molecular determinants of the species-selective inhibition of brain acetylcholinesterase
TRANSCRIPT

rOXICOLoCY AND APPLIED PHARMACOLOGY 104,246-258 ( 1990)
Molecular Determinants of the Species-Selective inhibition of Brain Acetylcholinesterase
JON R. KEMP AND KENDALL B. WALLACE’
Department of Pharmacology, University of Minnesota, School of Medicine, Duluth, Minnesota 55812
Received August 18, 1989; accepted March 6, 1990
Molecular Determinants of the Species-Selective Inhibition of Brain Acetylcholinesterase. KEMP, J. R.. AND WALLACE, K. B. (1990). Toxicol. Appl. Pharmacol. 104, 246-258. The objective of this investigation was to distinguish which of the catalytic features of enzyme action is principally responsible for conferring the observed insensitivity of trout brain acetylcholines- terase (AChE, EC 3.1.1.7) to in vitro inhibition by organophosphates. The experimental design consisted of comparing the kinetic constants for the hydrolysis of a series of acylthiocholine substrates as well as the inhibition constants for a homologous series of dialkyl pnitrophenyl phosphates among AChE from rats, hens, and trout. Chicken and rat brain AChE failed to distinguish between acetyl- and propionylthiocholine as inferred from the comparable Michae- lis-Menten constants (K,), whereas trout brain AChE exhibited a much higher affinity for ace- tylthiocholine than for either of the two larger analogs. Diethyl pnitrophenyl phosphate was the most potent inhibitor toward chicken and rat brain AChE, whereas the IC50 for the trout en- zyme increased progressively between dimethyl and di-n-propyl pnitrophenyl phosphate. The kinetic constants revealed that a significant determinant of inhibitor potency in the chicken and rat is steric exclusion as reflected by changes in the dissociation constant (&) which paralleled the changes in IC50 and k,. Conversely, Kdwas 120-to 1450-fold higher and did not vary signifi- cantly for trout brain AChE. Instead, the phosphorylation rate constant (I$,) for trout brain AChE decreased with progressive methylene substitutions. The kinetic data suggest that trout brain AChE not only possesses less steric tolerance, but also has a weaker nucleophile at the esteratic subsite, both ofwhich may be important factors in conferring the observed insensitivity of trout to acute organophosphate intoxication. o 1990 Academic PKSS, IW.
The relative insensitivity of most species of fish to acute intoxication by organophospho- rus compounds such as Parathion (O,O-di- ethyl p-nitrophenyl phosphorothionate) is well established; the acute intraperitoneal LD50 for the biologically active oxygen ana- log, Paraoxon, varies from 1.1 and 2.6 pmol/ kg in hens and rats, respectively, to 27.3 rmol/kg in sunfish (Benke et al., 1974; Chat- topadhyay et al., 1986). Attempts to elucidate the principal factor which confers this resis- tance to fish have focussed on species-related differences in pharmacokinetic variables, in-
’ To whom correspondence should be addressed.
eluding differences in the absorption, distri- bution and elimination of an acute dose, and species-related differences in metabolic trans- formation (activation or detoxification) of the organophosphates. Despite obvious ana- tomical and physiological differences, it has been concluded that the species-selective tox- icity of organophosphates reflects more than simply subtle differences in kinetic disposi- tion (Benke et al., 1974; Hodson, 1985). Studies designed to assess the significance of species-related differences in metabolic acti- vation or detoxification as major determi- nants of species-selectivity are complicated by the fact that the steady-state concentration
0041-008X/90 $3.00 Copyright 0 1990 by Academic Press, Inc. 246 All rights of reproduction in any form resewed.

INHIBITION OF BRAIN ACETYLCHOLINESTERASE 247
of the biologically active intermediate is a function of the relative activities of a group of nonspecific esterases acting both in series and in parallel to inactivate the reactive oxygen analog formed by the cytochrome P450- mediated oxidative desulfuration of Para- thion. Consequently, despite large differences among species in the activities of the mixed function oxidases and the nonspecific ester- ases responsible for the activation and detoxi- fication of organophosphonates, respectively, the significance of metabolic transformation as a major determinant of species selectivity in organophosphorus poisoning remains questionable (Potter and O’Brien, 1964; Murphy, 1966; Hitchcock and Murphy, 197 1; Benke et al., 1974; Wang and Murphy, 1982b; Wallace and Dargan, 1987).
Recent evidence suggests that the insensi- tivity of fish to organophosphate toxicity, rather than being a pharmacokinetic phe- nomenon, may reflect pharmacodynamic differences between species, manifested at the level of inhibition of acetylcholinesterase (AChE) by the active intermediate. The IC50 values for the inhibition of brain or plasma cholinesterase by Paraoxon vary over a 2000- fold range between species, with chickens be- ing the most sensitive, rats intermediate, and fish the least sensitive species (Dauterman and O’Brien, 1964; Murphy et al., 1968; Ecobichon and Comeau, 1973; Benke et al., 1974; Andersen et al., 1977; Wang and Mur- phy, 1982a,b; Johnson and Wallace, 1987). These differences are consistent with the differences in the in vivo sensitivity among species. Several groups of investigators (Becker et al., 1964; Darlington et al., 1971; Lotti and Johnson, 1978) reported direct cor- relations between the in vitro IC50 for AChE and the acute LD50 for a wide array of or- ganophosphates in rodents, insects, and chickens. Accordingly, the sensitivity of AChE to inhibition in vitro appears to be a principal determinant of the species-selective toxicity of anticholinesterase agents and may be employed as a convenient screen for inter- species comparisons.
On a molecular basis, these differences in enzyme sensitivity may reflect any one or combination of several catalytic properties of AChE; (1) the dimensions of the active site, (2) the strength of the nucleophilic center re- sponsible for interacting with the phosphoryl group, (3) the influence of a possible hydro- phobic subsite, and (4) the distance separat- ing the esteratic and anionic subsites. Struc- ture-activity studies have revealed isolated differences in some of these properties be- tween selected enzymes. Zahavi and co- workers (197 1) suggested that resistance in certain strains of mites to Malaoxon is con- ferred by a sterically smaller esteratic site, thereby excluding the large phosphoryl esters. Alternatively, species-related differences in sensitivity to inhibition may reflect differ- ences in the proximity of the anionic site to the esteratic site (O’Brien, 1963; Holling- worth et al., 1967). Moss and Famey (1978) reported that although fish AChE possesses both anionic and esteratic subsites, the posi- tive cooperativity between these regions is less than that exhibited by mammalian AChE, which is consistent with the resistance of the fish enzyme to inhibition by organo- phosphates. The objective of the present in- vestigation was to elaborate on the observed differences between rats, chickens, and rain- bow trout in in vitro sensitivity of AChE in an attempt to identify the principal catalytic feature of enzyme action responsible for the large idiosyncrasies between species. The in- vestigation was limited to assessing species- related differences in only those variables associated directly with the esteratic site of AChE (steric dimensions, nucleophilic strength, hydrophobic subsite). The paper ex- pands beyond previous reports (Wang and Murphy, 1982a) to include a larger series of five structurally related analogs of Paraoxon, allowing for more definitive deductions re- garding molecular and catalytic distinctions between different enzymes. Furthermore, the reactions were conducted at the respective physiological temperatures and, in order to eliminate possible confounding influences of

248 KEMP AND WALLACE
isozyme heterogeneity and allosterism be- tive in inhibiting AChE. Every 2 months neat dialkyl p
tween species, a homogeneous preparation of nitrophenyl phosphates were dissolved in methylene
detergent-solubilized membrane-associated chloride and washed with 5% sodium carbonate to re-
AChE was employed. Finally, the investiga- move hydrolysis products (pnitrophenol). From this or- eanoohosphate reserve. stock solutions were orepared in
tion employs a novel kinetic method (Hart absolute ethanol (stable for 1 week at -20”C)br in buffer and O’Brien, 197 3) to circumvent important containing 10% ethanol and 20% Triton X-100 (stable
restrictions imposed by the extreme rapidity for 8 hr at -20°C). Final concentrations of ethanol and
of the inhibition reaction. Triton X-100 in the reaction medium did not exceed 1 and 2.25%, respectively, neither of which affected the ac- tivity of AChE.
