on the role of drug penetration in amethopterin resistance of sarcoma-180 cells in vitro
TRANSCRIPT

198
BBA 45 158
BIOCHIMICA ET BIOPHYSICA ACTA
ON THE ROLE OF DRUG PENETRATION IN AMETHOPTERIN
RESISTANCE OF SARCOMA-ISO CELLS I N V I T R O
M A I R E T. H A K A L A
Department of Experimental Therape~ttics, Roswell Park/VIemorial Institute, Bzt.tJalo, N.Y. (U.S.A.)
(Received A u g u s t 4th, 1964)
SUMMARY
I. The measurement of the net rate of influx of amethopterin was based on the rate of formation of the intracellular folic acid reductase-amethopterin complex. This rate was found to be identical for the sensitive and resistant sublines of Sarcoma- 18o cells cultured in vitro. It was slow (4.6" IO -s cm/sec at 36°) and proportional to the concentration of amethopterin over a IOOO-fold range. The net rates of influx of amethopterin and aminopterin were the same.
2. It was mathematically proven that the degree of resistance to amethopterin would be directly proportional to the content of folic acid reductase in these cells if amethopterin were irreversibly bound by the enzyme. The lack of direct proportion- ality is caused by the intracellular dissociation of the enzyme-amethopterin complex. The final degree of resistance to amethopterin is determined by (a) the cellular folic acid reductase (EC 1.5.1.3) content, (b) the rate of growth, which determines the rate of synthesis of new enzyme, (c) the slow influx of amethopterin and its great affinity to folic acid reductase. These cause the concentration of the intracellular free amethopterin to be only a fraction of the extracellular amethopterin. Consequently, even a small degree of dissociation of the enzyme-amethopterin complex contributes to the degree of resistance to amethopterin. This dissociation becomes especially significant in the 3ooo-fold resistant subline in which folic acid reductase constitutes about 4 % of the total cellular protein. In this subline less than 1% of the total enzyme needs to be functional for normal growth.
INTRODUCTION
Resistance to folic acid analogs in mammalian cells has been shown in several instances to be associated with increased levels of folic acid reductase (EC 1 .5 .1 .3 )
in the resistant cells I 3. In the resistant sublines of S-I8O cells, although the quantity of this enzyme was increased, the Michaelis constant for folic acid and the turnover number per drug-binding site remained unchanged 1. Folic acid reductase "inacti-
Abbrev ia t ions : H T G - m e d i u m , folic acid-free EAGLE'S m e d i u m 8 con ta in ing IO % of horse s e rum and s u p p l e m e n t e d wi th hypoxa l l t h i ne (ioo itM), t h y m i d i n e (3o FYl) and glycine (30 ~tM) : A - H T G - m e d i u m % similar m e d i u m s u p p l e m e n t e d in addi t ion wi th a m e t h o p t e r i n ; S-I8O cells, Sarcoma-I8O cells.
Biochim. I3iophys. ~4cta, lO2 (1965) 198-2o9

D R U G P E N E T R A T I O N AND A M E T H O P T E R I N R E S I S T A N C E 199
va tes" the drug by binding it t ightly; at the same time enough of the enzyme is left free to carry on its normal functions.
The content of folic acid reductase in S-I8O cells in vitro increased with re- sistance. At lower levels of resistance the increase in folic acid reductase content followed closely the degree of resistance. At higher levels of resistance, however, the deviation from direct proportionality became quite pronounced. Thus, cells which were 300o times more resistant than the sensitive parent ceils contained 300-400 times more folic acid reductase than the parent cells 4. This led to the hypothesis that some additional changes, possibly changes in cell permeabili ty might be associated with higher levels of amethopterin resistance. Mechanisms of amethopterin resistance in- volving a decrease in permeabili ty have indeed been observed in mouse leukemic cells in vitro 5 and in vivo 6.
In the present paper an a t tempt has been made to determine the contributions of both the permeabili ty and the enzyme-inhibi tor interaction to the overall effect of amethopterin in growing cultures. No change in permeabili ty of resistant S-I8O cells to amethopterin was observed. Instead, dissociation of the enzyme-amethop te r in complex within the cells was found to play an important role in determining the degree of resistance. I t is this dissociation which causes the deviation from the direct pro- portionality between the degree of resistance and the level of folic acid reductase in the cells. A short communication on this subject has been published:.
M A T E R I A L AND M ETHODS
Origin and maintenance of cells The origin of S-I8O cells, the development of the amethopterin-resistant sublines
and their maintenance mediahave been previously described 4. Folic acid-free EAGLE'S medium s containing IO % of horse serum and supplemented with hypoxanthine (IOO ~M), thymidine (30 ~M) and glycine (30/zM) is designated as HTG-medium. Similar medium supplemented in addition with amethopterin is designated as A-HTG-medium 9.
