neural crest cell behavior in white and dark larvae of ambystoma mexicanum: differences in cell...
TRANSCRIPT

THE JOURNAL OF EXPERIMENTAL ZOOI,OGY 229:91-107 (1984)
Neural Crest Cell Behavior in White and Dark Larvae of Ambystoma mexicanurn: Differences in Cell Morphology, Arrangement, and Extracellular Matrix as Related to Migration
J. SPIETH AND R.E. KELLER Department of Biology, Indiana University, Bloomington, Indiana 47402 (J.S.) and Department of Zoology, University of California, Berkeley, Berkeley, California 94720 (R. E X )
ABSTRACT Melanocytes of white (did) larvae of the Mexican axolotl (Arn- bystoma mexicanurn) are confined to the dorsal midline of the trunk region, whereas in dark (D/-) larvae they are spread laterally on the flank as well, where they contribute to the normal pigment pattern of the trunk. Pigment cell migration in the subepidermal space of white larvae is inhibited by the white epidermis (Dalton '50; Keller et al., '82). The present scanning electron microscopic study describes a well-defined sequence of changes in shape and arrangement of neural crest cells during and after their segregation from the neural tube in both dark and white axolotls. The morphology of the neural crest cells migrating in the subepidermal pathway of dark larvae is correlated with their motile behavior and pattern of migration in vivo, as described by time-lapse cinemicrography (Keller and Spieth, '83). Also, the structures of the matrix material in the subepidermal space of dark and white axolotls differ in ways that may be related to the epidermal inhibition of migration in the latter. Numerous possibilities for contact guidance offered by the structure and topog- raphy of the substrata, neighboring cells, and the extracellular matrix in the migration path are described and discussed.
Larvae of the white strain of the Mexican axolotl (Ambystoma mexicanurn) bear melan- ophores only along the dorsal midline of the trunk region, whereas in dark larvae melan- ophores are spread across the flank as well. The white larval phenotype is due to the homozygous recessive condition (did) of the gene d, resulting in the failure of pigment cells, or their neural crest progenitors, to mi- grate laterally in the subepidermal space (see Dalton, '50; Keller et al., '82). Patches of white epidermis grafted to dark hosts block migration in the areas immediately beneath the grafted patches. Conversely, patches of dark epidermis grafted to white hosts sup- port migration in the area under the grafted patch. These results indicate that the epider- mis of the white axolotl has a local, inhibi- tory effect on migration.
In the present study, we find that neural crest cell morphogenesis in the axolotl in- volves distinct morphological phases during their segregation from the neural tube, dur- ing their elongation and alignment prior to
migration, and during their migration. These are characterized by specific cellular mor- phologies and packing patterns that differ in important ways from similar phases seen in the chick or the mouse. The morphology of migrating neural crest cells is correlated with their motility and pattern of migration in vivo, as seen by time-lapse cinemicrography (Keller and Spieth, '83). We describe similar- ities and differences between neural crest morphogenesis in dark and white axolotls. We also describe differences in the subepider- ma1 extracellular matrix of white and dark larvae that may be related to the inhibition of subepidermal migration in the white axo- lotl. Finally, a more complete analysis of the topography of the substrata suggests that the neural crest migration pathway is structur- ally more complex than previously thought and that it poses numerous possibilities for contact guidance of cell movement.
Address reprint requests to R E Keller, Department of Zool- ogy, University of California, Berkeley, CA 94720
0 1984 ALAN R. IJSS, INC.

92 J. SPIETH AND R.E. KELLER
MATERIALS AND METHODS
Dark (Did), D/D) and white (dd) embryos were obtained from natural spawnings in the Axolotl Colony at Indiana University. They were manually dejellied in 20% Steinberg’s solution and fixed overnight a t stages 16 through 38 (Detlaff, ’75) in fixatives contain- ing either 2.5% glutaraldehyde, or 1.5% glu- taraldehyde and 1.5% formaldehyde (from paraformaldehyde) in 0.1 M sodium cacodyl- ate buffer (pH 7.4). Both fixatives were also used with the addition of 1% cetyl pyridi- nium chloride (CPC). Dissections were done in buffer with a fine steel knife and watch- makers forceps. For cryofracturing, the spec- imens were placed in Parafilm tubes in absolute ethanol, frozen in liquid nitrogen, and fractured transversely on a steel block with a razor blade (Humphreys et al., ’74). Processing of otherwise intact larvae re- quired removal of the heads and tails during fixation to prevent an expansion of the sub- epidermal “space” that was observed to oc- cur without this treatment. The dissected or cryofractured specimens were dried in a crit- ical point dryer from absolute ethanol, coated with gold: palladium (60:40) in a vacuum evaporator or sputter coater, and observed and photographed with an Etec AutoScan electron microscope. Magnifications were corrected with a measured grid.
RESULTS Segregation of neural crest cells fiom the
neural tube The early development of the axolotl trunk
neural crest consists of three distinct phases, similar to those described by Tosney (‘78) for avian neural crest development: (1) segrega- tion from the neural tube; (2) elongation and alignment; and (3) migration. At any stage of development of the animal, these events are most advanced in the anterior region where the neural folds first meet (Fig. l a ) and progress posteriorly. The prospective neural crest can be recognized for the first time, prior to neural fold fusion, in trans- versely fractured specimens (Fig. lb) or in embryos from which the epidermis has been removed (Fig. lc) as a distinct population of cells in the dorsomedial region of the neural fold. They are characterized by a rounded, pleiomorphic shape and stand in contrast to the regular, elongate shape and radial orien- tation of the remaining cells of the prospec- tive neural tube (Fig. lb-e). These cells were identified by light microscopy as neural crest
cells (Raven, ’31; Detwiler, ’37). Noticeable intercellular spaces are often found between prospective neural crest cells and they bear numerous filiform protrusions that extend across the surfaces of adjacent cells (Fig. Id). In contrast, cells of the prospective neural tube are tightly packed and show few lateral protrusions (Fig. le).
