersAtActin2-F1 and AtActin2-R1; Table S1) was used as an internalcontrol to quantify the expression levels, and the lowest expres-sion level among transgenic lines was regarded as standard andthe relative level of gene expression was computed as describedabove.
To assay the expression of stress-responsive genes (AtSOD1,AtSOD2, AtCAT1, AtRD20, AtRD22 and AtAREB2) in transgenic Ara-bidopsis plants, qRT-PCR analysis was also performed with the RNAsamples isolated from four-week-old transgenic plants harvested insoil under normal conditions. Total RNA isolation and reversetranscription were performed as described above. PCR amplifica-tion was performed with gene specific primers (Table S1). Criteriafor designing primers were a primer size between 22 and 25, anoptimal Tm at 60 �C, and a product size ranging from 150 bp to250 bp. Amplification of AtActin2 was used as an internal control,and qRT-PCR experimental procedures were performed asdescribed above. For the entire qRT-PCR assay, three technicalreplicates were performed for each experiment and the expressionof each gene was investigated in three biological replicates.
2.4. Construction of plant expression vectors and development oftransgenic Arabidopsis lines
TheNtDnaJ1 coding sequencewas amplified and introduced intothe pART7 plasmid (Xia et al., 2012b) using primers NtDnaJ1-F2with BamHI restriction site (underlined) and NtDnaJ1-R2 with XbaIrestriction site (underlined) (Table S1) and was subsequentlyinserted downstream of the 35S promoter in the plasmid vectorpART7. The resulting expression cassette containing the 35S pro-moter and NtDnaJ1 coding sequence was cut and inserted into thebinary vector pART27, producing the transformation constructpART27-35S-NtDnaJ1.
The binary construct was introduced into Agrobacterium tume-faciens (strain GV3101) and then transformed into Arabidopsis (Col-0) via the floral dip method (Clough and Bent, 1998). Transgeniclines were selected by germinating seeds on medium containingMurashige and Skoog (MS) basal salt mixture (SigmaeAldrich, USA)and 50 mg/L kanamycin. After two weeks on selection medium,green seedlings (T1 plants) were transferred to soil pots and growntomaturity in a growth room. The presence of the transgene in eachplant was checked by PCR with genomic DNA from leaves of indi-vidual plant using primers 35SP-F and NtDnaJ1-R3 (the forwardprimer 35SP-F is from CaMV 35S promoter sequence) (Table S1).The PCR-positive plants as transgenes were grown to maturity andseeds were collected (T2 seed). T2 seeds were germinated onkanamycin selection medium again and the one-copy lines wereidentified by examining the segregation ratio (3:1) of the kana-mycin selectable marker. Each one-copy line was maintainedgrowth to set seeds until T3 generation. Five independent homo-zygous NtDnaJ1 transgenic lines (named OE-8, OE-14, OE-18, OE-22and OE-27) were developed. The expression levels of the transgene
Z. Xia et al. / Plant Physiology and Biochemistry 83 (2014) 100e106102
among the five lines were evaluated by qRT-PCR with the primersNtDnaJ1-F1 and NtDnaJ1-R1 (Table S1) as described above.
2.5. Analysis of NtDnaJ1-overexpressing Arabidopsis for osmoticand drought stress tolerance
For osmotic stress tolerance analysis, surface-sterilized seeds ofboth transgenic Arabidopsis lines (OE-18 and OE-22) or wild-typewere germinated and cultured on MS medium for 7 days, andthen the seedlings (30e40) were transferred to grow on MS me-dium supplemented with 0 or 250 mM mannitol in the growthchamber. The phenotype of seedlings was photographed after 7days of growth. The experiment was repeated at least three times.Survival rates (%) under mannitol treatment were determined asthe number of visibly green plants after 7 days.
For drought tolerance analysis, four-week-old plants were sub-jected to progressive drought by withholding water until a nearlylethal effect of dehydration (about 3 weeks) was observed. Thedrought stress experiment was performed at least three times.Seven days after re-watering, survival rates (%) under droughtstress were determined as described above.
