organochlorinated pesticide degrading microorganisms isolated from contaminated soil
TRANSCRIPT

Research
Pap
er
RESEARCH PAPER New Biotechnology � Volume 32, Number 1 � January 2015
Organochlorinated pesticide degradingmicroorganisms isolated fromcontaminated soilPetra Lovecka1, Iva Pacovska1, Petr Stursa1, Blanka Vrchotova1,Lucie Kochankova2 and Katerina Demnerova1
1Department of Biochemistry and Microbiology, Institute of Chemical Technology, Technicka 3, Prague 166 28, Czech Republic2Department of Environmental Chemistry, Institute of Chemical Technology, Technicka 3, Prague 166 28, Czech Republic
Degradation of selected organochlorinated pesticides (g-hexachlorocyclohexane – g-HCH,
dichlorodiphenyltrichloroethane – DDT, hexachlorobenzene – HCB) by soil microorganisms was
studied. Bacterial strains isolated from contaminated soil from Klatovy-Luby, Hajek and Neratovice,
Czech Republic, capable of growth on the selected pesticides were isolated and characterised. These
isolates were subjected to characterisation and identification by MS MALDI-TOF of whole cells and
sequence analysis of 16S rRNA genes. The isolates were screened by gas chromatography for their ability
to degrade the selected pesticides. Some isolates were able to degrade pesticides, and the formation of
degradation products (g-pentachlorocyclohexane (g-PCCH), dichlorodiphenyldichloroethylene (DDE)
and dichlorodiphenyldichloroethane (DDD)) observed in liquid culture confirmed their degradation
capability. The isolates and DNA samples isolated from the contaminated soil were also screened for the
bphA1 gene (encoding biphenyl-2,3-dioxygenase, the first enzyme in the PCB degradation pathway) and
its occurrence was demonstrated. The isolates were also screened for the presence of linA, encoding
dehydrochlorinase, the first enzyme of the HCH degradation pathway. The linA gene could not be found
in any of the tested isolates, possibly due to the high specificity of the primers used. The isolates with the
most effective degradation abilities could be used for further in situ bioremediation experiments with
contaminated soil.
IntroductionIncreased agricultural production to meet the ever increasing
demand for food, has been achieved thanks to the widespread
use of herbicides and insecticides. Thanks to these substances,
several diseases caused by insect vectors have been eradicated [1].
Throughout the 20th century organic pesticides such as organo-
chlorinated substances were primarily used, even though they can
survive in the environment for decades [2]. Organochlorinated
pesticides (OCPs) are generally white crystalline substances which
are poorly soluble in water, more soluble in organic solvents
and highly soluble in fats. Due to their chemical resistance and
Corresponding author:. Lovecka, P. ([email protected])
www.elsevier.com/locate/nbt
26
solubility in fats they accumulate in adipose tissue and subsequent
biomagnification to higher trophic levels occurs. The basic repre-
sentatives of OCPs include DDT, lindane, technical HCH (hexa-
chlorocyclohexane), dieldrin, aldrin, heptachlor, chlordane,
hexachlorobenzene (HCB) and others. These compounds were
used primarily as insecticides, but nowadays are banned in most
countries due to proven negative effects on human health and the
environment [3]. They occur more and more frequently as con-
taminants of soil, air and water sources, posing a risk both to
humans and to the environment. Increased attention has been
paid to persistent metabolites resulting from the partial degrada-
tion of OCPs, which is carried out mainly by microorganisms.
Factors affecting the rate of degradation include the compound
http://dx.doi.org/10.1016/j.nbt.2014.07.003
1871-6784/� 2014 Published by Elsevier B.V.