After the neural folds fuse and the epider- mis becomes continuous over the neural tube, a cord of cells, five to six tiers wide, compris- ing the definitive neural “crest” is found in the dorsal midline of the neural tube (Fig. 10. If these cells are mechanically removed (Fig. lg), or when the neural tube is viewed in transverse fractures (Fig. lh), it is evident that, immediately after fusion of the neural folds, the dorsal edges of the prospective neural tube are separated by a wedge of neural crest cells. The columnar cells of the tube first meet a t their inner (luminal) ends, and then their adjacent surfaces move to- gether, progressing dorsally from this point of first contact, and exclude the neural crest from the neural tube (Fig. lh,i). After exclu- sion, the columnar cells form a continuous surface-the roof of the neural tube-on which the neural crest cells rest (See Fig. 2d).
Elongation and alignment Beginning at stage 28 in the anterior (de-
velopmentally most advanced) region and proceeding posteriorly, the newly segregated neural crest cells become elongate and spin- dle shaped. Concurrently, they become aligned with one another, and oriented par- allel to the long axis of the neural tube (com- pare Fig. If and j). Newly segregated cells are loosely packed and have spaces between them, but during elongation and alignment, they draw together in a compact cylindrical cord and the spaces between them are re- duced (compare Fig. If and j). Segregation is complete, or nearly so, before elongation and alignment begin, and elongation and align- ment are complete before migration begins. The neural crest is completely extruded from the neural tube and well formed before lat- eral migration begins. It is during this period after their segregation and prior to migra- tion that the crest can be mechanically re- moved as an intact cord of cells and cultured or grafted [See Fig. 10 of Keller and Spieth (‘83)l.
Onset of migration At stage 29, neural crest cells in the ante-
rior trunk region of the dark axolotl embryo

AXOLOTL NEURAL CREST MIGRATION 93
begin to migrate laterally. The early migra- tion begins and continues only a t the basal region of the crest where it is in contact with the neural tube (Fig. 2a,b). The elongate, spindle-shaped cells of the basal region of the crest form large protrusions that extend lat- erally on the inner surface of the epidermis (Fig. 2b). These protrusions may form lat- erally, midway along the length of the cell or at either end (Fig. 2a), and the migrating cells become reoriented such that they lie perpendicular to the cells remaining in the crest (Fig. 2a,b). Only basal cells undergo this reorientation and begin migration.
Migration The migrating cells pass a short distance
between the dorsolateral surface of the neural tube and the overlying epidermis, us- ing one, the other, or perhaps both, as a sub- stratum. They are elongate and oriented parallel to the apices of the cells of the neural tube and perpendicular to the cells remain- ing in the crest (Fig. 2c,d). After traversing this common, initial path, they reach the apex of the somite where they either move ventrally between the somites and neural tube (the perineural migration route), or con- tinue laterally between the epidermis and the outer surface of the somites (the subepi- dermal migration route). Crest cells in the perineural route may be attached to the lat- eral wall of the neural tube (Fig. 2c,f) or the inner surface of the somite (Fig. 2e), though the latter cells can not be readily distin- guished from schlerotome cells beyond stage 32 (see Detwiler, '37). In both cases they are elongated and aligned parallel to their direc- tion of migration; this direction is parallel to the elongated apices of the cells of the tube (Fig. 2c,d), whereas it is perpendicualr to the anterioposterly aligned cells of the somite (Fig. 2e). The perineural migration appears to be normal in white larvae (Fig. 2c), in contrast to the lack of migration in the sub- epidermal route [see below and Keller et al., ('82)l.
The amorphous and fibrillar extracellular matrix in the perineural pathway extends just beyond the extent of migration at any given stage, whereas the surface of the tube ahead of the migration is clean (Fig. 2f,g). It is possible to find small areas in which the fibrils of matrix are perhaps aligned parallel to the migration route, but this is not a con- sistent feature (Fig. 2g).
Those cells that choose the subepidermal route continue laterally and then ventrally
on the outer surface of the somite (Fig. 3a) or innner surface of the epidermis (Fig. 3b). Time-lapse cinemicrography shows that these cells are highly polarized with nearly all pro- trusive activity localized at their ventral, leading edges, and that they migrate direc- tionally from dorsal to ventral with high ef- ficiency (Keller and Spieth, '83). As individuals, the cells vary in shape from elongate to rotund (Fig. 3a-c), but as a popu- lation they are elongate and oriented paral- lel to their path of migration [Fig. 4 of Keller and Spieth ('8311. The variation in shape as seen in the SEM reflects their fixation at various times in the cyclical process of cell shape change associated with translocation in vivo [see Figs. 5 and 6 of Keller and Spieth ('83)j. They usually have one to three large protrusions of the cell body a t their leading (ventral) edges that are attached locally to the substratum by small filiform or lamellar protrusions (Fig. 3b-e). The fine filiform pro- trusions appear to be continuous with a thin mat of fibrils on the inner surface of the epidermis (Fig. 3b,f-i) or on the somite (Fig. 3c). In contrast, the dorsal (trailing) edges are usually blunt or rounded and generally ap- pear to have fewer protrusions attached to the substratum (Fig. 3b,d,e). The formation of two or three large protrusions at the ven- tral, leading edge Wig. 3c,e; left cell in Fig. 3d) is common but is always followed by re- traction of all but one, which then leads the ventrally directed movement of the bipolar cell [See Figs. 3 and 6 of Keller and Spieth ('83)l.
Neural crest cells adhere to the inner sur- face of the epidermis and to the outer surface of the somite. They may use both as a sub- stratum. Some cells have protrusions broken in the process of dissection that suggest adhe- sion to both surfaces at the same time (Fig. 30 and others appear to be suspended in the fibrillar matrix (Fig. 3i; also see Fig. 5).
In the axolotl, the first neural crest cells to migrate into the subepidermal space form melanophores, whereas those following later form xanthophores (see Lehman, '57). Nearly all cells seen by SEM in the subepidermal space prior to stage 36 are developing melan- ocytes (see Keller et al., '82).