Preparation of cells for studies of the apparent rate of influx of amethopterin The determinations of the apparent rate of amethopterin influx are based on
the measurement of the rate of appearance in the cells of t ightly bound amethopterin, i.e., amethopterin-folic acid reductase complex. However, when the amethopterin- resistant cell lines are grown in their maintenance media*, i.e., in the presence of amethopterin, maj or portion (78-97 %) of the enzyme is already bound to amethopterin as is seen in Table I. In order to eliminate the enzyme-bound amethopterin, these cells were grown for lO-15 days in amethopterin-free medium (HTG). At the end of this period the amount of enzyme-bound amethopterin in the cells was not detectable.
Determination of the apparent rate of influx of amethopterin Cells prepared as described above and growing as a monolayer in Roux bottles
were used for these studies. At zero time the HTG-medium was replaced with arginine- free (to minimize growth) A-HTG-medium (IOO ml per Roux bottle). The concen- trat ion of amethopterin (o.1-3oo ~M) and tile duration of incubation (15 rain-6 h) at 36° were chosen so tha t at the end of the incubation an appreciable portion of
Biochirn. Biophys. Mcta, lO2 (1965) 198-2o9
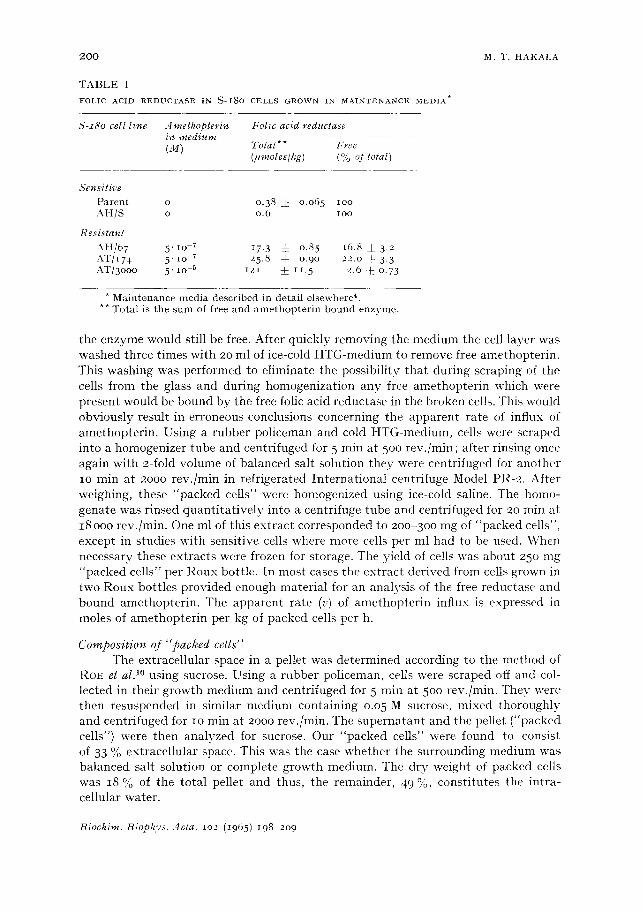
2 0 0 M . T . H A K A L A
T A B L E I
FOLIC ACID REDUCTASE IN S - I 8 o CELLS GROWN IN MAINTENANCE MEDIA ~
S-z8o cell l ine A methoplerin Folic acid reductase in med ium (M) Total** Free
(itmoles/kg) (°/o of total)
Sensit ive P a r e n t o o .38 ~: o .o65 lOO A H / S o 0 .6 IOO
Resis tant
A H / 6 7 5 ' Io 7 17. 3 ~_ 0.85 i6 .8 ~ 3.2 A T / 1 7 4 5 " I O v 25.8 ~ o .9o 22.0 ± 3.3 A T / 3 o o o 5" lO-5 i 2 i ± 1I . 5 2.6 ~_o.73
* M a i n t e n a n c e m e d i a d e s c r i b e d in d e t a i l e l s e w h e r O . ** T o t a l is t h e s u m of f ree a n d a m e t h o p t e r i n - b o u n d e n z y m e .
the enzyme would still be free. After quickly removing the medium the cell layer was washed three times with 2o ml of ice-cold HTG-medium to remove free amethopterin. This washing was performed to eliminate the possibility that during scraping of the cells from the glass and during homogenization any free amethopterin which were present would be bound by the free folic acid reductase in the broken cells. This would obviously result in erroneous conclusions concerning the apparent rate of influx of amethopterin. Using a rubber policeman and cold HTG-medium, cells were scraped into a homogenizer tube and centrifuged for 5 rain at 5oo rev./min; after rinsing once again with 2-fold volume of balanced salt solution they were centrifuged for another IO min at 2ooo rev./min in refrigerated International centrifuge Model PR-2. After weighing, these "packed cells" were homogenized using ice-cold saline. The homo- genate was rinsed quantitatively into a centrifuge tube and centrifuged for 2o min at 18 ooo rev./min. One ml of this extract corresponded to 2oo-3oo mg of "packed cells", except in studies with sensitive cells where more cells per ml had to be used. When necessary these extracts were frozen for storage. The yield of cells was about 25o mg "packed cells" per Roux bottle. In most cases the extract derived from cells grown in two Roux bottles provided enough material for an analysis of the free reductase and bound amethopterin. The apparent rate (v) of amethopterin influx is expressed in moles of amethopterin per kg of packed cells per h.
