orexin-a immunoreactivity and prepro-orexin mrna expression in hyperphagic rats induced by...
TRANSCRIPT

Orexin-A Immunoreactivity and Prepro-Orexin mRNA Expression inHyperphagic Rats Induced By Hypothalamic Lesions and Lactation
G. Sun, K. Narita, T. Murata, K. Honda and T. HiguchiDepartment of Physiology, Fukui Medical University, Matsuoka, Fukui, Japan.
Key words: orexin-A, prepro-orexin mRNA, ventromedial hypothalamus, paraventricular nucleus, lactation, immunohistochemistry,
lateral hypothalamus.
Abstract
Orexins are endogenous neuropeptides that potently facilitate appetite and food consumption. In thepresent study, we examined orexin immunoreactivity and prepro-orexin mRNA expression in thelateral hypothalamus by immunohistochemistry and competitive reverse transcription-polymerasechain reaction (RT-PCR) methods in different models of hyperphagia in rats. Hyperphagia wasinduced by lesions of either the ventromedial hypothalamus (VMHL) or the paraventricular nucleus(PVNL), and we also compared lactating rats to nonlactating controls. Both VMHL and PVNLincreased food intake and body weight compared to shams. On day 7 post lesion, serum leptin andinsulin concentrations exhibited 3.2- and 2.8-fold increases in VMHL rats, and nonsignificant 1.8- and1.8-fold increases in PVNL rats; there were significant decreases (48% and 33%) in lactating rats onday 12 postpartum compared to controls, respectively. Serum glucose concentrations were notsignificantly changed compared to controls in these rats. Quantification by image analysis suggeststhat VMHL significantly decreased the number and mean staining intensity of orexin-Aimmunoreactive neurones compared to those in the sham-lesioned group; while PVNL did notchange orexin-A immunoreactivity. Competitive RT-PCR analysis showed that VMHL significantlydecreased the prepro-orexin mRNA expression compared to those in the sham-lesioned group, andPVNL did not change it. Lactating rats on days 11–12 of lactation had significantly greater numberand mean staining intensity of orexin-A immunoreactive neurones, prepro-orexin mRNA expressionfood intake and body weight than nonlactating postpartum rats. Thus, changes in orexin-Aimmunoreactivity and prepro-orexin mRNA expression were not consistent between the hyperphagiamodels. These results suggest that the hyperphagia from VMHL or PVNL and lactating rats differ intheir involvement of orexin-A, and the change in circulating leptin and insulin concentrations may beinvolved in the change of orexin-A immunoreactivity in these rats.
Feeding behaviour is controlled by a complex feedbackmechanism involving both central and peripheral systems.The hypothalamus plays a central role in the integratedcontrol of feeding and energy homeostasis. Specifically, thelateral hypothalamic area (LHA) has long been considered asa feeding centre (1). LHA lesions cause anorexia (2) and caneven lead to the affected animal starving to death. Chemicaland electrical stimulation of LHA induce food intake (3) andthe neurones in this region fire spontaneously during naturallyoccurring feeding behaviour (4), as well as during hypogly-caemia (5). However, which neuropeptides or neuromodula-tors play the main role in this feeding behaviour was poorly
understood until orexins and melanin-concentrating hormonewere discovered and characterized in the LHA. At present,orexin-A and orexin-B subtypes have been identified in thebrain. Orexin-A when injected intracerebroventricularly (6, 7),even at low doses, into the LHA stimulates feeding (8). Inaddition, prepro-orexin mRNA expression is increased infasted rats (6), indicating that orexins are important cellularmediators that participate in the hypothalamic regulation offeeding behaviour.
Rats with either lesions of the ventromedial hypothalamus(VMHL) or the paraventricular nucleus (PVNL) have beenused as models for hyperphagia and obesity for a long time
Correspondence to: Guirong Sun, Department of Physiology, Fukui Medical University, Matsuoka, Fukui 910–1193, Japan (e-mail:
Journal of Neuroendocrinology, 2003, Vol. 15, 51–60
# 2003 Blackwell Publishing Ltd
(9, 10). The current understanding of the mechanisms ofhyperphagia, obesity and metabolic changes induced by bothlesions is very limited. In addition, lactation is a physiologicalstate characterized by a large energy demand (11) due to milkproduction. To meet the increased energy demand, foodintake is increased several-fold during lactation, associatingwith elevated neuropeptide Y (NPY) and its mRNA expres-sions in the hypothalamus (12, 13), and therefore the lactatinganimal is a physiological model of hyperphagia. However, themechanisms that drive hyperphagia during lactation are stillnot fully understood. Whether orexins are involved inhyperphagia induced by hypothalamic lesions and/or lacta-tion is not clear. The purposes of this study were to evaluatethe responses of orexin-A neurones in the LHA duringhyperphagia induced by VMHL, PVNL and lactation, and toexplore changes in orexin-A and prepro-orexin mRNAexpression using immunohistochemistry and competitivereverse transcription-polymerase chain reaction (RT-PCR)technique and the relationship with serum concentrations ofthe peripheral regulatory factors, leptin, insulin and glucose.
Materials and methods
Animals and treatment paradigm
Male Wistar rats, weighing 220–250 g, and female Wistar rats, weighing
210–250 g, were purchased from Charles-River (Yokohama, Japan). The rats
were kept in individual cages in air-conditioned rooms (23t2 uC) with lights
on from 06.00 to 18.00 h. Standard lab chow and tap water were available ad
libitum throughout the experiment. All the rats were allowed to adapt to the
laboratory for 7–10 days before being used for experiment. For the surgical
procedures of VMHL and PVNL, rats were anaesthetized with pentobarbital
sodium (45 mg/kg i.p.) and placed in a stereotaxic frame. The coordinates of
the VMH and PVNwere identified from an atlas of the rat brain with the upper
incisor bar 5.0 mm above the interaural line (14). For the VMHL, an electrode
was positioned at bregma, 0.8 mm lateral to the midsagittal line and 10.0 mm
ventral to the skull. For the PVNL, the electrode was placed 0.4 mm anterior
to bregma, 0.5 mm lateral to the midsagittal line and 8.4 mm ventral to the
skull. Bilateral VMHL and PVNL were produced by passing anodal direct
current (2.0 mA and 1.25 mA for 10 s, respectively) through the exposed tips of
insulated platinum electrodes with a rectal electrode serving as a cathode. For
sham lesions, electrodes were introduced 2.2 mm below the brain surface, but
no current was passed. On days 2, 7 and 21 post lesion, the rats were killed for
immunohistochemistry and verification of site of the VMHL and PVNL. For
lactation, oestrous cycles of rats were determined by daily vaginal smears and
proestrous rats were mated with conspecific male rats. Pregnant rats were
checked for the birth of pups in the mornings and the day of delivery was
considered as day 0 postpartum. All postpartum rats were divided into
nonlactating and lactating groups. For nonlactating rats, the pups were
removed on the day of delivery. For lactating rats, on day 2 postpartum, litters
were adjusted to 10 pups per dam. On day 11 or 12 postpartum, all the rats
were sacrificed.
Histological examination of VMHL and PVNL and orexin-A
immunohistochemistry
All the rats were deeply anaesthetized with pentobarbital and perfused
transcardially by a rapid bolus of 150 ml of 0.1 M phosphate buffer (PB) at
pH 7.4 followed by 500 ml of Zamboni’s solution between 09.00 and 11.00 h.
The brains were rapidly removed, postfixed in the same fixative for 24 h at 4 uCwith gentle shaking and submerged in 0.1 M PB containing 20% sucrose with
0.1% sodium azide at 4 uC. The brains were sectioned on a cryostat (Leica
CM1510, Nussloch, Germany) at 40 mm thick and processed for histological
examination of VMHL and PVNL and for orexin-A immunohistochemistry.
