nucleotide ribonucleic acid metabolism … & ribonucleic acid metabolism of corn seedlings'...
TRANSCRIPT

NUCLEOTIDE & RIBONUCLEIC ACID METABOLISM OF CORN SEEDLINGS'JOE H. CHERRY & R. H. HAGEMAN
DEPARTMENT OF AGRONOMY, UNIVERSITY OF ILLINOIS. URBAN.A
The consideration of nucleotides and ribonucleicacid (RNA)2 in metabolism has become increasinglyimiiportant in cellular biology during the past fewyears. The commonly occurring nucleotides of-wheat and corn plants have been identified by Bergk-N-ist (1) and Cherry and Hageman (3), respectively.The latter have shown that the nucleoside di- andtrip)hosphates in etiolated corn embryos shift to thentcleoside monophosphates as a function of growth.
Little is known about the metabolism of nucleicacid during seed germination. Oota et al (11, 12)have examined nucleic acid metabolism in germinatingsee(ls, and have found large amounts of RNA storedin the cotyledons of a bean. Germination resulted ina steady decrease in the RNA content of the coty-ledlons with an increased RNA content in the seed-ling. The presence of the embryo was essential forbreakdown of RNA in the cotyledons. If embrvosw-ere detached from the cotyledons, no RNA degrada-tioni occurred (13).
Recently, Cherry et al (4) have shown that RNAis synthesized in etiolated corn embryonic axis butdegraded in the scutellum with both reactions beinggreater in hybrid plants than in their inbred parents.
Oota and Takota (12) have proposed that therear-e at least two types of RNA-s i.e. functional andtransportable, in bean germ tissues. Their resultsappear to illustrate conversion steps from functionalRNA to transportable RNA.
Hanson (6) indicates that RNA in membranes isinlplicated in ion accumulation, solute retention, andoxidative phosphorylation.
The experiments reported here represent stu(diesoni the metabolism of nucleotides, RNA, an(d proteinin v-arious cytoplasmic fractions of corn seedlings.Ilcorporation of P32 into various soluble nucleotidesanl(l cytoplasmic RNA--s, an(l ADP-C814 uptake byexcised tissue were studied.
I Received August 5, 1960.2 The following abbreviations will be used: AMP,
adeniosine monophosphate; ADP, adenosine diphosphate;ATP, adenosine triphosphate; UMP, uridine monophos-phate; UDP, uridine diphosphate; UTP, uridine triphos-phate; GTP, guanosine triphosphate; CMP, cytosinemonophosphate; CTP, cytosine triphosphate; DPN, di-phosphopyridine nucleotide; TCA, trichloroacetic acid;RNase, ribonuclease; anid RNA, ribonuicleic acid.
1IATERIALS & METHODS
PLANT MATERIAL: Hybrid corn seed3 (Zea wRaysL. var. WF9XM14) were lightly dusted with a fungi-cide (Spergon, U. S. Rubber Co.) and approximately100 were placed embryo down on paper towellingsupported by a layer of gravel (400 ml) in a 2 quartpyrex utility dish. The gravel and towelling weresaturated with 200 ml of deionized water containing0.5 mc of P32 (5.2 /g) as potassium phosplhate. Theseeds were germinated in darkness in a humid atmos-phere at 290 C for 1, 2, 3, or 4 days. Additional de-ionized water was added to the dish during the germi-nation period to keep the gravel saturated. Seedlingswere harvested and washed several times with deion-ized water to remove excess P3 before use.
Plant material for the ADP-C,14 stu(lies was ob-tained by the above procedure except that the seedswere moistened with 10-4 A, CaCl2 insteadl of theradioactive phosphate solution.
ISOLATION OF CYTOPLASM'IIC PARTICULATES: Theplant tissue (about one gin) was homogenized for 3minutes in 5 ml of deionize(d vater with an ice-jacket-ed glass homogenizer with a power-driven Teflonpestle. The homogenate was cleared of cellulardebris by centrifugation at .500 g for 5 minutes. Analiquot of the debris-free homogenate was subjectedto differential centrifugation in a Spinco model Lultracentrifuge, each successive particulate fractionbeing obtained from the supernate of the precedingfraction. The centrifugal forces, time, and thearbitrary designation of the isolatedl fractions wereas follows: mitochondria, 22.000 g for 15 minutes:microsomes, 110,000 g for 1 hour; cytoplasmic sap,particulate-free supernate. The terms used to desig-nate the cytoplasmic fractions are operational, as listedabove, and do not necessarily mean the presence ofpure morphological entities.