Few pigment cells migrate laterally in the subepidermal space of white larvae (Keller et al., '82), and those that migrate, do so in the intersomitic furrows (Fig. 3j). Most have protrusions and morphologies similar to those of dark larvae (Fig. 3j,k) but their pro- trusive activity is sluggish [see Fig. 9 of
94 J. SPIETH AND R.E. KELLER

AXOLOTL NEURAL CREST MIGRATION 95
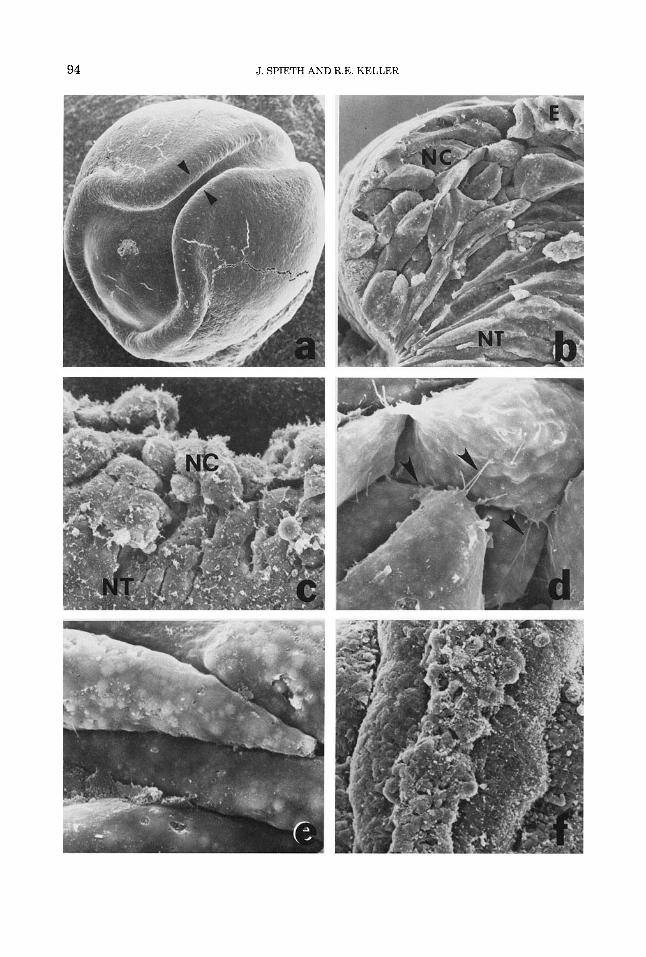
Fig. 1. An SEM of a stage 17 neurula (a) shows the region of first contact and fusion of the neural folds (pointers). A fracture 6) through the region marked by the pointers in (a) shows the contrasting shapes of the prospective neural crest cells (NO, those of the prospec- tive neural tube (NT), and the epidermis (E). The same are shown in a dorsolateral view of the neural fold with the epidermis removed (c). Cells of the neural crest are rounded and bear filiform protrusions (d), whereas those of the neural tube are closely packed and have fewer connections of this type (e). With closure of the tube, the neural crest appears to be excluded from the tube (0, but
removal of the crest cells shows that they are embedded in the dorsal wall of the tube (outlined by pointers, g). Transverse fractures show the position of the neural crest cells early (outlined by pointers, h) and late ii) in their extrusion from the neural tube. A dorsal view of a stage 28 dark embryo with the epidermis removed shows the compact cord of elongated, aligned, and tightly packed neural crest cells Q). Anterior is toward the top in f and g and to the right in j. Magnifications are: a, x30; b, x300; c, ~ 5 5 0 ; d and e, X1750; f, X 190; g, ~ 4 5 0 ; h, ~ 2 5 0 ; i, x380;.j, ~ 3 2 0 .
96 J. SPIETH AND R.E. KELLER

AXOLOTL NEURAL CREST MIGRATION 97
Keller and Spieth ('83)J and most never lose contact with the crest (Fig. 3k). The fibrillar matrix also forms in the white larvae (Fig. 3k). In both white and dark larvae, these fibrils are not oriented in some areas (Fig. 31) and are oriented parallel to the direction of migration in other areas (Fig. 3m).
From stage 36 onward, the majority of me- lanocytes in dark larvae become multipolar, spread centrifugally, occupy greater area, and form an anastomosing reticulum around a central cell body (Fig. 3n). These changes are correlated with a slowing and finally a cessation of movement, and with a loss of contact-induced retraction of cells after colli- sion as seen by time-lapse cinemicrography of cells in vivo and in culture (Keller and Spieth, '83).
Differences between neural crest cells of white and dark larvae at the onset of
migration The neural crest cells of dark and white
axolotls are similar in their emergence from the dorsal surface of the neural tube and in their initial morphology and arrangement (Fig. 4a,b). However, during elongation and alignment (farther anteriorly in Fig. 4), those of white larvae are more spherical and ap- pear to be covered with more fibrillar and amorphous matrix material (Fig. 4d), whereas those of the dark larvae are more elongate, and appear to have less material between them and on their surfaces (Fig. 44. Further anteriorly, the neural crest cells of dark larvae become elongated and aligned (Fig. 4e), whereas those of the white often show less elongation and alignment (Fig. 4f). In dark larvae, protrusions extend laterally across the dorsolateral surface of the neural
Fig. 2. A ventral view (a) of the crest (C) shows the basal cells of the crest turning laterally (pointers) on the inner surface of the epidermis (E). If most of the crest is removed, the basal cells (pointers) that have spread on the neural tube (NT) and have begun to migrate are seen (b). The elongated, migrating cells initially use the dor- solateral wall of the neural tube (c) or the apposed inner surface of the epidermis (a) as a substratum. The cells of the neural tube are aligned parallel to the direction of migration (d). Cells moving between the somite and the neural tube (perineural migration path) may he attached to the inner surface of the somite (pointers, e) or the lateral wall of the neural tube (0. A fibrillar, amorphous matrix, found up to the limit of the migration (pointers, 0, is shown at high magnification (g). Magnification and stages are: a , x275, 33; b, x310, 30; c, ~ 3 2 0 , 33; d, ~ 2 9 0 , 30; e, ~ 2 5 0 , 34; f, ~ 3 3 , 160; g, X33, 900. Photo- graphs b and c are of white larvae.