VMHL, PVNL and cytomorphological structure of sections were confirmed by
labelling one series of sections/rat from VMHL, PVNL and lactating rats
ranging from bregma x1.4 mm to bregma x3.8 mm with Neutral Red. The
rats with misplaced lesions were excluded and only those rats with typical
lesions (Fig. 1) were included in experiment and used for analyses. Following
the description of Broberger et al. (15) (that the most rostral orexin mRNA-
expressing cells were found in the sections x1.4 mm from the bregma and the
most caudal cells were observed in the sections x3.8 mm from Bregma)
sections ranging from bregma x1.4 mm to bregma x3.8 mm/rat from
VMHL, PVNL and lactating rats were processed for orexin-A immunohis-
tochemistry with their corresponding controls. Sections were pretreated with
1% hydrogen peroxide and then incubated in 1 : 50 normal goat serum
(Antibodies Incorporated, Davis, CA, USA) for 30 min. Rabbit antiorexin-A
antiserum (Peninsula Laboratories, San Carlos, CA, USA) was diluted in
1 : 20 000 and incubated for 24 h at 4 uC. The antibody used in this study is
specific to orexin-A of human, mouse and rat and has no cross reactivity with
orexin-B. After washing in phosphate-buffered saline (PBS), sections were
incubated in biotinylated antirabbit immunoglobulin G (Vector Laboratories,
Burlingame, CA, USA) at a dilution of 1 : 1000 containing 2% normal rat
serum for 1 h at room temperature. Sections were then incubated with avidin-
biotin complex (ABC kit, Vector Laboratories, Burlingame, CA, USA; 1 : 2000
in PBS) for 30 min, rinsed with PBS, and incubated in diaminobenzidine
tetrahydrochloride solution (DAB kit, Vector Laboratories) for 10 min. The
reaction was terminated by two successive rinses in 50 mM Tris-HCl buffer for
10 min. Sections were mounted onto gelatin-coated glass slides, dried at 37 uC,
(A) (B)
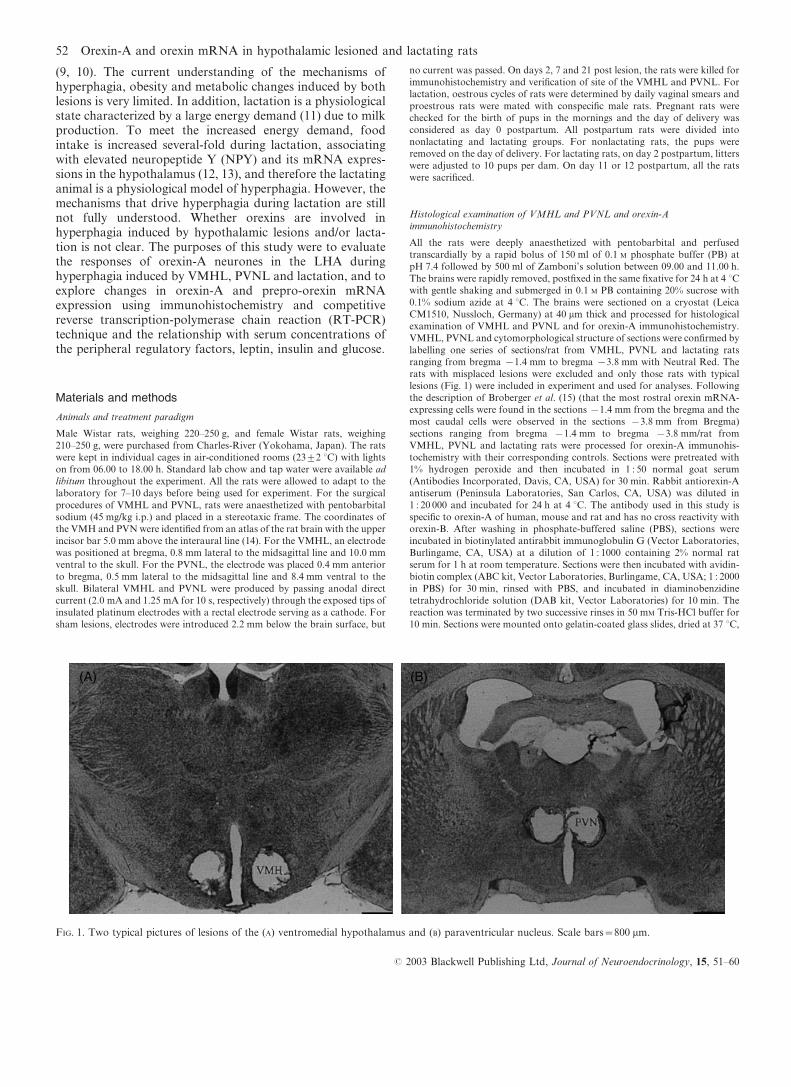
FIG. 1. Two typical pictures of lesions of the (A) ventromedial hypothalamus and (B) paraventricular nucleus. Scale bars=800 mm.
52 Orexin-A and orexin mRNA in hypothalamic lesioned and lactating rats
# 2003 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 15, 51–60

dehydrated in alcohol, cleared in xylene, and then coverslipped. The negative
controls were generated by omission of primary antiserum.
Quantitative analysis of immunoreactivity of orexin-A
The microscopic images (r40) of stained sections were transported to a
videomonitor using a digital camera (HC-2500 3CCD, Fujix, Tokyo, Japan)
attached to Vanox-S microscope (AH-2, Olympus, Tokyo, Japan) and
Fujifilm/Photograb-2500 software (version 1.1). The size of images taken by
digital camera was 1280r1000 pixels, which enclosed the population of
orexin-A neurones in unilateral hypothalamus of one section. For one section,
two images including left and right hypothalamic area were taken. To
distinguish positive orexin-A immunoreactive products, grey level threshold
was used (16). The MacSCOPE (Version 2.56, Mitani Corp., Fukui, Japan)
can distinguish 256 grey levels and give each pixel a grey level numbering
between 0 and 255, representing pure black (full light absorption) and pure
white (full light transmission), respectively. The grey level threshold defined the
upper grey level limit for selecting pixels for analysis; dark pixels that had grey
levels below the threshold were selected for analysis, whereas lighter pixels with
higher grey levels than the threshold were excluded. Grey level threshold was
determined by selecting the different positive immunoreactive products several
times so that it produced a colour overlay that covered individual stained
neurones. This grey level threshold was maintained throughout the analysis to
obtain consistent measurements. The MacSCOPE program determined the
average grey levels of individual positive neurones by summing values for all
pixels in a neurone divided by the total number of pixels and then the mean
grey level of orexin-A immunoreactive neurones in each image was calculated
by MacSCOPE program. Then the mean grey level was subtracted from 255 to
obtain values that were used to represent the staining intensity and were
directly rather than inversely related to staining intensity, and expressed in
arbitrary units (17). Neuronal counts were made for orexin-A labelled
neurones from each image (including the unilateral hypothalamic region)
according to the counting methods of MacSCOPE. The number of orexin-A
immunoreactive neurones counted represents the total number of neurones
labelled/image. Five sections (10 images) per rat, collected in every 200 mm of
tissue from bregma x2.6 mm to bregma x3.4 mm (18), were analysed.
Results are the mean of 10 images from five separate sections.
Competitive RT-PCR analysis of prepro-orexin mRNA expression
For competitive RT-PCR processing, VMHL and PVNL rats on day 7 post-
lesion and lactating rats on day 12 postpartum were killed by decapitation
from 10.00 to 11.00 h. The brain was quickly removed and the hypothalamus
was dissected from a frontal slice of brain by cutting between the optic chiasm
and the mammillary bodies, then coronally immediately below the anterior
commissure and finally sagittally through the perihypothalamic sulci and
immersed in liquid nitrogen and stored at x80 uC until used that enclose the
population of orexin neurones (6). In this experiment, only those VMHL and
PVNL rats with greater daily food intake and body weight gain on days 2 and 7
post-lesion were used for competitive RT-PCR processing. Total RNA was
isolated from individual hypothalamus using TRIzol Reagent (Gibco BRL,
Gaithersburg, MD, USA) according to the manufacturer’s instructions. The
total RNA concentration was determined by absorbance at 260 nm using a
spectrophotometer. Total RNA sample (1 mg/1 ml) was reverse transcribed
with 200 U SuperScript II reverse transcriptase according to the supplier’s
manual (Gibco BRL) and 10 pmol 9-mer random primer. For the construction
of normal and mutant DNAs of prepro-orexin and b-actin, the oligonucleotideprimers were used for prepro-orexin (19): 5k-CGGATTGCCTCTCCCT-
GAGC -3k for upper primer, 5k-GCGAGGAGAGGGGAAAGTTA-3k forlower primer and b-actin amplification: 5k-GTCACCCACACTGTGCC-
CATCT-3k for upper primer, 5k-ACAGAGTACTTGCGCTCAGGAG-3k forlower primer (20). Each PCR product was subcloned into pGEMT easy vector
(Promega, Madison, WI, USA). Mutated DNAs, as competitors for assay,
were prepared by deleting 30–40% nucleotides in length from normal DNA.
Deletion of normal DNA in prepro-orexin was performed by amplifying
plasmid with lower primer and mutant upper primer, 5k-CGGATTGCC-
TCTCCCTGAGCTCTCTACGAA-3k. Mutant primers were designed to
amplify shorter DNA than primary (normal) DNA amplified by RT-PCR
and were linked with 3k end of normal upper primer. Mutant DNAs for prepro-
orexin amplified with mutant upper primer and lower primer were subcloned
into pGEMT vectors. For deletion in b-actin, the inserted vector was digested
with Eco0190I and AvaI, reacted with T4 DNA polymerase (Gibco BRL) to
make the both restriction sides blunt and ligated with T4 ligase (Gibco BRL).
Normal and mutant DNAs were amplified with M13 primer and purified with
rapid PCR Clean-up system (Gibco BRL). Concentrations of amplified DNAs
were assessed with a spectrophotometer at 260 nm. Normal and mutant DNAs
were used as standard DNA and competitor, respectively. All the samples and
the standards in each experiment were assayed in a single run. Constant
amounts of competitor DNA for prepro-orexin (1.2 fM) and b-actin (5.5 pM)
were added to all samples and standards. The PCR amplification was carried
out in PCR buffer (50 mM KCl, 2 mM MgCl2 and 20 mM Tris-HCl, pH 8.4),
with 0.2 mmol primers and 2.5 U Taq polymerase (Perkin Elmer) in a total
volume of 20 ml. The optical conditions for amplification were: the initial
denaturation at 94 uC for 10 min followed by 40 cycles at 94 uC for 1 min, 62 uC(prepro-orexin) or 55 uC (b-actin) for 1 min and 72 uC for 1 min. An additional
extension was 10 min at 72 uC. The PCR products were electrophoresed in a
2% agarose gel, and then stained with ethidium bromide (0.5 mg/ml) and
analyzed using NIH mage software. The linear standard curve was obtained
and no band was detected when the PCR reaction was carried out with total
RNA sample without the reverse transcription to check contamination with
genomic DNA or non-specific reaction in each experiment. The ratio of native
prepro-orexin mRNA-derived DNA to competitor DNA-derived DNA of
each sample was obtained and calibrated from a standard curve as the amount
of standard total mRNA. The relative orexin mRNA levels in VMHL, PVNL
and lactating rats were expressed by taking, as reference 100, the mean value of
the orexin mRNA/b-actin mRNA ratio found in the control rats.
Measurements of serum hormone and glucose
Other sets of VMHL and PVNL rats on day 7 post lesion and lactating rats on
day 12 postpartum were decapitated at 10.00 to 11.00 h. After decapitation,
trunk blood samples were collected and the brain was removed quickly and
fixed in Zamboni’s solution for 72 h at 4 uC for histological examinations of
VMHL and PVNL. Sera were separated before 12.00 h and stored at x20 uC.Serum leptin concentrations were determined by a specific radioimmunoassay
kit (Linco Research Inc., St Charles, MO, USA) for determining rat leptin.
Serum insulin concentrations were measured by an enzyme immunoassay kit
(Mercodia AB, Seminariegatan, Uppsala, Sweden) for rat insulin assay
according to the protocols described in the kit instructions. Serum glucose
concentrations were determined by the glucose oxidase method (Iatron
Laboratories, Inc., Tokyo, Japan).
Statistical analysis
Results are presented as the meantSEM. In VMHL and PVNL experiments
the statistical significance for all analyses was determined by one-way ANOVA
followed by Scheffe’s F-test and, in lactating experiments, data were analysed
by Student’s t-test. P<0.05 was regarded as a statistically significant
difference.
Results
Effects of VMHL, PVNL and lactation on food intake andbody weight
Daily food intake and body weight were monitored on days 2,7 and 21 following the lesions and the food intake and bodyweight data from rats with misplaced lesions were discarded.As shown in Table 1, the daily food intake on days 2, 7 and 21post surgery in both VMHL and PVNL rats were muchgreater than those in sham controls (P<0.001). Both lesionedgroups gained significantly greater weight at the three post-surgery intervals than sham-lesioned controls (P<0.001), andthe VMHL rats gained more weight than the PVNL rats at thethree post surgery intervals (P<0.001). Lactating rats in ourstudy increased their daily food intake three-fold and gainedmore body weight on day 11 postpartum compared withnonlactating rats (P<0.001) (Table 2).
Orexin-A and orexin mRNA in hypothalamic lesioned and lactating rats 53
# 2003 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 15, 51–60
Effects of VMHL, PVNL and lactation on the number and
staining intensity of orexin-A immunoreactive neurones
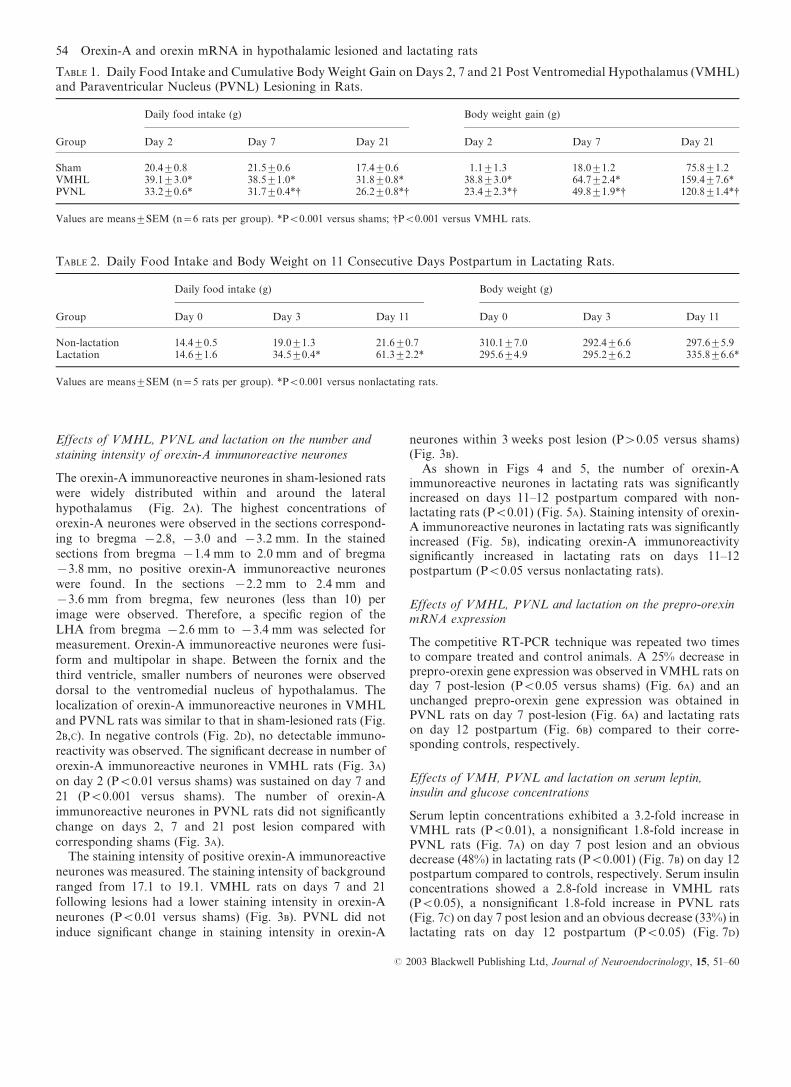
The orexin-A immunoreactive neurones in sham-lesioned rats
were widely distributed within and around the lateral
hypothalamus (Fig. 2A). The highest concentrations of
orexin-A neurones were observed in the sections correspond-
ing to bregma x2.8, x3.0 and x3.2 mm. In the stained
sections from bregma x1.4 mm to 2.0 mm and of bregma
x3.8 mm, no positive orexin-A immunoreactive neurones
were found. In the sections x2.2 mm to 2.4 mm and
x3.6 mm from bregma, few neurones (less than 10) per
image were observed. Therefore, a specific region of the
LHA from bregma x2.6 mm to x3.4 mm was selected for
measurement. Orexin-A immunoreactive neurones were fusi-
form and multipolar in shape. Between the fornix and the
third ventricle, smaller numbers of neurones were observed
dorsal to the ventromedial nucleus of hypothalamus. The
localization of orexin-A immunoreactive neurones in VMHL
and PVNL rats was similar to that in sham-lesioned rats (Fig.
2B,C). In negative controls (Fig. 2D), no detectable immuno-
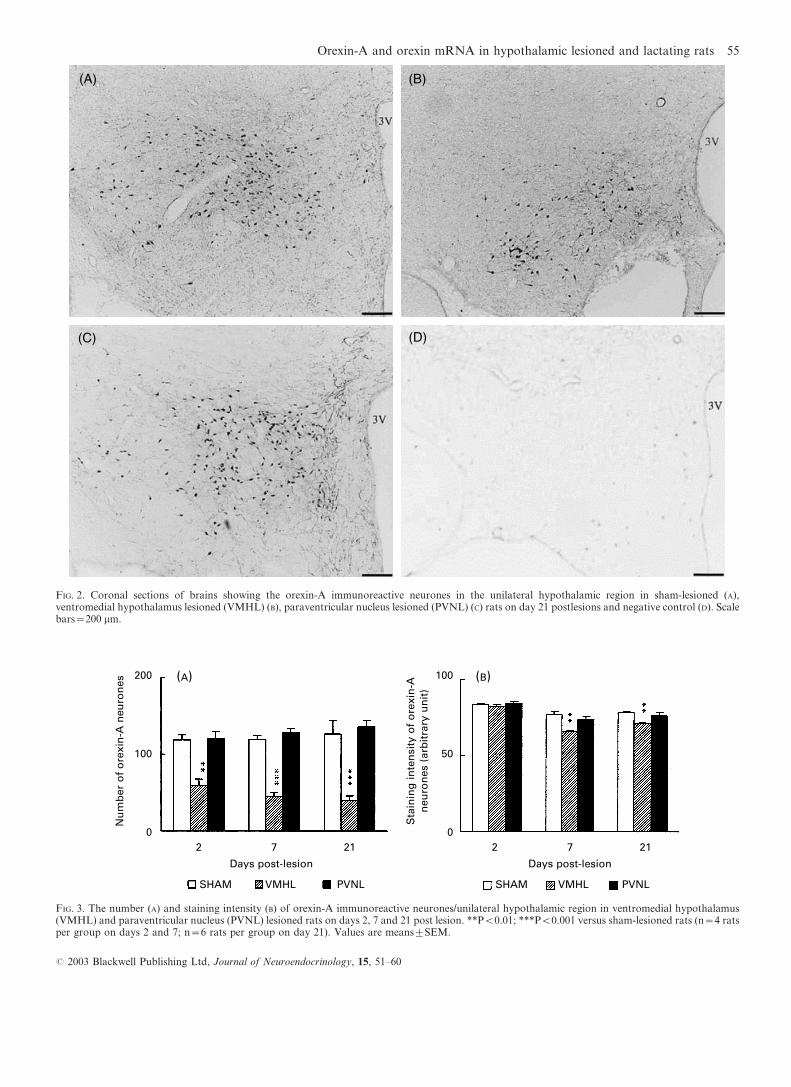
reactivity was observed. The significant decrease in number of
orexin-A immunoreactive neurones in VMHL rats (Fig. 3A)
on day 2 (P<0.01 versus shams) was sustained on day 7 and
21 (P<0.001 versus shams). The number of orexin-A
immunoreactive neurones in PVNL rats did not significantly
change on days 2, 7 and 21 post lesion compared with
corresponding shams (Fig. 3A).The staining intensity of positive orexin-A immunoreactive
neurones was measured. The staining intensity of background
ranged from 17.1 to 19.1. VMHL rats on days 7 and 21
following lesions had a lower staining intensity in orexin-A
neurones (P<0.01 versus shams) (Fig. 3B). PVNL did not
induce significant change in staining intensity in orexin-A
neurones within 3 weeks post lesion (P>0.05 versus shams)(Fig. 3B).
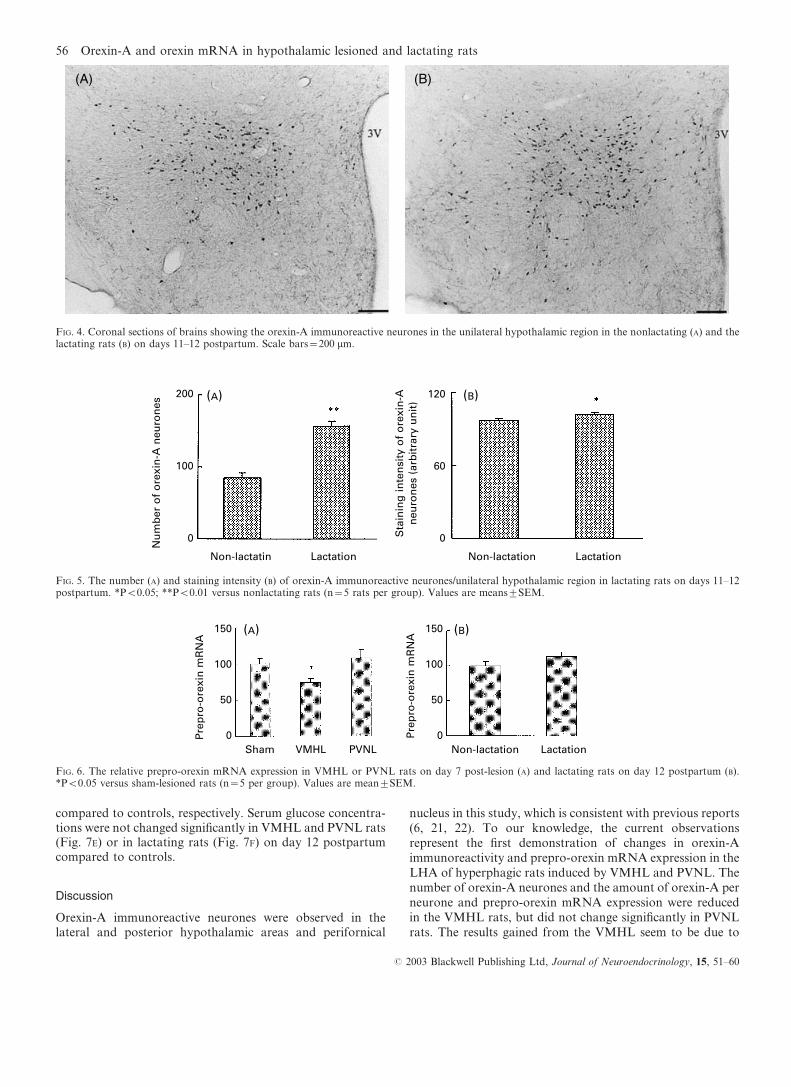
As shown in Figs 4 and 5, the number of orexin-Aimmunoreactive neurones in lactating rats was significantlyincreased on days 11–12 postpartum compared with non-lactating rats (P<0.01) (Fig. 5A). Staining intensity of orexin-A immunoreactive neurones in lactating rats was significantlyincreased (Fig. 5B), indicating orexin-A immunoreactivitysignificantly increased in lactating rats on days 11–12postpartum (P<0.05 versus nonlactating rats).
Effects of VMHL, PVNL and lactation on the prepro-orexinmRNA expression
The competitive RT-PCR technique was repeated two timesto compare treated and control animals. A 25% decrease inprepro-orexin gene expression was observed in VMHL rats onday 7 post-lesion (P<0.05 versus shams) (Fig. 6A) and anunchanged prepro-orexin gene expression was obtained inPVNL rats on day 7 post-lesion (Fig. 6A) and lactating ratson day 12 postpartum (Fig. 6B) compared to their corre-sponding controls, respectively.
Effects of VMH, PVNL and lactation on serum leptin,insulin and glucose concentrations
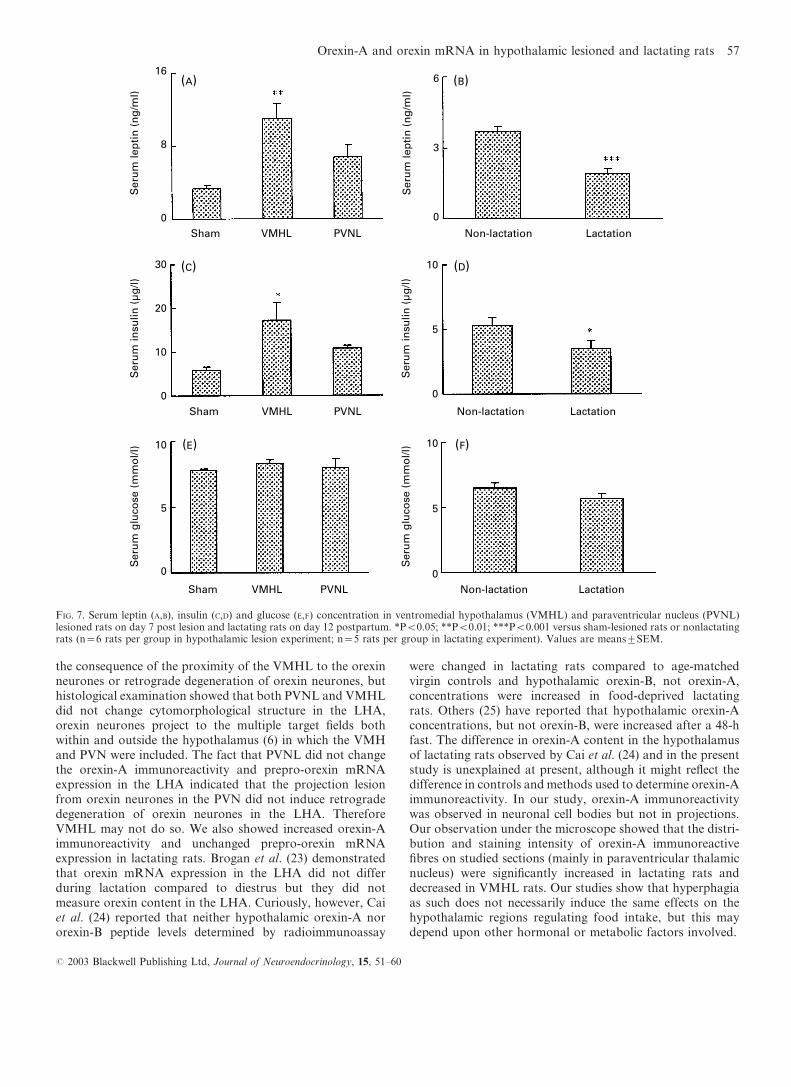
Serum leptin concentrations exhibited a 3.2-fold increase inVMHL rats (P<0.01), a nonsignificant 1.8-fold increase inPVNL rats (Fig. 7A) on day 7 post lesion and an obviousdecrease (48%) in lactating rats (P<0.001) (Fig. 7B) on day 12postpartum compared to controls, respectively. Serum insulinconcentrations showed a 2.8-fold increase in VMHL rats(P<0.05), a nonsignificant 1.8-fold increase in PVNL rats(Fig. 7C) on day 7 post lesion and an obvious decrease (33%) inlactating rats on day 12 postpartum (P<0.05) (Fig. 7D)
TABLE 2. Daily Food Intake and Body Weight on 11 Consecutive Days Postpartum in Lactating Rats.
Group
Daily food intake (g) Body weight (g)
Day 0 Day 3 Day 11 Day 0 Day 3 Day 11
Non-lactation 14.4t0.5 19.0t1.3 21.6t0.7 310.1t7.0 292.4t6.6 297.6t5.9Lactation 14.6t1.6 34.5t0.4* 61.3t2.2* 295.6t4.9 295.2t6.2 335.8t6.6*
Values are meanstSEM (n=5 rats per group). *P<0.001 versus nonlactating rats.
TABLE 1. Daily Food Intake and Cumulative BodyWeight Gain on Days 2, 7 and 21 Post Ventromedial Hypothalamus (VMHL)and Paraventricular Nucleus (PVNL) Lesioning in Rats.
Group
Daily food intake (g) Body weight gain (g)
Day 2 Day 7 Day 21 Day 2 Day 7 Day 21
Sham 20.4t0.8 21.5t0.6 17.4t0.6 1.1t1.3 18.0t1.2 75.8t1.2VMHL 39.1t3.0* 38.5t1.0* 31.8t0.8* 38.8t3.0* 64.7t2.4* 159.4t7.6*PVNL 33.2t0.6* 31.7t0.4*† 26.2t0.8*† 23.4t2.3*† 49.8t1.9*† 120.8t1.4*†
Values are meanstSEM (n=6 rats per group). *P<0.001 versus shams; †P<0.001 versus VMHL rats.
54 Orexin-A and orexin mRNA in hypothalamic lesioned and lactating rats
# 2003 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 15, 51–60
(A) (B)200
100
0
Nu
mb
er o
f o
rexi
n-A
neu
ron
es
Sta
inin
g in
ten
sity
of
ore
xin
-An
euro
nes
(ar
bit
rary
un
it)
Days post-lesion Days post-lesion
100
50
02 7 21 2 7 21
SHAM VMHL PVNL SHAM VMHL PVNL
FIG. 3. The number (A) and staining intensity (B) of orexin-A immunoreactive neurones/unilateral hypothalamic region in ventromedial hypothalamus(VMHL) and paraventricular nucleus (PVNL) lesioned rats on days 2, 7 and 21 post lesion. **P<0.01; ***P<0.001 versus sham-lesioned rats (n=4 ratsper group on days 2 and 7; n=6 rats per group on day 21). Values are meanstSEM.
(A) (B)
(C) (D)
FIG. 2. Coronal sections of brains showing the orexin-A immunoreactive neurones in the unilateral hypothalamic region in sham-lesioned (A),ventromedial hypothalamus lesioned (VMHL) (B), paraventricular nucleus lesioned (PVNL) (C) rats on day 21 postlesions and negative control (D). Scalebars=200 mm.
Orexin-A and orexin mRNA in hypothalamic lesioned and lactating rats 55
# 2003 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 15, 51–60
compared to controls, respectively. Serum glucose concentra-tions were not changed significantly in VMHL and PVNL rats(Fig. 7E) or in lactating rats (Fig. 7F) on day 12 postpartumcompared to controls.
Discussion
Orexin-A immunoreactive neurones were observed in thelateral and posterior hypothalamic areas and perifornical
nucleus in this study, which is consistent with previous reports(6, 21, 22). To our knowledge, the current observationsrepresent the first demonstration of changes in orexin-Aimmunoreactivity and prepro-orexin mRNA expression in theLHA of hyperphagic rats induced by VMHL and PVNL. Thenumber of orexin-A neurones and the amount of orexin-A perneurone and prepro-orexin mRNA expression were reducedin the VMHL rats, but did not change significantly in PVNLrats. The results gained from the VMHL seem to be due to
(A) (B)
Nu
mb
er o
f o
rexi
n-A
neu
ron
es
Non-lactatin Lactation Non-lactation Lactation
200
100
0
120
60
0Sta
inin
g in
ten
sity
of
ore
xin
-An
euro
nes
(ar
bit
rary
un
it)
FIG. 5. The number (A) and staining intensity (B) of orexin-A immunoreactive neurones/unilateral hypothalamic region in lactating rats on days 11–12postpartum. *P<0.05; **P<0.01 versus nonlactating rats (n=5 rats per group). Values are meanstSEM.
(A) (B)150
100
50
0Pre
pro
-ore
xin
mR
NA
Sham VMHL PVNL
150
100
50
0Pre
pro
-ore
xin
mR
NA
Non-lactation Lactation
FIG. 6. The relative prepro-orexin mRNA expression in VMHL or PVNL rats on day 7 post-lesion (A) and lactating rats on day 12 postpartum (B).*P<0.05 versus sham-lesioned rats (n=5 per group). Values are meantSEM.
(A) (B)
FIG. 4. Coronal sections of brains showing the orexin-A immunoreactive neurones in the unilateral hypothalamic region in the nonlactating (A) and thelactating rats (B) on days 11–12 postpartum. Scale bars=200 mm.
56 Orexin-A and orexin mRNA in hypothalamic lesioned and lactating rats
# 2003 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 15, 51–60
the consequence of the proximity of the VMHL to the orexinneurones or retrograde degeneration of orexin neurones, buthistological examination showed that both PVNL and VMHLdid not change cytomorphological structure in the LHA,orexin neurones project to the multiple target fields bothwithin and outside the hypothalamus (6) in which the VMHand PVN were included. The fact that PVNL did not changethe orexin-A immunoreactivity and prepro-orexin mRNAexpression in the LHA indicated that the projection lesionfrom orexin neurones in the PVN did not induce retrogradedegeneration of orexin neurones in the LHA. ThereforeVMHL may not do so. We also showed increased orexin-Aimmunoreactivity and unchanged prepro-orexin mRNAexpression in lactating rats. Brogan et al. (23) demonstratedthat orexin mRNA expression in the LHA did not differduring lactation compared to diestrus but they did notmeasure orexin content in the LHA. Curiously, however, Caiet al. (24) reported that neither hypothalamic orexin-A nororexin-B peptide levels determined by radioimmunoassay
were changed in lactating rats compared to age-matchedvirgin controls and hypothalamic orexin-B, not orexin-A,concentrations were increased in food-deprived lactatingrats. Others (25) have reported that hypothalamic orexin-Aconcentrations, but not orexin-B, were increased after a 48-hfast. The difference in orexin-A content in the hypothalamusof lactating rats observed by Cai et al. (24) and in the presentstudy is unexplained at present, although it might reflect thedifference in controls and methods used to determine orexin-Aimmunoreactivity. In our study, orexin-A immunoreactivitywas observed in neuronal cell bodies but not in projections.Our observation under the microscope showed that the distri-bution and staining intensity of orexin-A immunoreactivefibres on studied sections (mainly in paraventricular thalamicnucleus) were significantly increased in lactating rats anddecreased in VMHL rats. Our studies show that hyperphagiaas such does not necessarily induce the same effects on thehypothalamic regions regulating food intake, but this maydepend upon other hormonal or metabolic factors involved.
(A)16
8
0
Ser
um
lep
tin
(n
g/m
l)
Sham VMHL PVNL
(C)30
20
10
0
Ser
um
insu
lin (
µg
/l)
Sham VMHL PVNL
(E)10
5
0
Ser
um
glu
cose
(m
mo
l/l)
Sham VMHL PVNL
(B)6
3
0
Ser
um
lep
tin
(n
g/m
l)
(D)10
5
0S
eru
m in
sulin
(µ
g/l)
(F)10
5
0
Ser
um
glu
cose
(m
mo
l/l)
Non-lactation Lactation
Non-lactation Lactation
Non-lactation Lactation
FIG. 7. Serum leptin (A,B), insulin (C,D) and glucose (E,F) concentration in ventromedial hypothalamus (VMHL) and paraventricular nucleus (PVNL)lesioned rats on day 7 post lesion and lactating rats on day 12 postpartum. *P<0.05; **P<0.01; ***P<0.001 versus sham-lesioned rats or nonlactatingrats (n=6 rats per group in hypothalamic lesion experiment; n=5 rats per group in lactating experiment). Values are meanstSEM.
Orexin-A and orexin mRNA in hypothalamic lesioned and lactating rats 57
# 2003 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 15, 51–60

A regulatory role of leptin on orexin neurones has beenspeculated (26, 27). Food deprivation associated with amarked decrease in serum leptin concentration (28), increasedprepro-orexin mRNA in the LHA (6). The effect of fooddeprivation on prepro-orexin mRNA can be reversed byexogenous leptin administration (26) and chronic treatmentwith leptin for 7 days has been reported to significantlydecrease orexin-A concentration in the LHA (27). Funahashiet al. (29) also reported that leptin inhibited increase of[Ca2+]i of glucose-sensitive arcuate nucleus and LHAneurones and enhanced the increase of glucose-sensitiveVMH neurones induced by a reduction of the glucose con-centration via the leptin receptor expressed in these cells. Inaddition, recent data have indicated that orexin neuronesin the LHA express leptin receptors (30). Serum leptinconcentration in VMHL rats exhibited progressive increasesover the 21 days post lesion period in fed state (31) and theincrease in plasma leptin concentrations occurred in the fastedstate in VMHL rats (32). These results suggest that VMHLcan stimulate leptin production, even in the absence ofincreased adiposity or hyperphagia. As expected, we observedthat serum leptin concentration in VMHL rats increased3.2-fold on day 7 post lesion. Taken together, the presentresults suggest that increased leptin may contribute to thedecrease in orexin-A immunoreactivity in the LHA of VMHLrats through the activation of specific receptors located onorexin neurones. In addition, we observed a slight butinsignificant increase in serum leptin concentrations on day7 post lesion in PVNL rats compared with shams on the sameday. In previous studies, PVNL induced by colchicine micro-injection caused an increased plasma leptin concentration(P<0.05) on day 5 compared to shams on day 0 (33).Furthermore, Banks et al. (34) observed that obesity-inducinglesions of the PVN and posterodorsal amygdala, but not theVMH were associated with decreased uptake of leptin by theblood–brain barrier. Therefore, we suggest that unchanged orincreased leptin concentrations associated with decreaseduptake of leptin by the blood–brain barrier may be relatedto the unchanged orexin-A immunoreactivity in the LHAin PVNL rats. In lactating rats, the decrease in leptin con-centration in blood (35) and leptin receptor mRNA in VMH(23) may underlie the chronic increases in food intake duringlactation. Consistent with previous reports, the decrease inserum leptin concentration was also observed in lactating ratsin this study. Therefore, the lower circulating leptin concen-trations may contribute to the increase in orexin-A immuno-reactivity in lactating rats.
Insulin stimulates the expression of the ob gene inadipocytes, thereby increasing plasma leptin concentrations(36). In the present study, we observed that serum insulinconcentrations increased 2.8-fold in VMHL rats, 1.8-fold inPVNL rats on day 7 post lesion and decreased in lactating ratson day 12 postpartum. Changes in serum insulin concentra-tions in these rats paralleled their serum leptin concentrationsand are consistent with previous reports that hyperinsulin-aemia and hyperleptinaemia are characteristic in VMHL rats(31, 32). Plasma insulin concentration in obese PVNL rats onday 9 after lesion were approximately 1.9-fold increased andwere significantly lower than that in VMHL rats (37), andfasting and lactation decreased the insulin and leptin
concentrations in the circulation (38, 39). The importanceof the combined leptin and insulin increase is supported by astudy on the suppression of hypothalamic NPY mRNAexpression induced by chronic i.c.v. administration of NPY(40), which demonstrated that both are necessary for over-feeding-induced suppression of arcuate NPY mRNA expres-sion. Overall, these data suggest that the combination ofincreased leptin and insulin signalling may be required forsuppression of LHA orexin-A immunoreactivity.
Some (25%) LHA neurones are thought to be glucose-sensitive, being inhibited by increased circulating glucose andstimulated by decreased glucose (1, 41). Plasma glucose infasted rats is lower than controls (38), and orexin mRNAexpressions are up-regulated in fasted rats (6). Acute (6 h)hypoglycaemia induced by insulin without feeding alsoincreases prepro-orexin mRNA expression (38). Loweringglucose concentrations increases [Ca2+]i in orexin neuronesisolated from the rat LHA (42), and these data suggest thatplasma glucose concentrations may contribute to the negativeregulation of orexin immunoreactivity and prepro-orexinmRNA expression. However, we showed that orexin-Aimmunoreactivity and prepro-orexin mRNA expression inthe LHA decreased in VMHL rats and increased in lactatingrats in spite of their unchanged blood glucose concentration(Fig. 7). In addition, Tokunaga et al. (37) detected a lowerplasma glucose level in obese VMHL rats at 08.00 h,not at 17.00 h. Thus, it is difficult to explain the changes inorexin-A immunoreactivity and prepro-orexin mRNA interms of plasma glucose in these rats.
In addition, NPY and NPY mRNA expression inhypothalamus are elevated in underfeeding (43, 44) andlactating rats (12, 13) and decreased in electrolytic (31) orcolchicine-lesioned (45) VMHL rats. In this study, weobserved that orexin-A immunoreactivity in the LHA isincreased in lactating rats similar to fasted rats and decreasedin electrolytic VMHL rats. Thus, the involvement of orexin-Ain these rats appeared to be similar to NPY. However, neitherNPY nor orexin-A content exhibits consistent changes bywhich the hyperphagia observed in VMHL and lactating ratscan be explained. Considering both NPY and orexin-Aincreases in fasting and lactating rats, the decrease in orexin-Ain VMHL rats was unexpected. Dube et al. (31) proposed thatincreased sensitivity to NPY and leptin resistance contributedto the hyperphagia in VMHL rats, but orexigenic and/oranorexigenic factor(s) other than orexin and NPY may beinvolved in the suppressive action of the VMHL. Similarhypersensitivity to orexins in VMHL rats therefore remains tobe examined.
Although VMHL and PVNL produced obesity, they did soby inducing different changes in feeding patterns (34). PVNLincreased food intake during both dark and light periods,whereas VMHL resulted in increased food intake only duringthe light period and a tendency for reduced feeding during thedark period. These two hypothalamic obese syndromes aredifferent in many other respects, such as ambulatory activity(46) and sympathetic nerve activity to brown adipose tissue(47). In this study, we observed that orexin-A immuno-reactivity in the LHA was also different between VMHL andPVNL rats. Recently, it was reported that i.c.v. administratedorexin-A increases sympathetic nerve activity in rats (48).
58 Orexin-A and orexin mRNA in hypothalamic lesioned and lactating rats
# 2003 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 15, 51–60

These findings suggest that the decrease in orexin-Aimmunoreactivity in the LHA may be related to the changesin sympathetic nerve activity in VMHL rats and the lattermight explain the difference of orexin-A immunoreactivity inthe LHA between the two models of hyperphagia used in thepresent study.
In conclusion, we provide evidence that orexin-A immu-noreactivity was down-regulated in obese VMHL rats,unchanged in PVNL rats and up-regulated in lactating rats.Thus, orexin-A neurones did not show consistent changes inthe hyperphagia models induced by VMHL, PVNL andlactation. These differences are probably due to differences inmechanisms by which hyperphagia are induced in these rats.
Acknowledgements
The authors gratefully acknowledge comments and assistance with editing the
manuscript by Drs A. J. Douglas and C. Okere.
Accepted 15 August 2002
References
1 Bernardis LL, Bellinger LL. The lateral hypothalamlic area revisited:
Ingestive behavior. Neurosci Biobehav Rev 1996; 20: 189–287.
2 Wolf G. Orexins: a newly discovered family of hypothalamic regulators of
food intake. Nutr Rev 1998; 56: 172–173.
3 Shiraishi T. Noradrenergic neurons modulate lateral hypothalamic
chemical and electrical stimulation-induced feeding by sated rats. Brain
Res Bull 1991; 27: 347–351.
4 Katafuchi T, Oomura Y, Yoshimatsu H. Single neuron activity in the rat
lateral hypothalamus during 2-deoxy-D-glucose induced and natural
feeding behavior. Brain Res 1985; 359: 1–9.
5 Himmi T, Boyer A, Orsini JC. Changes in lateral hypothalamic neuronal
activity accompanying hyper- and hypoglycemias. Physiol Behav 1988; 44:
347–354.
6 Sakurai T, Amemiya A, Ishii M, Matsuzaki I, Chemelli RM, Tanaka H,
Williams SC, Richarson JA, Kozlowski GP, Wilson S, Arch JR,
Buckingham RE, Haynes AC, Carr SA, Annan RS, McNulty DE, Liu
WS, Terrett JA, Elshourbagy NA, Bergsma DJ, Yanagisawa M. Orexins
and orexin receptors: a family of hypothalamic neuropeptides and G
protein-coupled receptors that regulate feeding behavior. Cell 1998;
92: 573–585.
7 Edwards CM, Abusnana S, Sunter D, Murphy KG, Ghatei MA, Bloom
SR. The effect of the orexins on food intake. Comparison with
neuropeptide Y, melanin-concentrating hormone and galanin.
J Endocrinol 1999; 160: R7–R12.
8 Dube MG, Kalra SP, Kalra PS. Food intake elicited by central
administration of orexins/hypocretins: identification of hypothalamic
sites of action. Brain Res 1999; 842: 473–477.
9 Hetherington AW, Ranson SW. Hypothalamic lesion and adiposity in the
rats. Anat Res 1940; 78: 149–172.
10 Leibowitz SF, Hammer NJ, Chang K. Hypothalamic paraventricular
nucleus lesion produce overeating and obesity in the rats. Physiol Behav
1981; 27: 1031–1040.
11 Ota K, Yokoyama A. Body weight and food consumption of lactating
rats. Effects of ovariectomy and of arrest and resumption of suckling.
J Endocrinol 1967; 38: 251–261.
12 Pickavance L, Dryden S, Hopkins D, Bing C, Frankish H, Wang Q,
Vernon RG, Williams G. Relationships between hypothalamic neuropep-
tide Y and food intake in the lactating rat. Peptides 1996; 17: 577–582.
13 Smith MS. Lactation alters neuropeptide-Y and proopiomelanocortin
gene expression in the arcuate nucleus of the rat. Endocrinology 1993; 133:
1258–1265.
14 Pellegrino LJ, Pellegrino AS, Cushman AJ. A Stereotaxic Atlas of the Rat
Brain. New York: Plenum Press, 1979.
15 Broberger C, De Lecea L, Sutcliffe JG, Hokfelt T. Hypocretin/orexin-
and melanin-concentrating hormone-expressing cells form distinct
populations in the rodent lateral hypothalamus: relationship to the
neuropeptide Y and agouti gene-related protein systems. J Comp Neurol
1998; 402: 460–474.
16 Weaver JR, Au JL. Comparative scoring by visual and image analysis of
cells in human solid tumors labeled for proliferation markers. Cytometry
1997; 27: 189–199.
