mitochondrial defects confer tolerance against cellulose ... · mitochondrial defects confer...
TRANSCRIPT

Mitochondrial Defects Confer Tolerance againstCellulose Deficiency
Zhubing Hu,a,b,c Rudy Vanderhaeghen,a,b Toon Cools,a,b Yan Wang,d Inge De Clercq,a,b Olivier Leroux,e
Long Nguyen,f Katharina Belt,g A. Harvey Millar,g Dominique Audenaert,f Pierre Hilson,a,b,h Ian Small,g
Grégory Mouille,h Samantha Vernhettes,h Frank Van Breusegem,a,b James Whelan,d Herman Höfte,h andLieven De Veyldera,b,1
a Department of Plant Systems Biology, VIB, B-9052 Gent, BelgiumbDepartment of Plant Biotechnology and Bioinformatics, Ghent University, B-9052 Gent, BelgiumcCollege of Life Sciences, Nanjing Agricultural University, 210095 Nanjing, People’s Republic of ChinadDepartment of Botany, ARC Centre of Excellence in Plant Energy Biology, School of Life Science, La Trobe University, Bundoora3086, Victoria, Australiae Department of Biology, Ghent University, B-9000 Gent, BelgiumfCompound Screening Facility, VIB, B-9052 Gent, BelgiumgAustralian Research Council Centre of Excellence in Plant Energy Biology, University of Western Australia, Crawley 6009, Australiah Institut Jean-Pierre Bourgin, INRA, Centre National pour la Recherche Scientifique, AgroParisTech, Université Paris-Saclay,78026 Versailles Cedex, France
ORCID IDs: 0000-0002-1258-5531 (T.C); 0000-0001-8125-1239 (I.D.C.); 0000-0003-4617-1908 (P.H.); 0000-0001-5300-1216 (I.S.);0000-0002-3147-0860 (F.V.B); 0000-0001-5754-025X (J.W); 0000-0003-1150-4426 (L.D.V.)
Because the plant cell wall provides the first line of defense against biotic and abiotic assaults, its functional integrity needs tobe maintained under stress conditions. Through a phenotype-based compound screening approach, we identified a novelcellulose synthase inhibitor, designated C17. C17 administration depletes cellulose synthase complexes from the plasmamembrane in Arabidopsis thaliana, resulting in anisotropic cell elongation and a weak cell wall. Surprisingly, in addition tomutations in CELLULOSE SYNTHASE1 (CESA1) and CESA3, a forward genetic screen identified two independent defectivegenes encoding pentatricopeptide repeat (PPR)-like proteins (CELL WALL MAINTAINER1 [CWM1] and CWM2) as conferringtolerance to C17. Functional analysis revealed that mutations in these PPR proteins resulted in defective cytochrome cmaturation and activation of mitochondrial retrograde signaling, as evidenced by the induction of an alternative oxidase.These mitochondrial perturbations increased tolerance to cell wall damage induced by cellulose deficiency. Likewise,administration of antimycin A, an inhibitor of mitochondrial complex III, resulted in tolerance toward C17. The C17 tolerance ofcwm2 was partially lost upon depletion of the mitochondrial retrograde regulator ANAC017, demonstrating that ANAC017links mitochondrial dysfunction with the cell wall. In view of mitochondria being a major target of a variety of stresses, ourdata indicate that plant cells might modulate mitochondrial activity to maintain a functional cell wall when subjected tostresses.
INTRODUCTION
Cellulose is a main component of the plant cell wall, consisting ofa linear chain of several hundred to many thousands of b-1,4-linked D-glucose units (McFarlane et al., 2014). Its synthesis isachieved through a plasmamembrane-localized protein complexreferred to as the cellulose synthase complex (CSC) that hasa hexameric rosette-like structure (Endler and Persson, 2011).Each of the six lobes is thought to contain three distinct cellulosesynthase catalytic subunits (CESAs) corresponding to CESA1,CESA3, and CESA6 (or CESA6-like proteins CESA2, CESA5, and
CESA9) in theprimary cellwall (Desprezet al., 2007;Perssonet al.,2007; Vandavasi et al., 2016).The activity of the CSC is modulated by abiotic and biotic
stresses. For example, when exposed to osmotic stress, plantsdownregulate their cellulose production through the depletion ofCSCs from the plasmamembrane (Gutierrez et al., 2009; Lei et al.,2015). A similar depletion can be triggered by small moleculesproducedbypathogens, such as thaxtominA, apotent inhibitor ofcellulose biosynthesis produced by the plant pathogen Strepto-myces scabies responsible for the scab disease (Crowell et al.,2009). Recently, a previously unidentified CSC inhibitor, aceto-bixan, was isolated from small molecule secretions derived froma library of switchgrass endophytes (Xia et al., 2014).Because the cell wall is a key feature of plant cells, dedicated
systems have evolved to monitor its integrity and to triggerchanges in its composition and structure through tightly con-trolled enzymatic modifications and shifts in cellular metabolism(Hamann, 2015). Cell wall damage induces a wide range of
1 Address correspondence to [email protected] author responsible for distribution of materials integral to the findingspresented in this article in accordance with the policy described in theInstructions for Authors (www.plantcell.org) is: Lieven De Veylder ([email protected]).www.plantcell.org/cgi/doi/10.1105/tpc.16.00540
The Plant Cell, Vol. 28: 2276–2290, September 2016, www.plantcell.org ã 2016 American Society of Plant Biologists. All rights reserved.
responses, including ectopic lignin deposition, activation ofjasmonate and ethylene signaling pathways, and upregulation ofstress response genes (Caño-Delgado et al., 2000; Ellis andTurner, 2001; Ellis et al., 2002;Caño-Delgado et al., 2003). Severalproteins have been identified to control these responses (Wolfet al., 2012). Among these, THESEUS1 (THE1) is one of the best-studied components, belonging to the family of the Catharanthusroseus receptor-like kinases. THE1 was originally identified ina screen for the suppression of the elongation defect in the cel-lulose-deficient mutant cesa6/procuste1. Depletion of THE1partially restores growth inhibition and attenuates ectopic ligni-fication of cellulose-deficient mutants, but fails to rescue thecellulose deficiency (Hématy et al., 2007).
In addition to the cell wall, mitochondria have been consideredto be the target of a variety of stresses (Bartoli et al., 2004; Giraudet al., 2008). Generally, stresses can trigger the accumulation ofreactive oxygen species (ROS), which in turn result in oxidativedamage to mitochondria (Fujita et al., 2006; Gechev et al., 2006).Apart frombeing a target of ROS,mitochondria are amajor sourceofcellularROS (Rhoadsetal., 2006).Mitochondriapossessa largenumber of cellular enzymatic systems that catalyze the oxidationof various substrates and generate the reducing equivalents toreduce thepyridineandflavinnucleotidesNADandFAD.ReducedNAD and FAD in turn are oxidized by coenzyme Q in reactionscatalyzed by several enzyme complexes located in the innermembrane of mitochondria. Ultimately, the flux of electrons fromsubstrates through various redox carriers and centers is termi-nated in a four-electron reduction of molecular oxygen to water,catalyzed by the cytochrome c oxidase (Starkov, 2008). Stress-induced inhibition of the various mitochondrial electron transportprotein complexesmight result in an increase in the nonenzymatic
single-electron reduction of oxygen, converting it into superoxide,a progenitor ROS (Starkov, 2008; Millar et al., 2011; Huang et al.,2016). Although many mitochondrial mutants exhibit increasedROS levels (Zsigmond et al., 2008; Liu et al., 2010; He et al., 2012;Yang et al., 2014; Zhu et al., 2014), a recent study reported thatinhibiting mitochondrial activity might play a protective role toprevent the production of mitochondrial ROS and diminish theROS-induced damage (Wu et al., 2015). Moreover, perturbingmitochondrial functions activates signaling cascades from thisorganelle to the nucleus, resulting in the modulation of gene ex-pression (Rhoads andSubbaiah, 2007; Ng et al., 2014; Huang et al.,2016). In plants, several signaling components have been identifiedfor this process, which include the cyclin-dependent kinase E1(CDKE1), as well as the transcription factors ABSCISIC ACIDINSENSITIVE4,WRKY40,NOAPICALMERISTEM/ARABIDOPSISTRANSCRIPTION FACTOR/CUP-SHAPED COTYLEDON013(ANAC013), and ANAC017 (Giraud et al., 2009; De Clercq et al.,2013; Ng et al., 2013b; Ng et al., 2013a; VanAken et al., 2013). Thedepletion of either ANAC013 or ANAC017 affects mitochondrialretrograde signaling, resulting in plants being hypersensitive toabiotic stresses (De Clercq et al., 2013; Ng et al., 2013a), whichhighlights the importance of mitochondria in stress adaptation.Comparative analysis shows that among relatedNACs,ANAC017is almost solely responsible for transcript induction of mitochondrialretrograde marker genes after chemical inhibition of organellefunction (Van Aken et al., 2016).Despite the crucial role of the plant mitochondria and the cell
wall in stress response, it is unclear whether these two com-partments are functionally linked.Here,we identify aCSC inhibitornamed C17 that reduces cellulose production through pertur-bation of CSC activity. Surprisingly, C17-induced growth defects
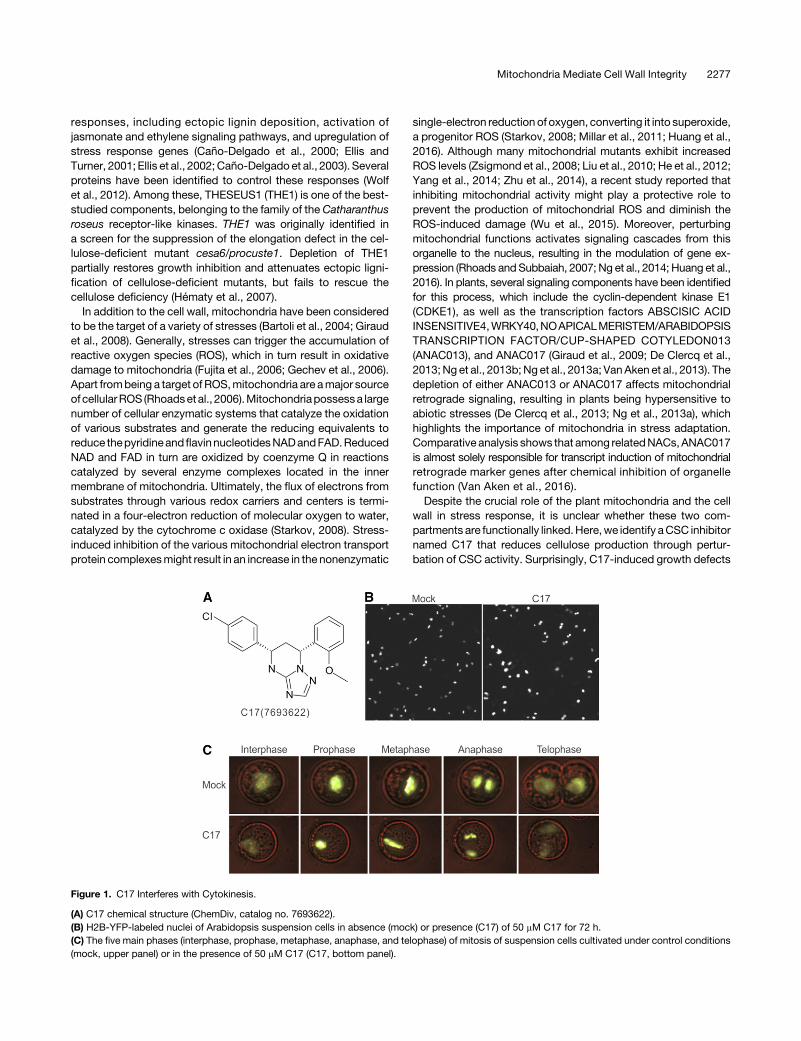
Figure 1. C17 Interferes with Cytokinesis.
(A) C17 chemical structure (ChemDiv, catalog no. 7693622).(B) H2B-YFP-labeled nuclei of Arabidopsis suspension cells in absence (mock) or presence (C17) of 50 mM C17 for 72 h.(C) The five main phases (interphase, prophase, metaphase, anaphase, and telophase) of mitosis of suspension cells cultivated under control conditions(mock, upper panel) or in the presence of 50 mM C17 (C17, bottom panel).
Mitochondria Mediate Cell Wall Integrity 2277
can be suppressed by perturbation of mitochondrial activity andby activation ofmitochondrial retrograde signaling, indicating thatmodulation of mitochondrial activity might be required for themaintenance of functional cell walls.