New Biotechnology �Volume 32, Number 1 � January 2015 RESEARCH PAPER
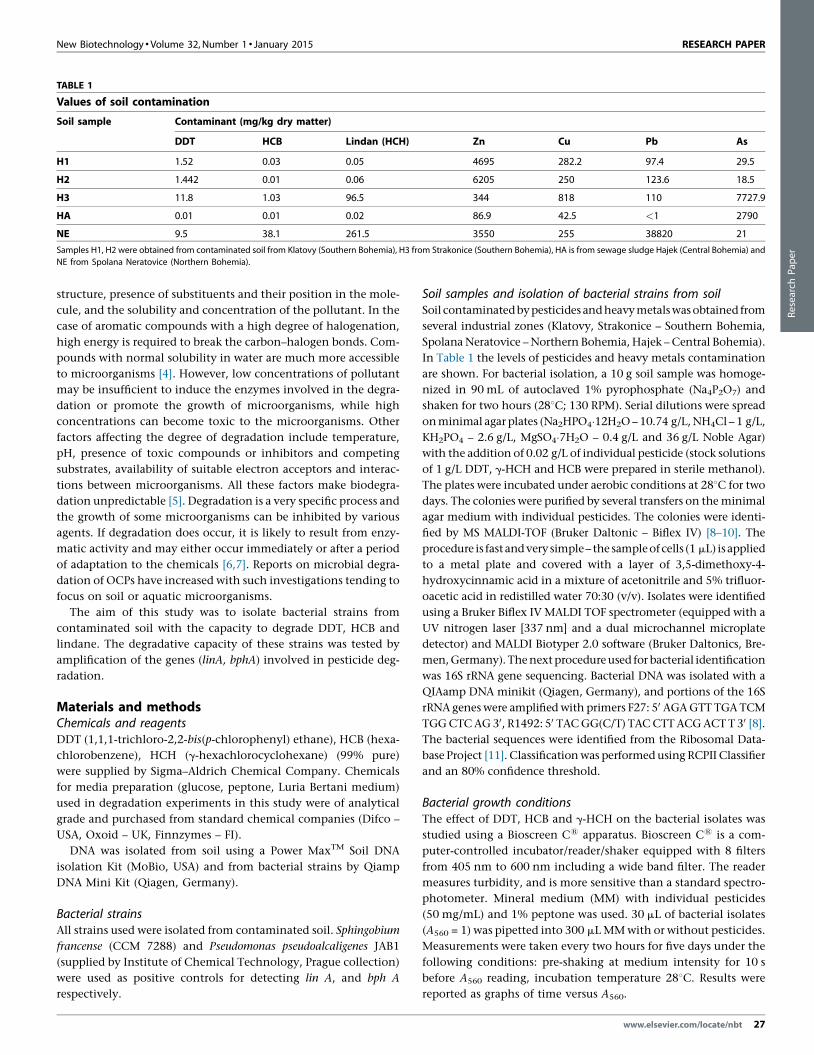
TABLE 1
Values of soil contamination
Soil sample Contaminant (mg/kg dry matter)
DDT HCB Lindan (HCH) Zn Cu Pb As
H1 1.52 0.03 0.05 4695 282.2 97.4 29.5
H2 1.442 0.01 0.06 6205 250 123.6 18.5
H3 11.8 1.03 96.5 344 818 110 7727.9
HA 0.01 0.01 0.02 86.9 42.5 <1 2790
NE 9.5 38.1 261.5 3550 255 38820 21
Samples H1, H2 were obtained from contaminated soil from Klatovy (Southern Bohemia), H3 from Strakonice (Southern Bohemia), HA is from sewage sludge Hajek (Central Bohemia) and
NE from Spolana Neratovice (Northern Bohemia).
ResearchPap
er
structure, presence of substituents and their position in the mole-
cule, and the solubility and concentration of the pollutant. In the
case of aromatic compounds with a high degree of halogenation,
high energy is required to break the carbon–halogen bonds. Com-
pounds with normal solubility in water are much more accessible
to microorganisms [4]. However, low concentrations of pollutant
may be insufficient to induce the enzymes involved in the degra-
dation or promote the growth of microorganisms, while high
concentrations can become toxic to the microorganisms. Other
factors affecting the degree of degradation include temperature,
pH, presence of toxic compounds or inhibitors and competing
substrates, availability of suitable electron acceptors and interac-
tions between microorganisms. All these factors make biodegra-
dation unpredictable [5]. Degradation is a very specific process and
the growth of some microorganisms can be inhibited by various
agents. If degradation does occur, it is likely to result from enzy-
matic activity and may either occur immediately or after a period
of adaptation to the chemicals [6,7]. Reports on microbial degra-
dation of OCPs have increased with such investigations tending to
focus on soil or aquatic microorganisms.
The aim of this study was to isolate bacterial strains from
contaminated soil with the capacity to degrade DDT, HCB and
lindane. The degradative capacity of these strains was tested by
amplification of the genes (linA, bphA) involved in pesticide deg-
radation.
Materials and methodsChemicals and reagentsDDT (1,1,1-trichloro-2,2-bis(p-chlorophenyl) ethane), HCB (hexa-
chlorobenzene), HCH (g-hexachlorocyclohexane) (99% pure)
were supplied by Sigma–Aldrich Chemical Company. Chemicals
for media preparation (glucose, peptone, Luria Bertani medium)
used in degradation experiments in this study were of analytical
grade and purchased from standard chemical companies (Difco –
USA, Oxoid – UK, Finnzymes – FI).
DNA was isolated from soil using a Power MaxTM Soil DNA
isolation Kit (MoBio, USA) and from bacterial strains by Qiamp
DNA Mini Kit (Qiagen, Germany).
Bacterial strainsAll strains used were isolated from contaminated soil. Sphingobium
francense (CCM 7288) and Pseudomonas pseudoalcaligenes JAB1
(supplied by Institute of Chemical Technology, Prague collection)
were used as positive controls for detecting lin A, and bph A
respectively.