tube as migration begins (see pointers, Fig. 4e), whereas in the white, few cells have such protrusions and generally the cord retains its cylindrical shape (Fig. 40. The alternate re- gions of difference in thickness of the crest along its length occurs in both white and dark larvae but is more prominent in the former (Fig. 4e,f). The significance of this pattern is not known. The anterioposterior banding pattern of alternate melanophore and xanthophore populations occurs in white as well as dark larvae (Keller and Spieth, '83) in spite of the fact that lateral migration does not occur in the former.
Structural organization of the extracellular matrix in the migration path
In transversely cryofractured specimens (Fig. 5a), many neural crest cells appear to be suspended in the subepidermal space, sur- rounded by an extracellular, fibrillar matrix (Fig. 5bXsee Lofberg et al., '80). In dark lar- vae, the subepidermal space contains fine fi- brils that bear numerous small spheres (Fig. 5b) averaging 0.98 pm in diameter (Table 1). Most are associated with the fibrils but some are found on the inner surface of the epider- mis and occasionally on the surfaces of the neural crest cells as well. They are found in large numbers from stage 32 to 36 in the pathway, laterally as far as migration has occurred. In white larvae, the fine fibrils are present (Fig. 5c) but bear fewer spherical bod- ies than the fibrils in dark larvae (Table 1). The fibrils in the white larvae appear to be more numerous and are densely packed and often appear to span the subepidermal space from the somite to the epidermis (Fig. 5c).
CPC appears to be necessary for the pres- ervation of the spherical bodies in dark lar- vae. Without it, the fibrils are less numerous and few of the fibril-associated bodies were seen in both dark and white larvae (see Fig. 5d). If differences between the subepidermal matrices of dark and white larvae are due to differences in preservation, these differences must nevertheless reflect differences be- tween dark and white larvae in the stability of the matrix rather than trivial differences in handling of dark and white specimens, since the matrix around the notochord, which is similar to the subepidermal matrix in ap- pearance, is nearly the same in white and dark larvae under any conditions of fixation and processing (compare Fig. 5e and f).
There was no consistent alignment or ori- entation of matrix fibrils in the subepider-

98 J. SPIETH AND R.E. KELLER
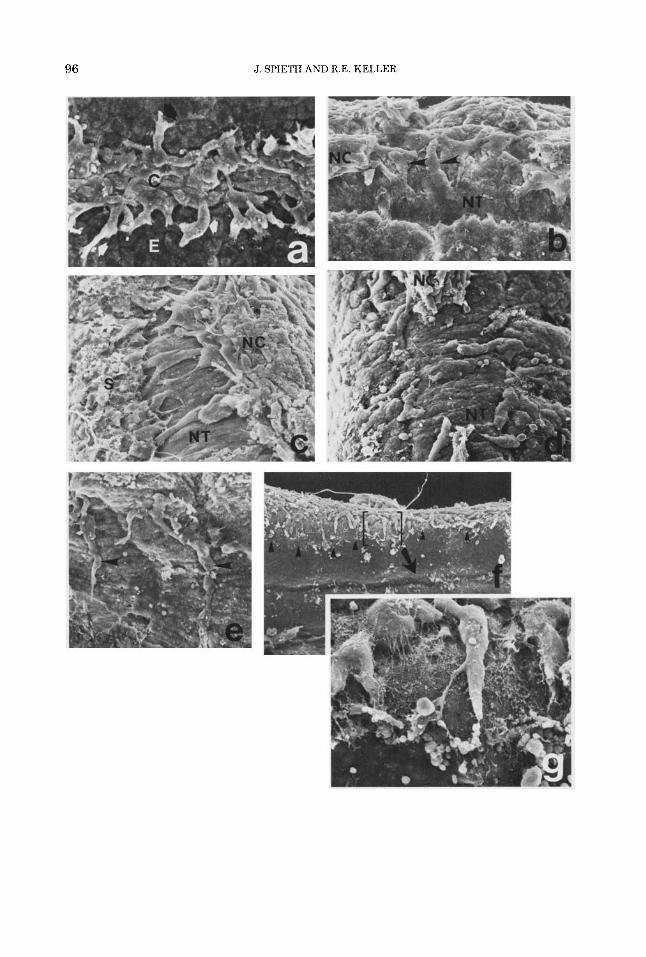
Fig. 3. The pointers (a) indicate cells that have left the neural crest (C) and are moving laterally and ven- trally on the outer surface of the somites (S). Others are attached to the inner surface of the epidermis (b, d, e, 0. Cells moving on the somite (c) or on the epidermis have one to three large protrusions at their ventral (leading) edges (pointers in c,d,e). These end in fine lamellipodia or, more commonly, filiform extensions (f,g,h), which merge with the fibrillar matrix (h,i) in the subepidermal space. A few neural crest cells of the white larvae move laterally in the intersomitic grooves (i), but most remain
attached to the crest fk). Fibrillar matrix on surfaces in the initial migration path may be unaligned (1) or aligned (m). Older neural crest cells that have differentiated into pigment cells (melanophores) form anastornosing reti- cula of large area (n). Magnifications and stages are: a, x270,34; b, x900,32; c, x700,33; d, x400,32; e, x700, 32; f, ~ 1 6 0 0 , 32; g, ~ 1 9 0 0 , 33; h, ~ 8 0 0 0 , 34; i, ~3500, 32; j, ~ 2 7 5 , 34; k, ~ 1 3 0 0 , 32; 1, X4000, 30; m; x2000, 32; n, ~ 1 4 0 0 , 39. Micrographs of j, k, 1, and m are of white larvae.

