molecular characterization of conditionally immortalized cell lines derived from mouse early...
TRANSCRIPT

PATTERNS & PHENOTYPES
Molecular Characterization of ConditionallyImmortalized Cell Lines Derived From Mouse EarlyEmbryonic Inner EarJohn A. Germiller,1,2†‡ Elizabeth C. Smiley,1† Amanda D. Ellis,1 Jessica S. Hoff,1 Ian Deshmukh,1
Susan J. Allen,1 and Kate F. Barald1*
Inner ear sensory hair cells (HCs), supporting cells (SCs), and sensory neurons (SNs) are hypothesized to develop fromcommon progenitors in the early embryonic otocyst. Because little is known about the molecular signals that controlthis lineage specification, we derived a model system of early otic development: conditionally immortalized otocyst(IMO) cell lines from the embryonic day 9.5 Immortomouse. This age is the earliest stage at which the otocyst caneasily be separated from surrounding mesenchymal, nervous system, and epithelial cells. At 9.5 days post coitum,there are still pluripotent cells in the otocyst, allowing for the eventual identification of both SN and HCprecursors—and possibly an elusive inner ear stem cell. Cell lines derived from primitive precursor cells can also beused as blank canvases for transfections of genes that can affect lineage decisions as the cells differentiate. It isimportant, therefore, to characterize the “baseline state” of these cell lines in as much detail as possible. Wecharacterized seven representative “precursor-like” IMO cell populations and the uncloned IMO cells, before cellsorting, at the molecular level by polymerase chain reaction (PCR) and immunocytochemistry (IHC), and one line(IMO-2B1) in detail by real-time quantitative PCR and IHC. Many of the phenotypic markers characteristic ofdifferentiated HCs or SCs were detected in IMO-2B1 proliferating cells, as well as during differentiation for up to 30days in culture. These IMO cell lines represent a unique model system for studying early stages of inner eardevelopment and determining the consequences of affecting key molecular events in their differentiation.Developmental Dynamics 231:815–827, 2004. © 2004 Wiley-Liss, Inc.
Key words: gene expression; hair cells; supporting cells; cell line models; auditory system development
Received 31 March 2004; Revised 21 July 2004; Accepted 21 July 2004
INTRODUCTION
Early Stages of OticDevelopment
The development of the vertebrateinner ear is a complex, highly orches-trated process. The mouse andchick epithelial otic placodes invag-inate to form the otocyst (otic vesi-cle), which appears to be a simple
epithelial sac. This structure will even-tually give rise to the complex mem-branous labyrinth with its enclosedsensory organs. However, even fromearly times, day 9 post coitum in themouse, the Hamburger and Hamil-ton (HH) stage 12 (Hamburger andHamilton, 1951) otic vesicle in thechick and the prim5 (24 hr) stage inthe cavitating (rather than invagi-
nating) zebrafish otocyst, the verte-brate otocyst contains cells that arecommitted to specific inner ear cellfates (reviewed in Barald and Kelley,2004). At these early stages, theunique environment surrounding theotocyst influences its development.Signals from the hindbrain, the undif-ferentiated mesenchyme surround-ing the otocyst (periotic mesen-
1Department of Cell and Developmental Biology, Program in Cell and Molecular Biology, Program in Neuroscience, University of Michigan,Ann Arbor, Michigan2Department of Otolaryngology-Head and Neck Surgery, University of Michigan, Ann Arbor, MichiganGrant sponsor: the NIH; Grant number: RO1DC04184; Grant number: R01 DC05939; Grant sponsor: NSF; Grant number: IBN9906424; Grantsponsor: the Deafness Research Foundation; Grant sponsor: the American Academy of Otolaryngology-HNS.†J.A. Germiller and E.C. Smiley contributed equally to this work.‡Dr. Germiller’s present address is Division of Otolaryngology, Children’s Hospital of Philadelphia, Philadelphia, PA.*Correspondence to: Kate F. Barald, Department of Cell and Developmental Biology, University of Michigan, 1150 W. Medical CenterDrive, Ann Arbor, MI 48109-0616. E-mail: [email protected]
DOI 10.1002/dvdy.20186Published online 29 October 2004 in Wiley InterScience (www.interscience.wiley.com).
DEVELOPMENTAL DYNAMICS 231:815–827, 2004
© 2004 Wiley-Liss, Inc.

chyme/POM), the notochord, andthe neural crest (NC) provide induc-tive and patterning information nec-essary for normal inner ear develop-ment (Fritzsch et al., 1997; Fritzschand Beisel, 2001; Fekete and Wu,2002; Liu et al., 2003b; Barald andKelley, 2004). By day 2 of chick em-bryonic development (embryonicday [E] 2; HH stage 13), genes impor-tant for otic development are re-gionalized, with well-defined bor-ders. Parallels to this organizationalso exist in mouse, zebrafish, andXenopus (Fekete, 1996, 2000a,b;Brigande et al., 2000a; Lang et al.,2000; Fekete and Wu, 2002; Baraldand Kelley, 2004).
Cell Specification andDifferentiation
Cells in the epithelium of the oticvesicle give rise to specialized sen-sory structures, among other celltypes. Hair cells (HCs) develop withinthe cochlear duct and in other innerear sensory organs, including vestib-ular organs. Primary sensory neurons(SNs) of the statoacoustic ganglion(SAG), which later contribute to cra-nial nerve VIII, delaminate as precur-sors from the otic crest (Hemondand Morest, 1991). It is still unclearwhether HCs, supporting cells (SCs),and SNs are derived from a com-mon precursor cell (Fritzsch andBeisel, 2001; Barald and Kelley,2004). Viral tracing studies did notfind any progeny of labeled cellsthat gave rise to both a HC and aSN, but the authors (Lang andFekete, 2001) also could not rule outthis possibility in their experiments.
Sensory epithelia are intricatelypatterned so that each HC is sepa-rated from other HCs by interveningSCs that form an invariant and alter-nating mosaic throughout sensoryorgans, including the cochlear duct(Echteler et al., 1994; Barald andKelley, 2004). This alternating patternis postulated to result from the pro-cess of lateral inhibition (Adam etal., 1998; Lanford et al., 1999), inwhich some cells within a collectiveassume a certain fate, and then in-hibit their neighbors from doing thesame (Lewis, 1996; Kimble and Simp-son, 1997; Kopan and Cagan, 1997;Lanford et al., 1999). Recent work
has implicated the Notch signalingparadigm in molecular control of thisprocess. In the developing cochlea,Notch-1 and its ligand jagged-2 areexpressed in alternating cell types,starting at E14.5 in the basal turn(Lanford et al., 1999), with jagged-2restricted to presumptive HCs.
Markers of HC and SC
During later developmental stages,the expression of several genes be-comes restricted to sensory organs,and then specifically to HCs and/orSCs; these genes have been used ascell phenotype markers in the innerear. One of the earliest-expressedgenes specifically in HCs is murineatonal homolog-1 (Math-1), a basichelix-loop-helix transcription factor,which first appears in the sensory epi-thelium of the utricle, saccule, semicir-cular canals, and cochlea, and be-comes restricted to HCs by the end ofgestation (Bermingham et al., 1999;Kawamoto et al., 2003). Another early“HC-specific” gene product is Brn3.1(POU4F3, Brn3c), a POU family tran-scription factor essential for HC differ-entiation and survival (Erkman et al.,1996; Ryan, 1997; Xiang et al., 1997).Mutations in Brn3.1 (DFNA15) wereshown to cause familial adult onsetprogressive hearing loss in humans(Vahava et al., 1998). Later, HCs ex-press unique �9 and �10 subunits ofthe nicotinic acetylcholine receptor(�9 and �10 AChRs; Elgoyhen et al.,1994, 2001; Glowatski et al., 1995). Sev-eral other genes are expressed exclu-sively by HCs, including myosin VIIA(Hasson et al., 1997), parvalbumin 3(PV3, Heller et al., 2002), and theNotch ligand jagged-2. Several geneshave been identified that are charac-teristic of SCs and absent from HCs.These genes include the cytokeratins,which are nonspecific epithelial mark-ers; the �- and �-tectorins (Legan etal., 1997); jagged-1; and the transcrip-tion factors HES-1 and HES-5. The lastthree genes mediate Notch signalingin presumptive SCs in early cochlearpatterning (Weir et al., 2000). Zicgenes have been postulated to af-fect the fate of precursor cells com-mon to neurons or sensory HCs by af-fecting expression of downstreamatonal class genes, which may them-selves be regulated by bone morpho-
genetic proteins (BMPs) and their an-tagonists (Warner et al., 2003; Baraldand Kelley, 2004).
BMP Signaling System
We are beginning to learn moreabout the molecular factors that reg-ulate inner ear development (re-viewed in Barald and Kelley, 2004).Recent evidence points to the BMP4signaling system as being of criticalimportance for inner ear develop-ment (Hogan et al., 1994; Wall andHogan, 1994). In the developing ear,BMP4 is one of the earliest genes andis expressed as early as the otic pla-code stage. It then becomes local-ized to distinct regions of the develop-ing otocyst (Hemmati-Brivanlou andThomsen, 1995; Oh et al., 1996; Morsliet al., 1998; Gerlach et al., 2000; Coleet al., 2000) and later to specific re-gions of the inner ear destined to pro-duce sensory tissue (Morsli et al., 1998;Barald and Kelley, 2004).