Soil samples and isolation of bacterial strains from soilSoil contaminated by pesticides and heavy metals was obtained from
several industrial zones (Klatovy, Strakonice – Southern Bohemia,
Spolana Neratovice – Northern Bohemia, Hajek – Central Bohemia).
In Table 1 the levels of pesticides and heavy metals contamination
are shown. For bacterial isolation, a 10 g soil sample was homoge-
nized in 90 mL of autoclaved 1% pyrophosphate (Na4P2O7) and
shaken for two hours (288C; 130 RPM). Serial dilutions were spread
on minimal agar plates (Na2HPO4�12H2O – 10.74 g/L, NH4Cl – 1 g/L,
KH2PO4 – 2.6 g/L, MgSO4�7H2O – 0.4 g/L and 36 g/L Noble Agar)
with the addition of 0.02 g/L of individual pesticide (stock solutions
of 1 g/L DDT, g-HCH and HCB were prepared in sterile methanol).
The plates were incubated under aerobic conditions at 288C for two
days. The colonies were purified by several transfers on the minimal
agar medium with individual pesticides. The colonies were identi-
fied by MS MALDI-TOF (Bruker Daltonic – Biflex IV) [8–10]. The
procedure is fast and very simple – the sample of cells (1 mL) is applied
to a metal plate and covered with a layer of 3,5-dimethoxy-4-
hydroxycinnamic acid in a mixture of acetonitrile and 5% trifluor-
oacetic acid in redistilled water 70:30 (v/v). Isolates were identified
using a Bruker Biflex IV MALDI TOF spectrometer (equipped with a
UV nitrogen laser [337 nm] and a dual microchannel microplate
detector) and MALDI Biotyper 2.0 software (Bruker Daltonics, Bre-
men, Germany). The next procedure used for bacterial identification
was 16S rRNA gene sequencing. Bacterial DNA was isolated with a
QIAamp DNA minikit (Qiagen, Germany), and portions of the 16S
rRNA genes were amplified with primers F27: 50 AGA GTT TGA TCM
TGG CTC AG 30, R1492: 50 TAC GG(C/T) TAC CTT ACG ACT T 30 [8].
The bacterial sequences were identified from the Ribosomal Data-
base Project [11]. Classification was performed using RCPII Classifier
and an 80% confidence threshold.
Bacterial growth conditionsThe effect of DDT, HCB and g-HCH on the bacterial isolates was
studied using a Bioscreen C1 apparatus. Bioscreen C1 is a com-
puter-controlled incubator/reader/shaker equipped with 8 filters
from 405 nm to 600 nm including a wide band filter. The reader
measures turbidity, and is more sensitive than a standard spectro-
photometer. Mineral medium (MM) with individual pesticides
(50 mg/mL) and 1% peptone was used. 30 mL of bacterial isolates
(A560 = 1) was pipetted into 300 mL MM with or without pesticides.
Measurements were taken every two hours for five days under the
following conditions: pre-shaking at medium intensity for 10 s
before A560 reading, incubation temperature 288C. Results were
reported as graphs of time versus A560.
www.elsevier.com/locate/nbt 27
RESEARCH PAPER New Biotechnology � Volume 32, Number 1 � January 2015
TABLE 2
Identification of chosen isolated bacteria by MS MALDI-TOF and analysis of 16S rRNA gene
Isolates Identification by MS MALDI-TOF Score valuea Identification by sequencing 16S rRNA gene RDP score
NE6 Aeromonas sp. 2.26 Pseudomonas sp. 0.92
H23 Rhodococcus sp. 2.29 Rhodococcus erythropolis 0.86
H16 Bacillus sp. 1.89 Bacillus pumilus 0.98
H1D7 Stenotrophomonas sp. 1.82 Stenotrophomonas sp. 0.99
NE15 Unidentified – Lysinibacillus fusiformis 0.99
NE22 Bacillus sp – Bacillus cereus 0.85
HA1 Bacillus sp 2.14 Bacillus cereus 0.91
a Range: 0.00–1.699: identification not reliable; 1.7–1.999: probable genus identification; 2.0–2.299: secure genus identification, probable species identification; 2.3–3.0: highly probable
species identification.
