naujausių mokslinių pasiekimų, naujausių mokslinių tyrimo...
TRANSCRIPT

Naujausių mokslinių pasiekimų, naujausių mokslinių tyrimo
metodų molekulinės biologijos srityje mokslinė studija
MOLEKULINĖ EKOLOGIJA
Prof. Aniolas Sruoga
Biologijos katedra
Vytauto Didžiojo universitetas
Kaunas – 2007

1. MOLEKULINĖ EKOLOGIJA: MOKSLO RAIDA IR TAIKYMAS
Šiuolaikinis požiūris į populiacijas, kaip vienos rūšies organizmų genetiškai
skirtingų individų grupes, formavosi visą šimtmetį nuo XX a. pradžios. Kadangi
įvairių sričių specialistai akcentuoja skirtingas pupuliacijų savybes, būtina apibrėžti ką
mes vadiname gamtine populiacija – tai vienos rūšies individų grupė, užimanti tam
tikrą teritoriją arba erdvę, kurioje jie keičiasi genetine informacija, palaiko
atitinkamą gausumą ir turi tik tai grupei būdingus požymius.
Visi organizmai, nepriklausomai nuo jų evoliucijos, su aplinka ir tarpusavyje
yra susieti ekologiniais ryšiais. Organizmų tarpusavio sąveika yra tokia sudėtinga, kad
dar ir dabar mažai žinoma apie konkrečius, tarpusavyje susijusius ir vienas nuo kito
priklausančius ekologinius procesus, vykstančius tarp įvairių individų, populiacijų,
biocenozių. Šių ryšių atskleidimas ir tyrimas – viena iš sudėtingiausių šiuolaikinės
biologijos problemų.
Be to dabartinėse sąlygose tai nėra tik natūraliai vykstantys procesai, nes
žmogaus praktinė veikla griauna evoliuciškai susiklosčiusius, natūralius mechanizmus
ir pirmiausiai tuos, kurie saugo ir palaiko genetinę įvairovę ir vidinę organizacinę
struktūrą. Antropogeninis poveikis, stipriai keičiantis gamtą, neišvengiamai veikia
populiacijų genetinę struktūrą ir pasireiškia genų dažnių poslinkiu. Be to žmogaus
poveikis landšaftui, keičia atrankos veiksnius, o tuo pačiu ir populiacijų struktūrą.
Nuo XX a. aštunto dešimtmečio stebimas klimato atšilimas bei pasikeitę
antropogeniniai veksniai turi poveikį ir Lietuvos gyvūnų populiacijoms. Pastebėti
migruojančių paukščių rūšinės sudėties pokyčiai, atskridimo-išskridimo laiko
skirtumai, nebūdingas žiemojimas, žuvų rūšinės sudėties, skaitlingumo, biomasės
pokyčiai, vabzdžių dinamikos pokyčiai, erkių pernešamų infekcinių ligų susirgimų
padažnėjimas dėl klimato kaitos.
Populiacijų genetinė struktūra ir jos genetinis kintamumas yra vienas iš
rodiklių, atspindinčių individų pokyčius besikeičiant įvairiam gyvenimo sąlygų
kompleksui ir leidžiančių įvertinti vykstančių pokyčių mastą (Nei 1987, Ridley 2004,
Beebee 2005).
Genetinis kintamumas yra ir individų adaptacijos įvairioms gyvenimo
sąlygoms rodiklis, todėl jo tyrimas gali būti indikatorius ekologinių sąlygų įvertinimui
ir suteikti informaciją apie genofondo kitimą gamtinėse populiacijose (genų dreifą,
migraciją, aplinkos poveikį). Jo įvertinimas taip pat gali suteikti duomenų apie
migruojančių gyvūnų (pvz., paukščių perinčių bendroje teritorijoje, o žiemojančių

skirtingose vietovėse, migruojančių į nerštavietes žuvų) populiacijų genetinę
struktūrą, paukščių pernešamų parazitų kilmės vietą, platinimą; apie introdukuotų ir
reintrodukuotų gyvūnų prisitaikymo mechanizmus.
Nūdienos biomedicinos ir fizikiniuose moksluose tvirtai įsigali
tarpdisciplininė prieiga. Paskutinio XX šimtmečio pabaigoje pasiekti rezultatai
parodė, kad neįmanoma išanalizuoti zoologinių, botaninių, ekologinių, evoliucijos
problemų be genetikos, chemijos, matematikos, informacinių technologijų pasiekimų.
Susiformavo naujos tarpdisciplininės mokslo šakos – molekulinė biologija,
bioinformatika, biomodeliavimas. Molekulinių metodų panaudojimas populiacijų
ekologiniuose tyrimuose šiandien akivaizdus. Terminas “molekulinė ekologija”
pradėtas plačiai naudoti įvairiuose kontekstuose, apimančiuose biogeocheminių ciklų,
biologiškai aktyvių organinių junginių, ar ekotoksikologijos tyrimus po tarptautinio
žurnalo Molecular Ecology pirmo numerio publikavimo 1992 m. Žurnalo redkolegija
nurodo, kad leidinyje publikuojami rezultatai, kurie panaudoja molekulinę biologiją
įvairių ekologijos ar populiacijų biologijos aspektų analizei, ypatingą dėmesį skiriant
natūralių ir introdukuotų populiacijų bei jų aplinkos tyrimams panaudojant
molekulinę technologiją, o taip pat rekombinantinių organizmų (kitaip genetiškai
modifikuotų organizmų, GMO) išleidimo į aplinką studijoms. Šiandieninių požiūriu
molekulinė ekologija apibrėžiama kaip molekulinių genetinių žymenų
panaudojimas tiriant ekologijos ir evoliucijos problemas, apimančias genetinių
ryšių tarp individų, populiacijų ir/ar rūšių tyrimus, įvertinant kaip patelės
pasirenka patinus, populiacijų prisitaikymo prie kintančios aplinkos
mechanizmus, efektyvaus populiacijos dydžio svyravimo vaidmenį adaptacijos
procese, naujų rūšių formavimosi mechanizmus, introdukuotų ir reintrodukuotų
rūšių poveikį vietinėms populiacijoms, rūšims, antropogeninį poveikį populiacijų
struktūrai, skirtingų genotipų adaptacijai.
Nuo 1992 m. leidžiamas tarptautinis žurnalas „Molecular ecology“.

2. TYRIMO METODAI MOLEKULINĖJE EKOLOGIJOJE
Šiandien, dėl žymiai padidėjusios žmogaus veiklos, ekosistemos pastoviai yra
veikiamos įvairių abiotinių ir biotinių veiksnių. Pagrindinis vaidmuo organizmų
prisitaikymui besikeičiančiose ekosistemose tenka konkrečioms organizmų
populiacijoms, jų sąveikai bendrijoje. Vertinant populiacijų kintamumo priežastis ir
dinamiką bei konkrečios organizmų populiacijos genetinės struktūros pokyčius laike
ir erdvėje tam tikroje buveinėje, gali būti panaudoti daugelis metodų leidžiančių
nustatyti individų genotipus. Visus juos galima suskirstyti į keletą grupių: 1)
morfologiniai-biometriniai metodai; 2) citogenetiniai; 3) imunogenetiniai metodai; 4)
biocheminis baltymų polimorfizmas; 5) DNR polimorfizmas.
Kiekybinei genetinio kintamumo charakteristikai bei tyrimo metodams
keliami keletas reikalavimų: 1) fenotipiniai skirtumai, sąlygoti alelių pakitimais
atskirame lokuse, turi būti nustatomi skirtinguose individuose; 2) alelių pakitimai
viename lokuse turi būti atskiriami nuo alelių pakitimų kitame lokuse; 3) žymi dalis
alelių pakitimų atskirame lokuse turi būti atskiriama viena nuo kitos; 4) tiriami
lokusai turi atspindėti atsitiktinę genų imtį pagal fiziologinį pasireiškimą ir
kintamumo laipsnį.

Kokius žymenis pasirinkti?
Pagrindinė populiacijų biologams iškylanti problema yra rasti tinkamiausius metodus
(Ecology: vol.79(2), 1998 p.372, table 3), kurie patikimai tiksliai atskleistu genetinį
kintamumą išsprendžiant konkrečias problemas, susijusias su:
• mažiausiu mėginių kiekiu,
• tyrimų išlaidos (santykinė kaina) ir
• pavojus sveikatai
Priimtiniausi yra tie metodai, kurie leidžia nustatyti
• Didelį alelių skaicių viename lokuse ir/arba
• Daug lokusų su dviem ar daugiau dažnai aptinkamais aleliais
Morfologinių požymių analizė tenkina pirmą ir antrą kriterijų, tačiau kitos
netenkinamos, kadangi tik dalis alelinių pakitimų sukelia morfologinius pakitimus.
Citogenetiniai metodai, plačiai taikomi antropogeninio poveikio
mutageniškumui įvertinti įvairiuose populiacijose, atspindi tik labai ryškius
chromosominius pakitimus – genomines ir chromosomines mutacijas, atsirandančias
dėl labai stipraus mutageninio poveikio organizmams. Nestiprūs veiksniai, sąveika
tarp skirtingų genotipų bendrijoje nėra registruojama.
Nors imunogenetiniai metodai plačiai taikomi gyvūnų selekcijoje (kaip
genetiniai žymenys), evoliucinių ryšių nustatyme, tačiau vien tiktai kraujo grupių
analizė neduoda pilnos informacijos apie populiacijos kintamumo laipsnį, kadangi
monomorfinių lokusų skaičius tokio tyrimo atveju lieka neaiškus.
XX a. sparčiai vystantis molekulinei biologijai ir atradus baltymų ir DNR
polimorfizmą, pastarieji du metodai padarė perversmą populiacijų ekologijoje,
sudarydami galimybę įvertinti rūšių ir populiacijų genetinį kintamumą laike ir
erdvėje, tirti rūšis, paveiktas ne tik natūralių gamtinių sąlygų, bet ir žmogaus veiklos
padarinių, neišimant individų iš gamtinės aplinkos, kas ypač svarbu tiriant retas ir
nykstančias rūšis.
2. 1. Baltymų polimorfizmas
Baltymų polimorfizmo analizė paremta baltymų elekrtocheminėmis
savybėmis: baltymų (taip pat ir fermentų) molekulių mišinys migruodamas gelyje ir
veikiamas elektrinio lauko išsiskiria į frakcijas priklausomai nuo krūvio, molekulinės

masės ir baltymų dydžio, kurį sąlygoja organizmo genotipas. Įvairių organizmų
audinių pavyzdžiai, norint išlaisvinti fermentus ir kitus baltymus iš ląstelių,
homogenizuojami (susmulkinami) įvairiuose ekstragavimo buferiuose,
centrifūguojami ir supernatanto (tirpios frakcijos) mėginiai užnešami ant gelių.
Šiandien elektroforezėje panaudojami ivairiūs tvirti nešėjai – popierius, agaras,
agarozė, krakmolas, poliakrilamidas, acetatceliuliozė ir kt. geliai. Po to, per gelį keletą
valandų leidžiama pastovi elektros srovė, kurios veikiami baltymai juda gelyje.
Judėjimo greitis ir kryptis priklauso nuo pasirinkto elektroforezės metodo bei pačių
baltymų elektrocheminių savybių. Po elektroforetinio frakcionavimo, panaudojus
specifinius histocheminius fermentų dažymo metodus, geliuose vizualiai galima
nustatyti izofermentų išsidėstymą.
Izofermentai arba jų subvienetai tarnauja kaip žymuo savo genui. Šiuo metu
biocheminės polimorfinės baltymų sistemos plačiai naudojamos molekulinės
ekologijos laboratorijose dėl kelių patogių priežasčių:
1) izofermentus galima nustatyti mažame ekstrakto kiekyje;
2) tuo pačiu metu ir tokiomis pat sąlygomis galima analizuoti daug mėginių;
3) tyrimo metodika standartizuota daugelyje pasaulio laboratorijų;
4) nereikia daug laiko tiriamų pavyzdžių analizei;
5) aleliniai genai vienas kito nemaskuoja – išryškėja kodominantiškai;
heterozigotose dalyvauja abu tipai subvienetų, homozigotose viena;
6) galima išskirti nedideles mutacijas, nepastebimas vizualiai;
7) galima analizuoti iškart keletą atskirų lokusų, t.y. keletą skirtingų požymių;
8) analizei reikalingi palyginti nebrangūs reagentai.
2.1.1. Izofermentai
Genetiniame lygmenyje populiacijos yra pakankamai stabilios sistemos, kurių
pastovumą užtikrina genetinės įvairovės optimumas, bet koks nuokrypis – įvairovės
sumažėjimas ar padidėjimas, sukelia nepalankias biologines pasekmes ir gali baigtis
populiacijos degradacija. Įprastinėje aplinkoje optimali genetinė įvairovė palaikoma
veikiant populiacinės dinamikos faktoriams – natūraliai atrankai, genų mutacijoms,
migracijai.
Genetinis baltymų polimorfizmas sudaro dalį genetinės įvairovės, stebimos
skirtingų rūšių gamtinėse populiacijose. Vykstant neutralioms mutacijoms alelinuose
genuose, formuojasi daugialypės baltymų formos. Kiekvienas individas tik jam

būdinga, unikalią baltymų sudėtį, todėl individų atsitiktinės imties polimorfizmo
tyrimai atspindi genetinio kintamumo procesus, kurie vyksta visoje populiacijoje.
Izofermentai gali būti panaudojami populiacinėje ir evoliucinėje genetikoje, taip pat
tiriant morfogeninius procesus, izofermentų spektro ekspresijos genetinius
mechanizmus (Glazko, 1985). Tiriant izofermentus galime:
1) Įvertinti vidurūšinį ir tarprūšinį genetinį kintamumą;
2) Išaiškinti filogenetinius ryšius tarp skirtingų taksoninių grupių;
3) Įvertinti genetinio kintamumo kiekį formuojantis rūšiai;
4) Tyrinėti mikroevoliucinio proceso pokyčius
Izofermentai – genėtinai žymenys. Bet kuris polipeptidas yra žymuo jį
koduojančiam genui, t.y. jei aptinkami kurio nors tipo subvienetai, tai tas genas yra
aktyviai ekspresuojamas. Populiacinės genetikos tyrimams dažniau pasirenkamos
izofermentinės sistemos, kadangi izofermentus lengviau aptikti dėl jiems būdingo
specifinio katalitinio aktyvumo. Juos patogu naudoti, nes izofermentus galima
nustatyti mažame ekstrakto kiekyje ir tuo pačiu analizuoti daug mėginių vienu metu
(Paulauskas ir kt., 2002).
Taip pat svarbu paminėti, kad aleliai išryškėja kodominantiškai t.y.
heterozigotų atveju vienas kito nemaskuodami. Heterozigotose dalyvauja abu
izofermentų subvieneto tipai, homozigotose tik vienas. Apie genotipą galime spręsti
pagal fenotipų požymį – izofermentų spektrą. Kodominavimas palengvina
elektrofotogramų išaiškinimą (iššifravimą), nes taip ypatingai patogu dirbti su
laukinėmis rūšims, kada hibridiologinė analizė yra apsunkinta arba visai neįmanoma.
Svarbus aspektas, kuris teikia izofermentams pranašumą, tai laikas, kadangi
tiriamų pavyzdžių analizė nereikalauja daug laiko bei rezultatai gaunami greitai bei
yra lengvai įvertinami, nereikia išskyrinėti DNR. Izofermentai leidžia aptikti
nedidelius genų mutacijas, nepastebimas vizualiai ir tuo pačiu metu analizuoti keletą
atskirų genų lokusų.
Šis metodas duoda pilna informaciją apie populiacijos kintamumo laipsnį.
Tačiau kaip ir kiekvienas metodas ir šis turi savotiškų trūkumų. Izofermentai pasižymi
mažu gausumu bei žemu polimorfizmo lygiu. Jų spektrus kartais sudėtinga
interpretuoti dėl susidarančių kompleksų, kurie atsiranda dėl genų poliploidijos arba
duplikacijos, bei integralinių heterodimerų susiformavimo.
Dirbant su izofermentais galimi netikslumai, kurie atsiranda pakitus
izofermentų sudėčiai skiriančioje terpėje. Be to, imant pavyzdžiui iš įvairių audimų ir

