most uv lesions are repaired by nucleotide excision repair (ner) stalled replication forks may be...
TRANSCRIPT
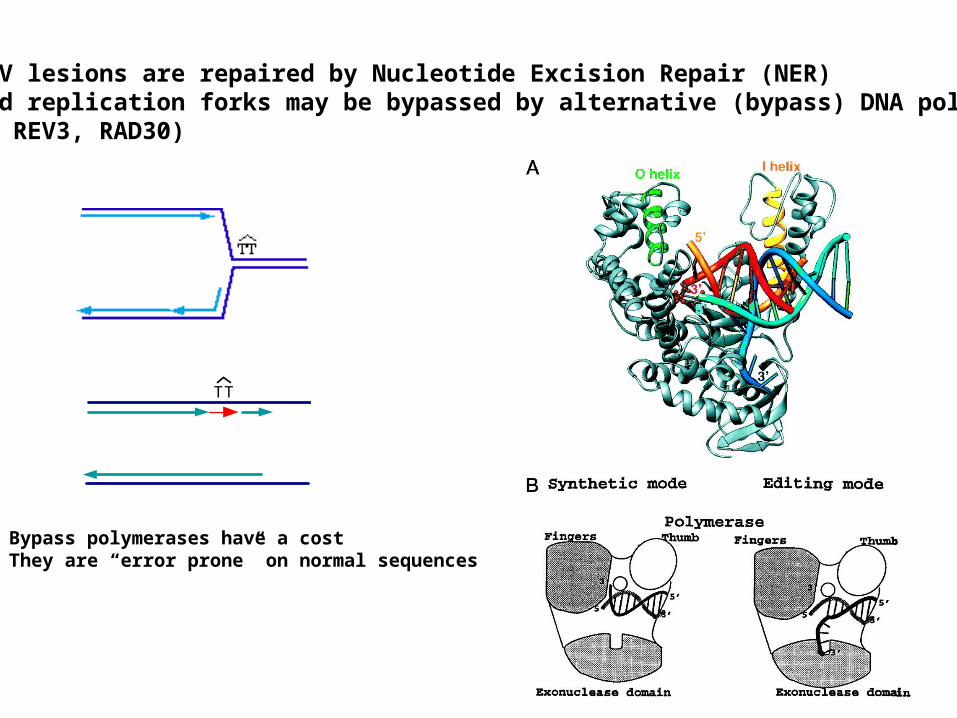
Most UV lesions are repaired by Nucleotide Excision Repair (NER)Stalled replication forks may be bypassed by alternative (bypass) DNA polymerases(REV1, REV3, RAD30)
TT
Bypass polymerases have a cost They are “error prone” on normal sequences
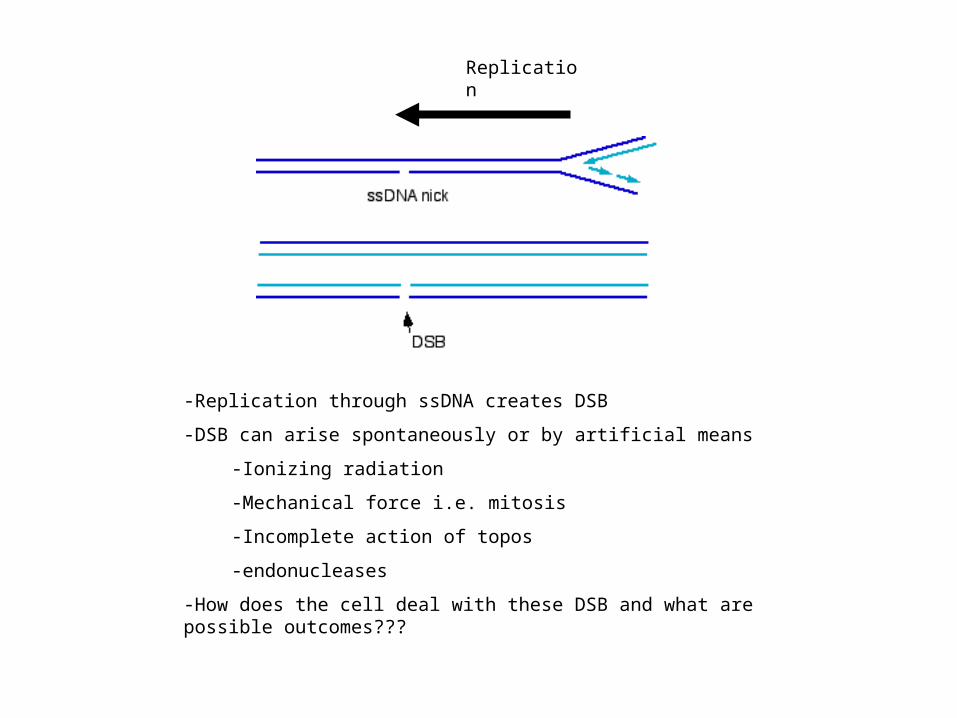
Replication
-Replication through ssDNA creates DSB
-DSB can arise spontaneously or by artificial means
-Ionizing radiation
-Mechanical force i.e. mitosis
-Incomplete action of topos
-endonucleases
-How does the cell deal with these DSB and what are possible outcomes???

Recombination as a source of genetic instability
A hallmark of cancer cells is their genetic instability
Most of these types of instability may be explained by various mechanisms of homologous and nonhomologous recombination