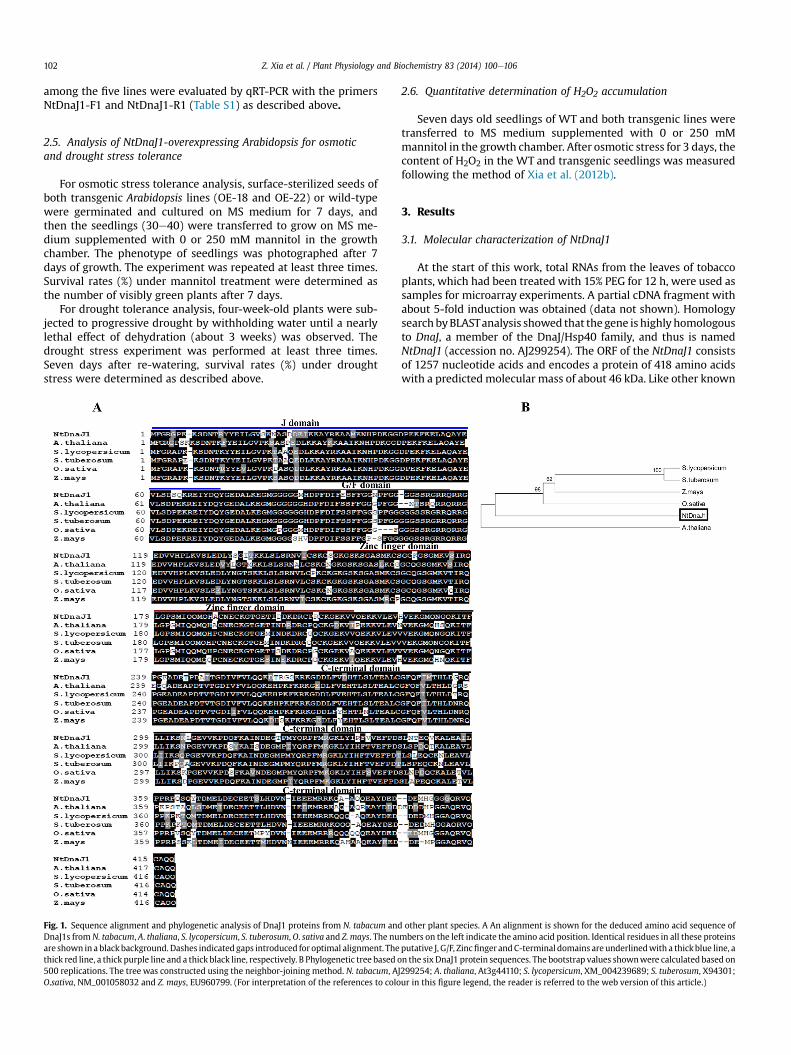
Fig. 1. Sequence alignment and phylogenetic analysis of DnaJ1 proteins from N. tabacum andDnaJ1s from N. tabacum, A. thaliana, S. lycopersicum, S. tuberosum, O. sativa and Z. mays. The nuare shown in a blackbackground. Dashes indicated gaps introduced for optimal alignment. Thethick red line, a thick purple line and a thick black line, respectively. B Phylogenetic tree based o500 replications. The tree was constructed using the neighbor-joining method. N. tabacum, AO.sativa, NM_001058032 and Z. mays, EU960799. (For interpretation of the references to colo
2.6. Quantitative determination of H2O2 accumulation
Seven days old seedlings of WT and both transgenic lines weretransferred to MS medium supplemented with 0 or 250 mMmannitol in the growth chamber. After osmotic stress for 3 days, thecontent of H2O2 in the WT and transgenic seedlings was measuredfollowing the method of Xia et al. (2012b).
3. Results
3.1. Molecular characterization of NtDnaJ1
At the start of this work, total RNAs from the leaves of tobaccoplants, which had been treated with 15% PEG for 12 h, were used assamples for microarray experiments. A partial cDNA fragment withabout 5-fold induction was obtained (data not shown). Homologysearch byBLASTanalysis showed that the gene is highly homologousto DnaJ, a member of the DnaJ/Hsp40 family, and thus is namedNtDnaJ1 (accession no. AJ299254). The ORF of the NtDnaJ1 consistsof 1257 nucleotide acids and encodes a protein of 418 amino acidswith a predictedmolecular mass of about 46 kDa. Like other known
other plant species. A An alignment is shown for the deduced amino acid sequence ofmbers on the left indicate the amino acid position. Identical residues in all these proteinsputative J, G/F, Zincfinger andC-terminal domains are underlinedwith a thickblue line, an the six DnaJ1 protein sequences. The bootstrap values shownwere calculated based onJ299254; A. thaliana, At3g44110; S. lycopersicum, XM_004239689; S. tuberosum, X94301;ur in this figure legend, the reader is referred to the web version of this article.)
Z. Xia et al. / Plant Physiology and Biochemistry 83 (2014) 100e106 103
type-I DnaJ proteins, the deduced amino acid sequence of theNtDnaJ1 exhibits typical structural characteristics with four func-tional domains, including a J domain (residues 1e72), a Gly/Phe (G/F) domain (residues 82e115), a CXXCXGXG zinc-finger domain(residues 148e216) and a less conserved C-terminal domain (resi-dues 229e418) (Fig. 1A). Amino acid sequence comparisons haverevealed that NtDnaJ1 exhibits high identity to counterpart proteinsfrom A. thaliana (79.1% identity), Solanum lycopersicon (84.3%identity), Symphytum tuberosum (84% identity), Oryza sativa (83.5%identity), and Zea mays (82.4% identity) (Fig. 1A).
A phylogenetic tree was established based on DnaJ1 proteinsequences available in GenBank from 6 plant species (Arabidopsis,tobacco, tomato, potato, rice, and maize) (Fig. 1B). As shown inFig. 1B, interestingly, DnaJ from Arabidopsis formed a subgroupdistinct from the other DnaJs subgroup, including the solanaceaeDnaJs from S. Lycopersicon, S.tuberosum and N. tabacum, and themonocot DnaJs from Oryza sativa and Z.mays. The NtDnaJ1 showedhigher identities with DnaJ proteins from tomato, potato, rice, andmaize, and thus was clustered into the same isoform subgroup. Thesolanaceae DnaJs from S. lycopersicon and S. tuberosum were clus-tered into the same isoform subgroup. These results clearlydemonstrated that NtDnaJ1 shares basic structural feature similarto the known DnaJ proteins from Arabidopsis and other crop plants,thus it could be identified as an ortholog of DnaJ protein.