RESULTS
C17 Interferes with Cytokinesis during Mitotic Division
Plants are vulnerable to environmental stress conditions thatinterfere with a series of physiological processes, ultimatelyresulting in acell cycle exit andgrowth inhibition.Compounds thatinterfere with cell division can be identified by assessment ofintracellular DNA accumulation. To screen for novel cell division-interfering compounds, a high-throughput chemical screen wasperformed using an Arabidopsis thaliana cell suspension lineproducing a translational fusion between the ArabidopsisHistone2B and Yellow Fluorescent Protein genes (H2B-YFP). Because H2B-YFP associates with chromatin and is fluorescently
marked, DNA accumulation can be indirectly measured as en-hanced YFP fluorescence. The H2B-YFP cells were treated witha chemical library of 12,000 organic molecules (DIVERSet;ChemBridge). Three compounds were found to induce poly-ploidy within 72 h of treatment, of which the synthetic moleculeC17 (5-(4-chlorophenyl)-7-(2-methoxyphenyl)-1,5,6,7-tetrahydro-[1,2,4]triazolo[1,5-a] pyrimidine; ChemBridge, catalogue no.7693622) was the most effective (Figure 1A). C17-treated cellsshowed larger nuclei and higher YFP fluorescence (Figure 1B).Through time-lapse imaging, C17 was found to inhibit cyto-kinesis without affecting mitosis, thus leading to endomitosis(Figure 1C; Supplemental Movies 1 and 2).
Mutations in CESA1 and CESA3 Confer C17 Tolerance
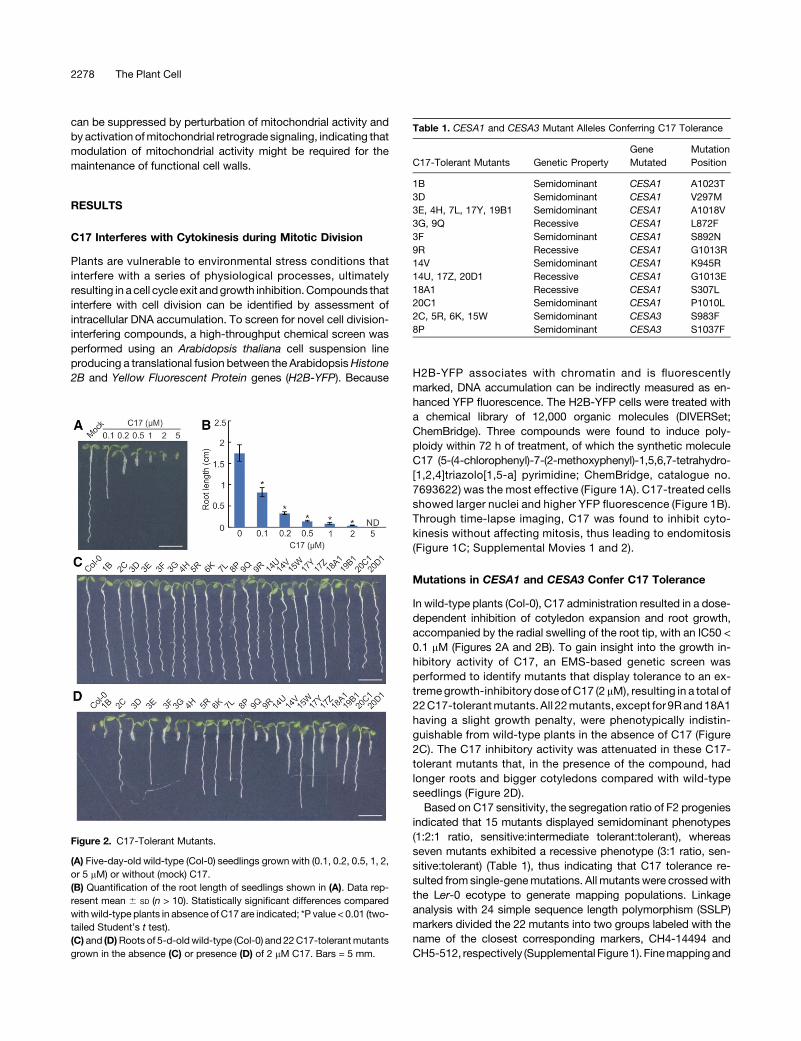
In wild-type plants (Col-0), C17 administration resulted in a dose-dependent inhibition of cotyledon expansion and root growth,accompanied by the radial swelling of the root tip, with an IC50 <0.1 mM (Figures 2A and 2B). To gain insight into the growth in-hibitory activity of C17, an EMS-based genetic screen wasperformed to identify mutants that display tolerance to an ex-tremegrowth-inhibitory dose of C17 (2 mM), resulting in a total of22C17-tolerantmutants. All 22mutants, except for 9Rand18A1having a slight growth penalty, were phenotypically indistin-guishable from wild-type plants in the absence of C17 (Figure2C). The C17 inhibitory activity was attenuated in these C17-tolerant mutants that, in the presence of the compound, hadlonger roots and bigger cotyledons compared with wild-typeseedlings (Figure 2D).Based on C17 sensitivity, the segregation ratio of F2 progenies
indicated that 15 mutants displayed semidominant phenotypes(1:2:1 ratio, sensitive:intermediate tolerant:tolerant), whereasseven mutants exhibited a recessive phenotype (3:1 ratio, sen-sitive:tolerant) (Table 1), thus indicating that C17 tolerance re-sulted from single-genemutations. All mutants were crossedwiththe Ler-0 ecotype to generate mapping populations. Linkageanalysis with 24 simple sequence length polymorphism (SSLP)markers divided the 22 mutants into two groups labeled with thename of the closest corresponding markers, CH4-14494 andCH5-512, respectively (Supplemental Figure1). Finemappingand
Figure 2. C17-Tolerant Mutants.
(A) Five-day-old wild-type (Col-0) seedlings grown with (0.1, 0.2, 0.5, 1, 2,or 5 mM) or without (mock) C17.(B) Quantification of the root length of seedlings shown in (A). Data rep-resent mean 6 SD (n > 10). Statistically significant differences comparedwith wild-type plants in absence of C17 are indicated; *P value < 0.01 (two-tailed Student’s t test).(C) and (D)Roots of 5-d-oldwild-type (Col-0) and 22C17-tolerantmutantsgrown in the absence (C) or presence (D) of 2 mM C17. Bars = 5 mm.
Table 1. CESA1 and CESA3 Mutant Alleles Conferring C17 Tolerance
C17-Tolerant Mutants Genetic PropertyGeneMutated
MutationPosition
1B Semidominant CESA1 A1023T3D Semidominant CESA1 V297M3E, 4H, 7L, 17Y, 19B1 Semidominant CESA1 A1018V3G, 9Q Recessive CESA1 L872F3F Semidominant CESA1 S892N9R Recessive CESA1 G1013R14V Semidominant CESA1 K945R14U, 17Z, 20D1 Recessive CESA1 G1013E18A1 Recessive CESA1 S307L20C1 Semidominant CESA1 P1010L2C, 5R, 6K, 15W Semidominant CESA3 S983F8P Semidominant CESA3 S1037F
2278 The Plant Cell
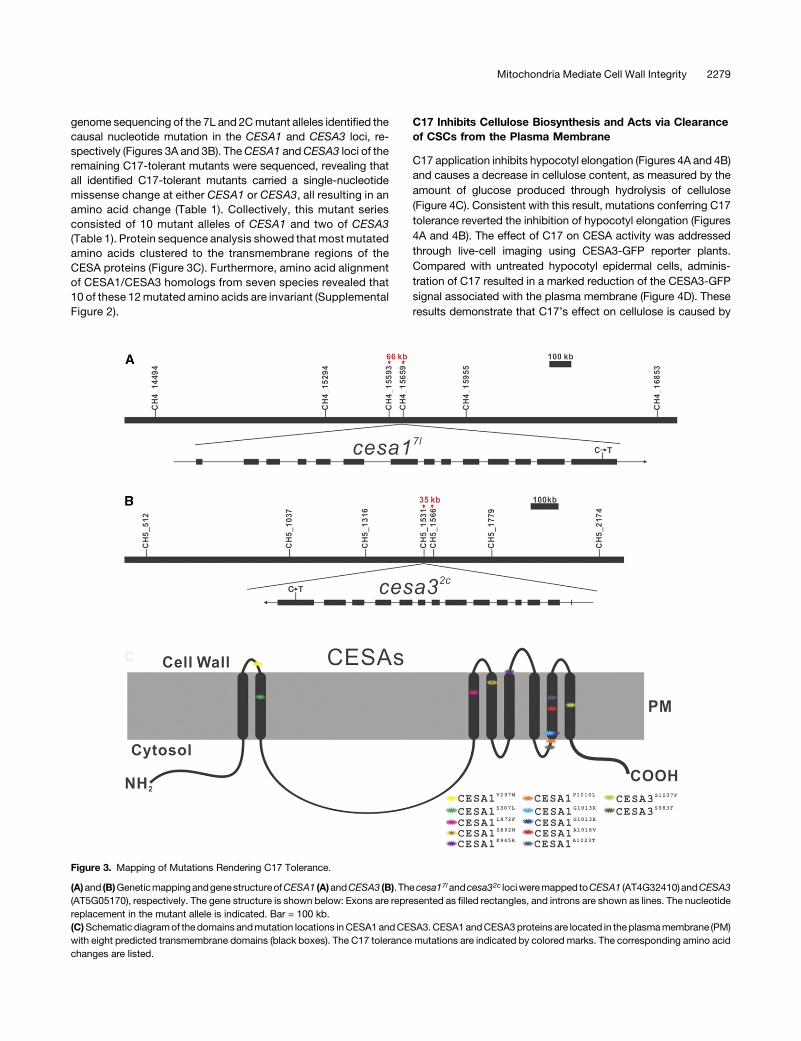
genome sequencing of the 7L and 2Cmutant alleles identified thecausal nucleotide mutation in the CESA1 and CESA3 loci, re-spectively (Figures 3A and 3B). TheCESA1 andCESA3 loci of theremaining C17-tolerant mutants were sequenced, revealing thatall identified C17-tolerant mutants carried a single-nucleotidemissense change at either CESA1 or CESA3, all resulting in anamino acid change (Table 1). Collectively, this mutant seriesconsisted of 10 mutant alleles of CESA1 and two of CESA3(Table 1). Protein sequence analysis showed that most mutatedamino acids clustered to the transmembrane regions of theCESA proteins (Figure 3C). Furthermore, amino acid alignmentof CESA1/CESA3 homologs from seven species revealed that10 of these 12mutated amino acids are invariant (SupplementalFigure 2).
C17 Inhibits Cellulose Biosynthesis and Acts via Clearanceof CSCs from the Plasma Membrane
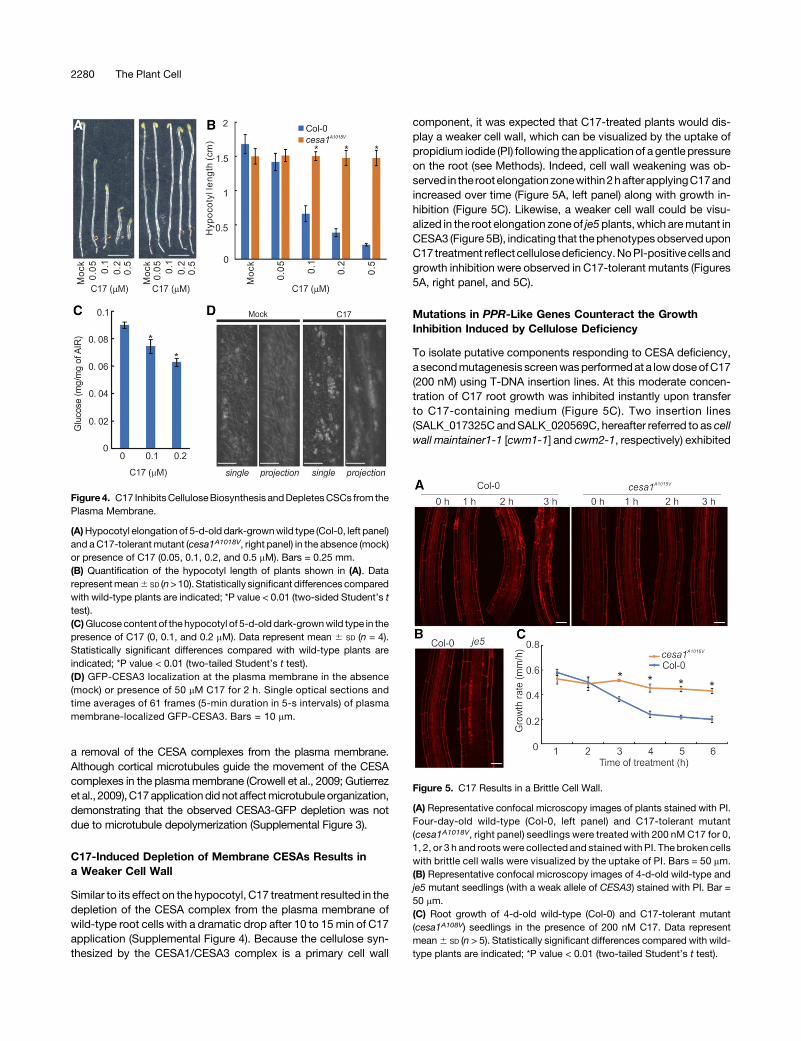
C17 application inhibits hypocotyl elongation (Figures 4A and 4B)and causes a decrease in cellulose content, as measured by theamount of glucose produced through hydrolysis of cellulose(Figure 4C). Consistent with this result, mutations conferring C17tolerance reverted the inhibition of hypocotyl elongation (Figures4A and 4B). The effect of C17 on CESA activity was addressedthrough live-cell imaging using CESA3-GFP reporter plants.Compared with untreated hypocotyl epidermal cells, adminis-tration of C17 resulted in a marked reduction of the CESA3-GFPsignal associated with the plasma membrane (Figure 4D). Theseresults demonstrate that C17’s effect on cellulose is caused by
Figure 3. Mapping of Mutations Rendering C17 Tolerance.
