morphometric correlates of the ovary and ovulatory … · morphometric correlates of the ovary and...
TRANSCRIPT

Morphometric Correlates of the Ovaryand Ovulatory Corpora in the Bowhead
Whale, Balaena mysticetusRAYMOND J. TARPLEY,1* DANIEL J. HILLMANN,2 JOHN C. GEORGE,3
JUDITH E. ZEH,4 AND ROBERT S. SUYDAM5
1Department of Veterinary Integrative Biosciences, College of Veterinary Medicine andBiomedical Sciences, Texas A&M University, College Station, Texas
2Department of Comparative Biomedical Sciences, School of Veterinary Medicine, Louisi-ana State University, Baton Rouge, Louisiana
3Department of Wildlife Management, North Slope Borough, Barrow, Alaska4Department of Statistics, University of Washington, Seattle, Washington
5Department of Wildlife Management, North Slope Borough, Barrow, Alaska
ABSTRACTGross morphology and morphometry of the bowhead whale ovary,
including ovulatory corpora, were investigated in 50 whales from theChukchi and Beaufort seas off the coast of Alaska. Using the presenceof ovarian corpora to define sexual maturity, 23 sexually immaturewhales (7.6–14.2 m total body length) and 27 sexually mature whales(14.2–17.7 m total body length) were identified. Ovary pair weightsranged from 0.38 to 2.45 kg and 2.92 to 12.02 kg for sexually imma-ture and sexually mature whales, respectively. In sexually maturewhales, corpora lutea (CLs) and/or large corpora albicantia (CAs) pro-jected beyond ovary surfaces. CAs became increasingly less interrup-tive of the surface contour as they regressed, while remainingidentifiable within transverse sections of the ovarian cortex. CLsformed large globular bodies, often with a central lumen, featuringgolden parenchymas enfolded within radiating fibrous cords. CAs,sometimes vesicular, featured a dense fibrous core with outwardfibrous projections through the former luteal tissue. CLs (never morethan one per ovary pair) ranged from 6.7 to 15.0 cm in diameter in 13whales. Fetuses were confirmed in nine of the 13 whales, with theassociated CLs ranging from 8.3 to 15.0 cm in diameter. CLs from fourwhales where a fetus was not detected ranged from 6.7 to 10.6 cm indiameter. CA totals ranged from 0 to 22 for any single ovary, and from1 to 41 for an ovary pair. CAs measured from 0.3 to 6.3 cm in diame-ter, and smaller corpora were more numerous, suggesting an accumu-lating record of ovulation. Neither the left nor the right ovarydominated in the production of corpora. Anat Rec, 299:769–797,2016. VC 2016 Wiley Periodicals, Inc.
Grant sponsors: North Slope Borough Department of WildlifeManagement, Barrow, AK, to Texas A&M University; Grantnumbers: C3469, E1621, 20177, 16840, 83-58, 58-58, 84-91, 85-62, 86-61, 87-123, 88-134, 93-293, 19980490, 19981343, and1999226; Grant sponsor: U.S. National Marine Fisheries Serv-ice Scientific Permits; Grant numbers: 345 and 519 (to T.F.Albert) and 780 (issued to R.J. Tarpley).
*Correspondence to: Raymond J. Tarpley, 701 Lincoln Ave.,College Station, TX 77840. Fax: 979-764-9193. E-mail: [email protected]
Received 24 January 2015; Revised 23 November 2015;Accepted 29 December 2015.
DOI 10.1002/ar.23337Published online 24 February 2016 in Wiley Online Library(wileyonlinelibrary.com).
THE ANATOMICAL RECORD 299:769–797 (2016)
VVC 2016 WILEY PERIODICALS, INC.

Key words: Cetacea; Balaena mysticetus; bowhead whale;ovary morphology; fetus
INTRODUCTION
The reproductive system of the great whales has his-torically been a focus of morphological study for thoseseeking to define the productivity and sustainability ofwhale populations being pressured by the commercialwhaling industry (Mackintosh and Wheeler, 1929; Mat-thews, 1937, 1938; Dempsey and Wislocki, 1941; vanLennep, 1950; Robins, 1954; Chittleborough, 1954; Laws,1957, 1961; Ohsumi, 1964; Slijper, 1949, 1966; Best,1967, 1977, 1982; Gambell, 1968, 1972; Lockyer, 1984,1987). Most of these investigations were conducted dur-ing the peak whaling periods of the early and mid-20thcentury when specimen materials suitable for accurateexamination and interpretation were more readily avail-able. Consequently, the literature reflects the species ofinterest to commercial whaling at the time, and therorquals (the balaenopterids) have figured most promi-nently in discussions of mysticete reproduction.
In contrast, commercial exploitation of the right whalefamily (Balaenidae), of which the bowhead whale is apart, occurred largely before adequate investigative tech-niques were available or attempted. As a result, lessdata address reproductive function in the balaenids, andthe need still exists to better define parameters ofestrous cyclicity. Since the 1970s, collection of reproduc-tive structures has been feasible in the bowhead whale,Balaena mysticetus, from subsistence-harvested whalestaken by Alaskan Eskimo hunters.
Such information is relevant to management decisionsconcerned with bowhead whale conservation. A recentabundance study estimated 16,820 bowheads in theBering, Chukchi, Beaufort seas, with a 95% confidenceinterval of 15,176 to 18,643 (Givens et al., 2016). Manage-ment strategies aimed at conserving bowhead whalestocks are founded upon knowledge of population sizeand trend, as well as biological information such as indi-ces of reproduction, including ovulation frequency, lengthof gestation, length of lactation, and the calving interval.Length at sexual maturity is also useful in aerial photo-grammetric surveys to estimate the proportion of matureanimals that are potentially contributing to populationgrowth (Angliss et al., 1995; Koski et al., 2006). An impor-tant preliminary consideration of these parameters waspresented by Nerini et al. (1984) based on data gatheredlargely from 1973 to 1981. Since that time, collection ofmaterials supporting ongoing studies of bowhead whalereproduction have continued under the supervision of theNorth Slope Borough Department of Wildlife Manage-ment in Barrow, Alaska (Albert, 1985; George et al.,2011). Here we report findings on gross morphology andmorphometric relationships in the bowhead whale ovary,including quantitative correlates of ovarian corpora.
MATERIALS AND METHODS
Ovary pairs were collected from 48 bowhead whales(and a single ovary from each of two additional whales)
at the harvest sites of seven northern and western Alas-kan villages during the spring and fall subsistence huntsfrom 1980 through 1993 (Table 1). After designatingright or left, if known (A or B if not), ovaries wereimmersed whole (without cutting) in 10% phosphate buf-fered formalin at the harvest site or at the field labora-tory. Total body length (measured in a straight line fromthe tip of the rostrum to the notch of the tail flukes) wasdetermined to the nearest decimeter after the whale waspulled onto the ice (spring) or beach (fall). Whales wererecorded as being pregnant, lactating, recently ovulated,or resting (neither pregnant nor lactating and lacking aCL) based on ovary morphology and observations at theharvest site (Table 1). Fetal body lengths and sex wererecorded.
Ovaries were later weighed, measured, (length, width,height), photographed, and examined grossly with par-ticular attention to externally-evident corpora lutea(CLs) and corpora albicantia (CAs) as indicators of sex-ual maturity (Tables 2 and 3). Forty-nine of 52 sexuallymature ovaries were sliced transverse to their long axesat approximately 5.0 mm intervals using an electricmeat cutter (supplemented with an electric knife asneeded) to expose the cortex and regressed CAs (Fig.1a,b). Where preservation was sufficient, CAs werecounted, serially tabulated and photographed along eachovary’s long axis.
CLs and CAs were identified and tabulated for all ova-ries collected among 13 and 23 whales, respectively. CAswere numbered in serial order through the length of theovary. The diameters of two CLs from two whales wereestimated in the field. Comparative quantitative meas-ures of 11 CLs from 11 whales and 257 CAs from 22whales were determined by tracing the approximatemidsection perimeter of each corpus using computer-based morphometry applications (Bioquant System IVVC;Universal ImagingVC ) to determine the total cross-sectional area of the corpus (as a proxy for corpus size),as well as its fractions of former luteal tissue and itscentral scar or lumen (Fig. 2a,b; Tables 4 and 5). Diame-ter equivalents were calculated (A 5 pr2) from areameasurements to facilitate comparisons with otherreports in the literature where corpora size classes arepresented as diameter values. CA diameters were usedto define regression size frequencies.
RESULTS
Ovary Morphology
Bowhead whale ovaries were elongate and bluntlyfusiform, tapering toward rounded poles, presenting twoextremities, two borders, and two surfaces, with specificphysical features varying with the degree of maturity.Sexually immature ovaries were smaller than matureones, with those least developed being considerably com-pressed between the two, relatively smooth, surfaces(Fig. 3). The larger immature ovaries were increasingly
770 TARPLEY ET AL.

robust, thickening between the two surfaces as the latterbecame more complex with a variable pattern of infold-ings (Fig. 4). Each of the two extremities (or poles) ofthe ovary was formed by the tapered convergence of thesurfaces and borders. The flattened mesovarial insertionextended nearly along the entire length of the hilar bor-der. The latter border produced two long edges as the
ovarian surface curved inward from each side to join themesovarium. Vessels and nerves passed into and out ofthe full length of the ovary’s medulla via the hilar bor-der, traveling within the fibrous and fatty mesovarium.
Mature ovaries were larger, more robust, and thick-ened, with much less compression between the surfaces.Surface infoldings were interrupted by fluid-filled
TABLE 1. Bowhead whale ovaries collected 1980–1992 (sorted by village, year, whale)
WhaleMonthcaught
Bodylength (m)
Ovariescollected
Maturitystatus
Estrouscycle phase
Fetuslength (cm)
Barrow80B7 May 10.0 AB I - -80B9 May 13.6 AB I - -81B1 May 10.7 LR I - -81B2 May 8.0 LR I - -81B3 May 8.4 LR I - -83B1 May 8.5 LR I - -83B2 May 14.2 LR I - -84B2 May 8.5 LR I - -84B4 May 8.3 LR I - -86B3 Apr 8.9 LR I - -86B6 May 12.3 LR I - -87B5 Jun 15.7 LR M Pregnant 40587B6 Oct 15.7 LR M Lactating -88B1 Apr 8.9 LR I - -88B4 Apr 9.0 LR I - -88B6 May 8.3 LR I - -88B7 May 8.2 LR I - -89B2 May 14.7 LR M Pregnant 40189B3 May 16.9 LR M Resting -90B4 May 14.9 LR M Pregnant 39092B2 May 15.6 R M Resting -92B3 Aug 14.6 AB M Resting -92B4 Sep 16.2 AB M Resting -92B7 Sep 14.2 AB M Pregnant 7592B8 Sep 15.7 AB M Pregnant 8492B9 Sep 15.0 AB M Resting -92B10 Sep 14.6 LR M Resting -92B12 Sep 12.0 LR I - -92B13 Sep 15.0 R M Pregnant 145
Gambell80G1 May 15.7 LR M Recent Ovulation -82G2 May 8.8 LR I - -88G1 Apr 15.7 AB M Pregnant 434
Pt. Hope81H2 May 11.7 LR I - -81H3 May 9.8 LR I - -
Kaktovik81KK1 Sep 17.4 LR M Resting -81KK3 Sep 16.2 LR M Resting -84KK1 Sep 9.8 AB I - -86KK1 Sep 7.6 LR I - -86KK2 Sep 17.2 LR M Resting -88KK1 Sep 14.9 LR M Pregnant 150
Nuiqsut87N1 Oct 15.2 AB M Pregnant 160
Savoonga81S2 Apr 14.2 LR M Recent Ovulation -82S1 Apr 13.4 AB I - -
Wainwright81WW3 May 16.2 LR M Recent Ovulation -82WW1 May 17.6 LR M Resting -82WW2 May 16.5 LR M Lactating -85WW1 May 16.2 LR M Resting -85WW2 May 15.1 LR M Recent Ovulation -86WW2 May 17.7 AB M Pregnant 2787WW4 Jun 11.4 AB I - -
CORRELATES OF OVARY MORPHOMETRICS IN THE BOWHEAD WHALE 771