juos homogenizuojant, gali išsiskirti fiziologiškai aktyvios medžiagos, kurios sukelia
ryškius pakitimus tiriamo fermento molekulėje.
Analizuojant izofermentus elekroforeziniais metodais išryškinami tiktai tokie
aleliai, kurie sąlygoja izofermentų, besiskiriančių elektroforeziniu judrumu. Dalis
pasikeitusių aminorūgščių gali keisti polipeptido elektrinį krūvį netiesiogiai keičiant
fermento antrinę, tretinę arba ketvirtinę struktūrą. Taigi izofermentai parodo tik dalį
egzistuojančio biocheminio polimorfizmo. Be to, tas pats elektroforezinis alelis gali
turėti keletą genetinių variantų (Žukas, 2005).
Baltymų polimorfizmo analizė paremta baltymų elektrocheminėmis
savybėmis: baltymų (taip pat fermentų) molekulių mišinys, migruodamas gelyje ir
veikiamas elektrinio lauko, išsiskiria į frakcijas priklausomai nuo krūvio, molekulinės
masės ir baltymų dydžio. Gauti rezultatai vadinami elektromorfais. Tai pastebėjo XX
amžiaus trečiajame dešimtmetyje A. Tisel. Priklausomai nuo subvienetų skaičiaus,
baltymai elektroforezės metu gali formuoti keletą elektromorfų tipų. Homozigotų
baltymai, koduojami vienodų vieno lokuso alelių, formuoja vieną frakciją.
Kartais elektromorfai, vienodai judantys gelyje, gali būti koduojami skirtingų
alelių, nes: 1) genetinis kodas yra išsigimęs – viena amino rūgštis gali būti koduojama
keliomis nukleotidų sekomis; 2) pakeitus kai kurias amino rūgštis, elektroforezinis
viso baltymo judrumas gali nepasikeisti; 3) gali atsirasti netikslumų dėl baltymo
sudėties pokyčių ruošiant medžiagą.
2.1.2. Izobaltymai
Polimorfizmas būdingas ne tik fermentams, bet ir daugeliui nekatalitinių
baltymų Hemoglobinas, mioglobinas, feritinas, aktinas ir miozinas turi po keletą
izoformų, kadangi yra oligomerai, jie sudaryti iš dviejų arba daugiau subvienetų tipų.
Nefermentinių baltymų daugybinės formos ištirtos mažiau nei fermentai, kadangi dėl
specifinių katalitinių savybių fermentus lengviau aptikti grubiuose ekstraktuose
(Žukas, 2005). Heterogeniškumo atveju, izobaltymams būdingos tos pačios genetinės
ir negenetinės priežastys. Kadangi baltymų polimorfinėms sistemoms būdingas
nesudėtingas išskyrimas, stabilumas, informatyvumas ir paprasta genetinė kontrolė,
jos plačiai naudojamos populiaciniuose tyrimuose.
Nespecifinio baltymo elektromorfuose išsiskiria makroglobulinai,
postransferinai, transferinai, pretransferinai, postalbuminai, albuminai ir
prealbuminai. Žmogaus ir gyvūnų kraujo plazmoje elektroforezės būdu geriausiai
išsiskiria albuminai, α1 - ,α2 -, β1-, β2- ir γ – globulinai ir transferinai.

Albuminai sudaro 54-58 % visų plazmos baltymų. Jie dalyvauja įvairių
medžiagų, turinčių hidrofobines savybes transporte bei osmosinio slėgio reguliacijoje
(Шубин, Ефимцева, 1988). Jų polimorfiškumas yra aprašomas remiantis daugeliu
naminių gyvūnų tyrimais: arklio, didelių raguotų galvijų, ožkų ir kiaulių. Baltymą
formuoja viena polipeptidinė grandinė.
Transferinai išskirti iš daugelio organizmų yra polimorfiniai, kontroliuojami
autosominio lokuso polialelinės sistemos.
2.1.3. Baltymų elektroforezė ir jos metodika
Elektroforezė yra vienas iš svarbiausių baltymo polimorfiškumo tyrimo
metodų. Elektroforezės tyrimų pradininku laikomas švedų mokslininkas Tisel (1937
m.), kuris nustatė, kad panaudojus elektroforezę laisvame tirpale kraujo serumo
globulinus galima išskaidyti į keletą frakcijų – elektromorfų. Elektroforezės metodas
pirmą aprašytą tiriant esterazių heterozigotiškumą (Market, Hunter, 1957). Krakmolo,
poliakrilamido gelių panaudojimas, histocheminio dažymo metodikos įsisavinimas
atvėrė plačiai duris augalų bei gyvūnų genetinio kintamumo tyrimui ir įvertinimui ne
tik tarp atskirų rūšių bet ir rūšies viduje. 1966 metais Harry, Johson, o ypač Lewontin
ir Hubby publikacijos elektroforezės srityje, tapo bendru metodu gamtinių populiacijų
struktūrai .
Tyrinėjant gamtinių populiacijų genetinę struktūrą, žymenimis naudojami
įvairūs baltymai ir fermentai, elektrofotogramose matomi histochemiškai dažant
gelius po elektroforezės. Baltymai, kaip ir juos sudarančios amino rūgštys, yra
amfoteriniai elektrolitai (gali būti anijonais ir katijonais priklausomai nuo pH). Dėka
baltymo dalelių judėjimo elektros lauke baltymų mišinius galima frakcionuoti. Per
tirpalą leidžiant elektros srovę, įelektrintos molekulės bei jonai juda elektrodo,
turinčio priešingą elektros krūvį link. Judėjimo greitis priklauso nuo judančios dalelės
krūvio dydžio, jos formos ir molekulinės masės, tirpiklio savybių, temperatūros ir kitų
veiksnių (Glemža, 1987). Dėl šių skirtumų elektrinio lauko veikiamos elektringos
dalelės tirpale yra atskiriamos. Tai ir sudaro elektroforezės pagrindą. (Mickevičius,
1999)
Prie kitų veiksnių, veikiančių judrumą priklauso :
• Elektrinis laukas. Jo įtaka dalelių atskyrimui priklauso nuo nuolatinės
srovės stiprumo, įtampos ir varžos. Jonų pernešimo greitis yra
proporcingas srovės stiprumui, o nueitas kelias – srovės laidumo laikui.

Migracijos greitis yra atvirkščiai proporcingas varžai. Aukštos įtampos
naudojamos žemamoliniams junginiams frakcionuoti. Išskiriant šilumai šie
dydžiai kinta, todėl būtina šaldymo sistema.
• Buferis palaiko ir stabilizuoja pH, įtakodamas medžiagos migracijos greitį.
Pradinės buferinių tirpalų savybės: joninė jėga, pH ir buferinė talpa.
Padidėjus joninei jėgai, keičiasi ir suminė srovė, todėl padidėja ir
išskiriamos šilumos kiekis. Jei naudojame buferines sistemas su žema
jonine jėga, srovė ir išsiskiriantis šilumos kiekis sumažėja, bet padidėja
difuzija. Labai svarbu įvertinti buferio pH, nes priklausomai nuo jo
keičiasi judančių junginių kryptis ir dydis. Buferinė talpa pasižymi
didesniu ar mažesniu sugebėjimu neutralizuoti elektrolizės produktus,
susidarančius elektroforezės metu.
• Absorbcija – (pavyzdžio molekulių sulaikymas ant nešėjų) Didžiausia
absorbcija pasižymi popierius, todėl mažėja metodo skiriamoji geba.
• Elekroosmozė – (krūvio atsiradimas tarp buferinio tirpalo vandens
molekulių ir nėšejaus paviršiaus). Iššaukia H30 susidarymą, kuris
pagreitina katijonų judrumą.
Be to, pirminės fermento struktūros pasikeitimas dėl paveldimoje medžiagoje
įvykusios mutacijos, dažnai nors ne visada, įtakoja ir suminį makromolekulės krūvį.
Todėl fermento molekulės, koduojamos skirtingų to paties geno alelių, dažniausiai
elektriniame lauke judės skirtingu greičiu ir užims skirtingas pozicijas po
elektroforezės (Aлтyхob, 1983).
Elektroforezių tipai:
1. Su judama riba – makromolekulės yra visame tirpalo tūryje. Tai analitinis
metodas, naudojamas nustatant baltymų judrumą ir izoelektrinį tašką;
2. Zoninė – tirpalas užnešamas dėmių arba juostų pavidalu. Naudojama
švarumo nustatymui, judrumo ir konformacijos analizavimui, taip pat
gryninimui.
3. Nepertraukiama – pavyzdžių užnešimas nepertraukiamose zonose
Elektroforezė gali būti atliekama skystoje terpėje. Šis metodas pagrįstas
skiriamųjų ribų tarp baltymų ir buferinio tirpalo pastebėjimu ir fiksavimu. Tokiu būdu
nustatoma ar baltymų tirpalas yra vienalytis. Šiuo metodu nustatyta sudėtinga kraujo
baltymų struktūrą ir daugelio fermentų elektroforezinis grynumas.

Tvirtuose nešikliuose: popieriaus, celiuliozės acetato, plonos silikagelio
aliuminio oksido arba celiuliozės sluoksnių, krakmolo, agaro bei poliakilamido.
Bendra visų šių nešiklių ypatybė yra ta, kad atskiriamos medžiagos juda aiškiomis
zonomis ir jose susitelkusias medžiagas po to lengva nustatyti kitais analizės metodais
(Mickevičius, 1999). Dėl gelių cheminių savybių (juose beveik nepasireiškę sorbcija
nei elekroosmozė), baltymų zonos neišsiplečia. Elektroforezė geliuose atliekama
dažniausiai tada, kai reikia analizuoti, o ne preparatyviai aiškinti baltymus (Glemža,
1987).
Agaro gelis dažniausia naudojamas kai reikia analizuoti nedidelius medžiagų
kiekius. Elektroforezė acetilceliuliozėje naudojama klinikiniuose tyrimuose, kai reikia
greitai diagnozuoti kai kuriuos susirgimus, pasireiškiančius eritrocitų ir kraujo serumo
izofermentų spektrų pokyčiais. Elektroforezę krakmolo gelyje kraujo serumo baltymų
analizei pirmasis panaudojo Smith, 1959 metais. Jis patogu naudoti dirbant su
nepatvariais fermentais, kurių aktyvumo išsaugojimas elektroforezės metu susijęs su
tam tikrais sunkumais.
Vienas iš dažniausiai naudojamų gelių yra poliakrilamido, kuri pasižymi
geromis fizinėmis ir cheminėmis savybėmis. Pirmasis šį gelį panaudojo Riamond su
kolegomis 1950 metais. Gelis gaunamas maišant ir polimerizuojant akrilamidą su N,
N′-metil-bis-akrilamidu terpėje, kurioje yra katalizatorius. Katalizatorius , kaip laisvų
radikalų šaltinis, naudojamas oksidacinės-redukcinės sistemos. Pavyzdžiui:
1. Amonio persulfatas -N,N,N,N′- tetrametiletilendiamidas (TEMED)
2. Amonio persulfatas 3 dimetilaminproprio-nitrilas ir kt.
Svarbiausia, kad katalizatorių sistemos neturi veikti nei buferinių tirpalų, nei
gelio elertolaidumo bei klampumo. Gelio klampumas, elastingumas ir tvirtumas
priklauso nuo poliakrilamido polimerizacijos laipsnio (nuo grandinių ilgio) ir nuo
susiuvimo laipsnio, t.y. nuo pridėto N, N′-metil-bis-akrilamido kiekio
Poliakrilamidinis gelis yra stabilus ir inertiškas, labilios struktūros (tai sudaro
galimybę gauti gelius su norimo dydžio poromis). Poliakrilamidinis gelis
neatsorbuojasi, neturi elektooosmozės, atsparus pH ir temperatūros kitimais, netirpsta
daugelyje tirpikliuose.
Atilikus elektroforezę fermentai yra dažomi. Specifiškumą apsprendžia
dažančio tirpalo sudėtyje esantis specifinis analizuojamam fermentui substratas ir
dažančios druskos, kurios reaguoja su fermento kanalizuojamos reakcijos produktu
(Aйaлa, 1984, Harry , Hopkinson, 1977).

Toje vietoje kur tiriamasis fermentas, vyksta cheminė reakcija :
Substratas → Produktas+ druska → spalvota substancija
Ilgesniam gelių saugojimui agaro, acetilceliuliozės ir krakmolo gelio
pavyzdžiai džiovinami, o poliakrilamido laikomi 7% acto rūgštyje po to džiovinami.
Elektroforezės svarbiausi privalumai:
1. Analizuojamas sudėtingas baltymų mišinys. Naudojant specifinius
histocheminius dažymo metodus, nebereikia papildomai jų frakcionuoti
2. Yra galimybė tyrinėt šviežiai išskirtą iš įvairių audinių medžiagą
3. Analizės paprastumas ir gaunamų rezultatų informatyvumas.
Amino rūgščių skirtumų išskyrimas baltymuose priklauso nuo biologinių ir
biocheminių elektroforezės sąlygų, tokių kaip tiriamas audinio ir gelio tipas. Jei ne
visada vienodi visiems individams ir laboratorijoms, kuriose yra atliekama genetinio
kintamumo analizė. Skirtumai sąlygose gali įtakoti kai kuriuos baltymų polimorfizmo
išskyrimo skirtumus (Глазко, 1993).
Tačiau ir baltymų polimorfizmas atspindi tik dalį visų skirtumų DNR
nukleotidų sekose, nes 1) skirtumai tarp sinoniminių kodonų nekeičia koduojamų
aminorūgščių; 2) 90% ar net daugiau DNR, kuriai priklauso intronai ir vieną nuo kito
geno skiriančios nukleotidų sekų dalys, yra netransliuojamos. Todėl paskutiniame
dešimtmetyje plačiai tiriamas DNR polimorfizmas.
2.2. DNR polimorfizmas
Įvertinti DNR polimorfizmo dydį pagal nukleotidus galima keliais būdais: tiesioginiais DNR sekvenavimo metodais nustatant konkrečios genomo dalies nukleotidų sekos įvairovę skirtinguose individuose, ir netiesioginiu pagal DNR restricinių fragmentų ilgio polimorfizmą (RFIP), panaudojus Southern blotingo metodą; itin variabilias DNR sekas, sutrumpintai žymimos VNTR, dar vadinami hipervariabilūs minisatelitai (DNR “pirštų atspaudai”, angl. fingerprints); labai trumpas pasikartojančios sekos STR, pavadintos mikrosatelitais, kurias galima identifikuoti genų introninėse dalyse arba speiserinėje DNR. Hipervariabilių sekų tyrimui pakanka labai nedaug pradinės DNR kiekio, nes panaudojus polimerazinės grandininės reakcijos (PGR) metodą, specifiniai DNR fragmentai pagal atitinkamus pradmenis yra padauginami (amplifikuojama) iki tokio kiekio, kurio užtenka elektroforezinei DNR analizei, nudažius etidžio bromidu ar sidabro nitratu.
Naudojant įvairias restrikcines endonukleazes ir DNR žymes, nustatytas daug didesnis kintamumas nei pagal kraujo grupes, izofermentus ir kitus požymius, paveldimus pagal Mendelio dėsnius.