(A)and (B)GeneticmappingandgenestructureofCESA1 (A)andCESA3 (B). Thecesa17landcesa32c lociweremapped toCESA1 (AT4G32410)andCESA3(AT5G05170), respectively. The gene structure is shown below: Exons are represented as filled rectangles, and introns are shown as lines. The nucleotidereplacement in the mutant allele is indicated. Bar = 100 kb.(C)Schematic diagramof thedomains andmutation locations inCESA1andCESA3.CESA1andCESA3proteins are located in theplasmamembrane (PM)with eight predicted transmembrane domains (black boxes). The C17 tolerance mutations are indicated by colored marks. The corresponding amino acidchanges are listed.
Mitochondria Mediate Cell Wall Integrity 2279
a removal of the CESA complexes from the plasma membrane.Although cortical microtubules guide the movement of the CESAcomplexes in the plasmamembrane (Crowell et al., 2009; Gutierrezet al., 2009),C17applicationdidnot affectmicrotubuleorganization,demonstrating that the observed CESA3-GFP depletion was notdue to microtubule depolymerization (Supplemental Figure 3).
C17-Induced Depletion of Membrane CESAs Results ina Weaker Cell Wall
Similar to its effect on the hypocotyl, C17 treatment resulted in thedepletion of the CESA complex from the plasma membrane ofwild-type root cells with a dramatic drop after 10 to 15 min of C17application (Supplemental Figure 4). Because the cellulose syn-thesized by the CESA1/CESA3 complex is a primary cell wall
component, it was expected that C17-treated plants would dis-play a weaker cell wall, which can be visualized by the uptake ofpropidium iodide (PI) following the application of a gentle pressureon the root (see Methods). Indeed, cell wall weakening was ob-served in the rootelongationzonewithin2hafterapplyingC17andincreased over time (Figure 5A, left panel) along with growth in-hibition (Figure 5C). Likewise, a weaker cell wall could be visu-alized in the root elongation zoneof je5plants, which aremutant inCESA3 (Figure 5B), indicating that the phenotypes observeduponC17 treatment reflectcellulosedeficiency.NoPI-positivecells andgrowth inhibition were observed in C17-tolerant mutants (Figures5A, right panel, and 5C).
Mutations in PPR-Like Genes Counteract the GrowthInhibition Induced by Cellulose Deficiency
To isolate putative components responding to CESA deficiency,a secondmutagenesis screenwasperformedat a lowdoseofC17(200 nM) using T-DNA insertion lines. At this moderate concen-tration of C17 root growth was inhibited instantly upon transferto C17-containing medium (Figure 5C). Two insertion lines(SALK_017325C andSALK_020569C, hereafter referred to as cellwall maintainer1-1 [cwm1-1] and cwm2-1, respectively) exhibited
Figure4. C17 InhibitsCelluloseBiosynthesis andDepletesCSCs from thePlasma Membrane.
(A)Hypocotyl elongationof 5-d-old dark-grownwild type (Col-0, left panel)and aC17-tolerantmutant (cesa1A1018V, right panel) in the absence (mock)or presence of C17 (0.05, 0.1, 0.2, and 0.5 mM). Bars = 0.25 mm.(B) Quantification of the hypocotyl length of plants shown in (A). Datarepresentmean6 SD (n>10). Statistically significant differences comparedwith wild-type plants are indicated; *P value < 0.01 (two-sided Student’s ttest).(C)Glucose content of the hypocotyl of 5-d-old dark-grownwild type in thepresence of C17 (0, 0.1, and 0.2 mM). Data represent mean 6 SD (n = 4).Statistically significant differences compared with wild-type plants areindicated; *P value < 0.01 (two-tailed Student’s t test).(D) GFP-CESA3 localization at the plasma membrane in the absence(mock) or presence of 50 mM C17 for 2 h. Single optical sections andtime averages of 61 frames (5-min duration in 5-s intervals) of plasmamembrane-localized GFP-CESA3. Bars = 10 mm.
Figure 5. C17 Results in a Brittle Cell Wall.
(A) Representative confocal microscopy images of plants stained with PI.Four-day-old wild-type (Col-0, left panel) and C17-tolerant mutant(cesa1A1018V, right panel) seedlings were treated with 200 nM C17 for 0,1, 2, or 3 h and roots were collected and stainedwith PI. The broken cellswith brittle cell walls were visualized by the uptake of PI. Bars = 50 mm.(B) Representative confocal microscopy images of 4-d-old wild-type andje5 mutant seedlings (with a weak allele of CESA3) stained with PI. Bar =50 mm.(C) Root growth of 4-d-old wild-type (Col-0) and C17-tolerant mutant(cesa1A108V) seedlings in the presence of 200 nM C17. Data representmean6 SD (n > 5). Statistically significant differences compared with wild-type plants are indicated; *P value < 0.01 (two-tailed Student’s t test).
2280 The Plant Cell
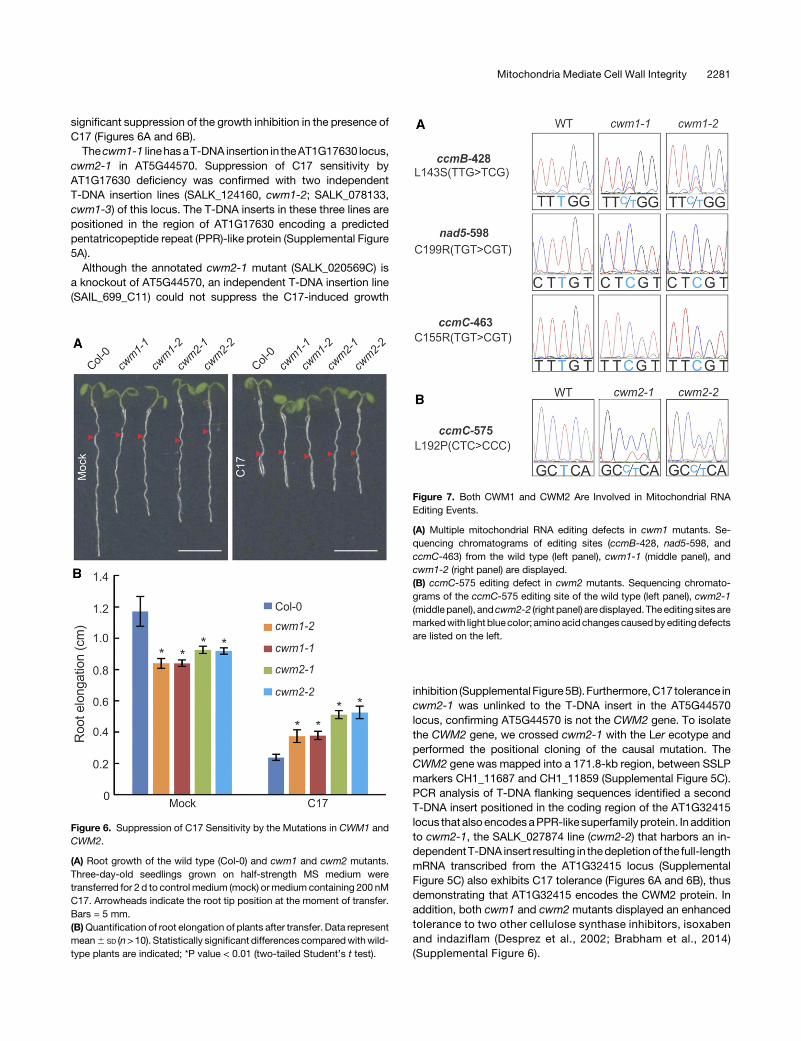
significant suppression of the growth inhibition in the presence ofC17 (Figures 6A and 6B).
Thecwm1-1 linehasaT-DNA insertion in theAT1G17630 locus,cwm2-1 in AT5G44570. Suppression of C17 sensitivity byAT1G17630 deficiency was confirmed with two independentT-DNA insertion lines (SALK_124160, cwm1-2; SALK_078133,cwm1-3) of this locus. The T-DNA inserts in these three lines arepositioned in the region of AT1G17630 encoding a predictedpentatricopeptide repeat (PPR)-like protein (Supplemental Figure5A).
Although the annotated cwm2-1 mutant (SALK_020569C) isa knockout of AT5G44570, an independent T-DNA insertion line(SAIL_699_C11) could not suppress the C17-induced growth
inhibition (Supplemental Figure5B).Furthermore,C17 tolerance incwm2-1 was unlinked to the T-DNA insert in the AT5G44570locus, confirming AT5G44570 is not the CWM2 gene. To isolatethe CWM2 gene, we crossed cwm2-1 with the Ler ecotype andperformed the positional cloning of the causal mutation. TheCWM2 gene was mapped into a 171.8-kb region, between SSLPmarkers CH1_11687 and CH1_11859 (Supplemental Figure 5C).PCR analysis of T-DNA flanking sequences identified a secondT-DNA insert positioned in the coding region of the AT1G32415locus that alsoencodesaPPR-like superfamily protein. In additionto cwm2-1, the SALK_027874 line (cwm2-2) that harbors an in-dependentT-DNA insert resulting in thedepletionof the full-lengthmRNA transcribed from the AT1G32415 locus (SupplementalFigure 5C) also exhibits C17 tolerance (Figures 6A and 6B), thusdemonstrating that AT1G32415 encodes the CWM2 protein. Inaddition, both cwm1 and cwm2 mutants displayed an enhancedtolerance to two other cellulose synthase inhibitors, isoxabenand indaziflam (Desprez et al., 2002; Brabham et al., 2014)(Supplemental Figure 6).
Figure 6. Suppression of C17 Sensitivity by the Mutations in CWM1 andCWM2.
(A) Root growth of the wild type (Col-0) and cwm1 and cwm2 mutants.Three-day-old seedlings grown on half-strength MS medium weretransferred for 2 d to control medium (mock) ormedium containing 200 nMC17. Arrowheads indicate the root tip position at the moment of transfer.Bars = 5 mm.(B)Quantification of root elongation of plants after transfer. Data representmean6 SD (n>10). Statistically significant differences comparedwithwild-type plants are indicated; *P value < 0.01 (two-tailed Student’s t test).
Figure 7. Both CWM1 and CWM2 Are Involved in Mitochondrial RNAEditing Events.
(A) Multiple mitochondrial RNA editing defects in cwm1 mutants. Se-quencing chromatograms of editing sites (ccmB-428, nad5-598, andccmC-463) from the wild type (left panel), cwm1-1 (middle panel), andcwm1-2 (right panel) are displayed.(B) ccmC-575 editing defect in cwm2 mutants. Sequencing chromato-grams of the ccmC-575 editing site of the wild type (left panel), cwm2-1(middle panel), andcwm2-2 (right panel) aredisplayed. Theeditingsites aremarkedwith light bluecolor; aminoacidchangescausedbyeditingdefectsare listed on the left.
Mitochondria Mediate Cell Wall Integrity 2281

Both cwm1 and cwm2 Mutations AffectMitochondrial Complexes
Protein domain analysis indicated that both CWM1 and CWM2proteins contain an N-terminal mitochondrial targeting peptide,strongly suggesting that CWM1 and CWM2 are localized in themitochondria. Since PPR proteins have been found to affectthe maturation, stability, or expression of transcripts encoded inthe mitochondrial genome (Barkan and Small, 2014), the mito-chondrial transcriptome of the cwm1 and cwm2 mutants wassequenced. Among the transcripts from cwm1, editing defectswere found in the cytochrome c maturation protein B (ccmB),ccmC, and NADH dehydrogenase subunit5 (nad5) sequences(Figure 7A), while an editing defect in the ccmC transcript wasidentified in cwm2 (Figure 7B). nad5 encodes a subunit of NADHdehydrogenase in mitochondrial complex I, whereas both ccmBand ccmC are required for the maturation of cytochrome c (Kranzet al., 2009; Millar et al., 2011). Consistent with this, a dramatic
decrease in cytochrome c content was observed in all cwmmutants,whereas the levelof theubiquinoneoxidoreductaseFe-Sprotein4 (NDUFS4, subunit of complex I) was affected only in thecwm1 mutants (Figures 8A and 8B). Three proteins were addi-tionally found to display altered abundances in all cwm mutants,with cytochrome oxidase subunit II (COXII, subunit of complex IV)and alternative oxidase (AOX) being detected at higher levels, andthe Rieske iron-sulfur protein (RISP, subunit of complex III) beingreduced (Figures 8A and 8B). No significant change in proteinabundance was observed for the beta subunit of ATP synthase(ATPb, subunit of complex V) and three tested mitochondrialimport inner membrane translocase subunits (Tim9, Tim17, andTim23) (Figures 8A and 8B).To determine the abundance of assembled respiratory com-
plexes, total membrane proteins were resolved by blue nativepolyacrylamide gel electrophoresis, and specific complexeswere detected by probing immunoblots with antibodies againstNDUFS4 (complex I), RISP (complex III), COXII (complex IV), and
Figure 8. Deficiency of CWM1 and CWM2 Perturbs Mitochondrial Function.