BMP4 signaling during develop-ment is modulated by the action ofseveral endogenous antagoniststhat block BMP4 downstream signal-ing. Three of these, noggin, DAN,and chordin, play a critical modula-tory role in BMP signaling during de-velopment of many organ systems(reviewed in Thomsen, 1997 andBarald and Kelley, 2004). Noggin isexpressed in the POM in the earlystage chick embryo (Gerlach et al.,2000). Furthermore, antagonism ofBMP4 by exogenously applied an-tagonists results in profound, specificmalformations of the inner ear, par-ticularly of the semicircular canals(SCC; Chang et al., 1999; Gerlach etal., 2000). DAN is expressed in theearly developing chick medial oticepithelium and in the mouse POM(Gerlach-Bank et al., 2002). Bothgain- and loss-of-function experi-ments show that DAN has a role inED/ES development and may helppartition the epithelium into theED/ES and the anterior and posteriorSCC (Gerlach-Bank et al., 2004).
Immortalized Otocyst CellLines as Models of Early OticDevelopment
Historically, molecular studies of thedeveloping and mature inner ear
816 GERMILLER ET AL.

have been hampered by difficultaccess to the temporal bone, thesmall size of the developing sensoryorgans, and in particular the ab-sence of good organ and cell cul-ture models of the early inner ear.Therefore, our laboratory (Barald etal., 1997) and other researchers (Hol-ley et al., 1997; Kalinec et al., 1999)developed conditionally immortal-ized cell lines from the mouse innerear. Our cell lines were derived fromthe earliest stage that otocysts couldbe excised from 9- to 9.5-day em-bryos. We used the H2KbtsA58transgenic mouse (Immortomouse),which carries a temperature-sensi-tive variant of the SV40 large T anti-gen under the control of a �-interfer-on–sensitive promoter (Jat et al.,1991; reviewed in Noble, 1999). Im-mortomouse otocyst (conditionallyimmortalized otocyst, IMO) cells pro-liferate in the presence of �-inter-feron at 32–33°C, the permissivetemperature for transgene expres-sion, but stop proliferating and differ-entiate after temperature shift to37°–39°C and withdrawal of �-inter-feron. Because the native otocystharbors progenitors of nearly all cellsin the inner ear, our hypothesis is thatimmortalized cells should, given theright culture conditions, eventuallydemonstrate characteristics of mul-tiple divergent sensory and nonsen-sory phenotypes as they differenti-ate. Initially, they would havecharacteristics of “common precur-sor” or multipotent cells. By varyingthe culture conditions, includingcell–cell interactions and/or trans-fecting genes hypothesized to pre-dispose these cells into specific lin-eages, we can use this model todetermine which genes are impor-tant for the development of identi-fied cell types.
A major aim of auditory system re-search is to discover the fundamen-tal differences that allow chicks toregenerate a sensory epithelium,whereas mammals cannot (Stoneand Rubel, 2000a,b). One notabledifference between chicks andmice is that mature HCs of chicksexpress BMP4, although SCs do not.The reverse is true in the maturemouse ear, where SCs but not HCsexpress BMP4. We hypothesize that,in both the chick and the mouse, the
common precursor cell (Brigande etal., 2000b) expresses BMP4 (Baraldand Kelley, 2004). We therefore tar-geted mouse IMO cells that expressBMP4 for initial characterization, be-cause they are excellent candi-dates for common precursor cellsand perhaps precursors to SN aswell. We analyzed the expression ofa panel of markers associated withdeveloping and mature HC, SC, andnonsensory inner ear epithelia in se-lected BMP4-expressing IMO celllines and in the uncloned, unsortedImmortomouse otocyst (UIMO), iso-lated from the original E9.5 mouseotocysts. The clonal cell lines werederived by fluorescence-activatedcell sorting (FACS)/light scatteringcloning (Barald et al., 1997).
We also tested the hypothesis thatspatially restricted cues such as cell–cell interactions in culture influencephenotype development on a locallevel, as is true in the living ear(Barald and Kelley, 2004). We havestudied expression of genes associ-ated with BMP4 and Notch signaling,including BMP4 and related growthfactors, BMP receptors, three BMPantagonists (noggin, chordin, andDAN), Notch-1, and its ligandjagged-2. We have also examinedthe expression of Zic genes, whichwe hypothesize are downstream ef-fectors of BMP4/chordin antagonis-tic cascades and potentially impor-tant for neuronal or HC specification(Warner et al., 2003; Barald andKelley, 2004). One of the cell lines,IMO-2B1, which we hypothesize is agood model for an early “precursorcell,” has been characterized in de-tail in this study by real-time quanti-tative polymerase chain reaction(PCR) with a panel of 50 primer pairscharacteristic of HCs, SCs, SNs, andpresumptive precursor cells. We wishto describe in as much detail as pos-sible the baseline state of these cellsbefore determining effects of spe-cific gene transfections, cell–cell in-teractions, or culture conditions. Wehave already successfully used thiscell line to identify a novel BMP4 pro-moter in intron 2, which our subse-quent studies demonstrated is alsoexpressed in the inner ears of bothchicks and mice (Thompson et al.,2003).
RESULTS AND DISCUSSION
Gene Expression Profiling byPCR
A PCR analysis was used to measureexpression for a panel of genes im-portant in inner ear development.These fell into four categories: (1)genes involved in BMP signaling; (2)other signaling genes, includingthose involved in Notch signaling; (3)markers for the differentiated HCs;and (4) SC phenotypes. Equal quan-tities of RNA were used for all genestested to provide a sense of relativeabundance of the various mRNAs.The E15 mouse embryo was used asa control for primer efficiency, notstaging of the inner ear. Seven differ-ent BMP4-expressing cell lines andthe UIMO cells were screened forgene expression by conventionalPCR at 0 (proliferative) and 15 daysof differentiation; IMO-2B1 cells weretested more frequently (0, 3, 7, 15,and 30 days of differentiating condi-tions; Table 1; Fig. 1).
In all of the IMO cell lines tested(Table 1), BMP4 and its receptorswere detected at very high levelsduring proliferation and levels wereunchanged after differentiation.Chordin and noggin were detectedin many of the cell lines, but expres-sion levels were variable. BMP signal-ing genes included secreted growthfactors of the BMP/transforminggrowth factor-� superfamily (BMPs 2,4, and 6, and activins �A and �B),the type 1A and type 2 BMP4 recep-tors, and three secreted antagonistsof BMP4: noggin, chordin, and DAN(Gerlach-Bank et al., 2004).
An expression profile was first es-tablished over time in UIMO cells,which represent a random cell sam-ple of the otocyst as a whole. A rep-resentative cell line, IMO-2B1 (Fig. 1)is one of the cell lines cloned fromthe UIMO by light scattering on aFACS (Barald et al., 1997). High levelsof expression of BMP4, the activins,and the BMP4 receptor mRNAs werefound in both UIMO and IMO-2B1cells, with expression remaining highthroughout differentiation (Fig. 1).The same pattern was found forchordin and DAN; noggin was foundin UIMO cells after differentiationand in both proliferating and differ-
MOUSE INNER EAR CELL LINES 817
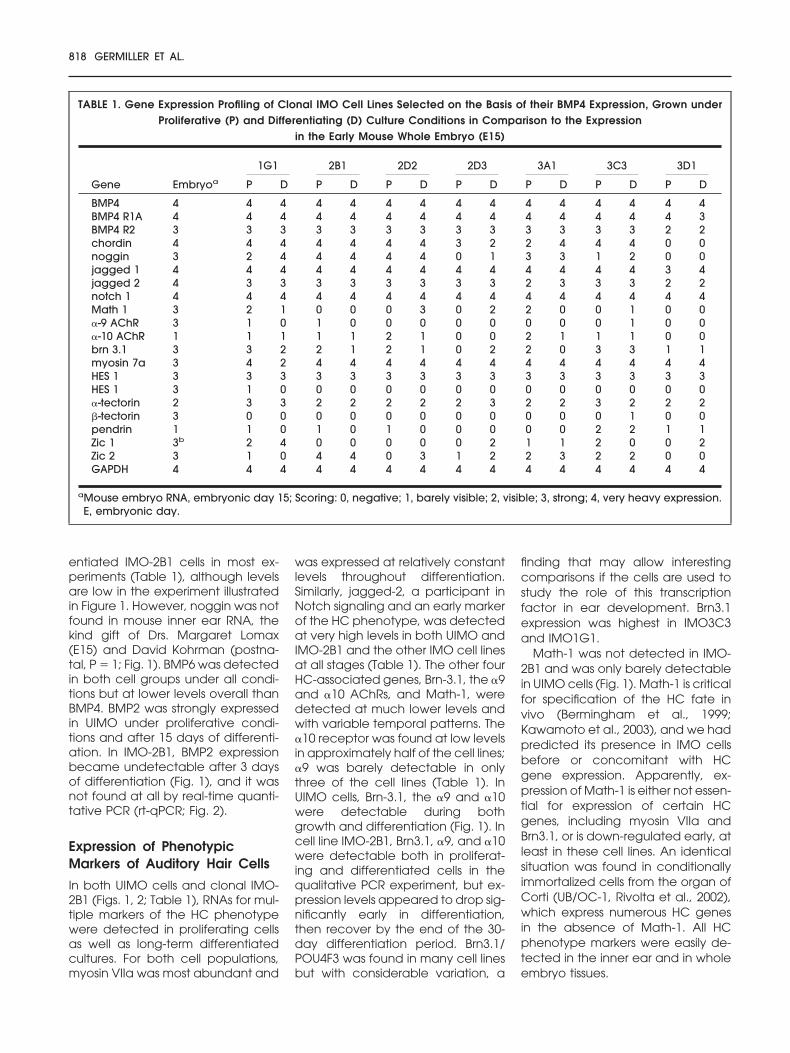
entiated IMO-2B1 cells in most ex-periments (Table 1), although levelsare low in the experiment illustratedin Figure 1. However, noggin was notfound in mouse inner ear RNA, thekind gift of Drs. Margaret Lomax(E15) and David Kohrman (postna-tal, P � 1; Fig. 1). BMP6 was detectedin both cell groups under all condi-tions but at lower levels overall thanBMP4. BMP2 was strongly expressedin UIMO under proliferative condi-tions and after 15 days of differenti-ation. In IMO-2B1, BMP2 expressionbecame undetectable after 3 daysof differentiation (Fig. 1), and it wasnot found at all by real-time quanti-tative PCR (rt-qPCR; Fig. 2).