17 Sudhaker Rao D, Han ZH, Phillips ER, Palnitkar S, Parfitt AM.
Reduced vitamin D receptor expression in parathyroid adenomas:
implications for pathogenesis. Clin Endocrinol 2000; 53: 373–381.
18 Paxinos G, Watson C. The Rat Brain in Stereotaxic Coordinates. Tokyo:
Academic Press, 1986.
19 Lopez M, Senaris R, Gallego R, Garcia-Caballero T, Lago F, Seoane L,
Casanueva F, Dieguez C. Orexin receptors are expressed in the adrenal
medulla of the rat. Endocrinology 1999; 140: 5991–5994.
20 Liu CX, Takahashi S, Murata T, Hashimoto K, Agatsuma T,
Matsukawa S, Higuchi T. Changes in oxytocin receptor mRNA in the
rat uterus measured by competitive reverse transcription-polymerase
chain reaction. J Endocrinol 1996: 15: 479–486.
21 Cutler DJ, Morris R, Sheridhar V, Wattam TA, Holmes S, Patel S, Arch
JR, Wilson S, Buckingham RE, Evans ML, Leslie RA, Williams G.
Differential distribution of orexin-A and orexin-B immunoreactivity in
the rat brain and spinal cord. Peptides 1999; 20: 1455–1470.
22 Peyron C, Tighe DK, van den Pol AN, de Lecea L, Heller HC,
Sutcliffe JG, Kilduff TS. Neurons containing hypocretin (orexin)
project to multiple neuronal systems. J Neurosci 1998; 18: 9996–10015.
23 Brogan RS, Grove KL, Smith MS. Differential regulation of leptin
receptor but not orexin in the hypothalamus of the lactating rat.
J Neuroendocrinol 2000; 12: 1077–1086.
24 Cai XJ, Denis R, Vernon RG, Clapham JC, Wilson S, Arch JR, Williams
G. Food restriction selectively increases hypothalamic orexin-B levels in
lactating rats. Regul Pept 2001; 97: 163–168.
25 Mondal MS, Nakazato M, Date Y, Murakami N, Yanagisawa M,
Matsukura S. Widespread distribution of orexin in rat brain and its regu-
lation upon fasting. Biochem Biophys Res Commun 1999; 256: 495–499.
26 Lopez M, Seoane L, Garcia MC, Lago F, Casanueva FF, Senaris R,
Dieguez C. Leptin regulation of prepro-orexin and orexin receptor
mRNA level in the hypothalamus. Biochem Biophys Res Commun 2000;
269: 41–45.
27 Beck B, Richy S. Hypothalamic hypocretin/orexin and neuropeptide
Y: divergent interaction with energy depletion and leptin. Biochem
Biophys Res Commun 1999; 258: 119–122.
28 Li H, MathenyM, Tumer N, Scarpace PJ. Aging and fasting regulation of
leptin and hypothalamic neuropeptide Y gene expression. Am J Physiol
1998; 275: E405–E411.
29 Funahashi H, Yada T, Muroya S, Takigawa M, Ryushi T, Horie S,
Nakai Y, Shioda S. The effect of leptin on feeding-regulating neurons in
the rat hypothalamus. Neurosci Lett 1999; 264: 117–120.
30 Hakansson M, de Lecea L, Sutcliffe JG, Yanagisawa M, Meister B.
Leptin receptor- and STAT3-immunoreactivities in hypocretin/orexin
neurones of the lateral hypothalamus. J Neuroendocrinol 1999; 11:
653–663.
31 Dube MG, Xu B, Kalra PS, Sninsky CA, Kalra SP. Disruption in
neuropeptide Y and leptin signaling in obese ventromedial hypothalamic-
lesioned rats. Brain Res 1999; 816: 38–46.
32 Suga A, Hirano T, Kageyama H, Kashiba M, Oka J, Osaka T, Namba Y,
Tsuji M, Miura M, Adachi M, Inoue S. Rapid increase in circulating
leptin in ventromedial hypothalamus-lesioned rats: role of hyperinsul-
inemia and implication for upregulation mechanism. Diabetes 1999;
48: 2034–2038.
33 Choi S, Sparks R, Clay M, Dallman MF. Rats with hypothalamic obesity
are insensitive to central leptin injections. Endocrinology 1999; 140:
4426–4433.
34 Banks WA, King BM, Rossiter KN, Olson RD, Olson GA, Kastin AJ.
Obesity-inducing lesions of the central nervous system alter leptin uptake
by the blood–brain barrier. Life Sci 2001; 69: 2765–2773.
35 Johnstone LE, Higuchi T. Food intake and leptin during pregnancy and
lactation. Prog Brain Res 2001; 133: 215–227.
36 Cusin I, Sainsbury A, Doyle P, Rohner-Jeanrenaud F, Jeanrenaud B. The
ob gene and insulin. A relationship leading to clues to the understanding
of obesity. Diabetes 1995; 44: 1467–1470.
37 Tokunaga K, Fukushima M, Kemnitz JW, Bray GA. Comparison of
ventromedial and paraventricular lesions in rats that become obese.
Am J Physiol 1986; 251: R1221–R1227.
Orexin-A and orexin mRNA in hypothalamic lesioned and lactating rats 59
# 2003 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 15, 51–60

38 Cai XJ, Widdowson PS, Harrold J, Wilson S, Buckingham RE, Arch JR,
Tadayyon M, Clapham JC, Wilding J, Williams G. Hypothalamic orexin
expression. modulation by blood glucose and feeding. Diabetes 1999;
48: 2132–2137.
39 Burnol AF, Leturque A, Ferre P, Kande J, Girard J. Increased insulin
sensitivity and responsiveness during lactation in rats. Am J Physiol 1986;
251: E537–E541.
40 McMinn JE, Seeley RJ, Wilkinson CW, Havel PJ, Woods SC,
Schwartz MW. NPY-induced overfeeding suppresses hypothalamic
NPY mRNA expression: potential roles of plasma insulin and leptin.
Regul Pept 1998; 75–76: 425–431.
41 Oomura Y, Ooyama H, Sugimori M, Nakamura T, Yamada Y. Glucose
inhibition of the glucose-sensitive neurone in the rat lateral hypothala-
mus. Nature 1974; 247: 284–286.
42 Muroya S, Uramura K, Sakurai T, Takigawa M, Yada T. Lowering
glucose concentrations increases cytosolic Ca(2+) in orexin neurons of the
rat lateral hypothalamus. Neurosci Lett 2001; 309: 165–168.
43 Jang M, Romsos DR. Neuropeptide Y and corticotropin-releasing
hormone concentrations within specific hypothalamic regions of lean but
not ob/ob mice respond to food-deprivation and refeeding. J Nutr 1998;
128: 2520–2525.
44 Chua SC Jr, Leibel RL, Hirsch J. Food deprivation and age modulate
neuropeptide gene expression in the murine hypothalamus and adrenal
gland. Brain Res Mol Brain Res 1991; 9: 95–101.
45 Kalra PS, Dube MG, Xu B, Kalra SP. Increased receptor sensitivity to
neuropeptide Y in the hypothalamus may underlie transient hypophagia
and body weight gain. Reg Peptides 1997; 72: 121–130.
46 Tokunaga K, Matsuzawa Y, Fujioka S, Kobatake T, Keno Y,
Odaka H, Matsuo T, Tarui S. PVN-lesined obese rats maintain
ambulatory activity and its circadian rhythm. Brain Res Bull 1991;
26: 393–396.
47 Sakaguchi T, Bray GA, Eddlestone G. Sympathetic activity following
paraventricular or ventromedial hypothalamic lesions in rats. Brain Res
Bull 1988; 20: 461–465.
48 Shirasaka T, Nakazato M, Matsukura S, Takasaki M, Kannan H.
Sympathetic and cardiovascular actions of orexins in conscious rats.
Am J Physiol 1999; 277: R1780–R1785.
60 Orexin-A and orexin mRNA in hypothalamic lesioned and lactating rats
# 2003 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 15, 51–60