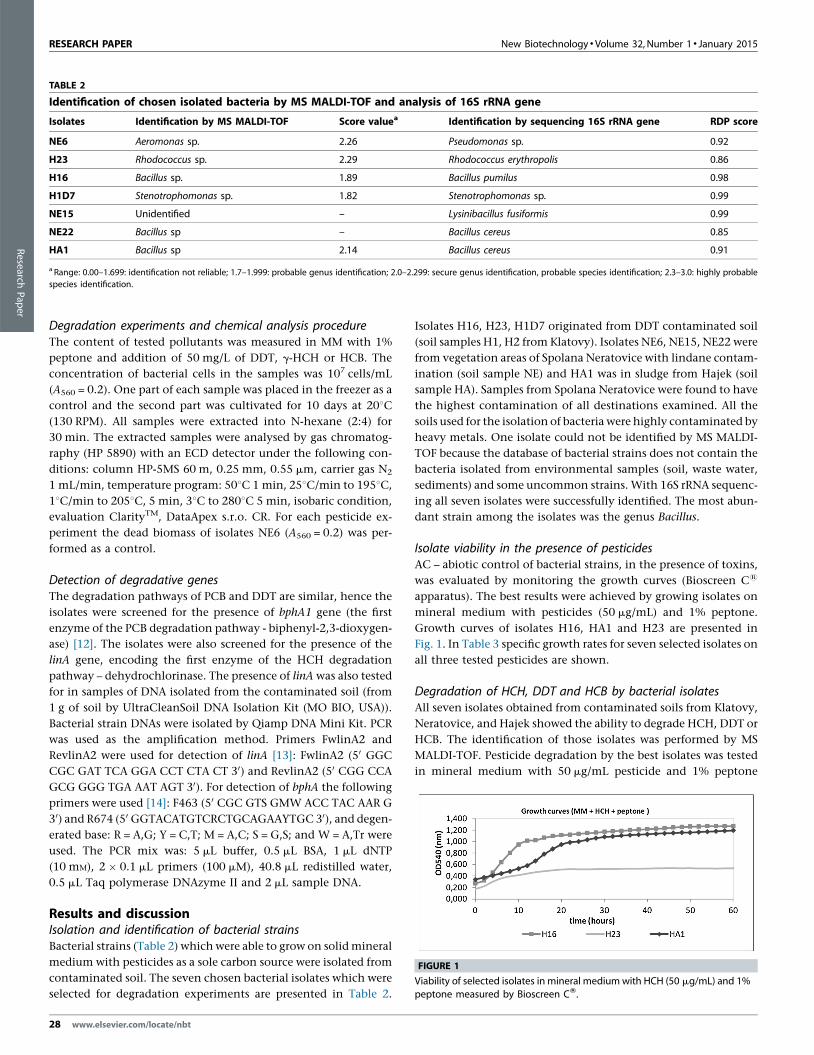
FIGURE 1
Viability of selected isolates in mineral medium with HCH (50 mg/mL) and 1%peptone measured by Bioscreen CW.
Research
Pap
er
Degradation experiments and chemical analysis procedureThe content of tested pollutants was measured in MM with 1%
peptone and addition of 50 mg/L of DDT, g-HCH or HCB. The
concentration of bacterial cells in the samples was 107 cells/mL
(A560 = 0.2). One part of each sample was placed in the freezer as a
control and the second part was cultivated for 10 days at 208C(130 RPM). All samples were extracted into N-hexane (2:4) for
30 min. The extracted samples were analysed by gas chromatog-
raphy (HP 5890) with an ECD detector under the following con-
ditions: column HP-5MS 60 m, 0.25 mm, 0.55 mm, carrier gas N2
1 mL/min, temperature program: 508C 1 min, 258C/min to 1958C,
18C/min to 2058C, 5 min, 38C to 2808C 5 min, isobaric condition,
evaluation ClarityTM, DataApex s.r.o. CR. For each pesticide ex-
periment the dead biomass of isolates NE6 (A560 = 0.2) was per-
formed as a control.
Detection of degradative genesThe degradation pathways of PCB and DDT are similar, hence the
isolates were screened for the presence of bphA1 gene (the first
enzyme of the PCB degradation pathway - biphenyl-2,3-dioxygen-
ase) [12]. The isolates were also screened for the presence of the
linA gene, encoding the first enzyme of the HCH degradation
pathway – dehydrochlorinase. The presence of linA was also tested
for in samples of DNA isolated from the contaminated soil (from
1 g of soil by UltraCleanSoil DNA Isolation Kit (MO BIO, USA)).
Bacterial strain DNAs were isolated by Qiamp DNA Mini Kit. PCR
was used as the amplification method. Primers FwlinA2 and
RevlinA2 were used for detection of linA [13]: FwlinA2 (50 GGC
CGC GAT TCA GGA CCT CTA CT 30) and RevlinA2 (50 CGG CCA
GCG GGG TGA AAT AGT 30). For detection of bphA the following
primers were used [14]: F463 (50 CGC GTS GMW ACC TAC AAR G
30) and R674 (50 GGTACATGTCRCTGCAGAAYTGC 30), and degen-
erated base: R = A,G; Y = C,T; M = A,C; S = G,S; and W = A,Tr were
used. The PCR mix was: 5 mL buffer, 0.5 mL BSA, 1 mL dNTP
(10 mM), 2 � 0.1 mL primers (100 mM), 40.8 mL redistilled water,
0.5 mL Taq polymerase DNAzyme II and 2 mL sample DNA.