AXOLOTL NEURAL CREST MIGRATION 99
100 J. SPIETH AND R.E. KELLER

AXOLOTL NEURAL CREST MIGRATION 10 1
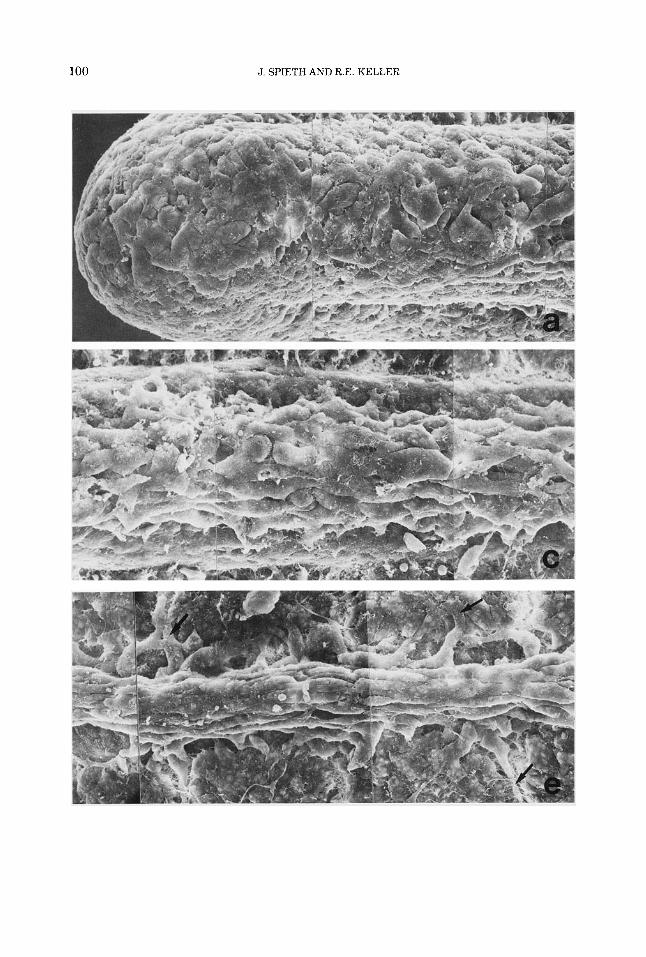
Fig. 4. SEM montages of the dorsal surface of the dark (left panel) and white (right panel) axolotl larvae at stage 32 with the epidermis removed, show the similar- ity of the neural crest cell populations in the two strains immediately after segregation from the tube (a and b). Farther anteriorly, from the mid-20th to the mid-16th or mid-15th somite, alignment and orientation of crest cells
have begun in the dark larva (c) but less so in the white fd). At more advanced stages of development (farther anteriorly, from somites 11 and 12 to 8 and 91, alignment and orientation are complete and lateral migration from the base of the neural crest is underway in the dark (pointers in e) but not in the white (0. Magnification is X425 for all.

102 J. SPIETH AND R.E. KELLER

AXOLOTL NEURAL CREST MIGRATION 103
TABLE I . Spherical bodies in the extracellular matrix of white and dark larvae
Darks (D/-) Whites (did) _________ _ _ _ - _ _ ~ - ~ _ _ _ _ _ _ _ _ ~
Size Suhepidermal space 0.98 pm k 0.24', n= 143 0.98 pm k 0.14, n = l l Perinotoehordnl space 1.36 pm k 0.36, n=36 1.57 pm i 0.49, n=36
Density Subepidermal space 311100 pm', n = 7 14/100 pm2, n=13
'Standard deviation.
ma1 space. In some cases, and in both white and dark larvae, the fibrils appear to be ori- ented across the subepidermal space and per- pendicular to the axis of migration (Figs. 5b,c; see Tosney, '82). In other cases, they are ori- ented parallel to the principal direction of cell movement (Fig. 6a). In most cases, how- ever, only the fibrils associated with protru- sions of the leading edges of the neural crest cells are consistently aligned with one an- other and oriented in the direction of migra- tion; adjacent regions show no such arrangement (Fig. 6b). Thus, alignment of fibrils may be a result of traction during mi- gration rather than preceding it (see Discussion).
The thin fibrillar matrix seen earlier (Fig. 3b) develops as a mat-like meshwork on the inner surface of the epidermis (Fig. 6d-g). At all stages, the matrix develops progressively from dorsal to ventral and anterior to poste- rior (compare Fig. 6d and e with f and g). The inner surfaces of the epidermis of white and dark larvae do not differ consistently in the number, size, arrangement, or appearance of the fibrils (compare Fig. 6d and g with e and f). The epidermal matrix is well developed by stage 39 (Fig. 3n) in contrast to the onset of migration (Fig. 3b-g), but the fibrils remain in random array.
Fig. 5. Cryofracturing of axolotl larvae transversely (a) exposes the neural tube (NT), the neural crest INC), the epidermis (E), the somites (S), the subepidermal space (SES), the notochord (N), and the perinotochordal space (PNS). The matrix that surrounds neural crest cells in a dark embryo 6) consists of a fine fihrillar network hear- ing spherical masses at irregular intervals, whereas the matrix in a white embryo (c) is similar hut does not hear as many of the spheres. In some cases they are nearly absent. If dark larvae are fixed without CPC in the fixative, the fihrillar matrix is retained hut the spheres are absent (d). The perinotochordal matrices in dark (el and white (0 larvae are similar. All micrographs are of stage 32 larvae. Magnifications are: a, X330; h, X3100; c, ~ 2 8 0 0 ; d, x 1900; e, ~ 2 8 0 0 ; f, X3000.