METHODS
Triton X- 100, acetyl-, propionyl-, and butyrylthiocho- line iodide were purchased from Sigma Chemical Co. (St. Louis, MO). All other chemicals were reagent grade as supplied by Aldrich Chemical Co. (Milwaukee, WI). Di- methyl, diethyl, di-n-propyl, diisopropyl, and di-n-butyl chlorophosphates were synthesized by a modification of the method of Fiszer and Michalski ( 1952) and M&or et al. ( 1956). Briefly, three equivalents of the appropriate alcohol were added dropwise to one equivalent of cold (5-1o’C) phosphorus trichloride in hexane; an icebath was used to maintain the reaction temperature below 10°C. Immediately after the addition of alcohol, one equivalent of sulfuryl chloride was introduced dropwise to the newly formed trialkylphosphite. The hexane was removed under reduced pressure (water pump aspira- tion) and the dialkyl chlorophosphate purified by vac- uum distillation.
White Leghorn hens (2-2.5 years old: 1300-2000 g) were supplied by Morterud’s Eggs, Inc. (Duluth, MN), and were killed by cervical dislocation. Male Sprague- Dawley rats (175-250 g) were purchased from Harlan, Inc. (Indianapolis, IN), and were housed under climate- controlled conditions on a I2-hr light cycle and provided food and water ad libitum. All rats were fasted for at least 12 hr prior to their use and were killed by decapitation. Unsexed rainbow trout (750-1250 g) were purchased from Cedar Bend Trout Farm (Scandia, MN) and main- tained at the U.S. EPA Environmental Research Labora- tory (Duluth, MN) under regulated conditions and killed by a blow to the head.
The various dialkyl pnitrophenyl phosphates were then prepared from the respective dialkyl chlorophos- phates and sodium pnitrophenoxide by a modification of the methods of Fletcher et al. ( 1950) Schrader ( 1952) and DeRoos and Toet (1958). Specifically, sodium pni- trophenoxide was suspended in ethanol (I .5 M) and neat dialkyl chlorophosphate was added dropwise, producing a faint exothermic reaction. After the addition ofthe dial- kyl chlorophosphate, the mixture was heated for 90 min at 87°C under a reflux apparatus. The reaction was cooled to room temperature then filtered to remove so- dium chloride. Ethanol was removed by rotoevaporation under reduced pressure (water pump aspirator) and the resulting product was dissolved in methylene chloride and washed with a 5% aqueous solution of sodium car- bonate to remove unreacted sodium pnitrophenoxide. The methylene chloride was dried with anhydrous mag- nesium sulfate, filtered, and then removed by rotoevapo- ration. The resulting dialkyl pnitrophenyl phosphate was then heated under reduced pressure (0.2 mm Hg) to remove traces of unreacted dialkyl chlorophosphate (Fletcher et al., 1950). Purity of the isolated dialkyl p- nitrophenyl phosphates were greater than 95% as deter- mined by GC-FID, and confirmed by GC-MS and HPLC-uv. The only detectable impurity was the corre- sponding chlorophosphate which was found to be inac-
Membrane-associated AChE from brain homogenates was solubilized in detergent to recover the majority of the enzyme. Solubilization of 100,OOOg pellet of brain ho- mogenates with Triton X-100 yields 90-95% of the total enzyme activity which, for rats and chickens, is almost exclusively in the form ofa globular 10 S or I 1 S tetramer of AChE (Rakonczay et al., 198 1; Vallette et al., 1983; Rotundo, 1984). Such a homogeneous enzyme prepara- tion allows for comparisons of AChE among species without concern for possible differences in the composi- tion of isozymes present and eliminates any possible differences in allosteric regulation of the enzyme due to species-related differences in membrane lipid composi- tion (Ott, 1985; Stieger and B&beck. 1985).
For each experiment, brain homogenates from the cor- responding species were prepared from a single animal, with the exception of rainbow trout (two brains were pooled for each experiment). The brain was quickly transferred to ice-cold pH 8.0 buffer [IO0 mM sodium phosphate, 400 IIIM sodium chloride, and 3 mM sodium ethylenediaminetetraacetic acid (EDTA)] and weighed. For hens and rats a 20% (w/v) brain homogenate was pre- pared in a Wheaton glass homogenizer with Teflon pes- tel; a 12.5% brain homogenate was prepared from trout brain. The homogenate was centrifuged at lOO,OOOg for 60 min (4’C). The resulting supematant was discarded and the pellet resuspended in an equal volume of pH 8.0 buffer containing 100 mM sodium phosphate, 3 mM so- dium EDTA, and 0.5% Triton X-100 and recentrifuged at 100,OOOg for 60 min. Sodium chloride was dissolved in the resulting supematant to a final concentration of 400 mrvr and the solution used that same day for all ki- netic analyses and IC50 determinations.

INHIBITION OF BRAIN ACETYLCHOLINESTERASE 249
Cholinesterase assays were performed spectrophoto- metrically utilizing the method of Ellman et al. (196 1) as previously described (Johnson and Wallace, 1987). Ace- tyl-, propionyl-, and butyrylthiocholines have proven to be representative substrates for a variety ofAChE sources (Andersen and Mikalsen, 1978), with Michaelis-Menten constants (K,,,) corresponding to the oxygen analogs. Pro- tein content for each enzyme preparation was deter- mined by the technique described by Lowry et al. (195 1) using bovine serum albumin as standard.
To determine K,,, and I’,,,,, for the entire series of sub- strates within each species, an excess of 5,5’-dithiobis(2- nitrobenzoic acid) (DTNB) was added to AChE in buffer. The reactions were initiated by adding small aliquots of varying concentrations of the respective substrate to yield a final volume of 2 ml. Concentrations of substrate were within a range of one-fifth to five times the esti- mated Km and the amount of AChE included in the assay was adjusted to yield a maximum rate of0.05 absorbance unit per minute (A&min) at the highest substrate con- centration employed. Corresponding blanks lacking AChE were subtracted to yield the enzymatic rate. Ki- netic incubations for rat and hen brain AChE were con- ducted at 37°C. The reactions for the trout enzyme were performed at 11°C which is the temperature at which the fish were acclimated. Finally, determinations of rat AChE kinetics were repeated at I 1 “C in order to compare the values with those for trout. The maximum velocity of substrate hydrolysis (V,,,,,) and the Michaelis-Menten constant (K,,,) were estimated by the double-reciprocal method of Lineweaver and Burk (1934), the line being calculated by a least-squares regression.