(A) Mitochondrial protein content was quantified using antibodies. Antibodies used detect ATP synthase (Atpb, subunit of complex V), ubiquinone ox-idoreductase Fe-S protein4 (NDUFS4, subunit of complex I), Rieske iron-sulfur protein (RISP, subunit of complex III), cytochrome oxidase subunit II (COXII,subunit of complex IV), cytochrome c (Cyt c), AOX, and mitochondrial import inner membrane translocase subunits (Tim9, Tim17, and Tim23).(B) Image of total mitochondrial proteins of the wild type, cwm1mutants (cwm1-1 and cwm1-2), and cwm2mutants (cwm2-1 and cwm2-2). Two or fourmicrograms of mitochondrial proteins were separated with SDS-PAGE and stained with Coomassie blue.(C)Mitochondrial complexes in thewild type,cwm1mutants (cwm1-1andcwm1-2), andcwm2mutants (cwm2-1andcwm2-2).Mitochondrialproteinswereseparated with blue native polyacrylamide gel electrophoresis. Atpb antibody was used to visualize complex V, RISP for complex III, COXII for complex IV,and NDUFS4 for complex I. The identities of protein complexes are indicated on the left or right of the blots: I, complex I; IV, complex IV; V, complex V; III2,dimeric complex III; I+III2, supercomplex composed of complex I and dimeric complex III; I2+III4, a dimer of supercomplex I+III2.
2282 The Plant Cell
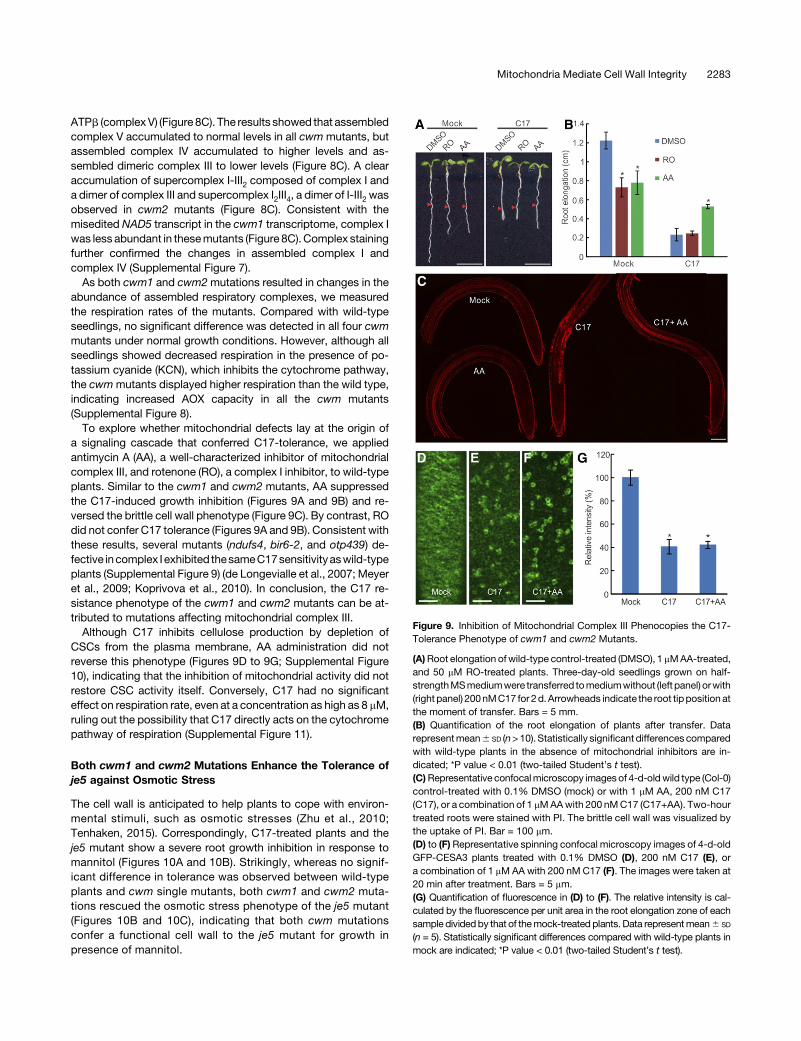
ATPb (complex V) (Figure 8C). The results showed that assembledcomplex V accumulated to normal levels in all cwm mutants, butassembled complex IV accumulated to higher levels and as-sembled dimeric complex III to lower levels (Figure 8C). A clearaccumulation of supercomplex I-III2 composed of complex I anda dimer of complex III and supercomplex I2III4, a dimer of I-III2 wasobserved in cwm2 mutants (Figure 8C). Consistent with themisedited NAD5 transcript in the cwm1 transcriptome, complex Iwas less abundant in thesemutants (Figure 8C). Complex stainingfurther confirmed the changes in assembled complex I andcomplex IV (Supplemental Figure 7).
As both cwm1 and cwm2mutations resulted in changes in theabundance of assembled respiratory complexes, we measuredthe respiration rates of the mutants. Compared with wild-typeseedlings, no significant difference was detected in all four cwmmutants under normal growth conditions. However, although allseedlings showed decreased respiration in the presence of po-tassium cyanide (KCN), which inhibits the cytochrome pathway,the cwmmutants displayed higher respiration than the wild type,indicating increased AOX capacity in all the cwm mutants(Supplemental Figure 8).
To explore whether mitochondrial defects lay at the origin ofa signaling cascade that conferred C17-tolerance, we appliedantimycin A (AA), a well-characterized inhibitor of mitochondrialcomplex III, and rotenone (RO), a complex I inhibitor, to wild-typeplants. Similar to the cwm1 and cwm2 mutants, AA suppressedthe C17-induced growth inhibition (Figures 9A and 9B) and re-versed the brittle cell wall phenotype (Figure 9C). By contrast, ROdid not confer C17 tolerance (Figures 9A and 9B). Consistent withthese results, several mutants (ndufs4, bir6-2, and otp439) de-fective incomplex I exhibited thesameC17sensitivityaswild-typeplants (Supplemental Figure 9) (de Longevialle et al., 2007; Meyeret al., 2009; Koprivova et al., 2010). In conclusion, the C17 re-sistance phenotype of the cwm1 and cwm2 mutants can be at-tributed to mutations affecting mitochondrial complex III.
Although C17 inhibits cellulose production by depletion ofCSCs from the plasma membrane, AA administration did notreverse this phenotype (Figures 9D to 9G; Supplemental Figure10), indicating that the inhibition of mitochondrial activity did notrestore CSC activity itself. Conversely, C17 had no significanteffect on respiration rate, even at a concentration as high as 8 mM,ruling out the possibility that C17 directly acts on the cytochromepathway of respiration (Supplemental Figure 11).
Both cwm1 and cwm2 Mutations Enhance the Tolerance ofje5 against Osmotic Stress
The cell wall is anticipated to help plants to cope with environ-mental stimuli, such as osmotic stresses (Zhu et al., 2010;Tenhaken, 2015). Correspondingly, C17-treated plants and theje5 mutant show a severe root growth inhibition in response tomannitol (Figures 10A and 10B). Strikingly, whereas no signif-icant difference in tolerance was observed between wild-typeplants and cwm single mutants, both cwm1 and cwm2 muta-tions rescued the osmotic stress phenotype of the je5 mutant(Figures 10B and 10C), indicating that both cwm mutationsconfer a functional cell wall to the je5 mutant for growth inpresence of mannitol.
Figure 9. Inhibition of Mitochondrial Complex III Phenocopies the C17-Tolerance Phenotype of cwm1 and cwm2 Mutants.
(A)Root elongation of wild-type control-treated (DMSO), 1 mMAA-treated,and 50 mM RO-treated plants. Three-day-old seedlings grown on half-strengthMSmediumwere transferred tomediumwithout (left panel) orwith(right panel) 200nMC17 for 2d.Arrowheads indicate the root tip position atthe moment of transfer. Bars = 5 mm.(B) Quantification of the root elongation of plants after transfer. Datarepresentmean6 SD (n>10). Statistically significant differences comparedwith wild-type plants in the absence of mitochondrial inhibitors are in-dicated; *P value < 0.01 (two-tailed Student’s t test).(C)Representative confocalmicroscopy imagesof 4-d-oldwild type (Col-0)control-treated with 0.1% DMSO (mock) or with 1 mM AA, 200 nM C17(C17), or a combination of 1 mMAAwith 200 nMC17 (C17+AA). Two-hourtreated roots were stained with PI. The brittle cell wall was visualized bythe uptake of PI. Bar = 100 mm.(D) to (F) Representative spinning confocal microscopy images of 4-d-oldGFP-CESA3 plants treated with 0.1% DMSO (D), 200 nM C17 (E), ora combination of 1 mM AA with 200 nM C17 (F). The images were taken at20 min after treatment. Bars = 5 mm.(G) Quantification of fluorescence in (D) to (F). The relative intensity is cal-culated by the fluorescence per unit area in the root elongation zone of eachsample divided by that of themock-treated plants. Data representmean6 SD
(n = 5). Statistically significant differences compared with wild-type plants inmock are indicated; *P value < 0.01 (two-tailed Student’s t test).
Mitochondria Mediate Cell Wall Integrity 2283
The C17 Tolerance Triggered by Dysfunctional MitochondriaPartially Depends on ANAC017
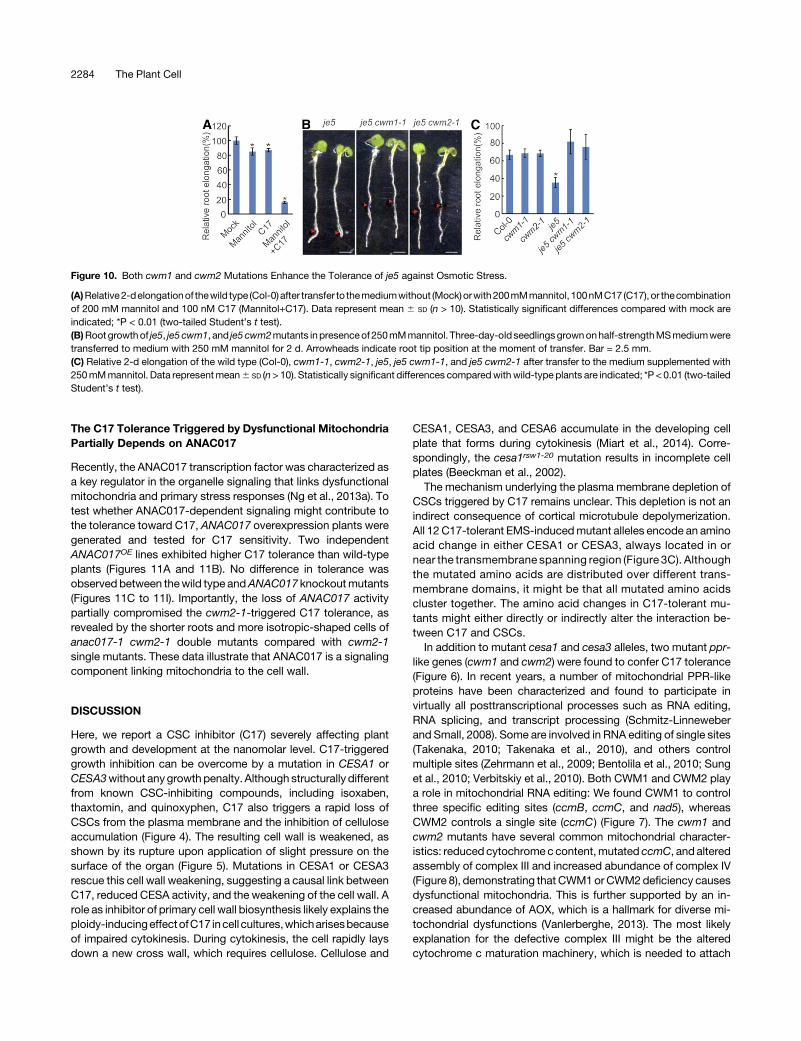
Recently, the ANAC017 transcription factor was characterized asa key regulator in the organelle signaling that links dysfunctionalmitochondria and primary stress responses (Ng et al., 2013a). Totest whether ANAC017-dependent signaling might contribute tothe tolerance toward C17, ANAC017 overexpression plants weregenerated and tested for C17 sensitivity. Two independentANAC017OE lines exhibited higher C17 tolerance than wild-typeplants (Figures 11A and 11B). No difference in tolerance wasobserved between thewild type andANAC017 knockoutmutants(Figures 11C to 11I). Importantly, the loss of ANAC017 activitypartially compromised the cwm2-1-triggered C17 tolerance, asrevealed by the shorter roots and more isotropic-shaped cells ofanac017-1 cwm2-1 double mutants compared with cwm2-1single mutants. These data illustrate that ANAC017 is a signalingcomponent linking mitochondria to the cell wall.