An isotonic grinding mledlium (0.5 M sucrose) wasalso used in the isolation of particulate fractions.Analysis of the particulates obtained from water and0.5 M sucrose extractions were comparable; therefore,only the results from water extraction will be reported.
3 We are indebted to Dr. W. J. Mumm, Crow's HybridCorn Co., Milford, Ill., for the corn seed used in this work.
163
www.plantphysiol.orgon July 6, 2018 - Published by Downloaded from Copyright © 1961 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
AN'ALYSIS OF CYTOPLASMNIC FRACTIONS: Thetotal acid-soluble nucleotides and RNA w!ere extractedand( measured by a method (5) similar to that of Oguran(l Rosen (10). Aliquots of each of the watersuspended particulate fractions (mlitochondlrial &micr-osomal) and the cytoplasmic fraction, and debris-free homogenate were acidified with 1.2 at HCIO.to a final molarity, of 0.2 whlliclIprecipitated the RNA.The RNA precipitates were isolated by centrifuga-tion an(l washed twice wvith 0.2 M HCIO. The en-tire extraction procedure for the acid soluble nuicleo-ti(les was perfornied at 20 C. The supernatalnt solu-tion and the two washes were combined, made toVolume, and the soluble nucleoticdes estimated bv thedifference of absorbanicv at 260 alndl 290 nii usingAMP as a standard.
The precipitated RNA pellets wvere extracted thricewith 2: 2: 1 ethanol-ether-chloroform to removelipids. RNA was hydrolyzed by incubating the lipid-free residue with 0.5 Af HCIO, at 70° C for 40millnltes. Absorbancv differelnces at 260 and 290 mynof the cleared supernatant solutionl were referred toa standard curve obtained frolmi similarly treatedyeast RNA.
Protein was determinied by the method of Lowryet al (8) on ice-cold 10 % TCA pr-ecipitates of eachof the c ytoplasmic fractions.
DETER-MINATION OF RADIOACTIVITY: Aliquots ofthe acid-soluble nucleotides an(l of the hydrolyzedRNA were neutralize(d with KOH anld the KCIO1was remove(d bv centrifugation. Each of thle nucleo-tidle and hydrolyzed RNA samples was dried and theradioactivity counted.
ADP-Cj14 UPTAKE: Root tips. 1 cim lonig, w\eresectionicd friom primary anld seconldary roots, apicalmesocotyl sections (1 cim), aind scutella were removedfroml 3-day-old corn seedlings. The tissue was rinsedin deionized water, blotted, and(1 rapi(lly weighed tothe nearest milligramii before use. Tissue sampleswere placed in petri (lislhes andl incubated in the lightat roonim temnperature with 30 ml of a solution contain-ing 1 %O sucrose, 10-3 -\r KH..PO1, and 1 uc ofADP-C514 (0.8 mg) adjusted to pH 6.3 withNH OH. After 8 hlours the tissue was removed,rinised sev-eral times with deioniized water, weighed,an(l frozen.
Samples of the fresh tissue were frozen for initialnucleotide and( RNA (letermiinations.
NUCLEOTIDE EXTRACTION & TON-EXCHANGE CHRO-\ATOGRAPHY: Scutellum and radicle tissues wereremoved from the etiolated corn seedlings (3 daysol(l) and the acid-soluble nucleotides extracted fromeach tissue witlh ice-cold 0.6 M\ HClO,. The pro-cedlures for extraction and column chromatography ofthe nucleotides were the same as previously described(3), except that Dowex-1 X 8 was used instead ofDowex-1 X 10. The eluate was collected in 10-mlvolumes. The nucleotide content was determined bythe absorption of light at 260 iinA and 1-ml aliquots
from each fractioin were (Iriedl alid counte(l for radio-activity.
PAPER CO-CIIROrMATOGRAPIIY OF ORGANIC PHOS-PHATES & ORTHOPIIOSPHATE: Samples obtained byconcentrating the eluate of peak fractions from columnlchromatography were co-chromiatograplhed on paperwith standard phosphate compounlds in (lifferenitsolvents. The solvents mnost usedl for chromatographywere n-propanol: NH,OH: H2O (60: 30: 10) andlisobutvric acid: NH1OH: FfLO (57: 4: 39). .Afterthe paper chromatograms were developedl and dIrie(l.phosphorus-colntaininig compoundls w-ere detecte(d withla molybdic acid spray reagent (2). Tn some instancesin wlhichl the unknown radioactive samiiples were toodilute to react with molvbdic acid, the paper was cut-into 2-cm strips and these were in turnl cut into 2-cimisquares. Each of the squar-es was counited for radio-activity, thereby establislhing the location of the tin-knownvn on the palper.