Composition of"packed cells" The extracellular space in a pellet was determined according to the method of
ROE et al) ° using sucrose. Using a rubber policeman, cells were scraped off and col- lected in their growth medium and centrifuged for 5 min at 5oo rev./min. Thev were then resuspended in similar medium containing o.o5 M sucrose, mixed thoroughly and centrifuged for IO min at 2o0o rev./min. The supernatant and the pellet ("packed ceils") were then analyzed for sucrose. Our "packed cells" were found to consist of 33 % extracellular space. This was the case whether the surrounding medium was balanced salt solution or complete growth medium. The dry weight of packed cells was 18 % of the total pellet and thus, the remainder, 49 %, constitutes the intra- cellular water.
Biochim. Biophys. Acla, loz (1965) 198 209

DRUG PENETRATION AND AMETHOPTERIN RESISTANCE 201
Properties of average single cell All of the permeability measurements in this and the following paper 15 were
made per kg of packed cells. In order to express the data per cell or per unit of cell surface it became necessary to determine the average cell weight in a pellet. Two independent methods were used. (I) Cells in a certain suspension were counted using a haemocytometer. The suspension was then centrifuged for IO min at 20o0 rev./min, and the weight of these "packed cells" was determined. The average weight per cell in such a pellet from several determinations was found to be 7" lO-9 g. This weight includes the weight of the extracellular space, which in such a pellet was shown to be 33 % (see above). (2) The diameter of rounded cells was measured in a microscope. The average from several such measurements was 19 ~ giving a radius (r) equal to 9.5 ~. This corresponds to a cell volume of 359 ° / ~ and a surface area of 1133/z 2. Assuming that the specific gravity is I.O, the weight of one cell is 3.6" lO -9 g. When the 33 % extracellular space in a pellet is added, the weight of one cell in a pellet becomes 5-4" lO-9 g. Since the values arrived at by these two independent methods were in reasonably good agreement, the mean, 6.2.IO -9 g, calculated from the two values was chosen to be used in these studies as the weight per cell in a pellet. The protein content per cell was found to be 4" Io-v mg (ref. II) .
Determination o f free folic acid reductase Suitable dilutions of the cell extracts prepared as described above were used
to determine the free folic acid reductase. This was done by titrating the enzyme with amethopterin as described 1. The basis of the assay is the enzymatic conversion of folic acid to tetrahydrofolic acid which after being spontaneously decomposed to p-aminobenzoylglutamic acid is determined by the BRATTON--MARSHALL reaction. Glucose 6-phosphate and glucose-6-phosphate dehydrogenase (EC 1.1.1.49) were used for regenerating TPNH in the reaction mixture of pH 6.0. The folic acid reductase content in the cells is generally expressed as molar equivalents of amethopterin per kg of "packed cells" or moles per mg of protein in the cell extract.
Determination of bound amethopterin The cell extract was heated for IO min in a boiling-water bath. Under such con-
ditions the enzyme is denatured and the drug originally bound to it is set free. The concentration of amethopterin in this solution was determined as described 12 using folic acid reductase preparation from pooled AT/3ooo cells.
Chemicals Amethopterin (4-amino-Nl°-methylpteroylglutamic acid), supplied as "Metho-
trexate powder" and aminopterin (4-aminopteroylglutamic acid) were products of American Cyanamide Co., Lederle Division.
RESULTS
Amethopterin binding by folic acid reductase I t had been demonstrated by WERKHEISER 13 that folic acid reductase is the
only agent in rat liver which is responsible for the tight binding of 4-amino-analogs of folic acid. It was important to establish whether this is also true for S-I8O cells
Biochim. Biophys. Acta, lO2 (1965) I98-2o9

2 0 2 M . T . H A K A L A
cul tured in vitro. Table I I presents the results of a dialysis experiment performed with an extract of AT/I74 cells. I t shows tha t the amoun t of free reductase which was l iberated by dialysis is equal to the bound amethopter in found in these cells before
TABLE II
F O L I C A C I D R E D U C T A S E A N D A M E T H O P T E R I N B I N D I N G IN R E S I S T A N T S - I S O C E L L S (AT/I74)
Cell extract ,4 methopterin Free folic acid (izmoles / kg) reductase*
(Fonoles/kg) Free Bound
Test Sample**
Before dialysis i.o 25.6 o After dialysis* * * o o 26.o
Control**
After dialysis** * o o 26.o
* Expressed as molar equivalents of amethopterin per kg of packed cells. ** Control and test cells were grown for 16 days in HTG-medium. Test cells were thereafter
grown for 6 days in A-HTG-mediunl (io #M amethopterin). *** Dialysis was performed at 4 ° for 65 h. The volume of the cell extract was about I it11; it
was dialyzed against ioo ml of o.25 M sucrose solution containing 4.36 mM folic acid; solvent was changed once at 45 h.
dialysis. This again was equal to the free enzyme present in the control extract handled similarly. Thus, also in S-I8O cells folic acid reductase is the only measurable agent responsible for the t ight b ind ing of amethopter in .