ICSO values were determined as described in detail by Johnson and Wallace (1987), wherein the enzyme was preincubated for 30 min with differing concentrations of the inhibitor prior to measuring residual AChE activity. For all reactions, the enzyme was diluted such that in the absence of inhibitor, the rate of acetylthiocholine hydro- lysis was approximately 0.1 A,,,/min. The data were ana- lyzed by linear regression of the natural log (In) of inhibi- tor concentration versus percentage remaining enzyme activity following incubation for 30 min in the presence of inhibitor. A value of 50% was substituted into the re- sulting equation to solve for ICSO.
The bimolecular inhibition constants for the organo- phosphates were estimated in one of two ways. The first method was employed to determine the inhibitor kinetic constants for rainbow trout brain AChE as described by Johnson and Wallace (1987),
[E] + [I] 2 [EI] 1 [EI’]
li,,,, = WI/(& + 14)
where k.pp is the observed velocity of inhibition; kp, the first-order phosphorylation rate constant; and Kd, the dissociation constant for the enzyme-inhibitor complex.
The bimolecular inhibition constant, k,, is equal to k,/ Kd. Ten microliters of the appropriate concentration of inhibitor was incubated with 90 ~1 of the AChE prepara- tion (which at Time zero yielded anA.&min ofO.l). The reactions were stopped at the indicated times by adding 1.5 ml of 1 mM acetylthiocholine- 1 mM DTNB and the
A&min was recorded immediately over 5 min. Addi- tion of excess substrate was effective at preventing the continued inhibition of AChE and provided reliable ki- netic measurements. This procedure was repeated with six different concentrations of each dialkyl p-nitrophenyl phosphate.
Although the above kinetic method worked well for the trout enzyme, it was unsatisfactory for rat or hen AChE. With these species, the reaction rates were so rapid that it was virtually impossible to employ inhibitor concentrations between one-fifth and five times the Kd
as required. According to the above equation, when [I] B Kd, kam becomes a direct function of [I]; k., = k, [I]/ Kd = k;[I]. As expected, double-reciprocal plots of the data intersected very near the origin yielding spuriously high estimates of both k, and Kd. Accordingly, it was nec- essary to modify the kinetic method for rat and hen AChE in order to approach the Kd for the enzyme and to obtain accurate estimates of the kinetic constants. This was accomplished by including a known concentration of acetylthiocholine in the reaction mixture to compete with, and thus retard, the inhibition of rat or hen AChE by the organophosphates (Hart and O’Brien, 1973). Ten microliters of the appropriate concentration of inhibitor was added to 1.5 ml of buffer containing 1 mM DTNB and 600 or 400 pM acetylthiocholine for the chicken or rat, respectively. This solution was equilibrated to 37°C then 90 ~1 of the corresponding AChE preparation (added at a concentration which in the absence of inhibi- tor yielded a rate of0. I Aalz/min) was added and the pro- gressive decrease in the rate of substrate hydrolysis was recorded over a IO-min period. This was repeated with at least six concentrations of each dialkyl pnitrophenyl phosphate. The reaction can be represented by
[E] + [I] 2 [E.] 2 [E-I]
kl where, Kds = Kd(l + [S]/K,)
k-* It kz
tES1 therefore, k.,, = kp’[ll Kis + [II
k,
[El + [PI 1 Kds 1 1
and G=k,‘cI]+k,’
where Kds is the apparent Kd for the enzyme-inhibitor complex determined in the presence of substrate, KS is the Michaelis-Menten constant (k-,/k,) determined for acetylthiocholine in the species under study, and [S] is the concentration of acetylthiocholine added to the reac- tion.

250 KEMP AND WALLACE
TABLE I
V,, (nmol/min . mg protein) Hen (37°C) Rat (37°C) Rat(ll”C) Trout ( 11 “C)
Km (FM) Hen (37°C) Rat (37°C) Rat(ll”C)
Trout ( 11 ‘C)
Acetylthiocholine Propionylthiocholine
350.1 f 44.4” 266.8 + 28.7h 289.5 + 6.7 185.8 + 5.3* 159.0 f 3.7 86.2 f 2.7 b 148.9 f 11.1 15.1 f 1.0*
105.2 * 7.0* 117.7+4.9* 77.5 * 3.3* 83.3 f 4.2’ 99.1 * 4.0* 92.3 f 9.3b
197.2 i 51.3’ 2078.5 2 203.2b
Butyrylthiocholine
16.7 f 1.2’ 8.7 + 0.5’ 2.4 kO.lb
n.d.’
23.0 k 1.2* 337.8 + 40.5’ 183.3 f 22.8*
n.d.’
’ Values represent the means ? SE of three separate enzyme preparations. Means underscored by the same line are not significantly different (p < 0.05).
* Significantly different from the corresponding values for all other enzyme preparations (p < 0.05). ’ n.d. signifies that the reaction proceeded too slow to allow for reliable estimation ofthe kinetic constants.
In this scheme, as with the method used for trout brain AChE, the k,, for each inhibitor concentration was de- termined by regressing residual enzyme activity @[El,/ [El,) against time (Fig. I): then the double-reciprocal plot was formed with the inverse of each inhibitor concentra- tion regressed against the inverse of -kapp (Fig. 2). The reciprocal of the y-intercept is equal to the k, and the negative reciprocal of the x-intercept is equal to I&; however, a correction factor must be intejected to deter- mine the actual Kd and thus the k, (Kd = K&/( I + [S]/K,) and k, = kp/Kd).
Statistical analyses of the log transformed data were accomplished by analysis of variance and the individual means compared by the method of Scheffe for all possible comparisons both within and across species (Sokal and Rohlf, I98 I). A probability of p < 0.05 was used as the criterion for statistical significance.
RESULTS
The kinetic constants (V,,, and K,,J de- scribing the hydrolysis of the three acylthio- choline substrates by AChE from each of the three species (including the rat at 11°C) are presented in Table 1. Within each species the V,,,, for the individual thiocholine substrates were significantly different from each other; however, in the trout it was impossible to measure either V,,,,, or K,,, for butyrylthio- choline as the limits of solubility for this com-
pound were reached before any appreciable rate of hydrolysis could be measured. For each species studied, acetylthiocholine was hydrolyzed at the fastest rate, followed by propionyl- and then butyrylthiocholine; however, there were large differences in the degree of substrate preference among species (Table 1). For rat and chicken AChE, the V,,,,, for propionylthiocholine was between 64 and 76% of that for acetylthiocholine. In contrast, the trout enzyme exhibited a more profound preference for the smaller analog, hydrolyzing propionylthiocholine at only 10% of the maximum rate for acetylthiocho- line. This idiosyncrasy of the trout brain en- zyme was not due to the different tempera- ture at which the reaction was conducted since the rat enzyme incubated at 11 “C exhib- ited a substrate specificity comparable to that observed at 37°C. Comparing substrate speci- ficity among species, there was no significant difference between the maximum rate of ace- tylthiocholine hydrolysis for the hen or rat AChE or between trout and rat when both ex- periments are performed at 11°C. For any given substrate, all other possible compari- sons across species were significantly dif- ferent.