All results reporte(l in this paper are base(l ontwo or more separate experimiielnts. Each assay wvasmia(le in d*uplicate.
RiESULTS
CHANGES DuRING. SEED GERTUINATIoN: Typical'dry weighlt changes of corn scutellumii andcl radicle aregiven in table T. The scutelluml (lecrease(l in (dr-weight about 18 % wvhereas the radicle increase(l illdry weight 42-fold in the 3-day' per-iod (1-4 days).
As corn seed germiniate there is a l)rodllction ofnucleoti(les in both scutellum andl ra(licle tissues (tableII). An interestinig facet of RNA miietabolism (ludrin¢gcorn see(l germination is that 57 %7, of the sctutellumiiiRNA wvas (legraded by- the 4th (lay whereas radicleRNA increase(l 182 %. Grow-vth (drv Nvt0 of the-radicle occurredl at the samce rate as nitcleoti(le syn-thesis, but RNA wvas synthesized at a mtuchi slowerrate.
Protein nmetabolism was sinmilar to RNA imietabo-lism in the scutellum an(l radicle tissues: hoNwever,the RNA to protein ratio wvas not constant lor eithertissue. The RNA to protein ratios (g/'mg) in thescutella were 67, 79, 57, an(l 51 from 1 to 4 days, in-clusivelv, whereas the ratios in the ra(licles were 160,143, 79 and 96.
Changes in protein content in the (lesignatedl frac-tions of radicle an(l scutellumll are represented in
TABLE IDRY WEIGHT CHANGES OF CORN SCUTELLUMI & RADICLE
TIssuE WITH GROWTH
AGE,DAYS
1234
DRY WT/SCUTELLUM DRY WT/RADICLEmg mg
24.5 0.623.1 6.821.0 i8.520.1 26.1
164
www.plantphysiol.orgon July 6, 2018 - Published by Downloaded from Copyright © 1961 American Society of Plant Biologists. All rights reserved.
CHERRY & HAGEMAN-NUCLEOTIDE & NUCLEIC ACID METABOLISM 165
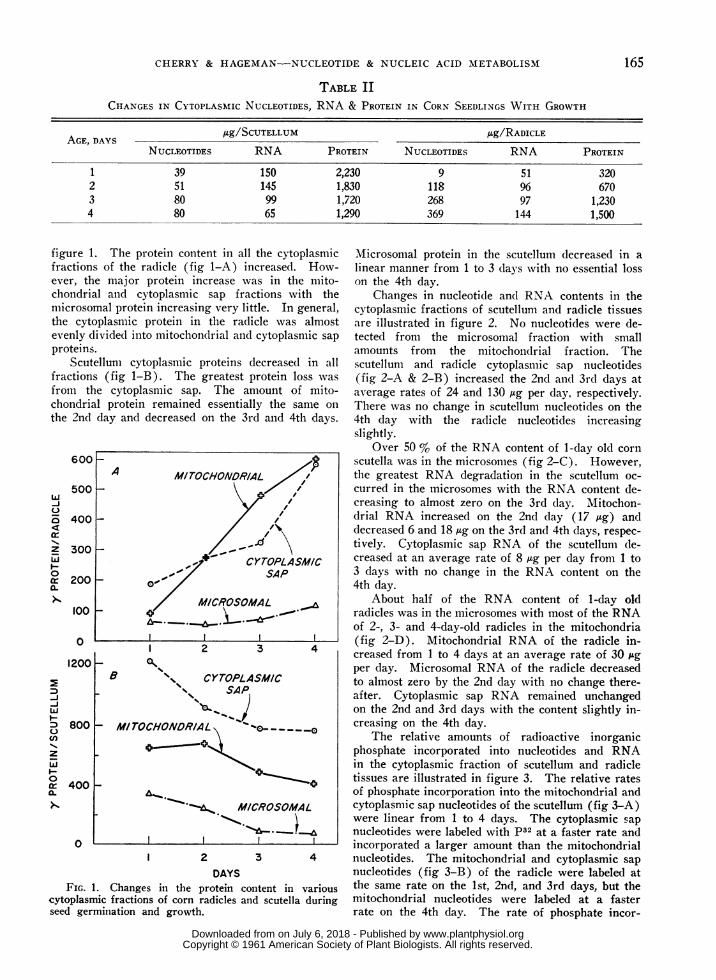
TABLE IICHANGES IN CYTOPLASMIC NUCLEOTIDES, RNA & PROTEIN IN CORN SEEDLINGS WITH GROWTH
ADg/SCUTELLUM Ag/RADICLEAGE, DAYSNUCLEOTIDES RNA PROTEIN NUCLEOTIDES RNA PROTEIN
1 39 150 2,230 9 51 320234
figure 1fractionever, tichondrinmicrosothe cytevenly cproteins
Scutfractionfrom tlchondri;the 2nd
FIG.cytoplasrseed ger
518080
1459965
1,8301,7201,290
118268369
9697144
6701,2301,500
1. The protein content in all the cytoplasmic Microsomal protein in the scutellum decreased in aIs of the radicle (fig 1-A) increased. How- linear manner from 1 to 3 days with no essential lossie major protein increase was in the mito- on the 4th day.al and cytoplasmic sap fractions with the Changes in nucleotide and RNA contents in themal protein increasing very little. In general, cytoplasmic fractions of scutellum and radicle tissuesoplasmic protein in the radicle was almost are illustrated in figure 2. No nucleotides were de-livided into mitochoncdrial and cytoplasmic sap tected from the microsomal fraction with smallS. amounts from the mitochondrial