Results and discussionIsolation and identification of bacterial strainsBacterial strains (Table 2) which were able to grow on solid mineral
medium with pesticides as a sole carbon source were isolated from
contaminated soil. The seven chosen bacterial isolates which were
selected for degradation experiments are presented in Table 2.
28 www.elsevier.com/locate/nbt
Isolates H16, H23, H1D7 originated from DDT contaminated soil
(soil samples H1, H2 from Klatovy). Isolates NE6, NE15, NE22 were
from vegetation areas of Spolana Neratovice with lindane contam-
ination (soil sample NE) and HA1 was in sludge from Hajek (soil
sample HA). Samples from Spolana Neratovice were found to have
the highest contamination of all destinations examined. All the
soils used for the isolation of bacteria were highly contaminated by
heavy metals. One isolate could not be identified by MS MALDI-
TOF because the database of bacterial strains does not contain the
bacteria isolated from environmental samples (soil, waste water,
sediments) and some uncommon strains. With 16S rRNA sequenc-
ing all seven isolates were successfully identified. The most abun-
dant strain among the isolates was the genus Bacillus.
Isolate viability in the presence of pesticidesAC – abiotic control of bacterial strains, in the presence of toxins,
was evaluated by monitoring the growth curves (Bioscreen C1
apparatus). The best results were achieved by growing isolates on
mineral medium with pesticides (50 mg/mL) and 1% peptone.
Growth curves of isolates H16, HA1 and H23 are presented in
Fig. 1. In Table 3 specific growth rates for seven selected isolates on
all three tested pesticides are shown.
Degradation of HCH, DDT and HCB by bacterial isolatesAll seven isolates obtained from contaminated soils from Klatovy,
Neratovice, and Hajek showed the ability to degrade HCH, DDT or
HCB. The identification of those isolates was performed by MS
MALDI-TOF. Pesticide degradation by the best isolates was tested
in mineral medium with 50 mg/mL pesticide and 1% peptone
New Biotechnology �Volume 32, Number 1 � January 2015 RESEARCH PAPER
TABLE 3
Specific growth rate (hour�1) of isolates for cultivation on mineral medium with peptone and individual pesticides (concentration50 mg/mL)
Isolates Identification Specific growth rate
with DDT (hour�1)
Specific growth rate
with HCB (hour�1)
Specific growth rate
with HCH (hour�1)
NE6 Pseudomonas sp. 0.5 0.4 0.2
H23 Rhodococcus erythropolis 0.5 0.5 0.5
H16 Bacillus pumilus 0.1 0.2 0.2
H1D7 Stenotrophomonas sp. 0.3 0.2 0.2
NE15 Lysinibacillus fusiformis 0.5 0.4 0.2
NE22 Bacillus cereus 0.4 0.4 0.5
HA1 Bacillus cereus 0.4 0.4 0.4
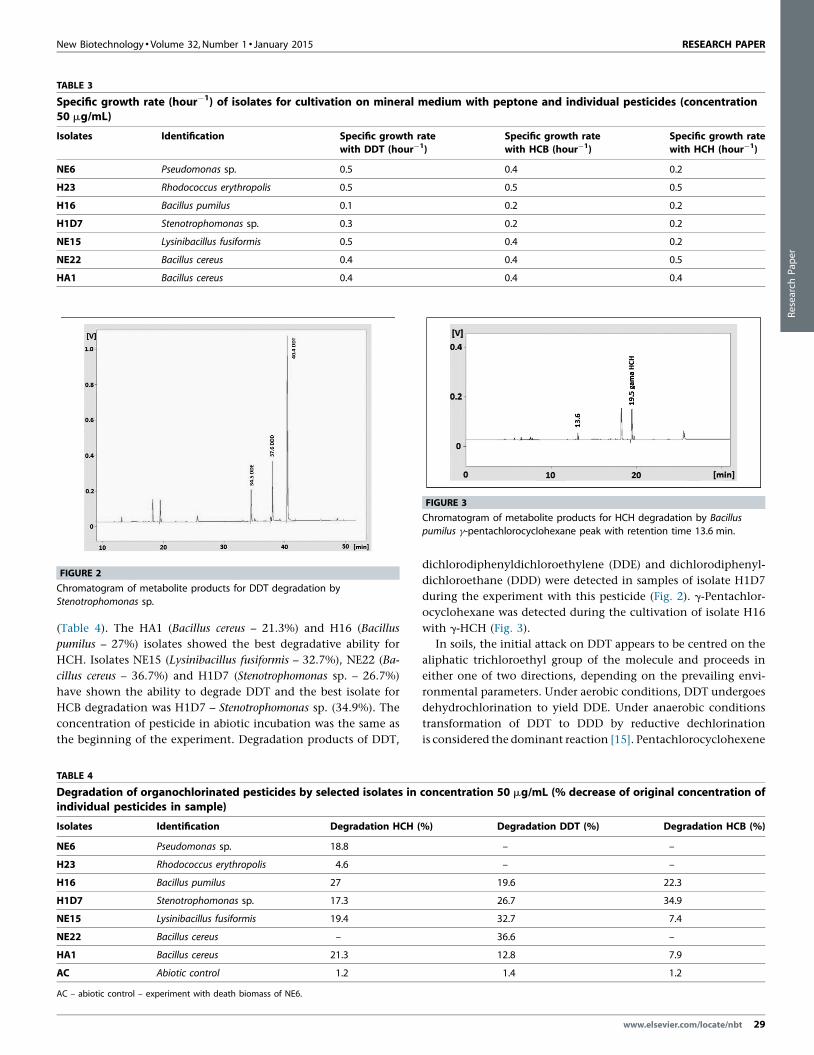
FIGURE 2
Chromatogram of metabolite products for DDT degradation by
Stenotrophomonas sp.