2.3. Rezultatų įvertinimas
Gautiems molekulinės ekologijos rezultatams įvertinti šiandien plačiai
naudojamas kompiuterinis duomenų apdorojimas. Izofermentinių sistemų analizės
rezultatų apskaičiavimui ir genetiniams panašumams bei skirtumams įvertinti
naudojamos kompiuterinės programos Biosys-1, Biosys-2 ir DNR - PopGene32,
TREECON. Statistiniams duomenų apdorojimams taip pat panaudojamos
kompiuterinės programos PRIMER 5.0, STATISTICA 6.0, taikomos tiek vienmatės
tiek daugiamatės analizės: Kruskalo-Voliso testas; dispersinės analizės (ANOVA,
MANOVA/MANCOVA), diskriminantinė analizė, multidimensinio skirstymo (MDS)
analizė bei RELATE ir 2STAGE analizės – pastarosios naudotos skaičiuojant
koreliaciją tarp dviejų matricų arba tarp daugiau nei dviejų atstumų matricų.
Dendrogramų sudarymui naudojamas UPGMA grupavimo metodas.

3. POPULIACIJOS, POPULIACINĖ GENETIKA
3.1 Populiacijos samprata ir tipai
Populiacija (lot. populiatio, populus – liaudis, minia) – tai vienos rūšies
genetiškai skirtingų individų grupė, užimanti tam tikrą teritoriją arba erdvę, kurioje
jie keičiasi genetine informacija. Populiacija yra elementari rūšies egzistavimo forma.
Populiacijų ekologijos terminą pirmą kartą panaudojo C. Elton (1927) savo
knygoje “Gyvūnų ekologija” teigdamas, kad populiacija yra svarbiausias ekologinis
vienetas, nes populiacijoje gausiai atsiskleidžia rūšies santykiai su aplinka ir
adaptacinės galimybės.
Populiacijų ekologijos (demekologijos) kūrėjais laikomi A. Macfelden (1965)
ir S. Švarc. Jie tvirtino, kad nėra kitos ekologijos, tik populiacijų ekologija, ir kad tik
populiacijų ekologijos pagalba galima išspręsti visų lygių ekologines problemas. Tik
tada, kai E. Odum (1983, 1986) pateikė ekosistemų mokslo koncepciją, tapo aišku,
kad populiacijų ekologija nėra visaapimantis mokslas.
Biologinių sistemų hierarchijoje rūšis yra aukščiau už populiaciją. Kiekviena
rūšis gali sudaryti daug populiacijų. Pavyzdžiui, žmonių populiacijų yra labai daug, o
rūšis viena – Homo sapiens.
Skirtumai tarp tos pačios biologinės rūšies populiacijų nėra dideli. Tos pačios
rūšies skirtingų populiacijų individai skiriasi morfologiškai. Gyvūnai skiriasi kūno
dydžiu, plaukų, akių spalva, augalai – stiebo forma, lapų, žiedų ir vaisių dydžiu,
forma bei spalva. Esminiai yra vidiniai skirtumai, slypintys genofonde. Kiekviena
populiacija turi savo genofondą. To paties genofondo individai yra genetiškai
nevienodi ir individualaus vystymosi metu nevienodai prisitaiko prie aplinkos sąlygų.
Tos pačios rūšies skirtingų populiacijų individai gali tarpusavyje kryžmintis ir keistis
genetine informacija.
Populiacijoje vyksta vienokia ar kitokia panmiksija – įvairių tipų gametų
kryžminimasis populiacijos viduje. Tarp skirtingų rūšių biologinių savybių,
gyvybingumo, sugebėjimo prisitaikyti vienokiose ar kitokiose sąlygose.
Populiacijų tipai nėra griežti. Pagal užimamas teritorijos dydį ir individų
skaičių populiacijos skirstomos į geografines, ekologines ir elementariąsias arba
mikropopuliacijas.

Geografinės populiacijos užima didžiausias teritorijas ir yra didžiausios
individų skaičiumi. Pavyzdžiui, septyntaškė boružė, užimanti didelį arealą Europoje,
Azijoje ir Šiaurės Amerikoje, sudaro keturias geografines populiacijas, kurios skiriasi
aktyvaus periodo trukme. Didžioji zylė sudaro penkias geografines populiacijas, kai
kurios iš jų net tarpusavyje nesikryžmina.
Ekologinės populiacijos – tai vietinės biotopinės populiacijos, gyvenančios
konkrečioje teritorijoje ar erdvėje. Pagal užimamą teritoriją ir pagal individų skaičių
jos daug mažesnės už geografines populiacijas ir silpnai izoliuotos.
Elementariosios arba mikropopuliacijos nėra savarankiška rūšies egzistavimo
forma. Jos skiriasi viena nuo kitos morfologiniais ir fiziologiniais požymiais, tačiau
tarp jų vyksta pilna genetinės informacijos kai俯 a.
Visos populiacijos yra dinamiškos. Kintant populiacijos individų skaičiui ir
užimamos teritorijos dydžiui, populiacijos tipas gali keistis.
Nuolat keliaujančių ar klajojančių rūšių, pavyzdžiui, šiaurės elnių,
populiacijos yra labai didelės, bet jų mažai. Sėslioms rūšims būdinga daug smulkių
populiacijų.
Pagal gebėjimą savarankiškai daugintis populiacijos skirstomos į
nepriklausomas, iš dalies priklausomas, priklausomas, pseudopopuliacijas ir laikinas
populiacijas.
Nepriklausomos arba pastovios populiacijos gali daugintis savarankiškai, be
kitų tos pačios rūšies individų imigracijos.
Iš dalies priklausomos populiacijos gali daugintis savarankiškai, bet individų
imigracija padidina populiacijos gausumą ir gyvybingumą.
Priklausomos populiacijos yra tokios, kuriose gimstamumas nekompensuoja
mirtingumo. Jeigu į tokią populiaciją neimigruoja kitų tos pačios rūšies populiacijų
individai, ji mažėja, degraduoja ir išnyksta.
Pseudopopuliacijos nesugeba savarankiškai reguliuoti savo narių skaičiaus ir
visiškai priklauso nuo individų imigracijos iš kitų populiacijų.
Laikinos arba periodiškos populiacijos susiformuoja iš nuo kitų populiacijų dėl
klimatinių ir mitybos veiksniu poveikio atskilusių individų.
Kiekvieną populiaciją apibūdina šie rodikliai:
• gausumas;
• tankumas;

• vidinė struktūra;
• gimstamumas;
• mirtingumas;
• augimo greitis;
• migracija.
Populiacijos gausumas yra bendras populiacijos individų skaičius jos
užimamoje teritorijoje.
Populiacijos tankumas tai populiacijos individų skaičius jos užimamos
teritorijos ploto (km2, m2, ha) arba erdvės tūrio (m3) vienete. Aukštesniųjų gyvūnų bei
žmonių populiacijų tankumas paprastai vertinamas individų skaičiumi kvadratiniame
kilometre.
Populiacijos gausumą ir tankumą reguliuoja aplinkos veiksniai: klimato
sąlygos, dirvožemio derlingumas, konkuruojančių rūšių poveikis, parazitai, plėšrūnai
ir kt. Vienos populiacijos koncentruojasi nedidelėse teritorijose, pavyzdžiui olandai,
japonai, kitos pasklinda plačiai, užimdamos dideles teritorijas (rusai). Įsisavindamos
naujas teritorijas ir didėdamos individų skaičiumi, populiacijos pretenduoja iš
elementarių pereiti į ekologines.
Populiacijos struktūra priklauso nuo abitinių, biotinių ir antropogeninių aplinkos
veiksnių visumos. Dėl morfologinių, fiziologinių, biocheminių, genetinių procesų,
populiacijos individų lyties, išsidėstymo užimamoje teritorijoje, amžiaus, dauginimosi
ypatumų, konkurencijos susiformuoja sudėtinga vidinė populiacijos struktūra.

3.2. Populiacinė genetika
Populiacinė genetika yra genetikos mokslo šaka, tirianti paveldimumo ir
kintamimo dėsningumus organizmų grupėse, t.y. populiacijose, kurie išreiškiami genų
dažnių paskirstymu ir kitimu įvairiose populiacijose.
Populiacija (lot. populatio – minia, liaudis) – tai istoriškai susiklosčiusi,
vienos biologinės rūšies genetiškai skirtingų individų grupė, gyvenanti tam tikroje
ribotoje rūšies arealo erdvėje ar teritorijoje ir turinti savybių, įgalinančių jai ilgai
egzistuot.
Populiacinėje genetikoje daugiausia dėmesio skiriama lokalaus inbrydingo
grupėms (kuo didesnėms ir geografiškai struktūrizuotoms) – demams, lokaliomis
populiacijomis, Mendelio populiacijoms. Būtent šiose grupėse ir vyksta sisteminiai
alelinių genų dažnių pasikeitimai.
Populiacija, kurioje bet kuris populiacijos narys gali atsitiktinai poruotis su bet
kuriuo kitu populicijos nariu (manoma, kad paprastai jie yra skirtingų lyčių) vadinama
Mendelio populiacija. Aukščiausio rango Mendelio populiacija yra rūšis.
Atsitiktinis kryžminimas – kryžminimas kai poprų susidarymui neturi įtakos
genetinė konstitucija. Asortatyvinis kryžminimas – porų susidarymą sąlygoja
genotipas.
Vienos biologinės rūšies individų grupė, gyvenanti tam tikroje ribotoje rūšies
arealo erdvėje ar teritorijoje dar vadinama lokalinė populiacija.
Organizmų pasiskirstymas rūšies areale nera vienalytis – organizmai jungiasi į
šeimynines grupes (po 4-10 individų). 20-40 šeimyninių grupių gyminystės ryšiais
susijungia į demus, o apie 4% kryžminimų įvyksta tarp organizmų iš skirtingų
lokalinės populiacijos demų.
Genofondu vadinama visų populiacijos individų genotipų visuma. Diploidinių
organizmų populiacijos, sudarytos iš n individų, genofondas susideda iš 2n genomų,
t.y. iš 2n kiekvieno lokuso genų ir n porų homologinių chromosomų.

3.3 Populiacijos genetinė struktūra: fenotipų, genotipų, alelių dažnių įvertinimas
populiacijoje
Populiacijos genetinę struktūrą ir genofondo kitimą nusako atskirų lokusų
alelių dažnis arba genotipų dažnis.
Vienetas, kuriuo operuoja populiacinė genetika, yra genas - fizinis vienetas,
kuris reprodukcijos procese yra perduodamas iš tėvų vaikams ir kuris nulemia pačias
įvairiausias organizmo savybes. Genui būdingos kelios alternatyvios savybės,
būsenos. Todėl tokie genai vadinami aleniniais. Molekuliniu požiuriu aleniniai genai
– tai skirtinga nukleotidų seka lokalizuota toje pačioje DNR atkarpoje.
Populiacinėje genetikoje laikoma , kad bet kuriame genetiniame lokuse yra
bent du aleliniai genai (genomas diploidinis). Jei jų nukleotidų seka yra identiška,
laikoma, kad individas pagal šį geną yra homozigotas, jei ne – heterozigotas, o
kartais sudėtinis (compound) heterozigotas.
Fenotipų dažnis – tai konkretaus feno dažnio pasireiškimas konkrečioje
individų grupėje, lygus individų pagal šį feną skaičiaus santykiui su tirtų individų
skaičiumi (n).
Jei žinomas ryšys tarp genotipų ir juos atitinkančių fenotipų, tai pagal
stebimus fenotipų dažnius galima apskaičiuoti genotipų dažnius. Konkretaus
genotipo dažnis randamas padalijus atitinkamų genotipų skaičių imtyje iš viso imties
genotipų skaičiaus n.
Fenotipų dažnis ne visada lygus genotipų dažniui. Jis lygus tik recesyvinių ir
kodominuojančių alelių genotipų dažniui.
Alelinis dažnis – tai tam tikro alelinio geno dažnis konkrečioje individų
grupėje. Bet kurio alelinio geno dažnis (p) tiriamoje individų grupėje yra lygus
dvigubo homozigotų (Hm) pagal šį alelinį geną skaičiaus (kadangi kiekvienas
homozigotas turi du tuos pačius alelinius genus) ir heterozigotų (Ht) pagal šį alelinį
geną skaičiaus sumai padalintai iš dvigubo individų skaičiaus (n) tiriamoje grupėje
(nes kiekvienas individas turi du alelinius genus), t. y.
p=(2 Hm+Ht)/2 n (3.1.1)
Abiejų alelinių genų dažnių suma populiacijoje visada lygi vienetui, t.y. (p+q=1).