fraction. Thetellum cytoplasmic proteins decreased in all scutellum and radicle cytoplasniic sap nucleotidesIs (fig 1-B). The greatest protein loss was (fig 2-A & 2-B) increased the 2nd and 3rd days athe cytoplasmic sap. The amount of mito- average rates of 24 and 130 /Ag per day, respectively.al protein remained essentially the same on There was no change in scutellum nucleotides on theday and decreased on the 3rd and 4th days. 4th day with the radicle nucleotides increasing
slightly.Over 50 % of the RNA content of 1-day old corn
scutella was in the microsomes (fig 2-C). However,A MITOCHONDR/AL / the greatest RNA degradation in the scutellum oc-
curred in the microsomes with the RNA content de-creasing to almost zero on the 3rd day. Mitochon-
_//,drial RNA increased on the 2nd day (17 ,ug) and//X \decreased 6 and 18 Ag on the 3rd and 4th days, respec-
_.d/-~\tively. Cytoplasmic sap RNA of the scutellum de-
+~ CYTOPLASM/C creased at an average rate of 8 Ag per day from 1to
SAP 3 days with no change in the RNA content on the- OS 4th day.
M/CROSOMAL About half of the RNA content of 1-day old_,:,\._ radicles was in the microsomes with most of the RNA
of 2-, 3- and 4-day-old radicles in the mitochondrial2 4
(fig 2-D). Mitochondrial RNA of the radicle in-42 creased from 1 to 4 days at an average rate of 30 ,Ag
l- A^ per day. Microsomal RNA of the radicle decreasedB \ CYTOPLASM/C to almost zero by the 2nd day with no change there-
ssSAPj after. Cytoplasmic sap RNA remained unchangedon the 2nd and 3rd days with the content slightly in-
-IMTOCHONOR/Af;L ^->____-o creasing on the 4th day.MITOCHONDRIA- - - - - -0 The relative amounts of radioactive inorganic
'' <phosphate incorporated into nucleotides and RNAin the cytoplasmic fraction of scutellum and radicle
_4< tissues are illustrated in figure 3. The relative ratesof phosphate incorporation into the mitochondrial and
MICROSOMAL cytoplasmic sap nucleotides of the scutellum (fig 3-A)were linear from 1 to 4 days. The cytoplasmic sapnucleotides were labeled with p32 at a faster rate and
I I I incorporated a larger amount than the mitochondrial1 2 3 4 nucleotides. The mitochondrial and cytoplasmic sap
DAYS nucleotides (fig 3-B) of the radicle were labeled at1. Changes in the protein content in various the same rate on the 1st, 2nd, and 3rd days, but themic fractions of corn radicles and scutella during mitochondrial nucleotides were labeled at a faster'mination and growth. rate on the 4th day. The rate of phosphate incor-
-J
a:c)
zLH0a:a-
n
D
-J-jwI-
(I)
zw0
a.
0
1200
800
400
0
9965
www.plantphysiol.orgon July 6, 2018 - Published by Downloaded from Copyright © 1961 American Society of Plant Biologists. All rights reserved.