Expression of PhenotypicMarkers of Auditory Hair Cells
In both UIMO cells and clonal IMO-2B1 (Figs. 1, 2; Table 1), RNAs for mul-tiple markers of the HC phenotypewere detected in proliferating cellsas well as long-term differentiatedcultures. For both cell populations,myosin VIIa was most abundant and
was expressed at relatively constantlevels throughout differentiation.Similarly, jagged-2, a participant inNotch signaling and an early markerof the HC phenotype, was detectedat very high levels in both UIMO andIMO-2B1 and the other IMO cell linesat all stages (Table 1). The other fourHC-associated genes, Brn-3.1, the �9and �10 AChRs, and Math-1, weredetected at much lower levels andwith variable temporal patterns. The�10 receptor was found at low levelsin approximately half of the cell lines;�9 was barely detectable in onlythree of the cell lines (Table 1). InUIMO cells, Brn-3.1, the �9 and �10were detectable during bothgrowth and differentiation (Fig. 1). Incell line IMO-2B1, Brn3.1, �9, and �10were detectable both in proliferat-ing and differentiated cells in thequalitative PCR experiment, but ex-pression levels appeared to drop sig-nificantly early in differentiation,then recover by the end of the 30-day differentiation period. Brn3.1/POU4F3 was found in many cell linesbut with considerable variation, a
finding that may allow interestingcomparisons if the cells are used tostudy the role of this transcriptionfactor in ear development. Brn3.1expression was highest in IMO3C3and IMO1G1.
Math-1 was not detected in IMO-2B1 and was only barely detectablein UIMO cells (Fig. 1). Math-1 is criticalfor specification of the HC fate invivo (Bermingham et al., 1999;Kawamoto et al., 2003), and we hadpredicted its presence in IMO cellsbefore or concomitant with HCgene expression. Apparently, ex-pression of Math-1 is either not essen-tial for expression of certain HCgenes, including myosin VIIa andBrn3.1, or is down-regulated early, atleast in these cell lines. An identicalsituation was found in conditionallyimmortalized cells from the organ ofCorti (UB/OC-1, Rivolta et al., 2002),which express numerous HC genesin the absence of Math-1. All HCphenotype markers were easily de-tected in the inner ear and in wholeembryo tissues.
TABLE 1. Gene Expression Profiling of Clonal IMO Cell Lines Selected on the Basis of their BMP4 Expression, Grown underProliferative (P) and Differentiating (D) Culture Conditions in Comparison to the Expression
in the Early Mouse Whole Embryo (E15)
Gene Embryoa
1G1 2B1 2D2 2D3 3A1 3C3 3D1
P D P D P D P D P D P D P D
BMP4 4 4 4 4 4 4 4 4 4 4 4 4 4 4 4BMP4 R1A 4 4 4 4 4 4 4 4 4 4 4 4 4 4 3BMP4 R2 3 3 3 3 3 3 3 3 3 3 3 3 3 2 2chordin 4 4 4 4 4 4 4 3 2 2 4 4 4 0 0noggin 3 2 4 4 4 4 4 0 1 3 3 1 2 0 0jagged 1 4 4 4 4 4 4 4 4 4 4 4 4 4 3 4jagged 2 4 3 3 3 3 3 3 3 3 2 3 3 3 2 2notch 1 4 4 4 4 4 4 4 4 4 4 4 4 4 4 4Math 1 3 2 1 0 0 0 3 0 2 2 0 0 1 0 0�-9 AChR 3 1 0 1 0 0 0 0 0 0 0 0 1 0 0�-10 AChR 1 1 1 1 1 2 1 0 0 2 1 1 1 0 0brn 3.1 3 3 2 2 1 2 1 0 2 2 0 3 3 1 1myosin 7a 3 4 2 4 4 4 4 4 4 4 4 4 4 4 4HES 1 3 3 3 3 3 3 3 3 3 3 3 3 3 3 3HES 1 3 1 0 0 0 0 0 0 0 0 0 0 0 0 0�-tectorin 2 3 3 2 2 2 2 2 3 2 2 3 2 2 2�-tectorin 3 0 0 0 0 0 0 0 0 0 0 0 1 0 0pendrin 1 1 0 1 0 1 0 0 0 0 0 2 2 1 1Zic 1 3b 2 4 0 0 0 0 0 2 1 1 2 0 0 2Zic 2 3 1 0 4 4 0 3 1 2 2 3 2 2 0 0GAPDH 4 4 4 4 4 4 4 4 4 4 4 4 4 4 4
aMouse embryo RNA, embryonic day 15; Scoring: 0, negative; 1, barely visible; 2, visible; 3, strong; 4, very heavy expression.E, embryonic day.
818 GERMILLER ET AL.
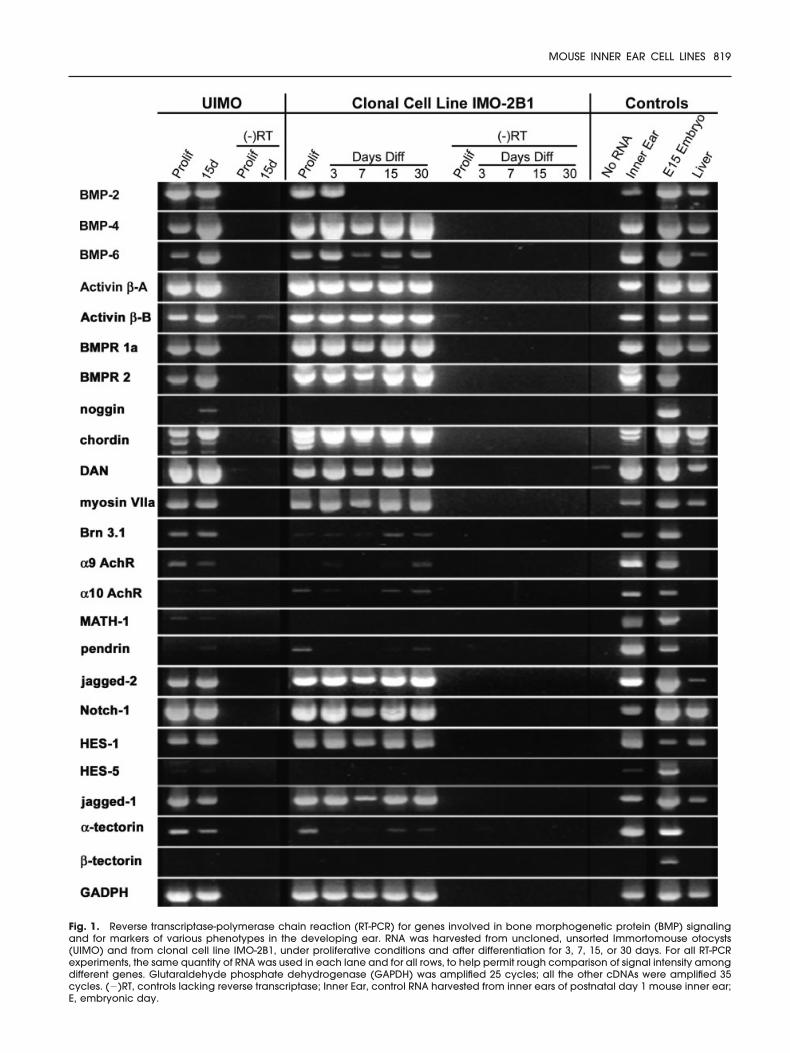
Fig. 1. Reverse transcriptase-polymerase chain reaction (RT-PCR) for genes involved in bone morphogenetic protein (BMP) signalingand for markers of various phenotypes in the developing ear. RNA was harvested from uncloned, unsorted Immortomouse otocysts(UIMO) and from clonal cell line IMO-2B1, under proliferative conditions and after differentiation for 3, 7, 15, or 30 days. For all RT-PCRexperiments, the same quantity of RNA was used in each lane and for all rows, to help permit rough comparison of signal intensity amongdifferent genes. Glutaraldehyde phosphate dehydrogenase (GAPDH) was amplified 25 cycles; all the other cDNAs were amplified 35cycles. (�)RT, controls lacking reverse transcriptase; Inner Ear, control RNA harvested from inner ears of postnatal day 1 mouse inner ear;E, embryonic day.
MOUSE INNER EAR CELL LINES 819
Expression of PhenotypicMarkers of Supporting Cellsand Nonsensory Epithelia
Expression of pendrin and manyother markers of the SC phenotype
were detected in both UIMO cellsand proliferative clonal IMO-2B1(Fig. 1; Table 1). In IMO-2B1, expres-sion of �-tectorin and pendrin fol-lowed a bimodal temporal patternsimilar to that of some of the HC
genes described above, with ex-pression dropping during the earlyphases of differentiation, then re-covering by 30 days. In UIMO, �-tec-torin was easily detected at bothstages, but pendrin only appeared
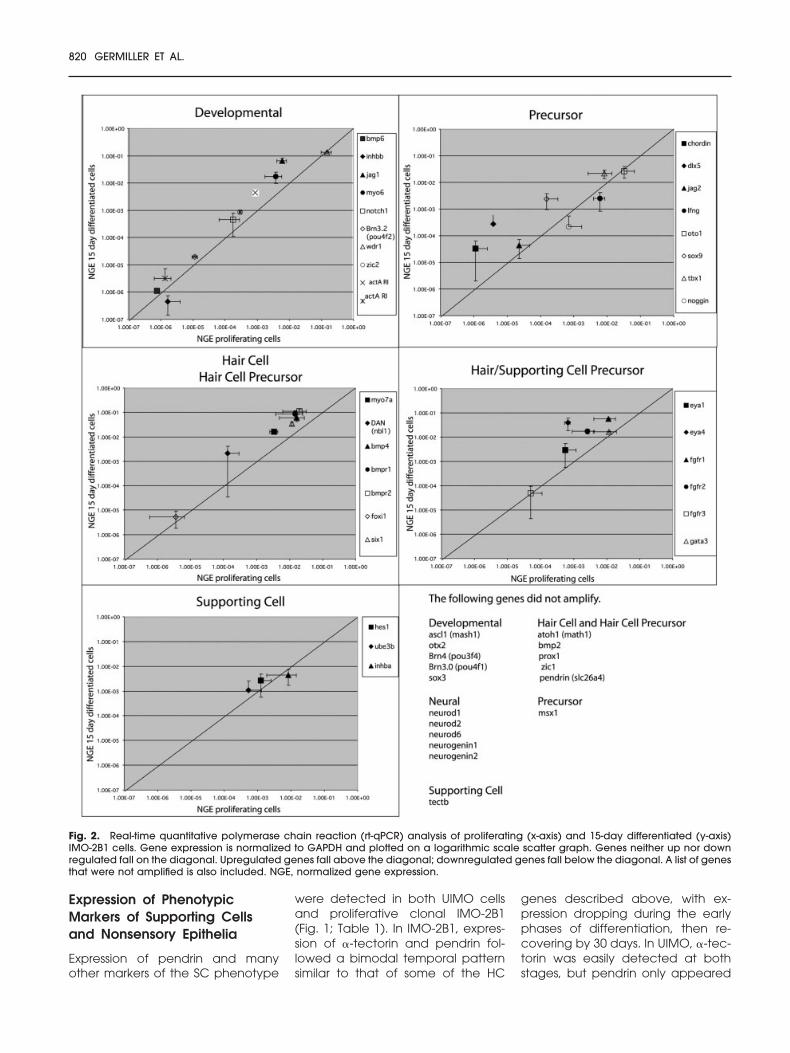
Fig. 2. Real-time quantitative polymerase chain reaction (rt-qPCR) analysis of proliferating (x-axis) and 15-day differentiated (y-axis)IMO-2B1 cells. Gene expression is normalized to GAPDH and plotted on a logarithmic scale scatter graph. Genes neither up nor downregulated fall on the diagonal. Upregulated genes fall above the diagonal; downregulated genes fall below the diagonal. A list of genesthat were not amplified is also included. NGE, normalized gene expression.
820 GERMILLER ET AL.

in the differentiated cells. At leastone cell line (IMO3C3) expressesenough pendrin to make it poten-tially useful for study of this gene,which is implicated in Pendred syn-drome (Table 1). �-tectorin was notdetected in either cell population byconventional PCR.
The cell lines were uniformly posi-tive for HES-1, Notch-1, andjagged-1 at relatively high levels;Notch-1 was particularly abundant.�-tectorin, a component of the tec-torial membrane, was detected atmoderate levels in all lines both be-fore and after differentiation. Instrong contrast is the finding that twoother supporting cell markers, �-tec-torin and HES-5, were not detect-able in UIMO or IMO-2B1 cells byconventional PCR. The finding of di-vergent gene expression profiles inIMO cell lines supports the idea thatphenotypic gene regulation may in-deed occur in IMO cells, i.e., theyare not simply expressing all theirgenes indiscriminately.
Expression of Zic Genes, WhichMay Be Involved in Cell FateDecisions
Expression of the zinc-finger-of-the-cerebellum gene family membersZic1 and Zic2 (Warner et al., 2003)were measured during the screen-ing of the seven clonal IMO cell lines(Table 1). Many cell lines expressedone or both Zic genes. Zic2 was par-ticularly abundant in IMO-2B1. Zic2expression increased significantlywith differentiation in three cell lines:1G1, 2D2, and 3D1. Whereas mostcell lines expressed Zic2, very few ex-pressed Zic1. The notable exceptionis IMO1G1, which has significant lev-els of Zic1 mRNA in both proliferativeand differentiated cultures. We hy-pothesize that the Zic genes are partof a signaling cascade that is down-stream of BMP4 and its molecularantagonist chordin (Elms et al., 2003)and that up-regulation of Zic1 in aprecursor cell common to both neu-rons and HCs in turn up-regulates theatonal gene neuroD1, leading to aneuronal fate. In contrast, we hy-pothesize that up-regulation of Zic2leads to up-regulation of a differentatonal class gene, Math1, resultingin a HC fate (Barald and Kelley,
2004). These cell lines provide anideal system to test and model thesemolecular regulatory cascades.
rt-qPCR
Because of the relatively high cyclenumbers (35), the qualitative PCR as-say could only be used to determinewhether a cell line expressed or didnot express the cDNA. Only rt-qPCRresults, using the methods of Stank-ovic and Corfas (2003), are quanti-tative. We compared these twomethods on the IMO-2B1 cell line(Table 1; Figs. 1, 2).
We performed rt-qPCR on bothproliferating and 15-day differentiat-ing IMO-2B1 cells using 50 primerpairs (Fig. 2). As with the conven-tional PCR, we found that Math-1,Zic1, and �-tectorin were not ex-pressed. We found that the followinggenes also were not expressed:Mash-1, BMP2, neurod1, neurod2and neurod6, neurogenin 1 and 2,otx2, msx1, POU3F4 (Brn4) andPOU4F1 (Brn3a), Prox1, and pendrin(slc26a4). Genes that were down-regulated slightly include inhibin b(4x), noggin (3x), and lunatic fringe(3x). Several genes were slightly up-regulated (Fig. 2). Genes up-regu-lated more than 10-fold includedDAN (nbl1)(16x), chordin (20x), sox9(15x), and jag 1 (11x). Dlx5, whichappears to be a key player in earlyinner ear induction (Liu et al., 2003b),was up-regulated more than 72-fold,and Eya4 was up-regulated 60-fold(Fig. 2; Barald and Kelley, 2004).Primers for Brn3.1, HES-5, and the �9and �10 AChRs are still under devel-opment.
Most of the Cell Lines Clonedto Date Express BMP4 andNotch/Delta Signaling Genes
Early in our analysis, it became clearthat most of the IMO cell lines wehad cloned from the very early oto-cyst (embryonic day 9.5) expressedvery high levels of BMP signalinggenes under both proliferating anddifferentiated conditions (Table 1;Figs. 1, 3). To date, we have foundonly 6 clonal lines of the 27 initiallyscreened that either do not expressBMP4 or express it at very low levels.The characterization of these BMP-
negative cell lines is in progress withrt-qPCR. BMP4 appears to be se-creted in active form in consider-able quantity, as we showed previ-ously with the cell line IMO3C6(Gerlach et al., 2000). Implantationof beads carrying cells that makethe BMP4 antagonist noggin into thedeveloping inner ear caused abnor-mal development of semicircularcanals (Gerlach et al., 2000). How-ever, coimplantation of the IMO3C6cell line, which produces largeamounts of BMP4, rescued the nog-gin-induced canal loss. Many IMOcells also express high levels of BMPreceptors and BMP antagonists inaddition to the growth factors them-selves. These cells should provideideal model systems for further studyof this very complex signaling sys-tem, with its multiple interactionsamong similar growth factors, re-ceptors, binding proteins, and feed-back loops.
IMO cells also appear to express,simultaneously, very high levels ofgene products involved in Notch sig-naling, including Notch-1 and its li-gand jagged-2. This, combined withtheir ability to express genes associ-ated with both HC and non-HC phe-notypes, makes them potentiallyuseful models for study of pheno-type specification by means of thissignaling paradigm.
Expression of Genes Found inMature HCs
The gene expression profiling exper-iments highlight two interesting andconsistent properties of these condi-tionally immortalized cells. First, de-spite their origins from the primitiveotocyst, many cell lines expressgenes that, in vivo, are typically as-sociated with more mature celltypes, such as the myosins, tectorins,and AChRs. Moreover, they do soeven when proliferating, i.e., underconditions permissive for expressionof the immortalizing transgene.Clearly, it would be too simplistic tothink of the proliferative IMO cells asbeing suspended in a primitive oto-cyst-like state under the culture con-ditions used here. However, it wouldbe equally simplistic to posit the op-posite, i.e., that proliferative cellshave already fully differentiated, be-
MOUSE INNER EAR CELL LINES 821
cause they continue to express highlevels of “early” inner ear genes,such as BMP4. Second, as a generalrule, we found little variation in ex-pression levels for most genes whenthe cells were switched to differenti-ating (nonpermissive) conditions, al-though there were exceptions. In fu-ture applications in inner earresearch, abundant gene expres-sion in proliferating cells would bedesirable, such as in dissection offactors regulating their transcription,because it is more convenient towork with stable proliferative cells.However, demonstration of changesin gene expression associated withcommitment to certain cell fates, willlikely require additional exogenoussignals, because the cells do notdemonstrate such shifts under oursimple “default” culture conditions.
Our otocyst-derived cells are notunique among inner ear cell lines intheir propensity to express maturephenotypes before being inducedto differentiate. Conditionally im-mortalized organ of Corti cells UB/OC-2 express many hair cell markersunder proliferative conditions, in-cluding myosin VIIa, �9 AChR, andBrn3.1, and continue to do so afterdifferentiation (Rivolta et al., 1998),whereas another cell line, UB/OC-1,by contrast, up-regulates the samegenes. Similarly, another HC maker,myosin VI, is strongly expressed byboth cell lines in proliferative condi-tions and is unchanged after differ-entiation (Rivolta et al., 1998). Al-though these findings parallel ourown results regarding expression ofsome HC genes, they may also re-flect the later stage at which thecells were derived (E13). Also inter-esting are findings from another Im-mortomouse cell line, UB/UE-1, de-rived from P2 utricle (Lawlor et al.,1999). These postnatal supportingcells were shown to acquire charac-teristics of HCs after differentiation.However, expression of several HC-specific genes was also found be-fore differentiation, including Brn3.1and myosin6, although others wereundetected (myosin VIIa, �9AChR).These results and our own remind usthat it should not be assumed thatthe gene expression profile of im-mortalized inner ear cells is identicalto that of their cells of origin.
Fig. 3.
822 GERMILLER ET AL.
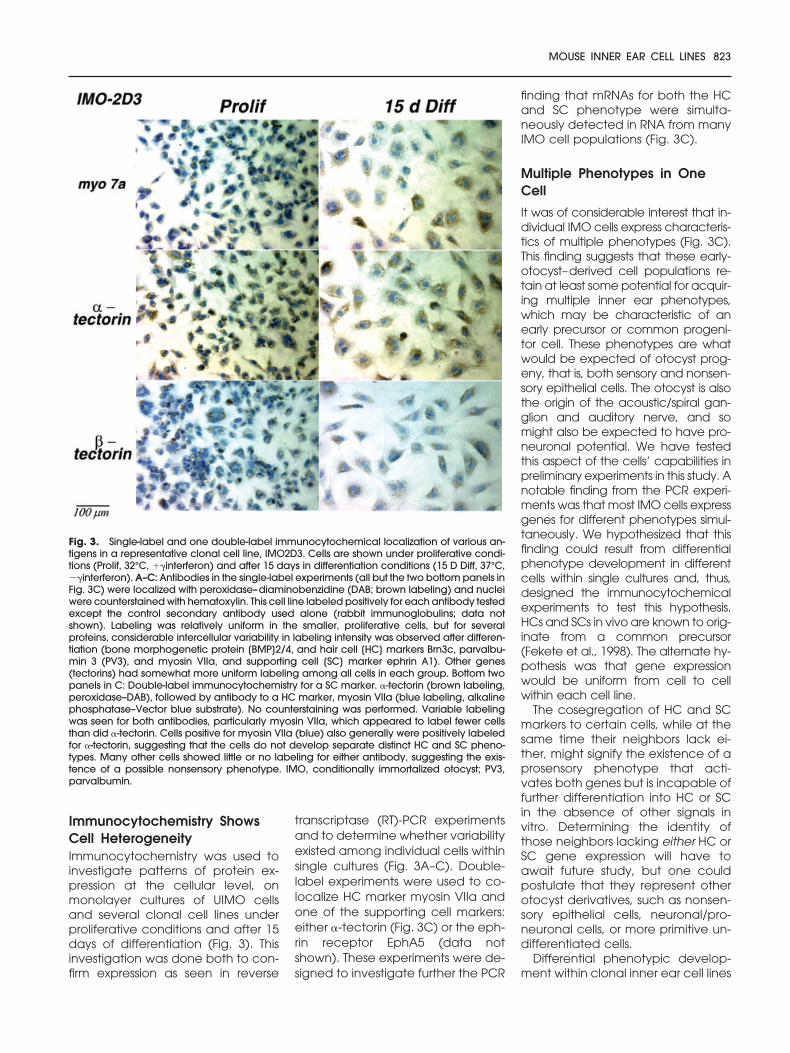
Immunocytochemistry ShowsCell HeterogeneityImmunocytochemistry was used toinvestigate patterns of protein ex-pression at the cellular level, onmonolayer cultures of UIMO cellsand several clonal cell lines underproliferative conditions and after 15days of differentiation (Fig. 3). Thisinvestigation was done both to con-firm expression as seen in reverse
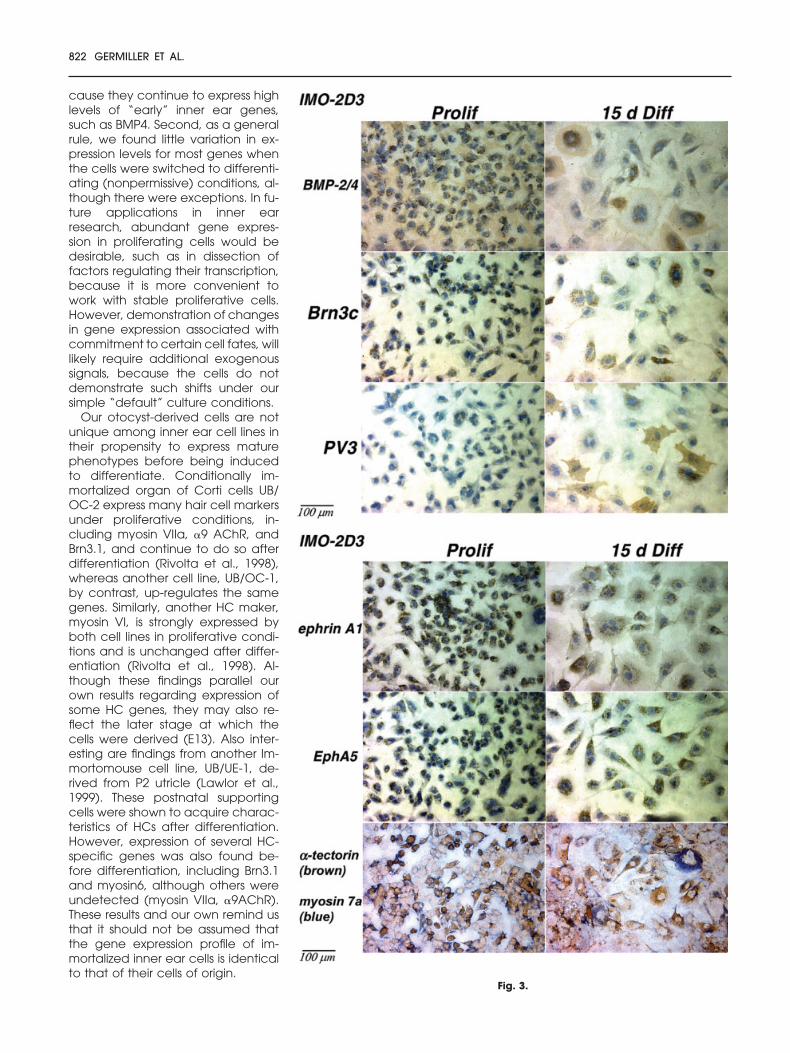
transcriptase (RT)-PCR experimentsand to determine whether variabilityexisted among individual cells withinsingle cultures (Fig. 3A–C). Double-label experiments were used to co-localize HC marker myosin VIIa andone of the supporting cell markers:either �-tectorin (Fig. 3C) or the eph-rin receptor EphA5 (data notshown). These experiments were de-signed to investigate further the PCR
finding that mRNAs for both the HCand SC phenotype were simulta-neously detected in RNA from manyIMO cell populations (Fig. 3C).