INHIBITION OF BRAIN ACETYLCHOLINESTERASE 251
TABLE 2
IC~OFORTHEINHIBITIONOFBRAINACETYLCHOLINESTERASEBYVARIOUS DIALKYL-p-NITROPHEN~LPHOSPHATES
Dialkyl-p-nitrophenyl phosphate IC50 (pM)
Methyl Ethyl n-Propyl n-Butyl Isopropyl
Species Hen (37°C) 0.037 + 0.003” 0.028 -+ 0.000 0.336 -+0.013’ 0.762 _+ 0.053 3.30+0.10b Rat (37°C) 0.029 f 0.00 1 0.026 + 0.001 0.228 f 0.004b 0.453 + 0.008 2.12 k 0.056
Rat(ll”C) 0.108~0.005b 0.109 k o.002b 3.280 t 0. I 16” 8.400 f 0.740’ 35.64 + 1.76’
Trout (11°C) 3.3 1 + 0.20” 70.04 * 4.95b 963.0-t 137.1b 763.6 + 142.7’ 12,779 rt 1002’
’ Values represent the means -C SE of three separate enzyme preparations. Means underscored by the same line are not significantly different (p < 0.05).
b Significantly different from the corresponding value for all other species (p < 0.05).
The chicken and rat enzyme failed to dis- tinguish between acetyl- and propionylthio- choline as indicated by the comparable val- ues for K, for the two substrates (Table I), whereas the trout was far (IO-fold) more se- lective for acetylthiocholine than for the pro- pionyl analog. For rat and trout AChE, buty- rylthiocholine yielded a significantly greater K,,, compared to the two smaller acylthiocho- lines; however, the K,,, for the butyryl analog in the chicken was 4.8 times lower than the K,,, for either the acetyl or propionyl deriva- tive. All possible comparisons of K,,, across species for a given acylthiocholine were sig- nificantly different.
Diethyl p-nitrophenyl phosphate was the most potent inhibitor for both chicken and rat brain AChE, with an IC50 of 28 and 26 nM, respectively (Table 2). Dimethyl p-nitro- phenyl phosphate was almost as potent in the terrestrial species, displaying an IC50 of 37 nM for chicken AChE and 29 nM for rat AChE. For rat and chicken AChE, a dramatic transition in potency occurred between di- ethyl and di-n-propyl pnitrophenyl phos- phate, the IC50 increasing 12- and 9-fold, respectively. Potency continued to de- crease with increasing methylene substitution through di-n-butyl p-nitrophenyl phosphate; diisopropyl p-nitrophenyl phosphate being the least potent inhibitor of the series for all
species studied. Repeating the IC50 determi- nations for rat AChE at 11°C yielded a trend similar to that observed at 37°C; IC50 in- creased with increasing methylene substitu- tion. The individual IC50 values for each in- hibitor were, however, greater at the lower temperature. For the chicken and rat AChE, all possible comparisons among the different inhibitors were significantly different (except when the rat enzyme was incubated at 11°C dimethyl and diethyl g-nitrophenyl phos- phate were not statistically different).
The rainbow trout brain AChE was most sensitive to inhibition by dimethyl pnitro- phenyl phosphate (Table 2). However, the IC50 was approximately 1 OO-fold higher than that for rat or chicken (IC50 = 3 PM). Unlike the rat or chicken enzyme, inhibitor potency toward trout AChE decreased 23-fold as the result of including a single additional methy- lene group (diethyi analog IC50 = 70 PM).
This trend continued in a stepwise fashion for the remaining inhibitors; however, di-n-butyl pnitrophenyl phosphate (IC50 = 764 PM) was found to be equipotent with di-n-propyl p-nitrophenyl phosphate (IC50 =963 PM). All other possible inhibitor IC50 compari- sons within the trout were significantly different from each other. When comparing each inhibitor across species, there was no difference in the IC50 for dimethyl, diethyl,
252 KEMP AND WALLACE
Minutes
-0.5
0 iii v
iii - -1.5
C
-2.5
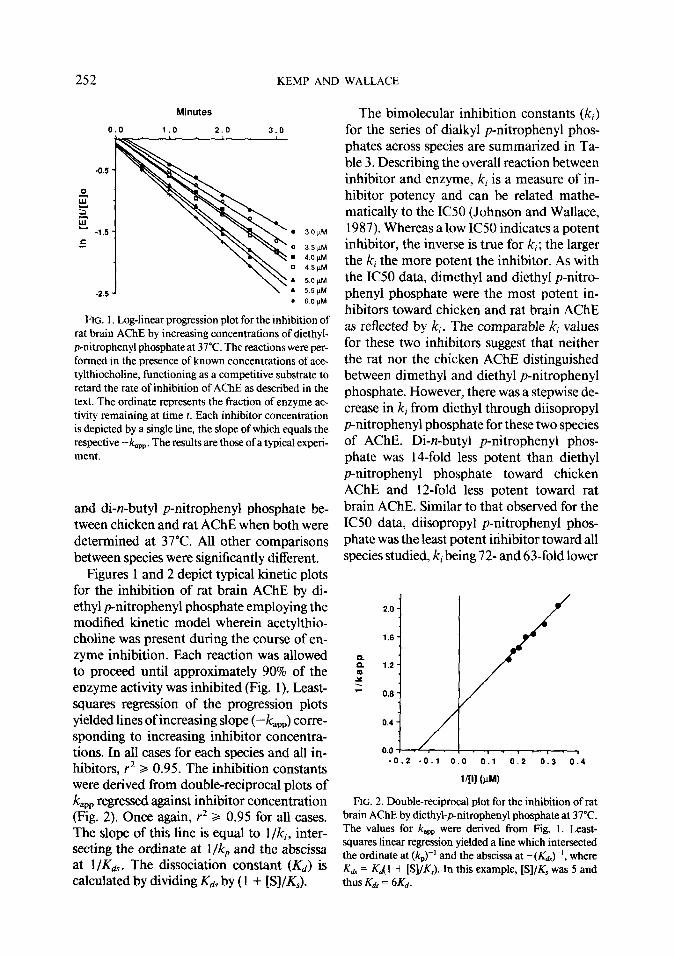
FIG. 1. Log-linear progression plot for the inhibition of rat brain AChE by increasing concentrations of diethyl- p-nitrophenyl phosphate at 37°C. The reactions were per- formed in the presence of known concentrations of ace- tylthiocholine, functioning as a competitive substrate to retard the rate of inhibition of AChE as described in the text. The ordinate represents the fraction of enzyme ac- tivity remaining at time f. Each inhibitor concentration is depicted by a single line, the slope of which equals the respective -Y&,~. The results are those of a typical experi- ment.
and di-n-butyl p-nitrophenyl phosphate be- tween chicken and rat AChE when both were determined at 37°C. All other comparisons between species were significantly different.