FIGURE 3
Chromatogram of metabolite products for HCH degradation by Bacillus
pumilus g-pentachlorocyclohexane peak with retention time 13.6 min.
ResearchPap
er
(Table 4). The HA1 (Bacillus cereus – 21.3%) and H16 (Bacillus
pumilus – 27%) isolates showed the best degradative ability for
HCH. Isolates NE15 (Lysinibacillus fusiformis – 32.7%), NE22 (Ba-
cillus cereus – 36.7%) and H1D7 (Stenotrophomonas sp. – 26.7%)
have shown the ability to degrade DDT and the best isolate for
HCB degradation was H1D7 – Stenotrophomonas sp. (34.9%). The
concentration of pesticide in abiotic incubation was the same as
the beginning of the experiment. Degradation products of DDT,
TABLE 4
Degradation of organochlorinated pesticides by selected isolates in
individual pesticides in sample)
Isolates Identification Degradation HCH (
NE6 Pseudomonas sp. 18.8
H23 Rhodococcus erythropolis 4.6
H16 Bacillus pumilus 27
H1D7 Stenotrophomonas sp. 17.3
NE15 Lysinibacillus fusiformis 19.4
NE22 Bacillus cereus –
HA1 Bacillus cereus 21.3
AC Abiotic control 1.2
AC – abiotic control – experiment with death biomass of NE6.
dichlorodiphenyldichloroethylene (DDE) and dichlorodiphenyl-
dichloroethane (DDD) were detected in samples of isolate H1D7
during the experiment with this pesticide (Fig. 2). g-Pentachlor-
ocyclohexane was detected during the cultivation of isolate H16
with g-HCH (Fig. 3).
In soils, the initial attack on DDT appears to be centred on the
aliphatic trichloroethyl group of the molecule and proceeds in
either one of two directions, depending on the prevailing envi-
ronmental parameters. Under aerobic conditions, DDT undergoes
dehydrochlorination to yield DDE. Under anaerobic conditions
transformation of DDT to DDD by reductive dechlorination
is considered the dominant reaction [15]. Pentachlorocyclohexene
concentration 50 mg/mL (% decrease of original concentration of
%) Degradation DDT (%) Degradation HCB (%)
– –
– –
19.6 22.3
26.7 34.9
32.7 7.4
36.6 –
12.8 7.9
1.4 1.2
www.elsevier.com/locate/nbt 29
RESEARCH PAPER New Biotechnology � Volume 32, Number 1 � January 2015
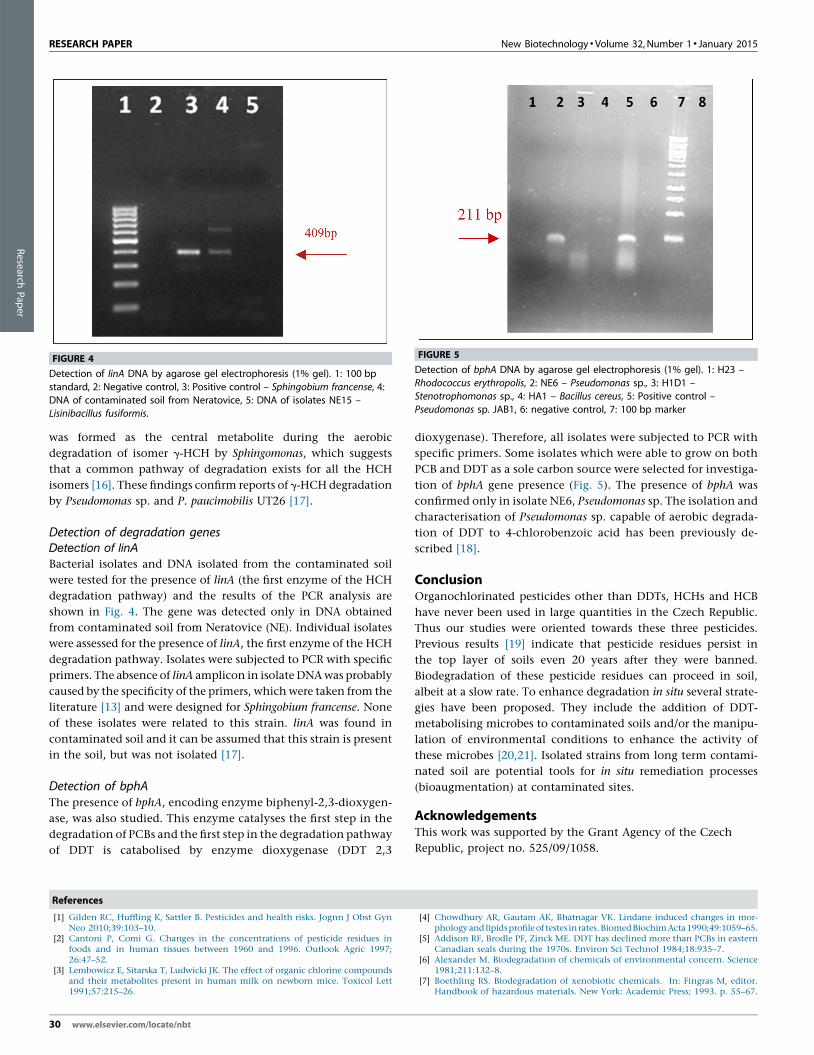
FIGURE 4
Detection of linA DNA by agarose gel electrophoresis (1% gel). 1: 100 bp
standard, 2: Negative control, 3: Positive control – Sphingobium francense, 4:
DNA of contaminated soil from Neratovice, 5: DNA of isolates NE15 –Lisinibacillus fusiformis.
FIGURE 5
Detection of bphA DNA by agarose gel electrophoresis (1% gel). 1: H23 –