Taip nustatytas alelinio geno dažnis yra tik empirinis ir gali skirtis nuo tikrosios šio
geno dažnio reikšmės konkrečioje populiacijoje. Todėl imtyje tiriamų individų
skaičius turi būti pakankamas (ne mažesnis 20).
Retais aleliniais genais vadinami tokie genai, kurių dažnis tiriamoje
populiacijoje yra mažesnis už 0,05. Daugelis retų alelinių genų yra žalingi ir dėl to
populiacijoje dažniausiai palaikomi tik pasikartojančių mutacijų.
Genų ir genotipų dažniai
Genotipų dažnis yra pakankamai pastovus populiacijos požymis, o genų dažniams
neaptinkama ypatinga populiacinė specifika (Benedictis de, 1978)
Genofondų kintamumas gali būti aprašomas tiek genų, tiek ir genotipų dažniais. Imtis
turi būti reprezentacinė visos populiacijos individų atžvilgiu.
Genofondas – visų populiacijos individų genotipų visuma. Diploidiniams
organizmams jis lygus 2N, kur N – individų skaičius. Kiekviename genome yra
užkoduota visa genetinė informacija, kurią tas individas gavo iš vieno iš tėvų.
Populiacijos genofondas – susideda iš 2N kiekvieno lokuso genų ir N homologinių
chromosomų porų, kur N – individų skaičius. Išimtis: lytinės chromosomos ir su
lytimi sukibę genai, kurie, kiekviename heterogametiniame organizme, egzistuoja po
vieną egzempliorių.
Genų dažniai:
A=(2*A) + AB, B=(2*B) +AB – alelių skaičius, išskaičiuojamas iš fenotipų;
Bendras alelių skaičius – 2n, nes kiekvienas individas neša po du vieno alelio
variantus;
Alelio A dažnis : a=A/2n, n – individų skaičius.
Polialelinių sistemų atveju skaičiavimų principas išlieka tas pats: homozigotiniai
individai neša po du tuos pačius alelius, heterozigotiniai – po vieną skirtingų tipų.
Viena priežasčių, kodėl populiacinėje genetikoje dažniau genetinis kintamumas
aprašomas naudojant genų o ne genotipų dažnius yra ta, kad alelinių variantų yra
žymiai mažiau nei genotipų. {jei skirtingų alelių skaičius lokuse yra k, tai įmanomų
genotipų skaičius yra k(k+1)/2} (Altūhov)
Sudėtingumas aprašant paveldimos informacijos visuma atsiranda dėl didelio
segreguojančių lokusų skaičiaus genome. Bet kokiu atveju, vienintelis būdas aprašyti
paveldimą informaciją – alelinių genų dažnių nustatymas kiekvienam lokusui. Žinant
šį dydį ir stebint jo pokyčius laike (erdvėje) atsiranda galimybė įvertinti genetinio
proceso vyksmus populiacijoje veikiant įvairiems išoriniams ir vidiniams faktoriams.
Kodominantinio paveldimumo atveju: genų dažnių įverčių patikimumas priklauso nuo
laukinės populiacijos imties dydžio. Būtinas imties dydis priklauso nuo populiacijos
genetinės struktūros ir turi būti nustatomas preliminarių tyrimų metu.
Tarkim, kad iš N diploidinių individų N1 turi alelį A, N2 – heterozigotos AB, N3 –
homozigotos BB; visų N1,2,3, suma lygi bendram individų skaičiui N. tokiu atveju
bendras genų skaičius yra 2N. kiekviena homozigota AA turi du A genus, kiekviena
heterozigota AB, po vieną A ir B geną. Bendras A genų skaičius populiacijoje A =
2N1+N2, o šio geno dažnumas: pA= (2N1+N2)/2N = (N1+1/2N2)/N. taip pat aprašomas
ir B alelio dažnumas qB. Tuomet q + p = 1. Tokiu pat principu skaičiuojama ir
polialelinėms sistemoms.
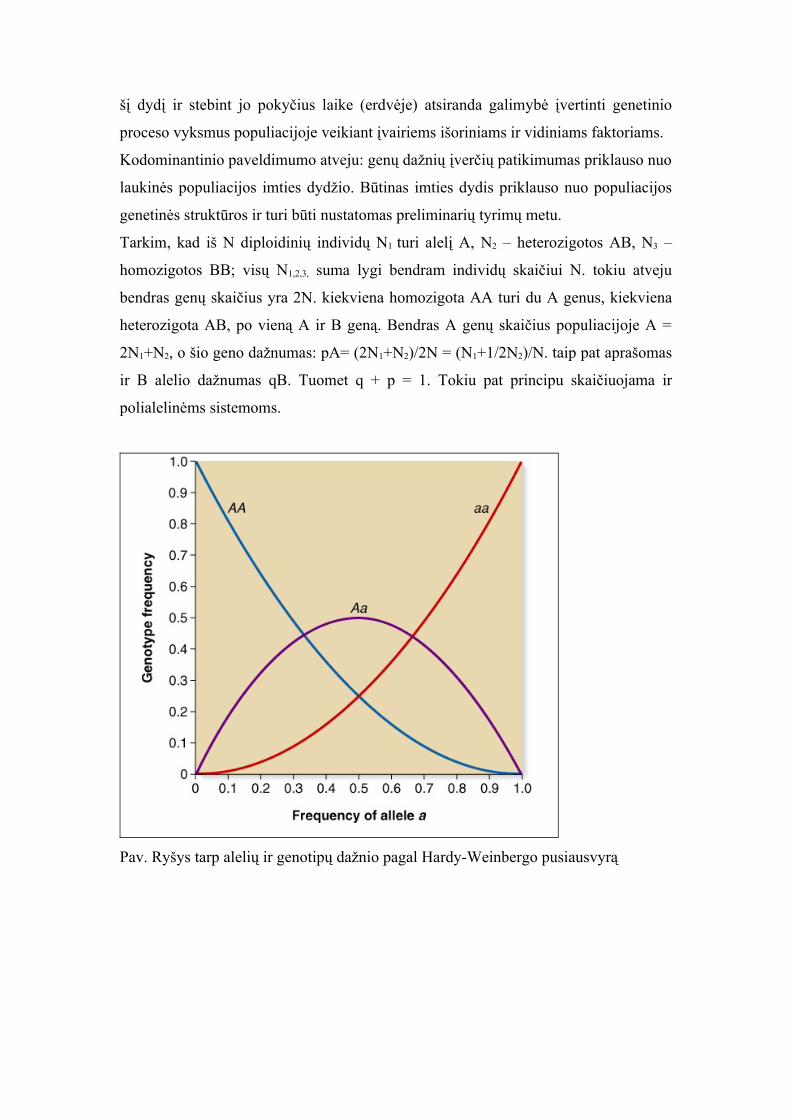
Pav. Ryšys tarp alelių ir genotipų dažnio pagal Hardy-Weinbergo pusiausvyrą

3.4. Populiacijos kintamumas
Genetinis kintamumas yra veiksnys padedantis prisitaikyti prie aplinkos
sąlygų pokyčio. Genetiniai procesai dalyvauja populiacijų dydžio reguliavime ir
kituose ekologiniuose procesuose. Atsiradę pakitimai gali nepasireikšti palikuonių
fenotipe, būti recesyvinėje arba heterozigotinėje formose. Tokiems pakitimams
pasireiškiant fenotipe, galima natūrali atranka, dėl kurios gali susiformuoti ir nauja
rūšys ( Aлтyхob, 1983).
Genetinis kintamumas plačiai paplitęs gamtinėse populiacijose, dėl ko yra palankios
sąlygos evoliuciniam kintamumui. Įvairių organizmų genetinis kintamumas yra
skirtingas (1 lentelė).
1 lentelė. Kai kurių gyvūnų ir augalų genetinis kintamumas gamtinėse populiacijose
(Ayala, 1984)
Organizmai Rūšių skaičius
Vidutinis lokusų
skaičius rūšyje
Vidutinis polimorfizma
s (P)
Vidutinis heterozigotiškumas(H)
Bestuburiai
Drosophila2
82
4 0,529 1,15
Vapsvos 6 15 0,243 0,062
Kiti vabzdžai 4 18 0,531 0,151
Jūrinai bestuburiai1
42
3 0,439 0,124
Sausumos sraigės 5 18 0,437 0,15
Stuburiniai
Žuvys1
42
1 0,306 0,078
Varlagyviai1
12
2 0,336 0,082
Ropliai 9 21 0,231 0,047
Paukščiai 4 19 0,145 0,042
Žinduoliai3
02
8 0,206 0,051
AugalaiSvidulkiniai 1 1 0,231 0,033

2 5Kryžmadulkia
i 5 17 0,344 0,078
Vidurkis
Bestuburiai5
72
2 0,496 0,134
Stuburiniai6
82
4 0,247 0,06
Augalai1
71
6 0,264 0,046
Pagrindinai genetinio kintamumo parametrai yra polimorfizmo lygis (P) ir
vidutinis (faktinis) individų heterozigotiškumas (Ho) bei alelių skaičius lokuse (A).
Šie parametrai naudojami atliekant skirtingų ar pakankamai tarpusavyje nutolusių
gyvūnų taksonominių grupių genetinio kintamumo lyginamąją charakteristiką.
Evoliucijos eigoje dėka mutacijų keičiasi genai ir populiacijoje sutinkami
nevienodoje, o keliose formose. Tokie genai kurie sudaryti daugiau nei iš vieno alelio
vadinami polimorfinais. Polimorfinai aleliai yra apsprendžiami dviejų ar daugiau,
ryškiai besiskiriančių formų koegzistavimą.
Populiacijų analizę galima vykdyti naudojant bet kokį polimorfinių sistemų
kiekį. Aišku, didinant baltymų sistemų kiekį padidėja ir galimybė aptikti skirtumus
tarp populiacijų.
Daug geresnis genetinio kintamumo matas gali būti vidutinis individų
heterozigotiškumas. Skirtingai nuo polimorfizmo šis matas pasižymi tuo, kad jame
nėra laisvumo ir netikslumo. Populiacijos heterozigotiškumas skaičiuojam dviem
etapais. Pirma susumuojama heterozigotus h pagal kiekvieną genetinį lokusą j, o po to
gautą skaičių padalijus iš tirtų individų skaičiaus n bei tirtų lokusų skaičiaus k vidurkį:
Polimorfiniu laikomas toks lokusas, kuriame esančio dažniausiai
pasitaikančio alelinio geno dažnis yra mažesnis už 0,95.
Lokusas, kuriame neaptikta skirtingų alelių nėra polimorfinis, jis vadinamas
monomorfiniu.
Populiacijos polimorfiškumas (polimorfinių lokusų dalis) nustatumas polimorfinių
lokusų skaičių dalijant iš tirtų lokusų skaičiaus k :
P = polimorf. lok. / k

Vidutinis heterozigotiškumas (faktinis) Ho nustatytas sumuojant
heterozigotus h pagal kiekvieną genetinį lokusą j ir gautą skaičių dalijant iš tirtų
individų skaičiaus n bei vidurkinant pagal tirtų lokusų skaičių:
kHo = 1/k ∑ Hj (3.2.1)
j
Hj = h/n (3.2.2)
kur, Hj – heterozigotiškumas j lokuse
Yra teigiama polimorfizmo dydžio ir heterozigotiškumo laipsnio koreliacija. Vidutinė
P reikšmė yra lygi 0,26 + 0,15; vidutinė H reikšmė lygi 0,07 + 0,05. Žmonėms
būdingos tos pačios H ir P reikšmės kaip ir kitiems žinduoliams. Ištyrus europiečius
pagal 87 genetinius lokusus pateikti tokie įverčiai: P = 0,38; H = 0,07.
Genetinio kintamumo įvertinimas.
Tradiciniai genetinės analizės metodai genetinio kintamumo populiacijų lygmenyje
neparodo, todėl vertinant genetinį kintamumą reikia:
- Nustatyti koks yra polimorfinių genų įnašas populiacijoje. Kadangi negalim
ištirti visų lokusų organizme (mes net nežinome tiksliai kiek tų lokusų ten yra), reikia
pasirinkti lokusų imtį, kuri turi būti atsitiktinė. Tokiu atveju rezultatus galima
pritaikyti visos populiacijos mastu. Norint įvertinti polimorfinių lokusų dalį, reikia
išanalizuoti nedidelį skaičių genų, kurie atstovauja nepriklausomą imtį iš visų lokusų
rinkinio. Naudojant tradicinės genetikos metodus tai yra neįmanoma, kadangi geno
buvimo faktas pas individą yra nustatomas atliekant kryžminimą tarp individų
pasižyminčių skirtingu fenotipu pagal to geno koduojamą požymį. Tuomet F1
nustačius kokią dalį užima individai su skirtingais fenotipais, galima išsiaiškinti, kiek
– vienas ar daugiau, genų dalyvauja jo formavime.
Tokiais metodais galima aptikti tik tuos genus kurie pasiduoda genetiniam
kintamumui. Tokiu atveju negalim gauti nepriklausomų genų imties duotajam
genomui, kadangi genai pagal kuriuos nevyksta kintamumas į imtį nepakliūna.

Išeitis atsirado panaudojus molekulinės biologijos metodus. Genetinė informacija
užkoduota DNR struktūrinių genų nukleotidinėse sekose, transliacijos metu
perduodama aminorūgščių sekai, susidaro polipeptidai.
Tyrimams imama baltymų imtis nieko nežinant apie jos populiacinį kintamumą.
Tokia imtis – tai nesimaišanti, visų duotojo organizmo struktūrinių genų, imtis.
Jei vienas ar kitas baltymas yra vienodas pas visus individus, vadinasi šis genas
nepasiduoda populiaciniam kintamumui. Jei stebimi skirtingi baltymo variantai –
genas pasižymi populiaciniu kintamumu.
Šiuo atfveju galima įvertinti ir genetinio kintamumo lygį t.y. nustatyti tiriamo baltymo
variacijų skaičių ir dažnį, kuriuo jos sutinkamos populiacijoje.
Polimorfinių lokusų dalis populiacijoje – P; neigiamos įverčio savybės – negriežtas
pasirinkimas ir netikslumas;
Polimorfinių lokusų skaičius priklauso nuo ištirtų individų skaičiaus. Kad išvengti
imties įtakos P įverčiui taikomi keli kriterijai: 0,95, 0,99 ir pan. Naudojant skirtingus
kriterijus gaunami skirtingi P įverčiai: jei lokusas monomorfinis pagal 95% kriterijų,
tai jis gali būti polimorfinis pagal 99% kriterijų ir pan.
Be viso šito P yra netikslus genetinio kintamumo įvertis – t.y. dėl, to, kad silpnai
polimorfiniai lokusai su labai žemu visų alelių dažniu priskiriami, pagal P, kaip
lygiaverčiai lokusams, kuriuose yra tik keli aleliai, pasižymintys normaliais dažniais.
Pvz.: 1 lokusas – a – 0,95; b – 0,05; 2 lokusas – 20 alelių su dažniais 0,05 . 2 lokusas
yra žymiai polimorfiškesnis, bet pagal 95% kriterijų abu lokusai patampa
lygiaverčiais.
Tikslesnis įvertis – H – heterozigotinių individų skaičius populiacijoje. Nėra
neigiamų P savybių.
Patikimumui H turi būti skaičiuojamas ne mažiau kaip 4 lokusams. {% išraiška}
Tai patikimas kintamumo įvertis, nes atspindi tikimybę, kad dvi duotojo lokuso alelės,
atsitiktinai paimtos iš genofondo, pasirodys esančios skirtingos.
Bet, H neatspindi genetinio kintamumo lygį populiacijose, kuriose vyksta giminingas
kryžminimąsis (“kraujomaiša”). Čia homozigotų bus daugiau nei populiacijose,
kuriose vyksta atsitiktinis kryžmiimąsis, nors alelių dažniai butų tokie patys abiejose
populiacijose. Šito išvengimui skaičiuojamas teorinis heterozigotiškumas, kuris
nustatomas pagal alelių dažnius, darant prielaidą, kad vyksta atsitiktinis
kryžminimąsis.