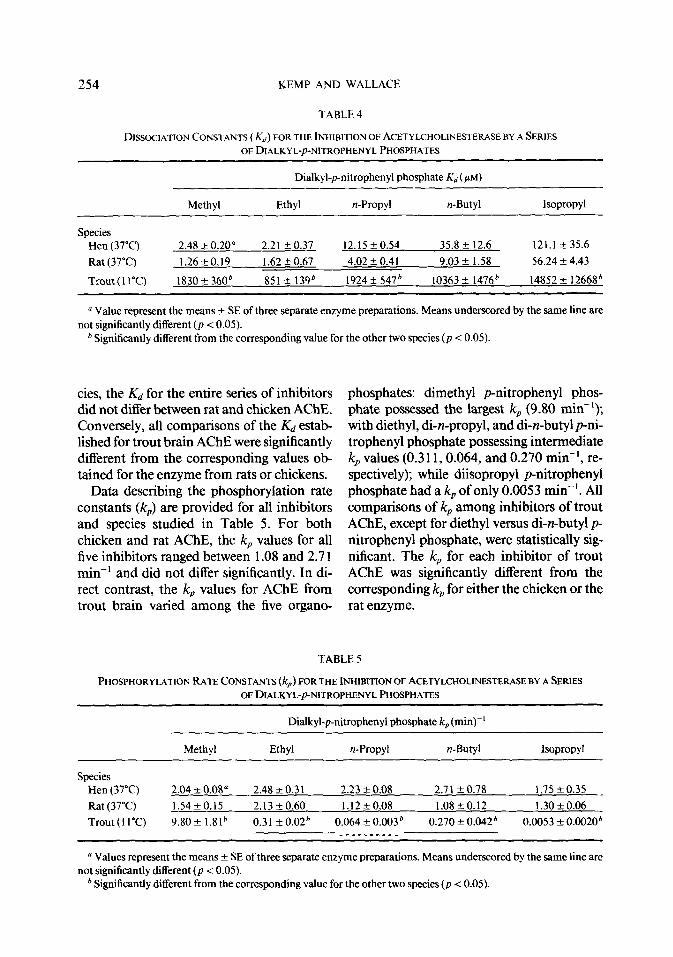
Figures 1 and 2 depict typical kinetic plots for the inhibition of rat brain AChE by di- ethyl p-nitrophenyl phosphate employing the modified kinetic model wherein acetylthio- choline was present during the course of en- zyme inhibition, Each reaction was allowed to proceed until approximately 90% of the enzyme activity was inhibited (Fig. 1). Least- squares regression of the progression plots yielded lines of increasing slope (-k,,,) corre- sponding to increasing inhibitor concentra- tions. In all cases for each species and all in- hibitors, r2 3 0.95. The inhibition constants were derived from double-reciprocal plots of kapp regressed against inhibitor concentration (Fig. 2). Once again, r2 3 0.95 for all cases. The slope of this line is equal to l/ki, inter- secting the ordinate at l/k, and the abscissa at l/Kds. The dissociation constant (Kd) is calculated by dividing K~s by ( 1 + [S]/K&
The bimolecular inhibition constants (k;) for the series of dialkyl p-nitrophenyl phos- phates across species are summarized in Ta- ble 3. Describing the overall reaction between inhibitor and enzyme, kj is a measure of in- hibitor potency and can be related mathe- matically to the IC50 (Johnson and Wallace, 1987). Whereas a low IC50 indicates a potent inhibitor, the inverse is true for k;; the larger the k, the more potent the inhibitor. As with the IC50 data, dimethyl and diethyl p-nitro- phenyl phosphate were the most potent in- hibitors toward chicken and rat brain AChE as reflected by ki. The comparable k, values for these two inhibitors suggest that neither the rat nor the chicken AChE distinguished between dimethyl and diethyl p-nitrophenyl phosphate. However, there was a stepwise de- crease in k, from diethyl through diisopropyl p-nitrophenyl phosphate for these two species of AChE. Di-n-butyl p-nitrophenyl phos- phate was 14-fold less potent than diethyl p-nitrophenyl phosphate toward chicken AChE and 12-fold less potent toward rat brain AChE. Similar to that observed for the IC50 data, diisopropyl p-nitrophenyl phos- phate was the least potent inhibitor toward all species studied, ki being 72- and 63-fold lower
J
2.0 -
1.6 -
.i/
0.4
i/l 0.04 .‘# ! . , . I , ,
-0.2 -0.1 0.0 0.1 0.2 0.3 0.4
1 Ml (PM)
FIG. 2. Double-reciprocal plot for the inhibition of rat brain AChE by diethyl-pnitrophenyl phosphate at 37°C. The values for Knapp were derived from Fig. 1. Least- squares linear regression yielded a line which intersected the ordinate at (&J’ and the abscissa at -(&)-I, where Kdr = KAI + [S]/K,). In this example, [S]/K, was 5 and thus K*= 6%.

INHIBITION OF BRAIN ACETYLCHOLINESTERASE 253
TABLE 3
BIMOLECULARRATECONSTANTS(~~)FORTHEINHIBITIONOFACE~LCHOLINESTERASEBYASERIES OFDIALKYL-p-NITROPHENYLPHOSPHATES
Dialkyl-pnitrophenyl phosphate k, (mM . min)-’
Methyl Ethyl n-Propyl n-Butyl Isopropyl
Species Hen (37’C) 828k34".b 1138+44 18324 79*56 16k3
Rat (37°C) 1238+59' 1463+20 282k12 1242 lib 23&l
Trout ( 11 “C) 5.37 IL 0.08' 0.375 +o.030b 0.042+0.015b 0.027 +O.OOlb 0.0013 + 0.0001~
0 Values represent the means -t SE ofthree separate enzyme preparations. Means underscored by the same line are not significantly different (p < 0.05).
b Significantly different from the corresponding value for the other two species (p < 0.05).
than that for diethyl pnitrophenyl phosphate in chicken and rat, respectively. For rat and chicken AChE, all possible comparisons of ki for the individual inhibitors were signifi- cantly different, with the only exception be- ing dimethyl and diethyl p-nitrophenyl phos- phate.
The kj data indicate that the entire series of dialkyl p-nitrophenyl phosphates were less potent toward the trout AChE than toward the other species (Table 3). Trout brain AChE exhibited the greatest sensitivity to dimethyl pnitrophenyl phosphate with a ki of 5.4 (mM. min))‘. From dimethyl pnitrophenyl phosphate there was a progressive decrease in k; with increasing methylene substitution: the diethyl analog having a ki of 0.38 (mM. min))‘, di-n-propyl and di-n-butyl possessing a k, of 0.042 and 0.027 (mM.min)-‘, respec- tively, while diisopropyl p-nitrophenyl phos- phate had the lowest ki- of 0.0013 (mM. min)-‘. Other than di-n-propyl and di-n-bu- tyl p-nitrophenyl phosphate, the individual ki values for the inhibitors against trout AChE were all statistically different from one an- other. An analysis across species for each in- hibitor showed no significant difference be- tween the ki observed in the chicken and rat for diethyl, di-n-propyl, or diisopropyl p-ni- trophenyl phosphate; all other possible com- parisons for each inhibitor across all three species were statistically different.
Table 4 illustrates the dissociation con- stants (&) for all inhibitors in the three spe- cies studied. Brain AChE from chicken and rat possessed the lowest Kd for dimethyl and diethyl pnitrophenyl phosphate, the values not being statistically different. There was, however, a two- to sixfold increase in the Kd between diethyl and di-n-propyl p-nitrophe- nyl phosphate in the chicken and rat and this trend continued in a stepwise fashion with the Kd increasing with successive methylene substitutions through di-n-butyl pnitrophe- nyl phosphate for both species of AChE. Both the chicken and rat enzyme failed to distin- guish between the Kd for the dimethyl and di- ethyl analogs; similarly, no difference was ob- served when comparing the Kd of chicken AChE for di-n-propyl and di-n-butyl p-nitro- phenyl phosphate. Rat brain AChE also did not distinguish between the Kd values for di- ethyl and di-n-propyl p-nitrophenyl phos- phate or between the di-n-propyl and the di- n-butyl analog. All other comparisons of Kd for the inhibitors of rat or chicken AChE were statistically significant. Rainbow trout brain AChE possessed Kd values which were two to three orders of magnitude larger than the cor- responding Kd for the rat or chicken enzyme (Table 4). However, in contrast to rat and chicken AChE, Kd for the trout enzyme did not vary significantly between the five inhibi- tors examined. When comparing across spe-
254 KEMP AND WALLACE
TABLE 4
DISS~CIATIONCONSTANTS( Kd) FORTHEINHIB~TIONOFACETYLCHOLINESTERASEBYASERIES OFDIALKYL-p-NITROPHENYLPHOSPHATES
Dialkyl-p-nitrophenyl phosphate Kd (PM)
Methyl Ethyl n-Propy! n-Buty! Isopropyl
Species Hen (37°C) 2.48 rt: 0.20” 2.21 2 0.37 12.15 20.54 35.8 -t 12.6 121.1 + 35.6 Rat (37°C) 1.26 +0.19 1.62 t 0.67 4.02 f 0.4 1 9.03 +_ 1.58 56.24 + 4.43
Trout(! 1°C) 1830t_360* 851 +_ t39b 1924 +- 547 b 10363 t 14761~ 14852 t- t2668b
y Value represent the means f SE of three separate enzyme preparations. Means underscored by the same line are not significantly different (p -c 0.05).
b Significantly different from the corresponding value for the other two species (p < 0.05).
ties, the Kd for the entire series of inhibitors did not differ between rat and chicken AChE. Conversely, all comparisons of the & estab- lished for trout brain AChE were significantly different from the corresponding values ob- tained for the enzyme from rats or chickens.
Data describing the phosphorylation rate constants (k,) are provided for all inhibitors and species studied in Table 5. For both chicken and rat AChE, the kp values for all five inhibitors ranged between 1.08 and 2.7 1 min-’ and did not differ significantly. In di- rect contrast, the kp values for AChE from trout brain varied among the five organo-
phosphates: dimethyl p-nitrophenyl phos- phate possessed the largest kp (9.80 min-‘); with diethyl, di-n-propyl, and di-n-butyl p-m- trophenyl phosphate possessing intermediate kp values (0.3 11,0.064, and 0.270 mm-‘, re- spectively); while diisopropyl pnitrophenyl phosphate had a kp of only 0.0053 min-‘. All comparisons of kp among inhibitors of trout AChE, except for diethyl versus di-n-butyl p nitrophenyl phosphate, were statistically sig- nificant. The kp for each inhibitor of trout AChE was significantly different from the corresponding k,, for either the chicken or the rat enzyme.
TABLE 5
PHOSPHORYLATIONRATECONSTANTS (k,) FORTHEINHIBITIONOFACETYLCHOLINESTERASEBYASERIES OFDIALKYL-~NITROPHENYLPHOSPHATES
Dialkyl-p-nitrophenyl phosphate k,, (min)) ’
Species Hen (37°C) Rat (37°C) Trout (I 1°C)
Methyl Ethyl
2.04 + 0.08” 2.48 + 0.31 1.54t0.15 2.13f0.60 9.80 f l.81b 0.3 1 2 0.02b
n-Propy!
2.23 ST 0.08 1.12*0.08
0.064 -+ 0.003 * _----_---_
n-Buty!
2.7 1 + 0.78 1.08 kO.12
0.270 + 0.042’
Isopropyl
1.75 + 0.35 1.30 + 0.06
0.0053 +- 0.00206
LI Values represent the means f SE of three separate enzyme preparations. Means underscored by the same line are not significantly different (p c 0.05).
b Significantly different from the corresponding value for the other two species (p c 0.05).

INHIBITION OF BRAIN ACETYLCHOLlNESTERASE 255
DISCUSSION
The linearity of the kinetic plots is consis- tent both with a first-order process with re- spect to inhibitor concentration and with the existence of a single major isozyme of AChE (Chemnitius, 1982). The preferential hydro- lysis of acetylthiocholine by rat, chicken, and trout brain AChE as well as the relative veloc- ities using propionyl- (60-75% for rats and hens, and 10% for trout) and butyryl- (10% for rats and hens, and <I% for trout) com- pared to acetylthiocholine agree with previ- ous reports (Andersen and Mikalsen, 1978; Hobden and Klaverkamp, 1977). The low K,,, for acetylthiocholine compared to that of the other substrates is consistent with the rat and chicken enzyme being “true” acetylcholines- terases (EC 3.1-l .7). Furthermore, the values of K,,, for acetylthiocholine hydrolysis by rat and trout brain AChE of 78 and 197 PM, re- spectively, compare with those reported by Moss and Famey ( 1978) for rats and goldfish (55 and 250 PM, respectively). The low K, for butyrylthiocholine hydrolysis by chicken AChE is unusual, but not unprecedented. Rotundo (1984) reported a similar relation- ship for substrate Km for chicken brain AChE, where the Km for butyrylthiocholine hydroly- sis was approximately one-third that for ace- tylthiocholine despite a slow reaction veloc- ity. The significance of these observations has yet to be resolved.
The bimolecular inhibition constants for the series of organophosphates complement the IC50 data. Predicted IC50 values were calculated from the corresponding ki value for each of the five inhibitors in each species (Wang and Murphy, 1982a; Johnson and Wallace, 1987). The calculated ~150 values (n = 45) were regressed by least-squares against the observed ~150, yielding a line with slope = 1.040, y-intercept = 0.035, and a regression coefficient (r2) = 0.986 (data not shown). This correlation between the experi- mental ~150 and that calculated from the ki value has been reported previously (Wang and Murphy, 1982a; Johnson and Wallace,
1987) and substantiates the validity of the methods employed to estimate the various in- hibition constants, regardless of the species of enzyme or the specific kinetic model em- ployed.
The substrate and inhibitor kinetic data support the thesis that trout AChE possesses less steric tolerance at the catalytic site than do the enzymes from either hens or rats. This conclusion is based on the observation that the trout enzyme exhibited high dissociation constants and far greater discrimination be- tween the hydrolysis of acetyl- and propionyl- thiocholine as well as far greater inhibitory sensitivity to dimethyl than to diethylpnitro- phenyl phosphate compared to either rat or chicken AChE. In contrast, rat and chicken brain AChE both failed to distinguish be- tween dimethyl and diethyl p-nitrophenyl phosphate based on the Kd data (Table 4). These differences were inherent to the en- zymes studied and not simply a manifesta- tion of the different reaction temperatures. A similar hypothesis regarding species-related differences in steric hindrance at the active site was proposed by Anderscn et al. (1977) to account for the resistance of frog brain AChE, compared to that from rats or chickens, to in- hibition in vitro by various substituted phos- phates. A smaller esteratic site of trout brain AChE would, by excluding bulky molecules, confer a degree of resistance to the enzyme to inhibition by phosphoryl esters and, conse- quently, insensitivity to intoxication by or- ganophosphate insecticides. Similar trends of decreasing inhibitor potency with increasing carbon number were previously reported for homologous series of pyrophosphates and fluorophosphates, and O-substituted diethyl- mercaptosuccinyl phosphinothiolates and p- nitrophenyl phosphates (Aldridge, 1953; Becker et al., 1964; Chiu et al., 1969). The fact that diisopropyl p-nitrophenyl phosphate was a much weaker inhibitor of AChE than was either di-n-propyl or diethyl pnitrophe- nyl phosphate suggests that molecular vol- ume, rather than linear dimension, is the lim- iting factor for the association of the inhibitor

256 KEMP AND WALLACE
with the enzyme for all three species exam- ined.