DISCUSSION Patterns of change in cell morphology and
arrangement Axolotl neural crest cells go through a se-
ries of discrete, separate, and well-defined morphological phases during their segrega- tion and migration that may be important in the analysis of how neural crest morphogen- esis is regulated. Each is characterized by specific changes in cell shape and arrange- ment and is probably the result of a specific pattern of cell behavior. Moreover, this series of events differs in important ways from that found in the mouse and in the chick and thus suggests that the basic cellular events in crest morphogenesis may be organized in dif- ferent ways in various vertebrates.
First, the fact that prospective neural crest cells of the axolotl can be identified as rounded, pleiomorphic cells in the fold, prior to neural fold fusion, suggests that these cells do not participate in the elongation that is characteristic of prospective neural tube cells medial to them, nor the flattening that is characteristic of the epidermal cells lateral to them (also see Raven, '31; Detwiler, '37). This behavior is the first morphological expression of their fate. A similar pattern is seen in Xenopus (Schroeder, '70) and in the chick (Tosney, '78, '82). In the mouse, how- ever, the trunk neural crest cells are not morphologically distinguishable prior to neural fold fusion; they appear to participate in neural tube formation and then they crawl out of the tube after its closure (Erickson and Weston, '83).
Second, exclusion of the neural crest cells from the neural tube appears to be a morpho- genetic process that is separate and distinct from migration. They do not begin migration during or immediately after their exclusion from the neural tube, nor do they have the protrusions characteristic of migratory cells during their exclusion. This is in contrast to
104 J. SPIETH AND R.E. KELLER
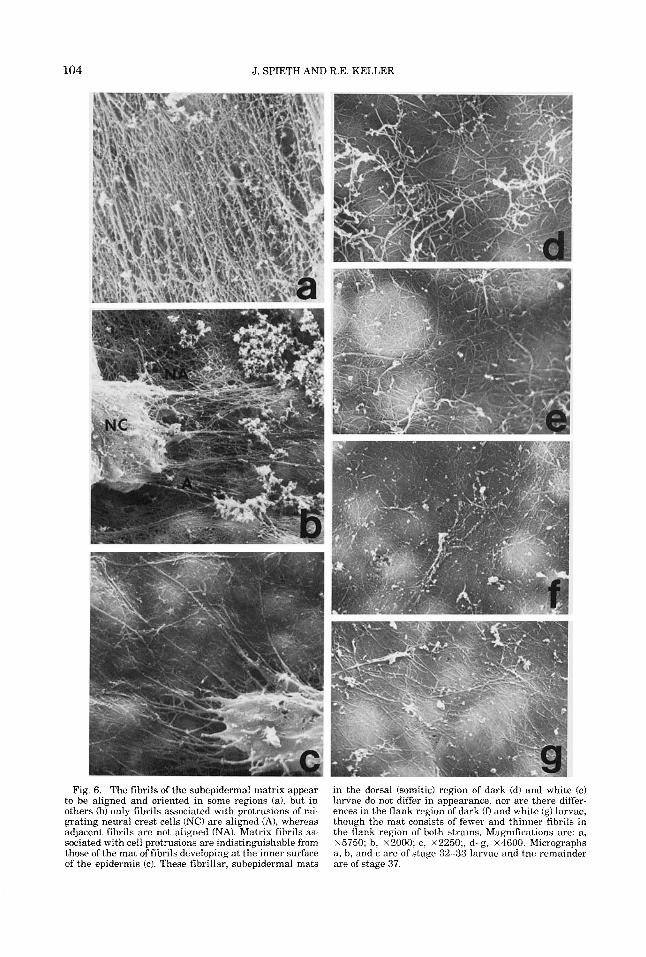
Fig. 6. The fibrils of the subepidermal matrix appear to be aligned and oriented in some regions (a), but in others (b) only fibrils associated with protrusions of mi- grating neural crest cells ( N O are aligned (A), whereas adjacent fibrils are not aligned (NA). Matrix fibrils as- sociated with cell protrusions are indistinguishable from those of the mat of fibrils developing at the inner surface of the epidermis (c). These fibrillar, subepidermal mats
in the dorsal (somitic) region of dark (d) and white (e) larvae do not differ in appearance, nor are there differ- ences in the flank region of dark (D and white (g) larvae, though the mat consists of fewer and thinner fibrils in the flank region of both strains. Magnifications are: a, ~ 5 7 5 0 ; b, ~ 2 0 0 0 ; c, x2250;, d-g, ~ 4 6 0 0 . Micrographs a, b, and c are of stage 32-33 larvae and the remainder are of stage 37.

AXOLOTL NEURAL CREST MIGRATION 105
the mouse (Erickson and Weston, '831, where the neural crest cells actively migrate out of the tube and immediately continue their mi- gration. Third, the axolotl neural crest cells show a unique behavior in which they be- come elongated rostrocaudally, acquire a spindle shape, become closely packed, and orient parallel to the anterioposterior axis. This packing pattern was described by Lof- berg and Ahlfors ('78). Such elongation and alignment does not occur in Xenopus laevis (Spieth, unpublished observations), in the chick (Bancroft and Bellairs, '75, '76; Tosney, '781, or in the mouse (Erickson and Weston, '83). A comparable phase in the chick in- volves the formation of a loose array of mul- tipolar cells, which then condenses into an array of cells that is tightly packed and flat- tened tangentially with respect to the neural tube (Tosney, '78). In the mouse, no such pos- tsegregation, premigration array exists (Er- ickson and Weston, '83).
Fourth, these elongated, spindle-shaped cells of the axolotl crest turn laterally and begin their migration, but in normal devel- opment this initial migration occurs only from the basal region of the crest where it contacts the neural tube. Later, the array of elongated and aligned cells breaks up and the whole crest may be involved in migra- tions. Why the initial migration involves only basal cells in the axolotl is not known. Per- haps only these have access to a suitable substratum-the dorsal neural tube and the apposed epidermis-or perhaps only basal cells are ready to migrate (see below).