DISCUSSION
Here, we report a CSC inhibitor (C17) severely affecting plantgrowth and development at the nanomolar level. C17-triggeredgrowth inhibition can be overcome by a mutation in CESA1 orCESA3without anygrowth penalty. Although structurally differentfrom known CSC-inhibiting compounds, including isoxaben,thaxtomin, and quinoxyphen, C17 also triggers a rapid loss ofCSCs from the plasma membrane and the inhibition of celluloseaccumulation (Figure 4). The resulting cell wall is weakened, asshown by its rupture upon application of slight pressure on thesurface of the organ (Figure 5). Mutations in CESA1 or CESA3rescue this cell wall weakening, suggesting a causal link betweenC17, reduced CESA activity, and the weakening of the cell wall. Arole as inhibitor of primary cell wall biosynthesis likely explains theploidy-inducingeffectofC17 incell cultures,whicharisesbecauseof impaired cytokinesis. During cytokinesis, the cell rapidly laysdown a new cross wall, which requires cellulose. Cellulose and
CESA1, CESA3, and CESA6 accumulate in the developing cellplate that forms during cytokinesis (Miart et al., 2014). Corre-spondingly, the cesa1rsw1-20 mutation results in incomplete cellplates (Beeckman et al., 2002).The mechanism underlying the plasma membrane depletion of
CSCs triggered by C17 remains unclear. This depletion is not anindirect consequence of cortical microtubule depolymerization.All 12 C17-tolerant EMS-inducedmutant alleles encode an aminoacid change in either CESA1 or CESA3, always located in ornear the transmembrane spanning region (Figure 3C). Althoughthe mutated amino acids are distributed over different trans-membrane domains, it might be that all mutated amino acidscluster together. The amino acid changes in C17-tolerant mu-tants might either directly or indirectly alter the interaction be-tween C17 and CSCs.In addition to mutant cesa1 and cesa3 alleles, two mutant ppr-
like genes (cwm1 and cwm2) were found to confer C17 tolerance(Figure 6). In recent years, a number of mitochondrial PPR-likeproteins have been characterized and found to participate invirtually all posttranscriptional processes such as RNA editing,RNA splicing, and transcript processing (Schmitz-Linneweberand Small, 2008). Some are involved in RNA editing of single sites(Takenaka, 2010; Takenaka et al., 2010), and others controlmultiple sites (Zehrmann et al., 2009; Bentolila et al., 2010; Sunget al., 2010; Verbitskiy et al., 2010). Both CWM1 and CWM2 playa role in mitochondrial RNA editing: We found CWM1 to controlthree specific editing sites (ccmB, ccmC, and nad5), whereasCWM2 controls a single site (ccmC) (Figure 7). The cwm1 andcwm2 mutants have several common mitochondrial character-istics: reduced cytochrome c content,mutated ccmC, and alteredassembly of complex III and increased abundance of complex IV(Figure 8), demonstrating that CWM1orCWM2deficiency causesdysfunctional mitochondria. This is further supported by an in-creased abundance of AOX, which is a hallmark for diverse mi-tochondrial dysfunctions (Vanlerberghe, 2013). The most likelyexplanation for the defective complex III might be the alteredcytochrome c maturation machinery, which is needed to attach
Figure 10. Both cwm1 and cwm2 Mutations Enhance the Tolerance of je5 against Osmotic Stress.
(A)Relative2-delongationof thewild type (Col-0) after transfer to themediumwithout (Mock)orwith200mMmannitol, 100nMC17 (C17), or thecombinationof 200 mM mannitol and 100 nM C17 (Mannitol+C17). Data represent mean 6 SD (n > 10). Statistically significant differences compared with mock areindicated; *P < 0.01 (two-tailed Student’s t test).(B)Rootgrowthof je5, je5cwm1, and je5cwm2mutants inpresenceof 250mMmannitol. Three-day-old seedlingsgrownonhalf-strengthMSmediumweretransferred to medium with 250 mM mannitol for 2 d. Arrowheads indicate root tip position at the moment of transfer. Bar = 2.5 mm.(C) Relative 2-d elongation of the wild type (Col-0), cwm1-1, cwm2-1, je5, je5 cwm1-1, and je5 cwm2-1 after transfer to the medium supplemented with250mMmannitol. Data representmean6 SD (n> 10). Statistically significant differences comparedwithwild-type plants are indicated; *P < 0.01 (two-tailedStudent’s t test).
2284 The Plant Cell
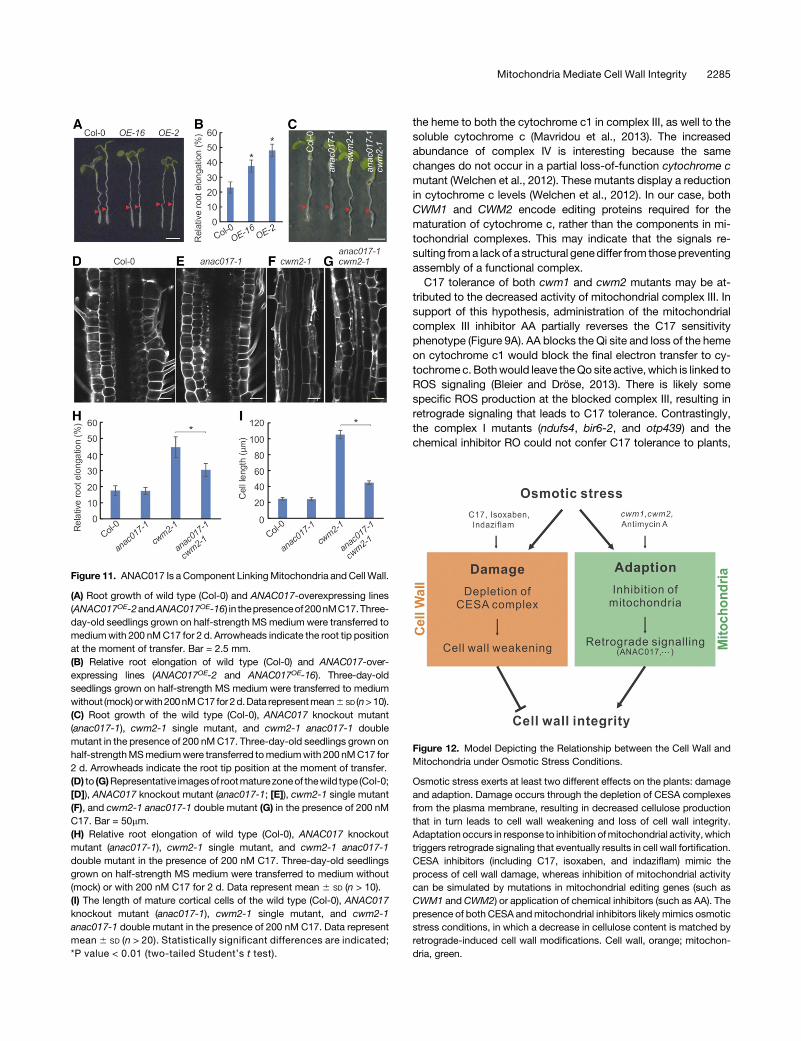
the heme to both the cytochrome c1 in complex III, as well to thesoluble cytochrome c (Mavridou et al., 2013). The increasedabundance of complex IV is interesting because the samechanges do not occur in a partial loss-of-function cytochrome cmutant (Welchen et al., 2012). These mutants display a reductionin cytochrome c levels (Welchen et al., 2012). In our case, bothCWM1 and CWM2 encode editing proteins required for thematuration of cytochrome c, rather than the components in mi-tochondrial complexes. This may indicate that the signals re-sulting froma lack of a structural genediffer from thosepreventingassembly of a functional complex.C17 tolerance of both cwm1 and cwm2 mutants may be at-
tributed to the decreased activity of mitochondrial complex III. Insupport of this hypothesis, administration of the mitochondrialcomplex III inhibitor AA partially reverses the C17 sensitivityphenotype (Figure 9A). AA blocks the Qi site and loss of the hemeon cytochrome c1 would block the final electron transfer to cy-tochrome c. Bothwould leave theQo site active, which is linked toROS signaling (Bleier and Dröse, 2013). There is likely somespecific ROS production at the blocked complex III, resulting inretrograde signaling that leads to C17 tolerance. Contrastingly,the complex I mutants (ndufs4, bir6-2, and otp439) and thechemical inhibitor RO could not confer C17 tolerance to plants,
Figure 12. Model Depicting the Relationship between the Cell Wall andMitochondria under Osmotic Stress Conditions.
Osmotic stress exerts at least two different effects on the plants: damageand adaption. Damage occurs through the depletion of CESA complexesfrom the plasma membrane, resulting in decreased cellulose productionthat in turn leads to cell wall weakening and loss of cell wall integrity.Adaptation occurs in response to inhibition ofmitochondrial activity, whichtriggers retrograde signaling that eventually results in cell wall fortification.CESA inhibitors (including C17, isoxaben, and indaziflam) mimic theprocess of cell wall damage, whereas inhibition of mitochondrial activitycan be simulated by mutations in mitochondrial editing genes (such asCWM1 and CWM2) or application of chemical inhibitors (such as AA). Thepresence of both CESA andmitochondrial inhibitors likely mimics osmoticstress conditions, in which a decrease in cellulose content is matched byretrograde-induced cell wall modifications. Cell wall, orange; mitochon-dria, green.
Figure 11. ANAC017 Is a Component LinkingMitochondria and Cell Wall.
(A) Root growth of wild type (Col-0) and ANAC017-overexpressing lines(ANAC017OE-2andANAC017OE-16) in thepresenceof200nMC17.Three-day-old seedlings grown on half-strength MSmedium were transferred tomediumwith 200 nMC17 for 2 d. Arrowheads indicate the root tip positionat the moment of transfer. Bar = 2.5 mm.(B) Relative root elongation of wild type (Col-0) and ANAC017-over-expressing lines (ANAC017OE-2 and ANAC017OE-16). Three-day-oldseedlings grown on half-strength MS medium were transferred to mediumwithout (mock)orwith200nMC17 for 2d.Data representmean6 SD (n>10).(C) Root growth of the wild type (Col-0), ANAC017 knockout mutant(anac017-1), cwm2-1 single mutant, and cwm2-1 anac017-1 doublemutant in the presence of 200 nMC17. Three-day-old seedlings grown onhalf-strengthMSmediumwere transferred tomediumwith 200 nMC17 for2 d. Arrowheads indicate the root tip position at the moment of transfer.(D) to (G)Representative imagesof rootmaturezoneof thewild type (Col-0;[D]), ANAC017 knockout mutant (anac017-1; [E]), cwm2-1 single mutant(F), and cwm2-1 anac017-1 double mutant (G) in the presence of 200 nMC17. Bar = 50mm.(H) Relative root elongation of wild type (Col-0), ANAC017 knockoutmutant (anac017-1), cwm2-1 single mutant, and cwm2-1 anac017-1double mutant in the presence of 200 nM C17. Three-day-old seedlingsgrown on half-strength MS medium were transferred to medium without(mock) or with 200 nM C17 for 2 d. Data represent mean 6 SD (n > 10).(I) The length of mature cortical cells of the wild type (Col-0), ANAC017knockout mutant (anac017-1), cwm2-1 single mutant, and cwm2-1anac017-1 double mutant in the presence of 200 nM C17. Data representmean 6 SD (n > 20). Statistically significant differences are indicated;*P value < 0.01 (two-tailed Student’s t test).
Mitochondria Mediate Cell Wall Integrity 2285

suggesting that a defective mitochondrial complex I in cwm1mutants is not linked to C17 tolerance. These observations ex-clude the possibility that the alleviated deleterious effects in cwmmutants are caused by reduced growth as a result of reducedrespiration with dysfunctional mitochondria. Although we cannotrule out the possibility that C17 acts on mitochondria, it seemsunlikely because a high dosage of C17 did not cause inhibition ofthe cytochrome pathway of respiration (Supplemental Figure 11).Additionally, the increased tolerance of cwm1 and cwm2 towardtwoothercellulosesynthase inhibitorsemphasizes thatcwm1andcwm2 mutations counteract the growth inhibition induced bycellulose deficiency rather than other effects triggered by C17.
Perturbing mitochondria alters nuclear gene expression viaorganelle signaling regulation (Rhoads and Subbaiah, 2007; Nget al., 2014). Organelle signaling is active in the cwm1 and cwm2mutants, as marked by AOX accumulation, and might mediatea crosstalk between the mitochondria and the cell wall. This hy-pothesis is supported by the observation that ANAC017 over-expression confers C17 tolerance, whereas ANAC017 depletionresults in a partial loss of the cwm2-1-triggered C17 tolerancephenotype (Figure 11). ANAC017 is the core cellular component inmitochondrial organelle signaling, particularly for retrogradesignaling, accounting for more than 85% of H2O2-mediated pri-mary stress responses in plants (Ng et al., 2013a). BecauseANAC017 is latent in theendoplasmic reticulum (ER)andactivatedby mitochondrial dysfunction (Ng et al., 2013a), C17 toleranceupon inhibition of mitochondrial activity might find its origin in theER; all the cell wall-synthesizing enzymes are transported via theER. ANAC017 deficiency only partially abolished the C17 toler-ance triggered by cwm2-1mutation. It is possible that ANAC017functions redundantly with another ER-bound NAC transcriptionfactor, such as ANAC013 (De Clercq et al., 2013) or ANAC017-independent organelle signaling cascades. Indeed, at least twoother stress-responsive systems in the ER are known, includingthe activation of bZIP transcription factors by the S1P and S2Pproteases and the IRE1 and IRE2 splicing systems (Deng et al.,2013; Howell, 2013; Srivastava et al., 2013).