In all probability the initiating lesion on DNA that leads to these changes in chromosomes can be attributed to a double-strand break (DSB)
Loss of heterozygosityReciprocal and nonreciprocal translocationsDeletions TruncationsChromosome loss
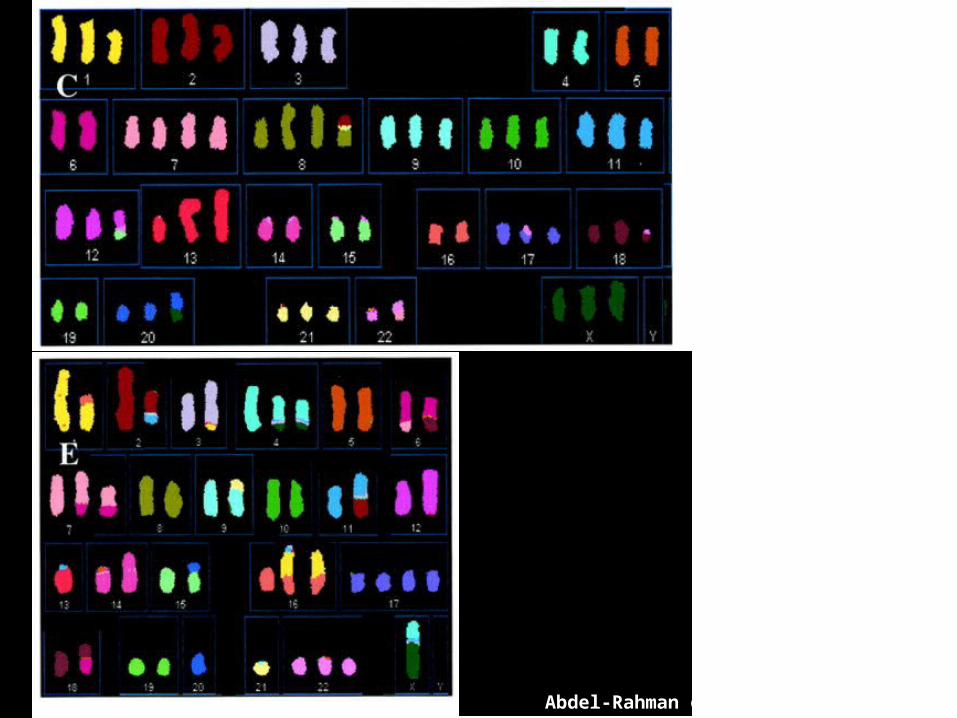
Abdel-Rahman et al. PNAS 98: 2538
Genome instabilityin tumorcells
TruncationsTranslocationsInversionsDuplicationsAmplifications
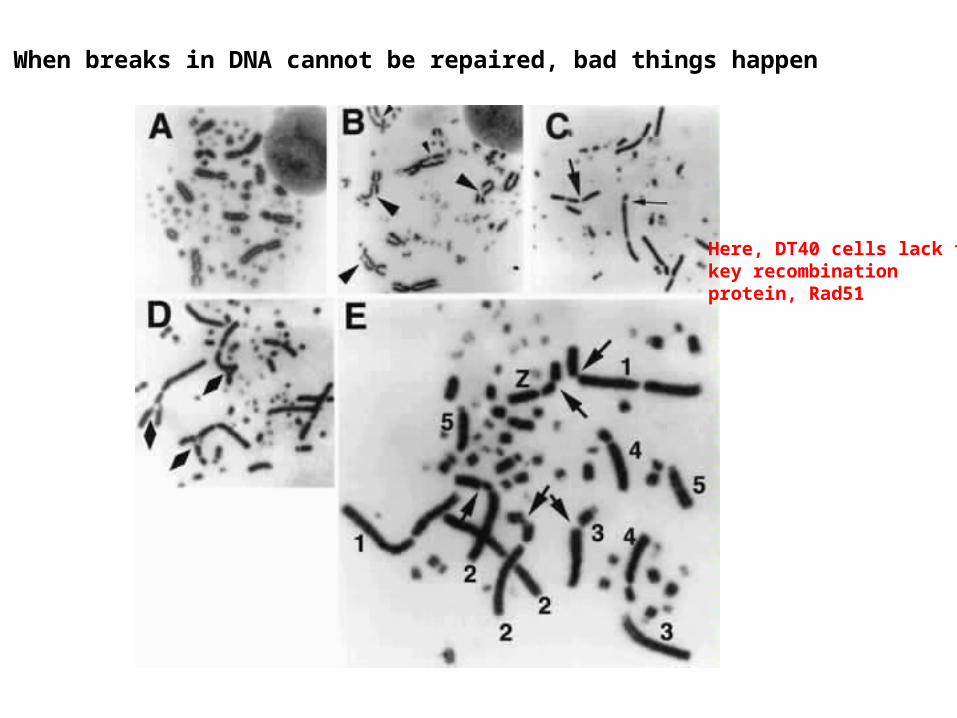
When breaks in DNA cannot be repaired, bad things happen
Here, DT40 cells lack thekey recombinationprotein, Rad51
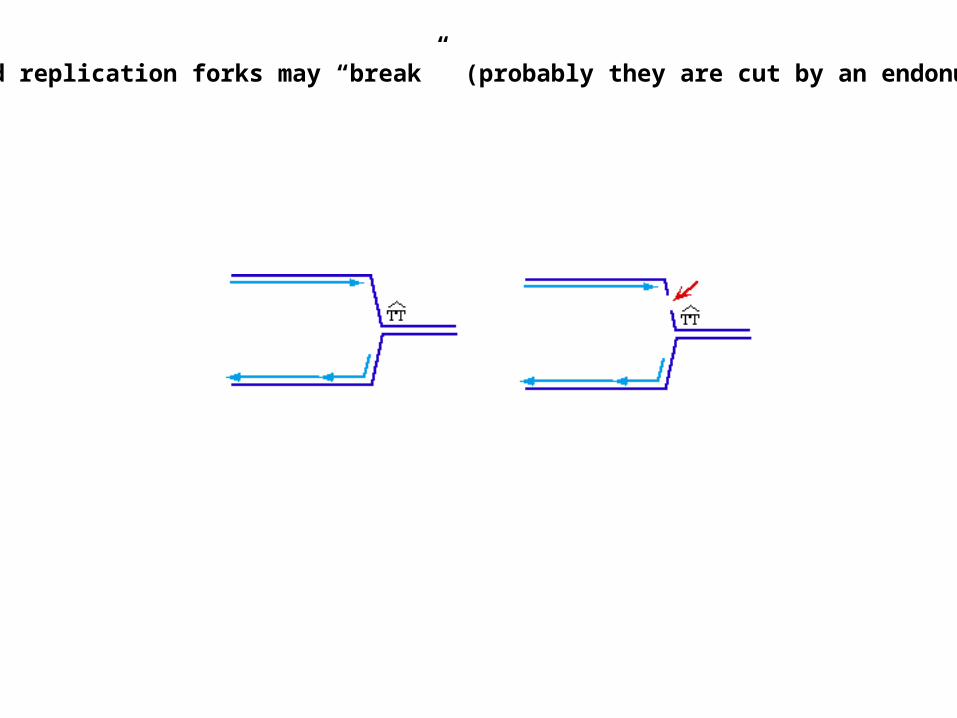
Stalled replication forks may “break” (probably they are cut by an endonuclease)
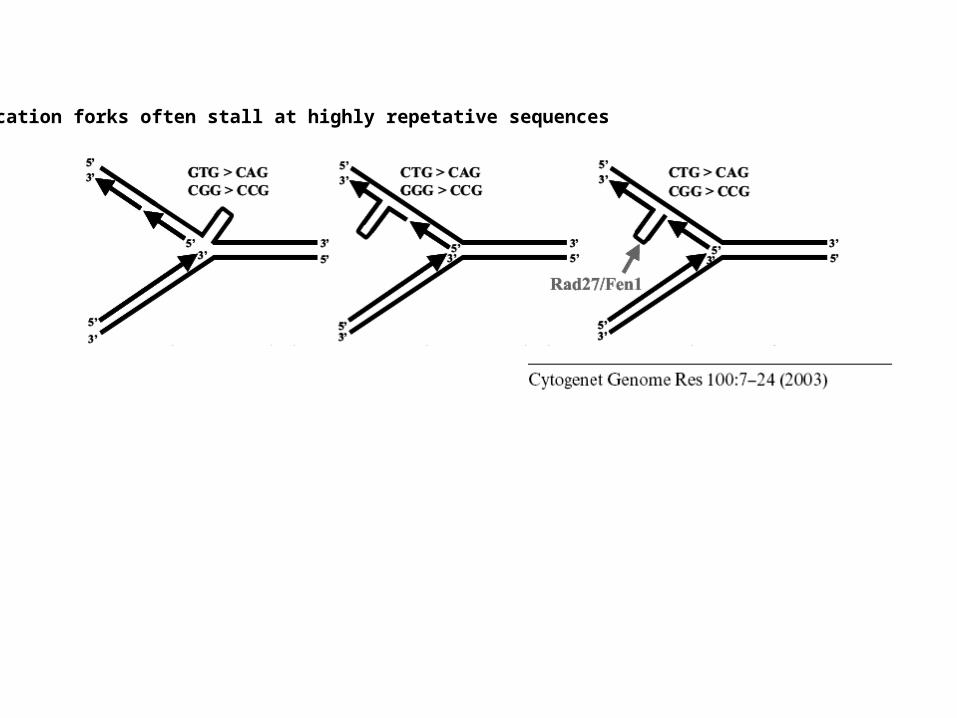
Replication forks often stall at highly repetative sequences
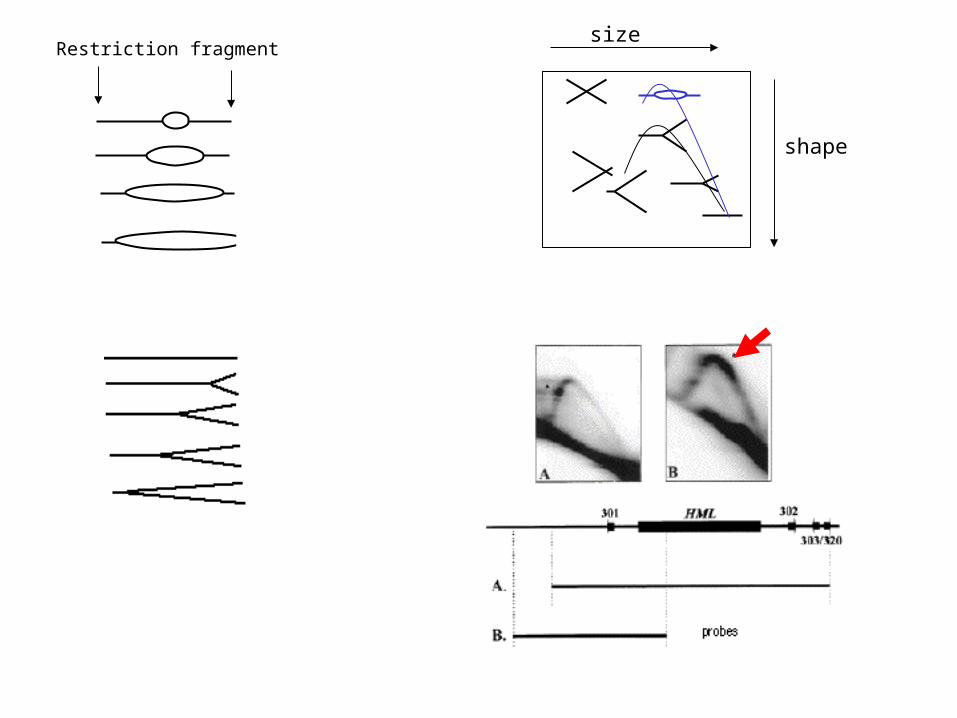
size
shape
Restriction fragment
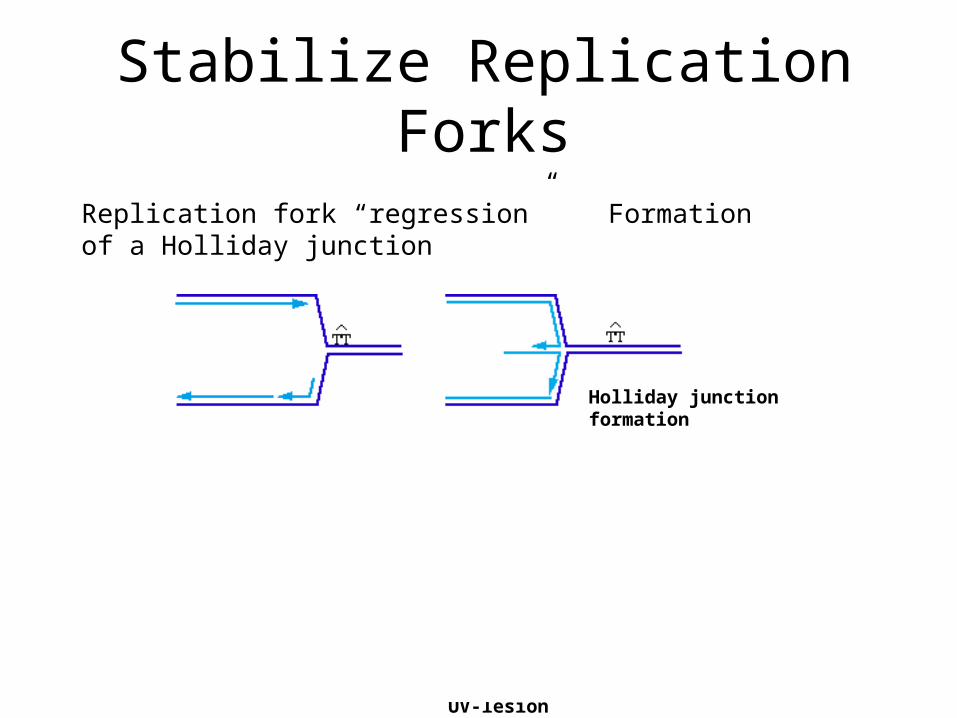
Replication fork “regression” Formation of a Holliday junction
Extension of leading strandBranch migration reverses Holliday junction and allows bypass of the UV-lesion
Holliday junction formation
Stabilize Replication Forks
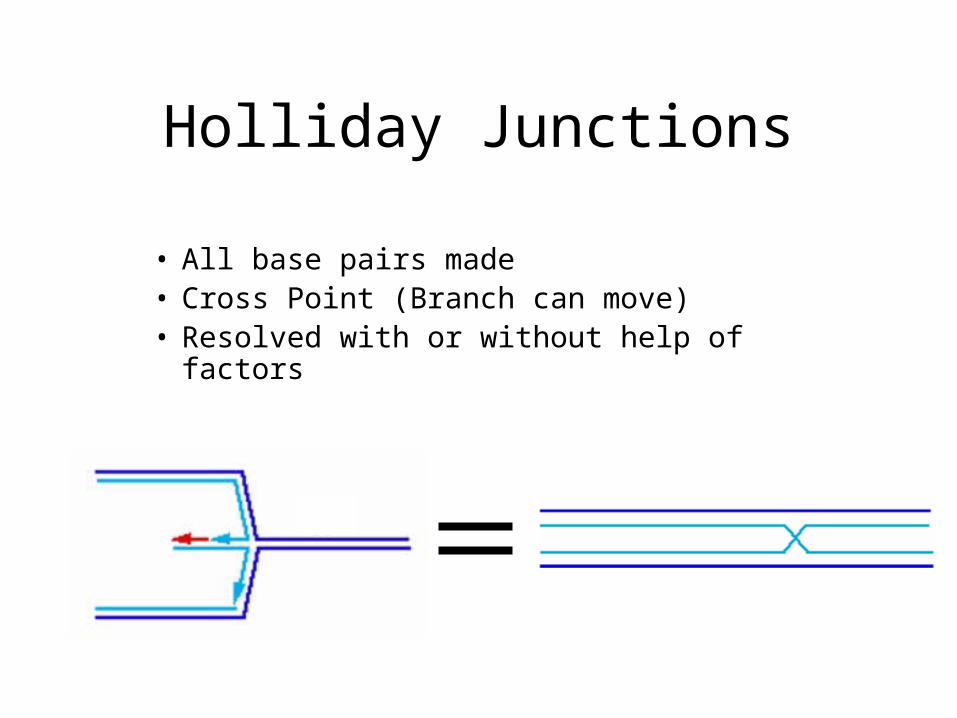
Holliday Junctions
• All base pairs made• Cross Point (Branch can move)• Resolved with or without help of factors
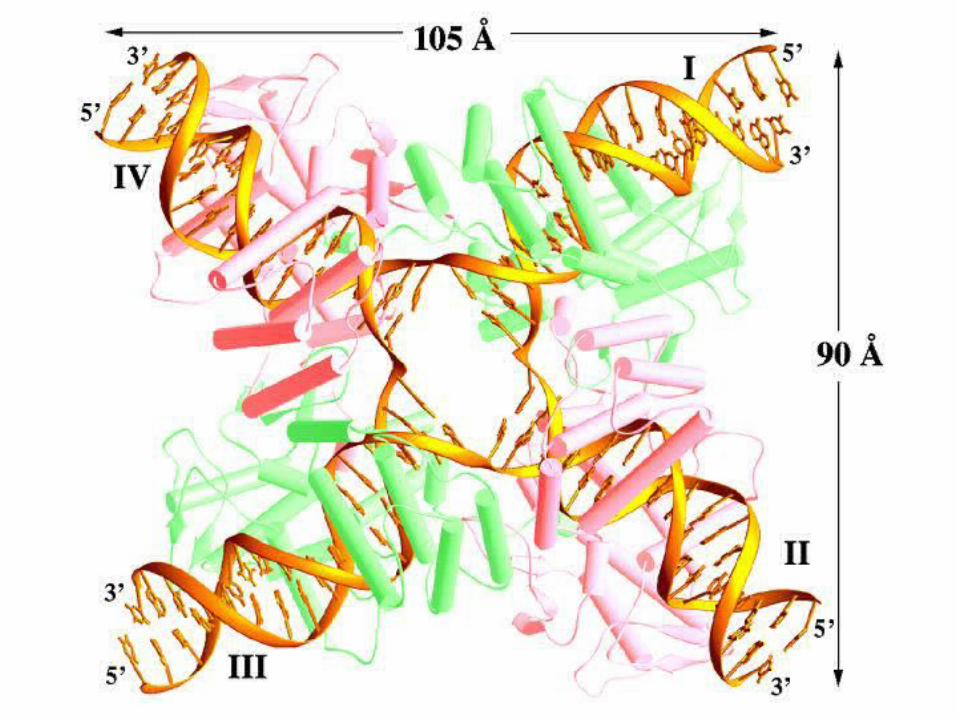

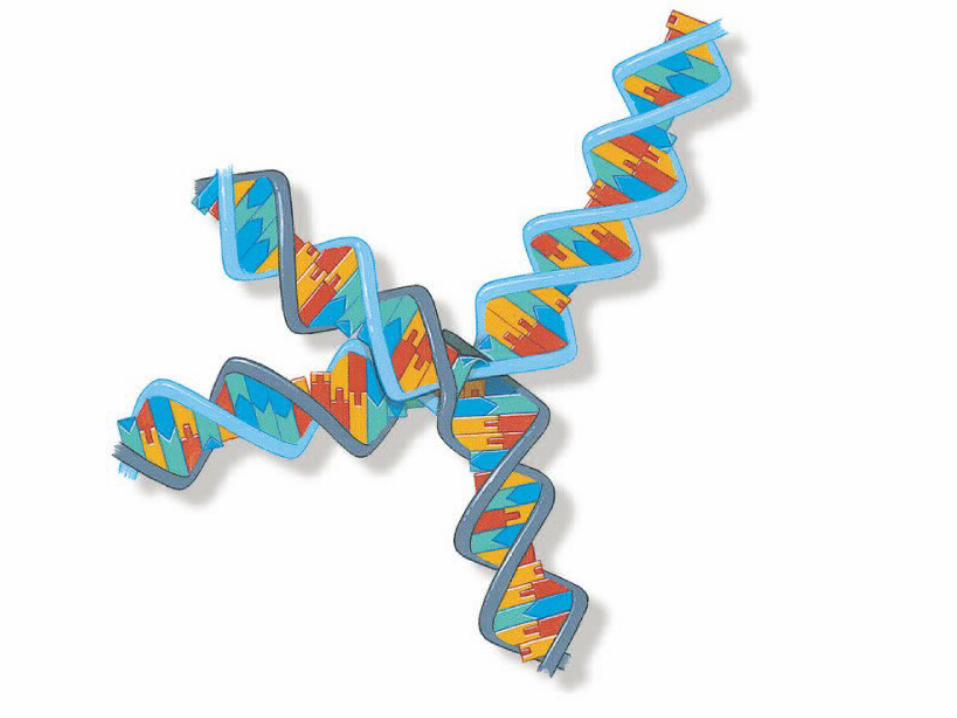
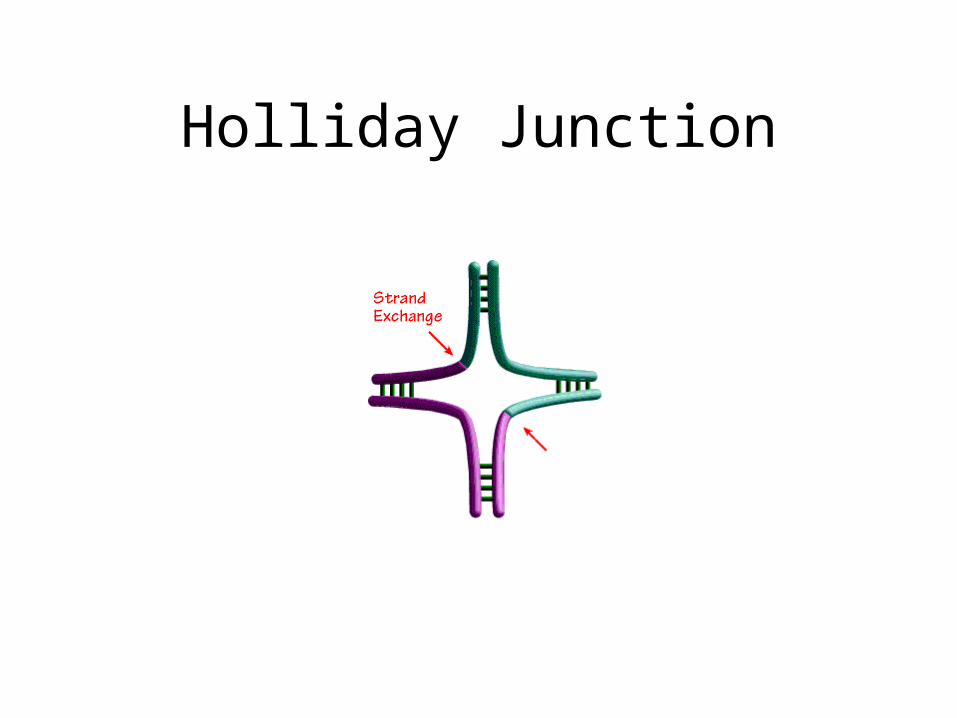
Holliday Junction
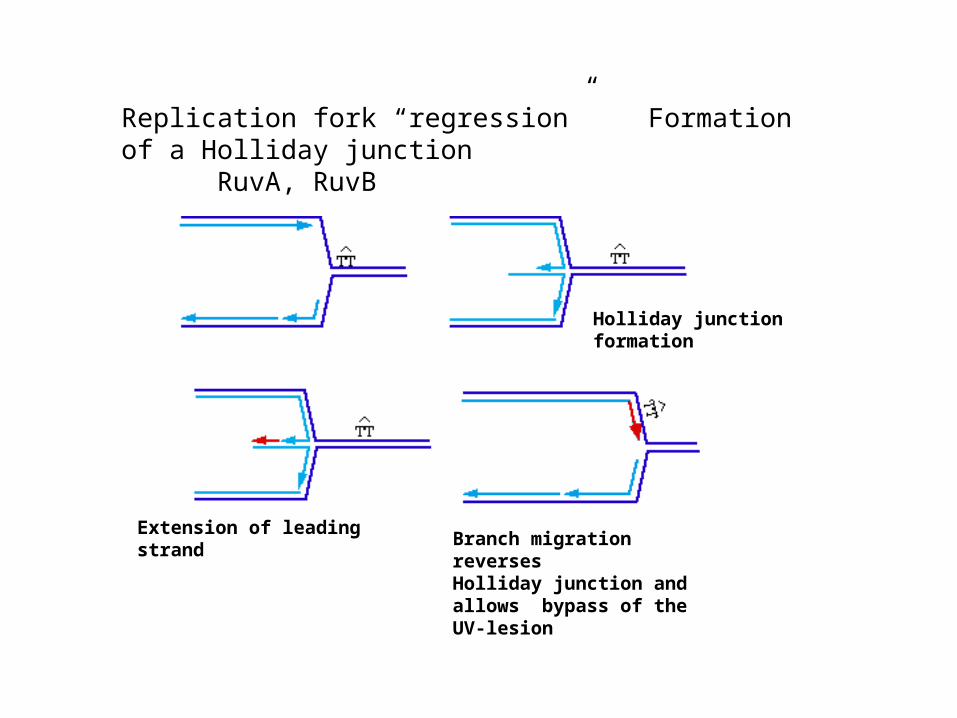
Replication fork “regression” Formation of a Holliday junctionRuvA, RuvB
Extension of leading strandBranch migration reverses Holliday junction and allows bypass of the UV-lesion
Holliday junction formation
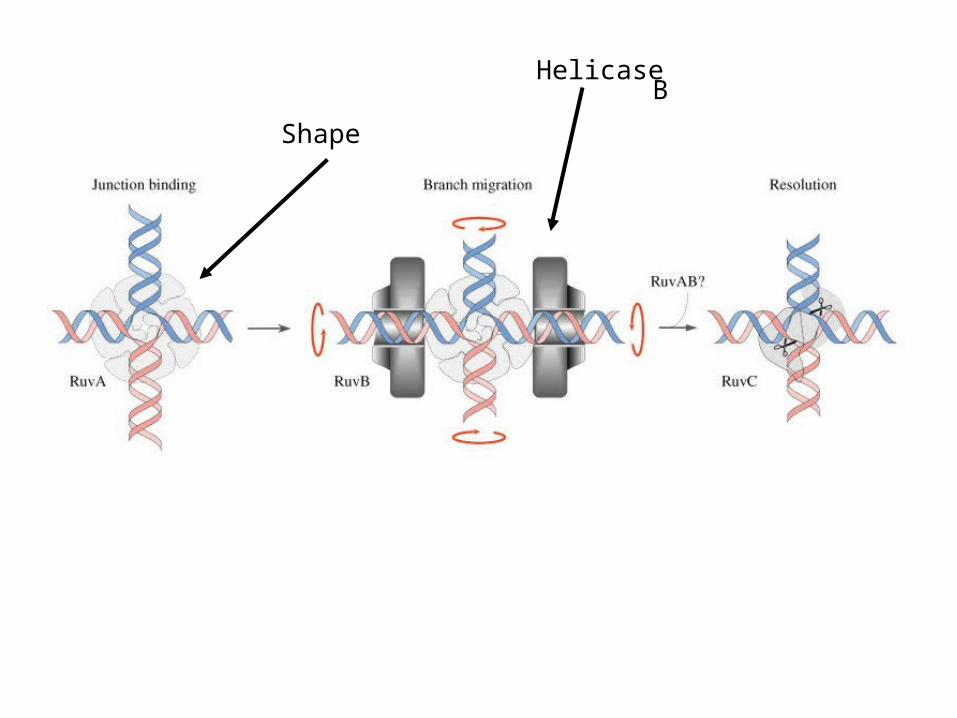
B
Shape
Helicase
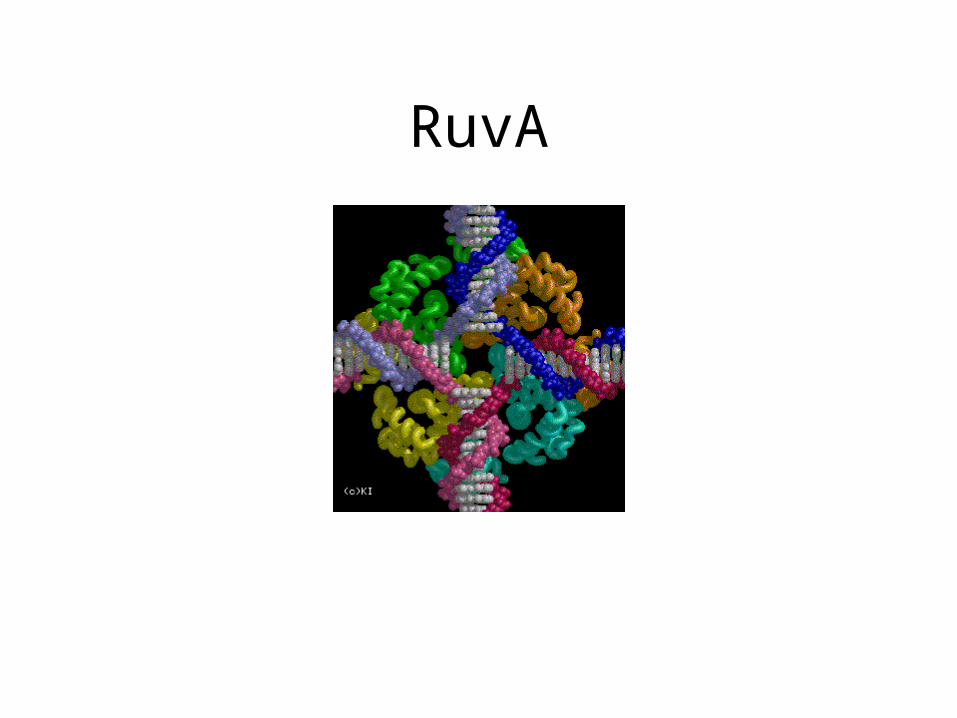
RuvA
GO TO: http://www.sdsc.edu/journals/mbb/ruva.html