Įvertinant heterozigotiškumą reikia: 1) ištirti pakankamai didelį skaičių individų,
kad imties dydžiu apsprendžiamas nuokrypis būtų minimalus; 2) ištirti lokusus,
koduojančius skirtingas funkcijas; 3) Tyrimus būtina vykdyti kik galima didesniam
lokusų skaičiuje (15 – 50 lokus ). Tyrinėjant heterozigotiškumą, svarbiausia ne ištirtų
individų skaičius, bet ištirtų lokusų skaičius. Kuo jis didesnis, tuo paklaida būna
mažesnė. Kiekybiškai genetinio kintamumo analizei turi būti ne mažiau kaip 14
lokusų.

3.5 KASTLO - HARDY - WAINBERGO DĖSNIS
1903 m.W. Kastlas (W.E. Castle), o 1908 m. G.H.Hardy ir W.Weinberg’as,
nepriklausomai vienas nuo kito, suformulavo alelių pusiausvyros dėsnį
panmiksinėms populiacijoms, pagal kurį matematiškai įvertinami fenotipų, genotipų ir
alelių dažniai populiacijose:
jei populiacija tenkina panmiksinės populiacijos sąlygas
1. Organizmai tiriamoje populiacijoje yra diploidiniai;
2. Populiacijos reprodukcija vyksta lytiniu keli;
3. Kartos populiacijoje nesutampa;
4. Kryžminimasis populiacijose yra atsitiktinis;
5. Populiacija yra labai didelė;
6. Migracijos poveikis nereikšmingas;
7. Ignoruojamas mutacinio proceso poveikis;
8. Tiriamo genetinio lokuso neveikia natūrali atranka
tai 1) keičiantis kartoms alelinių genų (p ir q) dažnis nesikeičia ir jų suma yra
lygi vienetui
kadangi p+q=1, tai p2+pq=p (p+q)=p (3.3.1)
2) bet kurios kartos genotipų AA, Aa ir aa pusiausvyriniai dažniai
nesikeičia iš kartos į kartą ir yra atitinkamai lygūs p2, 2pq ir q2 ; jų dažnumų
reikšmės gaunamos alelių dažnumų sumą pakėlus kvadratu:
(p+q)2=p2+2pq+q2=1 (3.3.2)
3) genotipų dažnumas pusiausvyrą pasiekia per vieną palikuonių kartą.
Hardy - Wainberg’o- Kastlo dėsningumai teisingi bet kuriam alelių skaičiui k.
(p+q+r)2=p2+2pq+q2+2pr+2rq+r2=1
(p+q+...+z)2=p2+2pq...+q2+2pz+2zq+z2=1
Molekulinė ekologija tiria populiacijų genetinės struktūros išsilaikymo ir kitimo
erdvėje ir laike dėsningumus. H-W taisyklė atspindi panmiksinės populiacijos,
neapribotos individų skaitlingumu ir egzistuojančios ne specifinėje terpėje, genetinės
struktūros stabilumo būseną. Tokioje populiacijoje genotipų proporcijos įgyja
pusiausvyrą jau pirmoje laisvai besikryžminančioje palikuonių kartoje. Kadangi
laisvas kryžminimas tai atsitiktinis gametų apsijungimas, p2+2pq+q2 =1, kai p+q=1.

Jei nėra neigiamų faktorių veikiančių į populiaciją, tai genotipų ir genų dažnių
charakteristikos skaitlingumas išlieka nepakitęs neribotoje kartų kaitoje, t.y. genetinė
dinamika lygi 0. Tai idealus atvejis ir gamtoje praktiškai nesutinkamas, nes visada yra
natūralūs faktoriai (genų dreifas, migracijos, mutacijos, natūrali atranka), kurie išveda
populiaciją iš pusiausvyros ir pažeidžia populiacijos stabilumą. Tai evoliucijos
veiksniai.
Hardy-Wainbergo-Kastlo dėsnio taikymas
A. Genų ir genotipų skaičiavimas, kai ne visi genotipai identifikuojami dėl
dominavimo poveikio
Pvz. Žmogaus albinizmas populiacijoje aptinkamas 1/10 000 dažnumu.
Pagal H-W-K dėsnį genotipo aa dažnumas
q2=0,0001, ⇒ alelio a dažnumas q=(0,0001)1/2=0,01
⇒ alelio A – p=1-0,01=0,99 , o genotipo AA dažnumas p2=0,992=0,98
⇒ genotipo Aa dažnumas 2pq=2x0,99x0,01≈0,02
Išvada: retas alelis populiacijoje egzistuoja heterozigotinėje , o ne homozigotinėje
būsenoje
Jei 2pq / q2 = p / q ⇒ kai q⇒∞ , o p = 1 – q , tai
2pq / q2 = p / q ≈ 1 / q ⇒ vadinasi, kuo mažesnis q, tuo didesnis
heterozigotų skaičius populiacijoje.
Palikuonių kartų skaičius t = 1 / qt - 1 / qo
B. Genų ir genotipų skaičiavimas, kai genai sukibę su lytimi
Homogametinės lyties (moters, drozofilos patelių, paukščių patinų) palikuonių
genotipų dažnumas sutampa su pusiausvyriniais autosominių genų ir genotipų
dažnumais:
Jei A ≡ p ir a ≡ q, tai AA – p2 , Aa – 2pq , aa – q2
Hemizigotinės lyties (vyrų, drozofilos patinų, paukščių patelių) palikuonių genotipų
dažnumas sutampa su genų dažnumais buvusiais pas homogametinę lytį:

A - p ir a - q,
Kai recesyvinis geno dažnumas q, tai hemizigotinio fenotipo dažnumas bus q,o
homogametinio - q2, tada
q / q2 = 1 / q
ir kuo mažesnis q (q⇒∞) , tuo hemizigotinio fenotipo dažnumas bus didesnis.
Išvada: Fenotipai, sąlygoti recesyvinių genų, pas hemizigotas populiacijoje aptinkami
dažniau nei pas homozigametas
Pvz. Daltonizmo genas žmogaus populiacijose paplitęs dažnumu q=0,08.
Kadangi q / q2 = 1 / q = 1/0,08 = 12,5 , todėl vyrai 12,5 karto dažniau serga
daltonizmu nei moterys.
Problemos.
Hemofilijos genas žmogaus populiacijose paplitęs dažnumu q=0,0001. Kokiu
dažnumu hemofiliza pasireikš pas moteris?

4 ELEMENTARŪS EVOLIUCIJOS PROCESAI
Alelių dažnumą keičiantys procesai:
1. Genetinį kintamumą sąlygojantys procesai
a) Mutacijos
b) Rekombinacijos
2. Procesai padedantys genetinį kintamumą perduoti iš kartos į kartą
a) Naturali atranka
b) Migracija
c) Genų dreifas
Genotipų dažnumas keičiasi ir dėl asortatyvinio (neatsitiktinio) porų susidarymo
4.1 Natūrali atranka
Naturali atranka molekulinėje ekologijoje – pagrindinis evoliucijos veiksnys,
iššaukiantis adaptacinius pokyčius populiacinėje struktūroje.
Wt – populiacijos prisitaikymas t.t. laiko momentu.
Wt=Nt/No t.y. prieš tai buvusios kartos ir ateinančios kartos skaitlingumų santykiai;
Wt>1, populiacija auga;
Wt <1, mažėja;
Wt =1, nekinta
Genetinis populiacijos krūvis (Muller, 1950): silpno letalumo genai sugeba atnešti
populiacijai žymiai didesnį nuostolį, nei mutantiniai genai, pasižymintys stipriu
letalumu. Maksimalų genetinį krūvį viename dialeliniame lokuse populiacija neš
tuomet, kai abi homozigotos letalios; tokiu atveju kiekvienoje kartoje žūva 50%
palikuonių.

Atranka heterozigotų naudai
Atranka kai homozigotos turi žemesnį nei heterozigotos prisitaikymą vadiname
superdominavimu arba heteroze
AA Aa aa viso a dažnumaspradinis zigotų dažnumas
p2 2pq q2 1 q
Prisitaikymas, w 1-s 1 1-t
kiekvieno genotipo indėlis į F1
p2(1-s) 2pq q2(1-t) 1-sp2-tq2
normalizuotas dažnumas
p 2 (1-s) /(1-sp2-tq2)
2 pq /(1-sp2-tq2)
q 2 (1-t)/ (1-sp2-tq2)
1 q1 = ( q -t q 2 )/ (1-sp2-tq2)
Alelio dažnumo pasikeitimas:∆q = pq(sp-tq)/(1-sp2-tq2)
Gamtinė atranka heterozės atveju sąlygoja stabilios polimorfinės pusiausvyros
susidarymą
∆q = 0 , kai pq(sp-tq) = 0
bet kai populiacijoje egzistuoja du aleliai, t.y. p ir q ≠ 0 , pusiausvyra nusistovi kai
sp=tq ⇒ s(1-q) = tq ⇒ s=q(s + t)
q=s/(s+t) ir p=t/ (s+t)
Pusiausvyriniai dažnumai heterozės atveju apsprendžiami santikinėmis atrankos
koeficientų reikšmėmis, bet ne apsoliutinėmis
Pvz. Žmogaus heterozė pjautuvinės anemijos atvejuHbAHbA AbAHbS HbSHbSMaliarijai imlūs maliarijai atsparūs žūsta nesulaukę
lytinės brandosw 0,88 1 0,13
IŠVADA: genotipų prisitaikymas priklauso nuo aplinkos sąlygų
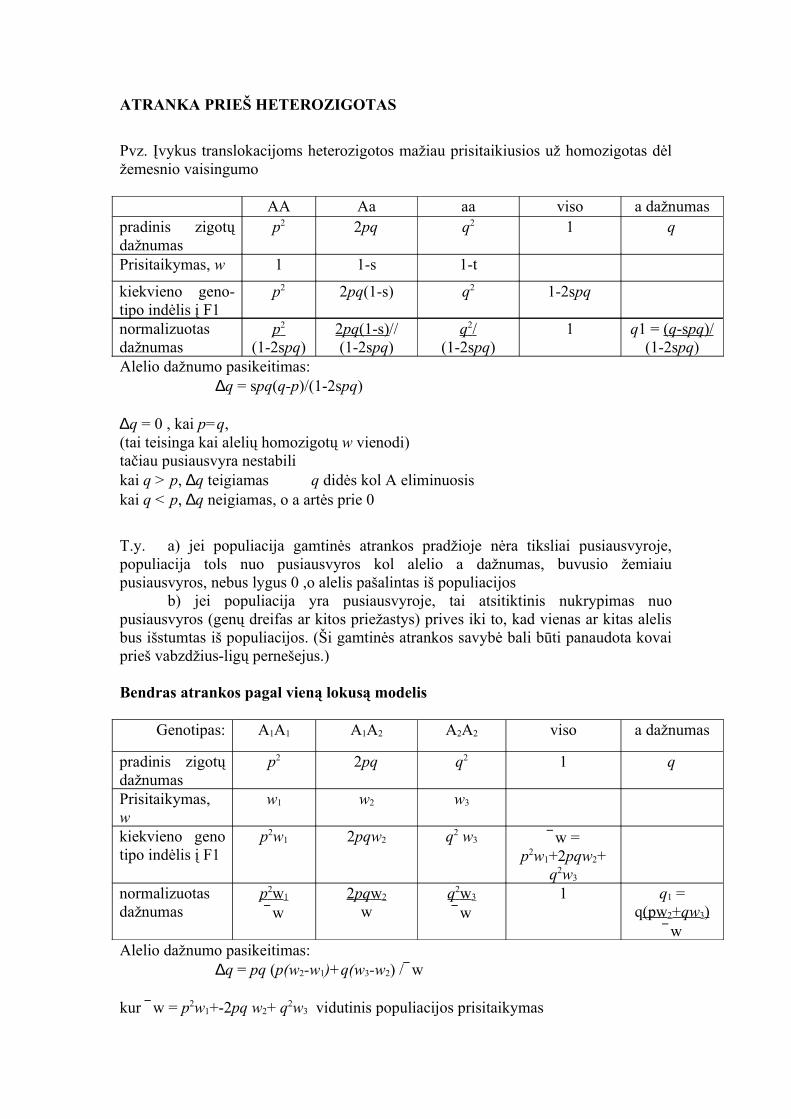
ATRANKA PRIEŠ HETEROZIGOTAS
Pvz. Įvykus translokacijoms heterozigotos mažiau prisitaikiusios už homozigotas dėl žemesnio vaisingumo
AA Aa aa viso a dažnumaspradinis zigotų dažnumas
p2 2pq q2 1 q
Prisitaikymas, w 1 1-s 1-t
kiekvieno geno- tipo indėlis į F1
p2 2pq(1-s) q2 1-2spq
normalizuotas dažnumas
p 2 (1-2spq)
2 pq (1-s) //(1-2spq)
q 2 / (1-2spq)
1 q1 = ( q -s pq )/ (1-2spq)
Alelio dažnumo pasikeitimas:∆q = spq(q-p)/(1-2spq)
∆q = 0 , kai p=q, (tai teisinga kai alelių homozigotų w vienodi)tačiau pusiausvyra nestabilikai q > p, ∆q teigiamas q didės kol A eliminuosiskai q < p, ∆q neigiamas, o a artės prie 0
T.y. a) jei populiacija gamtinės atrankos pradžioje nėra tiksliai pusiausvyroje, populiacija tols nuo pusiausvyros kol alelio a dažnumas, buvusio žemiaiu pusiausvyros, nebus lygus 0 ,o alelis pašalintas iš populiacijos
b) jei populiacija yra pusiausvyroje, tai atsitiktinis nukrypimas nuo pusiausvyros (genų dreifas ar kitos priežastys) prives iki to, kad vienas ar kitas alelis bus išstumtas iš populiacijos. (Ši gamtinės atrankos savybė bali būti panaudota kovai prieš vabzdžius-ligų pernešejus.)
Bendras atrankos pagal vieną lokusą modelis
Genotipas: A1A1 A1A2 A2A2 viso a dažnumas
pradinis zigotų dažnumas
p2 2pq q2 1 q
Prisitaikymas, w
w1 w2 w3
kiekvieno geno tipo indėlis į F1
p2w1 2pqw2 q2 w3 w = p2w1+2pqw2+
q2w3
normalizuotas dažnumas
p 2 w 1
w2 pq w 2
wq 2 w 3
w1 q1 =
q(pw2+ qw 3)w
Alelio dažnumo pasikeitimas:∆q = pq (p(w2-w1)+q(w3-w2) /w
kur w = p2w1+-2pq w2+ q2w3 vidutinis populiacijos prisitaikymas

Atranka priklausanti nuo genotipų dažnumo
Taip pat palaiko ir atveda populiaciją prie stabilaus genetinio polimorfizmo
Gamtinė atranka priklauso nuo genotipų dažnumo, kai genotipų prisitaikymas keičiasi
priklausomai nuo jų dažnumo
1) Pw atranka didelė kai genotipas retas ir maža, kai genotipas labai paplitęs
populiacijoje. Jei egzistuoja dažnumas kai abiejų alelių genotipų w vienodas, bus
pasiekta subalansuota popimorfinė pusiausvyra, net nesant heterozei.
Pvz. pupelės Phaseolus lumetus trijų genotipų SS, Ss, ss prisitaikymas w kečiasi iš
kartos į kartą, keičiantis genotipų dažnumui. WSs
2) Nuo dažnumo priklausanti lytinė atranka atsiranda kai kryžminimosi
tikimybė priklauso nuo genotipo dažnumo. (Pvz. Šiaurės tautelių papročiai).
Viduržemio vyrams labiau patinka blondinės, skandinavams – briunetės: pirmenybė
teikiama retų (populiacijoje) alelių nešiotojams
Išvada: a) nuo dažnumo priklausanti atranka retų genotipų naudai yra vienas iš
mechanizmų palaikančių genetinį polimorfizmą populiacijose, kadangi genotipo
prisitaikymas w didėja, mažėjant genotipo dažnumui;
b) nuo dažnumo priklausanti lytinė atranka gali būti ypač reikšminga esant
migracijai, nes nauji genai, atnešti į populiaciją, išlaikomi.