Our data suggest that the inhibition ofmitochondria contributesto the maintenance of a functional cell wall under osmotic stressconditions. In agreement, CWM1 or CWM2 loss of function incellulose-deficient plants increased the tolerance to osmoticstresses (Figure 10), indicating that the cell wall in these plantscopes better with stress. Enhanced osmotic tolerance was ob-servedonly inacellulose-deficientbackground, indicating that theconnection between mitochondrial function and cell wall ismasked in wild-type plants. Alternatively, regarding the fact thatosmotic stress naturally results in decreased cellulose productionby depletion of membrane-associated CESAs (Crowell et al.,2009), these changes might be dependent on the cell wall status.Taken together, our results suggest that perturbingmitochondrialactivity results in attenuationof thegrowth inhibition that iscausedby CESA depletion from the plasma membrane, likely throughachange incellwall composition.Moreover, in viewof the fact thatmitochondrial activity is inhibited under osmotic stresses (Skiryczet al., 2010;Vanderauweraetal., 2012), ourdatapoint toapossiblemechanism underlying adaptation to osmotic stress, in which themodulation of mitochondrial activity and subsequent retrogradesignaling controls themaintenanceof cellwall integrity (Figure 12).
METHODS
Plant Materials and Growth Conditions
Arabidopsis thaliana plants were grown under long-day conditions (16 hof light/8 h of darkness) at 22°C on half-strength Murashige and Skoog(MS) germination medium (Murashige and Skoog, 1962). The cwm1-1(SALK_017325C), cwm1-2 (SALK_124160), cwm1-3 (SALK_078133),cwm2-1 (SALK_020569C), cwm2-2 (SALK_027874), and SAIL_699_C11were acquired from the ABRC. The otp439, bir6-2, ndufs4, je5, andanac017-1 mutants, the GFP-CESA3 line in the je5 background, and theGFP-CESA3mCherry-MBDdouble fluorescence line havebeendescribedpreviously (de Longevialle et al., 2007; Desprez et al., 2007; Meyer et al.,2009; Koprivova et al., 2010; Ng et al., 2013a; Miart et al., 2014).
Chemical Treatments
AA and RO were purchased from Sigma-Aldrich and applied from 50 mMstock solutions in DMSO to the final concentration described in the text.Mock treatments of 0.1% (v/v) DMSO were used as a control. For thetreatments of chemical compounds, plants were grown on the controlmedium for three days and then transferred to medium without or with theindicated drugs for 2d. The2-d elongation of seedling roots after treatmentwas measured.
Generation of Transgenic Arabidopsis Plants
Overexpressing plants were generated by cloning the open reading frameofANAC017 into pK7WG2D (Karimi et al., 2002). Cloning primers are listedin Supplemental Table 1. Constructs were transformed into ArabidopsisCol-0 by Agrobacterium tumefaciens-mediated floral dipping (Clough andBent, 1998).
Chemical Screen
The PSB-L_H2B-YFP cell line was obtained through cocultivation of theArabidopsis PSB-L cell suspension culture (Landsberg erecta) (May andLeaver, 1993) with an Agrobacterium strain carrying a binary T-DNA vectorcontaining a transgene encoding a translational fusion between Arabi-dopsis Histone 2B (AT5G22880) and Yellow Fluorescent Protein (H2B-YFP) (Boisnard-Lorig et al., 2001). Protoplasts were isolated from 100 mL3-d-old Arabidopsis PSB-L_H2B-YFP cell suspension culture expressingthe H2B-YFP transgene. Cells were harvested through sedimentation in2 3 50 mL tubes (Corning) and removing the supernatants. After cell walldigestionwith 100mLcellulase (15mgmL21; YakultOnozukaR10) in 0.4Mmannitol, 5 mM MES, pH 5.7, for 3 to 4 h at 25°C, 50 rpm in a large (142/20 mm) vented Petri dish (Greiner), isolated cells were transferred to 2 3
50mL tubes (Corning) andwashedwith and resuspended in 25mLMSMOcontaining 0.35Mmannitol, 0.5mgL21NAA, and0.05mgL21 kinetin usingcentrifugation at 145g. Protoplasts were then filtered through a 40-mmmesh and incubated overnight in the dark at room temperature for partialcell wall regeneration in MSMO containing 0.35 M mannitol, 0.5 mg L21
NAA, and 0.05 mg L21 kinetin. Dead cells were removed by sucrosegradient (0.5 M) decantation at 100g during 10 min. Subsequently, theprotoplasts were washed in 23 40 mL and resuspended in 23 2.5-5 mLMSMO containing 0.35 M mannitol, 0.5 mg L21 NAA, and 0.05 mg L21
kinetin in 23 50 mL tubes. Cell density was estimated with a cell counter(Countess Automated Cell Counter; Invitrogen), and subsequently dilutedto obtain a standard density of 105 cells mL21. Diluted cells were thenseeded inaNunc96-wellCC2CoverglassBottomPlate (ThermoScientific)at 100 mL per well using a Biomek 2000 robot (Beckman Coulter). Sub-sequently, compounds of the DIVERSet library (12,000 compounds) wereadded with a Te-MO robot (Tecan) to a final concentration of 50 mM (1%[v/v] DMSO). Plates were shaken for 3 min to homogenize the medium.
2286 The Plant Cell

Imagesof theplatewereacquiredusing theScan^Rscreeningstation fromOlympus. This platform includes an epifluorescencemicroscope IX81witha MT20 illumination system, an automated motorized stage and a CCDcamera. YFP fluorescence was recorded using the fluorescein iso-thiocyanate parameters (YFP FITC, DAPI/FITC/TxRED filter cube, fluo-rescence illumination, FITC 492/18 excitation filter), the UPLSAPO 103objective and an exposure time of 15 ms. Time-lapse movies were re-corded with a resolution of 30 min between the pictures for up to 120 h.Because cells need between 60 and 90min to divide in optimal conditions,two to three imageswereobtainedcapturing themitoticfiguresofeachcell.This time resolution allowed the acquisition of four pictures for each of the96 wells, covering ;10% of the well surfaces.
Screen of C17-Tolerant Mutants and Map-Based Cloning
To obtain mutants tolerant to C17, seeds from an EMS-treated seedcollectionwithCol-0backgroundwereplatedonhalf-strengthMSmediumcontaining 2mMC17. After growing for 7 d under long-day conditions (16 hof light/8 h of darkness) at 22°C, plants with a long root were identified asC17- tolerant mutants. From 300,000 independent EMS-mutagenizedseeds (divided over 20 pools), 22 mutants were isolated (from 12 in-dependent pools). To define the mutations underlying C17 tolerance, allC17- tolerant mutants were backcrossed with wild-type (Col-0) plants toobtainF1progenies.F2progeniesfromself-pollinatedF1plantsweregrowninthepresenceof 2mMC17 for 7 d, allowing calculation of the segregation ratioof C17 tolerance within the F2 progenies. All mutants were used to generatemapping populations through crossing with another ecotype (Ler-0). SSLPmarkers (Supplemental Table 1)wereused tomap thepositionof themutatedgenes in the Arabidopsis genome. Subsequently, the mutant genes wereidentified through candidate gene sequencing.
For the cwm mutant screen, seeds of a set of confirmed T-DNA lines(CS27941) were germinated on half-strength MS medium. Three-day-oldseedlings were transferred to medium with a low dose of C17 (200 nM),a concentration at which root growth was inhibited instantly upon transferto C17-containing medium. Two insertion lines (SALK_017325C andSALK_020569C) were obtained with significant suppression of the growthinhibition in presence of C17. Because the T-DNA insert within theAT5G44570 locus in SALK_020569C was not linked with the C17-tolerantphenotype, SALK_020569C was crossed with the Ler-0 ecotype to gen-erate a mapping population. SSLP markers (Supplemental Table 1) wereused tomap the position of themutated genes in the Arabidopsis genome.Subsequently, using PCR, a second T-DNA insert was found positioned atthe coding region of AT1G32415.
Cellulose Measurement
The analysis of glucose content was performed on an AIR (alcohol-insoluble residue) prepared as follows. One hundred milligrams (freshweight) of ground 4-d-old dark-grown seedlings were washed twice in4 volumes of absolute ethanol for 15 min, then rinsed twice in 4 volumes ofacetone at room temperature for 10 min and left to dry under a fume hoodovernight at room temperature. AIR was then submitted to hydrolysis in2.5 M trifluoroacetic acid for 1.5 h at 100°C. To determine the cellulosecontent, the residual pellet obtained after the trifluoroacetic acid hydrolysiswas rinsed twice with 10 volumes of water and hydrolyzed with H2SO4 asdescribed previously (Updegraff, 1969). The released glucose was diluted500 times and then quantified using a high-performance anion exchangechromatographywithpulsedamperometricdetectionasdescribedpreviously(Harholt et al., 2006). Quantification was done with biological quadruplicatesand the average values and standard errors are indicated in the graphs.
Detection of Cell Wall Weakening
For the detection of cell wall weakening, 3-d-old seedlings grown on half-strength MS medium were transferred to liquid medium without or with
200 nM C17. The root tips were stained with 10 mg mL21 PI for 3 min. Thestained root tips were put on the Nunc Lab-Tek Chambered Coverglass(catalog no. 155361) without pressure or on a microscope slide with a coverslip that gently exerted pressure. Using confocal laser scanning microscopy(LSM710; Zeiss), the brittle cells could be visualized by the uptake of PI.
Spinning-Disk Microscopy and Image Analysis
For live-cell imaging, hypocotyls of 3-d-old etiolated je5 seedlings har-boring theGFP-CESA3marker were treatedwithout or with 50 mMC17 for3 h were analyzed on an Axiovert 200Mmicroscope (Zeiss) equipped witha Yokogawa CSU22 spinning disk, Zeiss 100/1.4 numerical aperture oilobjective, and Andor EMCCD iXon DU 895 camera (Plateforme d’ImagerieDynamique, Institut Pasteur, Paris, France). A 488-nm diode-pumpedsolid-state laser was used for excitation, and emissionwas collected usingband-pass 488/25 for GFP. For root tip observations, 3-d-old light-grownseedlings were treated without or with the indicated drugs and the changeof GFP were observed with a time-series scanning from 0 to 20 min.
Isolation of Mitochondria from Hydroponic Cultures
Mitochondria were isolated from 14-d-old hydroponically grown Arabi-dopsis plants as previously described (Millar et al., 2001) with slightmodifications. Plant material was homogenized in grinding buffer (0.3 Msucrose, 25 mM tetrasodium pyrophosphate, 1% [w/v] PVP-40, 2 mMEDTA, 10 mM KH2PO4, 1% [w/v] BSA, and 20 mM ascorbic acid, pH 7.5)using mortar and pestle for 2 to 5 min twice. The homogenate was filteredthrough four layers of Miracloth and centrifuged at 2500g for 5 min; theresulting supernatant was then centrifuged at 14,000g for 20 min. Theresulting pellet was resuspended in sucrosewashmedium (0.3M sucrose,0.1% [w/v] BSA, and 10mM TES, pH 7.5) and carefully layered over 35mLPVP-40 gradient (30%Percoll, 0 to 4%PVP). The gradientwas centrifugedat 40,000g for 40 min. The mitochondrial band was collected and washedthree times in sucrose wash buffer without BSA by 20,000g for 20 min.
Oxygen Consumption of Whole Seedlings and Isolated Mitochondria
Oxygen consumption of 6-d-old Arabidopsis seedlings wasmeasured usinga computer-controlled Clark-type O2 electrode (Hansatech Instruments). Allreactionswereperformedat25°Cusing2mLofwhole tissue reactionmedium(10mMHEPES, 10mMMES, and2mMCaCl2, pH7.2) and 40 to 60mgplantmaterial. To investigate AOX-dependent respiration rate, 1 mM KCN wasadded. To inhibit AOX activity, 2 mM SHAM was added.
To measure oxygen uptake of isolated mitochondria, 1 mL reaction me-dium (0.3Msucrose,10mMTES,10mMNaCl, 4mMMgSO4,and0.1%[w/v]BSA, pH7.2) togetherwith 50mgmitochondria proteinwasused.Then, 5mMsuccinate and 1 mM NADHwere added. To investigate the effect of C17 onisolated mitochondria, concentrations ranging from 1 to 8 mMwere added tomitochondria using succinate and NADH as substrates and compared withthe effect of 100 mM KCN as a cytochrome pathway inhibitor.
RNA Editing Analysis
For analysis of RNA editing, total RNA was isolated from the root tips (3 to5mm)of5-d-oldseedlingsusinganRNeasyplantmini kit (Qiagen)and treatedwith DNase I (Invitrogen). DNA-free RNA (2 mg) was reverse transcribed andsequences including the editing sites were amplified by PCR. Primers toamplify themitochondrial transcripts are describedpreviously (Bentolila et al.,2013). The RT-PCR products were sequenced immediately.