3.2. Transcript levels of NtDnaJ1 in various organs of tobacco andits responses to PEG-induced water stress
Transcriptional patterns of NtDnaJ1 were examined in five or-gans (roots, stems, leaves, flowers, and immature fruits) by qRT-PCR. As shown in Fig. 2A, NtDnaJ1 mRNA was detected in roots,stems, leaves, flowers, and immature fruits. The NtDnaJ1 transcriptlevels were significantly high in leaves, flowers and stems. Incontrast, NtDnaJ1 transcripts were low in roots and immature fruits
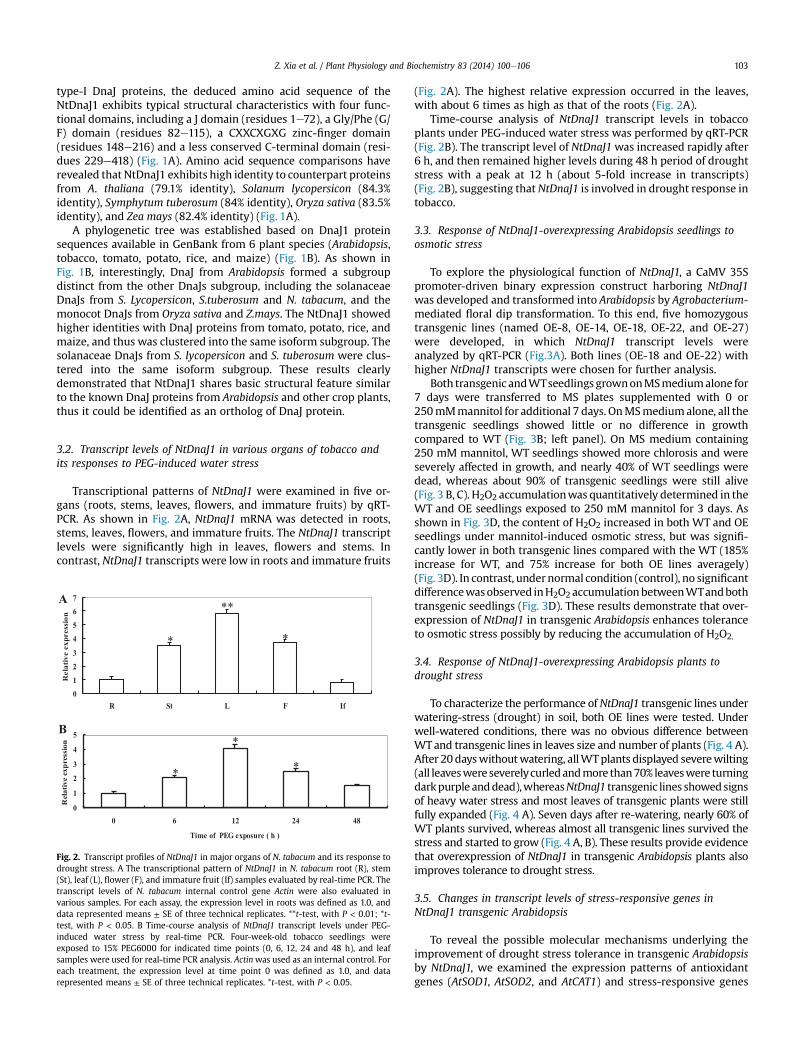
Fig. 2. Transcript profiles of NtDnaJ1 in major organs of N. tabacum and its response todrought stress. A The transcriptional pattern of NtDnaJ1 in N. tabacum root (R), stem(St), leaf (L), flower (F), and immature fruit (If) samples evaluated by real-time PCR. Thetranscript levels of N. tabacum internal control gene Actin were also evaluated invarious samples. For each assay, the expression level in roots was defined as 1.0, anddata represented means ± SE of three technical replicates. **t-test, with P < 0.01; *t-test, with P < 0.05. B Time-course analysis of NtDnaJ1 transcript levels under PEG-induced water stress by real-time PCR. Four-week-old tobacco seedlings wereexposed to 15% PEG6000 for indicated time points (0, 6, 12, 24 and 48 h), and leafsamples were used for real-time PCR analysis. Actinwas used as an internal control. Foreach treatment, the expression level at time point 0 was defined as 1.0, and datarepresented means ± SE of three technical replicates. *t-test, with P < 0.05.
(Fig. 2A). The highest relative expression occurred in the leaves,with about 6 times as high as that of the roots (Fig. 2A).
Time-course analysis of NtDnaJ1 transcript levels in tobaccoplants under PEG-induced water stress was performed by qRT-PCR(Fig. 2B). The transcript level of NtDnaJ1was increased rapidly after6 h, and then remained higher levels during 48 h period of droughtstress with a peak at 12 h (about 5-fold increase in transcripts)(Fig. 2B), suggesting that NtDnaJ1 is involved in drought response intobacco.