Multiple Phenotypes in OneCell
It was of considerable interest that in-dividual IMO cells express characteris-tics of multiple phenotypes (Fig. 3C).This finding suggests that these early-otocyst–derived cell populations re-tain at least some potential for acquir-ing multiple inner ear phenotypes,which may be characteristic of anearly precursor or common progeni-tor cell. These phenotypes are whatwould be expected of otocyst prog-eny, that is, both sensory and nonsen-sory epithelial cells. The otocyst is alsothe origin of the acoustic/spiral gan-glion and auditory nerve, and somight also be expected to have pro-neuronal potential. We have testedthis aspect of the cells’ capabilities inpreliminary experiments in this study. Anotable finding from the PCR experi-ments was that most IMO cells expressgenes for different phenotypes simul-taneously. We hypothesized that thisfinding could result from differentialphenotype development in differentcells within single cultures and, thus,designed the immunocytochemicalexperiments to test this hypothesis.HCs and SCs in vivo are known to orig-inate from a common precursor(Fekete et al., 1998). The alternate hy-pothesis was that gene expressionwould be uniform from cell to cellwithin each cell line.
The cosegregation of HC and SCmarkers to certain cells, while at thesame time their neighbors lack ei-ther, might signify the existence of aprosensory phenotype that acti-vates both genes but is incapable offurther differentiation into HC or SCin the absence of other signals invitro. Determining the identity ofthose neighbors lacking either HC orSC gene expression will have toawait future study, but one couldpostulate that they represent otherotocyst derivatives, such as nonsen-sory epithelial cells, neuronal/pro-neuronal cells, or more primitive un-differentiated cells.
Differential phenotypic develop-ment within clonal inner ear cell lines
Fig. 3. Single-label and one double-label immunocytochemical localization of various an-tigens in a representative clonal cell line, IMO2D3. Cells are shown under proliferative condi-tions (Prolif, 32°C, ��interferon) and after 15 days in differentiation conditions (15 D Diff, 37°C,��interferon). A–C: Antibodies in the single-label experiments (all but the two bottom panels inFig. 3C) were localized with peroxidase–diaminobenzidine (DAB; brown labeling) and nucleiwere counterstained with hematoxylin. This cell line labeled positively for each antibody testedexcept the control secondary antibody used alone (rabbit immunoglobulins; data notshown). Labeling was relatively uniform in the smaller, proliferative cells, but for severalproteins, considerable intercellular variability in labeling intensity was observed after differen-tiation (bone morphogenetic protein [BMP]2/4, and hair cell [HC] markers Brn3c, parvalbu-min 3 (PV3), and myosin VIIa, and supporting cell [SC] marker ephrin A1). Other genes(tectorins) had somewhat more uniform labeling among all cells in each group. Bottom twopanels in C: Double-label immunocytochemistry for a SC marker. �-tectorin (brown labeling,peroxidase–DAB), followed by antibody to a HC marker, myosin VIIa (blue labeling, alkalinephosphatase–Vector blue substrate). No counterstaining was performed. Variable labelingwas seen for both antibodies, particularly myosin VIIa, which appeared to label fewer cellsthan did �-tectorin. Cells positive for myosin VIIa (blue) also generally were positively labeledfor �-tectorin, suggesting that the cells do not develop separate distinct HC and SC pheno-types. Many other cells showed little or no labeling for either antibody, suggesting the exis-tence of a possible nonsensory phenotype. IMO, conditionally immortalized otocyst; PV3,parvalbumin.
MOUSE INNER EAR CELL LINES 823

has been demonstrated by others atlater stages of immortalization. Inpostnatal utricle cell line UB/UE-1,clear segregation among neighbor-ing cells is seen in their loss of celllabeling for cytokeratin upon differ-entiation, signifying the onset of dis-tinct HC-like cells (Rivolta and Holley,2002a). Moreover, only 50% of cellsacquired slow-acting rectifier cur-rents similar to HCs, with only a sub-set developing the fast inward recti-fier currents, and finally onlyapproximately 3% develop actinring structures, all suggesting thevariable expression of HC pheno-type and/or the presence of multi-ple phenotypes.
Kalinec et al. (1999) report differ-ential expression of nestin in P14 Im-mortomouse organ of Corti cell linesafter 1 week of differentiation, withconsiderable differences appearingeven between neighboring cells.Both UB/UE-1 and the Kalinec OCcells also appear to demonstrateasymmetric cell divisions at the timeof differentiation (Rivolta and Holley,2002b; Kalinec et al., 1999), whichmay be one mechanism by whichclonal cells develop divergent phe-notypes. By contrast, the well-stud-ied organ of Corti cell lines UB/OC-1and UB/OC-2 demonstrate uniformphenotypes when studied by immu-nocytochemistry (Rivolta et al.,1998).
The effects of the immortalizingoncogene that were discussed ingreat detail by the laboratory thatoriginated the transgenic mousefrom which these and many otherkinds of cell lines were derived (No-ble, 1999) apply to these cells as wellas to cell lines from other organs thatwere derived from these animals.However, a study of the regenera-tive capacity of these cells in thepresence of an immortalizing onco-gene has not yet been done in thisor any other system.
CONCLUSIONS
Otic Cell Lines Provide aGood Model System for InnerEar Development
Our results indicate that IMO celllines can express a broad repertoireof genes found in both the develop-
ing and mature inner ear, includingimportant signaling genes andmarkers of various inner ear pheno-types. The results of these studiessuggest that IMO cell lines are capa-ble of developing characteristics ofmultiple cell types of the mature in-ner ear in culture, but the relation-ships between phenotype develop-ment and differentiation status ofthe IMO cells are not simple.
Many researchers who want touse these cell lines ask how closelythey approximate the phenotypi-cally differentiated HC or SC. The an-swer is that they do not bear a phe-notypic resemblance to these cells.We never see stereocilia on thesecells unless we transfect themwith specific cytoskeletal elements(Cleveland et al., manuscript inpreparation); nor do they expressdifferentiated SC markers. However,we believe that they serve as an ex-cellent model system to study earlycues and influences on multipotentprecursor or progenitor cells thatcan direct these cells into differentdifferentiated cell pathways.
IMO cells are easy to maintain inlarge volumes, and their gene ex-pression patterns remain reproduc-ible over many experiments/pas-sages. Moreover, they express manyinner-ear–specific genes at high lev-els and do not require prolonged dif-ferentiation periods to do so. Thesecells are useful for predictive molec-ular studies, for example, analysis ofinner-ear–specific regulation of theirgene promoter regions (Thompsonet al., 2003). Although all of the indi-vidual cell lines characterized hereexpress BMP4, they present differentgene expression profiles for othermarkers. For example, the 3D1 cellline expresses neither chordin nornoggin at any stage, while theIMO1G1 cell line expresses bothBMP4 inhibitors, and noggin in-creases upon differentiation (Table1). IMO1G1 is also the only cell linethat increases its expression of Zic1to high levels on differentiation (Ta-ble 1). IMO-2B1 cells increase expres-sion of Zic2 on differentiation, asshown by conventional PCR (Table1) and by rt-qPCR (Fig. 2).
These IMO cell lines provide a reli-able, renewable, manipulable modelsystem in which to study gene regula-
tion in the early auditory system. Wehave used this system to good effectto discover a novel intron 2 promoterin the BMP4 gene expressed in theinner ear, which is down-regulated byall-trans retinoic acid (Thompson etal., 2003). Initial molecular studies inthe IMO-2B1 cell line were then veri-fied in “real” inner ears of chicks andmice. These findings can now explainwhy the results of exogenous RA ap-plication to the early developing innerear (Choo et al., 1998) and BMP4 an-tagonist application (Chang et al.,1999; Gerlach et al., 2000) result in thesame phenotypic abnormalities andsolve a long-standing controversy inthe literature.
We hypothesized that certain celllines might recapitulate the differen-tiation of mature auditory cells fromtheir primitive precursors. We deter-mined the phenotypic potential ofsome of these isolated cell lines,both in a global sense, as synchro-nized whole cultures, and in a localsense, as individual cells acting un-der influence of their neighbors. Wefound that a given cell line can ex-press markers of multiple differenti-ated auditory cell types (Fig. 3C). In-dividual cells within the clone vary inexpression of these markers. We be-lieve that these cell lines represent atleast multipotential precursor popu-lations capable of responding to lo-calized molecular signals from theirneighbors and the environment. Thenull hypotheses, which these datado not support, is that IMO cells ei-ther maintain a primitive phenotypein culture or proceed to differentiatein a uniform manner into a singlemature fate. The latter, in particular,is important to test in clonal cell lines,because each line represents theprogeny of a single cell from theoriginal otocyst. If the ultimate fateof otic cells were already deter-mined by the otocyst stage, theneach clonal cell line might be ex-pected to proceed uniformly to asingle phenotype. This process wasclearly not the case in UIMO cells.Several clonal lines expressed char-acteristics of multiple cell types ofthe developing inner ear.
One obvious question is how theculture conditions elicit responsesfrom the cells. Another issue is theinfluence that neighboring cells
824 GERMILLER ET AL.

have on the differentiation patternsof specific cells in the culture. Con-siderations include how many cell–cell contacts a given cell encoun-ters as it differentiates (colonyeffects), the sequence of a specificcell’s differentiated gene expres-sion, as well as limitations on differ-entiation presented by diminishingconcentrations of certain factors, in-cluding serum factors and cell-de-rived growth factors and �-inter-feron, but also cell-secreted factorsthat we have yet to identify.
EXPERIMENTAL PROCEDURES
Otocyst Cell Lines
Conditionally immortalized cells de-rived from the otocyst of 9.5-dayembryos of the H2kbtsA58 Immorto-mouse were maintained as eitherproliferative or differentiated cells aspreviously described (Barald et al.,1997; Thompson et al., 2003). Un-cloned, unsorted Immortomouseotocyst (UIMO) cells, representing arandom sample of the entire oto-cyst, and clonal cell lines (e.g., IMO-2B1, IMO2D3, and IMO1G1; Table 1)derived from this uncloned pop-ulation were analyzed. Chinesehamster ovary cells were used asnegative controls for immunocyto-chemistry experiments and weregrown under the proliferative anddifferentiation conditions defined forIMO cells. These cells do not expressBMP4 (Gerlach et al., 2000).
RNA Preparation, ReverseTranscription, and PCR
RNA preparation, reverse transcrip-tion, PCR, and rt-qPCR were per-formed with appropriate controlsand cycle conditions as previouslydescribed (Thompson et al., 2003) byusing primers designed for eitherconventional PCR or quantitativePCR. We designed the primer pairsfor rt-qPCR study to unique areas ofeach gene using the Beacon De-signer program (Bio-Rad) using atarget Ta of 55°C, length 18–22,and amplicon size 70–200. Theprimers were synthesized from ourdesigned sequences by InVitrogenLife Technologies (Carlsbad, CA).For the quantitative study, we used
SYBRgreen-based detection on aBio-Rad iCycler. All of these primersequences are available by emailrequest ([email protected]).
Normalized Gene Expressionfor rt-qPCR
The efficiency (E) of each primer setwas determined using dilutions ofday-12 mouse embryo cDNA. Eachprimer set was then used to amplifyIMO-2B1 cDNA made from proliferat-ing cells or from 15-day differentiatedcells and the critical threshold (Ct)was determined. The gene expressionwas normalized to the housekeepinggene glutaraldehyde phosphate de-hydrogenase (GAPDH) using the for-mula described by Stankovic andCorfas (2003):
NGE �(Etarget)�Cttarget
(EGAPDH)�CtGAPDH
. (1)
The normalized gene expressionwas plotted on a logarithmic scalescatter graph on which it is easy todepict both up- and down-regulationand the relative abundance of thegenes compared with GAPDH.Genes neither up- nor down-regu-lated fall on the diagonal. Up-regu-lated genes fall above the diagonal;down-regulated genes fall below thediagonal. A list of genes that were notamplified is also included (Fig. 2).
Cell Preparation forImmunocytochemistry
IMO cells were plated on eight-wellglass slides (Nunc LabTekII), and al-lowed to proliferate under permis-sive conditions until they were ap-proximately 70–80% confluent, atwhich point some cultures werefixed for immunocytochemical anal-ysis of the proliferative phenotype,and others were switched to differ-entiation conditions for an addi-tional 15 days (as in Barald et al.,1997). Fixation, permeabilization,binding and blocking conditions,and single and double labelingwere performed as previously de-scribed (Gardner and Barald, 1991).
Antibodies
Affinity-purified rabbit anti-humanmyosin VIIa, specific for a portion of
the tail (AA 880-1070), provided byTama Hasson through an MTA fromUniversity of California at San Diego,was used at 10 �g/ml; rabbit anti-serum to PV3, the kind gift of StefanHeller, Harvard University, was usedat 1:2,000; rabbit polyclonal anti-serum to Brn 3.1/Brn3c (BabCo-Co-vance PRB-249c) at 1:500; rabbit an-tisera to the tectorins (R9 anti–�-tectorin, R7 anti–�-tectorin) were thekind gift of Guy Richardson, Univer-sity of Sussex; each was used at1:200; rabbit anti–phrin A1 and rab-bit anti-EphA5 were from Santa Cruz,both were used at 1:100; rabbit iso-type control (Zymed, 08-6199) andmouse isotype control (Zymed, 08-6599) were both used undiluted;monoclonal mouse anti-BMP2/4(H3B2/17.8.1) were obtained fromGenetics Institute (GI) through anMTA and were used at 10 �g/ml. Un-der our MTA agreement with GI,Wyeth (who absorbed GI) was al-lowed to examine this manuscript for30 days before its submission. Mono-clonal anti-pan cytokeratin (SigmaC2562) was used at 1:100 (157 �g/ml; as in Rivolta et al., 2002).
ACKNOWLEDGMENTSWe thank Drs. Margaret Lomax, Uni-versity of Michigan, and MatthewKelley, NIH, for helpful discussionsand comments on the manuscript.Thanks to Dr. David Molea for helpwith Adobe Photoshop. K.F.B. wasfunded by the NIH and NSF; J.A.G.and K.F.B. were funded by the Deaf-ness Research Foundation, andJ.A.G. received a Resident ResearchAward from the American Acad-emy of Otolaryngology-HNS. J.S.H.was supported by an NSF REU sup-plement to K.F.B.
REFERENCES
Adam J, Myat A, Le Roux I, Eddison M,Henrique D, Ish-Horowicz D, Lewis J.1998. Cell fate choices and the expres-sion of Notch, Delta and Serrate homo-logues in the chick inner ear: parallelswith Drosophila sense-organ develop-ment. Development 125:4645–4654.
Barald KF, Kelley MW. 2004. From placodeto polarization: new tunes in inner eardevelopment. Development 131:4119–4130.
Barald KF, Lindberg KH, Hardiman K,Kavka AI, Lewis JE, Victor JC, GardnerCA, Poniatowski A. 1997. Immortalized
MOUSE INNER EAR CELL LINES 825

cell lines from embryonic avian andmurine otocysts: tools for molecularstudies of the developing inner ear. IntJ Dev Neurosci 15:523–540.
Bermingham NA, Hassan BA, Price SD,Vollrath MA, Ben-Arie N, Eatock RA,Bellen HJ, Lysakowski A, Zoghbi HY.1999. Math1: an essential gene for thegeneration of inner ear hair cells. Sci-ence 284:1837–1841.
Brigande JV, Kiernan AE, Gao X, Iten LE,Fekete DM. 2000a. Molecular genetics ofpattern formation in the inner ear: docompartment boundaries play a role?Proc Natl Acad Sci U S A 97:11700–11706.
Brigande JV, Iten LE, Fekete DM. 2000b. Afate map of chick otic cup closure re-veals lineage boundaries in the dorsalotocyst. Dev Biol 227:256–270.
Chang W, Nunes FD, De Jesus-EscobarJM, Harland R, Wu DK. 1999. Ectopicnoggin blocks sensory and nonsensoryorgan morphogenesis in the chickeninner ear. Dev Biol 216:369–381.
Choo D, Sanne JL, Wu DK. 1998. The dif-ferential sensitivities of inner ear struc-tures to retinoic acid during develop-ment. Dev Biol 204:136–150.
Cole LK, Le Roux I, Nunes F, Laufer E, LewisJ, Wu DK. 2000. Sensory organ genera-tion in the chicken inner ear: contribu-tions of bone morphogenetic protein 4,serrate1, and lunatic fringe. J CompNeurol 424:509–520.
Echteler SM, Fay RR, Popper AN. 1994.Structure of the mammalian cochlea.In: Fay RR, Popper AN, editors. Com-parative hearing: mammals. Springerhandbook of auditory research. Vol. 4.New York: Springer. p 134–171.
Elgoyhen AB, Johnson DS, Boulter J, VetterDE, Heinemann S. 1994. Alpha9: an ace-tylcholine receptor with novel pharma-cological properties expressed in rat co-chlear hair cells. Cell 79:705–715.
Elgoyhen AB, Vetter DE, Katz E, Rothlin CV,Heinemann SF, Boulter J. 2001. Alpha 10: adeterminant of nicotinic cholinergic re-ceptor function in mammalian vestibularand cochlear mechanosensory hair cells.Proc Natl Acad Sci U S A 98:3501–3506.
Elms P, Siggers P, Napper D, Greenfield A,Arkell R. 2003. Zic2 is required for neuralcrest formation and hindbrain pattern-ing during mouse development. DevBiol 264:391–406.
Erkman L, McEvilly RJ, Luo L, Ryan AK, Hoo-shmand F, O’Connell SM, Keithley EM,Rapaport DH, Ryan AF, Rosenfeld MG.1996. Role of transcription factors Brn-3 1and Brn-3 2 in auditory and visual systemdevelopment. Nature 381:603–606.
Fekete DM. 1996. Cell fate specificationin the inner ear. Curr Opin Neurobiol6:533–541.
Fekete DM. 2000a. Making sense of mak-ing hair cells. Trends Neurosci 23:386.
Fekete DM. 2000b. Ear rings: FGF3 in-volvement comes full circle. TrendsNeurosci 23:332.
Fekete DM, Wu DK. 2002. Revisiting cellfate specification in the inner ear. CurrOpin Neurobiol 12:35–42.