Isolation of Mitochondria and Immunoblots
For analysis of mitochondrial protein content through immunoblotting,mitochondria were isolated from 2-week-old hydroponically grown
Mitochondria Mediate Cell Wall Integrity 2287

Arabidopsis seedlings, according to the method described previously(Murcha and Whelan, 2015). Mitochondrial proteins were separated bySDS-PAGE (Bio-Rad) or blue-native gels as described previously (Eubelet al., 2005), followed by transfer to Hybond-C extra nitrocellulose (Bio-Rad). Immunodetections were performed as described previously (Wanget al., 2012). To ensure linearity of detection, two dilutions of mitochondriawere loaded. Antibodies used were raised against Ndufs4 (Meyer et al.,2009), AOX (Elthon et al., 1989), Tim9 (Wang et al., 2012), Tim17 (Wanget al., 2012), RISP (Duncan et al., 2011), and Tim23 (Wang et al., 2012). Theantibodies against to Atpb (catalog number AS05 085), Cyt c (AS08 343A),and COXII (AS04 0543A) were obtained from Agrisera.
For mitochondrial complex staining, mitochondrial proteins (20 mg)were solubilized with digitonin (5.0 g/g protein final) in digitonin extractionbuffer (30 mMHEPES. 150 mMK-Acetate, and 10% [v/v] glycerol, pH 7.4)and incubatedon ice for 20min. Thesampleswerecentrifuged for 10minat15,000g, and Serva BlueG (0.2% [v/v] final) was added to the supernatant.The samples were loaded onto NativePAGE Novex 4% to 16% Bis-Trisgels (Life Technologies). Gels were washed twice for 10 min with distilledwater and incubated in complex I staining medium (0.1 M Tris, pH 7.4,0.14 mM NADH, and 1 mg mL21 Nitro tetrazolium blue) and in peroxidasestaining medium (10 mM phosphate buffer, pH 6.0, 20 mM guaiacol, and0.03%H2O2). After 2 to 3hof staining, gelswere transferred toCoomassie-colloidal fixing solution (40% methanol and 10% acetic acid) to stop thereactions.
Accession Numbers
Sequence data from this article can be found in the Arabidopsis GenomeInitiative or GenBank/EMBL databases under the following acces-sion numbers: CWM1 (AT1G17630), CWM2 (AT1G32415), CESA1(AT4G32410), CESA3 (AT5G05170), ANAC017 (AT1G34190), andSLO2 (AT2G13600).
Supplemental Data
Supplemental Figure 1. Rough map position on the Arabidopsisgenome of the mutated genes rendering C17 tolerance.
Supplemental Figure 2. Sequence alignment of CESA1 and CESA3of several plant species.
Supplemental Figure 3. C17 does not trigger microtubule polymer-ization.
Supplemental Figure 4. C17 results in the depletion of CSCs from theroot plasma membrane.
Supplemental Figure 5. Isolation of cwm1 and cwm2 mutants.
Supplemental Figure 6. Both cwm1-1 and cwm2-1 mutationscounteract the growth inhibition induced by cellulose deficiency.
Supplemental Figure 7. Staining of respiratory protein complexesfrom the wild type, cwm1, and cwm2.
Supplemental Figure 8. Respiration rates of cwm1 and cwm2mutants.
Supplemental Figure 9. C17 sensitivity of the mutants with defectivemitochondrial complex I.
Supplemental Figure 10. Inhibition of mitochondrial activity did notrestore CSC activity.
Supplemental Figure 11. C17 does not directly inhibit isolatedmitochondrial respiration.
Supplemental Table 1. Primer sequences used for mapping andcloning.
Supplemental Movie 1. Time-lapse imaging of Arabidopsis H2B-YFPsuspension cells in absence of C17.
Supplemental Movie 2. Time-lapse imaging of Arabidopsis H2B-YFPsuspension cells in the presence of 50 mM C17.
ACKNOWLEDGMENTS
We thank Annick Bleys for help preparing manuscript. This work wassupported by grants from the Integrated Project AGRONOMICS, in theSixth Framework Programme of the European Commission (LSHG-CT-2006-037704), Research Foundation-Flanders (G.0236.16N), and the In-teruniversity Attraction Poles Programme (IUAP P7/29 “MARS”), initiatedby the Belgian Science Policy Office. T.C. and I.D.C. are PostdoctoralFellows of the Research Foundation-Flanders. I.D.C. is also supported byFWO travel grant 12N2415N. F.V.B. is supported by grants from theInteruniversity Attraction Poles Programme (IUAP P7/29 ‘MARS’) initiatedby theBelgianSciencePolicyOfficeandGhentUniversity (MultidisciplinaryResearch Partnership ‘Biotechnology for a Sustainable Economy’, Grant01MRB510W). A.H.M., K.B., Y.W., I.S., and J.W. were funded by the ARCCentre of Excellence Plant Energy Biology (CE140100008).
AUTHOR CONTRIBUTIONS
Z.H., J.W., H.H., and L.D.V. conceived and designed the research. Z.H.,R.V., T.C., Y.W., I.D.C., O.L., K.B., G.M., and S.V. performed the experi-ments. Z.H., I.S., A.H.M., S.V., F.V.B., J.W., H.H., and L.D.V. analyzedthe data. P.H. provided the platform for high-throughput chemical screen.Z.H. and L.D.V. wrote the article. All authors read, revised, and approvedthe article.
Received July 8, 2016; revised August 8, 2016; accepted August 19, 2016;published August 19, 2016.
REFERENCES
Barkan, A., and Small, I. (2014). Pentatricopeptide repeat proteins inplants. Annu. Rev. Plant Biol. 65: 415–442.
Bartoli, C.G., Gómez, F., Martínez, D.E., and Guiamet, J.J. (2004).Mitochondria are the main target for oxidative damage in leaves ofwheat (Triticum aestivum L.). J. Exp. Bot. 55: 1663–1669.
Beeckman, T., Przemeck, G.K.H., Stamatiou, G., Lau, R., Terryn,N., De Rycke, R., Inzé, D., and Berleth, T. (2002). Genetic com-plexity of cellulose synthase a gene function in Arabidopsis em-bryogenesis. Plant Physiol. 130: 1883–1893.
Bentolila, S., Knight, W., and Hanson, M. (2010). Natural variation inArabidopsis leads to the identification of REME1, a pentatricopeptiderepeat-DYW protein controlling the editing of mitochondrial transcripts.Plant Physiol. 154: 1966–1982.
Bentolila, S., Oh, J., Hanson, M.R., and Bukowski, R. (2013).Comprehensive high-resolution analysis of the role of an Arabi-dopsis gene family in RNA editing. PLoS Genet. 9: e1003584.
Bleier, L., and Dröse, S. (2013). Superoxide generation by complexIII: from mechanistic rationales to functional consequences. Bio-chim. Biophys. Acta 1827: 1320–1331.
Boisnard-Lorig, C., Colon-Carmona, A., Bauch, M., Hodge, S.,Doerner, P., Bancharel, E., Dumas, C., Haseloff, J., and Berger,F. (2001). Dynamic analyses of the expression of the HISTONE:YFPfusion protein in Arabidopsis show that syncytial endosperm is di-vided in mitotic domains. Plant Cell 13: 495–509.
Brabham, C., Lei, L., Gu, Y., Stork, J., Barrett, M., and DeBolt, S.(2014). Indaziflam herbicidal action: a potent cellulose biosynthesisinhibitor. Plant Physiol. 166: 1177–1185.
2288 The Plant Cell

Caño-Delgado, A., Penfield, S., Smith, C., Catley, M., and Bevan,M. (2003). Reduced cellulose synthesis invokes lignification anddefense responses in Arabidopsis thaliana. Plant J. 34: 351–362.
Caño-Delgado, A.I., Metzlaff, K., and Bevan, M.W. (2000). The eli1 mu-tation reveals a link between cell expansion and secondary cell wallformation in Arabidopsis thaliana. Development 127: 3395–3405.
Clough, S.J., and Bent, A.F. (1998). Floral dip: a simplified method forAgrobacterium-mediated transformation of Arabidopsis thaliana.Plant J. 16: 735–743.
Crowell, E.F., Bischoff, V., Desprez, T., Rolland, A., Stierhof, Y.-D.,Schumacher, K., Gonneau, M., Höfte, H., and Vernhettes, S. (2009).Pausing of Golgi bodies on microtubules regulates secretion of cellulosesynthase complexes in Arabidopsis. Plant Cell 21: 1141–1154.
De Clercq, I., et al. (2013). The membrane-bound NAC transcription factorANAC013 functions in mitochondrial retrograde regulation of the oxida-tive stress response in Arabidopsis. Plant Cell 25: 3472–3490.
de Longevialle, A.F., Meyer, E.H., Andrés, C., Taylor, N.L., Lurin, C.,Millar, A.H., and Small, I.D. (2007). The pentatricopeptide repeatgene OTP43 is required for trans-splicing of the mitochondrial nad1Intron 1 in Arabidopsis thaliana. Plant Cell 19: 3256–3265.
Deng, Y., Srivastava, R., and Howell, S.H. (2013). Endoplasmic re-ticulum (ER) stress response and its physiological roles in plants.Int. J. Mol. Sci. 14: 8188–8212.
Desprez, T., Vernhettes, S., Fagard, M., Refrégier, G., Desnos, T.,Aletti, E., Py, N., Pelletier, S., and Höfte, H. (2002). Resistanceagainst herbicide isoxaben and cellulose deficiency caused bydistinct mutations in same cellulose synthase isoform CESA6. PlantPhysiol. 128: 482–490.
Desprez, T., Juraniec, M., Crowell, E.F., Jouy, H., Pochylova, Z.,Parcy, F., Höfte, H., Gonneau, M., and Vernhettes, S. (2007).Organization of cellulose synthase complexes involved in primarycell wall synthesis in Arabidopsis thaliana. Proc. Natl. Acad. Sci.USA 104: 15572–15577.
Duncan, O., Taylor, N.L., Carrie, C., Eubel, H., Kubiszewski-Jakubiak, S., Zhang, B., Narsai, R., Millar, A.H., and Whelan, J.(2011). Multiple lines of evidence localize signaling, morphology,and lipid biosynthesis machinery to the mitochondrial outer mem-brane of Arabidopsis. Plant Physiol. 157: 1093–1113.
Ellis, C., and Turner, J.G. (2001). The Arabidopsis mutant cev1 hasconstitutively active jasmonate and ethylene signal pathways andenhanced resistance to pathogens. Plant Cell 13: 1025–1033.
Ellis, C., Karafyllidis, I., Wasternack, C., and Turner, J.G. (2002).The Arabidopsis mutant cev1 links cell wall signaling to jasmonateand ethylene responses. Plant Cell 14: 1557–1566.
Elthon, T.E., Nickels, R.L., and McIntosh, L. (1989). Monoclonalantibodies to the alternative oxidase of higher plant mitochondria.Plant Physiol. 89: 1311–1317.
Endler, A., and Persson, S. (2011). Cellulose synthases and synthesisin Arabidopsis. Mol. Plant 4: 199–211.
Eubel, H., Braun, H.-P., andMillar, A.H. (2005). Blue-native PAGE in plants:a tool in analysis of protein-protein interactions. Plant Methods 1: 11.
Fujita, M., Fujita, Y., Noutoshi, Y., Takahashi, F., Narusaka, Y.,Yamaguchi-Shinozaki, K., and Shinozaki, K. (2006). Crosstalkbetween abiotic and biotic stress responses: a current view fromthe points of convergence in the stress signaling networks. Curr.Opin. Plant Biol. 9: 436–442.
Gechev, T.S., Van Breusegem, F., Stone, J.M., Denev, I., and Laloi, C.(2006). Reactive oxygen species as signals that modulate plant stressresponses and programmed cell death. BioEssays 28: 1091–1101.
Giraud, E., Van Aken, O., Ho, L.H.M., and Whelan, J. (2009). Thetranscription factor ABI4 is a regulator of mitochondrial retrogradeexpression of ALTERNATIVE OXIDASE1a. Plant Physiol. 150: 1286–1296.
Giraud, E., Ho, L.H.M., Clifton, R., Carroll, A., Estavillo, G., Tan, Y.-F.,Howell, K.A., Ivanova, A., Pogson, B.J., Millar, A.H., and Whelan, J.(2008). The absence of ALTERNATIVE OXIDASE1a in Arabidopsis re-sults in acute sensitivity to combined light and drought stress. PlantPhysiol. 147: 595–610.
Gutierrez, R., Lindeboom, J.J., Paredez, A.R., Emons, A.M.C., andEhrhardt, D.W. (2009). Arabidopsis cortical microtubules position cellu-lose synthase delivery to the plasma membrane and interact with cellu-lose synthase trafficking compartments. Nat. Cell Biol. 11: 797–806.
Hamann, T. (2015). The plant cell wall integrity maintenance mecha-nism-concepts for organization and mode of action. Plant CellPhysiol. 56: 215–223.
Harholt, J., Jensen, J.K., Sørensen, S.O., Orfila, C., Pauly, M.,and Scheller, H.V. (2006). ARABINAN DEFICIENT 1 is a putativearabinosyltransferase involved in biosynthesis of pectic arabinanin Arabidopsis. Plant Physiol. 140: 49–58.
He, J., Duan, Y., Hua, D., Fan, G., Wang, L., Liu, Y., Chen, Z., Han, L.,Qu, L.J., and Gong, Z. (2012). DEXH box RNA helicase-mediatedmitochondrial reactive oxygen species production in Arabidopsismediates crosstalk between abscisic acid and auxin signaling. PlantCell 24: 1815–1833.