An alternative interpretation of the in- creasing Kd with successive methylene substi- tutions is a possible role of hydrophobic inter- actions: hydrophobicity increasing with lin- ear carbon number. Consequently, one may conclude that the enzymes either lack a hy- drophobic subsite or possess a hydrophilic subsite and that the relationship among the values of Kd for each enzyme reflects the pro- gressive exclusion of the large hydrophobic inhibitors from an aqueous environment of the active site. However, the octanol/water partition coefficient (calculated theoretically and substantiated by retention on silica HPLC) for diisopropyl p-nitrophenyl phos- phate is virtually identical to that for the di- ethyl analog. In view of the large difference in inhibitor potency for these analogs in all three species, there does not appear to be a direct relationship between enzyme inhibition and hydrophobicity. Consequently, we conclude that although a hydrophobic region may ex- ist, it plays only a minor role in discriminat- ing among the various inhibitors employed in this investigation and is of nominal signifi- cance in accounting for the large differences in inhibitor sensitivity exhibited by AChE from the different species.
The observation that it was the dissociation constant (&) and not the phosphorylation rate constant (k,) which was the primary de- terminant for differentiating between inhibi- tory potencies for both rat and chicken brain AChE is consistent with previous reports for AChE isolated from numerous sources, in- cluding bovine erythrocytes, electric eel, or from monkey, guinea pig, rat, chicken, rab- bit, catfish, or frog brain (Main and Iverson, 1966; Chiu et al., 1969; Andersen et al., 1977; Wang and Murphy, 1982a; For&erg and Putt, 1984; Gray and Dawson, 1987). This suggests that the association of the inhibitor with AChE is the principal factor in distin- guishing among potencies for the homolo- gous series of organophosphates. Once the complex is formed, phosphorylation pro-
ceeds rapidly and independent of the specific inhibitor. Implicit to this interpretation is that a strong nucleophile exists within the ac- tive site of AChE from these species and is the sole determinant of the nucleophilic attack on the phosphoryl group, kp being indepen- dent of the electronic properties of the inhib- itor.
In direct contrast, the inhibition of trout brain AChE by the different inhibitors varied as a function of the k, (Table 5), Kd not being significantly different for individual com- pounds despite a 4000-fold difference in the ki between dimethyl and diisopropyl pnitro- phenyl phosphate. For rat or chicken AChE, the kp values for the various inhibitors were not significantly different, ranging between 1.1 and 2.7 min-‘. For the trout enzyme, k, varied from 9.8 min-’ for dimethyl to 0.005 min-’ for diisopropyl p-nitrophenyl phos- phate (a 2000-fold range). This dependency on k, suggests that, in contrast to rat or chicken AChE, trout brain AChE possesses a fairly weak nucleophile within the esteratic site, the phosphorylation ofthe enzyme being proportional to the acidity of the phosphoryl group of the inhibitor. As the acidity (electro- philicity) of the phosphoryl group diminishes due to the electron-releasing inductive effects of successive methylene substitutions, k, and consequently the inhibitory potency (k;) de- crease. This variation between k, for the vari- ous inhibitors of trout AChE confirms the findings by Jar-v and Aaviksaar (1986) who suggest that the rate of phosphorylation of co- bra venom AChE by nonionic organophos- phates is governed solely by the inductive effects of alkyl substituents on the phosphor-y1 esters. Furthermore, this electronic interac- tion between the phosphoryl group of the in- hibitor and the nucleophilic center of AChE may contribute to the binding constant. If so, the proposed weaker nucleophilic strength of the active site of trout AChE may be partially responsible for the lower affinity (as reflected by 1 /Kd) of the enzyme for the various inhibi- tors compared to that for either chicken or rat brain AChE (Table 4).

INHIBITION OF BRAIN ACETYLCHOLINESTERASE 257
In conclusion, comparison of the kinetics of substrate hydrolysis and enzyme inhibi- tion for membrane-associated brain AChE from rats, chickens, and trout reveal substan- tial differences in catalytic properties. The re- sistance of trout brain AChE to in vitro inhi- bition by dialkyl esters ofpnitrophenyl phos- phate may be a manifestation of the greater steric hindrance, possibly reflecting a smaller active site of this enzyme, coupled with a weak nucleophilic center. As a result, bulky substrates or inhibitors are more effectively excluded by the finite dimensions of the ac- tive site of trout AChE. In addition, phos- phorylation of the trout enzyme is dependent on the acidity of the phosphoryl group of the inhibitor. This contrasts with chicken or rat AChE wherein the principal feature which distinguishes among inhibitor potencies is the association of the inhibitor in the esteratic site, phosphorylation occurring rapidly and independent of the inhibitor. These differ- ences in kinetic mechanism not only provide insight into the molecular variables confer- ring resistance to fish against organophos- phate intoxication, but also suggest possible strategies for improving on the species-selec- tive targeting of anticholinesterase agents. Depending on the catalytic characteristics of AChE from the species of concern, specificity may be improved by selectively modifying ei- ther the steric dimensions or the electronic properties of the prospective inhibitor.
ACKNOWLEDGMENTS
The authors appreciate the expert assistance of Robert M. Carlson and Robert J. Liukkonen in the synthesis and chemical characterization of the organophosphorus compounds employed in this investigation. Susan J. Kurki provided excellent clerical assistance in the prepa- ration of this manuscript. This work was supported in part by a grant-in-aid from the University of Minnesota Graduate School.
REFERENCES
ALDRIDGE, W. N. ( 1953). The differentiation of true and pseudo cholinesterase by organophosphorus com- pounds. Biochemistry 53,62-67.
ANDERSEN, R. A., AARAAS, I., GAARE, G., AND FON- NUM, F. (1977). Inhibition of acetylcholinesterase from different species by organophosphorus com- pounds, carbamates and methylsulphonylfluoride. Gen. Pharrnacol. 8,33 l-334.
ANDERSEN, R. A., AND MIKALSEN, A. (1978). Substrate specificity, effect of inhibitors and electrophoretic mo- bility of brain and serum cholinesterase from frog, chicken and rat. Gen. Pharmacol. 9, 177- 18 1.
BECKER, E. L., PUNTE, C. L., AND BARBERO, J. F. (1964). Acute toxicity of alkyl and phenylalkylphos- phonates in the guinea pig and rabbit in relation to their anticholinesterase activity and their enzymatic inactivation. B&hem. Pharmacol. 13, 1229- 1232.
BENKE, G. M., CHEEVER, K. L., MIRER, F. E., AND MURPHY, S. D. (1974). Comparative toxicity, anticho- linesterase action and metabolism of methyl parathion and parathion in sunfish and mice. Toxicol. Appl. Pharmacol. 28,97-109.
CHATTOPADHYAY, D. P., DIGHE, S. K., NASHIKKAR, A. B., AND DUBE, D. K. (1986). Species differences in the in vitro inhibition of brain acetylcho!inesterase and carboxylesterase by mipafox, paraoxon, and so- man. Pestic. B&hem. Physiol. 26,202-208.
CHEMNITIUS, J.-M., HASELMEYER, K-.H., AND ZECH, R. ( 1982). Identification of isozymes in cholinesterase preparations using kinetic data of organophosphate in- hibition. Anal. Biochem. 125,442-452.
CHIU, Y. C., MAIN, A. R., AND DAUTERMAN, W. C. ( 1969). Affinity and phosphorylation constants of a se- ries of QOdialkyl-malaoxons and paraoxons with acetylcholinesterase. Biochem. Pharmacol. 18, 2 17 l- 2177.
DARLINGTION, W. A., PARTOS, R. D., AND RA-rrs, K. W. (I 97 1). Correlation of cholinesterase inhibition and toxicity in insects and mammals. I. ethylphospho- nates. Toxicol. Appl. Pharmacol. 18,542-547.
DAUTERMAN, W. C., AND O’BRIEN, R. D. (1964). Cho- linesterase variation as a factor in organophosphate se- lectivity in insects. J. Agric. Food Chem. 12,3 18-3 19.