RuvB

RuvC Necessary for resolution
• RuvA/B sufficient to resolve HJ with an end nearby
• RuvC cleaves symmetrically on 2 homologous DNA segments
The EMBO Journal (1997) 16, 1464–1472, doi: 10.1093/emboj/16.6.1464
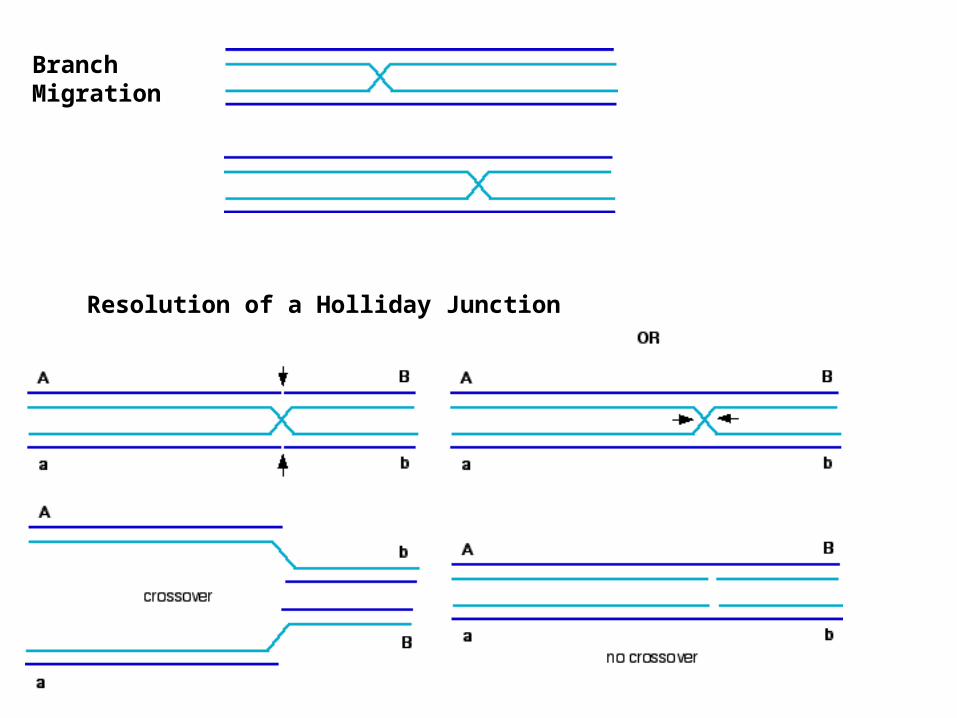
Branch Migration
Resolution of a Holliday Junction
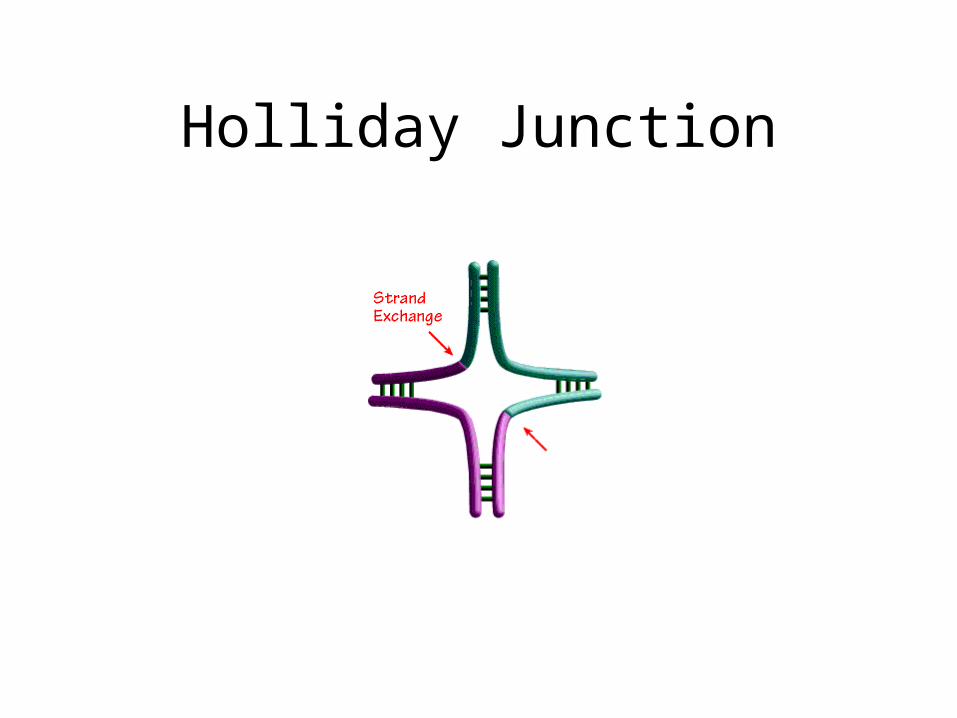
Holliday Junction
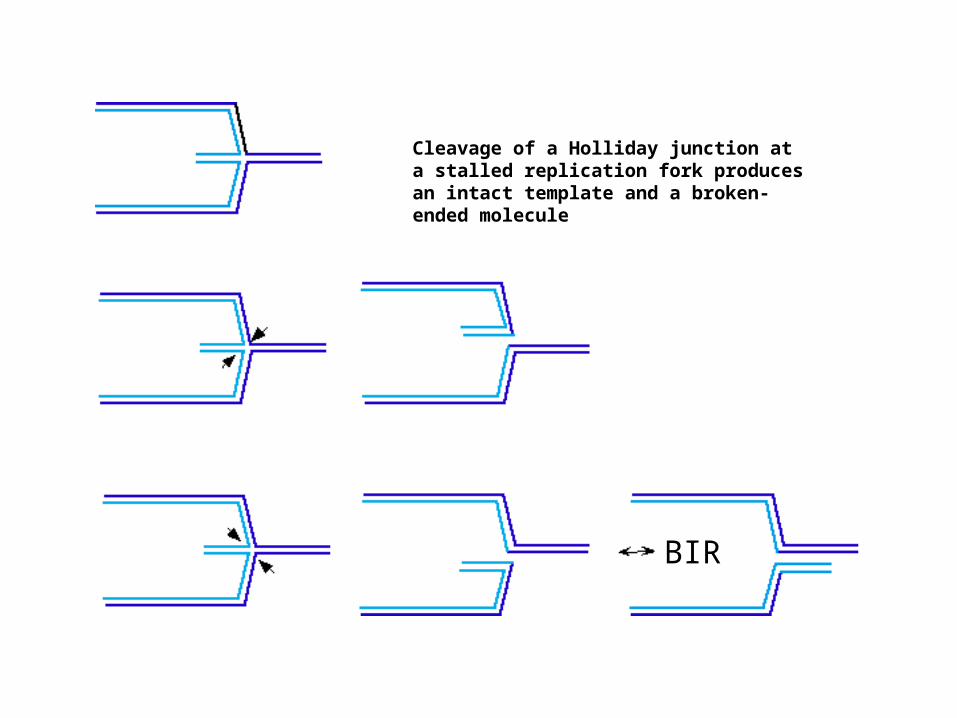
Cleavage of a Holliday junction at a stalled replication fork produces an intact template and a broken-ended molecule
BIR
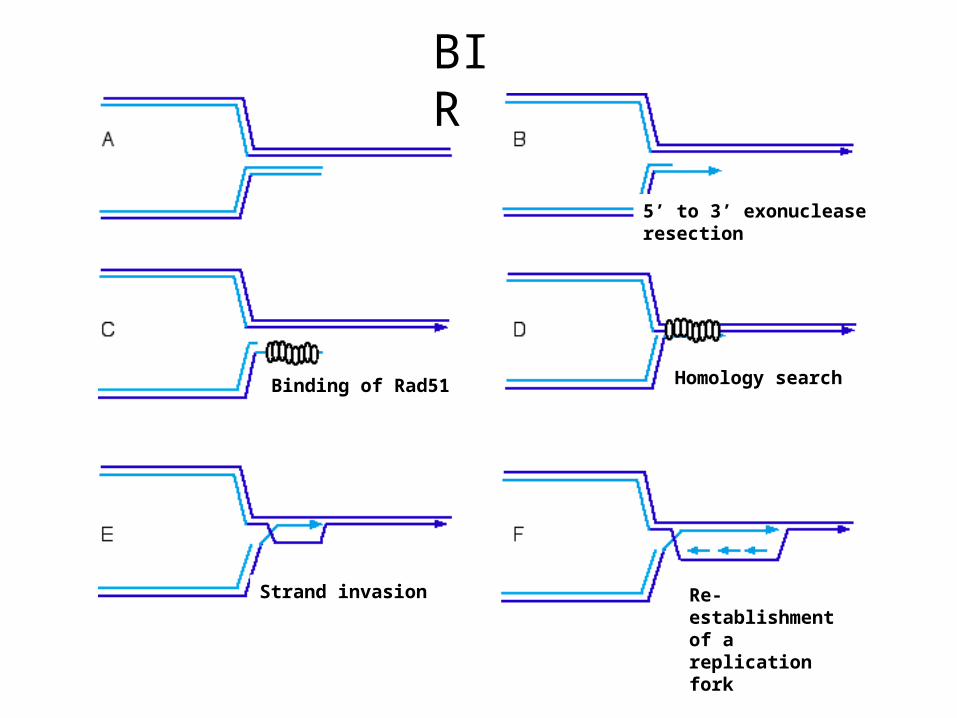
Binding of Rad51
Strand invasion
5’ to 3’ exonuclease resection
Homology search
Re-establishment of a replication fork
BIR
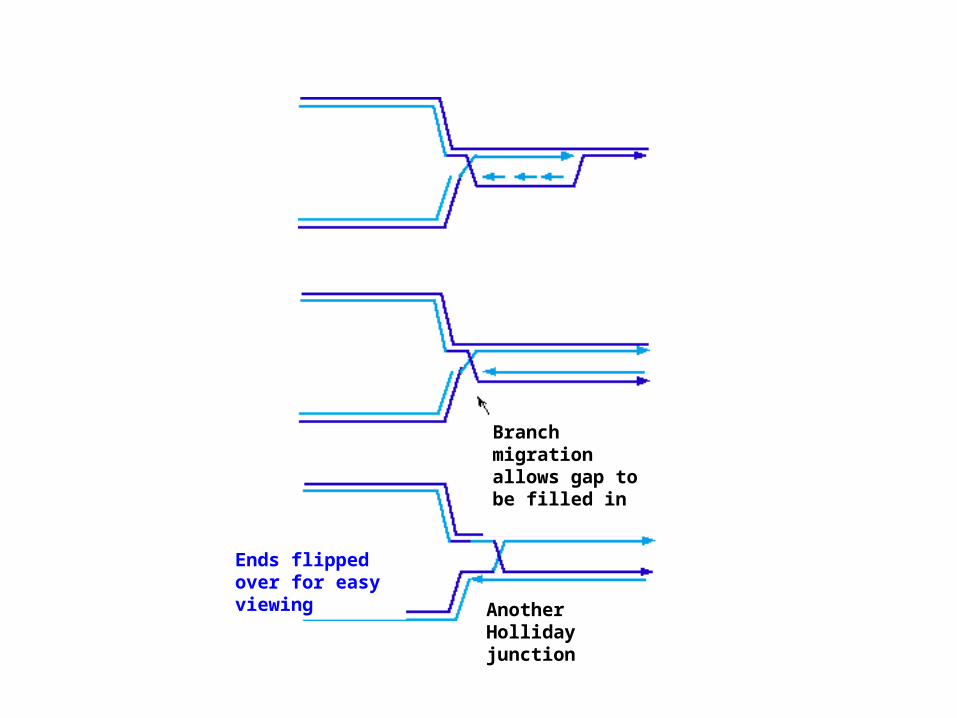
Branch migration allows gap to be filled in
Another Holliday junction
Ends flipped over for easy viewing
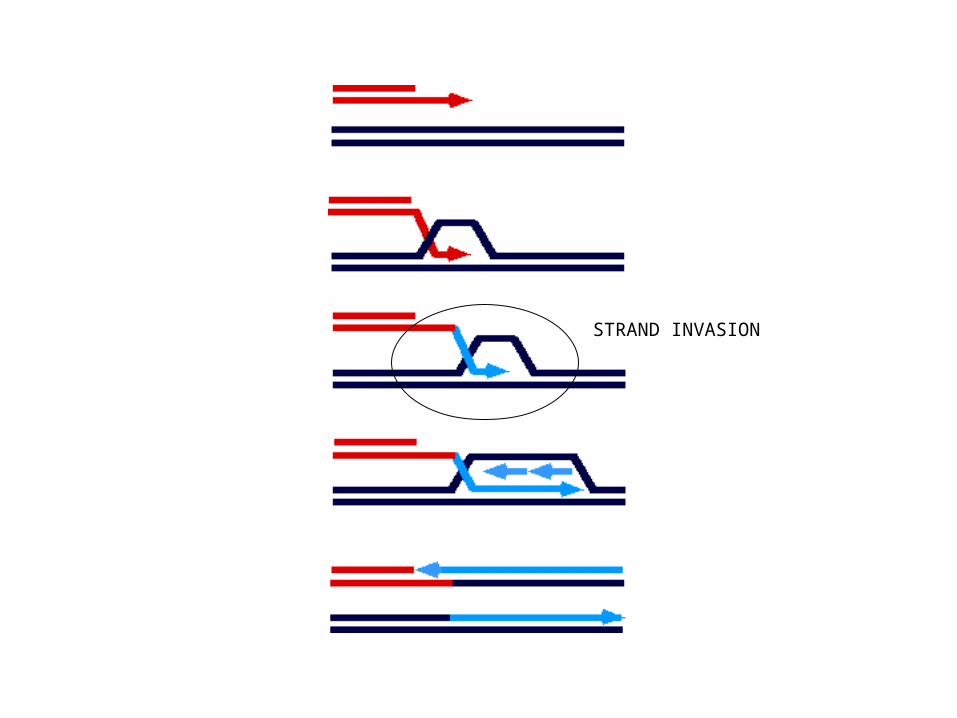
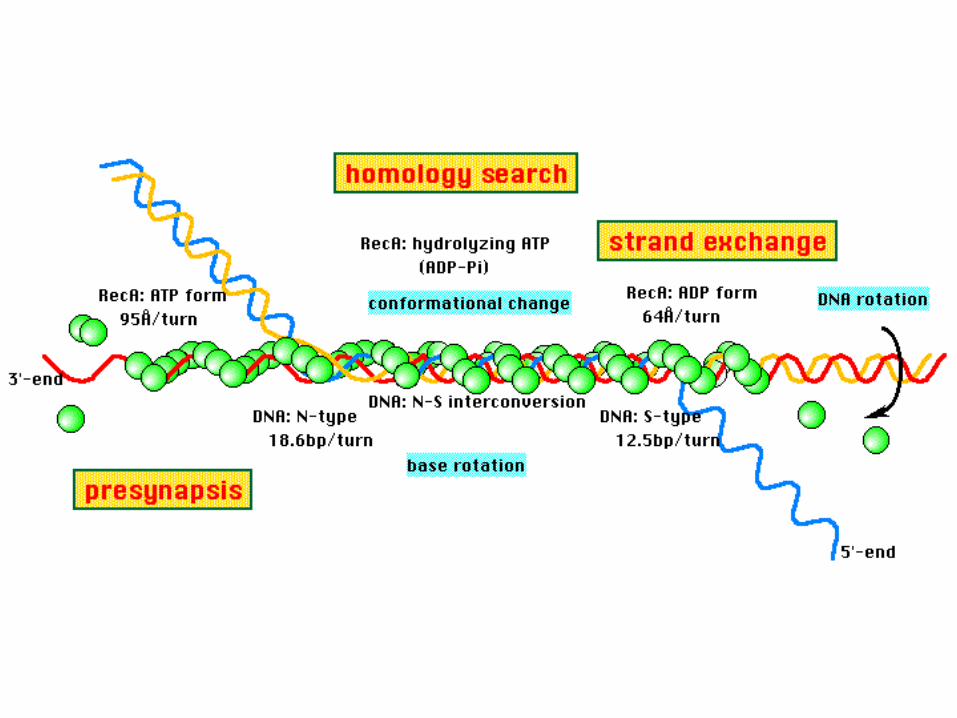
STRAND INVASION
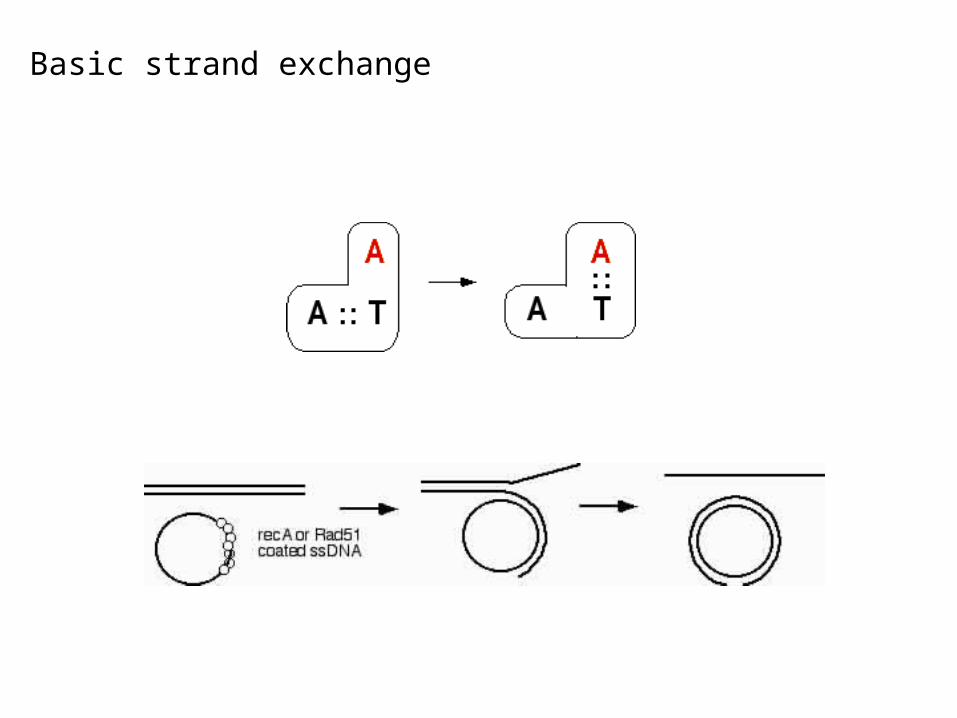
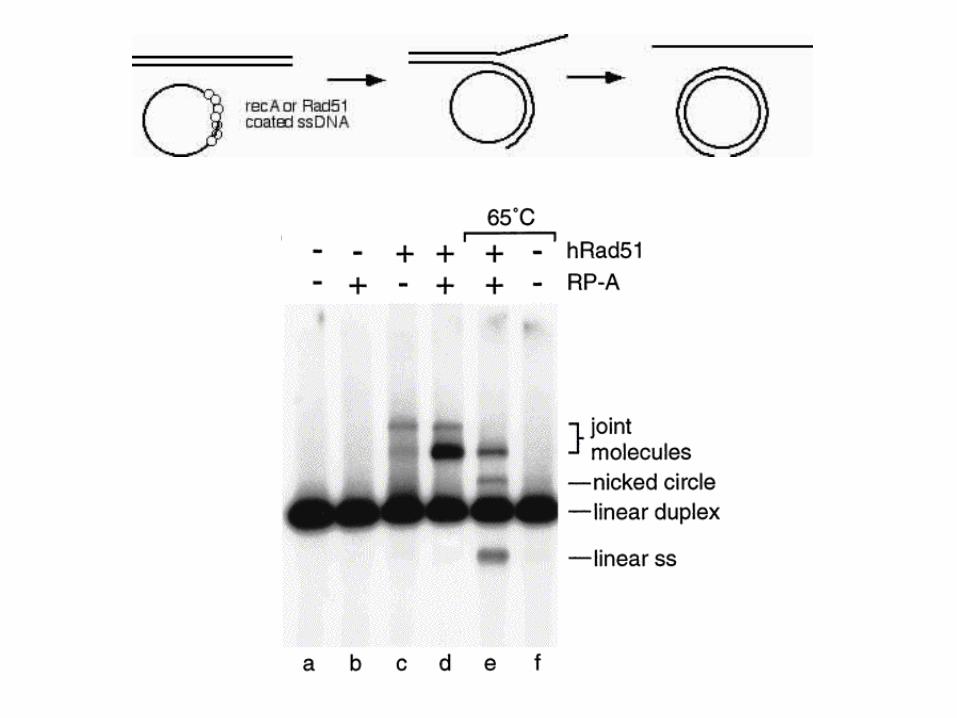
Basic strand exchange
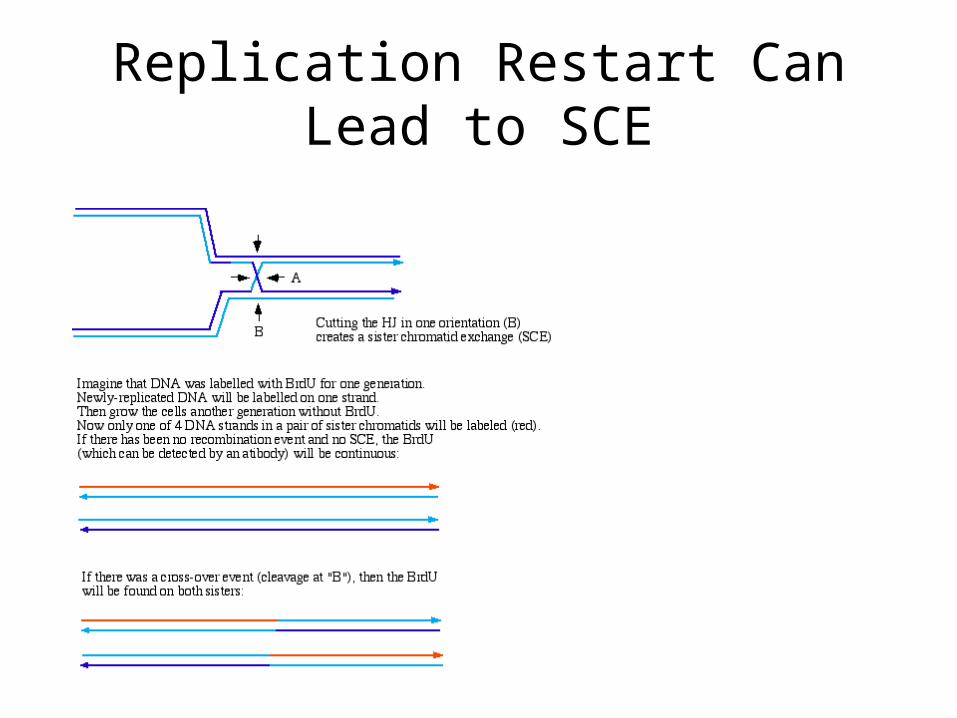
Replication Restart Can Lead to SCE


Recombination as a source of genetic instability
A hallmark of cancer cells is their genetic instability
Most of these types of instability may be explained by various mechanisms of homologous and nonhomologous recombination
In all probability the initiating lesion on DNA that leads to these changes in chromosomes can be attributed to a double-strand break (DSB)
Loss of heterozygosityReciprocal and nonreciprocal translocationsDeletions TruncationsChromosome loss
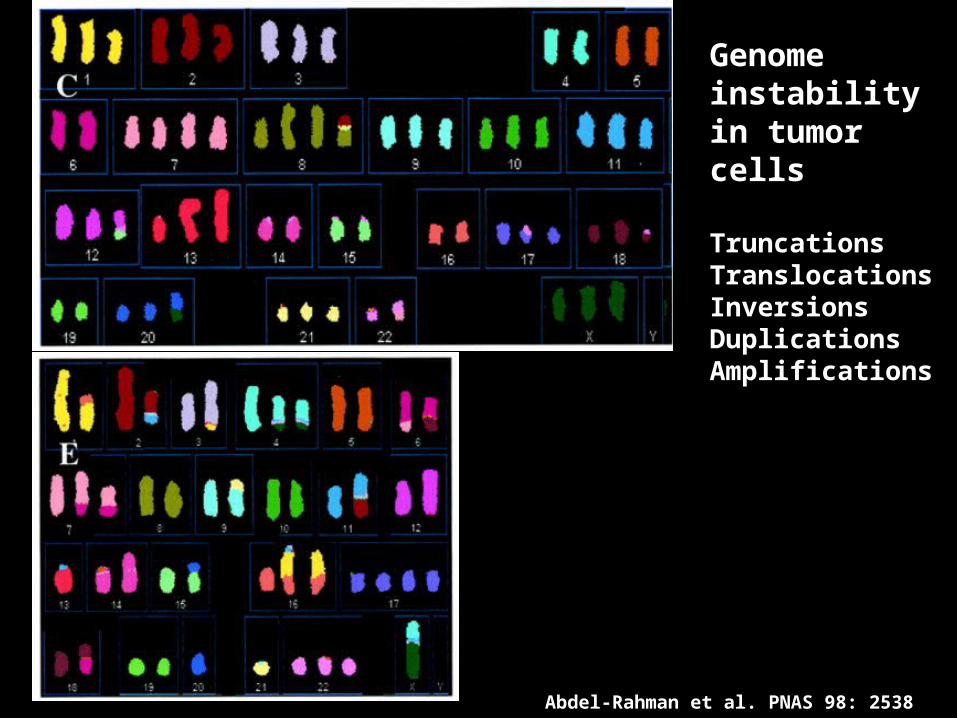
Abdel-Rahman et al. PNAS 98: 2538
Genome instabilityin tumorcells
TruncationsTranslocationsInversionsDuplicationsAmplifications

Eukaryotic recombination machinery complex
• Large number of proteins necessary– Rad51, Rad52, 5 Rad51
paralogs (load Rad51)
– Rad54 – strand invasion
– BRCA1 and BRCA2
• Rad51, BRCA1/2 KOs lethal
• Rad51 paralogs – less SCE

HOMOLOGOUS RECOMBINATION
Gene Conversion w/o Crossover
Break-Induced Replication
Gene Conversion w/ Crossover
DSB
NONHOMOLOGOUS RECOMBINATION
New telomere formation
NH End-Joining
Single-strand annealing
Nonreciprocal translocation
Gene amplification
Chromosomes and Chromatids break other times than during replication
Repaired BIR and Gene Conversion
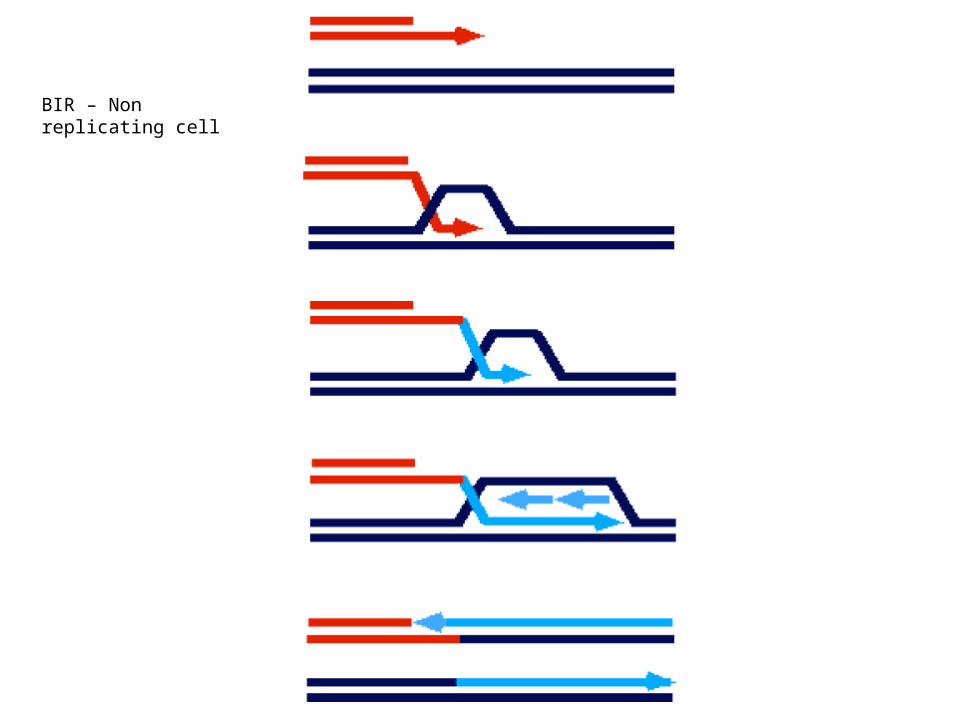
BIR – Non replicating cell
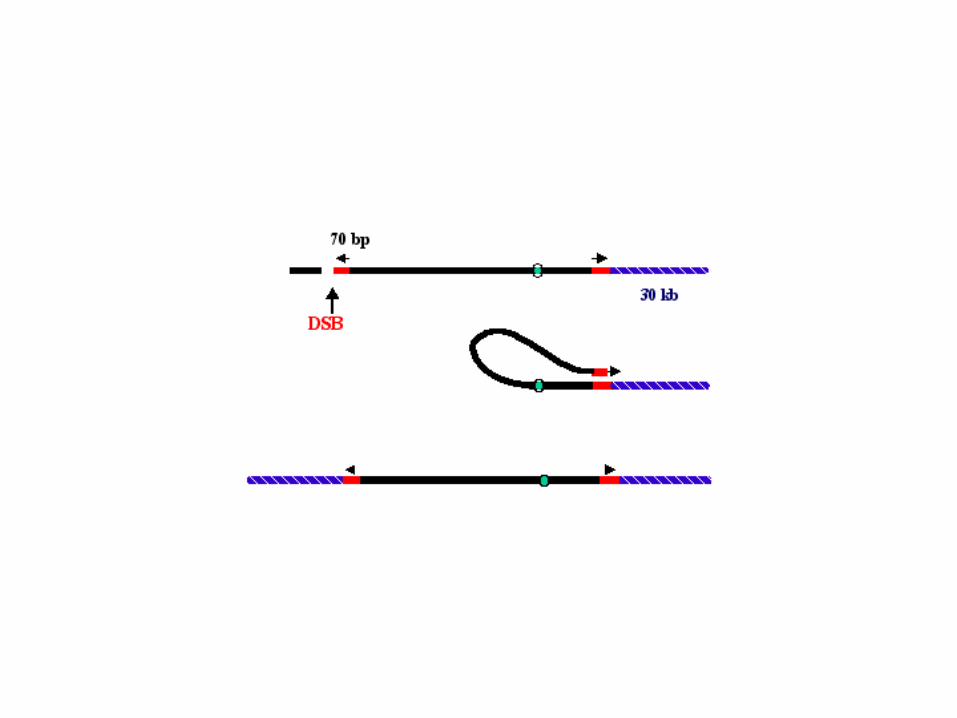
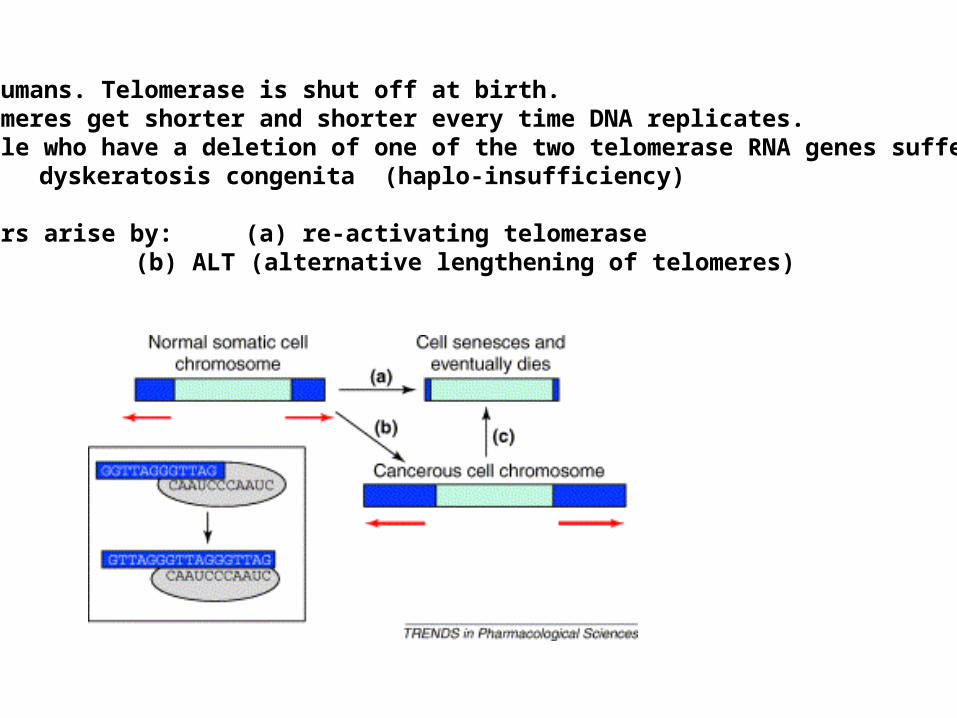
In humans. Telomerase is shut off at birth. Telomeres get shorter and shorter every time DNA replicates.People who have a deletion of one of the two telomerase RNA genes suffer from
dyskeratosis congenita (haplo-insufficiency)
Tumors arise by: (a) re-activating telomerase(b) ALT (alternative lengthening of telomeres)

ALT
• Extension telomeres by HR demonstrated yeast
• Deleted telomerase enzyme– Shorten 10nt/generation– Die 30-50 generations– Few telomerase independent survivors
• Eliminated Rad52
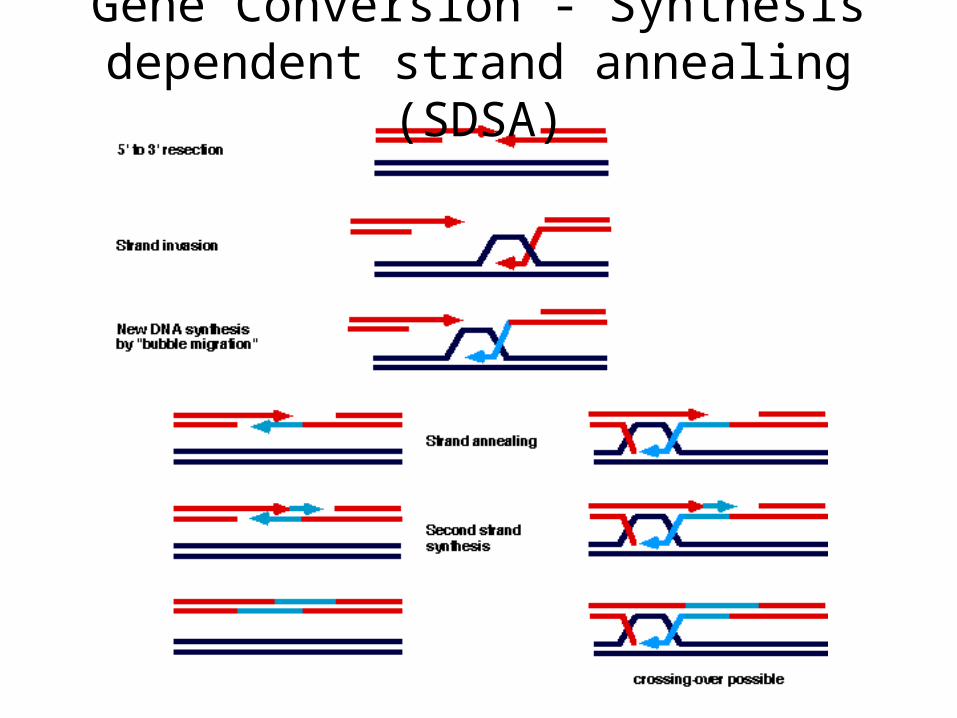
Gene Conversion - Synthesis dependent strand annealing (SDSA)
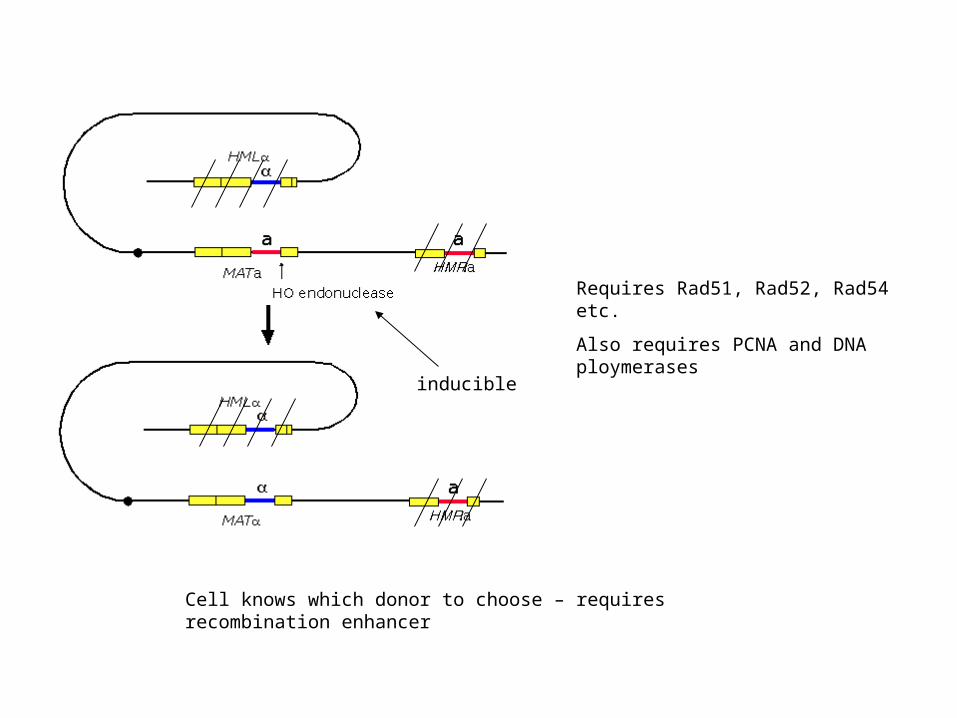
inducible
Requires Rad51, Rad52, Rad54 etc.
Also requires PCNA and DNA ploymerases
Cell knows which donor to choose – requires recombination enhancer
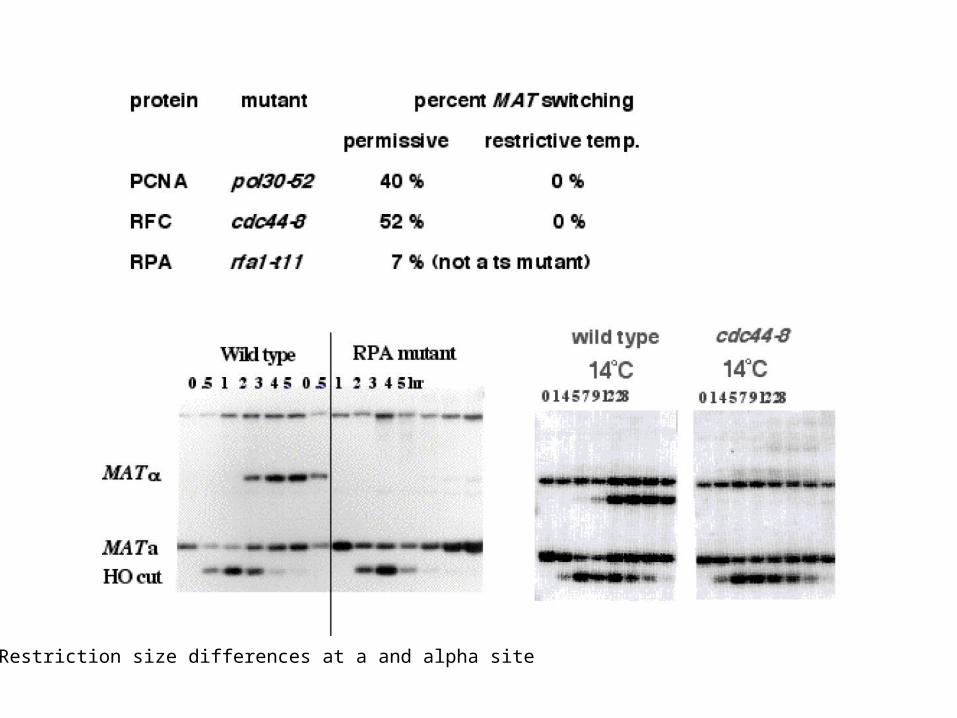
Restriction size differences at a and alpha site

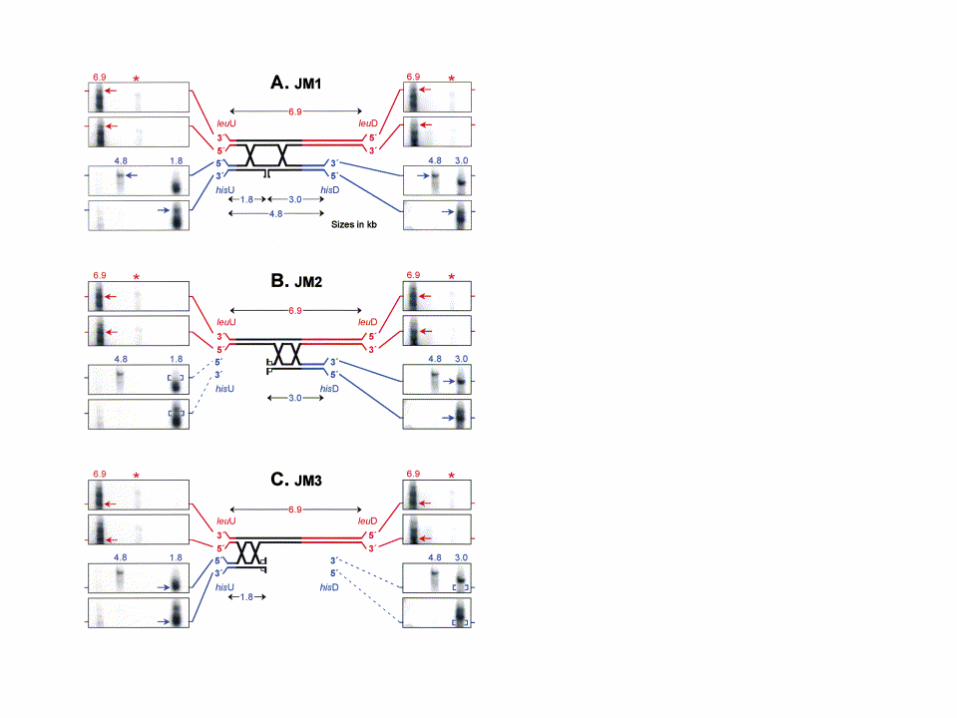
Can look for presence of HJs• Studied using 2D Gels hotspots
• Looking for differences novel spots not part of replicating arc of DNA
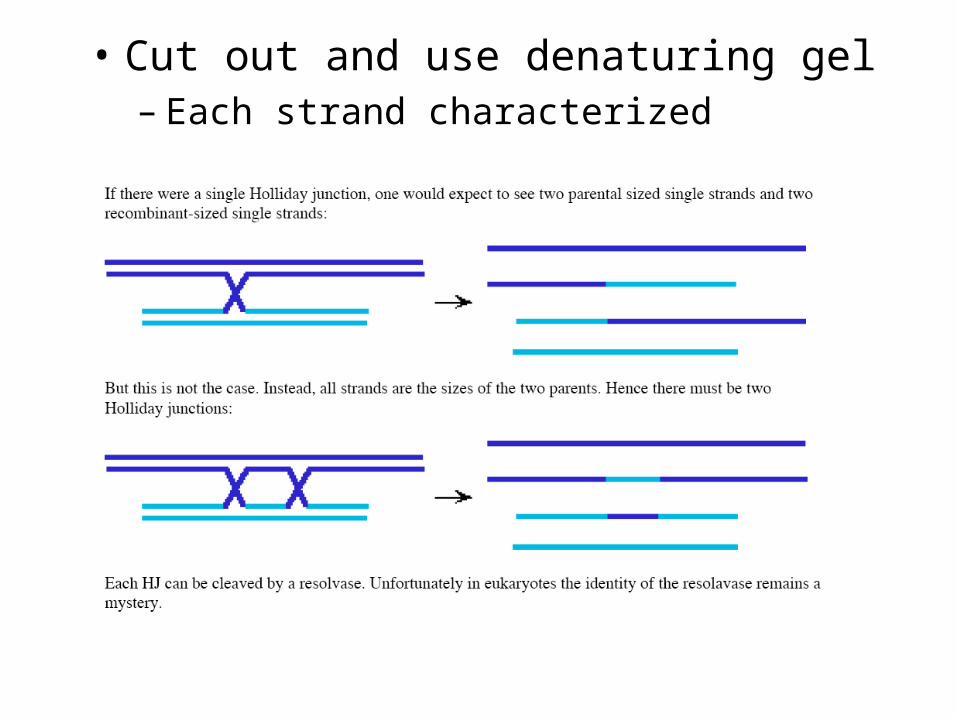
• Cut out and use denaturing gel– Each strand characterized
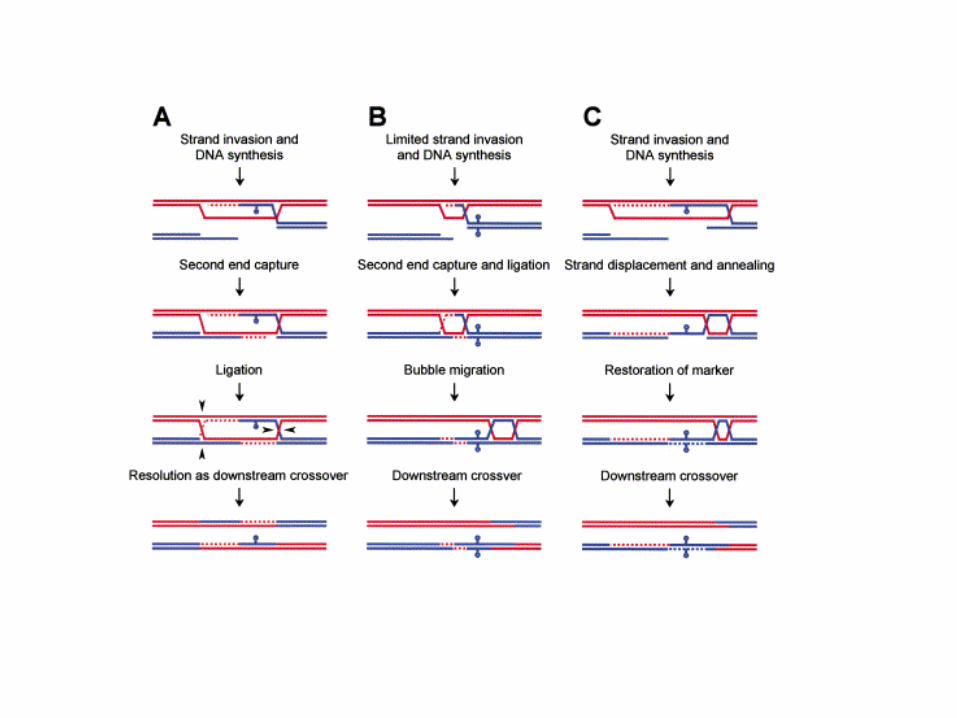