PLANT PHYSIOLOGY
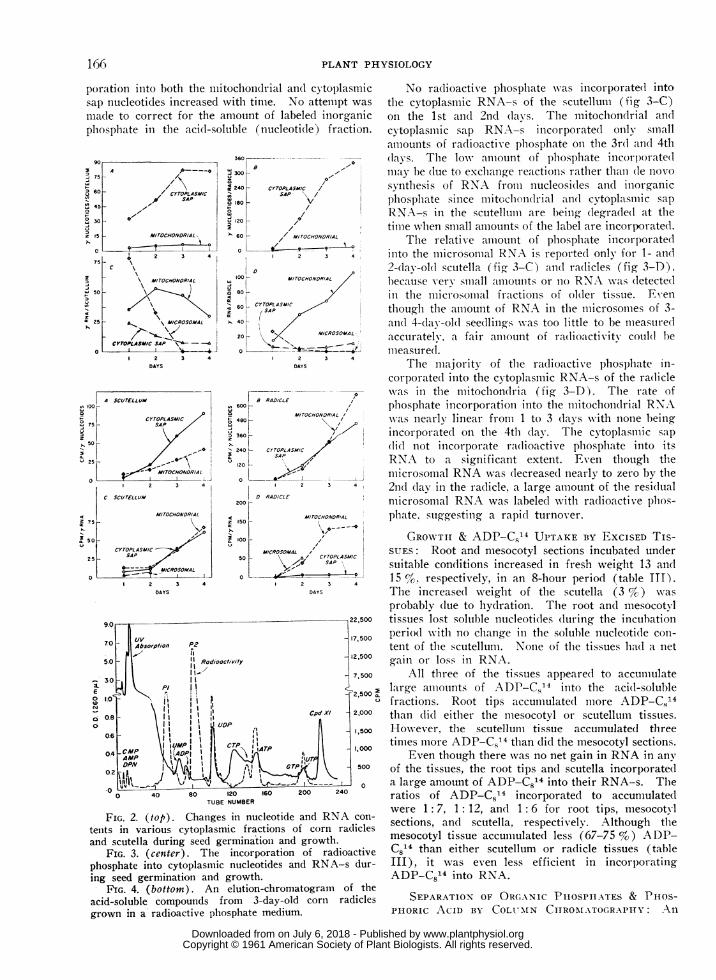
poration iinto both the mitochonidrial anl( cytoplasmicsap nucleotides increased with time. No attemlpt was
mlade to correct for the amount of labeled inorganicphosphate in the acid-soluble (nucleotide) fraction.
A
-75/
060 A ,DMo-n ,e/ SAP145
°30J
15 - M/rOCHON/OR/AL
2 3 4,75
\ M/TrOCCHONOR/AL
25-MI\ MICROSOMAL
cYroPLASM/C SAP
01 2 3 4
DAYS
A SCu/TELLUM
OsooLF75 ~cyroPLAsMIC f
7 5 SAP ,,
/ 4 A/TOCNO$VOR/AL
50
25 SA
C SCUrTOLLUM
~~~~~M/TOCHONDRIAL75
5.0
CYTOPLASMIC25
SAP
M/ICROSOMAL
0I2 3 4
DAYS
360
S0300- /
240 -CYrOLAsM/
10SAP
o ~~~/120 /
z/60 - / MIrOCHONDR/AL
2 3 4
100 - MI rOCH/ONDR/AL
o80
60 _ CYrOPLASM/C(SAP
40
MICROSOMAL
0
2 3 4
DAYS
9.0 22,500
70 _ UV P2 17,500Absor'pfiOnl
5.0 - |, Radiooctivity _2,500
7,500
10 41
~~~~~~CpdX/ 2,000
~~~~~~~~~~1500
0m r
0 0
TUBE NUMBER
FIG. 2. (top). Changes in nucleotide and RNA con-tents in various cytoplasmic fractions of corn radicles
and scutella during seed germination and growth.
FIG. 3. (center). Tile incorporation of radioactive
phosphate into cytoplasIic nucleotides and RNA-s dur-
ing seed germination and growth.FIG. 4. (bottom). An elution-chromatogram of the
acid-soluble compoulnds from 3-day-old corti radiclesgrown in a radioactive phosphate medium.
No radioactive phosphlate \vas incorporate(l intothe cytoplasmiiic RNA-s of the scutellum (fig 3-C)on the 1st and 2nd davs. The mitochondrial and
cytoplasmaic sap RNA-s incorporated only smnallamiioulnts of radioactive plhosplhate o01 the 3rd and 4th(las. The low amiiouint of phosphate incorp)orate(lmay he (lue to exclhange reactionis rather than (le llOVOsyntlhesis of RNA froml nucleosi(les anid inorganicplhosplhate since mitochondrial an(l cytoplasmic sapRNA-s in the scutelltiumi are being (legra(le(l at thetimiie wlhen slmiall amiiounts of the label are incorporated.