4.2 MUTACIJOS IR MIGRACIJOS
Mutacija – nekryptingi atsitiktiniai genetinės medžiagos pokyčiai, vykstantys
spontaniškai arba veikiant ypatingiems fiziniams, cheminiams ar biologiniams
faktoriams. Spontaninis atsiradimas – tai pagrindas naujoms alelėms, padidinančioms
populiacijos genetinį kintamumą.
Dauguma atsirandančių mutacijų yra žalingos organizmui (Muller, 1950), bet naujai
atsiradusi alelė, nedaranti žalos heterozigotinėje būsenoje, palaipsniui gali įsitvirtinti
rūšies genofonde. Homozigotinių žalingų alelių atveju veikia natūrali atranka.
Vyksta ir neutralios mutacijos ir šis procesas gali nukreipti populiacijos struktūrą nuo
HdyWnb pusiausvyros (Kimura, 1985).
Naujai atsiradusios mutacijos praradimas – negrįžtamas procesas ir jos neįsitvirtina
populiacijoje.
Mutacijos nuolat, kiekvienoje kartoje atsiranda iš naujo, ir neatmetama tikimybė, kad
taip vadinamos naujos mutacijos nebuvo atsiradusios anksčiau.
Kuomet alelio dažnis populiacijoje yra mažas, jo mutavimo į kitą aleli aptikti
praktiškai neįmanoma.
Analizuojant mutacijų įtaką į populiacijų genetinę pusiausvyrą, reikia atsižvelgti į tai,
kad:
1. tiesioginių genetinių mutacijų greitis yra viena eile didesnis nei grįžtamų
2. nors mutacijų tempas yra skirtingas kiekviename lokuse, tačiau alelio A virtimo į
a greitis yra pakankamai žemas – vidutiniškai 10-5-10-6 eilės (genui/per karta).
Migracijos – labai svarbus populiacinės dinamikos faktorius, kadangi kiekviena
populiacija gamtoje egzistuoja ne tik pati sau, bet taip pat sąveikauja ir su kitomis
populiacijomis (nebent yra pilna izoliacija). Vyksta genų kaita. Jei inmigrantai skiriasi
genetiškai, jie gali iššaukti atitinkamus alelių dažnių pokyčius jau pirmoje kartoje. Jei
genų imigracija ir emigracija yra vienodoi intensyvumo, populiacijoje nusistovi
stacionarus procesas.

4.3 ATSITIKTINIS GENŲ DREIFAS.
Atsitiktinis genų dreifas tai stochastinis genų dažnių pokytis sekančiose kartose
vykstantis dėl riboto, bet kurios realios populiacijos skaitlingumo.
Alelių dažnių kitimas, vykstant kartų kaitai, iššaukiamas atsitiktinių priežasčių, pvz.:
mažu populiacijos skaitlingumu. Tai visiškai atsitiktinis procesas, kuris priklauso t.t.
reiškinių klasei, vadinamai imties klaida. Kuo mažesnė imtis, tuo didesnė paklaida,
t.y. kuo mažiau individų kryžminasi tarpusavyje, tuo daugiau pokyčių, nulemiamų
genų dreifo, vyks alelių dažniuose. Kuo didesnis skaičius individų dalyvauja sukuriant
kitą kartą, tuo artimesnis teoriniam yra alelių dažnis.
Čia svarbiausia yra efektyvioji populiacijos dalis, kuri ir duoda pradžią sekančiai
kartai.
Kadangi alelių dažnių pokyčiai vyksta visomis kryptimis, tendencija alelio dažnio
sumažėjimui/padidėjimui visada gali pakisti į priešingą pusę, kol alelio dažnis
nepasiekia 1 arba 0. Jei alelis prarandamas (0) arba fiksuojamas (1), procesas baigiasi.
Alelių dažnis nebekinta tol, kol dėka atsitiktinės mutacijos neatsiranda naujas alelis.
Jei žinom tėvinių individų skaičių bazinėje populiacijoje ir alelių dažnį jame, galim
išskaičiuoti tikimybė gauti vienus ar kitus alelių dažnius F1 kartoje. Variansa –
kintamumo įvertis, gaunamas lyginant dvi imtis. Jei yra du aleliai su dažniais p ir q,
tėvinių individų skaičius lygus N (bazinės populiacijos genų skaičius bus 2N), tuomet
alelių dažnių variansa pirmoje kartoje: s2=pq/2N, o standartinis nuokrypis s – šaknis iš
pq/2N. šios formulės atspindi atvirkštinė priklausomybę tarp imties dydžio (2N) ir
teoriškai tikėtino alelių dažnio kintamumo.
Kumuliatyvinis efektas – atsitiktinio genų dreifo metu atsirandančių pokyčių kaupimo
efektas, kuris veda prie to, kad aleliai fiksuojasi populiacijose.
Tuo atveju kai alelių dažnių duotajame lokuse neveikia jokie kiti procesai (mutacijos,
migracijos ar atranka), evoliucija priveda prie to, kad vienas alelių bus fiksuojamas, o
visi kiti alternatyvūs – eliminuojami. Jei populiacijoje vyksta tik genų dreifas, tai
tikimybė kad duotasis alelis galiausiai fiksuosis yra lygi to alelio pradiniam dažniui.
Jei įvyksta naujo alelio atsiradimas (mutacija) tai jo dažnis 1/2N ir tai tokia tikimybė,
kad jis kada nors įsitvirtins toje populiacijoje.
Vertinant atsitiktinį genų dreifą labai svarbu įvertinti Ne – genetiškai efektyvią
populiacijos dalį.

Homozigotų sukaupimas vykstant genų dreifui parodo jo tiesioginį ryšį su inbrydingu,
t.y. apribotoje skaitlingumu populiacijoje yra sukaupiama neatsitiktinė gametų
asociacija. Toks nuokrypis nuo panmiksijos – stiprėja; išauga “kraujo” giminystės
lygis tarp populiacijos individų. {eiti į inbrydingą}.

4.4 INBRYDINGO KOEFICIENTAS
Hardy-Weinbergo dėsnis teisingas tik tuo atveju, kai vyksta atsitiktinis
kryžminimasis. Jam nesant, vyksta asortatyvinis kryžminimasis, t.y. individai su
tam tikrais genotipais tarpusavy susieina dažniau, nei tikėtina atsitiktinio
kryžminimosi atveju. Asortatyvinis kryžminimas nekeičia genų dažnių, bet keičia
genotipų dažnį. Asortatyvus kryžminimasis vyksta tada, kai dažniau kryžminasi
panašių fenotipinių bruožų individai Disasortatyvus kryžminimasis vyksta tada, kai
tarpusavyje dažniau kryžminasi skirtingų fenotipų individai
Kryžminimasis ir genotipai
Inbrydingas yra genetiškai susijusių individų kryžminimasis
Outbrydingas yra genetiškai nesusijusių individų kryžminimasis
Nesant kitų evoliucijos veiksnių, inbrydingas ar outbrydingas nekeičia alelių dažnio,
tačiau dėl tokio kryžminimosi yra pažeidžiamas genotipų balansas ir jis nebeatitinka
Hardy-Weinbergo pusiausvyros
Labai įdomi asortatyvinio kryžminimo forma – inbrydingas: kryžminimąsis
giminingų individų tarpe yra dažnesnis nei galima tikėtis atsitiktinio kryžminimosi
atveju.
Kadangi giminingi individai yra labiau genetiškai panašūs tarpusavyje, nei
negiminingi, tai inbrydingas veda prie homozigotų padidėjimo ir heterozigotų
sumažėjimo, lyginant su teoriniais duomenimis atsitiktiniam kryžminimui, nors ir
nekeičia alelių dažnio.
Inbrydingas padidina žalingų recesyvinių alelių pasireiškimo tikimybę.
Inbrydingo koeficientas – F – tikimybė, kad kurio nors individo duotame lokuse
atsiras dvi alelės, identiškos pagal savo kilmę (t.y. tikslios protėvinio alelio kopijos).
Dvi alelės, identiškos pagal savo struktūrą, nebūtinai yra identiškos ir pagal kilmę, ir
gali būti paveldėtos iš protėvių, neturinčių jokių giminystės ryšių.
Jei kiekvienoje kartoje naudojamas vienas ir tas pats inbrydinio kryžminimo tipas, tai
inbrydingo koeficientas auga sulig kiekviena karta. Kas kiekviena karta F išauga puse,
prieš tai buvusios kartos, heterozigotų dažnio įverčiu. Inbrydingo rezultate,
homozigotų dažnis populiacijoje išauga heterozigotinių individų sąskaita. Atsitiktinio
kryžminimosi populiacijose, kuriose aptinkami aleliai A ir a su dažniais p ir q,
heterozigotų dažnis bus 2pq. Populiacijose, kuriose yra inbrydingo koeficientas F,
heterozigotų dažnis bus (1-F) nuo jų dažnio atsitiktiniai besikryžminančioje
populiacijoje.

Kai F=0, inbrydingo nėra, genotipų dažniai atitinka HdyWnb pusiausvyrą.
F atspindi individų, homozigotinių pagal kurį nors lokusą, perteklių populiacijoje; taip
pat atspindi homozigotinių lokusų dalies padidėjimą atskirų individų genotipuose.
Inbrydingo koeficientas taip pat vadinamas fiksacijos koeficientu. Tai
paaiškina simbolį F. Fiksacijos koeficientas yra tikimybė, kad alelis populiacijoje
bus fiksuotas homozigotinėje būsenoje
Į populiaciją kurioje vyksta inbrydingas galima žiūrėti kaip į dviejų dalių
bendruomenę, iš kurių vienoje yra visiškas inbrydingas, o kita – visiškai panmiksinė.
Tokiu atveju, dviejų gametų A apsijungimo tikimybė: laisvo kryžminimosi
bendruomenėje yra p2, o esant inbrydingui turi būti didesnė kažkokiu teigiamu dydžiu
ε t.y. p2+ε (yra daugiau homozigotinių individų )
gametų a apsijungimo tikimybe: q2+ε;
gametų a ir A – 2pq -2ε
Genetinės pusiausvyros atveju inbrydingo koeficientas (lygys koreliacijos koeficientui
tarp apsijungiančių gametų):
F= ε/pq t.y. ε = Fpq
Tokiu atveju zigotų dalis: AA: Q1 = p2+Fpq ; Aa: Q2 = 2pq(1-F); AA: Q3 = q2+Fpq
Kai Q1+Q2+Q3 =1.
Jei didelė populiacija yra padalinta į k panmiksinių grupių, tai tokioje visumoje yra
stebimas efektas, panašus į inbrydingo pasekmes esančias nepadalintoje populiacijoje:
homozigotų dalis išauga tarpopuliacin\ės genų dažnių variansos dydžiu, heterozigotų
dalies sumažėjimo sąskaita. Li (1978): “populiacijų padalijimas į atskiras
besikryžminančias grupes formaliai ekvivalentiškas inbrydingo poveikiui visos
populiacijos mastu”. Tokios diferenciacijos laipsnis tiesiogiai susijęs su
tarpopuliacinių genų dažnių pločiu (variansa) – kuo stipriau genetiškai skiriasi
subpopuliacijos, tuo didesnė variansos Vq vertė.
Wright’o F-statistika (1943,1951) – nustatė keletą F-koeficientų, kurie apibūdina
genetinę diferenciaciją:
1. FIT – individo inbrydingo koeficientas visos populiacijos atžvilgiu
2. FIS – individo inbrydingo koeficientas subpopuliacijos, kuriai jis priklauso,
atžvilgiu

3. FST- subpopuliacijos inbrydingo koeficientas visos padalintos populiacijos
atžvilgiu
FST=Vq/vid.q(1-vid.q),
q-geno dažnis padalintoje populiacijoje
Santykis tarp šių koeficientų: FIT =F ST+(1-F ST)F IS
Visi šie indeksai rodo nuokrypį nuo panmiksijos atsirandantį dėl apsijungiančių
gametų koreliacijos ir, galiausiai, nustatomi homozigotinių ir heterozigotinių genotipų
santykiu.
FST labai svarbi biologinė prasmė: stacionariomis sąlygomis atspindi populiacijų
genofondų diferenciacijos ir integracijos procesų balansą.
GST (Nei,75): F-statistikos ekvivalentas, surišantis bendrą (H) ir vidupopuliacinę (H)
genų įvairovę tokia formule:
GST= (HT -vid.HS )/HT,
kur HT =1-Σvid.pi2
vid.Hs =1/nΣHs
Hs=1-ΣpIS2,
kur pIS - i-ojo alelio dažnis S subpopuliacijoje, o
vid.pi – vidutinis i-ojo alelio dažnis visoje padalintoje populiacijoje,
kurią sudaro n subpopuliacijų.
Vid.Hs-vidutinis subpopuliacijos heterozigotiškumas;
- HT - visos padalintos populiacijos heterozigotiškumas, laikant kad ta
populiacija atstovauja vieningaim panmiksinei bendruomenei
Inbrydingo koeficientas skaičiavimas
Inbrydingo koeficientas (F) gali būti suskaičiuotas genealogijoje analizuojant
giminingumo laipsnį.
Inbrydingo koeficientas skaičiuojamas taip:
1. Nustatomas bendrų protėvių skaičius
Bendras protėvis yra tas, kuris yra bendras abiems individo tėvams
IV-1 turi vieną bendrą protėvį I-2
2. Nustatomas inbrydingo kelių skaičius

Inbrydingo kelias = trumpiausias genealogijos kelias, jungiantis abudu tėvus ir
bendrą protėvį.
Kiekvieno inbrydingo kelio ilgis suskaičiuojamas sudedant visus individus,
sudarančius kelią, išskyrus tiriamąjį individą
Pateiktame pavyzdyje yra tik vienas kelias:
IV-1 à III-2 à II-2 à I-2 à II-3 à III-3
Jį sudaro penki nariai.
Pav. Žmogaus genealoginis medis, rodantis inbrydingą
3. Skaičiavimams naudojat formulę
F = Σ (1/2)n(1 + FA)
Kur F yra tiriamojo individo inbrydingo koeficientas,n yra individų kiekis inbrydingo kelyje (išskyrus inbredinį individą), FA yra bendro protėvio inbrydingo koeficientas, Σ rodo sumą dydžių (1/2)n(1 + FA), paskaičiuotų kiekvienam inbrydingo keliui
Pateiktame pavyzdyje yra tik vienas bendras protėvis. Kadangi nieko nežinoma apie
jo paveldėjimą, daroma prielaida, kad FA=0
F = Σ (1/2)n(1 + 0)
F= (1/2)5 = 1/32
F = 3.125%
(F = 3.125%) Rodo geno homozigotiškumo tikimybę individe IV-1. Tai atsitinka dėl
paveldėjimo iš bendro protėvio I-2