Control of amphibian neural crest morphogenesis
It is not clear to what extent each of these morphological phases reflects intrinsic pro- grams of change in neural crest cell affinities or behavior, and to what extent they reflect environmental changes. In favor of the first alternative, the neural crest pigment cell de- rivatives-melanophores and xanthophores at these stages-leave the neural tube and spread on the substratum, in that order, whether in vivo or isolated in culture (Leh- man, '57; Keller, unpublished work). Second, in culture premigratory cells adhere to one another but will not adhere to or spread on tissue culture plastic that will support spread of migratory cells (see Keller and Spieth, '83). Finally, the interaction of axolotl xantho- phores and melanophores that produces barred larval pigment pattern in the axolotl is an intrinsic property of the pigment cells themselves and will occur even when these
cells are grafted to a species that is unbarred (Lehman, '57). In favor of the second alter- native, the results of heterochronic grafting experiments suggest that the onset of migra- tion is controlled by the epidermis (Lofberg and Nynas-McCoy, '81). Moreover, the epi- dermis, rather than the neural crest cells, is responsible for the failure of the subepider- ma1 migration in the white axolotl (see Keller et al., '82). There is some evidence that the white allele is a temporal mutation that does not destroy but simply delays the epidermal capacity to support migration to a time be- yond which the neural crest cells are compe- tent to migrate (Bogomolova and Korochkin, '73). In fact, it appears that some aspects of amphibian neural crest morphogenesis is in- trinsic to the cells and others are environ- mentally controlled. The transition of cell morphology from bipolar and elongate to multipolar and circular seems to occur on schedule, in culture, independent of the in vivo environment, but stability and orienta- tion of protrusive activity are apparently de- pendent on conditions in vivo (Keller and Spieth, '83). These results and the work of Twitty and Niu ('54) and Lehman ('57) sug- gest that in amphibians both intrinsic prop- erties of neural crest cells and environmental influences are important in determining spe- cific aspects of the amphibian pigment pattern.
Structural patterns in the migration pathway Although there is some evidence for sub-
strate organization suitable for contact guid- ance (Lofberg et al., '811, there are inconsistencies that do not favor this mecha- nism as having a major role in guidance. There are many structural patterns in the neural crest pathway that would be expected to act on cells in conflicting ways, given what is known about contact guidance (see Harri- son, '14; Weiss, '45; Dunn and Heath, '76; Dunn and Ebendal, '78; Dunn, '83). For ex- ample, the curved dorsal and dorsolateral surfaces of the neural tube might function in alignment of the elongated cells and perhaps even in confining them in this region, prior to migration (see Dunn, '83), given the evi- dence that elongate cells are sensitive to cur- vature (see Dunn and Heath, '76). However, these elongated neural crest cells then turn and move laterally in the direction of maxi- mum curvature, around the dorsolateral neural tube requiring bending of the cells. Either their initial orientation does not re- flect sensitivity to curvature or their sensi-

106 J. SPIETH AND R.E. KELLER
ble than those in the dark larvae and are ganglia and vertebral arches in Ambystoma. Am. J.
tivity to curvature must change prior to migration. Likewise, as they migrate on the surface of neural tube, their orientation is parallel to that of the elongated apices the cells of the neural tube. But if any use the nearby surface of the somite as a substra- tum, they will be moving perpendicular to the anterioposteriorly elongated and aligned somitic cells. The effect of mechanical and structural properties of the substrate on cell behavior is complex (see Dunn, '831, and given the above inconsistencies, little will be gained by postulating mechanisms of contact guidance in vivo until more is known about the sensitivity of specific cell types to struc- tural patterns and whether these sensitivi- ties are modified in development.
Prealigned or traction-aligned matrix fibrils? Some areas of aligned fibrils have been
seen on the neural tube in this study and by others (Lofberg, et al., '80). However, we find no evidence for consistently aligned and ori- ented extracellular matrix preexisting in the path of the crest cells. Fibrils oriented paral- lel to the migration were usually seen at the ends of protrusion at the leading, ventral edges of migrating neural crest cells. This suggests migration-associated, traction alignment of matrix similar to that seen (in cell culture) by Stopak and Harris ('82), rather than a preexisting, migration-guiding alignment. Likewise, Bancroft and Bellairs ('76) and Tosney ('78) found no evidence of alignment and orientation of fibrils in the chick trunk region. Tosney ('82) found that many fibrils are aligned normal to the mi- gration path in the cranial region of the chick, and we find that the same is true in some regions of the axolotl subepidermal space.
Blockage of subepidermal migration in the white axolotl
Pigment cells fail to migrate laterally in white axolotl larvae because the white epi- dermis blocks or fails to support neural crest migration in the subepidermal region imme- diately beneath it (Keller et al., 1982). In the present study, we find that the number of spherical bodies found on the fibrillar matrix of white larvae is substantially less than that found in dark larvae, but only in the subepi- dermal region where the crest cells that form pigment cells must pass. Either they are not Dresent in white larvae or they are less sta-
extracted during processing. Either way, one must conclude that the subepidermal matrix of the white axolotl larvae is abnormal. The spherical bodies are similar to interstitial bodies (Low, '70) and are found in homolo- gous positions in the axolotl and the chick (Tosney, '78, '82). Tosney ('82) found them to be more abundant in the neural crest path- way than elsewhere. The system of fibrils and interstitial bodies consists of proteogly- cans and fibronectin (Mayer et al., '81) that may be involved in neural crest migration (see Pratt et al., '75; Derby, '78; Pintar, '78; Erickson et al., '80; Newgreen and Thiery, '80; Mayer et al., '81). One of these or related substances may be altered or missing in the white axolotl, and thus result in the altered appearance of the matrix and perhaps in the failure of the white epidermis to support migration.
ACKNOWLEDGMENTS
We thank Ms. Fran Briggs and the Indiana University Axolotl Colony for generously supplying axolotl larvae. Ms. Dot Barone is gratefully acknowledged for her technical as- sistance and Professor Robert Briggs for reading the manuscript and for his support. This work was supported by U. S. Public Health Service Grant RO1 GM 05850-21 to R.W. Briggs and NSF Grant PCM81-10985 to R.E. Keller. J. Spieth was supported by an American Cancer Society Postdoctoral Fel- lowship, PF 1463.