Fekete DM, Muthukumar S, Karagogeos D.1998. Hair cells and supporting cellsshare a common progenitor in thechicken inner ear. J Neurosci 18:7811–7821.
Fritzsch B, Beisel KW. 2001. Evolution anddevelopment of the vertebrate ear.Brain Res Bull 55:711–721.
Fritzsch B, Silos-Santiago I, Bianchi LM, Fa-rinas I. 1997. The role of neurotrophicfactors in regulating the developmentof inner ear innervation. Trends Neuro-sci 20:159–164.
Gardner CA, Barald KF. 1991. The cellularenvironment controls the expression ofengrailed-like protein in the cranial neu-roepithelium of quail/chick-chimericembryos. Development 113:1037–1048.
Gerlach LM, Hutson MR, Germiller JA,Nguyen-Luu D, Victor JC, Barald KF.2000. Addition of the BMP4 antagonist,noggin, disrupts avian inner ear devel-opment. Development 127:45–54.
Gerlach-Bank LM, Ellis AD, Noonen B,Barald KF. 2002. Cloning and expres-sion analysis of the chick DAN gene, anantagonist of the BMP family of growthfactors. Dev Dyn 224:109–115.
Glowatski E, Wild K, Brandle U, Fakler G,Fakler B, Zenner HP, Ruppersberg JP.1995. Cell-specific expression of the al-pha9-n-Ach receptor subunit in audi-tory hair cells revealed by single-cellRT-PCR. Proc R Soc Lond B 262:141–147.
Hamburger V, Hamilton VL. 1951. A seriesof normal stages in the development ofthe chick embryo. J Morphol 88:49–92.
Hasson T, Gillespie PG, Garcia JA, Mac-Donald RB, Zhao Y, Yee AG, MoosekerMS, Corey DP. 1997. Unconventionalmyosins in inner ear sensory epithelia.J Cell Biol 137:1287–1307.
Heller S, Bell AM, Denis CS, Choe Y, Hud-speth AJ. 2002. Parvalbumin 3 is anabundant Ca2� buffer in hair cells. JAssoc Res Otolaryngol 3:488–498.
Hemmati-Brivanlou A, Thomsen GH. 1995.Ventral mesodermal patterning in Xe-nopus embryos: expression patternsand activities of BMP2 and BMP4. DevGenet 17:78–89.
Hemond SG, Morest DK. 1991. Ganglionformation from the otic placode andthe otic crest in the chick embryo: mi-tosis, migration, and the basal lamina.Anat Embryol (Berl) 184:113.
Hogan BL, Blessing M, Winnier GE, SuzukiN, Jones CM. 1994. Growth factors indevelopment: the role of TGF-beta re-lated polypeptide signaling moleculesin embryogenesis. Dev Suppl :53–60.
Holley MC, Nishida Y, Grix N. 1997. Con-ditional immortalization of hair cellsfrom the inner ear. Int J Dev Neurosci15:541–552.
Jat SJ, Noble MD, Ataliotis PM, Tanaka Y,Yannoutsos N, Larsen L, Kioussis D. 1991.Direct derivation of conditionally im-mortal cell lines from an H2KbtsA58transgenic mouse. Proc Natl Acad SciU S A 88:5096–5100.
Kalinec F, Kalinec G, Boukhvalova M, Ka-char B. 1999. Establishment and char-acterization of conditionally immortal-
ized organ of Corti cell lines. Cell Biol Int23:175–184.
Kawamoto K, Ishimoto S, Minoda R,Brough DE, Raphael Y. 2003. Math1gene transfer generates new cochlearhair cells in mature guinea pigs in vivo.J Neurosci 23:4395–4400.
Kimble J, Simpson P. 1997. The LIN12/Notchsignaling pathway and its regulation.Ann Rev Cell Dev Biol 13:333–361.
Kopan R, Cagan R. 1997. Notch on thecutting edge. Trends Genet 13:465–467.
Lanford PJ, Lan Y, Jiang R, Lindsell C,Weinmaster G, Gridley T, Kelley MW.1999. Notch signaling pathway medi-ates hair cell development in mamma-lian cochlea. Nat Genet 21:289–292.
Lang H, Fekete DM. 2001. Lineage anal-ysis in the chicken inner ear shows dif-ferences in clonal dispersion for epithe-lial, neuronal, and mesenchymal cells.Dev Biol 234:120–137.
Lawlor P, Marcotti W, Rivolta MN, Kros CJ,Holley MC. 1999. Differentiation of mam-malian vestibular hair cells from condi-tionally immortal, postnatal supportingcells. J Neurosci 19:9445–9458.
Legan PK, Rau A, Keen JN, RichardsonGP. 1997. The mouse tectorins. Modularmatrix proteins of the inner ear homol-ogous to components of the sperm-egg adhesion system. J Biol Chem 272:8791–8801.
Lewis J. 1996. Neurogenic genes and ver-tebrate neurogenesis. Curr Opin Neu-robiol 6:3–10.
Liu W, Oh SH, Kang YkY, Li G, Doan TM,Little M, Li L, Ahn K, Crenshaw EB III,Frenz DA. 2003a. Bone morphogeneticprotein 4 (BMP4): a regulator of cap-sule chondrogenesis in the developingmouse inner ear. Dev Dyn 226:427–438.
Liu D, Chu H, Maves L, Yan Y-L, MorcosPA, Postlethwait JH, Westerfield M.2003b. Fgf3 and Fgf8 dependent andindependent transcription factors arerequired for otic placode specification.Development 130:2213–2224.
Morsli H, Choo D, Ryan A, Johnson R, WuDK. 1998. Development of the mouseinner ear and origin of its sensory or-gans. J Neurosci 18:3327–3335.
Noble M. 1999. Production and growth ofconditionally immortal cell lines fromthe H-2KbtsA58 transgenic mouse.Methods Mol Biol 97:139–158.
Oh SH, Johnson R, Wu DK. 1996. Differen-tial expression of bone morphogeneticproteins in the developing vestibularand auditory sensory organs. J Neuro-sci 16:6463–6475.
Rivolta MN, Holley MC. 2002a. Cell lines ininner ear research. J Neurobiol 53:306–318.
Rivolta MN, Holley MC. 2002b. Asymmet-ric segregation of mitochondria andmortalin correlates with the multi-lin-eage potential of inner ear sensory cellprogenitors in vitro. Brain Res Dev BrainRes 133:49–56.
Rivolta MN, Grix N, Lawlor P, Ashmore JF,Jagger DJ, Holley MC. 1998. Auditoryhair cell precursors immortalized from
826 GERMILLER ET AL.

the mammalian inner ear. Proc R ScoLond B Biol Sci 265:1595–1603.
Rivolta MN, Halsall A, Johnson CM, TonesMA, Holley MC. 2002. Transcript profil-ing of functionally related groups ofgenes during conditional differentia-tion of a mammalian cochlear hair cellline. Genome Res 12:1091–1099.
Ryan AF. 1997. Transcription factors andthe control of inner ear development.Semin Cell Dev Biol 8:249–256.
Stankovic KM, Corfas G. 2003. Real-timequantitative RT-PCR for low-abun-dance transcripts in the inner ear: anal-ysis of neurotrophic factor expression.Hearing Res 185:97–108.
Stone JS, Rubel EW. 2000a. Cellular stud-ies of audatory hair cell regeneration inbirds. Proc Nat Acad Sci USA 97:11714–11721.
Stone JS, Rubel EW. 2000b. Temporal,spatial, and morphologic features ofhair cell regeneratoin in the avian basi-lar papilla. J Comp Neurol 417:1–16.
Thomsen GH. 1997. Antagonism withinand around the organizer: BMP inhibi-tors in vertebrate body patterning.Trends Genet 13:209–211.
Thompson D, Gerlach-Bank LM, BaraldKF, Koenig RJ. 2003. Retinoic acid re-pression of BMP4 in inner ear develop-ment. Mol Cell Biol 23:2277–2286.
Vahava O, Morell R, Lynch ED, Weiss S,Kagan ME, Ahituv N, Morrow JE, LeeMK, Skvorak AB, Morton CC, Blumen-feld A, Frydman M, Friedman TB, KingMC, Avraham KB. 1998. Mutation intranscription factor POU4F3 associatedwith inherited progressive hearing lossin humans. Science 279:1950–1954.
Wall NA, Hogan BL. 1994. TGF-beta re-lated genes in development. Curr OpinGenet Dev 4:517–522.
Warner SJ, Hutson MR, Oh SH, Gerlach-Bank LM, Lomax MI, Barald KF. 2003.Expression of ZIC genes in thedevelopment of the chick inner earand nervous system. Dev Dyn 226:702–712.
Weir J, Rivolta M, Holley MC. 2000. Notchsignaling and the emergence of audi-tory hair cells. Arch Otol Head NeckSurg 126:1244–1248.
Xiang M, Gan L, Li D, Chen ZY, Zhou L,O’Malley BWJ, Klein W, Nathans J.1997. Essential role of POU-domain fac-tor Brn-3c in auditory and vestibularhair cell development. Proc Natl AcadSci U S A 94:9445–9450.
MOUSE INNER EAR CELL LINES 827