The relative amiiounit of phosplhate incorp)oratedinto the mlicrosolnlal RNA is reported only for 1- and
2-day-old scutella (fig 3-C) and raldicles (fig 3-D),l)ecause verv snlall amotunts or no RNA wvas (letectedin the microsomnal fractions of ol(ler tissue. Even
thouglh the amiiount of RNA in tile mlicrosomiies of 3-ani(l 4-day-old seedlings was too little to be imieasuriedaccurately, a fair amiiount of radioactivity could( he
ilieasure(l.The nllajoritv of the radioactive phosphate in-
corporate(l iilto the cytoplasmic RNA-s of the ra(liclewvas in the lllitocllon(lr-ia (fig 3-D). The rate ofpliospllate iilcorporationi into the illitochoildrial RNAwas nearlv linear fronm 1 to 3 dlays \ith none heingincorporated oIl tlle 4th day. The cytoplasmlic sap
li(l lot incorporate ra(lioactive pllospllate iilto itsRNA to a significant extent. Even tiouigll themicrosoimal RNA was (lecreased ilearly to zero by the21(1 (lay in the radicle, a large aillount of the residualnlicrosoillal RNA was labeled witli radioactive pllos-phate. suiggestinig a rapid turnover.
GROWThr & ADP-C914 UPTAKE BY EXCISED Tis-SUES: Root and mesocotyl sections incubated undersuitable conditions increased in fresh weight 13 and
15 %. respectively, in an 8-hour period (table ITI).The increase(l weight of the scutella (3 %) was
probably (lue to iy(dration. The root and illesocotvltissues lost soluble nucleoti(les (luring the incubationperiod witil no cilange in the soluble ilucleotide coil-
teilt of the scutellulll. 'None of the tissues lla(l a netgain or loss in RNA.
All tilree of the tissues appeare(l to accuillulatelarge aillounts of ADP-C514 inlto the aci(l-solublefractions. Root tips accumulated niore ADP-C514than diil eitlier tile imlesocotyl or scutellum tissues.How,vever, tile scutelluinl tissue accunlulate(1 tilreetimes more ADP-C514 than did the mesocotyl sections.
Even though there was no net gain in RNA in any
of the tissues, the root tips and scutella incorporateda large amount of ADP-C814 into their RNA-s. Tileratios of ADP-C814 incorporated to accunmulatedwere 1: 7, 1: 12, and 1: 6 for root tips, niesocotylsections, anci scutelia, respectively. Although themesocotyl tissue accullulated less (67-75 %) ADP-C814 than eitller scutellum or radicle tissues (tableIII), it was even less efficient in incorporatingADP-C814 into RNA.
SEPARATION OF ORG\NIC PHIOSPIIATES & PHOS-PIIORIC ACID BY COLUMN CHROMAITOGRAPHY: An
166
2 3
DAYS
4
www.plantphysiol.orgon July 6, 2018 - Published by Downloaded from Copyright © 1961 American Society of Plant Biologists. All rights reserved.

CHERRY & HAGEMAN-NUCLEOTIDE & NUCLEIC ACID METABOLISM
elution-chronmatogram of the ultraviolet absorbingand P32 labeled components from 3-day-old radicletissue which had been grown in a radioactive phos-
phate medium is given in figure 4. Similar elution-chromatograms were obtained from 3-day-old scutel-lum tissue; however, due to the likeness to theirradicle elution-chromatograms only the latter will bepresented. The identification of the nucleotides was
established by the order and position in which theywere eluted from the column as previously described(3). All the comlmonly occurring nucleotides pre-
viously identified were labeled with p32 by the 3rdday.
In general, the radioactive peaks were muchsharper than the ultraviolet peaks (O.D.. 260 myA)which suggests that other ultraviolet absorbing com-
poundls, not nucleotides, are adsorbed to the Dowexand are eluted from it contaminating the nucleotidepeaks and thus interfering with quantitative nucleo-tide measurements. Generally, UDP, ATP, andUTP contained most of the nucleotide P32 label.There were two radioactive peaks (P1 & P2) whichdid not correspond to any of the ultraviolet peaks.P1 and P2 contained 10 and 42 % of the total radio-activity added to the column, respectively. Thecomposited eluates of P1 and PO were each lypholyzedto dryness and co-chromatographed on paper usingtwo different solvents (Methods). Each of the uin-known samples (P1 & P,) contained only one com-
ponent. The Rf values of P2 in both solvents usedwere the same as that of phosphoric acid. The identi-ty of P2 as phosphoric acid was further verified byadsorbing radioactive phosphate on a Dowex-1 X 8column using ADP as a marker and eluting with thesame system as used for column chromatography ofthe acid-soluble nucleotides (3). Radioactive phos-phoric acid was eluted from the column at the same
tube number as was P2.The identity of P1 was not established; however,
its migration in both solvents was similar to thehexose monophosphates. Due to the small amount ofP1 obtained from the column, a sugar moiety was
not determined. The position of P1 on the elution-chromatogram suggests that it may be a sugar phos-phate.