LITERATURE CITED
Bancroft, M., and R. Bellairs (1975) Differentiation of the neural plate and neural tube in the young chick embryo: A study of scanning and transmission electron microscopy. Anat. Embryol., 147:309-335.
Bancroft, M., and R. Bellairs (1976) The neural crest cells of the trunk region of the chick embryo studies by SEM and TEM. Zoon,4:73-85.
Bogomolova, V., and L. Korochkin (1973) Development of pigmentation after transplantation of presumptive epidermis between embryos of white axolotls Ambys- toma mexicanurn of different ages. Ontogene, 4:420- 424.
Dalton, H.C. (1950) Inhibition of chromatoblast migra- tion as a factor in the development of genetic differ- ences in pigmentation in white and dark axolotls. J. Exp. Zool., 215:151-170.
Derby, M.A. (1978) Analysis of glycosaminoglycans within the extracellular environments encountered by migrating neural crest cells. Dev. Biol., 66:321-336.
Detlaff, T.H. (1975) Stages of the Normal Development of the Mexican Axolotl. Translated from Russian in the Axolotl Colony Bulletin, Indiana University, Bloomington, Indiana.
Detwiler, S.R. (1937) Observations upon the migration of neural crest cells. and uuon the develoument of suinal

AXOLOTL NEURAL CREST MIGRATION 107
Anat., 61:63-94. Dunn, G. (1983) Contact guidance of cultured tissue cells:
A survey of potentially relevant properties of the suh- stratum. In: Cell Behavior. R. Bellairs, A. Curtis and G. Dunn, ed. Cambridge University Press, Cambridge.
Dunn, G., and T. Ebendal (1978) Contact guidance on oriented collagen gels. Exp. Cell Res., 1llt475-479.
Dunn, G., and J.P. Heath (1976) A new hypothesis of contact guidance in tissue cells. Exp. Cell Res., 101:l- 14.
Erickson, C.A., K. Tosney and J. Weston (1980) Analysis of migratory behavior of neural crest and fibroblastic cells in embryonic tissues. Dev. Biol., 77t142-156.
Harrison, R.G. (1914) The reaction of embryonic cells to solid structure. J . Exp. Zool., 17:521-544.
Humphreys, W., B. Spurlock and J. Johnson (1974) Crit- ical point drying of ethanol-infiltrated, cryofractured biological specimens for scanning electron microscopy. In: Scanning Electron Microscopy 1974. 0. Johari and J. Corwin, ed. IITRI, Chicago. pp. 276-282.
Keller, R., and J. Spieth (1983) Neural crest cell behavior in white and dark embryos of Amhystoma mexicanum: A time-lapse cinemicrographic analysis in vivo and in vitro. J. Exp. Zool., 229t109-126.
Keller, R., J. Lofberg and J. Spieth (1982) Neural crest cell behavior in white and dark embryos ofAmbystoma mexicanum: Epidermal inhibition of pigment cell mi- gration in the white axolotl. Dev. Biol., 89:179-195.
Lehman, H.E. (1957) The developmental mechanics of pigment pattern formation in the black axolotl, Am- bystoma mexicanum J. Exp. Zool., 135:355-386.
Lofberg, J., and K. Ahlfors (1978) Extracellular matrix organization and early neural crest cell migration in the axolotl embryo. Zoon, 6:87-107.
Lofberg, J., and A. Nynas-McCoy (1981) Epidermal stim- ulation of initial neural crest cell migration in the amphibian embryo. Oral Presentation, ISDB Interna- tional Congress, Basel.
Lofberg, J., K. Ahlfors, and C. Fallstrom (1980) Neural crest migration in relation to extracellular matrix or-
ganization in the embryonic axolotl trunk. Dev. Biol., 75148-167.
Low, F.N. (1970) Interstitial bodies in the early chick embryo. Am. J. Anat., 128:45-56.
Mayer, B.W., E.D. Hay and R.O. Hynes (1981) Immuno- cytochemical localization of fibronectin in embryonic chick trunk and area vasculosa. Dev. Biol., 82.267-
Newgreen, D., and J.P. Thiery (1980) Fibronectin in early avian embryos: Synthesis and distribution along the migration pathways of neural crest cells. Cell Tissue Res., 211:269-291.
Pintar, J.E. (1978) Distribution and synthesis of glycos- aminoglycans during quail neural crest morphogene- sis. Dev. Biol., 67r444-464.
Pratt, R.M., M.A. Larsen and M.C. Johnston (1975) Mi- gration of cranial neural crest cells in a cell-free hyalu- ronate-rich matrix. Dev. Biol., 44398-305.
Raven, C.P. (1931) Zur Entwicklung der Ganglienleiste. I. Die Kinematik der Ganglienleistenentwicklung bei den Urodelen. Arch. f. Entwicklung., I25:200.
Schroeder, T. (1970) Neurulation in Xenopus Zaeuis. An analysis and model based on light and electron micros- copy. J . Embryol. Exp. Morphol. 23:427-462.
Stopak, D., and A.K. Harris (1982) Connective tissue morphogenesis by fibroblast traction. I. Tissue culture observations. Dev. Biol., 90:283-298.
Tosney, K.W. (1978) The early migration of neural crest cells in the trunk region of the avian embryo: An electron microscopic study. Dev. Biol., 62t317-333.
Tosney, K.W. (1982) The segregation and early migration of cranial neural crest cells in the avian embryo. Dev. Biol., 89r13-24.
Twitty, V., and M.C. Niu (19541 The motivation of cell migration, studied hy isolation of embryonic pigment cells singly and in small groups in vitro. J. Exp. Zool., 125:541-573.
Weiss, P. (1945) Experiments on cell and axon orienta- tion in vitro: The role of colloidal exudates in tissue organization. J . Exp. Zool., 100:353-386.
286.