Inbrydingo pasekmės gali būti įvertintos ir populiacijoje.
Tegul alelių A ir a dažnis yra p ir q. Tada genotipų dažniai bus nusakomi taip
p2 + Fpq atitinka AA dažnį
2pq(1 – F) atitinka Aa dažnį
q2 + Fpq atitinka aa dažnį
Tegul p = 0.8, q = 0.2 ir F = 0.25, tada genotipų dažniai bus
AA = p2 + Fpq = (0.8)2 + (0.25)(0.8)(0.2) = 0.68
Aa = 2pq(1 – F) = 2(0.8)(0.2)(1 – 0.25) = 0.24
aa = q2 + Fpq = (0.2)2 + (0.25)(0.8)(0.2) = 0.08
Esant inbrydingui bus: 68% AA homozigotų, 24% heterozigotų, 8% aa homozigotų
Nesant inbrydingo (t.y., F=0) bus: p2 = 64% AA homozigotų, 2pq = 32%
heterozigotų, q2 = 4% aa homozigotų
Išvada: Taigi, inbrydingas didina homozigotų dalį ir mažiną heterozigotų dalį.
Gamtinėse populiacijose inbrydingo koeficientas didėja, mažėjant populiacijos
dydžiui.
Inbrydingas gali turėti tiek teigiamų, tiek ir neigiamų pasekmių populiacijai.
Teigiamos pasekmės stebimos žemės ūkio kultūrose - inbrydingas gali padidinti
proporciją homozigotų, turinčių reikalingą požymį. Kaip neigiama pasekmė yra tai,
kad padidėja paveldimų recesyvinių ligų - inbrydingas padidina homozigotiškumo,
tuo pačiu ir ligos, tikimybę.
Inbrydinė depresija ir heterozė.
Selekcijos metu siekiama išvesti linijas pasižyminčias pačiomis geriausiomis
savybėmis, todėl tėviniai individai atrenkami su ryškiausiais požymiais. Vyksta
dirbtinė atranka. Dažnai vykdomas sistemingas inbrydingas, padidinantis
homozigotiškumą…bet…inbrydingas dažnai priveda prie labai svarbių gyvybiškų
savybių, kaip reprodukcija, gyvybingumas, suprastėjimo pas palikuonis. Tai
vad(inama inbrydine depresija. Ji atsiranda dėl letalių recesyvinių alelių
homozigotiškumo.
Heterozės efektas – priešingas inbrydinei depresijai. Pasiekiamas kryžminant
nepriklausomų inbrydinių linijų atstovus. Hibridai pasižymi geresnėmis savybėmis.
Inbrydinės linijos dažnai būna homozigotinės pagal kurį nors reikiamą “teisingą”
alelį, todėl jas sukryžminus, homozigotiškumas pagal dirbtinai atrinktus požymius
išlieka, o “neteisingi” aleliai pereina į heterozigotinę būseną.

5 GENETINIS PANAŠUMAS I IR DISTANCIJA D
Turint daugiau nei dvi imtis, ir analizuojant kelias polimorfines sistemas reikia
įvertinti genetinio panašumo indeksą (I).
Molekulinėje ekologijoje labai svarbu žinoti koks skaičius genų sutampa ir koks
yra skirtingas tarpe dviejų tiriamų populiacijų. Tokios žinios duotų atsakymą į
klausimą, kokio kiekio reikia genų pakaitalų, kad dvi palikuonių linijas atpažinti kaip
naujas rūšis. Išanalizavus ryšį tarp genų ar DNA cheminės struktūros ir baltymus
sudarančių amino rūgščių segmentų atsirado galimybė studijuoti genetinius skirtumus
analizuojant baltymų amino rūgščių seką. Daugelis tyrėjų analizavo to paties baltymo
amino rūgščių segmentų skirtumus pas skirtingus organizmus ir nustatė amino rūgščių
ir genų pakitimų per t.t. laiko vienetą konstantas. Šis metodas labai pasitarnavo tiriant
ilgalaikę evoliuciją, kaip šeimų, eilių ir klasių evoliuciją. Rūšių ir porūšių lygyje šis
metodas nėra labai sėkmingas, kadangi genetiniai pokyčiai šiame lygmenyje per
metus yra per maži. Kad šis metodas būtų naudingas analizuojant rūšių evoliucija turi
būti ištiriama labai daug baltymu sistemų, o amino rūgščių sekų tyrimai yra
pakankamai brangus.
Galimas ir kitas metodas kuris yra greitesnis tačiau ne toks tikslus. Grubus
panašumo įvertinimas gali būti gaunamas analizuojant baltymų elektroforetinį
judrumą. Šis metodas buvo panaudotas Hubbio ir Throckmortono studijose
analizuojant genetines distancijas tarp dvieju skirtingų Drosophila rušių. Išanalizavus
didelį kiekį baltyminių sistemų buvo parodyta kad sibsams budinga didesnis kiekis
bendrų baltymu negu ne-sibsams, bei morfologinių požymių koreliacija su baltymų ir
genų skirtumais. Kadangi amino rūgščių pakitimai per metus pasirodė besą konstanta
ir būdinga daugeliui skirtingų rūšių, pasiūlyta išvada kad sibsams būdinga didesnė
divergencija vieniem nuo kitų, nei ne-sibsams.
M. Nei pasiūlė patogų būdą, pagal kurį, turint elektroforezinius duomenis
galima įvertinti populiacijoje vykstančią genetinę diferenciaciją. Naudojami du
įverčiai:
Genetinis panašumas (I), įvertinantis struktūrinių genų dalį, kurie identiški
abiejose populiacijose.
Genetinė distancija (D), įvertina vidutinišką alelinių pokyčių skaičių
kiekviename lokuse, įvykusių šių populiacijų atskiros evoliucijos metu.

Aleliniai pokyčiai atsiranda tuomet, kai alelinių mutacijų vykstančių atskiruose
lokusuose metu, alelis yra pakeičiamas kitu aleliu arba pasikeis visas alelinių genų
rinkinys iš karto.
Šis metodas įvertina tai, kad alelių pakeitimas gali būti nepilnas, t.y. kažkurioje
populiacijos dalyje naujas alelis gali pakeisti seną, kuris, su didesniu ar mažesniu
dažniu, vis tiek yra aptinkamas populiacijoje.
0<I<1, 0- lyginamose populiacijose nėra bendrų alelių; 1- alelių dažniai yra
vienodi abiejose populiacijose;
0<D<∞, 0 - nėra jokių alelinių pokyčių. Ši reikšmė gali būti didesnė už 1
kadangi evoliucinio proceso metu, vykstančio ilgą laiką, kiekvieno lokuso aleliai gali
būti ne vieną kartą pilnai pakeičiami.
Ik=∑ai bi / √∑ai2 ∑ bi
2,
k- lokusas kurio atžvilgiu populiacija yra polimorfinė pagal i skirtingų alelių; A
ir B – dvi populiacijos; ai ir bi, i-ojo alelio dažniai atitinkamai A ir B (laisvai
besikryžminančios) populiacijose. Jei abi populiacijos monomorfinės pagal tą patį
alelį lokuse, t.y. a ir b lygūs 1, gemnetinis panašumas irgi lygus 1, t.y. populiacijos yra
tapačios pagal šį lokusą; jei lokusas monomorfinis pagal skirtingus alelius (a-0, b-1), I
bus 0, t.y. nėra visiškai jokio tapatumo. ___
Kelių lokusų atveju
Iab=Σaibi, Ia=Σai, Ib=Σbi, tada I=Iab/√IaIb
O genetinė distancija
D= - lnI
Jei nevyksta selekcija ir alelės atsiranda dėl protėvinėje populiacijoje įvykusios
mutacijos Ia,
Ib yra lygūs Wright’o inbrydingo koeficientui.
Genetinės distancijos ir panašumo įvertinimui užtenka 3 lokusų (min)
Pvz.: I-0,525, D-0,644, dviejų populiacijų nepriklausomos evoliucijos eigoje
kiekviename iš 100 lokusų vidutiniškai įvyko 64,4 aleliniai pakitimai (0,64 pakitimai
vienam lokusui).
Taikant Wehrhahns’o(1975) {šis išrado tikimybinį metodą, kuriuo yra tiriami
homologinių baltymų tarpe atsirandantys krūvių pokyčiai; minusas – daryta prielaida,
kad populiacija prasideda nuo vieno alelio.} populiacinės divergencijos numerinius
apskaičiavimus Nei (1972) distancijos patikrinimui, buvo parodyta, kad ši D
išskaičiuota iš elektroforezinių duomenų yra maždaug 10% mažesnė nei tikėtinas

aminorūgščių pakitimų, iššaukiančių krūvio pokyčius ankstyvojoje populiacijų
divergencijos stadijoje, skaičius ir tai gali nulemti nepakankamų įverčius lyginant dvi
rūšis.
Individų imtis įvertinant vid.H gali būti labai maža, jei yra išanalizuojamas
didelis lokusų skaičius ir vid.H yra žemas. Individų skaičius D įvertinimui taip pat
gali būti labai mažas jei D yra didelė, o vidH tarp dviejų lyginamų rūšių yra žemas.
Paišant dendrogramas, remiamasi D skirtumais tarp skirtingų rūšių porų. Jei šie
skirtumai maži, D turi būti išstudijuota preciziškiau. Tokiu atveju turi būti ištirtas
pakankamai didelis individų skaičius kiekvienam tiriamam lokusui. Jei skirtumai labai
dideli, net vieno individo gali užtekti, kad nustatyti jo tikslią dendrogramos
topologiją. Organizmuose, kuriuose vidH didesnis už 0,1 turi būti išanalizuotas
sąlyginai dielis individų skaičius, kad dendrograma būtų teisinga.
Nei unbiased vidH ir D įverčius galima taikyti bet kokiam individų skaičiui, ir
tie įverčęiai bus kokybiški, nepriklausomai nuo to kiek lokusų yra išanalizuota.
Skirtumai tarp biased ir unbiased įverčių yra labai maži kai ištirtų individų skaičius
yra labai didelis (50 ir daugiau).
MinD=-ln [vidGab/√vidGavidGb],
vidGa ir vidGb vidurkinės reikšmes (2naIa-1)/(2na-1) ir (2nbIb-1)/(2nb-1) per visus
ištirtus lokusus r, atitinkamai, o vidGab = Iab. n – individų skaičius (A(a) ir B(b)
populiacijose). Ga ,Gb ir Gab yra Σpi2,Σqi
2, Σ piqi reikšmės per visus lokusus genome. Ia
, Ib ir Iab individų geno identiškumas: Σai2,Σbi
2, Σ aibi vidurkinės reikšmės r ištirtiems
lokusams.
UnbH naudojama jei yra mažas individų skaičius, ir nedidelis skaičius lokusų
išanalizuotas (normoje turi buti atliekama min50 lokusų analizė). Pavyzdžių variansa
įvertina ytarplokusinę ir vidulokusinę variansas.
Genetinis polimorfizmas stebimas gamtinėse populiacijose yra įvairių
evoliucinių jėgų ilgalaikio poveikio rezultatas. Tokiu atveju, teorija aiškinanti
genetinio polimorfizmo esmę turėtų paaiškinti ir evoliucinius pokyčius populiacijoje
vykusius praeityje. Li ir Nei (1975): laikantis mutacijų dreifo hipotezės, tikėtina kad
H koreliacija silpnėja greičiau, nei genetinis I, iklgejant evoliucijos laikui.

Kitų autorių formulės
Cavalli-Sfoza et al(1967):
greičiausiai, ją reik naudoti kai analizuojama populiacijų divergencija rūšies
viduje veikiant atsitiktiniam genų dreifui.
I=(2√2/π)√(1-r),
m
kur r=Σ√piqi, r<1; r=1, kai lyginamos populiacijos yra identiškos. i=1
Edwards (1971):
8(1-r)/ (1+Σ√pi/m)(1+ Σ√qi/m)
m – suminis alelių skaičius lyginamose populiacijose.
________________________
Nei (1972)
m m m
I=Σ(piqi)/ √ ((Σpi2 )(Σqi
2)), i=1 i=1 i=1
gali blogai įvertinti panašumą, jei populiacija ryškiai skiriasi pagal retus alelius
(20%). Jei A ir B populiacijose sutampa 20% alelių tai I rodo visišką panašumo
nebuvimą.
_________________
Sokal, Sneath, 1963:
I=Σpiqi, kuo polimorfiškesnės identiškos populiacijos tuo mažesnė I reikšmė.
_______________________
Rogers (1972):
D=√Σ(pi -qi)2,
Kuo polimorfiškesnės lyginamos populiacijos tuo rodo mažesnę D

2. lentelė. Genų dažnumų p, q bei genotipų dažnumų p2, 2pq, q2 reikšmės.
p p2 2pq q2 q p p2 2pq q2 q.01 .0001 .0198 .9801 .99 .26 .0676 .3848 .5476 .74.02 .0004 .0392 .9604 .98 .27 .0729 .3942 .5329 .73.03 .0009 .0582 .9409 .97 .28 .0784 .4032 .5184 .72.04 .0016 .0768 .9216 .96 .29 .0841 .4118 .5041 .71.05 .0025 .0950 .9025 .95 .30 .0900 .4200 .4900 .70.06 .0036 .1128 .8836 .94 .31 .0961 .4278 .4751 .69.07 .0049 .1302 .8649 .93 .32 .1024 .4352 .4624 .68.08 .0064 .1472 .8464 .92 .33 .1089 .4422 .4489 .67.09 .0081 .1638 .8281 .91 .34 .1156 .4488 .4356 .66.10 .0100 .1800 .8100 .90 .35 .1225 .4550 .4225 .65.11 .0121 .1958 .7921 .89 .36 .1296 .4608 .4096 .64.12 .0144 .2112 .7744 .88 .37 .1369 .4662 .3969 .63.13 .0169 .2262 .7569 .87 .38 .1444 .4712 .3844 .62.14 .0196 .2408 .7396 .86 .39 .1521 .4758 .3721 .61.15 .0225 .2550 .7225 .85 .40 .1600 .4800 .3600 .60.16 .0256 .2688 .7056 .84 .41 .1681 .4838 .3481 .59.17 .0289 .2822 .6889 .83 .42 .1764 .4872 .3364 .58.18 .0324 .2952 .6724 .82 .43 .1849 .4902 .3249 .57.19 .0361 .3078 .6561 .81 .44 .1936 .4928 .3136 .56.20 .0400 .3200 .6400 .80 .45 .2025 .4950 .3025 .55.21 .0441 .3318 .6241 .79 .46 .2116 .4968 .2916 .54.22 .0484 .3432 .6084 .78 .47 .2209 .4982 .2809 .53.23 .0529 .3542 .5929 .77 .48 .2304 .4992 .2704 .52.24 .0576 .3648 .5776 .76 .49 .2401 .4998 .2601 .51.25 .0625 .3750 .5625 .75 .50 .2500 .5000 .2500 .50