HOMOLOGOUS RECOMBINATION
Gene Conversion w/o Crossover
Break-Induced Replication
Gene Conversion w/ Crossover
DSB
NONHOMOLOGOUS RECOMBINATION
New telomere formation
NH End-Joining
Single-strand annealing
Nonreciprocal translocation
Gene amplification
Chromosomes and Chromatids break other times than during replication
Repaired BIR and Gene Conversion

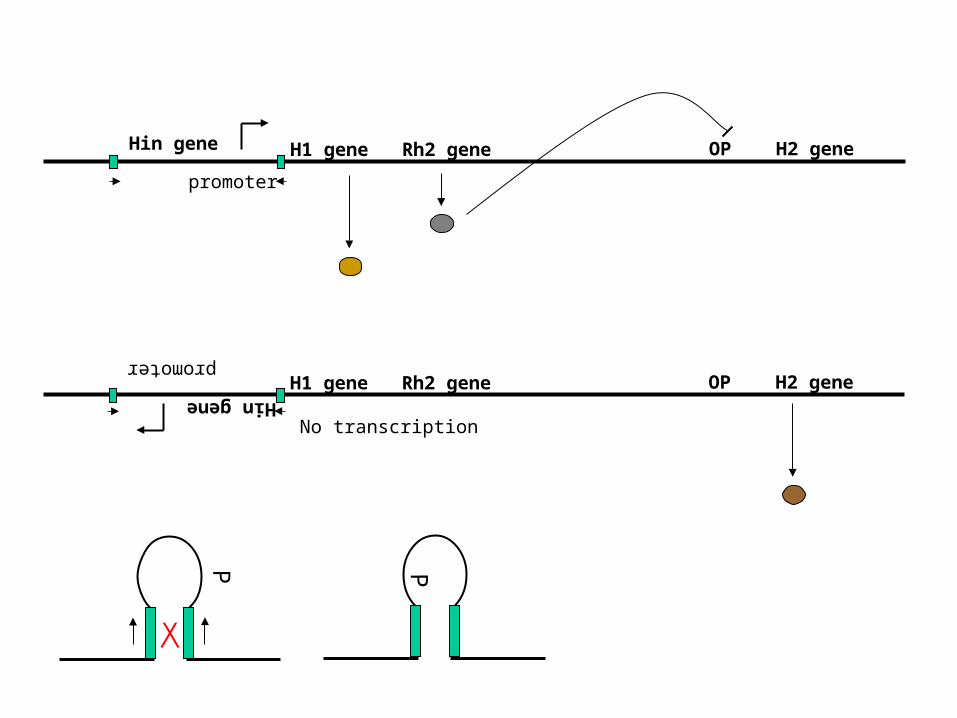
Hin gene
promoter
H1 gene Rh2 gene OP H2 gene
Hin gene
promoter
H1 gene Rh2 gene OP H2 gene
PP
No transcription
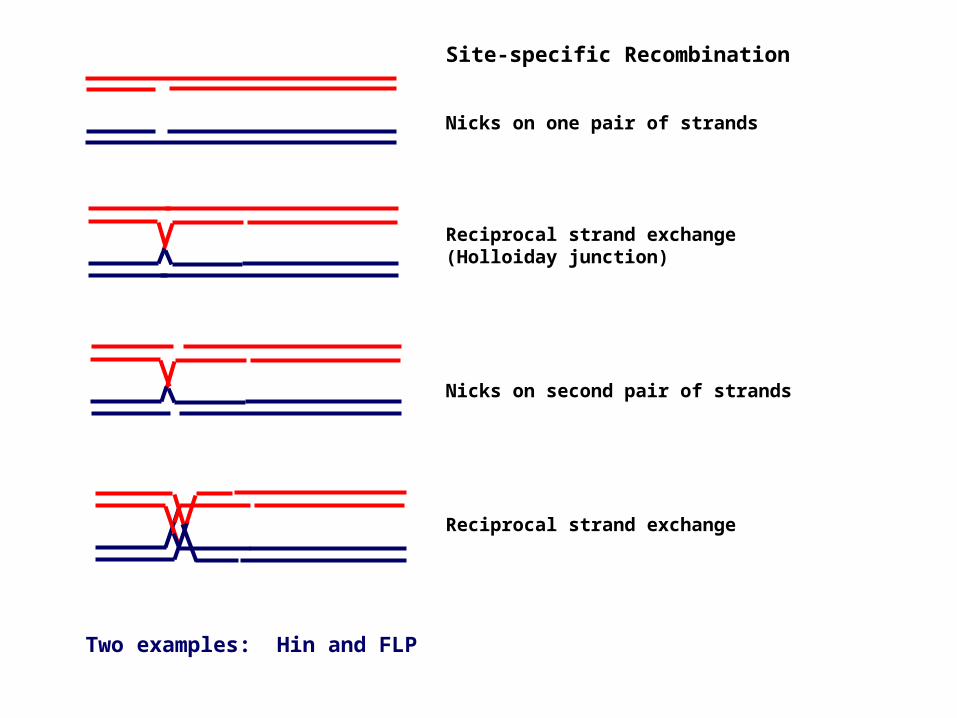
Site-specific Recombination
Nicks on one pair of strands
Reciprocal strand exchange(Holloiday junction)
Nicks on second pair of strands
Reciprocal strand exchange
Two examples: Hin and FLP

Site-specific recombination
One example: phase variation in Salmonella
Flagella composed of H1 flagellin
Anti H1 antibodyeliminates 99.99%
Survivors have alternativeH2 flagellin