3.3. Response of NtDnaJ1-overexpressing Arabidopsis seedlings toosmotic stress
To explore the physiological function of NtDnaJ1, a CaMV 35Spromoter-driven binary expression construct harboring NtDnaJ1was developed and transformed into Arabidopsis by Agrobacterium-mediated floral dip transformation. To this end, five homozygoustransgenic lines (named OE-8, OE-14, OE-18, OE-22, and OE-27)were developed, in which NtDnaJ1 transcript levels wereanalyzed by qRT-PCR (Fig.3A). Both lines (OE-18 and OE-22) withhigher NtDnaJ1 transcripts were chosen for further analysis.
Both transgenic andWTseedlings grownonMSmediumalone for7 days were transferred to MS plates supplemented with 0 or250mMmannitol for additional 7 days. OnMSmediumalone, all thetransgenic seedlings showed little or no difference in growthcompared to WT (Fig. 3B; left panel). On MS medium containing250 mM mannitol, WT seedlings showed more chlorosis and wereseverely affected in growth, and nearly 40% of WT seedlings weredead, whereas about 90% of transgenic seedlings were still alive(Fig. 3 B, C). H2O2 accumulationwas quantitatively determined in theWT and OE seedlings exposed to 250 mM mannitol for 3 days. Asshown in Fig. 3D, the content of H2O2 increased in both WT and OEseedlings under mannitol-induced osmotic stress, but was signifi-cantly lower in both transgenic lines compared with the WT (185%increase for WT, and 75% increase for both OE lines averagely)(Fig. 3D). In contrast, undernormal condition (control), no significantdifferencewasobserved inH2O2accumulationbetweenWTandbothtransgenic seedlings (Fig. 3D). These results demonstrate that over-expression of NtDnaJ1 in transgenic Arabidopsis enhances toleranceto osmotic stress possibly by reducing the accumulation of H2O2.
3.4. Response of NtDnaJ1-overexpressing Arabidopsis plants todrought stress
To characterize the performance ofNtDnaJ1 transgenic lines underwatering-stress (drought) in soil, both OE lines were tested. Underwell-watered conditions, there was no obvious difference betweenWTand transgenic lines in leaves size and number of plants (Fig. 4 A).After 20dayswithoutwatering, allWTplants displayed severewilting(all leaveswere severelycurledandmore than70% leaveswere turningdarkpurple anddead),whereasNtDnaJ1 transgenic lines showedsignsof heavy water stress and most leaves of transgenic plants were stillfully expanded (Fig. 4 A). Seven days after re-watering, nearly 60% ofWT plants survived, whereas almost all transgenic lines survived thestress and started to grow (Fig. 4 A, B). These results provide evidencethat overexpression of NtDnaJ1 in transgenic Arabidopsis plants alsoimproves tolerance to drought stress.
3.5. Changes in transcript levels of stress-responsive genes inNtDnaJ1 transgenic Arabidopsis
To reveal the possible molecular mechanisms underlying theimprovement of drought stress tolerance in transgenic Arabidopsisby NtDnaJ1, we examined the expression patterns of antioxidantgenes (AtSOD1, AtSOD2, and AtCAT1) and stress-responsive genes
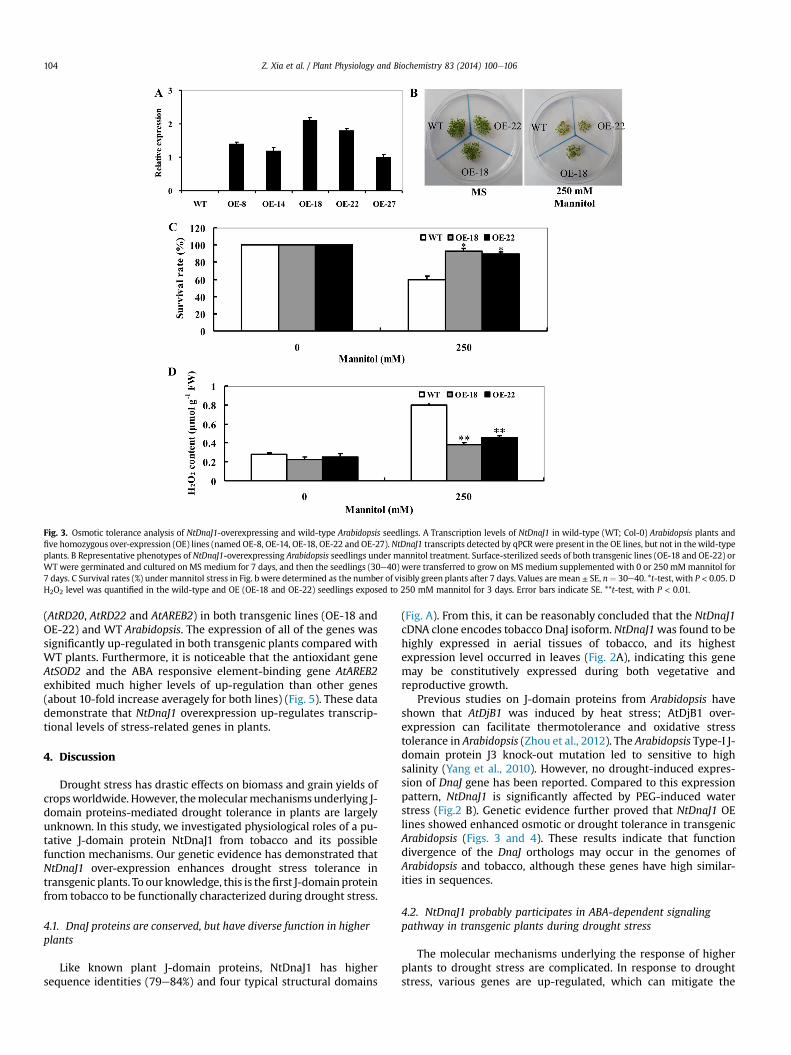