follicles, CLs and/or CAs, giving the ovary a “lumpy,”irregular external appearance (Fig. 5). Like the imma-ture ovaries, the flattened mesovarium extended nearlythe entire length of the hilar border (Fig. 6). The pres-ence of CLs and/or CAs was used to confirm sexualmaturity. On this basis, 23 whales were categorized assexually immature, and 27 whales were designated assexually mature. Corpora varied considerably in size,depending on the stage of involution, from a fully devel-oped CL to a small CA represented by a hyaline scar(Figs. 7 and 8). CLs and the larger CAs were externallyevident as protruding masses or as spherical or ovalendurated areas marked by the former site of ovulation,or stigma (Fig. 5). In transverse section, sexually matureovaries featured fluid-filled follicles and regressing cor-pora along the peripheral contour within the corticalregion.
All CLs were globular, although specific shapes var-ied, and some possessed a central lumen (Fig. 9).Grossly, all CLs were distinguished by their large rela-tive size and a yellowish parenchyma typical of luteinpigment (Figs. 5 and 7). A single CL was present in eachof 13 whales; CLs were sectioned in 11 whales andranged in cross-sectional area from 35.1 to 175.7 cm2
and in calculated diameter from 6.7 to 15.0 cm; CLswere field-measured in two whales with diameters of9.6 cm and 15.0 cm. The two 15.0 cm CLs supportedearly-gestational (spring-caught) and mid-gestational(fall-caught) fetuses. Nine of the 13 whales with a CLwere spring-caught; the remaining four were taken dur-ing the fall. A fetus was confirmed in five spring-caughtwhales and in four fall-caught whales. Fetuses rangedfrom 27.0 to 434.0 cm in total body length. Fetuses(n 5 5) with body lengths in the lower and upper rangeof spectrum (27 cm or 390–434 cm, respectively) wererecovered from spring-caught whales. Fetuses (n 5 4)that were mid-range in body length (75–160 cm) werefrom fall-caught whales. All four whales having a CLwith no detected fetus were spring-caught.
All CAs were smaller than any of the CLs, ranging insize from 0.3 to 6.3 cm, generally consisting of a collage-nous scar with radiating trabeculae that penetrated anarray of spiral arteries representing the former lutealparenchyma. Some had a central cavity, but most had asolid core of collagen (Fig. 10a,b). The larger CAs pro-truded prominently from the ovarian surface, marked bythe former site of ovulation (stigma) (Fig. 11). The shapeof CAs was variable (Fig. 12). The largest CA (6.3 cm)was almost spherical, whereas others (e.g., those in theother ovary of the same whale) were somewhat flattenedin one dimension (Fig. 11). The position of some of theregressing CAs that no longer protruded above the
TABLE 2. Body and ovary morphometrics in sexuallyimmature bowhead whales (sorted by body length)
Whale
Bodylength
(m) Ovarya
Ovarylength(cm)
Ovaryweight
(kg)
86KK1 7.6 L 21.0 0.23R 21.0 0.29
L&R 42.0 0.5281B2 8.0 L 21.5 0.28
R 20.5 0.33L&R 42.0 0.61
88B7 8.2 L 21.4 0.45R 23.0 0.45
L&R 44.4 0.9084B4 8.3 L 21.5 0.74
R 23.8 0.78L&R 45.3 1.52
88B6 8.3 L 17.4 0.36R 17.6 0.36
L&R 35.0 0.7281B3 8.4 L 18.5 0.26
R 19.0 0.33L&R 37.5 0.59
83B1 8.5 L 23.8 0.55R 23.0 0.51
L&R 46.8 1.0684B2 8.5 L 17.5 0.23
R 17.8 0.30L&R 35.3 0.53
82G2 8.8 L 17.0 0.17R 21.0 0.21
L&R 38.0 0.3886B3 8.9 L 17.2 0.45
R 18.0 0.34L&R 35.2 0.79
88B1 8.9 L 20.2 0.41R 21.6 0.41
L&R 41.8 0.8288B4 9.0 L 25.2 0.59
R 24.0 0.45L&R 49.2 1.04
81H3 9.8 L 24.5 0.45R 19.0 0.40
L&R 43.5 0.8584KK1 9.8 A 18.5 0.29
B 16.0 0.29A&B 34.5 0.58
80B7 10.0 A 21.0 0.62B 18.0 0.74
A&B 39.0 1.3681B1 10.7 L 24.5 0.57
R 19.0 0.51L&R 43.5 1.08
87WW4 11.4 A 13.6 0.34B 14.5 0.36
A&B 28.1 0.7081H2 11.7 L 28.0 0.65
R 31.0 0.77L&R 59.0 1.42
92B12 12.0 L 26.7 0.88R 29.0 0.85
L&R 55.7 1.7386B6 12.3 L 32.0 1.14
R 30.0 1.31L&R 62.0 2.45
82S1 13.4 A 27.0 0.65B 29.0 0.61
A&B 56.0 1.2680B9 13.6 A 28.5 0.63
B 31.0 0.71
TABLE 2. (continued).
Whale
Bodylength
(m) Ovarya
Ovarylength(cm)
Ovaryweight
(kg)
A&B 59.5 1.3483B2 14.2 L 19.6 0.68
R 20.1 0.68L&R 39.7 1.36
aOvaries are designated left (L) or right (R) when known; Aor B when not known.
772 TARPLEY ET AL.
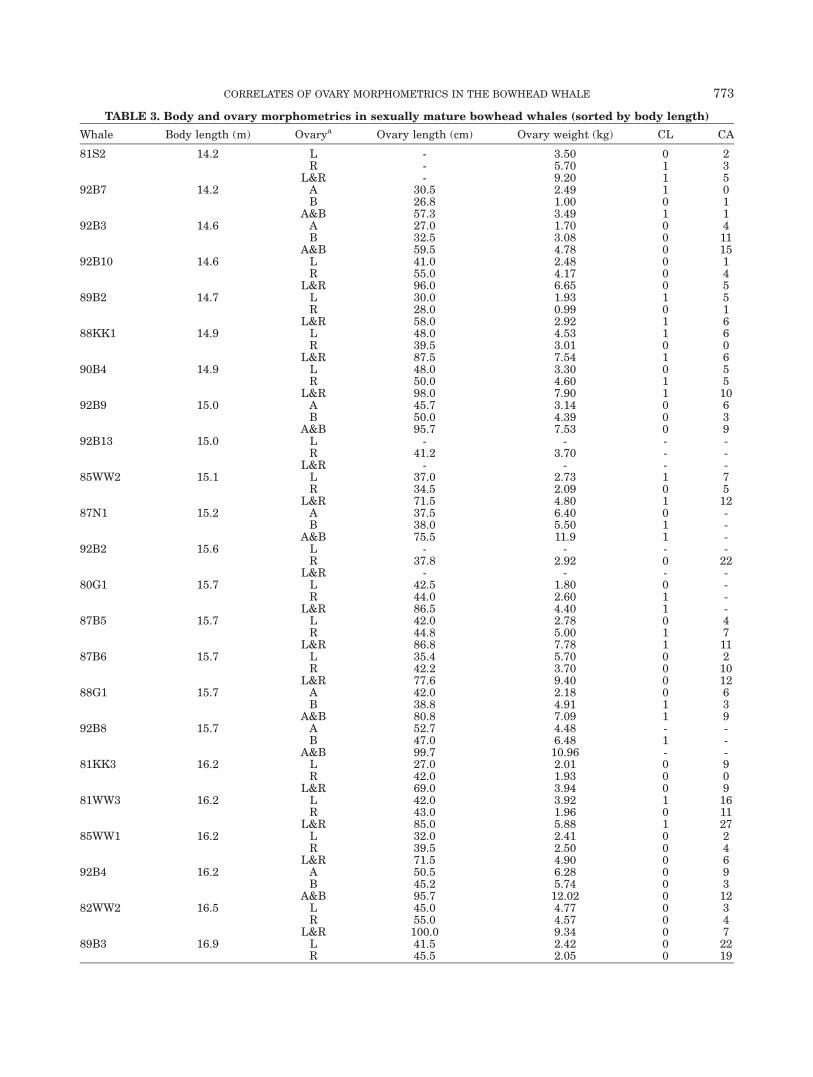
TABLE 3. Body and ovary morphometrics in sexually mature bowhead whales (sorted by body length)
Whale Body length (m) Ovarya Ovary length (cm) Ovary weight (kg) CL CA
81S2 14.2 L - 3.50 0 2R - 5.70 1 3
L&R - 9.20 1 592B7 14.2 A 30.5 2.49 1 0
B 26.8 1.00 0 1A&B 57.3 3.49 1 1
92B3 14.6 A 27.0 1.70 0 4B 32.5 3.08 0 11
A&B 59.5 4.78 0 1592B10 14.6 L 41.0 2.48 0 1
R 55.0 4.17 0 4L&R 96.0 6.65 0 5
89B2 14.7 L 30.0 1.93 1 5R 28.0 0.99 0 1
L&R 58.0 2.92 1 688KK1 14.9 L 48.0 4.53 1 6
R 39.5 3.01 0 0L&R 87.5 7.54 1 6
90B4 14.9 L 48.0 3.30 0 5R 50.0 4.60 1 5
L&R 98.0 7.90 1 1092B9 15.0 A 45.7 3.14 0 6
B 50.0 4.39 0 3A&B 95.7 7.53 0 9
92B13 15.0 L - - - -R 41.2 3.70 - -
L&R - - - -85WW2 15.1 L 37.0 2.73 1 7
R 34.5 2.09 0 5L&R 71.5 4.80 1 12
87N1 15.2 A 37.5 6.40 0 -B 38.0 5.50 1 -
A&B 75.5 11.9 1 -92B2 15.6 L - - - -
R 37.8 2.92 0 22L&R - - - -
80G1 15.7 L 42.5 1.80 0 -R 44.0 2.60 1 -
L&R 86.5 4.40 1 -87B5 15.7 L 42.0 2.78 0 4
R 44.8 5.00 1 7L&R 86.8 7.78 1 11
87B6 15.7 L 35.4 5.70 0 2R 42.2 3.70 0 10
L&R 77.6 9.40 0 1288G1 15.7 A 42.0 2.18 0 6
B 38.8 4.91 1 3A&B 80.8 7.09 1 9
92B8 15.7 A 52.7 4.48 - -B 47.0 6.48 1 -
A&B 99.7 10.96 - -81KK3 16.2 L 27.0 2.01 0 9
R 42.0 1.93 0 0L&R 69.0 3.94 0 9
81WW3 16.2 L 42.0 3.92 1 16R 43.0 1.96 0 11
L&R 85.0 5.88 1 2785WW1 16.2 L 32.0 2.41 0 2
R 39.5 2.50 0 4L&R 71.5 4.90 0 6
92B4 16.2 A 50.5 6.28 0 9B 45.2 5.74 0 3
A&B 95.7 12.02 0 1282WW2 16.5 L 45.0 4.77 0 3
R 55.0 4.57 0 4L&R 100.0 9.34 0 7
89B3 16.9 L 41.5 2.42 0 22R 45.5 2.05 0 19
CORRELATES OF OVARY MORPHOMETRICS IN THE BOWHEAD WHALE 773
ovarian surface could often still be located externally bytheir hardened consistency and the presence of theirrespective former stigmata (Fig. 5). Involution of others,however, was sufficiently advanced to preclude detectionbefore sectioning. CAs were found throughout the lengthof the ovaries, varying in number from 1 to 41 as a com-bined total for an ovarian pair. In 10 of the 17 whaleswhere both ovaries were collected and identified as rightor left, the right ovary contained the greatest number ofcorpora. Ovarian corpora did not appear to favor distri-bution toward one ovarian pole or the other.
Ovary Morphometrics
Linear measures and weights were recorded for sexu-ally immature and mature ovaries, and ovary lengthsand weights were examined relative to whale total bodylengths (all sexually immature whales were �14.2 m intotal body length, while all sexually mature whales were14.2 m or greater in total body length) (Tables 2 and 3).Immature ovaries ranged from 13.6 to 32.0 cm in indi-vidual length (n 5 46) and from 28.1 to 62.0 cm as com-bined lengths for a pair (n 5 23); they were from 5.0 to10.8 cm at greatest width and 1.9 to 9.2 cm at greatestdepth through the mesovarial plane (n 5 46). Weights ofindividual immature ovaries varied from 0.17 to 1.31 kg(n 5 46), while ovary pair weights for each whale rangedfrom 0.38 to 2.45 kg (n 5 23). Weight asymmetrybetween ovaries of a pair was assessed in 18 sexuallyimmature whales. There was no significant differencebetween the weight of the right and left ovaries; theheavier ovary occurred in eight of the right ovaries and
six of the left. Ovaries were equal in weight in fourwhales.
Sexually mature ovaries varied from 26.8 to 55.0 cmin individual length (n 5 50) and from 57.3 to 100.0 cmas combined lengths for a pair (n 5 24); they were from9.8 to 19.0 cm at greatest width and 5.5 to 13.8 cm atgreatest depth in the mesovarial plane (n 5 50). Matureovaries weighed from 0.99 to 6.48 kg (n 5 52) individu-ally and from 2.92 to 12.02 kg as a pair (n 5 25). For 12of the 13 whales with a CL, the heaviest ovary in a paircontained the CL. To counter weight bias introduced byCLs or large CAs for the purposes of body length correla-tions, mature ovary weight groups were also isolated for(1) paired ovary weights for whales lacking a CL (4.47–12.02 kg; n 5 13) and (2) weight of the single ovary (in apair) lacking a CL or the largest CA (1.70–5.74 kg;n 5 13).
Ovary lengths (single ovaries and combined lengthsfor ovaries of a pair) were examined in relation to whaletotal body length (Table 6). While there was a tendencyfor single and paired ovary lengths to be shorter relativeto total body lengths in immature whales than inmature individuals, there was considerable overlapbetween the two groups such that these values weremore often concordant than distinctive.
Ovary pair weights (single ovaries and combinedweights for ovaries of a pair) were also examined in rela-tion to whale total body length (Table 6). In maturewhales, three categories of ovary weights were examinedin order to consider possible influence caused by CLs orlarge CAs: (1) single ovary weights (using the ovary of apair lacking a CL or the largest CA; n 5 24), (2) paired
TABLE 3. (continued).
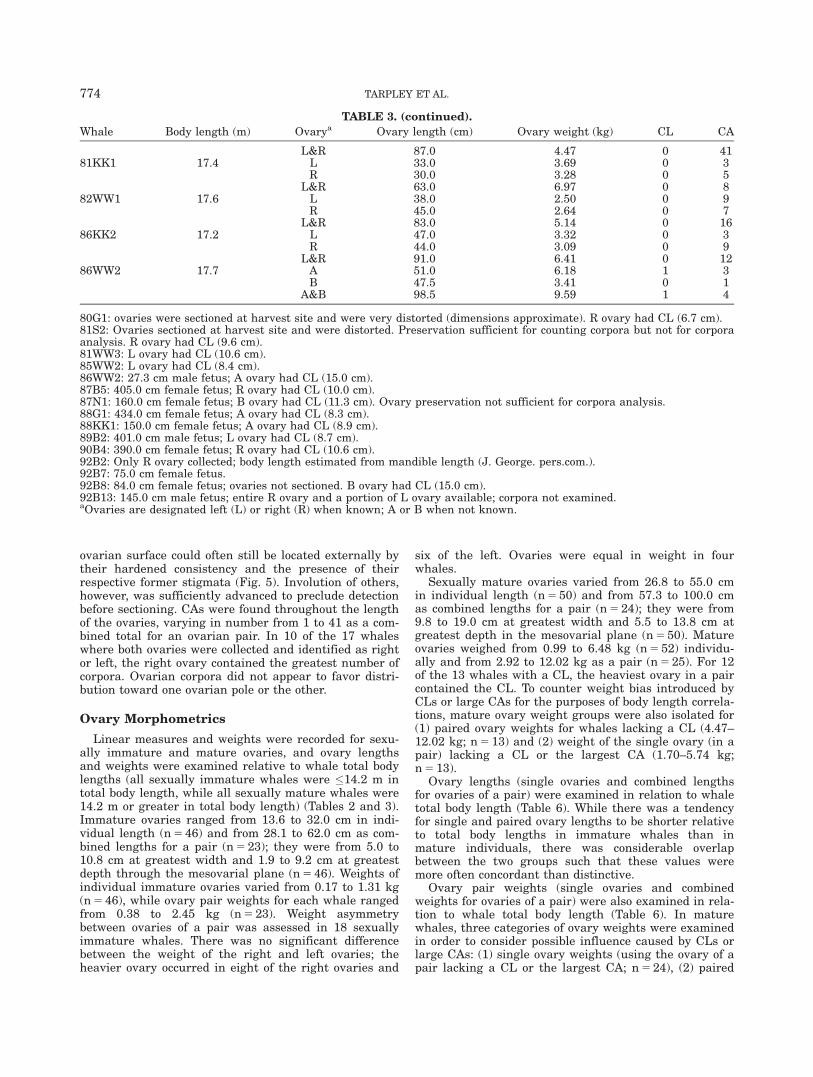
Whale Body length (m) Ovarya Ovary length (cm) Ovary weight (kg) CL CA
L&R 87.0 4.47 0 4181KK1 17.4 L 33.0 3.69 0 3
R 30.0 3.28 0 5L&R 63.0 6.97 0 8
82WW1 17.6 L 38.0 2.50 0 9R 45.0 2.64 0 7
L&R 83.0 5.14 0 1686KK2 17.2 L 47.0 3.32 0 3
R 44.0 3.09 0 9L&R 91.0 6.41 0 12
86WW2 17.7 A 51.0 6.18 1 3B 47.5 3.41 0 1
A&B 98.5 9.59 1 4
80G1: ovaries were sectioned at harvest site and were very distorted (dimensions approximate). R ovary had CL (6.7 cm).81S2: Ovaries sectioned at harvest site and were distorted. Preservation sufficient for counting corpora but not for corporaanalysis. R ovary had CL (9.6 cm).81WW3: L ovary had CL (10.6 cm).85WW2: L ovary had CL (8.4 cm).86WW2: 27.3 cm male fetus; A ovary had CL (15.0 cm).87B5: 405.0 cm female fetus; R ovary had CL (10.0 cm).87N1: 160.0 cm female fetus; B ovary had CL (11.3 cm). Ovary preservation not sufficient for corpora analysis.88G1: 434.0 cm female fetus; A ovary had CL (8.3 cm).88KK1: 150.0 cm female fetus; A ovary had CL (8.9 cm).89B2: 401.0 cm male fetus; L ovary had CL (8.7 cm).90B4: 390.0 cm female fetus; R ovary had CL (10.6 cm).92B2: Only R ovary collected; body length estimated from mandible length (J. George. pers.com.).92B7: 75.0 cm female fetus.92B8: 84.0 cm female fetus; ovaries not sectioned. B ovary had CL (15.0 cm).92B13: 145.0 cm male fetus; entire R ovary and a portion of L ovary available; corpora not examined.aOvaries are designated left (L) or right (R) when known; A or B when not known.
774 TARPLEY ET AL.

ovary weights (for all ovary pairs; n 5 25), and (3) pairedovary weights (for whales lacking a CL; n 5 12).
All ovary pairs from sexually mature whales wereheavier than those from sexually immature animals.However, for both immature and mature whales, therewas considerable variation in ovary pair weights for agiven body length. Therefore, while there was no overlapbetween immature and mature whales with regard topaired ovary weight, there was less indication that ovaryweights increased in proportion to whale body lengthwhen immature (r2 5 0.38; P 5 0.0019) or mature(r2 5 0.01; P 5 0.7059) categories were considered sepa-rately (Fig. 13). There was greater variation in ovarypair weight relative to body length among the maturewhales, and the heavier ovary pairs from mature whaleswere not always explained by the presence of a CL. Theratio of individual ovary weights to total body length
was more variable than that of ovary pairs for bothimmature and mature whales.
It should be noted that the accuracy of all morphomet-ric correlations of ovary data relative to whale bodylength in our study is subject to any errors associatedwith whale length measurements related to bodystretching during haul-out (George et al., 2004). Forexample, the measured length of a whale after it hasbeen pulled out of the water onto ice or beach couldresult in some degree of stretching during pull-out. Thedegree of stretching can be influenced (1) by the methodof haul-out (e.g., by a hand-driven rope and pulley sys-tem onto ice or whether pulled onto hard ground by trac-tor), (2) by the size and weight of the whale (with largerwhales likely stretching more relative to a given length),or (3) by the posture of the whale (i.e., on its ventrum,side or dorsum) during pull-out. We have not attempted
Fig. 1. a. Sexually mature ovaries were cut into transverse sections from pole to pole at approximately5 mm intervals on a commercial meat slicer. Corpora were identified and totaled for each ovary. b. Ovarysections were strung together in serial order for ovary archiving.
CORRELATES OF OVARY MORPHOMETRICS IN THE BOWHEAD WHALE 775

Fig. 2. Each corpus was photographed and scaled at its approxi-mate mid-section to estimate the size of its structural divisions, includ-ing the former luteal area, central scar and central lumen, if present.a. For the corpus luteum (CL) from the right ovary of whale 90B4, theouter and inner margins of the luteal tissue (L) have been traced usinga computer morphometry program, followed by the outer margin ofthe central lumen (V). From this, the respective areas of the luteal tis-
sue, inner collagenous core and central lumen could be determined.b. For the corpus albicans (CA) pictured (the third CA from the leftovary of whale 90B4), the inner and outer margins of the regressingformer luteal area (consisting largely of spiral arteries) has been tracedto quantify its area. The inner margin defines the area of the centralcollagenous core. All area measurements for CLs and CAs were con-verted to diameter values to facilitate comparisons from the literature.
776 TARPLEY ET AL.

to propose correction factors for such variables relativeto the correlations presented in this study since oursmall data set is not likely to provide sufficient resolu-tion to define any impacts related to the range of varia-bles attending whale length measurements due tostretching.
Corpora Counts
The number of corpora (CLs and/or CAs) within indi-vidual ovaries ranged from 0 to 22 (mean 6, median 4,n 5 45). Combined totals for an ovary pair varied from 2to 41 with a mean of 11.5 (median 9.5) per whale in thesample (n 5 22). The number of corpora in an ovary pair(n 5 22) had a low positive correlation with total bodylength (r2 5 0.11; P 5 0.1286) (Fig. 14). Only a singlelarge CL was found in an ovary pair from pregnantwhales (n 5 9) or in an ovary pair with a putative CL ofovulation (i.e., no fetus was detected). No accessory CLswere identified.
To explore whether the left or the right ovary is moreactive in the production of corpora, 16 whales where theovaries were identified as left or right at the time of col-lection were examined. In making this assessment, bothCLs and CAs were included in the total corpora count.The right ovary contained more corpora than the left innine ovary pairs while the left ovary contained more cor-pora in seven ovary pairs. On this basis, the McNemar’schi-square test found no significant difference betweenright and left corpora production (v2 5 0.06, df 5 1,P 5 0.807). Further, of the 200 corpora found in these 16whales, 103 (51.5%) were from the left and 97 from theright ovary. With respect to CLs specifically, for preg-nant whales where the ovaries were identified as rightor left (n 5 4), the CL was found in two left ovaries andtwo right ovaries. For CLs of ovulation where the ova-ries were identified as right or left (n 5 3), the CL wasfound in two left ovaries and one right ovary. Thus,when our sample was examined as a whole, there wasrobust balance in the production of corpora (i.e., ovula-tion) between left and right ovaries, even while there
was considerable variation in the numbers of corporabetween ovaries of a pair for individual whales, with aslight suggestion of increased right-left ovary corporabalance as more corpora accumulated (Fig. 15).
Variability in the numbers of corpora between ovariesof a pair from individual whales could be examined in 22whales since it was not necessary for the question ofbetween-ovary disparities to identify ovaries as rightand left but only to know which ovaries formed a pair.Greatest disparity was found in whale 81KK3 wherenine corpora were recognized in the left ovary, whilenone were found in the right. Least disparity was foundin whale 92B7 where there was a single corpus in eachovary. Thus the 22 between-ovary disparities in corporacounts for ovaries of a pair ranged from 0 to 9. Theirmean was 3.9 (median 3), demonstrating a tendency forthese disparities to cluster in the lower part of theirrange. Since one whale with 41 corpora had only a dif-ference of 3 corpora between left and right ovaries, itmust be considered that disparity might lessen with cor-pora accumulation, but our small data set does not per-mit an interpretation.
Finally, disparity in the production of corpora betweenleft and right ovaries was viewed through an examina-tion of the number of corpora produced in the left ovaryas a proportion of total corpora in the pair (Table 7). Forall 16 whales where left and right ovaries were identi-fied, left corpora as a percentage of total corpora in anovary pair ranged from 17 to 100% with a mean of 50%(median 44%). That is, the left proportions averaged0.50, as did the right proportions.
Since the confidence interval includes 0.5 for the first15 whales, the null hypothesis that the left and rightovaries are equally productive would not be rejected forthose whales at the 1% level. That hypothesis would berejected for 81KK3 at the 1% level because her confi-dence interval does not include 0.5. The 1% rather thanthe 5% level was used in order to control the probabilityof rejection of a true null hypothesis over the 16 tests.This overall level is approximately 0.16, so there isabout a 16% chance that the null hypothesis of equal
TABLE 4. Bowhead whale corpus luteum diametersa (sorted by increasing diameter)
OvaryCorpus
diameter (cm)Scar
diameter (cm)Lumen
diameter (cm)Capture dayand month
Fetuslength (cm)
80G1R 6.7 – - 4 May b
88G1A 8.3 2.4 - 16 Apr 43485WW2L 8.4 3.5 2.8 18 May b
89B2L 8.7 2.9 1.5 15 May 40188KK1L 8.9 3.3 1.8 24 Sep 15092B7A 9.0 3.1 - 2 Sep 7581S2 9.6 – - 21 Apr b
87B5R 10.0 4.0 2.5 15 Jun 40581WW3L 10.6 4.8 2.8 28 May b
90B4R 10.6 4.7 4.0 19 May 39087N1B 11.3 5.8 2.1 5 Oct 16086WW2A 15.0 12.1 9.8 10 May 2792B8 15.0 – - 3 Sep 84
aDiameter values are inclusive of divisional subsets (i.e., corpus diameters include both scar and lumen diameters, andscar diameters are inclusive of lumen diameters).bNo fetus recovered; possible corpus luteum of ovulation.
CORRELATES OF OVARY MORPHOMETRICS IN THE BOWHEAD WHALE 777
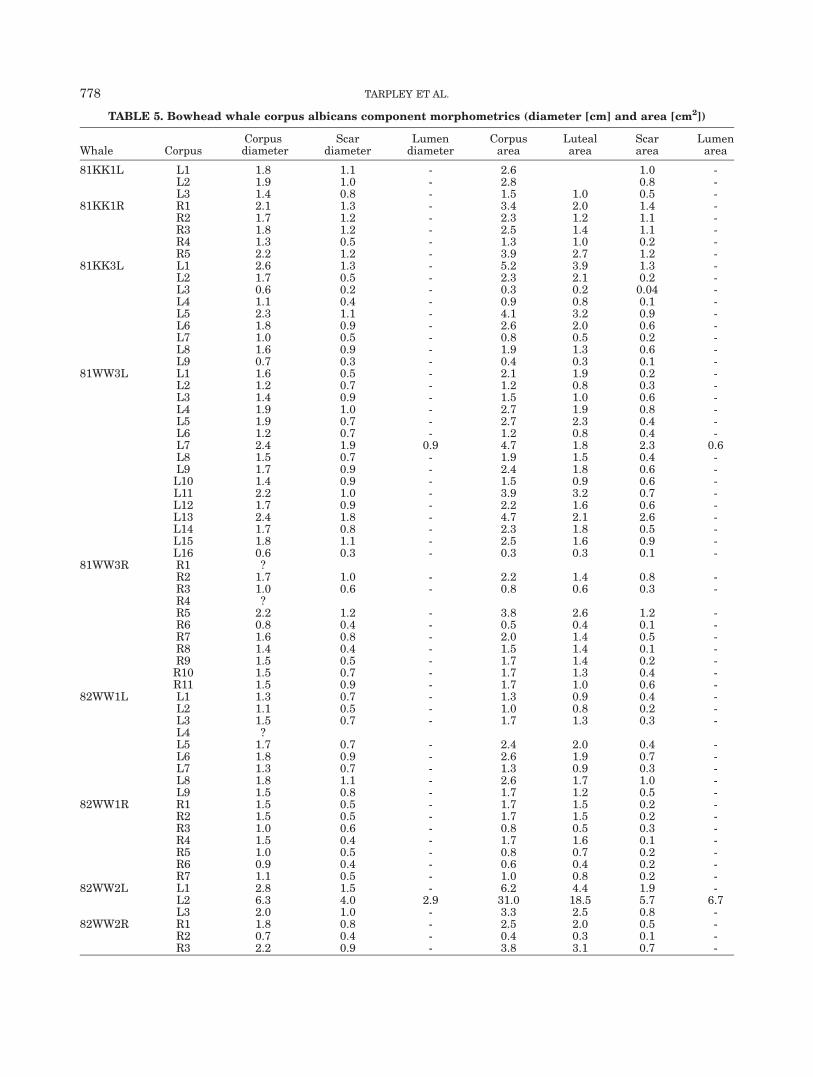
TABLE 5. Bowhead whale corpus albicans component morphometrics (diameter [cm] and area [cm2])
Whale CorpusCorpusdiameter
Scardiameter
Lumendiameter
Corpusarea
Lutealarea
Scararea
Lumenarea
81KK1L L1 1.8 1.1 - 2.6 1.0 -L2 1.9 1.0 - 2.8 0.8 -L3 1.4 0.8 - 1.5 1.0 0.5 -
81KK1R R1 2.1 1.3 - 3.4 2.0 1.4 -R2 1.7 1.2 - 2.3 1.2 1.1 -R3 1.8 1.2 - 2.5 1.4 1.1 -R4 1.3 0.5 - 1.3 1.0 0.2 -R5 2.2 1.2 - 3.9 2.7 1.2 -
81KK3L L1 2.6 1.3 - 5.2 3.9 1.3 -L2 1.7 0.5 - 2.3 2.1 0.2 -L3 0.6 0.2 - 0.3 0.2 0.04 -L4 1.1 0.4 - 0.9 0.8 0.1 -L5 2.3 1.1 - 4.1 3.2 0.9 -L6 1.8 0.9 - 2.6 2.0 0.6 -L7 1.0 0.5 - 0.8 0.5 0.2 -L8 1.6 0.9 - 1.9 1.3 0.6 -L9 0.7 0.3 - 0.4 0.3 0.1 -
81WW3L L1 1.6 0.5 - 2.1 1.9 0.2 -L2 1.2 0.7 - 1.2 0.8 0.3 -L3 1.4 0.9 - 1.5 1.0 0.6 -L4 1.9 1.0 - 2.7 1.9 0.8 -L5 1.9 0.7 - 2.7 2.3 0.4 -L6 1.2 0.7 - 1.2 0.8 0.4 -L7 2.4 1.9 0.9 4.7 1.8 2.3 0.6L8 1.5 0.7 - 1.9 1.5 0.4 -L9 1.7 0.9 - 2.4 1.8 0.6 -L10 1.4 0.9 - 1.5 0.9 0.6 -L11 2.2 1.0 - 3.9 3.2 0.7 -L12 1.7 0.9 - 2.2 1.6 0.6 -L13 2.4 1.8 - 4.7 2.1 2.6 -L14 1.7 0.8 - 2.3 1.8 0.5 -L15 1.8 1.1 - 2.5 1.6 0.9 -L16 0.6 0.3 - 0.3 0.3 0.1 -
81WW3R R1 ?R2 1.7 1.0 - 2.2 1.4 0.8 -R3 1.0 0.6 - 0.8 0.6 0.3 -R4 ?R5 2.2 1.2 - 3.8 2.6 1.2 -R6 0.8 0.4 - 0.5 0.4 0.1 -R7 1.6 0.8 - 2.0 1.4 0.5 -R8 1.4 0.4 - 1.5 1.4 0.1 -R9 1.5 0.5 - 1.7 1.4 0.2 -R10 1.5 0.7 - 1.7 1.3 0.4 -R11 1.5 0.9 - 1.7 1.0 0.6 -
82WW1L L1 1.3 0.7 - 1.3 0.9 0.4 -L2 1.1 0.5 - 1.0 0.8 0.2 -L3 1.5 0.7 - 1.7 1.3 0.3 -L4 ?L5 1.7 0.7 - 2.4 2.0 0.4 -L6 1.8 0.9 - 2.6 1.9 0.7 -L7 1.3 0.7 - 1.3 0.9 0.3 -L8 1.8 1.1 - 2.6 1.7 1.0 -L9 1.5 0.8 - 1.7 1.2 0.5 -
82WW1R R1 1.5 0.5 - 1.7 1.5 0.2 -R2 1.5 0.5 - 1.7 1.5 0.2 -R3 1.0 0.6 - 0.8 0.5 0.3 -R4 1.5 0.4 - 1.7 1.6 0.1 -R5 1.0 0.5 - 0.8 0.7 0.2 -R6 0.9 0.4 - 0.6 0.4 0.2 -R7 1.1 0.5 - 1.0 0.8 0.2 -
82WW2L L1 2.8 1.5 - 6.2 4.4 1.9 -L2 6.3 4.0 2.9 31.0 18.5 5.7 6.7L3 2.0 1.0 - 3.3 2.5 0.8 -
82WW2R R1 1.8 0.8 - 2.5 2.0 0.5 -R2 0.7 0.4 - 0.4 0.3 0.1 -R3 2.2 0.9 - 3.8 3.1 0.7 -
778 TARPLEY ET AL.