Rhodococcus erythropolis, 2: NE6 – Pseudomonas sp., 3: H1D1 –
Stenotrophomonas sp., 4: HA1 – Bacillus cereus, 5: Positive control –Pseudomonas sp. JAB1, 6: negative control, 7: 100 bp marker
Research
Pap
er
was formed as the central metabolite during the aerobic
degradation of isomer g-HCH by Sphingomonas, which suggests
that a common pathway of degradation exists for all the HCH
isomers [16]. These findings confirm reports of g-HCH degradation
by Pseudomonas sp. and P. paucimobilis UT26 [17].
Detection of degradation genesDetection of linA
Bacterial isolates and DNA isolated from the contaminated soil
were tested for the presence of linA (the first enzyme of the HCH
degradation pathway) and the results of the PCR analysis are
shown in Fig. 4. The gene was detected only in DNA obtained
from contaminated soil from Neratovice (NE). Individual isolates
were assessed for the presence of linA, the first enzyme of the HCH
degradation pathway. Isolates were subjected to PCR with specific
primers. The absence of linA amplicon in isolate DNA was probably
caused by the specificity of the primers, which were taken from the
literature [13] and were designed for Sphingobium francense. None
of these isolates were related to this strain. linA was found in
contaminated soil and it can be assumed that this strain is present
in the soil, but was not isolated [17].
Detection of bphAThe presence of bphA, encoding enzyme biphenyl-2,3-dioxygen-
ase, was also studied. This enzyme catalyses the first step in the
degradation of PCBs and the first step in the degradation pathway
of DDT is catabolised by enzyme dioxygenase (DDT 2,3
30 www.elsevier.com/locate/nbt
dioxygenase). Therefore, all isolates were subjected to PCR with
specific primers. Some isolates which were able to grow on both
PCB and DDT as a sole carbon source were selected for investiga-
tion of bphA gene presence (Fig. 5). The presence of bphA was
confirmed only in isolate NE6, Pseudomonas sp. The isolation and
characterisation of Pseudomonas sp. capable of aerobic degrada-
tion of DDT to 4-chlorobenzoic acid has been previously de-
scribed [18].
ConclusionOrganochlorinated pesticides other than DDTs, HCHs and HCB
have never been used in large quantities in the Czech Republic.
Thus our studies were oriented towards these three pesticides.
Previous results [19] indicate that pesticide residues persist in
the top layer of soils even 20 years after they were banned.