Fig. 3. Osmotic tolerance analysis of NtDnaJ1-overexpressing and wild-type Arabidopsis seedlings. A Transcription levels of NtDnaJ1 in wild-type (WT; Col-0) Arabidopsis plants andfive homozygous over-expression (OE) lines (named OE-8, OE-14, OE-18, OE-22 and OE-27). NtDnaJ1 transcripts detected by qPCR were present in the OE lines, but not in thewild-typeplants. B Representative phenotypes of NtDnaJ1-overexpressing Arabidopsis seedlings under mannitol treatment. Surface-sterilized seeds of both transgenic lines (OE-18 and OE-22) orWT were germinated and cultured on MS medium for 7 days, and then the seedlings (30e40) were transferred to grow on MS medium supplemented with 0 or 250 mMmannitol for7 days. C Survival rates (%) under mannitol stress in Fig. b were determined as the number of visibly green plants after 7 days. Values are mean ± SE, n¼ 30e40. *t-test, with P < 0.05. DH2O2 level was quantified in the wild-type and OE (OE-18 and OE-22) seedlings exposed to 250 mM mannitol for 3 days. Error bars indicate SE. **t-test, with P < 0.01.
Z. Xia et al. / Plant Physiology and Biochemistry 83 (2014) 100e106104
(AtRD20, AtRD22 and AtAREB2) in both transgenic lines (OE-18 andOE-22) and WT Arabidopsis. The expression of all of the genes wassignificantly up-regulated in both transgenic plants compared withWT plants. Furthermore, it is noticeable that the antioxidant geneAtSOD2 and the ABA responsive element-binding gene AtAREB2exhibited much higher levels of up-regulation than other genes(about 10-fold increase averagely for both lines) (Fig. 5). These datademonstrate that NtDnaJ1 overexpression up-regulates transcrip-tional levels of stress-related genes in plants.
4. Discussion
Drought stress has drastic effects on biomass and grain yields ofcropsworldwide. However, themolecularmechanismsunderlying J-domain proteins-mediated drought tolerance in plants are largelyunknown. In this study, we investigated physiological roles of a pu-tative J-domain protein NtDnaJ1 from tobacco and its possiblefunction mechanisms. Our genetic evidence has demonstrated thatNtDnaJ1 over-expression enhances drought stress tolerance intransgenic plants. To our knowledge, this is thefirst J-domainproteinfrom tobacco to be functionally characterized during drought stress.
4.1. DnaJ proteins are conserved, but have diverse function in higherplants
Like known plant J-domain proteins, NtDnaJ1 has highersequence identities (79e84%) and four typical structural domains
(Fig. A). From this, it can be reasonably concluded that the NtDnaJ1cDNA clone encodes tobacco DnaJ isoform. NtDnaJ1was found to behighly expressed in aerial tissues of tobacco, and its highestexpression level occurred in leaves (Fig. 2A), indicating this genemay be constitutively expressed during both vegetative andreproductive growth.
Previous studies on J-domain proteins from Arabidopsis haveshown that AtDjB1 was induced by heat stress; AtDjB1 over-expression can facilitate thermotolerance and oxidative stresstolerance in Arabidopsis (Zhou et al., 2012). The Arabidopsis Type-I J-domain protein J3 knock-out mutation led to sensitive to highsalinity (Yang et al., 2010). However, no drought-induced expres-sion of DnaJ gene has been reported. Compared to this expressionpattern, NtDnaJ1 is significantly affected by PEG-induced waterstress (Fig.2 B). Genetic evidence further proved that NtDnaJ1 OElines showed enhanced osmotic or drought tolerance in transgenicArabidopsis (Figs. 3 and 4). These results indicate that functiondivergence of the DnaJ orthologs may occur in the genomes ofArabidopsis and tobacco, although these genes have high similar-ities in sequences.