DEROOS, A. M., AND TOET, H. J. (1958). The prepara- tion of some dialkyl-pnitrophenyl phosphates. Re- cueil77,946-955.
ECOBICHON, D. J., AND COMEAU, A. M. (1973). Pseudo- cholinesterases of mammalian plasma: Physicochemi- cal properties and organophosphate inhibition in eleven species. Toxicol. Appl. Pharmacol. 24,92- 100.
ELLMAN, G. L., COURTNEY, K. D., ANDRE& V., JR., AND FEATHERSTONE, R. M. (196 1). A new and rapid calorimetric determination of acetylcholinesterase ac- tivity. Biochem. Pharmacol. 7,88-9X
FISZER, B., AND MICHALSKI, J. (1952). A simplified pro- cedure for preparation of dialkylchlorophosphates. Rocz. Chem. 26,688-689.
FLETCHER, J. H., HAMILTON, J. C., HECHENBLEIKNER, I., HOEGBERG, E. I., SERTYL, B. J., AND CASSADAY, J. T. (1950). The synthesis of parathion and some

258 KEMP AND WALLACE
closely related compounds. J. .dmer. Chem. Sot. 72,
246 I-2464.
FORSBERG, A., AND Puu, G. (1984). Kinetics for the in- hibition of acetylcholinesterase from the electric eel by some organophosphates and carbamates. Eur. J. Bio-
them. 140,153-156.
GRAY, P. .I., AND DAWSON, R. M. (1987). Kinetic con- stants for the inhibition of eel and rabbit brain acetyl- cholinesterase by some organophosphates and carba- mates of military significance. To~icol. Appl. Pharma-
col. 91, 140-144.
HART, G. J., AND O’BRIEN, R. D. (1973). Recording spectrophotometric method for determination of dis- sociation and phosphorylation constants for the inhi- bition of acetylchohnesterase by organophosphates in the presence of substrate. Biochemisrry 12, 2940-
2945.
HOBDEN, B. R., AND KLAVERKAMP, J. F. (1977). Aphar- macological characterization of acetylcholinesterase from rainbow trout (Salmo Gairdneri) brain. Comp.
Biochem. Physiol. C 57, 13 1 -I 33. HODSON, P. V. ( 1985). A comparison of the acute toxic-
ity of chemicals to fish, rats and mice. J. Appl. To.xicol.
5,220-226.
HOLLINGWORTH, R. M., FUKUTO, T. R., AND MET- CALF, R. L. (1967). Selectivity of sumithion compared with methyl parathion. Influence of structure on anti- cholinesterase activity. J. Agric. Food Chem. 15,235- 241.
JARV, J., AND AAVIKSAAR, A. (1986). Leaving group effects in binding and reaction steps of acetylcholines- terase inhibition by o,o-diethylthiophosphates. Bio-
org. Chem. 14,222-227.
JOHNSON, J. A., AND WALLACE, K. B. (1987). Species- related differences in the inhibition of brain acetylcho- linesterase by paraoxon and malaoxon. Toxicol. Appl. Pharmacol. 88,234-241.
LINEWEAVER, H., AND BURK, D. (1934). The determi- nation of enzyme dissociation constants. J. Amer. Chem. Sot. 56,658-666.
Lorry, M., AND JOHNSON, M. K. (1978). Neurotoxicity of organophosphorus pesticides: Predictions can be based on in vitro studies with hen and human en- zymes, Arch. Toxicol. 41,2 15-22 I.
LOWRY, 0. H., ROSEBROUGH, H. J., FARR, A. L., AND RANDALL, R. J. ( 195 I). Protein measurement with the folin phenol reagent. J. Biol. Chem. 193,265-275.
MAIN, A. R., AND IVERSON, F. (1966). Measurement of the affinity and phosphorylation constants governing irreversible inhibition of cholinesterases by di-isopro- pyl phosphofluoridate. Biochem. J. 100,525-53 1.
MCIVOR, R. A., MCCARTHY, G. D., AND GRANT, G. A. (1956). Preparation and toxicity of some alkyl thiopy- rophosphates. Canad. J. Chem. 34,18 19- 1832.
MOSS, D. E., AND FAHRNEY, D. (1978). Kinetic analysis of differences in brain acetylcholinesterase from fish or
mammalian sources. Biochem. Pharmacol. 27,2693- 2698.
MURPHY, S. D. (1966). Liver metabolism of thiophos- phate insecticides in mammalian, avian and piscine species. Proc. Sot. Exp. Biol. Med. 123,392-398.
MURPHY, S. D.. LAUWERYS, R. R., AND CHEEVER, K. L. (I 968). Comparative anticholinesterase action of organophosphorus insecticides in vertebrates. Toxicol.
Appl. Pharmacol. 12,22-35. O’BRIEN, R. D. (1963). Binding of organophosphates to
cholinesterase. J. Agric. Food Chem. 11, 163-166. OTT, P. (1985). Membrane acetylcholinesterases: Puri-
fication, molecular properties and interactions with amphiphilic environments. Biochim. Biophys. Acfa 822,375-392.
POTTER, J. L.. AND O’BRIEN, R. D. (1964). Parathion activation by livers of aquatic and terrestrial verte- brates. Science 144,55-56.
RAKONCZAY, Z., MALLOL, J., SCHENK, H., VINCEN- DON, G., AND ZANETTA, J. P. (198 1). Purification and properties of the membrane-bound acetylcholinester- ase from adult rat brain. Biochem. Biophys. Acta 657, 243-256.
ROTUNDO, R. L. (I 984). Purification and properties of the membrane-bound form of acetylcholinesterase from chicken brain. J. Biol. Chem. 259, 13,186- 13,194.
SCHRADER, G. (1952). Die entwicklung insektizide auf gnmdlage ogranishchler fluor- und phosphor-verbin- dungen. Monogr. Zu Angewandfe Chem. 62,48.
SOKAL, R. R., AND ROHLF, F. J. (198 I). Biometry. Free- man, New York.
STIEGER, S., AND BRODBECK, U. (1985). Amphiphilic detergent-soluble acetylcholinesterase from torpedo marmorata: Characterization and conversion by pro- teolysis to a hydrophilic form. J. Neurochem. 44,48- 56.
VALLETTE, F. M., MARSH, D. J., MULLER, F., MASSOU- LIE. J., MARCOT, B., AND VIEL, C. (1983). Compara- tive affinity chromatography of acetylcholinesterase from five vertebrate species. J. Chromatogr. 257,285- 296.
WALLACE, K. B.. AND DARGAN, J. E. (1987). Intrinsic metabolic clearance of parathion and paraoxon by liv- ers from fish and rodents. Toxicol. Appl. Pharmacol. 90,235-242.
WANG, C.. AND MURPHY, S. D. (1982a). Kinetic analy- sis of species differences in acetylcholinesterase sensi- tivity to organophosphate insecticides. Toxicol. Appl. Pharmacol. 66,409-4 19.
WANG, C., AND MURPHY, S. D. (1982b). The role of non-critical binding proteins in the sensitivity of ace- tylcholinesterase from different species to diisopropyl- flourophosphate (DFP), in vitro. Life Sci. 31, 139- 149.
ZAHAVI, M., TAHORI, A. S., AND KLIMER, F. (197 1). In- sensitivity of acetylchohnesterases to organophospho- rus compounds as related to size of esteratic site. Mol. Pharmucol. 7,6 1 l-6 19.