TABLE 5. (continued).
Whale CorpusCorpusdiameter
Scardiameter
Lumendiameter
Corpusarea
Lutealarea
Scararea
Lumenarea
R4 2.7 1.8 - 5.8 3.2 2.6 -85WW1L L1 1.2 0.6 - 1.1 0.9 0.3 -
L2 1.6 0.8 - 1.8 1.3 0.6 -85WW1R R1 1.6 0.9 - 2.0 1.4 0.7 -
R2 2.1 1.5 - 3.5 1.6 1.9 -R3 2.6 1.9 - 5.3 2.6 2.7 -R4 1.6 0.8 - 2.1 1.7 0.5 -
85WW2L L1 0.7 0.1 - 0.3 0.3 0.01 -L2 1.5 0.6 - 1.8 1.5 0.3 -L3 2.0 1.0 - 3.0 2.3 0.8 -L4 1.6 1.1 - 2.1 1.2 1.0 -L5 1.0 0.4 - 0.8 0.7 0.1 -L6 1.3 0.4 - 1.3 1.2 0.1 -L7 1.6 1.0 - 2.0 1.1 0.9 -
85WW2R R1 1.6 0.5 - 2.0 1.8 0.2 -R2 1.4 0.6 - 1.6 1.3 0.3 -R3 1.0 0.5 - 0.8 0.6 0.2 -R4 0.6 0.1 - 0.3 0.3 0.01 -R5 1.3 0.6 - 1.4 1.1 0.3 -
86KK2L L1 0.9 0.1 - 0.6 0.6 0.01 -L2 1.0 0.1 - 0.9 0.8 0.01 -L3 2.8 1.0 - 6.2 5.4 0.9 -
86KK2R R1 1.4 0.6 - 1.6 1.3 0.3 -R2 1.8 0.7 - 2.7 2.3 0.3 -R3 4.9 3.9 3.5 18.6 6.4 2.8 9.4R4 1.4 0.4 - 1.5 1.4 1.0 -R5 1.4 0.4 - 1.6 1.5 1.0 -R6 1.8 0.9 - 2.5 1.8 0.6 -R7 1.5 0.1 - 1.9 1.9 0.01 -R8 1.9 0.8 - 2.8 2.4 0.5 -R9 1.9 0.5 - 2.9 2.7 0.2 -
86WW2A A1 2.5 1.4 0.5 4.7 3.1 1.5 0.2A2 2.5 1.3 - 4.9 3.5 1.4 -A3 2.2 1.5 - 3.8 2.0 1.8 -
86WW2B B1 2.8 1.9 - 6.1 3.4 2.7 -87B5L L1 2.1 1.4 - 3.3 1.7 1.6 -
L2 0.9 0.4 - 0.6 0.4 0.1 -L3 0.8 0.4 - 0.5 0.4 0.1 -L4 0.9 0.4 - 0.6 0.5 0.1 -
87B5R R1 0.5 0.2 - 0.2 0.1 0.04 -R2 0.7 0.3 - 0.4 0.3 0.9 -R3 1.9 1.0 - 2.8 2.1 0.7 -R4 0.8 0.5 - 0.5 0.3 0.2 -R5 2.0 0.9 - 3.1 2.5 0.6 -R6 0.8 0.5 - 0.5 0.3 0.2 -R7 0.7 0.3 - 0.4 0.3 0.1 -
87B6L L1 2.7 1.1 - 5.5 4.6 0.9 -L2 3.4 1.5 - 9.1 7.2 1.9 -
87B6R R1 3.2 1.7 - 8.2 6.0 2.1 -R2 2.6 1.1 - 5.3 4.4 0.9 -R3 2.1 1.3 - 3.6 2.3 1.3 -R4 2.1 0.7 - 3.5 3.1 0.4 -R5 2.1 1.6 - 3.5 1.6 1.9 -R6 2.1 1.1 - 3.5 2.5 1.0 -R7 2.1 0.9 - 3.4 2.7 0.6 -R8 2.4 1.0 - 4.6 3.8 0.7 -R9 2.3 0.8 - 4.2 3.7 0.5 -R10 2.1 0.6 - 3.5 3.2 0.3 -
88G1A A1 1.9 0.6 - 2.8 2.4 0.3 -A2 2.7 1.8 - 5.6 3.1 2.5 -A3 1.8 0.7 - 2.6 2.3 0.4 -A4 2.0 1.1 - 3.1 2.2 0.9 -A5 1.8 0.8 - 2.6 2.1 0.5 -A6 1.6 0.6 - 2.1 1.9 0.3 -
88G1B B1 1.5 0.9 - 1.8 1.1 0.6 -B2 1.4 0.5 - 1.5 1.4 0.2 -
CORRELATES OF OVARY MORPHOMETRICS IN THE BOWHEAD WHALE 779

TABLE 5. (continued).
Whale CorpusCorpusdiameter
Scardiameter
Lumendiameter
Corpusarea
Lutealarea
Scararea
Lumenarea
B3 0.8 0.3 - 0.5 0.5 0.1 -88KK1L L1 1.4 0.3 - 1.6 1.5 0.1 -
L2 1.7 0.7 - 2.2 1.9 0.4 -L3 0.8 0.3 - 0.5 0.4 0.1 -L4 0.5 0.2 - 0.2 0.2 0.02 -L5 1.4 0.4 - 1.6 1.5 0.1 -L6 0.7 0.4 - 0.3 0.2 0.1 -
89B2L L1 0.5 0.2 - 0.2 0.1 0.1 -L2 0.8 0.4 - 0.5 0.3 0.1 -L3 0.9 0.4 - 0.7 0.5 0.2 -L4 0.5 0.3 - 0.2 0.1 0.1 -L5 0.6 0.3 - 0.3 0.2 0.1 -
89B2R R1 0.4 0.2 - 0.2 0.1 0.03 -89B3L L1 1.9 1.1 - 2.9 1.9 1.0 -
L2 1.8 1.3 0.8 2.6 1.3 0.8 0.5L3 0.9 0.4 - 0.6 0.5 0.1 -L4 1.2 0.6 - 1.2 0.9 0.3 -L5 1.3 0.7 - 1.3 0.9 0.4 -L6 1.3 0.7 - 1.4 1.0 0.4 -L7 1.1 0.6 - 1.0 0.7 0.3 -L8 1.1 0.6 0.1 0.9 0.7 0.3 0.01L9 1.2 0.6 0.3 1.1 0.8 0.2 0.1L10 1.4 0.6 - 1.5 1.2 0.3 -L11 1.7 0.8 - 2.3 1.7 0.* -L12 1.3 1.* - 1.3 0.6 0.7 -L13 1.7 1.2 - 2.3 1.2 *.2 -L14 1.4 0.8 - 1.6 1.1 0.5 -L15 1.3 0.6 - 1.3 1.0 0.3 -L16 0.7 0.3 - 0.3 0.3 0.1 -L17 1.5 0.8 0.2 1.8 1.2 0.5 0.04L18 1.1 0.7 0.3 1.0 0.6 0.3 0.1L19 2.2 1.5 - 3.9 2.1 1.8 -L20 1.0 0.5 0.1 0.8 0.6 0.2 0.01L21 1.1 0.6 0.2 0.9 0.6 0.3 0.02L22 1.6 1.0 0.5 2.1 1.3 0.6 0.2
89B3R R1 0.8 0.4 - 0.5 0.3 0.1 -R2 1.2 0.7 - 1.2 0.8 0.4 -R3 0.5 0.2 - 0.2 0.2 0.03 -R4 1.3 0.7 - 1.4 1.0 0.4 -R5 1.0 0.4 - 0.7 0.6 0.1 -R6 0.9 0.3 - 0.6 0.5 0.01 -R7 1.1 0.6 - 1.0 0.7 0.3 -R8 1.0 0.5 - 0.8 0.6 0.2 -R9 0.4 0.2 - 0.1 0.1 0.03 -R10 1.0 0.5 - 0.8 0.6 0.2 -R11 1.3 0.5 - 1.3 1.1 0.2 -R12 0.9 0.5 - 0.6 0.5 0.2 -R13 1.4 0.6 - 1.6 1.3 0.3 -R14 1.3 0.8 - 1.3 0.9 0.5 -R15 0.9 0.2 - 0.6 0.6 0.03 -R16 1.1 0.4 - 0.9 0.7 0.1 -R17 1.3 0.6 - 1.2 0.9 0.3 -R18 1.0 0.5 - 0.8 0.6 0.2 -R19 1.6 1.0 - 1.9 1.2 0.7 -
90B4L L1 0.5 0.2 - 0.2 0.1 0.1 -L2 0.8 0.3 0.1 0.4 0.4 0.1 0.01L3 1.5 0.9 - 1.7 1.0 0.6 -L4 1.3 0.7 - 1.3 0.9 0.4 -L5 1.9 1.0 0.3 2.7 1.9 0.8 0.1
90B4R R1 1.5 0.3 - 1.8 1.8 0.1 -R2 0.8 0.5 0.3 0.5 0.4 0.1 0.1R3 0.8 0.4 - 0.5 0.4 0.1 -R4 0.7 0.2 - 0.4 0.3 0.04 -R5 0.7 0.3 - 0.3 0.3 0.1 -
92B2R R1 0.8 0.3 - 0.5 0.5 0.04 -R2 1.6 0.6 - 2.1 1.8 0.3 -
780 TARPLEY ET AL.

TABLE 5. (continued).
Whale CorpusCorpusdiameter
Scardiameter
Lumendiameter
Corpusarea
Lutealarea
Scararea
Lumenarea
R3 1.9 0.8 - 2.8 2.3 0.5 -R4 1.8 1.0 - 2.5 1.8 0.7 -R5 1.8 0.7 - 2.5 2.1 0.4 -R6 1.8 0.5 - 2.5 2.3 0.2 -R7 1.9 0.7 - 2.8 2.4 0.4 -R8 1.8 0.9 - 2.6 1.9 0.7 -R9 0.8 0.2 - 0.5 0.5 0.04 -R10 1.4 0.3 - 1.5 1.4 0.05 -R11 1.4 0.5 - 1.5 1.3 0.2 -R12 1.7 0.7 - 2.2 1.8 0.4 -R13 2.1 1.1 - 3.5 2.6 0.9 -R14 1.7 0.6 - 2.3 2.0 0.3 -R15 0.6 0.3 - 0.3 0.2 0.05 -R16 1.0 0.3 - 0.8 0.7 0.05 -R17 0.9 0.2 - 0.6 0.6 0.03 -R18 1.9 0.6 - 2.8 2.5 0.3 -R19 1.3 0.4 - 1.4 1.3 0.1 -R20 1.8 0.6 - 2.6 2.3 0.3 -R21 1.5 0.7 - 1.7 1.3 0.4 -R22 1.8 0.6 - 2.4 2.1 0.3 -
92B3A A1 1.4 0.8 - 1.5 1.0 0.5 -A2 1.6 0.4 - 2.1 2.0 0.1 -A3 2.1 1.0 - 3.5 2.7 0.8 -A4 0.3 0.1 - 0.1 0.1 0.005 -
92B3B B1 2.2 0.6 - 4.0 3.8 0.2 -B2 2.0 0.5 - 3.2 3.0 0.2 -B3 2.2 0.3 - 4.0 3.9 0.1 -B4 2.4 0.9 - 4.4 3.8 0.6 -B5 1.6 0.7 - 1.9 1.5 0.4 -B6 3.0 1.3 - 7.0 5.7 1.3 -B7 1.1 0.3 - 0.9 0.8 0.1 -B8 2.0 0.3 - 3.3 3.2 0.1 -B9 4.0 0.8 3.0 12.5 4.8 0.5 7.2B10 1.9 0.4 - 2.8 2.7 0.1 -B11 2.8 0.6 - 6.2 5.9 0.3 -
92B4A A1 2.2 1.2 - 3.4 2.2 1.2 -A2 3.6 2.6 0.5 10.4 5.0 5.2 0.2A3 1.9 0.3 - 2.9 2.8 0.1 -A4 2.1 0.4 - 3.4 3.2 0.2 -A5 2.3 0.2 - 4.2 4.2 0.04 -A6 1.6 0.7 - 1.9 1.5 0.4 -A7 1.7 0.2 - 2.3 2.3 0.03 -A8 2.2 1.1 - 3.7 2.8 0.9 -A9 1.8 0.6 - 2.5 2.3 0.2 -
92B4B B1 1.6 0.7 - 2.0 1.6 0.4B2 1.9 0.8 - 2.9 2.4 0.5 -B3 2.0 0.9 - 3.2 2.6 0.6 -
92B7B B1 0.9 0.5 - 0.7 0.5 0.2 -92B9A A1 2.0 0.8 - 3.2 2.7 0.5 -
A2 1.9 0.6 - 2.8 2.6 0.2 -A3 2.2 0.5 - 3.7 3.5 0.2 -A4 2.7 0.9 - 5.9 5.3 0.6 -A5 2.8 1.3 - 6.0 4.7 1.3 -A6 2.7 0.6 - 5.7 5.3 0.4 -
92B9B B1 2.0 0.6 - 3.2 2.9 0.3B2 1.9 0.4 - 2.9 2.8 0.1 -B3 2.2 0.7 - 3.9 3.5 0.4 -
92B10L L1 2.5 1.1 0.2 4.9 4.0 0.9 0.0292B10R R1 4.5 2.2 1.0 16.2 11.6 3.8 0.8
R2 2.4 0.9 - 4.6 4.0 0.6 -R3 1.3 0.3 - 1.3 1.2 0.06 -R4 2.3 0.7 - 4.3 3.9 0.4 -
CORRELATES OF OVARY MORPHOMETRICS IN THE BOWHEAD WHALE 781