Biodegradation of these pesticide residues can proceed in soil,
albeit at a slow rate. To enhance degradation in situ several strate-
gies have been proposed. They include the addition of DDT-
metabolising microbes to contaminated soils and/or the manipu-
lation of environmental conditions to enhance the activity of
these microbes [20,21]. Isolated strains from long term contami-
nated soil are potential tools for in situ remediation processes
(bioaugmentation) at contaminated sites.
AcknowledgementsThis work was supported by the Grant Agency of the Czech
Republic, project no. 525/09/1058.
References
[1] Gilden RC, Huffling K, Sattler B. Pesticides and health risks. Jognn J Obst GynNeo 2010;39:103–10.
[2] Cantoni P, Comi G. Changes in the concentrations of pesticide residues infoods and in human tissues between 1960 and 1996. Outlook Agric 1997;26:47–52.
[3] Lembowicz E, Sitarska T, Ludwicki JK. The effect of organic chlorine compoundsand their metabolites present in human milk on newborn mice. Toxicol Lett1991;57:215–26.
[4] Chowdhury AR, Gautam AK, Bhatnagar VK. Lindane induced changes in mor-phology and lipids profile of testes in rates. Biomed Biochim Acta 1990;49:1059–65.
[5] Addison RF, Brodle PF, Zinck ME. DDT has declined more than PCBs in easternCanadian seals during the 1970s. Environ Sci Technol 1984;18:935–7.
[6] Alexander M. Biodegradation of chemicals of environmental concern. Science1981;211:132–8.
[7] Boethling RS. Biodegradation of xenobiotic chemicals. In: Fingras M, editor.Handbook of hazardous materials. New York: Academic Press; 1993. p. 55–67.

New Biotechnology �Volume 32, Number 1 � January 2015 RESEARCH PAPER
arch
Pap
er
[8] Lane DJ. 16S/23S rRNA sequencing. In: Nucleic acid techniques in bacterialsystematics. Wiley; 1991: 115–75.
[9] Stursa P, Junkova P, Strejcek M, Macek T, Mackova M. MALDI-TOF MS simpleand rapid tool for identification of bacteria isolated from the environment. LCaR2010;11:412–3.
[10] Holland RD, Wilkes JG, Sutherland JB, Persons CC, Voorhees KJ, Lay Jr Jo. Rapididentification of intact whole bacteria based on spectral patterns using matrixassisted laser desorption/ionization with time-of-flight mass spectrometry. Rap-id Commun Mass Spectrom 1996;10:1227–32.
[11] Cole J, Chai B, Farris RJ, Wang Q, Kulam SA, McGarrell MD, et al. The RibosomalDatabase Project (RDP-II): sequences and tools for high-throughput rRNAanalysis. Nucleic Acids Res 2005;33:D294–6.
[12] Abramowitz DA. Aerobic and anaerobic biodegradation of PCBs: a review. CritRev Biotechnol 1990;10:241–51.
[13] Wu J, Hong Q, Sun Y, Hong Y, Yan Q, Li S. Analysis of the role of LinA and LinBin biodegradation of d-hexachlorocyclohexane. Environ Microbiol 2007;9:2331–40.
[14] Uhlık O, Strejcek M, Junkova P, Sanda M, Hroudova M, Vlcek C, et al. Matrix-assisted laser desorption ionization (MALDI)-time of flight mass spectrometry- and
MALDI Biotyper-based identification of cultured biphenyl metabolizing bacteriafrom contaminated horseradish rhizosphere soil. Appl Environ Microbiol2011;77:6858–66.
[15] Aislabie JM, Richards NK, Boul HL. Microbial degradation of DDT and itsresidues—a review. N Z J Agric Res 1997;40:269–82.
[16] Johri AK, Dua M, Tuteja D, Saxena R, Saxena DM, Lal R. Degradation of alpha,beta, gamma and delta-hexachlorocyclohexane by Sphingomonas paucimobilis.Biotechnol Lett 1998;20:885–7.
[17] Manickam N, Misra R, Mayilraj S. A novel pathway for the biodegradation of g-hexachlorocyclohexane by a Xanthomonas sp strain ICH12. J Appl Microbiol2007;102:1468–78.
[18] Kamanavalli CM, Ninnekar H. Biodegradation of DDT by a Pseudomonas species.Curr Microbiol 2004;48:10–3.
[19] Shegunova P, Klanova J, Holoubek I. Residues of organochlorinated pesticides insoils from the Czech Republic. Environ Pollut 2007;146:257–61.
[20] Renner R. Natural remediation of DDT, PCBs Debat. Environ Sci Technol1998;32:360–3.
[21] Sharma HD, Reddy KR. Geoenvironmental engineering. New Jersey: John Wiley& Sons, Inc; 2004.
www.elsevier.com/locate/nbt 31
Rese