4.2. NtDnaJ1 probably participates in ABA-dependent signalingpathway in transgenic plants during drought stress
The molecular mechanisms underlying the response of higherplants to drought stress are complicated. In response to droughtstress, various genes are up-regulated, which can mitigate the
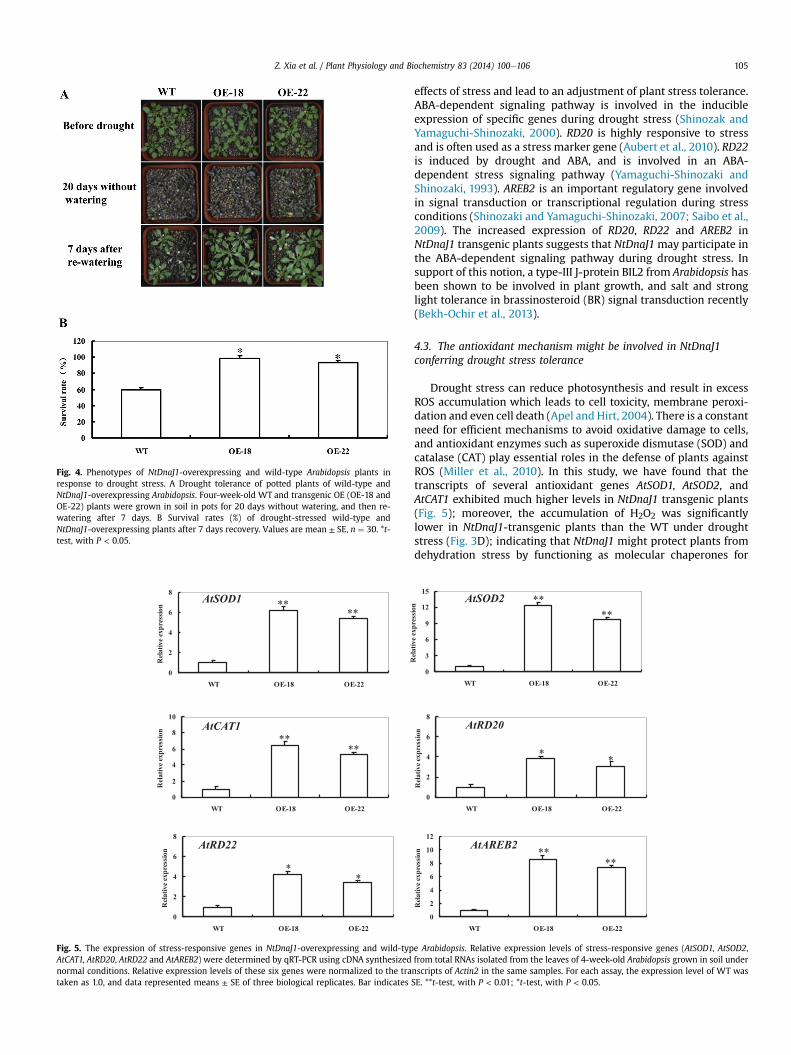
Fig. 4. Phenotypes of NtDnaJ1-overexpressing and wild-type Arabidopsis plants inresponse to drought stress. A Drought tolerance of potted plants of wild-type andNtDnaJ1-overexpressing Arabidopsis. Four-week-old WT and transgenic OE (OE-18 andOE-22) plants were grown in soil in pots for 20 days without watering, and then re-watering after 7 days. B Survival rates (%) of drought-stressed wild-type andNtDnaJ1-overexpressing plants after 7 days recovery. Values are mean ± SE, n ¼ 30. *t-test, with P < 0.05.
Fig. 5. The expression of stress-responsive genes in NtDnaJ1-overexpressing and wild-typAtCAT1, AtRD20, AtRD22 and AtAREB2) were determined by qRT-PCR using cDNA synthesizednormal conditions. Relative expression levels of these six genes were normalized to the trataken as 1.0, and data represented means ± SE of three biological replicates. Bar indicates
Z. Xia et al. / Plant Physiology and Biochemistry 83 (2014) 100e106 105
effects of stress and lead to an adjustment of plant stress tolerance.ABA-dependent signaling pathway is involved in the inducibleexpression of specific genes during drought stress (Shinozak andYamaguchi-Shinozaki, 2000). RD20 is highly responsive to stressand is often used as a stress marker gene (Aubert et al., 2010). RD22is induced by drought and ABA, and is involved in an ABA-dependent stress signaling pathway (Yamaguchi-Shinozaki andShinozaki, 1993). AREB2 is an important regulatory gene involvedin signal transduction or transcriptional regulation during stressconditions (Shinozaki and Yamaguchi-Shinozaki, 2007; Saibo et al.,2009). The increased expression of RD20, RD22 and AREB2 inNtDnaJ1 transgenic plants suggests that NtDnaJ1may participate inthe ABA-dependent signaling pathway during drought stress. Insupport of this notion, a type-III J-protein BIL2 from Arabidopsis hasbeen shown to be involved in plant growth, and salt and stronglight tolerance in brassinosteroid (BR) signal transduction recently(Bekh-Ochir et al., 2013).
4.3. The antioxidant mechanism might be involved in NtDnaJ1conferring drought stress tolerance
Drought stress can reduce photosynthesis and result in excessROS accumulation which leads to cell toxicity, membrane peroxi-dation and even cell death (Apel and Hirt, 2004). There is a constantneed for efficient mechanisms to avoid oxidative damage to cells,and antioxidant enzymes such as superoxide dismutase (SOD) andcatalase (CAT) play essential roles in the defense of plants againstROS (Miller et al., 2010). In this study, we have found that thetranscripts of several antioxidant genes AtSOD1, AtSOD2, andAtCAT1 exhibited much higher levels in NtDnaJ1 transgenic plants(Fig. 5); moreover, the accumulation of H2O2 was significantlylower in NtDnaJ1-transgenic plants than the WT under droughtstress (Fig. 3D); indicating that NtDnaJ1 might protect plants fromdehydration stress by functioning as molecular chaperones for
e Arabidopsis. Relative expression levels of stress-responsive genes (AtSOD1, AtSOD2,from total RNAs isolated from the leaves of 4-week-old Arabidopsis grown in soil undernscripts of Actin2 in the same samples. For each assay, the expression level of WT wasSE. **t-test, with P < 0.01; *t-test, with P < 0.05.