left and right ovary productivity was incorrectly rejectedfor 81KK3.
Thus, while there can be considerable left/right imbal-ance in the number of corpora found in an individualwhale at the time it is sampled, whales were collectivelyevenly divided regarding which ovary was favored.Although the sample is small, the suggestion is that nei-ther the left nor right ovary appears to dominate in theproduction of corpora in the bowhead whale populationoverall.
Corpora Morphometrics
Size measures for 13 CLs included total area and cal-culated diameter (n 5 11), field diameter measurement(n 5 2), central scar area and calculated diameter(n 5 10), and any lumen area and calculated diameter(n 5 8) (Table 4). Size measures for 257 CAs includedtotal area, calculated diameter, and size fractions of thecentral scar and any central lumen, representing 22whales (eight of the 265 CAs counted from 23 whaleswere not measured) (Tables 5 and 8). For one of the
Fig. 3. The left ovary from whale 84B2 is typical of the most immature ovaries, featuring a smooth sur-face with minimal external irregularities.
Fig. 4. The left ovary from whale 84B4 illustrates a more fully developed immature ovary, more typicalof whales approaching puberty.
782 TARPLEY ET AL.

whales (92B2), CAs could only be examined in a singleovary. The 13 CLs ranged in diameter from 6.7 to15.0 cm. For confirmed pregnant whales (n 5 9), CLsranged from 8.3 to 15.0 cm (mean 5 10.8 cm) (Table 9).
CL diameters for early gestational (spring-caught)fetuses, midgestational (fall-caught) fetuses, and lategestational (spring-caught) fetuses were 15.0 cm (n 5 1),8.9 to 15.0 cm (mean 5 11.1, n 5 4) and 8.3 to 10.6 cm
Fig. 5. This ovary from a pregnant whale (92B8) presents several features found only in sexually maturewhales, including Graafian follicles (F), a corpus luteum (CL), and endurated surface areas depictingregressing corpora albicantia (CA), each marked by its site of ovulation (stigma).
Fig. 6. The hilar border of the bowhead whale ovary features an elongated groove formed as each ofthe surfaces and poles curve inward to meet the flattened mesovarium (arrows), which extends along theentire groove. The mesovarium has been transected in this specimen where it meets the ovary to isolatethe ovary for weighing. Between its mesothelial layers, the mesovarium contains fatty and fibrous tissuessupporting vessels and nerves servicing the organ through the ovarian medulla.
CORRELATES OF OVARY MORPHOMETRICS IN THE BOWHEAD WHALE 783

(mean 5 9.4, n 5 4), respectively (Fig. 16). CLs where nofetus was detected (n 5 4) were from spring-caughtwhales and ranged in diameter from 6.7 to 10.6 cm(mean 5 8.8 cm) (Table 9).
Of the 12 CLs that were sectioned, 8 contained centrallumens and were designated as vesicular. Three of theCLs lacked lumens, and for one CL, the existence of alumen was undetermined. Six of the eight were associ-ated with confirmed pregnancies. For the two vesicularCLs with no confirmed fetus, pregnancy could not beruled out from field reports since a fetus could havegone undetected or aborted. Of the four nonvesiculatedCLs, two were linked to known pregnancies. All five CLs10.0 cm or more in diameter (four with confirmedfetuses) were vesicular. Although the largest absoluteand relative lumen size (9.8 cm) was found in the largestCL (15 cm diameter), there was no clear indicationamong the eight vesicular CLs of a positive correlationbetween corpus diameter and lumen diameter.
The diameters of 257 CAs from 22 whales rangedfrom 0.3 to 6.3 cm (mode 5 1.8 cm) (Fig. 17). Most (78%)measured 2.0 cm or less in diameter. CA diametersranged from 0.3 to 4.9 cm in 10 resting whales; from 0.4to 2.8 cm in seven pregnant whales; from 0.7 to 6.3 cmin two lactating whales; from 0.6 to 2.4 cm in tworecently ovulated whales; and 0.6 to 2.1 cm in one whaleof uncertain estrous cycle status (not pregnant or lactat-ing, but since only a single ovary was collected, no dis-tinction between resting or recently ovulated could bemade). While there were greater numbers of small CAs,there was a gradual decline in the frequency of thosecorpora with diameters less than the modal diameter of1.8 cm.
Of the 257 CAs measured in 22 whales, 19 (7.0%, rep-resenting 11 ovaries and 9 whales) had lumens. In eightof the 11 ovaries having one or more vesicular CAs, alumen was featured in the largest CA. Fourteen of the19 lumens were found in CAs �2.5 cm. One whale(89B3), which had the greatest number of CAs (41) in anovary pair, contributed eight of the 19 vesicular CAs, allin the left ovary with 22 total CAs. None of the 19 CAsin the right ovary of whale 89B3 had lumens, eventhough the CAs were similar in size distribution to thosein the left.
DISCUSSION
Ovary Morphology and Morphometrics
Gross morphology and coloration of bowhead whaleovaries in this study agree generally with those reportedby Kenney et al. (1981) and with accounts in severalspecies of balaenopterid whales, where immature ova-ries are relatively flattened and smooth-surfaced withminor furrows, but where protruding follicles and cor-pora give a “knobby” irregularity to the surface of themore thickened and robust ovaries in mature whales(Mackintosh and Wheeler, 1929; Ommanney, 1932; Mat-thews, 1937; Dempsey and Wislocki, 1941; Chittlebor-ough, 1954; Laws, 1957, 1961; Gambell, 1968; Lockyer,1987). While there was a definite constriction at thejunction of bowhead CLs and the surface of the ovary,the connection was fairly broad and not pendulous assometimes noted in fin whale ovaries (Laws, 1961).Even as a corpus is regressing, and the basal constric-tion becomes less distinct, bowhead whale corporaremain easily distinguishable, along with their stigmata,
Fig. 7. The fully functional corpus luteum from pregnant whale 92B7 demonstrates the complex infold-ing of luteal tissue (L) colored by the lutein pigment of active luteal cells. The former site of ovulation(stigma, s) is apparent as a nodule on the outer surface of the corpus luteum. The remainder of the ovaryfeatures cortical tissues with a few fluid-filled follicles around the outer margins, enveloping the innermedulla (M) and its vessels.
784 TARPLEY ET AL.

on the surface of the ovary for an undetermined periodbeyond their decline as an endocrine organ. The pro-nounced expression of mature follicles and recent cor-pora in baleen whales contrasts with that of the greatsperm whale, Physeter macrocephalus, where folliclesand corpora rarely project beyond the ovarian surface innonpregnant animals and only the recently-formed,endocrine-active corpora are readily apparent externally(Best, 1967).
While we found that absolute individual and pairedlengths of ovaries tended to be greater in mature indi-viduals, a mid-range overlap between immature andmature whales prevented these measures from accu-rately assessing maturity. The ratio of ovary lengths rel-ative to body length had even less resolution in makingthis distinction. Absolute ovary weights (inclusive of ova-ries with CLs) were more effective than lengths in dis-tinguishing sexually immature and mature bowheadwhales; paired mature weights were always greaterthan those from immature whales, indicating that, whilethere was some increase in ovary length in maturewhales, it was the greater bulk attained in mature ova-ries that signaled the differentiation between immature
and mature whales. However, our small sample size mayoverstate the distinction between immature and maturewhales. In their investigations of blue and fin whales,Mackintosh and Wheeler (1929) demonstrated consider-able distinction between immature and mature ovarygroups on the basis of weight, although there was someoverlap at the margins. Gambell (1968), however, deter-mined that it was not possible to precisely separate sex-ually immature and mature whales strictly on the basisof ovary weights in sei whales. Chittleborough (1954)found an overall increase in ovary pair weights withincreasing body length in humpback whales (though lessstriking for mature whales); but because of wide varia-tion in ovary weights in late immaturity and early matu-rity, a clear distinction between immature and maturewhales was not possible on the basis of ovary weights inhis sample—his mature ovary weight values wererestricted to paired weights from resting whales toreduce bias introduced by CLs. In sperm whales, Best(1967), comparing immature ovary pair weights withmature pairs from resting, nonpregnant, and nonlactat-ing whales, reported considerable overlap betweenimmature and mature ovary weights, eliminating this
Fig. 8. Three corpora albicantia from the left ovary of whale 82WW2 illustrate the former luteal tissue (L)surrounding a white collagenous core. The largest also features a central cavity in this case. These cor-pora also demonstrate successive stages of regression with the largest (and presumably youngest) to theright protruding considerably beyond the ovarian surface, while the corpus albicans on the left hasregressed farthest into the ovarian cortex, no longer extending beyond the surface contour.
CORRELATES OF OVARY MORPHOMETRICS IN THE BOWHEAD WHALE 785

Fig. 9. While generally globular, there was considerable variability in corpora lutea shape. Eleven of 13 corpora lutea were sectioned and areshown here in schematic profiles at their approximate mid-sections. Three of these were possible corpora lutea of ovulation since no fetus wasrecovered (pictured in the upper row), while the remaining eight came from pregnant whales. Luteal tissue (dark gray) surrounds the central scar(light gray), which in turn contains, where present, the central lumen (white).
786 TARPLEY ET AL.

Fig. 10. Regressing corpora albicantia presented an outer envelope of former luteal tissue (L), now com-prised primarily of a dense network of spiral arteries. The central collagenous core (C) projected fibrous tra-beculae into the arterial network. Corpora shapes varied, comprised of essentially spherical compartments(a) to an irregular configuration of collagenous projections, enveloped by the former luteal region (b).
CORRELATES OF OVARY MORPHOMETRICS IN THE BOWHEAD WHALE 787

Fig. 11. The ovary pair from bowhead whale 82WW2 demonstratesseveral corpora albicantia in relation to the ovarian surface. In theupper ovary, two corpora (A, B) are visible in different stages ofregression. In the lower ovary, three corpora (A–C) are evident, but areessentially equal in size. Large arrowheads depict sites of small antral
follicles. The small arrowhead in the upper ovary and the small arrowsin the lower ovary indicate the former site of ovulation (stigma) in theirrespective corpora. The mesovarium is indicated in both upper (M)and lower (m) photographs.
788 TARPLEY ET AL.

criterion as a means of determining maturity in spermwhales.
We found no significant weight asymmetry betweenright and left immature bowhead ovaries, in contrast toTetsuka et al. (2004) who reported that the right ovary
was three times as likely to be the heavier ovary in apair for sexually immature Antarctic minke whales.
Although sexually immature bowhead ovary pairsweighed less than those of mature ovaries, ovaryweights varied �7.5-fold and �6.5-fold for individualand paired weights, respectively, throughout the prepu-bertal growth period. While there was an increase inovary weights with body length in immature whales, thecorrelation (r2 5 0.38) was driven primarily by gains inweight during imminent pubescence. Chittleborough(1954) similarly found little change in ovary weightsduring the earlier prepubertal period of humpbackwhales, but found that weights began increasing aspuberty approached, albeit with greater variation inovary weights relative to body length. Tetsuka et al.(2004) also noted a lack of consistency in ovary weightswithin immature body length classes, with 10-fold differ-ences in ovary weights among recently weaned Antarcticminke whale calves (<6 m in length) and sixfold differ-ences in ovary weights among prepubertal Antarcticminke whales (�8 m in length). Still, they reported thatovary weights approximately doubled overall during thecourse of prepubertal development (6–7 years), which wedid not find in immature bowheads.
In mature bowhead whales, there was a fourfold dis-parity of combined ovary weights between whales, againwith increasing weights failing to track with increasingbody length. Shorter mature whales were as likely aslonger ones to have lighter or heavier ovary pairs. Mack-intosh and Wheeler (1929) described an increase inovary weights with increasing body length in bluewhales, although they believed ovary weights declinedin older whales. We also examined mature ovary pairweights relative to body length by restricting our sampleto ovary pairs that lacked a CL. For some studies in thebalaenopterid whales, where larger sample sets wereavailable, the study group could be limited to restingfemales, thus allowing the use of the combined weight ofthe pair without corporeal bias (Matthews, 1937; Chittle-borough, 1954; Gambell, 1968).
Matthews (1937, 1938) reported a decline in ovaryweight with increasing whale length in humpback andsei whales. Chittleborough (1954), using ovary pairweights from resting females along with those in latelactation, reported a tendency for the weight of sexuallymature ovary pairs to increase (though with wide varia-tion) with body length in humpback whales. Best (1967)also restricted his sample to mature ovaries lacking aCL and found an increase in ovary pair weights withbody length in mature sperm whales. Gambell (1968)found that ovary weights in sei whales continued toincrease following sexual maturity (at least until anaccumulation of 14–16 corpora).
Looking only at whales lacking a CL restricted ourstudy to a very small sample, and no ovary weightchange correlating with increasing whale length couldbe demonstrated. To enlarge our sample size while stillreducing bias that could be introduced by a large corpus,we examined mature ovary weight only in the ovary of apair that lacked the largest corpus (CL or largest CA),although in some whales it was the smaller ovary in apair that contained the largest CA. Rationale for thisapproach is supported in Gambell’s sei whale study(1968) in which he compared single ovary weights (lack-ing the large corpus) between resting and pregnant
Fig. 12. Variability in corpora albicantia shapes is demonstrated inthis schematic profile series from the approximate mid-sections of thecorpora albicantia from the left and right ovaries of whale 82WW1.
CORRELATES OF OVARY MORPHOMETRICS IN THE BOWHEAD WHALE 789