6. MOLEKULINĖS EKOLOGIJOS LABORATORINIAI DARBAI
. Baltymų įvairovės tyrimas
Tikslas: Nustatyti tam tikros vietovės gyvūnų izofermentinį kintamumą.
Tyrimo medžiaga. Baltymų elektroforezės metodu tyrimui naudojama po 20-
30 individų iš 4 (ar daugiau) rajonų, 4-8 skirtingų biotopų, audinių (kraujo, raumenų
ar kepenų) mėginiai.
Homogenizavimas: Atšildytas kepenų audinio gabaliukas homogenizuojamas
stikliniame homogenizatoriuje pridedant 0,6 ml homogenato tirpalo.
Homogenizavimo terpė ruošiama iš:
1% Tritonas X–100 -1 ml
MgCl2 x 6H2O – 0,4 g
0,2 M Tris – HCl buferis pH 8 - iki 100 ml
(Tris – 0,6 g; 1 N HCl - 27,5 ml; vanduo iki 100ml)
Saugant biologinius pavyzdžius nuolat vyksta natyvios baltymų struktūros
pakitimai, dėl:
• denatūracijos (polipeptidinių grandinių struktūriniai pokyčiai);
• agregacijos (kelių molekulių susijungimo);
• dezagregacijos į subvienetus (ketvirtinės struktūros praradimas);
• polipeptidinių grandinių degradacijos (pirminės struktūros pokyčiai dėl
proteolitinių fermentų veikimo).
Visi išvardyti procesai nulemia baltymų biologinių savybių praradimą
(fermentinį aktyvumą ir kt. funkcijas) ir tuo įtakoja jų elektroforezinį paslankumą.
Paruoštus homogenatus iki bandymų ir tarp jų laikomi -20°C temperatūroje,
kad fermentai nedegraduotų, neapsikrėstų bakterijomis ir grybeliais, įnešančius
papildomo proteazių aktyvumo.
Centrifugavimas: Jo metu pavyzdžiai paruošiami tyrimui. Centrifugavimo
greitis priklauso nuo pavyzdžio savybių. Centrifuguojama 15 sek. 1,5 tūkst.
apsisukimų per minutę greičiu.
Elektroforezė poliakrilamidiniame gelyje (PAAG)
Aparatūra:

Nespecifinių baltymų ir izofermentų elektroforezei PAAG gelio bloke buvo
naudojama vertikalios gel-elektroforezės aparatai „HIMFILL“, Tallin (4 pav.),
kuriuos sudaro:
- Nejudrūs plieno ir platininis elektrodai
- Kameros buferiui. Suformuojamos įstačius rėmelius nešėjui.
- Rėmeliai nešėjui, kurį kiekvieną sudaro korpusas ir du stiklai
(115x120x1,5 mm).
- Standartinės „šukutės“ pavyzdžių užnešimo vietoms („šulinėliams“)
formuoti gelyje.„Šukutės“ sudaro 13 „šulinėlius“ formuojančių plokštelių
- Tekančio vandens principu veikianti šaldymo sistema ( palaikoma +4ºC
temperatūra)
Darbo metu naudojami 2301 Mcrodrive 1 Power Supply ir УИП-1
maitinimo šaltiniai.
4 pav. Vertikalios gel-elektroforezės aparatai
PAAG bloko paruošimas:
Elektroforezė, priklausomai nuo tiriamos izofermentų sistemos, buvo
atliekama dvisluoksniame vertikaliame 5 / 7,5 % ir vienasluoksniame 5 % PAA
gelyje. Geliai ruošiami remiantis Harry ir Hopkinson (1975) metodinėmis
rekomendacijomis su kai kuriomis modifikacijom.
Pirmiausiai buvo paruošiama 20% AKA:
Akrilamidas 96g
Bis 4g
Dist.vanduo iki 500ml
Paruoštas tirpalas nufiltruojamas ir laikomas tamsoje prie 3-4ºC
temperatūroje.
Tai monomerų tirpalas naudojamas akrilamido linijų polimerų susiuvimui.
Nuo jo santykio priklauso gelio elastingumas ir poringumas.

Taip pat reikia pasiruošti gelinį buferį (Tris-glicininis buferis, pH 8,3):
0,0495 M Tris 6g
0,383 M Glicininas 28,8g
Dist.H2O iki 1000ml
Tris- EDTA-H3BO3 buferis, pH ~8,3 – 8,4 (Peacock et al., 1965)
0,9 M Tris 109,3 g
0,025 M Na2EDTA* 2 H2O (trilonas B) 9,30 g
0,89 M H3BO3 55,04 g
H2O dist. iki 1000 ml
Darbinis buferis:
Tris- EDTA-H3BO3 buferis, pH 8,3 -8,4 200 ml
H2O dist. iki 2000 ml
Gelinis buferis palaiko ir stabilizuoja nešėjo pH, taip įtakodamas medžiagos
migracijos greitį. Be to jis taip pat naudojamas kaip tiriamo pavyzdžio tirpiklis.
Naudojant didelės joninės jėgos buferius, pavyzdžių migracijos greitis mažėja,
sumarinė srovė didėja, išsiskiria šilumos kiekis. Mažinant buferio joninę jėgą
migracijos greitis didėja, sumarinė srovė ir išsiskiriantis šilumos kiekis mažėja, didėja
difuzija ir mažėja skiriamoji geba.
Paruošiami PAAG su skiriamosiomis AKA koncentracijomis:
TEMED (N,N,N,N′- tetrametiletilendiamidas) naudojamas kaip polimerazinės
reakcijos katalizatorius.
PCA naudojamas kaip polimerizacijos iniciatorius.
Ruošiant dvisluoksnį PAAG, koncentruojantis sluoksnis sudaro 1/3-1/4
bendro galutinio tirpalo, o skiriantysis 2/3-3/4. MDH (Malato dehidrogenazė), ME
(Malik fermentas) ir G6PDG (Gliukozė-6- fosfato dehidrogenazė) sistemas tiriant
Komponentai Gelio koncentracija 5% 7,50%20 % AKA 20 ml 30 ml
Pagrindinis buferis 8 ml 8 mlTEMED 0,08 ml 0,08 mlDist.H2O iki 75 75 mlPCA 0,08 g 0,08 g
Galutinė išeiga 80 ml 80 ml

buvo naudojam vienasluoksnis 5 % PAA gelis, o NB (Nespecifinis baltymas) ir EST
(Esterazės) dvisluoksnius 5 % / 7,5% PAA gelius.
Pagrindinės savybės suteikiančios PAAG privalumą prieš kitus nešėjus:
• Pagal porų dydį galima varijuoti plačiose ribose. Gelio procentingumas
pasirenkamas empiriškai, atsižvelgiant į frakcionuoto objekto molekulinę
masę.
• Naudojamo buferio pH neturi ryškios įtakos gelio polimerizacijos
procesui, kas suteikia galimybę eksperimento metu pasirinkti pačias
palankiausias sąlygas.
• PAAG būdinga žemo laipsnio absorbcija ir elektroendoosmozė, dėl ko
makromolekulių migracijos metu dažniausiai išvengiama šalutinių efektų.
• Makromolekulių frakcionavimo laikas PAAG yra sąlyginai trumpas
lyginant su kitais nešėjais.
Prieš sustingstant geliui į jį įdedama „ šukutės“kurios prieš eksperimentą
išimamos ir formuoja „kišenes“ į kurias įnešamas tiriamas homogenatas. Gelių
polimerizacija trunka apie 15-20 minučių.
Elektroforezės paruošimas:
Kamera elektroforezei buvo paruošta sekant nurodymus instrukcijoje.
Elektrodiniai indai užpilami atitinkamu elektrodiniu buferiu. Naudojam šaldymo
sistema, kuri elektroforezės kamerose palaiko 3-5ºC temperatūrą. Elektroforezės metu
bloko pašildymas gali sukelti fermentų inaktyvaciją bei rezultatų iškraipymą.
Paleidžiama preelektroforezę (160V, 30 minučių), kurios metu iš gelių išvaromas
PCA (nes jis yra vienintelis kenksmingas komponentas, kuris veikia fermentus, todėl
prieš pradedant frakcionavimą pašalinamas iš gelio preelektroforezės būdu).
Prieš eksperimentą buvo pasiruošiama dešimt bandinių su 7 µl 40 %
sacharozės tirpalo, kuris suteikia klampumą ir neleidžia pavyzdžiui išplaukti
apsaugodamas nuo išplaukimo efekto pavyzdžių užnešimo į „kišenes“ metu. Taip pat
įlašinama lašas bromfenolio mėlynojo. Tai vadinamas lyderinis dažas (angl. tracker
dye) kurio migracijos greitis yra didesnis nei frakcionuojamų sistemų, todėl pagal jį
stebima elektroforezės eiga. Taip pat imama 10 µl bandinio ir sumaišius su sacharoze
bei bromfenolio mėlynuoju užnešama individualiai į jam skirtą „kišenę“.
Elektroforezė vykdoma tokiu režimu:

I etapas: Pavyzdžių išėjimas iš „kišenių“ (40mA, 20 minučių). Šio etapo metu
baltymai koncentruojasi buferio/gelio riboje ir glaustesnėje formoje patenka į gelį.
II etapas: Darbinis režimas. Srovė pakeliama iki 250V ir frakcionavimas
atliekamas 1-2 valandų.
Baltymų dažymas
Dažymo principas:
Po elektroforezės izofermentų lokalizacija nustatoma dažymo metu.
Specifiškumą apsprendžia dažančio tirpalo sudėtyje esantis specifinis analizuojamam
fermentui substratas ir druskos, kurios reaguoja su fermento kanalizuojamos reakcijos
produktu :
Substratas + fermentas → produktas+ druska → spalvota substancija
Nespecifinio baltymo ir esterazių dažymo metu spalvota substancija susidaro
vykstant druskų kondensacijai su aromatiniu diazotiniu komponentu (FBRR, CBB).
Nespecifinės esterazės (EST) pasižymi labai didele genetine įvairove.
Dehidrogenazių dažymo metu, vyksta tetazolio (NBT) oksidacijos - redukcijos
reakcija, susidaro formazanas, netirpus tamsiai mėlynos-violetinės spalvos junginys.
Dažai gaminami prieš pat dažymą. Bandymo metu buvo dažomi šie fermentai:
• MDH ( Malato dehidrogenazė);
• ME (Malik-fermentas);
• G6PDG ( Gliukozė-6- fosfato dehidrogenazė);
• NSP ( Nespecifinis baltymas)
• EST ( Esterazės)
• LDH ( Laktato dehidrogenazė)
• XDH ( Ksantino dehidrogenazė)
Dažų paruošimas:
Malik-fermentas [ME, E.C. 1.1.1.40
Pagal Ayala et. (1972), Cross et al..(1979) metodikas su tam tikromis
modifikacijomis:
0,05 M Tris-HCl buferis, pH 8,5 100 ml ( 25 ml 0,2 M Tris + 15 ml 0,1 N HCl
+ dist. H2O iki 100ml) arba ( 0,6 g Tris į 25 ml H2O + 15 ml 0,1 N HCl + dist. H2O
iki 100ml)
Malic acid Na2 salt 0,1 g

MgCl2 ∙ 6 H2O 0,025 g
PMS 0,002 g
NBT 0,03 g
NADP 0,04 g
Inkubuojama 37ºC temperatūroje tamsoje 2-4 valandas. ME izofermentai
dažosi tamsiai violetine spalva.
Reakcija: Malic acid Na2 salt + NADP ↔ piruvatas + CO2+ NADPH + H
Malato dehidrogenazė [MDH, E.C. 1.1.1.37]
Pagal Brewer ( 1970) metodika su tam tikromis modifikacijomis:
0,05 M Tris-HCl buferis, pH 8,5 100 ml ( 25 ml 0,2 M Tris + 15 ml 0,1 N HCl
+ dist. H2O iki 100ml) arba ( 0,6 g Tris į 25 ml H2O + 15 ml 0,1 N HCl + dist. H2O
iki 100ml)
Natrio malatas 0,1 g
NAD 0,03 g
NBT 0,03 g
PMS 0,002 g
Inkubuojama 37ºC temperatūroje tamsoje 2 valandas. MDH izofermentai
dažosi tamsiai violetine spalva.
Reakcija: Natrio malatas +NAD ↔ oksalacetatas +NADH +H
Glukozo-6-fosfato dehidrogenazė [G6PDH, E.C. 1.1.1.49]
Pagal Brewer ( 1970) metodika su tam tikromis modifikacijomis:
0,5 M Tris-HCl, pH 7,1 25 ml (1,2g Tris + 45 ml 0,1 N HCl iki pH7 + dist.
H2O iki 100ml)
NADP 0,03 g
NBT 0,02 g
PMS 0,002 g
D-glucose-6-phosphate disodium salt 0,2 g
H2O iki 90ml
Gelis inkubuojamas 37ºC temperatūroje tamsoje apie 1 valandą. Aktyvumas
pasireiškę tamsiai mėlynomis juostomis.
Esterazės [EST, nespecifinės]
Pagal Brewer ( 1970) metodika su tam tikromis modifikacijomis:
α-Naftilacetato tirpalas 0,06 g
Acetonas 3 ml

FBRR ( angl. Fast blue RR salt) 0,06 g
H2O iki 100ml
Inkubuojama 30 minučių. Esterazės pasireiškia rausvos- rudos spalvos
zonomis, naudojant α- izomera – tamsiai rudos spalvos.
Nespecifinis baltymas [NSP]
Comasie blue G-250 arba R-250 0,2 g
HClO4 17,5 ml
H2O iki 500ml
Inkubuojama kambario temperatūroje apie vieną parą. Baltymų frakcijos
nusidažo tamsiai mėlyna spalva.
Laktato dehidrogenazė [LDH, E.C. 1.1.1.27]
0,05 M Tris-HCl buferis, pH 7,5 100ml
Na laktatas 0,2 g
PMS 0,005 g
NBT 0,03 g
NAD 0,02 g
Inkubuojama prie 37ºC temperatūroje tamsoje apie valandą. dažosi tamsiai
mėlyna spalva.
Ksantino dehidrogenazė [XDH, E.C. 1.1.1.204] monomeras arba dimeras.
0,05 M Tris- HCl buferis, pH 8,5 100ml
Hypoxanthine 0,1 g
NAD 0,03 g
NBT 0,02 g
PMS 0,002 g
Inkubuojama prie 37ºC temperatūroje tamsoje.
Po dažymo geliai fiksuojami 7% acto rūgšties tirpale ir įdedami į uždaromą
maišelį.