It is interesting that very little of the radioactivephosphate was incorporated into the nucleoside mono-phosphates. No trace of radioactivity was found inCompound XI which agrees witlh previous work (3)that Compound XI is not a nucleotide.
DISCUSSIONThe evidence gained from these experiments indi-
cates that there is a dynamic change in the particulatesof radicles and scutella as corn seedlings grow. TheRNA measurements, which may be regarded as an in-dex to particulate mass, show that the microsomessedimenting at 110,000 g rapi(dly decrease with seed-ling growth (fig 2). The small microsomal par-ticulates appear to be either completely degraded orattached to larger or similar particulates so thatsome sediment with mitochondlria as the plant ma-terial matures. If the microsomes are attached toother particulates so that they have a mass similarto mitochondria or if they are attached to themitochondria, it is not known whether or not thecytological function of the microsomes is also changed.Although the cytoplasmic particulates undergo a dy-namic change, it seems unlikely that the microsomesare completely degraded since protein is synthesizedon the 4th day. The rapidl clecrease in microsomalRNA with growth is probably due to the decline innumber of active cells as compared to mature cells.
Investigations by Lund et al (9) clearly showthat there are changes in cytoplasmic particulatesduring growth and maturation of root cells whichcorrelate with changes in the composition and phys-iological activity of these cells.
The mitochondrial RNA of the scutellum andradicle tissues may be correlated with the changes ofthe mitochondria. Lund et al (9) reported that im-mature mitochondria develop cristae and reach ma-
turity as roots grow. Hanson et al (7) reported thatthe mitochondrial mass of corn scutellum increasedwith germination for 3 days, then began to decline.During germination of corn seed the respiratoryactivity of the scutellum (7) increased for 3 to 4 dayswith a decline in respiration thereafter. There ob-servations on the development of mitochondria in
TABLE IIICHANGES IN FRESH WEIGHT, SOLUBLE NUCLEOTIDES, & RNA, & ADP-C814 UPTAKE BY
SCUTELLUM & ROOT TiP & MESOCOTYL SECTIONS IN 8-HOUR PERIOD
TISSUE Ag/gm FRESH WEIGHT ADP-C814 UPTAKE1TISSUE GROWTH SOLUBLE NUCLEOTIDE RNA (cpm/gm FRESH WT)
(INCREASE) INITIAL FINAL INITIAL FINAL SOLUBLE RNANUCLEOTIDES RN
Root tips (1 cm) 12.8 1,630 1,375 2,290 2,260 11,875 2,033Mesocotyl section
(1 cm) 15.2 1,530 1,235 813 835 3,370 317Scutella 3.0 959 963 2,170 2,185 9,210 1,980
1 The sum of radioactivity in the soluble nucleotide and RNA fractions is considered as the amount accumulated.
167
www.plantphysiol.orgon July 6, 2018 - Published by Downloaded from Copyright © 1961 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
corn scutellum can be associated with the increase inmitochondclrial RNA (fig 2).
As corn seed germinates there is a steady declinein scutellum RNA and( a synthesis of radicle RNA(table IT). The loss of scutellum RNA is greaterthan the increase in soluble nucleotides. A possibleexplanatioll for the mode of RNA utilization is thatthe RNA is (legraded to nucleotides and used forgrowvth of the embryonic plant or that a transportableRNA is moved to the growing tissue where it is used.
SU MAAIARY
The clhange in amounts of nucleoti(les aln(l RNAin corn see(llings during germination has been studied.Normally , as corn seeds germinate there is a produc-tion of nucleotides in the scutellum and in the radicletissues. Both scutellum and radicle tissues incorpo-ratedl fairly large amounts of radioactive phosphateinto the acid-soluble nucleoti(les w-ith the radicle ac-
cumulatinig anid incorporating radioactiye phosphateat a faster- rate thani the scutellum.
RNA of the scutellunm w\as rapidly (legralded ap-
parently at the expense of the microsomes. Mito-chond(lrial RNA w-as higlhest oIn the 2nd an(d 3rd dayswvith rapi(ldegradationi occurring on the 4th (lay.
The amiiount of radioactive phosplhate incorporatedinto the scutelluml RNA-s appears to be (lue to ex-
chanige reactions rather than (le novo synthesis.Generally, all of the RNA of the radicles was in
the mitochonidria except for 1-day-old radlicles whichlhad abotut hallf of the RNA conlteint in the mllicrosoimles.All of the radioactive phosplhate incorporated intothe cytoplasimiic RNA-s in the radicles was ultimiatelylocalize(d in the mlitoclhonid(ria. The cytoplasmic sap
RNA incorporated no ra(lioactive phosphate.The protein content increase(d in all the cyto-
plasmiiic fractions in the radlicle froml 1 to 4 (layswhereas the protein contenit in the scutellunm cyto-plasnmic fractions decrease( with germlinatioin.