individuals and found no significant difference betweenthem, suggesting that single ovaries could be used toconsider change in ovary weight relative to body length.Still, even with this third approach, we found noincrease in ovary weight with increasing body length inmature bowhead whales.
Corpora Morphology and Morphometrics
The methodology used in this study followed that ofearlier investigators in order to permit comparisons of
TABLE 6. Ratio of ovary lengths and weights in relation to whale total body length
Status Single ovaries to body length Ovary pairs to body length
Immature Length (m/m) 0.012-0.029 (n 5 46) 0.025-0.055 (n 5 23)Mature 0.017-0.038 (n 5 50) 0.036-0.066 (n 5 24)Immature Weight (kg/m) 0.019-0.107 (n 5 46) 0.043-0.199 (n 5 23)Mature 0.067-0.421(n 5 24)a 0.264-0.783 (n 5 25)Mature 0.327-0.742 (n 5 12)b
aUsing the ovary of a pair lacking a CL or the largest CA.bFor ovary pairs lacking a CL.
Fig. 13. In our dataset, all paired ovary weights from sexuallymature whales (open circles) exceeded those from immature whales(closed circles), and all sexually immature whales were �14.2 m intotal body length, while all sexually mature whales were 14.2 m orgreater in total body length. Therefore, combined weights for an ovarypair served to distinguish sexually immature and mature whales. How-ever, there was considerable variation in ovary pair weights relative tospecific body lengths within immature and mature categories consid-ered separately. While there was some tendency for ovary pairweights to increase with body length in immature whales, this wasless true for mature animals.
Fig. 14. Total corpora for an ovary pair ranged from 2 to 41. Therewas only a slight positive correlation between the number of corporain an ovary pair and whale total body length.
Fig. 15. Total corpora (inclusive of both CLs and CAs) were examinedin the left and right ovaries of 16 whales to explore the question of ovu-latory dominance in one ovary of a pair versus the other. (Note that thesquare symbol represents two whales, both having four corpora in theright ovary and two corpora in the left ovary.) While there was consider-able variation in the number of corpora between ovaries of an individualwhale, there was a preliminary suggestion that whales with larger num-bers of corpora may show less disparity between ovaries; that is, ascorpora accumulate, there are more opportunities for left and right ova-ries to equally share the ovulatory load. Our small sample does not per-mit a conclusion in this regard, but it does imply value in furtherexploring this question with a larger sample of corpora-rich whales.
790 TARPLEY ET AL.

our dataset with their results in the context of relativecorpora size and regression that could address theapplied questions of qualitative morphological compari-sons and corpora persistence through time (e.g., Mackin-
tosh and Wheeler, 1929; Laws, 1961, Gambell, 1968). Werecognize that making a literally accurate assessment ofcorpora size and their subcompartments would requirestereological techniques that account for attending vari-ables that are artifacts of processing and sampling (Gun-dersen et al., 1988).
There was considerable diversity in bowhead whaleCL and CA morphology with regard to size, shape, cen-tral scar patterns, the amount of luteal (or remnantluteal) tissue around the scar, and the presence orabsence of a lumen. This appears to agree with accountsin other mysticetes, such as the drawings of Laws (1961)for fin whales.
Our sample of 13 CLs represented 13 whales; that is,no more than a single large CL, with no recognizedaccessory CLs, was found in any ovary or ovary pairfrom a single whale. Lockyer (1987) reported the absenceof accessory CLs in her collection of 39 CLs from 67minke whale ovary pairs. Chittleborough (1954) found asingle pregnant humpback whale with two functionalCLs, each of which was smaller than normal. In finwhales, Laws (1961) classified 3.7% of 760 CLs as acces-sory CLs (ranging from 0.4 to 8.5 cm in diameter). Sincesome of his proposed CLs were very small, and, sincevariable amounts of lutein pigment appear to be grosslyevident in some of our smaller bowhead corpora, the pos-sibility of a population of smaller CLs cannot be ruledout in bowhead whales and should be further investi-gated. But within our sample, one large CL was by farthe dominant organ supporting the pregnancy. In othercetaceans, such as the beluga whale among the odonto-cetes, accessory CLs are thought to occur (Brodie, 1971,1972; Mossman and Duke, 1973).
The largest corpus in each of nine pregnant bowheadwhales ranged from 8.3 to 15.0 cm in diameter(mean 5 10.8 cm), inclusive of early, mid, and late preg-nancies; all featured the grossly-evident, golden paren-chyma typical of active CLs reported in other whales(Dempsey and Wislocki, 1941). The bowhead CL 10.8 cmmean was smaller than means reported in blue whales(13.7 cm, Mackintosh and Wheeler, 1929), in humpback
TABLE 7. Corpus production disparity between left and right ovaries
Whale
Corpora CountsLeft ovaryproportion
Exact binomial 99%confidence intervalLeft Right Total
87B6 2 10 12 0.167 (0.009–0.573)92B10 1 4 5 0.200 (0.001–0.815)86KK2 3 9 12 0.250 (0.030–0.655)81S2 2 4 6 0.333 (0.019–0.856)85WW1 2 4 6 0.333 (0.019–0.856)87B5 4 8 12 0.333 (0.062–0.728)81KK1 3 5 8 0.375 (0.047–0.830)82WW2 3 4 7 0.429 (0.055–0.882)90B4 5 6 11 0.455 (0.114–0.831)89B3 22 19 41 0.537 (0.330–0.735)82WW1 9 7 16 0.563 (0.236–0.853)81WW3 17 11 28 0.607 (0.351–0.827)85WW2 8 5 13 0.615 (0.245–0.906)89B2 6 1 7 0.857 (0.315–0.999)88KK1 7 0 7 1.000 (0.469–1.000)81KK3 9 0 9 1.000 (0.555–1.000)Total 103 97 200 0.515
Corpora counts for the 16 whales with left and right ovaries identified. The proportion from the left ovary, with an exactbinomial 99% confidence interval, is also given (Beyer, 1968). The table is ordered by left ovary proportion.
Fig. 16. Corpora luteal (CL) diameters from the nine pregnantwhales were compared relative to associated fetal body lengths. Apreliminary suggestion from this small sample is that CLs (1) attainedmaximum diameter during the spring when supporting early fetuses(square), (2) were decreasing in diameter when associated with mid-sized fall-collected fetuses (open circles), and (3) reached a reduceddiameter when found with term fetuses in the spring (diamonds).
TABLE 8. Bowhead whale CA diameter frequenciesfor all ovaries (22 whales)
CA diameters (cm) Number of CAs Frequency (%)
0.3-6.3 257a 100�3.0 250 94�2.0 200 73
aThere were actually 265 corpora albicantia counted among23 whales examined, but eight corpora were not measured(two from the right ovary of 81WW3, one from the left ovaryof 82WW1, and five from the ovaries of 81S2).
CORRELATES OF OVARY MORPHOMETRICS IN THE BOWHEAD WHALE 791
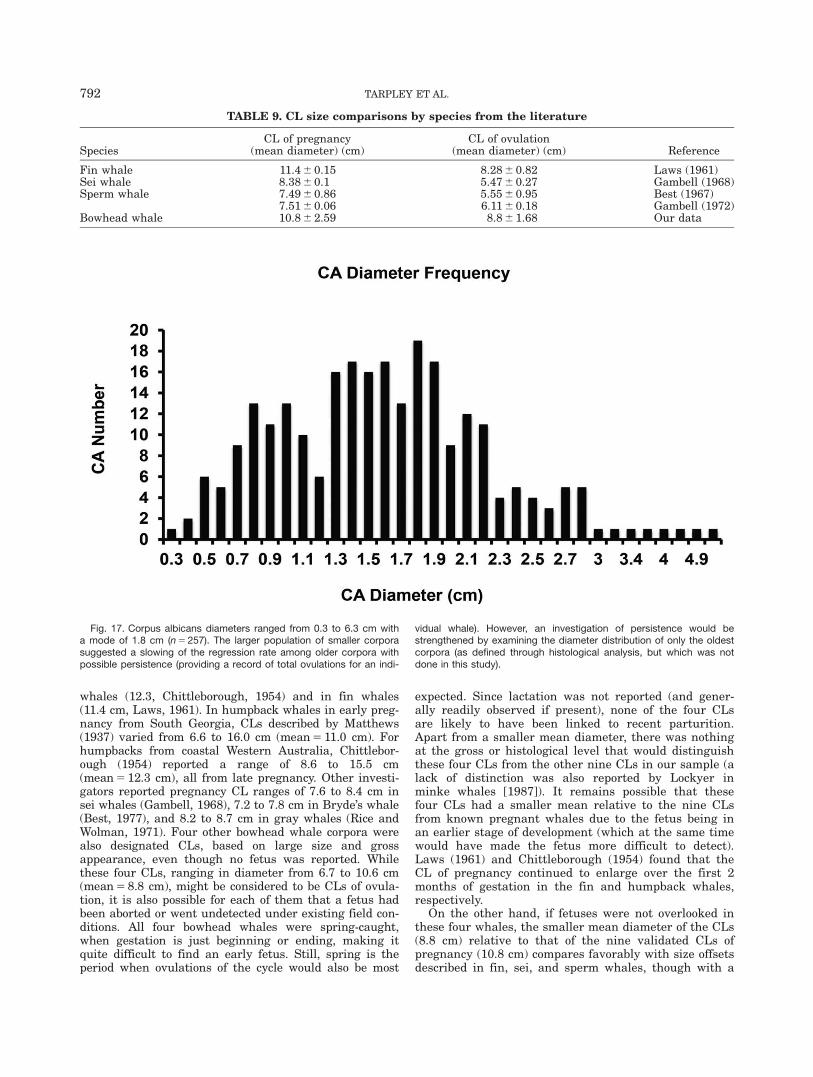
whales (12.3, Chittleborough, 1954) and in fin whales(11.4 cm, Laws, 1961). In humpback whales in early preg-nancy from South Georgia, CLs described by Matthews(1937) varied from 6.6 to 16.0 cm (mean 5 11.0 cm). Forhumpbacks from coastal Western Australia, Chittlebor-ough (1954) reported a range of 8.6 to 15.5 cm(mean 5 12.3 cm), all from late pregnancy. Other investi-gators reported pregnancy CL ranges of 7.6 to 8.4 cm insei whales (Gambell, 1968), 7.2 to 7.8 cm in Bryde’s whale(Best, 1977), and 8.2 to 8.7 cm in gray whales (Rice andWolman, 1971). Four other bowhead whale corpora werealso designated CLs, based on large size and grossappearance, even though no fetus was reported. Whilethese four CLs, ranging in diameter from 6.7 to 10.6 cm(mean 5 8.8 cm), might be considered to be CLs of ovula-tion, it is also possible for each of them that a fetus hadbeen aborted or went undetected under existing field con-ditions. All four bowhead whales were spring-caught,when gestation is just beginning or ending, making itquite difficult to find an early fetus. Still, spring is theperiod when ovulations of the cycle would also be most
expected. Since lactation was not reported (and gener-ally readily observed if present), none of the four CLsare likely to have been linked to recent parturition.Apart from a smaller mean diameter, there was nothingat the gross or histological level that would distinguishthese four CLs from the other nine CLs in our sample (alack of distinction was also reported by Lockyer inminke whales [1987]). It remains possible that thesefour CLs had a smaller mean relative to the nine CLsfrom known pregnant whales due to the fetus being inan earlier stage of development (which at the same timewould have made the fetus more difficult to detect).Laws (1961) and Chittleborough (1954) found that theCL of pregnancy continued to enlarge over the first 2months of gestation in the fin and humpback whales,respectively.
On the other hand, if fetuses were not overlooked inthese four whales, the smaller mean diameter of the CLs(8.8 cm) relative to that of the nine validated CLs ofpregnancy (10.8 cm) compares favorably with size offsetsdescribed in fin, sei, and sperm whales, though with a
TABLE 9. CL size comparisons by species from the literature
SpeciesCL of pregnancy
(mean diameter) (cm)CL of ovulation
(mean diameter) (cm) Reference
Fin whale 11.4 6 0.15 8.28 6 0.82 Laws (1961)Sei whale 8.38 6 0.1 5.47 6 0.27 Gambell (1968)Sperm whale 7.49 6 0.86 5.55 6 0.95 Best (1967)
7.51 6 0.06 6.11 6 0.18 Gambell (1972)Bowhead whale 10.8 6 2.59 8.8 6 1.68 Our data
Fig. 17. Corpus albicans diameters ranged from 0.3 to 6.3 cm witha mode of 1.8 cm (n 5 257). The larger population of smaller corporasuggested a slowing of the regression rate among older corpora withpossible persistence (providing a record of total ovulations for an indi-
vidual whale). However, an investigation of persistence would bestrengthened by examining the diameter distribution of only the oldestcorpora (as defined through histological analysis, but which was notdone in this study).
792 TARPLEY ET AL.