Z. Xia et al. / Plant Physiology and Biochemistry 83 (2014) 100e106106
ROS-scavenging proteins to maintain higher levels of antioxidantactivities. Supporting these interesting findings, mutation of theArabidopsis type-II J-protein AtDjiB1 decreased thermotoleranceunder heat stress by down-regulating activities of antioxidant en-zymes and accumulating ROS levels (Zhou et al., 2012); implyingthat the antioxidant systemmay be involved in J-protein-mediatedabiotic stress tolerance in plants. In the future, further study will beinteresting to clarify themechanism of activation of the antioxidantsystem in NtDnaJ1-transgenic plants under drought stress.
Acknowledgments
This work was financially supported by the science and tech-nology R&D project from Henan Tobacco Company (grant no.HYKJ201010).
Appendix A. Supplementary data
Supplementary data related to this article can be found at http://dx.doi.org/10.1016/j.plaphy.2014.07.023.
Authors' contributions
ZX designed the study andwrote themanuscript; ZX, XZ, J-Li, XSand J-Liu performed the experiments and analyzed the data; ZXcontributed reagents/materials/analysis tools.
References
Apel, K., Hirt, H., 2004. Reactive oxygen species: metabolism, oxidative stress, andsignal transduction. Annu Rev. Plant Biol. 55, 373e399.
Aubert, Y., Vile, D., Pervent, M., Aldon, D., Ranty, B., Simonneau, T., Vavasseur, A.,Jean- Philippe, G., 2010. RD20, A stress-inducible caleosin, participates in sto-matal control, transpiration and drought tolerance in Arabidopsis thaliana. PlantCell. Physiol. 51, 1975e1987.
Bekh-Ochir, D., Shimada, S., Yamagami, A., Kanda, S., Ogawa, K., Nakazawa, M.,Matsui, M., Sakuta, M., Osada, H., Asami, T., Nakano, T., 2013. A novel mitochon-drial DnaJ/Hsp40 family protein BIL2 promotes plant growth and resistanceagainst environmental stress in brassinosteroid signaling. Planta 237,1509e1525.
Caplan, A.J., Cyr, D.M., Douglas, M.G., 1993. Eukaryotic homologs of Escherichia coliDnaj-a diverse protein family that functions with Hsp70 stress proteins. Mol.Biol. Cell. 4, 555e563.
Cheetham, M.E., Caplan, A.J., 1998. Structure, function and evolution of DnaJ: conser-vation and adaptation of chaperone function. Cell. Stress Chaperones 3, 28e36.
Clough, S.J., Bent, A.F., 1998. Floral dip: a simplified method for Agrobaterium-mediated transformation of Arabidopsis thaliana. Plant J. 16, 735e743.
Craig, E.A., Huang, P., Aron, R., Andrew, A., 2006. The diverse roles of J-proteins, theobligate Hsp70 co-chaperone. Rev. Physiol. Biochem Pharmacol. 156, 1e21.
de Crouy-Chanel, A., Kohiyama, M., Richarme, G., 1995. A novel function ofEscherichia coli chaperone DnaJ protein-disulfide isomerase. J. Biol. Chem. 270,22669e22672.
Du, Y., Zhao, J., Chen, T., Liu, Q., Zhang, H.,Wang, Y., Hong, Y., Xiao, F., Zhang, L., Shen, Q.,Liu, Y., 2013. Type I J-domain NbMIP1 proteins are required for both tobaccomosaic virus infection and plant innate immunity. PLoS Pathog. 9, e1003659.
Goffin, L., Georgopoulos, C., 1998. Genetic and biochemical characterization ofmutations affecting the carboxy-terminal domain of the Escherichia coli mo-lecular chaperone DnaJ. Mol. Microbiol. 30, 329e340.
Hennessy, F., Boshoff, A., Blatch, G.L., 2005. Rational mutagenesis of a 40 kDa heatshock protein from Agrobacterium tumefaciens identifies amino acid residuescritical to its in vivo function. Int. J. Biochem Cell. Biol. 37, 177e191.
Kneissl, J., Wachtler, V., Chua, N.H., Bolle, C., 2009. OWL1: an Arabidopsis J-domainprotein involved in perception of very low light fluences. Plant Cell. 21,3212e3225.
Liu, J.Z., Whitham, S.A., 2013. Overexpression of a soybean nuclear localized type IIIDnaJ domain-containing HSP40 reveals its roles in cell death and diseaseresistance. Plant J. 74, 110e121.
Livak, K.J., Schmittgen, T.D., 2001. Analysis of relative gene expression data usingreal- time quantitative PCR and the 2(-Delta Delta C (T)) Method. Methods 25,402e408.
Mayer, M.P., Bukau, B., 2005. Hsp70 chaperones: cellular functions and molecularmechanism. Cell. Mol. Life Sci. 62, 670e684.
Miernyk, J.A., 2001. The J-domain proteins of Arabidopsis thaliana: an unexpectedlylarge and diverse family of chaperones. Cell. Stress Chaperones 6, 209e218.
Miller, G., Suzuki, N., Ciftci-Yilmaz, S., Mittler, R., 2010. Reactive oxygen specieshomeostasis and signaling during drought and salinity stresses. Plant Cell.Environ. 33, 453e457.