Content o f ro l ic acid reductase and amethopterin resistance The increase observed in amethopte r in -b ind ing capacity with increase in re-
sistance was the basis for the previous s tudy 1 on the relationship between ametho- pter in resistance and folic acid reductase content of cells. At lower levels of resistance the increase in folic acid reductase content follows closely the degree of resistance. At higher levels of resistance, however, the deviat ion from direct propor t ional i ty is quite pronounced 4. W h y does this deviat ion exist ? One answer could be a decrease in the rate of permeabi l i ty into these highly resis tant cells.
Apparent rate of influx of amethopterin into different sublines of S-z8o cells Studies of the apparen t rate of influx of amethopter in into all sublines of S-i80
cells were performed under slow-growing condit ions (in the absence of arginine) and using short- t ime incubat ions (15 ra in-6 h). The exper imental condit ions were otherwise as described in METHODS. The measurement of the apparent rate of influx was based on the fact tha t folic acid reductase b inds amethopter in so t ight ly 13 and presumably so rapidly tha t all of the drug which enters into a cell is immedia te ly taken up by the enzyme. Thus, the rate of format ion of the e n z y m e - a m e t h o p t e r i n complex is used here as the measure of the apparen t rate of influx, v (moles/h" kg packed cells).
Fig. I shows the dependence of the apparen t init ial rates of influx on the amethopter in concentra t ion in the medium for both resis tant and sensitive sublines
Biochim. Biophys. dcta, lO2 (1965) 198 209

DRUG PENETRATION AND AMETHOPTERI2¢ RESISTANCE 203
TABLE II I
C H A N G E IN T H E C O N S T A N T F O R T H E A P P A R E N T R A T E OF I N F L U X W I T H A M E T H O P T E R I N
C O N C E N T R A T I O N AT 3 6 o
The apparent rate of influx of amethopterin is dependent on amethopterin concentration as seen in Eqn. I, v = K[A0]moles/h'kg of packed cells.
[A 0] K (M) (h -1)
1 o - s o.5i lO -7 0.43 IO -s 0.36 io -s 0.30 io -4 0.25 io -~ o.21
of S-I8O cells. I t r evea l s no c h a n g e in t he r a t e of a p p a r e n t in f lux a s soc ia t ed w i t h res i s tance . I t also shows t h a t t he r e is a s t r a igh t - l i ne r e l a t i onsh ip b e t w e e n t h e l o g a r i t h m
of t h e a p p a r e n t r a t e of inf lux, a n d the l o g a r i t h m of t h e s u r r o u n d i n g d r u g concen- t r a t i o n , [A0] (M). Th i s l ine i nd i ca t e s t h a t for all p r a c t i c a l pu rposes t h e a p p a r e n t r a t e
of in f lux is d i r e c t l y p r o p o r t i o n a l to t h e e x t e r n a l a m e t h o p t e r i n c o n c e n t r a t i o n .
v = K[A0] (moles/h-kg) (I)
T h e c h a n g e in K w i t h [A0] is so s l ight (Table I I I) t h a t in i t i a l ly i t was o v e r l o o k e d 7 ,14.
O n l y e x t e n s i v e s tudies , a l t o g e t h e r n e a r l y one h u n d r e d d e t e r m i n a t i o n s , f ina l ly con-
f i r m e d a s l igh t c u r v a t u r e on l inear scale. W h i l e a m e t h o p t e r i n c o n c e n t r a t i o n is in- c r ea sed i o ooo- fo ld t h e r a t e c o n s t a n t decreases on ly b y 50 %. Th i s s l ight d e v i a t i o n f rom
d i r ec t p r o p o r t i o n a l i t y m i g h t i nd i ca t e f ac i l i t a t ed dif fus ion w i t h v e r y low af f in i ty c o n s t a n t . I t is m o r e l ike ly , h o w e v e r , t h a t i nc reas ing a m e t h o p t e r i n c o n c e n t r a t i o n in
I 0 0
=
(a
"" I 0
o=
O I 10-7 10-6 10-5 10-4
A M E T H O P T E R I N ( M )
x ~ 1 0 o o ¢~.
o
i i i
0 1 2 3 4 - A M I N O - A N A L O G (M x I 0 ' )
Fig. I. Dependence of the apparent initial rate of influx (v) of amethopterin into the sensitive parent S-180 cells and three resistant sublines on amethopterin concentration in the medium at 36 °. Numbers next to certain points indicate the number of individual determinations at that concen- tration of amethopterin: the point in such a case represents the mean of the individual values.
Fig. 2. Comparison of the rates of apparent initial influx (v) of amethopterin (0) , and aminopterin (E3) into AT/I74 cells at 36°.
Biochim. Biophys. Acta, lO2 (1965) 198-2o9

204 M.T. HAKALA
the presence of a cons tan t level of Ca °~+ causes an a p p a r e n t decrease in the ra te "con- s t a n t " K (ref. 15). Since most of these s tudies were per formed at concent ra t ions of ame thop t e r i n a round lO .5 M, K will be considered to be a cons tan t equal to o.3o h -L This is in fact , the mean value from all the ind iv idua l K values. The mean ing of K in t e rms of the ind iv idua l ra te cons tan t s is discussed in de ta i l in the following paper ~5.
The magn i tude of v at 360 reveals the ve ry slow na tu re of ame thop te r in influx (Fig. I) . This a p p a r e n t ra te of influx can be expressed in pe rmeab i l i t y units , P (moles per t~ 2. sec), for a concen t ra t ion difference of I M and resul ts in:
P - K × 15.3-to-19
= 4.6. IO -19 moles/#2.sec at 36 ° (2)
I t should be no ted t ha t a concen t ra t ion difference of I M in case of a me thop t e r i n is an abs t r ac t one and b e y o n d prac t ica l feasibi l i ty. However , i t can be concluded on the basis of the d a t a in Table I I I t ha t a t t ha t pa r t i cu l a r concen t ra t ion P would stil l be smaller , a p p a r e n t l y 1. 9 • lO -19 moles/ / ,"-see at 36° . W h e n the a p p a r e n t ra te of influx of ame thop t e r i n is expressed in the uni ts of veloci ty 16 one gets 4.6" IO -8 cm/sec at 36°.
The apparent rate of influx of aminopterin I t was of in teres t to compare the a p p a r e n t ra te of influx of aminop te r in and
ame thop te r in . Fig. 2 presents the resul ts of this s t u d y and indica tes t h a t the a p p a r e n t ra te of influx is the same for the two compounds .
On the stability of enzvme-amethopterin complex in growing cultures If ame thop t e r i n were i r revers ib ly bound b y folic acid reductase then the pro-
por t ion of the t o t a l enzyme bound b y amethop te r in , [EA!/ [Et ] (EA, e n z y m e - a m e - t hop te r in complex ; Et, t o t a l enzyme), in growing cul ture when a s t e ady s ta te is reached could be easi ly predic ted . This ra t io would depend ent i re ly on the ra te of influx of ame thop t e r i n (KIA0]), the folic acid reductase conten t of the cell line (c) and the cell genera t ion t ime (~). The expression for d [EA! /d~Et ] when t approaches inf ini ty was der ived as follows:
Tota l folic acid reductase in a growing cul ture increases logar i thmica l ly as does the cell mass (m). The ra te of this increase depends on the folic acid reductase con ten t of the cell line as well as on the genera t ion t ime.
diet] ln2 -- cm - - (moles/h)
dt z [gt] (moles); c (moles/kg); m (kg); r (h) (3)
The ra te of increase in the to ta l amoun t of drug which has pe ne t r a t e d ( identical wi th b o u n d drug, EA) is p ropor t iona l to the appa ren t ra te of influx (K[A0]) and the cell mass.
d[EA~ -- / t ~ EAo] m (moles/h)
dl EEA] (moles); K EAo] (moles/h.kg); m (kg) (4)
Bo th Eqns. 3 and 4 are i n t eg ra t ed to get the amoun t of to ta l and bound enzyme as t approaches infini ty. The ra t io of these two as t approaches inf ini ty becomes:
Biochim. Biophys. Acta, io2 (1965) 198-2o9

DRUG PENETRATION AND AMETHOPTERIN RESISTANCE 205
d[EA) ~ [EA] _ K [Ao]
-d[Et]-], -. oo [E , ] c ln2 (5)
This equation indicates that the proportion of the total enzyme which is bound by the drug at a steady state increases with the rate of influx of amethopterin and with the generation time but decreases with increasing enzyme content. Indeed, if ametho- pterin were bound irreversibly by folic acid reductase the degree of resistance would be directly proportional to the enzyme content of the cell line when the permeability and the growth rate are unchanged as they are in the present case.
Eqn. 5 offers the means of testing whether in the live and multiplying S-I8O cells the drug is indeed so tightly bound by the enzyme that this binding would appear irreversiblel By growing ceils in A-HTG-medium one can relate the free and amethopterin-bound folic acid reductase with the amethopterin concentraton in the medium under such conditions that the function of this enzyme is not necessary for the survival of the cells 9. Therefore, growth is not limited and the rate of growth is normal. When cells are grown for a sufficiently long period (7 days or longer) in such a medium the proportion of the total enzyme bound by the drug was found to stay constant and to depend on the concentration of amethopterin in the medium. We have designated this steady state as "dynamic equilibrium". These cultures were then rinsed, collected and cell extracts were prepared as described for influx rate studies in METHODS. Under these experimental conditions a 6-I2-fold increase in cell mass occurs in I week. This gives a mean generation time in the range of 45 to 60 h. It is likely, however, that a rapid rate of growth (z < 45 h) occurs early during the 7-day growth period, while at the time of sampling the rate of growth has reached a plateau (z > 60 h). Therefore, two T values were chosen, 50 and 30o h. In Fig. 3 the two theo- retical curves (dotted lines) based on Eqn. 5 are compared with the experimental data for cell line AT/3ooo. No likeness can be detected between the data and the two curves. In fact, the experimental points seem to fall on a line which resembles a dissociation curve quite closely. This relationship was studied in the other cell lines, AH/67 and AT/I74 , also, and the observations were consistent. Thus, the binding of amethopterin by folic acid reductase in S-I8O cells does not appear to be irreversible.
Dynamic equilibrium Let us define
[Ao] [E] KDy~ -- (M) (6)
lEA]
[E] = free enzyme concentration (moles/kg) ; [EA] = concentration of enzyme- ame- thopterin complex (moles/kg) ; [Ao] ---- amethopterin concentration (M) in the medium.
When the value of KDy~ is calculated for each experimental point in Fig. 3, the mean is 8.16 • Io -~ M. This means that when AT/3ooo ceils are growing in the presence of o.816/~M amethopterin only one-half of all of the reductase in these cells is bound by amethopterin. The solid line in Fig. 3 is drawn for this value of KDy~. The cor- responding values of KDy~ for the other resistant cell lines are listed in Table IV.
It is interesting to observe Fig. 3 from the point of view of growth rate. It is logical to assume that in the case of irreversible binding a lower concentration of amethopterin would be necessary for a certain fraction of total enzyme to appear as enzyme-amethopterin complex than would be if the enzyme-amethopterin complex
Biochim. Biophys. Acta, lO2 (1965) 198-2o9

200 M.T. HAKALA
dissociates. Consequently, one finds that at the t ime when the ceils were harvested for the determinat ion of K D y ~ these cells must have been growing very s lowly indeed.
w 1 0 0 / ~ . -
/ "r = 3 0 0 h
" =50h ® 5 0 /
o
. . . . . n - - i 10-8 10-7 10-6 10-5 10-4
M AMETHOPTERIN IN MEDIUM (Ao)
Fig. 3. The fraction of intracellular folic acid reductase bound by amethopterin in growing cultures at a steady state. AT/3ooo cells were grown for 7 days or longer at 3 6o at the indicated concen- trations of amethopterin (A-HTG-medium). At low concentrations of the drug ( ~ I /*M) the cells used as inoculum were free of bound amethopterin. The solid line is drawn on the basis of the mean (8.16. IO ~ M) of individual KnyN-values (Eqn. 6) based on each experimental point. The dotted lines are drawn assuming irreversible binding on the basis of Eqn. 5 (K = o.3o h- l ; c = 1.2- io -4 moles/kg) for two T values as indicated.
TABLE IV
D Y N A M I C E Q U I L I B R I U M ( / ~ ' D Y N ) F O R D I F F E R E N T S U B L I N E S O F S - I 8 O C E L L S
S-z8o cell line KDyN
Experimental* Calculated** EAo]/[A,~*** ( M × r o 7) ( M ~ z o 9)
Resistant AT/3ooo AT/174 AH/67
Sensitive AH/S Parent
8.16 81 600 1.36 13 600 1.o 4 IO 400
3.58 358 2.26 226
* This is the mean calculated from experimental data according to Eqn. 6. ** Calculated on the basis of direct proportionality between the folic acid reductase content
(Table I) and the experimental KD~N-values. *** Calculated on the basis of Eqn. 8, assuming KDiss -- io 11.
W h e n t h e t h r e e cu rve s in Fig. 3 are c o m p a r e d it b e c o m e s c lear t h a t t h e dis-
soc i a t ion of e n z y m e - a m e t h o p t e r i n c o m p l e x , w h e n t h e m a j o r i t y of t h e e n z y m e is
b o u n d , b e c o m e s so s igni f icant , t h a t v e r y h igh c o n c e n t r a t i o n s of t h e d r u g are n e c e s s a r y
for s a t u r a t i o n of t h e e n z y m e a n d t heo re t i c a l l y , t o t a l s a t u r a t i o n can n e v e r be ach i eved .
I f one were dea l ing w i t h an e q u i l i b r i u m in t he usua l sense of t h e word , one were
to f ind j u s t one va lue of KDYN for all cell l ines. Th i s w o u l d be d e t e r m i n e d solely b y
p e r m e a b i l i t y a n d b y d i s soc i a t i on of t h e e n z y m e - a m e t h o p t e r i n c o m p l e x . H o w e v e r , in
t h e p r e s e n t case /£DyN r e p r e s e n t s a d y n a m i c s t e a d y s t a t e w h i c h i n v o l v e s n o t on ly
d i s soc i a t i on b u t also c o n s t a n t g r o w t h a n d c o n s t a n t s y n t h e s i s of n e w free e n z v m e .
Th i s will t e n d to m a k e K D y ~ a p p e a r l a rge r t h a n i t w o u l d be in r e s t i n g cells a n d i t a lso
Biochim. Biophys..4cla, io2 (1965) 198-2o9

DRUG PENETRATION AND AMETHOPTERIN RESISTANCE 207
resul ts in different/~DYI~I values for cell l ines wi th different enzyme con ten t (Table IV). A c t u a l l y one finds t h a t KDYN and the enzyme con ten t are d i rec t ly propor t iona l . This allows the ca lcula t ion of a p p r o x i m a t e KDYN values for the sensi t ive cell lines (Table IV). The de t e rmina t i on of the l a t t e r values would be expe r imen ta l ly unfeasible.
The dissocia t ion cons tan t of the e n z y m e - a m e t h o p t e r i n complex inside the cell is:
[Ai~ [EJ KDISS -- (7)
[EA]
When Eqns. 6 and 7 are combined one finds that:
[A0~ __ KDyN (8) [Ai~ KDISS
Studies with rat liver enzyme in vitro have indicated that KDiss for folic acid re- ductase-amethopterin complex at pH 7-5 is about io - ' I (ref. 17). When one sub- stitutes this KDISS value and the KDYN values from Table IV into Eqn. 8, one finds that the intracellular concentration of amethopterin is only a fraction of the extra- cellular concentration. KDy~ is not a true dissociation constant, but is determined by such factors as the rate of growth and by the enzyme content of the cells. Therefore, [A0]/[AI ] is not directly related to efflux and influx rates. The one definite conclusion that can be drawn from these data is, that the actual rate of amethopterin influx must be very slow when compared with the rate of the enzyme- amethopterin complex formation and/or the rate of efflux of amethopterin.
DISCUSSION
The ve ry t igh t b ind ing of ame thop t e r i n to folic acid reductase13,17 forms the basis for the m e t h o d o l o g y app l ied here to s t u d y the pe rmeab i l i t y of m a m m a l i a n cells to ame thop te r in . The measu remen t of the a p p a r e n t ra te of influx of a me thop t e r i n was based on the ra te of fo rma t ion of the e n z y m e - a m e t h o p t e r i n complex wi th in the cells. Using this me thodo logy i t was es tab l i shed t h a t no change in pe rmeab i l i t y occurred when cells became res i s tan t to ame thop te r in . In o ther ins tances involv ing resis tance to th is drug in mouse leukemic cells in vitro, a decrease in pe rme a b i l i t y to the in- h ib i to r was i nd i ca t ed as a mechan i sm of res is tance 5. Var ied pe rmeab i l i t y has also been sugges ted as the basis for different ia l effects of 4-aminofola te an tagon is t s i n vivo6,1s.
In the presen t s t u d y the a p p a r e n t ra te of influx was found to be p ropor t iona l to the concen t ra t ion of ame thop t e r i n over a thousand- fo ld range. W h e n conver ted into pe rmeab i l i t y uni ts one finds t h a t P ~ 4.6. lO -19 moles/tzz, sec for a concent ra t ion difference of I M a t 36°. The weight per cell, inc luding 33 % of ex t race l lu la r space, in a pel let of " p a c k e d " S-I8O cells, which was app l ied here, was 6.2- lO -9 g. This f igure is the average of two va lues (5.3" IO 9 and 7.0. lO -9 g) a r r ived at using two independen t me thods of measu remen t as descr ibed in METHODS. I t agrees well wi th the resul t of REGELSON* who found 2"lO s cells in I ml of packed ascites S-I80 cells. However , a large d i sc repancy is no ted be tween these values and t h a t r epor ted b y SCHRECKER et al. 19 s t a t ing t h a t I ml of S-I80 cells conta ins (1-2). 109 cells. The l a t t e r i nves t iga to r s
* Dr. W. I{EGELSON Of this Institute, personal communication.
Biochim. Biophys. Acta, lo2 (1965) 198-2o9

208 Vi. T. HAKALA
based the i r e s t ima te on the measu remen t of an average cell d i ame te r in a publ i shed photomicrograph2°, ". In general , the a p p a r e n t ra te of influx of a me thop t e r i n is ve ry slow when compared wi th the ra te of inf lux of o ther an t imetabol i t es . F o r example , azaser ine is t aken up abou t 3oo t imes fas ter b y the p l a sma cell neoplasm of mouse 21. To i l lus t ra te the slowness of ame thop t e r i n influx one can es t imate t ha t a t I ~M concen t ra t ion of the drug i t would require I h and 16 min to s a tu ra t e the enzyme in the sensi t ive pa ren t S-I8O cells, 2 h in AH/S , 58 h in AH/67, 86 h in AT/ I74 and over 4oo h in AT/3ooo. Moreover, this would be t rue only if the cells were cons t an t ly exposed to this concen t ra t ion (I/~M) of the drug and ne i ther g rowth nor dissociat ion of the complex would occur. If the ra te of pe rmeab i l i t y of o ther m a m m a l i a n cells to ame thop t e r i n were of the same order of magn i tude as of S-I8O cells one could pred ic t t ha t i t would be difficult to s a tu ra t e the enzyme even of sensi t ive cells in vivo. Under condi t ions in vivo a I /~M concen t ra t ion of drug is ha rd to ma in t a in for any length of t ime.
The most i m p o r t a n t po in t revealed by this s t u d y is the fact t ha t the dissociat ion of the e n z y m e - a m e t h o p t e r i n complex, however small , is significant in de te rmin ing the degree of resis tance. I t is this dissociat ion which is the cause of the lack of d i rec t p ropo r t i ona l i t y be tween the level of enzyme in the cells and the degree of resis tance. The dev ia t ion from direct p ropor t i ona l i t y becomes especial ly s t r ik ing in cell lines such as AT/3ooo which conta in large amoun t s of folic acid reductase (4 % of t o t a l cell protein) and consequent ly have the c a p a c i t y to form more of the e n z y m e - a m e t h o - p te r in complex. There is an a p p a r e n t con t rad ic t ion be tween these f indings and those on dissociat ion of the e n z y m e - a m e t h o p t e r i n complex in vitro 17. The present s t u d y (KDYN) shows t ha t the ex t race l lu la r concent ra t ions of ame thop te r in requi red for half s a tu ra t i on of in t race l lu la r folic acid reductase are much higher than requi red in an enzyme assay. This indica tes e i ther t ha t this dissociat ion cons tan t does not app ly in t race l lu la r ly and /o r t h a t the p e r m e a b i l i t y is ve ry much in favor of drug efflux. I t will be shown 15 t ha t the ra te cons tan t for efflux is indeed larger t han t ha t for influx. The na tu re of the pe rmeab i l i t y process of a m e t h o p t e r i n as a whole will be descr ibed in more de ta i l in the following pape r ~5.
A C K N O W L E D G E M E N T S
The au tho r wishes to express her apprec ia t ion to Dr. C. A. NICHOL for his in te res t and suppor t and to Miss E. TAYLOR, Miss L. PUCCETTI and Miss D. SUGG for the i r skillful technica l assistance. This inves t iga t ion was suppor t ed in pa r t b y research g ran t CA-o 4175 from the Na t iona l Cancer I n s t i t u t e of the U.S. Publ ic Hea l th Service.
REFERENCES
i M. T. HAKALA, S. F. ZAKRZEWSKI AND C. A. NICHOL, J. Biol. Chem., 236 (1961) 952. 2 G. A. FISCrtER, Biochem. Pharmacol,, 7 (1962) 75. 3 D. K. MISRA, S. R. HUMPHREYS, M. FRIEDKIN, A. GOLDIN AND E. J, CRAWFORD, Nature,
189 (1961) 39- 4 M. T. HAKALA AND T. ISHIHARA, Cancer Res., 22 (1962) 987 . 5 G. A. FISCHER, Biochem. Pharmacol., I1 (1962) 1233. 6 ~T. C. WERNHEISER, Cancer Res., 23 (1963) 1277. 7 M. T. HAKALA, Federation Proc., 21 (1962) 236.
* Dr. D. HUTCHISON, Sloan Kettering Institute, private connnunication.
Biochim. Biophys. Acta, ioz (1965) 198-2o9

DRUG PENETRATION AND AMETHOPTERIN RESISTANCE 209
8 H. EAGLE, Science, 13o (1959) 432. 9 M. T. HAKALA AND E. TAYLOR, f . Biol. Chem., 234 (1959) 126.
IO J. H. ROE, J. H. EPSTEIN AND N. P. A. GOLDSTEIN, J. Biol. Chem., 178 (1949) 839. i i V. I. OYAMA AND H. EAGLE, Proc. Soc. Exptl. Biol. Med., 91 (1956) 305 . 12 W. C. WERKHEISER, S. F. ZAKRZEWSKI AND C. A. NICHOL, J. Pharmacol. Exptl . Therap.,
137 (1962) 162. 13 W. C. WERKHEISER, J. Biol. Chem., 236 (1961) 888. 14 M. T. HAKALA, Federation Proc., 22 (1963) 183. 15 M. T. HAKALA, Biochim. Biophys. Acta, lO2 (1965) 21o. 16 H. DAVSON, The Textbook of General Physiology, Lit t le , B rown and Co., Boston, 196o, p. 224. 17 D.-H. WANG AND W. C. WERKHEISER, Federation Proc., 23 (1964) 324 . 18 W. C. WERKHEISER, L. W. LAW, R. A. ROOSA AND C. A. •ICHOL, Proc. Am. Assoc. Cancer Res.,
4 (1963) 282. 19 A. W. SCHRECKER, J. M. VENDITTI, N. H. GREENBERG, J. L. BRIDLER, D. L. ROBINSON AND
D. J. HUTCHISON, J. Natl. Cancer Inst., 31 (1963) 557- 20 G. E. FOLEY AND D. P. DROLET, Proc. Soc. Exptl. Biol. Med., 92 (1956) 347. 21 E. P. ANDERSON AND J. A. JACQUEZ, Cancer Res., 22 (1962) 27.
Biochim. Biophys. Acta, lO2 (1965) 198-2o9