wider standard deviation due to our small sample size(Table 9).
Conversely, Lockyer (1987) reported a CL of ovulationthat was larger than the mean of pregnancy CLs(though based only on a single CL of ovulation) in minkewhales. There is considerable variance around the meanfor CLs in our small bowhead whale data set, and thelarger mean diameter of CLs from the nine confirmedpregnant whales is influenced by the two largest CLs(15.0 cm). Taken individually, the sizes of the four CLswhere no fetus was reported are distributed ratherevenly among the total sample of 13 CLs. Therefore,while suggestive, our data do not conclusively demon-strate that CL diameters distinguish between CLs ofpregnancy and ovulation in the bowhead whale. Overlapin the range of CL diameters for ovulation and preg-nancy has also been reported in fin whales (Laws, 1961)and sei whales (Gambell, 1968).
The bowhead gestational CL mean diameters of15.0 cm, 11.1 cm, and 9.4 cm in early, mid, and latepregnancies, respectively, suggest attainment of maxi-mal CL size in early gestation that may decrease duringthe remainder of the pregnancy. Mackintosh andWheeler (1929) thought that CLs decreased in size withadvancing pregnancy in blue and fin whales. Laws(1961), however, also studying fin whales with the bene-fit of more data, contested this conclusion. He found, asdid Gambell (1968) in the sei whale, Chittleborough(1954) in the humpback whale and Best (1967) in thesperm whale, that a maximum size was attained earlyand maintained throughout pregnancy. Laws suggestedthat the CL expanded until the fetus reached 10 to20 cm in length (an age of 1–2 months), then stabilizedthrough the remainder of pregnancy (although he gavesome evidence for a slight increase in CL size in latepregnancy). In the humpback whale, the CL grew duringthe first 2 months of gestation, then enlarged moreslowly to achieve maximum size in late pregnancy(Chittleborough, 1954). Lockyer (1987) reported anincrease in mean CL diameter as pregnancy progressedin the minke whale. However, her sample size was onlyslightly larger than our own, and there was considerablevariation around the mean. While the temporal meansfrom our bowhead sample suggested a decrease in CLsize as pregnancy progressed, our dataset is consideredtoo small to firmly establish trends in CL size duringpregnancy.
There has been longstanding interest among whalebiologists in finding a way to distinguish between CLsand CAs of ovulation and those of pregnancy, since(assuming corpora persist as a permanent record of ovu-lation in large whales as widely thought) the ability toidentify those ovulations linked to pregnancy would pro-vide a record of a whale’s lifetime productivity (Peters,1939; van Lennep, 1950; Robins, 1954). Peters (1939)proposed that CL color and texture could be used tomake this differentiation in fin and blue whales, whilevan Lennep (1950) thought the arrangement of CL tra-beculae and CL size (both more robust in pregnancy CLsin his view) might allow a distinction in these two cor-pora. No attempt was made in our study of bowhead cor-pora to histologically characterize the stage of corpusinvolution relative to size. Apart from CL diameter, sincevarious studies have shown overlap in size between pre-sumed CLs from ovulation and pregnancy, Robins (1954)
proposed from examinations of humpback whale CLsthat the presence of a vesiculated corpus (i.e., one with alumen) might differentiate corpora of ovulation andpregnancy, with the larger CL of pregnancy more likelyto be vesicular.
Laws (1961) and Gambell (1968) reported a tendencyfor the larger CLs of pregnancy to contain lumens in finand sei whales, respectively. We were able to examinelumen incidence in 12 of 13 bowhead CLs and found asimilar result since a greater fraction (0.75) of those CLsassociated with known pregnancies (n 5 8;mean 5 10.2 6 2.2 cm) had lumens when compared withthe fraction (0.50) for those CLs where no fetus wasreported (n 5 4; mean 5 8.8 6 1.7 cm). But the absence oflumens in some gestational CLs and the presence oflumens in some of those lacking a fetus defeats the useof this feature in bowhead whale ovaries to make thedistinction. Likewise, Lockyer (1987), in her study ofminke whales, found no reliable link between vesicula-tion and CLs of pregnancy, reporting lumens in only25% of early gestational CLs, 33.3% of midgestationalCLs and none in those of late gestation. Marsh (1985)described a single minke whale that was believed tohave a CL of ovulation (since it had been isolated in alagoon for 3 months before necropsy), yet possessed acentral, fibrin-filled lumen.
Gambell (1968) reported a decrease in the number ofvesicular corpora as pregnancy progressed, suggestingthat large corpora tend to have lumens that close overtime to leave a solid hyaline scar, preventing inferenceof an earlier lumen when examining smaller corpora. Incontrast, the presence of several small CAs with lumensin our sample (14 of 19 CAs with lumens were �2.5 cm)argues that lumen persistence is possible in the bowheadwhale, at least in some individuals, regardless of whatpromotes lumens to form. However, since these 14 CAswere contributed by only five whales (75% of thosewhales with vesicular CAs), and eight of the 14 werefound in a single whale, there may be a tendency for cer-tain individuals to generate vesicular CAs that has littlerelevance to the incidence of lumens in the wider popu-lation. Variability is further compounded by the fact thatthe whale with the greatest number of vesicular CAs inone ovary (8 of 22 total CAs) had no CAs with lumens inthe opposite ovary (19 total CAs).
It could be suggested that large CL size, rather thanthe presence of a fetus per se, favors lumen development.That is, if a CL is sufficiently large, a lumen might beexpected even in a CL of ovulation. Much of the theoreti-cal basis for the concept that a vesicular corpus mightbe indicative of a corpus of pregnancy was put forwardby Dempsey and Wislocki (1941); they suggested thatcavity formation in large CLs resulted from the inabilityof capillaries to reach and nourish the center of so largea mass. Our data appear to challenge this idea since,although the largest vesiculated CL on our sample(15 cm) also had the largest absolute and relative lumen(9.8 cm), there was otherwise no clear indication in ourCL sample of lumen size scaling up with CL size,although Laws (1961) did report larger lumens as CLsize increased in fin whales. Further, our sample con-tained four CLs (largest 5 9.6 cm) without lumensamong the total 12 CLs. Large CLs lacking lumens werealso reported in the blue, fin, humpback and spermwhales (van Lennep, 1950; Chittleborough, 1954; Laws,
CORRELATES OF OVARY MORPHOMETRICS IN THE BOWHEAD WHALE 793

1961; Best, 1967). Laws found nonvesicular CLs up to16.0 cm and 19.0 cm in diameter in fin and blue whales,respectively, with cavities in only 17.1% of fin whaleCLs. In contrast, 75% of bowhead CLs had lumens,inclusive of CLs of pregnancy and ovulation. Similarly,Best found cavities in some CLs of ovulation as well assome of pregnancy and suggested that “the occurrence ofvesicular and nonvesicular types of CLs may simply bethe result of a different degree of rupture of the follicleat ovulation.” Cavities have commonly been described inCLs of ovulation or pregnancy less than 1 cm in diame-ter in women (Dubreuil and Riviere, 1947, cited inLaws, 1961).
The absence of a lumen in the largest CA (3.4 cm) ofone of the lactating bowhead whales (87B6) also sug-gests that CAs representing pregnancies do not alwayshave lumens (assuming lumens do not close soon afterparturition). In fact, none of the 12 CAs in 87B6 werevesicular, an unlikely finding if CLs of pregnancy alwayshave lumens since it would indicate that (1) the whalehad never been pregnant (despite numerous CAs) orthat (2) CA lumens close over time. Neither of these twocircumstances seems likely. While it could be arguedthat lumen closures had occurred in this whale, thisseems doubtful given that several of the smallest CAs inour sample remained vesiculated. Such closure was sug-gested by Dempsey and Wislocki (1941) since theybelieved the lumens were eventually filled with a dense,hyalinized connective tissue scar. Nevertheless, if thelumen can be lost at some point along the regressionpath of a vesicular corpus, even for some of the corpora,the presence of a lumen is even further reduced in itspower to signal CAs from past pregnancies. Finally,while our bowhead whale data suggest a tendency forlarge corpora to have lumens, and that lumens can per-sist even in small corpora, the absence of vesiculatedCAs in 13 out of 22 bowhead whales renders it unlikelyin the bowhead whale, as in the balaenopterids previ-ously examined, that the presence of a lumen holds anypromise for distinguishing CLs of pregnancy.
Corpora Counts
There was no indication in our bowhead sample thatone ovary dominated the other either in the productionof CLs of pregnancy specifically or in corpora generally,notwithstanding several instances of count disparitiesbetween ovaries of a pair for individual whales. In thefour bowhead CLs associated with known pregnancieswhere ovaries had been identified as right or left, CLswere evenly split between right and left ovaries. Best(1967) reported no significance difference in CLs of preg-nancy in sperm whales with 12 in the left ovary andeight in the right ovary; for corpora overall, he deter-mined that 50.8% occurred in the left ovary. Also insperm whales, Ohsumi (1964) found 51.6% of corpora inthe left ovary. Lockyer (1987) found no heightened activ-ity in one ovary than the other in minke whales, agree-ing with Larsen (1984) for that species. In contrast,Laws (1961) combined data with those of Slijper (1949)that indicated greater frequency of CLs of pregnancy inthe right ovary in fin and blue whales, with an incidenceof 59.2% and 55.8%, respectively. When Laws (1961) con-sidered the combined totals for CLs and CAs, ovulatoryevents were closer to parity, with a frequency of 52.4%
in the right ovary in both fin and blue whales. In oursmall sample, a slight suggestion that ovulatory balancebetween ovaries of a pair might increase with increasednumbers of corpora in an ovary pair needs to be exploredfurther.
In odontocetes, the left ovary has been considered themore frequent ovulator (Slijper, 1966). This impressionrelates in part to the high incidence of left horn preg-nancies in odontocetes, though Slijper notes that not allsuch left-sided implantations reflect fertilization of anovum produced in the left ovary since zygote implanta-tion may occur in the horn contralateral to the ovaryproducing the successfully-fertilized ovum. In fact,Slijper maintains that even when fertilizations are asso-ciated with right-sided ovulations, subsequent implanta-tion occurs preferentially in the left horn. In pilotwhales Globicephala melas, Sergeant (1962) found that70% of ovulations occurred in the left ovary from anexamination of 42 ovary pairs.
Corpora Regression, Accumulation, andPersistence
Addressing the question of corpora persistencedepends upon an accurate count and measurement ofcorpora number and size. In this study, we followed theconvention of earlier investigators in transversely sec-tioning the ovaries at approximately 0.5 mm intervals toexpose corpora (e.g., Mackintosh and Wheeler, 1929;Laws, 1961, Gambell, 1968). Since this was accom-plished using a commercial meat slicer, section thicknesscould only be approximated, and it is possible that verysmall corpora could be missed. However, given the largenumber of slices made, it would be likely that corporaless than 0.5 mm in diameter, had they been frequent,would have often been revealed. Since only 3 out of 257corpora exposed and measured were less than 0.5 mm incalculated diameter (one was 0.3 mm and two were0.4 mm), the assumption was made for the purposes ofthis study that few such small corpora were beingmissed.
With respect to corpora accumulation, the importantquestion to ask is whether corpora numbers increase asa whale ages, providing preliminary evidence of persist-ence. The number of corpora in an ovary pair in ourstudy did not correlate with total body length. That is,shorter whales appeared as likely to have as many cor-pora as longer whales. At least two interpretations arepossible: (1) CAs do not accumulate or permanently per-sist with age or, (2) there is sufficient individual varia-tion in adult length that shorter whales may have beenreproductively active for as long or longer than longerwhales. The presence of so many corpora in some of thewhales argues against the lack of accumulation unlessthose whales were for some reason uncommonly produc-tive in numbers of ovulations (or inefficient in concep-tion) over time. Regressing corpora numbers against agewould be more appropriate than against length. Varia-tion in adult body size is well-established among individ-uals of a given species, and length cannot be assumed tocorrelate with age in the bowhead whale. While there isnow some evidence supporting age-related markers inthe bowhead (Rosa et al., 2013), no ages were availablefor the whales in this study.
794 TARPLEY ET AL.

Ninety-seven percent of the CAs in our sample of 257were 3.0 cm or less in diameter, demonstrating either arapid decline in CA size from that attained by the CLsor an extended time interval between ovulations—orboth. Laws (1961) divided regressing fin whale corporainto “young,” “medium,” and “old” classes based on mor-phological appearance. The “young” group had a rangeof 1.5 to 7.5 cm, with a mean of 4.01 cm, while the“medium” group ranged from 0.7 to 5.5 cm(mean 5 2.94 cm). Therefore, in both groups he identifiedmany CAs greater than 3 cm in diameter. Since hereported fin whale CL means of 11.4 cm and 8.28 cm forCLs of pregnancy and ovulation, respectively, and sinceCL size in our bowhead sample was similar, we mightexpect to find more of the younger and larger regressingCAs in our sample than we did. In only 6 of 22 whalesdid we find CAs greater than 3.0 cm: 82WW2 (6.3 cm),86KK2 (4.9 cm), 92B10 (4.5 cm), 92B3 (4.0 cm), 92B4(3.6 cm), 87B6 (3.2 and 3.4 cm). Therefore, even allowingfor an initial rapid involution, the rarity of larger CAssuggests a prolonged period of CA regression and accu-mulation between ovulations in the bowhead reproduc-tive cycle when compared with the balaenopterids.
For ovulating and pregnant bowhead whales, CAsranged in size from 0.6 to 2.4 cm and 0.4 to 2.8 cm,respectively. This group of whales represents those forwhom we would expect CA regression to have been inprogress longest preceding the most recent ovulation,and this seems to be confirmed by the small size of theirCAs. In our two lactating whales, CAs ranged from 0.7to 6.3, with the largest CA in each whale measuring6.3 cm (82WW2) and 3.4 cm (87B6), respectively. Forthese two whales, the larger CAs would be assumed torepresent the previous CLs of pregnancy and would stillbe in the earlier stages of regression. For resting whalesoverall, CAs ranged from 0.3 to 4.9 cm. Yet for manyindividual resting whales, the CA range was restrictedto the smaller diameters. For example, the 16 CAs inwhale 82WW1 ranged only from 0.9 to 1.8 cm. Theimpression is that resting whales with the largest CAswere just entering the resting period, while those withonly the smallest CAs were further into a protractedresting period. This may explain the rather large frac-tion of captured whales in our sample (approximately 1/3) found to be neither pregnant nor lactating. The rest-ing period, and therefore the calving interval, may berelatively long (�3–4 years) in the bowhead (Koskiet al., 1993; George et al., 2011) compared with some ofthe other mysticete species.
Approximately 3=4 of CAs were 2.0 cm or less in diame-ter, with a mode of 1.8 cm (range for all CAs was 0.3–6.3 cm). The greater number of small CAs suggests anoverall slowing of regression rate as the corpus ages.But there is also an increasing decline in the frequencyof those CAs with diameters less than the modal diame-ter of 1.8 cm. No CAs were recognized which were lessthan 0.3 cm in diameter, suggesting the possibility thatin the bowhead whale, as in other mysticetes that havebeen studied, the very small CAs remain as permanentmarkers of ovulation, or alternatively, that such smallcorpora could not be grossly detected. The decline in thenumber of CAs less than the modal 1.8 cm could resultfrom: (1) an acceleration of resorption in the final regres-sion stages, (2) failure to identify very small CAs, or (3)persistence of corpora with the smaller diameters repre-
senting the left tail of a normal distribution for the clus-ter of smaller corpora.
Similar results have been noted in balaenopterid stud-ies that support the probability of corpora persistence(Laws, 1961; Gambell, 1968; Lockyer, 1987). Theseinvestigators divided the corpora into three subjectivetemporal groups (“young,” “medium,” and “old”) on thebasis of macroscopic appearance (amounts of scar tissuecompared with old luteal tissue), in order to assess therelative rates of group accumulation. Since the popula-tion of “old” corpora formed a normal distributionalcurve, it was suggested that these CAs represented acomplete, and therefore persistent, CA population. CAsegregation into temporal groups was not attempted inour study and would likely be less reliable because ofthe small number of CAs available for analysis (althoughsuch segregation should be approached histologically toconsider the possibility of definitive characterizations).An attempt was made to age-characterize bowheadwhale corpora at the macroscopic level by consideringthe proportion of each corpus devoted to scar tissue rela-tive to inactive luteal tissue. However, no age-relevantpattern emerged.
In the context of extracting the “old” CAs from theyounger regressing corpora, it is interesting to examinethe frequency profile of CA size in bowhead whale 89B3where the relatively large number of corpora (41) spansa size range of 0.4 to 2.2 cm. The age estimate for thiswhale was reported as 149 yr (SE 41 yr) (George et al.,2011). The absence of larger CAs suggests ovulatory qui-escence for some period of time [e.g., in the fin whale,Laws (1961) determined that regression to the “old” CAcategory required about 3 years], tending to reflectaccordingly a population of CAs that is relatively “old.”This supposition is supported by the smaller modaldiameter for 89B3 CAs (1.3 cm) relative to that for com-bined corpora (1.8 cm) within our study sample. ThatCAs in a similar stage of regression are represented issuggested by the essentially bell-shaped curve of the dis-tribution around a mean (1.3 cm) in agreement with themode (1.3 cm). While the distribution is somewhatskewed to the left with the mean slightly less than themode, allowing the possibility that the CAs are continu-ing to regress to some degree, the nearly normal distri-bution tentatively supports the argument (in this onewhale at least) for CA persistence as presented in blueand fin whales (Laws, 1961), sei whales (Gambell, 1968),minke whales (Best, 1982; Lockyer, 1987), and spermwhales (Best, 1967; Gambell, 1972). That is, CAs smallerthan 1.3 cm in diameter should still be readily recogniz-able with greater skewing of the distribution to the left.Because the sample size of bowhead whale corpora willalways remain necessarily small, further substantiationof corpora persistence through the analysis of “old” CAsmay rest on examination of additional individual whaleswhere CA numbers are large.
ACKNOWLEDGEMENTS
The authors thank the Alaska Eskimo Whaling Commis-sion for its cooperation and the following subsistencehunters for the use of specimens collected fromharvested whales: Jacob Adams, Tom Agiak, BenAhmaogak, Sr., Fred Ahmaogak, George Ahmaogak, Sr.,Lawrence Ahmaogak, Jonathan Aiken, Herman
CORRELATES OF OVARY MORPHOMETRICS IN THE BOWHEAD WHALE 795