Qiu, X.B., Shao, Y.M., Miao, S., Wang, L., 2006. The diversity of the DnaJ/Hsp40family, the crucial partners for Hsp70 chaperones. Cell. Mol. Life Sci. 63,2560e2570.
Rajan, V.B., D'Silva, P., 2009. Arabidopsis thaliana J-class heat shock proteins: cellularstress sensors. Funct. Integr. Genomics 9, 433e446.
Saibo, N.J.M., Lourenço, T., Oliveira, M.M., 2009. Transcription factors and regulationof photosynthetic and related metabolism under environmental stresses. Ann.Bot. 103, 609e623.
Shen, L., Kang, Y.G., Liu, L., Yu, H., 2011. The J-domain protein J3 mediates theintegration of flowering signals in Arabidopsis. Plant Cell. 23, 499e514.
Shinozak, I.K., Yamaguchi-Shinozaki, K., 2000. Molecular responses to dehydrationand low temperature: differences and cross-talk between two stress signalpathways. Curr. Opin. Plant Biol. 3, 217e223.
Shinozaki, K., Yamaguchi-Shinozaki, K., 2007. Gene networks involved in droughtstress response and tolerance. J. Exp. Bot. 58, 221e227.
Silver, P.A., Way, J.C., 1993. Eukaryotic Dnaj homologs and the specificity of Hsp70activity. Cell 74, 5e6.
Szabo, A., Korszun, R., Hartl, F.U., Flanagan, J., 1996. A zinc finger-like domain of themolecular chaperone DnaJ is involved in binding to denatured protein sub-strates. EMBO J. 15, 408e417.
Szyperski, T., Pellecchia, M., Wall, D., Georgopoulos, C., Wuthrich, K., 1994. NMRstructure determination of the Escherichia coli DnaJ molecular chaperone:secondary structure and backbone fold of the N-terminal region (residues2e108) containing the highly conserved J domain. Proc. Natl. Acad. Sci. U. S. A.91, 11343e11347.
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., Kumar, S., 2011. MEGA5:molecular evolutionary genetics analysis using maximum likelihood, evolu-tionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28,2731e2739.
Walsh, P., Bursac, D., Law, Y.C., Cyr, D., Lithgow, T., 2004. The J-protein family:modulating protein assembly, disassembly and translocation. EMBO Rep. 5,567e571.
Wang, W., Vinocur, B., Shoseyov, O., Altman, A., 2004. Role of plant heat-shockproteins and molecular chaperones in the abiotic stress response. TrendsPlant Sci. 9, 244e252.
Wang, G., Cai, G., Kong, F., Deng, Y., Ma, N., Meng, Q., 2014. Overexpression of tomatochloroplast-targeted DnaJ protein enhances tolerance to drought stress andresistance to Pseudomonas solanacearum in transgenic tobacco. Plant Physiol.Biochem 82, 95e104.
Xia, Z., Su, X., Wu, J., Wu, K., Zhang, H., 2012. Molecular cloning and functionalcharacterization of a putative sulfite oxidase (SO) ortholog from Nicotianabenthamiana. Mol. Biol. Rep. 39, 2429e2437.
Xia, Z., Sun, K., Wang, M., Wu, K., Zhang, H., 2012. Overexpression of a maize sulfiteoxidase gene in tobacco enhances tolerance to sulfite stress via sulfite oxidationand CAT-mediated H2O2 scavenging. PLoS One 7, e37383.
Xia, Z., Wei, Y., Sun, K., Wu, J., Wang, Y., Wu, K., 2013. The maize AAA-type proteinSKD1 confers enhanced salt and drought stress tolerance in transgenic tobaccoby interacting with Lyst-interacting protein 5. PLoS One 8, e69787.
Yamaguchi-Shinozaki, K., Shinozaki, K., 1993. The plant hormone abscisic acidmediates the drought-induced expression but not the seed-specific expressionof rd22, a gene responsive to dehydration stress in Arabidopsis thaliana. Mol.Gen. Genet. 238, 17e25.
Yang, K., Xia, C., Liu, X., Dou, X., Wang, W., Chen, L., Zhang, X., Xie, L., He, L., Ma, X.,Ye, D., 2009. A mutation in Thermosensitive Male Sterile 1, encoding a heatshock protein with DnaJ and PDI domains, leads to thermosensitive gameto-phytic male sterility in Arabidopsis. Plant J. 57, 870e882.
Yang, Y., Qin, Y., Xie, C., Zhao, F., Zhao, J., Liu, D., Chen, S., Fuglsang, A.T.,Palmgren, M.G., Schumaker, K.S., Deng, X.W., Guo, Y., 2010. The Arabidopsischaperone J3 regulates the plasma membrane Hþ-ATPase through interactionwith the PKS5 kinase. Plant Cell. 22, 1313e1332.
Zhou, W., Zhou, T., Li, M., Zhao, C., Jia, N., Wang, X., Sun, Y., Li, G., Xu, M., Zhou, R.,Li, B., 2012. The Arabidopsis J-protein AtDjB1 facilitates thermotolerance byprotecting cells against heat-induced oxidative damage. New. Phytol. 194,364e378.