The nucleotide contenit of excise(d root tips andmlesocotyls decreased in an 8-hour incubation periodwith ADP-C814. The scutellunm niucleoti(les (lid notchange in the same perio(l. The RNA contents ofthe root tips, mesocotyl sectionis, andl scutella remiiainedthe samle throughout the inicubation perio(l. however,the root tips and scutella incorporate(d a fairly largeamount of ADP- '4 into their RNA-s. Mlesocotylsections dlid not incorporate ADP-C5'4 into its RNAas Nell as root tips or scutella.
The organic phosphates andl phosplhoric acidlabele(d with P32 from 3-day-old radicle tissue were
separated by ion-exclhailge chromatography. Thenucleosi(le di- an(d triphosphates contained most ofthe nucleotide radioactive phosphate. Two radlio-active peaks were separated that did not correspondto any nucleotide peaks. The first radioactive peak
was not identifiedl by paper co-chromatography butit is believed to be a sugar phosphate. The sec n1 1peak was identified as phosphoric aci(d using paperco-chromllatography andl ion-exclhange chromliatog-raphy .
ACKNOWLEDGIMEN\Ve recor(l our appreciation to the Atomic Energy
Commissioni (grant no. 791 ) for its support.The authors are indebtedl to Mrs. Donna Flesher-
al(l i\frs. Katherine Trupil for- techniiical assistalce.
LITERATURE CITE4 D1. BERGKVIST, R. 1956. Acid-soluble niucleotidles of
wheat plants. Acta Chemica Scan(l. 10: 1303-1316.2. BLOCK, R. J. 1952. Paper Chromotographv, A
Laboratory -Manual. Academic Press, Inc. NewxYork. P. 86-90.
3. CTIERRY, J. H. & R. H. IHA(GEMiAN. 1960. Separa-tion & identification of soluble nucleotides fromletiolated corn seedlings as a functioni of grow tl.Plant Physiol. 35: 343-353.
4. CHERRY, J. H., R. H. HAGEMAN, J. N. RUTGER, &J. B. JONES. The acid-soluble nucleotides & ribo-nucleic acid of different corn inbreds & single-crosshybrids. Cro1) Sci. (In Press)
5. HANSON, J. B. 1958. The effect of ribonuclease onoxidative p)hosphorylation by mitoclhondria. T.Biol. Chem. 234: 1303-1306.
6. HANSON, J. B. 1960. Impairmlent of respiration,ion accumulation, & ionl retenltioni in root tisstetreated wvith ribonuclease & etlhyleniedliaminie tetra-acetic acid. Plant Physiol. 35: 372-379.
7. HANSON, J. B., A. E. VATTER, M(. E. FISTFII. &
R. F. BILS. 1959. The (levelop)ment of mitoclloni-(Iria in the scutellum of germinating corn. _\gron.J. 51: 295-301.
8. OwARY, O. H., N. J. RoSEBROUGII. .A. L. FARR, &R. J. RANDALL. 1951. Protein mleasuiremnent witlthe Folin-phenol reagent. J. Biol. Cllemii. 193:265-275.
9. Ix.ND, H. A., A. F. VATTF-ER & T. B. HANSON. 1958.Biochemical & cytological chaniges accompanyingg,rowth and differentiation in the roots of Ztia n11m1as.J. Biophys. Bioclhelm. Cy'tolo-y. 4: 87-98.
10. OGUR, M. & MI. ROSEN. 1950. The nucleic .cid.9of plant tissues. I. The extraction & estinmationof deoxypentose nucleic acid & pentose nucleic aci(d.Arch. Biochem. 25: 262-276.
11. OOTA, Y., R. Fuc.Ir. & S. OSAwVA. 1953. Clhangesin chemical constituenits during the germinationstage of a bean, Vigna scsqm'ipd(dlis. J. Bioclhem.(Tokyo). 40: 6494061.
12. OOTA, Y. & K. TAKATA. 1959. Chlaniges iil imicro-.somal ribonucleoproteins in the time course of thegermiination stage as revealed by electrophoresis.Physiol. Plantarum. 12: 518-525.
13. OSAwVA, S., & Y. OOTA. 1954. Growth & pentosenucleic acid content of bean embryo. Experientia9: 96-98.
168
www.plantphysiol.orgon July 6, 2018 - Published by Downloaded from Copyright © 1961 American Society of Plant Biologists. All rights reserved.