Aishanna, Isaac Akootchook, Ralph Apatiki, Sr., HenryAttungana, Archie Brower, Arnold Brower, Sr., ArnoldBrower, Jr., Eugene Brower, Ronald Coates, Ray Golo-gergen, Edward Hopson, Hansen Irrigoo, Ben Itta,Edward Itta, Lloyd Kittick, Luke Koonook, Sr., LutherLeavitt, Jr., Alfred Linn, Sr., Roxy Oyagak, Sr., SimeonPatkotak, Sr., Rossman Peetook, Nolan Soloman, BillyTagarook, Raymond Toolie, Alfred Tukle, Joash Tukle,Patsy Tukle, and Alex Oozeva. Participating villagesincluded Barrow, Gambell, Kaktovik, Nuiqsut, PointHope, Savoonga and Wainwright. The authors are espe-cially grateful to Dr. Tom Albert who coordinated the sci-entific effort in the North Slope Borough Department ofWildlife Management during the study collection periodand to the following field personnel who gathered mate-rials for this study: Billy Adams, Susie Akootchook,Perry Anashugak, Leonard Apanagalook, Linda Ban-nish, David Barnard, John Basler, Karen Brewster,Charlie Brower, Joe Burgener, Geoff Carroll, RonClarke, Jim Cubbage, Carol Dalton, Les Dalton, DianaDronenburg, Tom Ford, Lane Franich, Darold Frankson,Aggie Frankson, Jean Craighead George, T. LukeGeorge, Carolyn Glass, Cyd Hanns, Craig Harms, MarkHarrington, Katherine Hazard, Tim Holmes, TammyHughes, Henry Huntington, Clarence Itta, Gordon Jar-rell, Darrell Kava, Luke F. Kagak, Richard Kovalsky,Mark Leinberger, Gary Miller, Paul Nader, BennyNageak, Charles Nageak, Charles Nayakik, Silas Nego-vanna, Scott Newman, Leland Nuehring, Hiram Okhto-kiyuk, Forrest Dean Olemaun, Conrad Oozeva, JackPanik, Mike Philo, Mike Pielorz, Ginny Pierce, LaurieQuakenbush, Dave Ramey, Ketil Reitan, Elijah Rock,Sr., Dave Smith, Nolan Soloman, Gary Tabor, GeorgeTagarook, George Taalak, Sam Taalak, Harry Tazruk,Kim Vinette, Jennifer Williams, Stephen Wood, SallyYackley, Joanne Young and Cort Zachel.Authors appreciate the early support of BenjaminNageak, Warren Matumeak, Charles Brower, formerDirectors of the North Slope Borough Department ofWildlife Management in Barrow, Alaska, through allphases of the study as well as the logistical assistance ofthe department’s office staff, including Dolores Vinasand Liza Dela Rosa. Noe Texeira, also of the wildlifedepartment, assisted with ovary morphometry. Weappreciate the support of current Director Taqulik Hepafor supporting the recent submission of this three-decade project.At Texas A&M University the authors are indebted toDr. George G. Stott for his work on follicular analysisand Barbara Merka for histological preparations, TonyRydzewski for photographic preparations and BarbaraGuidry for assistance in accessing the research litera-ture. Authors appreciate editorial reviews of the manu-script by Mike Dossey and Christine Curry.
LITERATURE CITED
Albert TF. 1985. Scientific Research Activities Concerning the Col-lection and Distribution of Marine Mammal Specimen Materialsunder Authority of Permit #345 issued by the National MarineFisheries Service. Report to the National Marine Fisheries Serv-ice from the Department of Conservation and Environmental Pro-tection. North Slope Borough, Barrow, Alaska: Department ofConservation and Environmental Protection. p 56.
Angliss RP, Rugh DJ, Withrow DE, Hobbs RC. 1995. Evaluations ofaerial photogrammetric length measurements of the Bering-Chukchi-Beaufort seas stock of bowhead whales (Balaena mystice-tus). Rep Int Whal Commn 45:313–324.
Best PB. 1967. The sperm whale (Physeter catodon) off the westcoast of South Africa. 1. Ovarian changes and their significance.Invest Rep Div Sea Fish S Afr 61:1–27.
Best PB. 1977. Two allopatric forms of Bryde’s whale off SouthAfrica. Rep Int Whal Commn Special Issue 1:10–34.
Best PB. 1982. Seasonal abundance, feeding, reproduction, age andgrowth in minke whales off Durban (with incidental observationsfrom the Antarctic). Rep Int Whal Commn 32:759–786.
Brodie PF. 1971. A reconsideration of aspects of growth, reproduc-tion and behavior of the white whale (Delphinapterus leucas)with reference to the Cumberland Sound, Baffin Island popula-tion. J Fish Res Board Can 28:1309–1318.
Brodie PF. 1972. Significance of accessory corpora lutea in odonto-cetes with reference to Delphinapterus leucas. J Mamm 53:614–616.
Chittleborough RG. 1954. Studies on the ovaries of the humpbackwhale, Megaptera nodosa (Bonnaterre), on the western Australiancoast. Aust J Mar Freshw Res 5:35–63.
Beyer WH, editor. CRC handbook of tables for Probability andStatistics. 2nd ed. 1968. Chemical Rubber Company. p 230–235.
Dempsey EW, Wislocki GB. 1941. The structure of the ovary of thehumpback whale (Megaptera nodosa). Anat Rec 80:243–257.
Gambell R. 1968. Seasonal cycles and reproduction in sei whales ofthe southern hemisphere. Disc Rep 35:31–134.
Gambell R. 1972. Sperm whales off Durban. Discov Rep 35:199–358.
George JC, Koski WR, Suydam R, Rugh D. 2004. Body stretching ofbowhead whales during hauling and butchering during the sub-sistence hunt. Paper SC/56/BRG9 presented to the Scientific Com-mittee of the International Whaling Commission 19–22 July,Sorrento, Italy.
George JC, Follmann E, Zeh J, Sousa M, Tarpley R, Suydam R,Horstmann-Dehn L. 2011. A new way to estimate the age of bow-head whales (Balaena mysticetus) using ovarian corpora counts.Can J Zool 89:840–852.
Givens GH, Edmondson SL, George JC, Suydam R, Charif RA,Rahaman A, Hawthorne D, Tudor B, DeLong RA, Clark CW.2016. Horvitz-Thompson whale abundance estimation adjustingfor uncertain recapture, temporal availability variation, and inter-mittent effort. Environmetrics (Wileyonlinelibrary.com) DOI:10.1002/env.2379.
Gundersen HJG, Bendtsen TF, Korbo L, Marcussen NA, MoIler A,Nielsen K, Nyengaard JR, Pakkenberg B, Sorensen FB, VesterbyA, West MJ. 1988. Some new, simple and efficient stereologicalmethods and their use in pathological research and diagnosis.Apmis 96:379–394.
Kenney RM, Garcia MC, Everitt JI. 1981. The biology of the repro-ductive and endocrine systems of the bowhead whale, Balaenamysticetus, as determined by evaluation of tissues and fluids fromsubsistence-killed whales. In: Albert T, editor. Tissue structuralstudies and other investigations of the biology of endangeredwhales in the Beaufort Sea. Final Report to the Bureau of LandManagement from the Department of Veterinary Science. Univer-sity of Maryland, College Park, MD: Department of VeterinaryScience. p 89–156.
Koski WR, Davis RA, Miller GW, Withrow DE. 1993. Reproduction.In: Burns JJ, Montague JJ, Cowles CJ, editors. The bowheadwhale. Special Publication Number 2. The Society for Mammal-ogy. Allen Press, P.O. Box 368, Lawrence, Kansas 66044, p 239–274.
Koski WR, Rugh DJ, Punt AE, Zeh J. 2006. An approach to mini-mize bias in estimation of the length-frequency distribution ofbowhead whales (Balaena mysticetus) from aerial photogrammet-ric data. J Cetacean Res Manage 8:45–54.
Larsen F. 1984. Reproductive parameters of the minke whale,Balaenoptera acutorostrata, off West Greenland. Rep. int. Whal.Commn. (Special Issue 6): 233–236.
796 TARPLEY ET AL.

Laws RM. 1957. Polarity of whale ovaries. Nature 179:1011–1012.Laws RM. 1961. Reproduction, growth and age of southern fin
whales. Disc Rep 31:327–486. 1 plates 4-7.Lennep EWv. 1950. Histology of the corpora lutea in blue and
fin whale ovaries. P K Ned Akad Wetensc Amsterdam 53:593–599.
Lockyer C. 1984. Review of baleen whale (Mysticeti) reproductionand implications for management. In: Perrin WF, Brownell RL,DeMasster DP, editors. Reproduction in whales, dolphins and por-poises. Cambridge: Rep Int Whal Commn (Special Issue 6). p 27–50.
Lockyer C. 1987. Observations on the ovary of the southern minkewhale. Sci Rep Whales Res Inst Tokyo 38:75–89.
Mackintosh NA, Wheeler JFG. 1929. IV. The reproductive organs.Disc Rep 1:379–471.
Marsh H. 1985. Observations on the ovaries of an isolated minkewhale: evidence for spontaneous sterile ovulation and structure ofthe resultant corpus. Sci Rep Whales Res Inst 36:35–39.
Matthews LH. 1937. The humpback whale, Megaptera nodosa. DiscRep 17:7–92.
Matthews LH. 1938. The sei whale, Balaenoptera borealis. Disc Rep17:183–290.
Mossman HW, Duke KL. 1973. Comparative mophology of themammalian ovary. Madison: University of Wisconsin Press. p372.
Nerini MK, Braham HW, Marquette WM, Rugh DJ. 1984. Life His-tory of the bowhead whale, Balaena mysticetus (Mammalia:Ceta-cea). J Zool Lond 204:443–468.
Ohsumi S. 1964. Comparison of maturity and accumulation rate ofcorpora albicantia between the left and right ovaries in Cetacea.Sci Rep Whales Res Inst Tokyo 18:123–148.
Ommanney FD. 1932. The urino-genital system of the fin whale(Balaenoptera physalus). Disc Rep 5:363-466 1 plates 2–3.
Peters N. 1939. Zool Anz 127:193.Rice DW, Wolman AA. 1971. The life history and ecology of the gray
whale, Eschrichtius robustus. Special Publication No. 3. AmericanSociety of Mammalogists, Stillwater, Oklahoma.
Robins JP. 1954. Ovulation and pregnancy corpora lutea in the ova-ries of the humpback whale. Nature 173:201–203.
Rosa C, Zeh J, George JC, Botta O, Zauscher M, Bada J, O’Hara T.2013. Age estimates based on aspartic acid racemization for bow-head whales (Balaena mysticetus) harvested in 1998-2000 and therelationship between racemization rate and body temperature.Marine Mamm Sci 29:424–445.
Sergeant DE. 1962. The biology of the pilot or pothead whale Globi-cephala malaena (Traill) in Newfoundland waters. Bull Fish ResBoard Can 132:1–84.
Slijper EJ. 1949. On some phenomena concerning pregnancy andparturition in cetacean. Bijd Dierk 28:416.
Slijper EJ. 1966. Functional Morphology of the Reproductive Sys-tem in Cetacea. In: Norris KH, editor. Whales, dolphins, and por-poises. Los Angeles: University of California Press. p 277–319.
Tetsuka M, Asada M, Mogoe T, Fukui Y, Ishikawa H, Ohsumi S.2004. The pattern of ovarian development in the prepubertal Ant-arctic minke whale (Balaenoptera bonaerensis). J Reprod Dev 50:381–389.
CORRELATES OF OVARY MORPHOMETRICS IN THE BOWHEAD WHALE 797