Hématy, K., Sado, P.-E., Van Tuinen, A., Rochange, S., Desnos, T.,Balzergue, S., Pelletier, S., Renou, J.-P., and Höfte, H. (2007). Areceptor-like kinase mediates the response of Arabidopsis cells tothe inhibition of cellulose synthesis. Curr. Biol. 17: 922–931.
Howell, S.H. (2013). Endoplasmic reticulum stress responses inplants. Annu. Rev. Plant Biol. 64: 477–499.
Huang, S., Van Aken, O., Schwarzlander, M., Belt, K., and Millar, A.H.(2016). Roles of mitochondrial reactive oxygen species in cellular sig-nalling and stress response in plants. Plant Physiol. 171: 1551–1559.
Karimi, M., Inzé, D., and Depicker, A. (2002). GATEWAY vectors forAgrobacterium-mediated plant transformation. Trends Plant Sci. 7:193–195.
Koprivova, A., des Francs-Small, C.C., Calder, G., Mugford, S.T.,Tanz, S., Lee, B.R., Zechmann, B., Small, I., and Kopriva, S.(2010). Identification of a pentatricopeptide repeat protein impli-cated in splicing of intron 1 of mitochondrial nad7 transcripts.J. Biol. Chem. 285: 32192–32199.
Kranz, R.G., Richard-Fogal, C., Taylor, J.-S., and Frawley, E.R.(2009). Cytochrome c biogenesis: mechanisms for covalent mod-ifications and trafficking of heme and for heme-iron redox control.Microbiol. Mol. Biol. Rev. 73: 510–528.
Lei, L., Singh, A., Bashline, L., Li, S., Yingling, Y.G., and Gu, Y.(2015). CELLULOSE SYNTHASE INTERACTIVE1 is required for fastrecycling of cellulose synthase complexes to the plasma membranein Arabidopsis. Plant Cell 27: 2926–2940.
Liu, Y., He, J., Chen, Z., Ren, X., Hong, X., and Gong, Z. (2010). ABAoverly-sensitive 5 (ABO5), encoding a pentatricopeptide repeatprotein required for cis-splicing of mitochondrial nad2 intron 3, isinvolved in the abscisic acid response in Arabidopsis. Plant J. 63:749–765.
Mavridou, D.A.I., Ferguson, S.J., and Stevens, J.M. (2013). Cyto-chrome c assembly. IUBMB Life 65: 209–216.
May, M.J., and Leaver, C.J. (1993). Oxidative stimulation of gluta-thione synthesis in Arabidopsis thaliana suspension cultures. PlantPhysiol. 103: 621–627.
McFarlane, H.E., Döring, A., and Persson, S. (2014). The cell biologyof cellulose synthesis. Annu. Rev. Plant Biol. 65: 69–94.
Meyer, E.H., Tomaz, T., Carroll, A.J., Estavillo, G., Delannoy, E.,Tanz, S.K., Small, I.D., Pogson, B.J., and Millar, A.H. (2009).Remodeled respiration in ndufs4 with low phosphorylation effi-ciency suppresses Arabidopsis germination and growth and alterscontrol of metabolism at night. Plant Physiol. 151: 603–619.
Mitochondria Mediate Cell Wall Integrity 2289

Miart, F., Desprez, T., Biot, E., Morin, H., Belcram, K., Höfte, H., Gonneau,M., and Vernhettes, S. (2014). Spatio-temporal analysis of cellulose syn-thesis during cell plate formation in Arabidopsis. Plant J. 77: 71–84.
Millar, A.H., Sweetlove, L.J., Giegé, P., and Leaver, C.J. (2001).Analysis of the Arabidopsis mitochondrial proteome. Plant Physiol.127: 1711–1727.
Millar, A.H., Whelan, J., Soole, K.L., and Day, D.A. (2011). Organi-zation and regulation of mitochondrial respiration in plants. Annu.Rev. Plant Biol. 62: 79–104.
Murashige, T., and Skoog, F. (1962). A revised medium for rapidgrowth and bio assays with tobacco tissue cultures. Physiol. Plant.15: 473–497.
Murcha, M.W., and Whelan, J. (2015). Isolation of intact mitochon-dria from the model plant species Arabidopsis thaliana and Oryzasativa. Methods Mol. Biol. 1305: 1–12.
Ng, S., De Clercq, I., Van Aken, O., Law, S.R., Ivanova, A., Willems, P.,Giraud, E., Van Breusegem, F., andWhelan, J. (2014). Anterograde andretrograde regulation of nuclear genes encoding mitochondrial proteinsduring growth, development, and stress. Mol. Plant 7: 1075–1093.
Ng, S., et al. (2013a). A membrane-bound NAC transcription factor,ANAC017, mediates mitochondrial retrograde signaling in Arabi-dopsis. Plant Cell 25: 3450–3471.
Ng, S., et al. (2013b). Cyclin-dependent kinase E1 (CDKE1) providesa cellular switch in plants between growth and stress responses.J. Biol. Chem. 288: 3449–3459.
Persson, S., Paredez, A., Carroll, A., Palsdottir, H., Doblin, M.,Poindexter, P., Khitrov, N., Auer, M., and Somerville, C.R. (2007).Genetic evidence for three unique components in primary cell-wallcellulose synthase complexes in Arabidopsis. Proc. Natl. Acad. Sci.USA 104: 15566–15571.
Rhoads, D.M., and Subbaiah, C.C. (2007). Mitochondrial retrograderegulation in plants. Mitochondrion 7: 177–194.
Rhoads, D.M., Umbach, A.L., Subbaiah, C.C., and Siedow, J.N.(2006). Mitochondrial reactive oxygen species. Contribution to oxidativestress and interorganellar signaling. Plant Physiol. 141: 357–366.
Schmitz-Linneweber, C., and Small, I. (2008). Pentatricopeptiderepeat proteins: a socket set for organelle gene expression. TrendsPlant Sci. 13: 663–670.
Skirycz, A., De Bodt, S., Obata, T., De Clercq, I., Claeys, H., De Rycke,R., Andriankaja, M., Van Aken, O., Van Breusegem, F., Fernie, A.R.,and Inzé, D. (2010). Developmental stage specificity and the role ofmitochondrial metabolism in the response of Arabidopsis leaves to pro-longed mild osmotic stress. Plant Physiol. 152: 226–244.
Srivastava, R., Deng, Y., Shah, S., Rao, A.G., and Howell, S.H.(2013). BINDING PROTEIN is a master regulator of the endoplasmicreticulum stress sensor/transducer bZIP28 in Arabidopsis. PlantCell 25: 1416–1429.
Starkov, A.A. (2008). The role of mitochondria in reactive oxygenspecies metabolism and signaling. Ann. N. Y. Acad. Sci. 1147: 37–52.
Sung, T.-Y., Tseng, C.-C., and Hsieh, M.-H. (2010). The SLO1 PPRprotein is required for RNA editing at multiple sites with similarupstream sequences in Arabidopsis mitochondria. Plant J. 63: 499–511.
Takenaka, M. (2010). MEF9, an E-subclass pentatricopeptide repeatprotein, is required for an RNA editing event in the nad7 transcript inmitochondria of Arabidopsis. Plant Physiol. 152: 939–947.
Takenaka, M., Verbitskiy, D., Zehrmann, A., and Brennicke, A.(2010). Reverse genetic screening identifies five E-class PPR pro-teins involved in RNA editing in mitochondria of Arabidopsis thali-ana. J. Biol. Chem. 285: 27122–27129.
Tenhaken, R. (2015). Cell wall remodeling under abiotic stress. Front.Plant Sci. 5: 771.
Updegraff, D.M. (1969). Semimicro determination of cellulose in bi-ological materials. Anal. Biochem. 32: 420–424.
Van Aken, O., Zhang, B., Law, S., Narsai, R., and Whelan, J. (2013).AtWRKY40 and AtWRKY63 modulate the expression of stress-responsive nuclear genes encoding mitochondrial and chloroplastproteins. Plant Physiol. 162: 254–271.
Van Aken, O., De Clercq, I., Ivanova, A., Law, S.R., Van Breusegem,F., Millar, A.H., and Whelan, J. (2016). Mitochondrial and chloro-plast stress responses are modulated in distinct touch and chemicalinhibition phases in Arabidopsis. Plant Physiol. 171: 2150–2165.
Vandavasi, V.G., et al. (2016). A structural study of CESA1 catalyticdomain of Arabidopsis cellulose synthesis complex: evidence forCESA trimers. Plant Physiol. 170: 123–135.
Vanderauwera, S., Vandenbroucke, K., Inzé, A., van de Cotte, B.,Mühlenbock, P., De Rycke, R., Naouar, N., Van Gaever, T., VanMontagu, M.C., and Van Breusegem, F. (2012). AtWRKY15 per-turbation abolishes the mitochondrial stress response that steersosmotic stress tolerance in Arabidopsis. Proc. Natl. Acad. Sci. USA109: 20113–20118.
Vanlerberghe, G.C. (2013). Alternative oxidase: a mitochondrial respiratorypathway to maintain metabolic and signaling homeostasis during abioticand biotic stress in plants. Int. J. Mol. Sci. 14: 6805–6847.
Verbitskiy, D., Zehrmann, A., van der Merwe, J.A., Brennicke, A., andTakenaka, M. (2010). The PPR protein encoded by the LOVASTATININSENSITIVE 1 gene is involved in RNA editing at three sites in mito-chondria of Arabidopsis thaliana. Plant J. 61: 446–455.
Wang, Y., Carrie, C., Giraud, E., Elhafez, D., Narsai, R., Duncan, O.,Whelan, J., and Murcha, M.W. (2012). Dual location of the mito-chondrial preprotein transporters B14.7 and Tim23-2 in complex Iand the TIM17:23 complex in Arabidopsis links mitochondrial ac-tivity and biogenesis. Plant Cell 24: 2675–2695.
Welchen, E., Hildebrandt, T.M., Lewejohann, D., Gonzalez, D.H., andBraun, H.-P. (2012). Lack of cytochrome c in Arabidopsis decreasesstability of Complex IV and modifies redox metabolism without affectingComplexes I and III. Biochim. Biophys. Acta 1817: 990–1001.
Wolf, S., Hématy, K., and Höfte, H. (2012). Growth control and cellwall signaling in plants. Annu. Rev. Plant Biol. 63: 381–407.
Wu, J., et al. (2015). Deficient plastidic fatty acid synthesis triggerscell death by modulating mitochondrial reactive oxygen species.Cell Res. 25: 621–633.
Xia, Y., Lei, L., Brabham, C., Stork, J., Strickland, J., Ladak, A., Gu,Y., Wallace, I., and DeBolt, S. (2014). Acetobixan, an inhibitor ofcellulose synthesis identified by microbial bioprospecting. PLoSOne 9: e95245.
Yang, L., Zhang, J., He, J., Qin, Y., Hua, D., Duan, Y., Chen, Z., andGong, Z. (2014). ABA-mediated ROS in mitochondria regulate rootmeristem activity by controlling PLETHORA expression in Arabi-dopsis. PLoS Genet. 10: e1004791.
Zehrmann, A., Verbitskiy, D., van der Merwe, J.A., Brennicke, A.,and Takenaka, M. (2009). A DYW domain-containing penta-tricopeptide repeat protein is required for RNA editing at multiplesites in mitochondria of Arabidopsis thaliana. Plant Cell 21: 558–567.
Zhu, J., Lee, B.H., Dellinger, M., Cui, X., Zhang, C., Wu, S., Nothnagel,E.A., and Zhu, J.K. (2010). A cellulose synthase-like protein is requiredfor osmotic stress tolerance in Arabidopsis. Plant J. 63: 128–140.
Zhu, Q., Dugardeyn, J., Zhang, C., Mühlenbock, P., Eastmond,P.J., Valcke, R., De Coninck, B., Oden, S., Karampelias, M.,Cammue, B.P., Prinsen, E., and Van Der Straeten, D. (2014). TheArabidopsis thaliana RNA editing factor SLO2, which affects themitochondrial electron transport chain, participates in multiplestress and hormone responses. Mol. Plant 7: 290–310.
Zsigmond, L., Rigó, G., Szarka, A., Székely, G., Otvös, K., Darula,Z., Medzihradszky, K.F., Koncz, C., Koncz, Z., and Szabados, L.(2008). Arabidopsis PPR40 connects abiotic stress responses tomitochondrial electron transport. Plant Physiol. 146: 1721–1737.
2290 The Plant Cell

DOI 10.1105/tpc.16.00540; originally published online August 19, 2016; 2016;28;2276-2290Plant Cell
VeylderMouille, Samantha Vernhettes, Frank Van Breusegem, James Whelan, Herman Höfte and Lieven DeNguyen, Katharina Belt, A. Harvey Millar, Dominique Audenaert, Pierre Hilson, Ian Small, Grégory Zhubing Hu, Rudy Vanderhaeghen, Toon Cools, Yan Wang, Inge De Clercq, Olivier Leroux, Long
Mitochondrial Defects Confer Tolerance against Cellulose Deficiency
This information is current as of July 9, 2018
Supplemental Data /content/suppl/2016/08/19/tpc.16.00540.DC1.html /content/suppl/2016/08/31/tpc.16.00540.DC2.html
References /content/28/9/2276.full.html#ref-list-1
This article cites 76 articles, 39 of which can be accessed free at:
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists