micrognathozoa: a new class with complicated jaws...
TRANSCRIPT

Micrognathozoa:A New Class With Complicated Jaws LikeThose of Rotifera and GnathostomulidaReinhardt Møbjerg Kristensen1* and Peter Funch2
1Department of Invertebrate Zoology, Zoological Museum, University of Copenhagen,Copenhagen Ø, Denmark2Department of Zoology, Institute of Biological Sciences, University of Åarhus, Åarhus C, Denmark
ABSTRACT A new microscopic aschelminth-like ani-mal, Limnognathia maerski nov. gen. et sp., is describedfrom a cold spring at Disko Island, West Greenland, andassigned to Micrognathozoa nov. class. It has a complex ofjaws in its pharynx, and the ultrastructure of the mainjaws is similar to that of the jaws of advanced scleropera-lian gnathostomulids. However, other jaw elements ap-pear also to have characteristics of the trophi of Rotifera.Jaw-like structures are found in other protostome taxa aswell—for instance, in proboscises of kalyptorhynch platy-helminths, in dorvilleid polychaetes and aplacophoranmollusks—but studies of their ultrastructure show thatnone of these jaws is homologous with jaws found inGnathostomulida, Rotifera, and Micrognathozoa. The lat-ter three groups have recently been joined into the mono-phylum Gnathifera Ahlrichs, 1995, an interpretation sup-ported by the presence of jaw elements with cuticular rodswith osmiophilic cores in all three groups. Such tubularstructures are found in the fulcrum of all Rotifera and inseveral cuticular sclerites of both Gnathostomulida andMicrognathozoa. The gross morphology of the pharyngealapparatus is similar in the three groups. It consists of aventral pharyngeal bulb and a dorsal pharyngeal lumen.The absence of pharyngeal ciliation cannot be used as anautapomorphy in the ground pattern of the Gnathiferabecause the Micrognathozoa has the plesiomorphic alter-native with a ciliated pharyngeal epithelium.
The body of Limnognathia maerski nov. gen. et sp. con-sists of a head, thorax, and abdomen. The dorsal and
lateral epidermis have plates formed by an intracellularmatrix, as in Rotifera and Acanthocephala; however, theepidermis is not syncytial. The ventral epidermis lacksinternal plates, but has a cuticular oral plate withoutciliary structures. Two ventral rows of multiciliated cellsform a locomotory organ. These ciliated cells resemble theciliophores present in some interstitial annelids. An adhe-sive ciliated pad is located ventrally close to a caudalplate.
As in many marine interstitial animals—e.g., gnatho-stomulids, gastrotrichs, and polychaetes—a special formof tactile bristles or sensoria is found on the body. Twopairs of protonephridia with unicellular terminal cells arefound in the trunk; this unicellular condition may be theplesiomorphic condition in Bilateria. Only specimens withthe female reproductive system have been found, indicat-ing that all adult animals are parthenogenetic females.We suggest that 1) jaws of Gnathostomulida, Rotifera, andthe new taxon, Micrognathozoa, are homologous struc-tures; 2) Rotifera (including Acanthocephala) and the newgroup might be sister groups, while Gnathostomulidacould be the sister-group to this assemblage; and 3) thesimilarities to certain gastrotrichs and interstitialpolychaetes are convergent. J. Morphol. 246:1–49, 2000.© 2000 Wiley-Liss, Inc.
KEY WORDS: Micrognathozoa; new class; Gnathifera;Limnognathia; taxonomy; ecology; phylogeny; Arctic;Greenland; freshwater meiofauna; cold spring
The rich vegetation near the more than 1,000 ho-mothermic springs of Disko Island, West Greenland,has been considered to contain a southern relictelement from a period with a warmer climate whenthe ranges of southern elements in the flora ex-tended beyond the island of Disko (Porsild, 1920;Kristensen, 1987). In the same way the marine fau-nal elements in Greenland springs have been re-garded as relicts from a postglacial hypsithermalperiod, when many springs were below sea level(Kristensen, 1977).
Measurements of the abiotic parameters in morethan 100 homothermic springs on Disko Island in-dicate that the springs can be separated into at least
three different types (Kristensen, 1982). The marinefaunal elements are only found in warm electrolyte-
Publication from the Danish Arctic Station, University of Copenha-gen, Denmark.
Contract grant sponsor: the Carlsberg Foundation; Contract grantnumber: 970345/30 - 488; Contract grant sponsor: the Danish Re-search Agency; Contract grant numbers: 9701589, 9801880.
*Correspondence to: R.M. Kristensen, Department of InvertebrateZoology, Zoological Museum, University of Copenhagen, Univer-sitetsparken 15, DK-2100 Copenhagen Ø, Denmark.E-mail: [email protected]
JOURNAL OF MORPHOLOGY 246:1–49 (2000)
© 2000 WILEY-LISS, INC.

rich, radioactive springs (“salt springs”). The domi-nant type of spring on Disko Island is an electrolyte-poor spring with only “normal freshwater” species.The “southern” vegetation around the springs is aresult of the so-called “greenhouse effect” broughtabout by the snow and ice cover in winter.
Compared with the “warm” springs of Disko Is-land, very few investigations had been carried out incold springs, which may be frozen up to 8 months ofthe year. The freshwater fauna of the cold (hetero-thermic) springs differ greatly from those of thehomothermic springs. The flora and fauna of a coldspring (Isunngua/Mudderbugten) was comparedwith that of the relatively cold homothermic springs(Sullorsuaq/Kvandalen) during a field course in Arc-tic Biology, 1994, at Disko Island. To our surprise,we found a new type of animal in the cold spring(Kristensen, 1995; Kristensen and Funch, 1995) ep-iphytic on water mosses. Here we fully describe thenew taxon, discuss its phylogenetic position, andcorrect some misconceptions about this unique ani-mal (see Ahlrichs, 1997; Herlyn and Ehlers, 1997).
The first specimens of the new species were col-lected in 1979 at Disko Island. These three speci-mens were then labeled Rotifera because they werenot observed alive and were strongly contracted inthe formaldehyde/glycerol preparation. Later, werecognized the complicated jaw apparatus. The newanimal has several superficial similarities to mono-gonont rotifers, especially in the pharyngeal appa-ratus. Therefore, we compared the extremely com-plex jaw apparatus with the more simple mastax ofseveral species of Monogononta by scanning electronmicroscopy (SEM). Recently, excellent SEM analysisof the sclerite system of the rotifer mastax has beenpublished (Markevich, 1989; De Smet, 1996, 1997).Surprisingly the jaw apparatus of the new animalclearly has elements similar to the scleroperaliangnathostomulids (Kristensen and Nørrevang, 1977,1978; Herlyn and Ehlers, 1997), especially in themain jaws (articularium and dentarium). We com-pare the lamellarization of the dentarium and thefibularization of the apophysis in the family Gnatho-stomulidae (Riedl and Rieger, 1972) with similarstructures in the main jaws of the new animal.
Gnathostomulida was described by Ax (1956) asan order of Turbellaria (Platyhelminthes). Later,Riedl (1969) established a new phylum for thegnathostomulids and Sterrer (1972) confirmed thenew status of the group, but he also mentioned thatgnathostomulids share characteristics with bothPlatyhelminthes and Aschelminthes. In the excel-lent review article of Lammert (1991) the Gnatho-stomulida was also included in the Aschelminthes.Zoology textbooks, such as Ruppert and Barnes(1994), follow the idea of Sterrer et al. (1985) andLammert (1991) and include the Gnathostomulidain the Aschelminthes. Nielsen (1995) included themin Annelida. Littlewood et al. (1998) were the first tosequence 18S ribosomal DNA from a species of
Gnathostomulida. In comparing this sequence tothat of other phyla they came to the conclusion thatthe Gnathostomulida is a member of a Nematoda 1Chaetognatha clade. Zrzavy et al. (1998) came toanother conclusion. They used the 18S ribosomalDNA data of Gnathostomulida from Littlewood et al.(1998) and combined these with morphological char-acters in a total-evidence approach. Analysis of thishuge dataset indicated a monophylum Neotrichozoaconsisting of Gnathostomulida and Gastrotricha. Sofar, the only agreement is that gnathostomulids areprotostomian worms.
Recently, Rieger and Tyler (1995) and Ahlrichs(1995) cited ultrastructural evidence for a sister-group relationship of Gnathostomulida withRotifera-Acanthocephala. Ahlrichs (1995) estab-lished a new monophylum Gnathifera for thesegroups. Later, Ahlrichs (1997) included our newtaxon as “New group A” in his phylogenetic diagramof Gnathifera. He cited our unpublished Danish re-port from the Arctic Field Course (Kristensen andFunch, 1995). Unfortunately, the same new taxonwas also mentioned as “New group 1” (Kristensen,1995). “New group A” for our unnamed taxon and itsinclusion in Gnathifera were mentioned again in thearticle of Herlyn and Ehlers (1997). They arguedthat the trophi of rotifers and the cuticular jaws ofgnathostomulids are homologous. Their conclusionwas based on a transmission electron microscopy(TEM) investigation of the pharynx of the sclerop-eralian gnathostomulid, Gnathostomula paradoxa.Recently, Sørensen (2000) made the first SEM-investigation of the two main type jaws (“compact”and “basket” type) within the Gnathostomulida. Hisstudy gives further support for a closer relationshipwith rotifers by virtue of similarities with the jawsand trophi of advanced monogonont rotifers. Sø-rensen shows that the pseudofulcrum of Rastro-gnathia macrostoma resembles the fulcrum in therotiferan family Dicranophoridae. The cuticular el-ements, the sclerofibrillae, are almost identical inthe two groups. Our new taxon from Greenland be-longs to Gnathifera and supports the monophyly ofthis taxon comprising the Gnathostomulida, Rotif-era (including Seison), and Acanthocephala. Conse-quently, Gnathostomulida is not as closely related tothe Turbellaria (Platyhelminthes) as suggested byAx (1956, 1985, 1989, 1996).
Finally, we considered other possibilities for aphylogenetic relationship for the “New Group 1,”because we doubt that Aschelminthes is monophy-letic, a hypothesis that has gained support fromrecent phylogenetic analyses using morphologicaldata (Nielsen et al., 1996; Sørensen et al., 2000),molecular data and combined data (Giribet et al.,2000). There are similarities in outer morphology—e.g., the ventral trunk ciliation and sensoria—between certain gastrotrichs (Hyman, 1951; Rup-pert, 1991b) and the new animal. The phylogeneticsignificance of these structures is briefly discussed.
2 R. M. KRISTENSEN AND P. FUNCH

The free-swimming chordoid larva of Cycliophora(Funch and Kristensen, 1995; Funch, 1996) has agastrotroch of compound cilia as in the ventral cili-ation of the new animal. This is probably an adap-tation to crawling and not a homology. We discussthe hypothesis of polyphyletic origin of Aschel-minthes (Winnepenninckx et al., 1995) in the con-text of groups, such as the interstitial polychaetefamilies Diurodrilidae and Dorvilleidae (see Kris-tensen and Niilonen, 1982; Eibye-Jacobsen andKristensen, 1994) having ventral ciliophores both onthe head and abdomen, as does our new animal.Furthermore, the family Dorvilleidae has a cuticularjaw in the pharyngeal apparatus. The recently de-scribed species of dorvilleid, Neotenotrocha sterreri,has a swimming behavior like that of a rotifer and ajaw apparatus similar to the trophi of rotifers. Inlight of the similarities between Neotenotrocha,“New Group 1,” and Rotifera the old theory of Sem-per (1872) that the rotifers are simply neotenic an-nelids is discussed.
MATERIALS AND METHODSSubstrate and Abiotic Factorsin the Isunngua Spring
The sources of the Isunngua spring are located1,030–1,120 m from the coast at 50 m above sealevel in a cold moor vegetation. The outflow runsthrough two more moors at 315–345 m and 200–296m from the coast. Between the last two moor areasand the sea the outflow runs in a 30–50 cm-deepwater channel in well-sorted sand from the Creta-ceous. The outlet from the second moor is the typelocality at a position: 69°43.799’N and 51°56.549’W.The spring temperature on 5 August 1994 was 5°C,the conductivity 62 mmho, and the pH in the fieldwas 6.4. The radioactivity was only slightly higherthan the background.
The new animal was found only in the last twomoors and the outlets from the moors. More than100 specimens were collected epiphytic in themosses on 22 July 1994, 25 July 1994, 5 August1994, and 20 May 1995. About 5 kg moss and soilwere collected 5 August 1994 and carried to Copen-hagen and kept in culture in a 4°C refrigerator withconstant light and airing. The animals reproducedin the moss culture until 28 February 1997, when anunknown fungus overgrew the culture. The mossspecies on which the new animal lives epiphyticallycomprise the following species: Aulocommiumpalustre, Calliergon sarmenfosum (the dominantspecies), Drepanocladus intermedius, Paludellasquarrosa, and Tomenthypnum nitens (soil species).
Living Materials
Mosses with the rhizoids kept at 4°C weresqueezed in spring water for detritus and meiofaunaand the rinsings decanted through a 32-mm mesh
net. The epiphytic meiofauna was placed in springwater or distilled water in Petri dishes at roomtemperature and inspected under a stereomicro-scope using magnifications of 340–3100. Afterabout a half hour the new animal could be observedfree-swimming in the water column. The animalsadhere strongly to Pasteur pipettes, so Irwin loopswere use to transfer specimens to the microslides,where they were mounted in spring water and pho-tographed in the differential interference contrastmicroscope (DIC, Nomarski technique) or video-taped with a camera mounted on a Zeiss phase con-trast microscope.
The holotype (Fig. 1) and one paratype (Fig. 2)were drawn using camera lucida (magnification32,000). These habitus drawings were made at 4°C,when the animals were still alive but had beenslightly squeezed. The measurements of total lengthand width of living specimens are about 20% largerthan the same animal later fixed with osmium-tetroxide vapor. The drawings and measurements ofall sensoria, ciliophores, adhesive ciliated pad, andthe dorsal plates could only be made on living butslightly squeezed animals (Figs. 3, 4). Some animalsfrom the culture were prepared for SEM (Figs. 5, 6)and a prominent oral plate that was not included inthe drawings was observed.
Some animals laid eggs in small salt cellars andsome of these eggs were prepared for SEM (Fig. 7).
Wholemount Preparations
Twenty-eight specimens were fixed with a drop of4% formaldehyde (buffered with borax) in the springwater or with 1% osmium-tetroxide vapor directedonto the hanging drop of water on the microslides.Thereafter the specimens were mounted with a cov-erslip in the fixative and the fixative was replaced by2% glycerol in distilled water that evaporated toglycerol over several days; the mounts were finallysealed with Glyceelt.
Seven living mounted specimens were treatedwith 2% sodium hypochlorite, using the techniquedescribed by Riedl and Rieger (1972) for isolatingthe jaws of Gnathostomulida. The specimen wasmounted under a coverslip in spring water in asqueezed preparation. Thereafter, sodium hypochlo-rite was sucked in with bibulous paper. The jawsand the basal plate (Figs. 8, 9) were strongly resis-tant to oxidation; however, the outer oral plate, thedifferent lamellae, ligaments, and symphyses be-came transparent and disappeared under thebleaching process.
The photomicrographs were taken during thebleaching process of the jaw apparatus (Figs. 10–15). After bleaching, the tissue debris was removedwith distilled water, glycerol was added, and thecoverslip was sealed with Glyceelt. All wholemountswere examined with phase contrast and DIC opticsand drawn using a camera lucida. For drawings of
3MICROGNATHOZOA: A NEW CLASS

the pharyngeal apparatus and the complex jaw sys-tem (Figs. 8, 9), a Wild drawing tube mounted on aWild M20 microscope was used, allowing a drawingmagnification of 310,000.
Electron Microscopical Techniques
Adult specimens and eggs used for SEM werefixed in 4% buffered formaldehyde or osmium-tetroxide vapor. Specimens were transferredthrough an acetone dehydration series and werecritical-point dried using carbon dioxide; thereafter,they were mounted on aluminum SEM stubs withdouble-sided tape.
A new technique was developed for SEM observa-tions of the jaw apparatus. For this, several livinganimals were placed in a small drop of distilledwater directly on the aluminum SEM stub. Whenthe distilled water evaporated, the dried animalsadhered strongly to the SEM stub. The soft tissuewas then removed by 2% sodium hypochlorite andthe cleaned jaw apparatus was rinsed several timeswith distilled water. All SEM preparations weresputter-coated with gold and examined in a JEOLJSM-840 scanning electron microscope (Figs. 16,17).
For TEM, six adult animals were fixed in a mix-ture of three aldehydes, a so-called trialdehyde fix-ation (Lake, 1973), in 0.1 M sodium cacodylatebuffer (see Kalt and Tandler, 1971). All specimenswere postfixed in 1% osmium-tetroxide with 0.1 Msodium cacodylate buffer for 1 h at 20°C. After fix-ation, the animals were dehydrated in an ethanolseries, transferred to propylene oxide, and finallyembedded in epoxy resin type TAAB 812t. The ul-trathin serial sections were stained with uranyl ac-etate and lead citrate (Reynolds, 1963). TEM exam-inations (Figs. 18–33) were performed with a JEOLJEM-100SX transmission electron microscope.
DESCRIPTION
Phylum: Gnathifera Ahlrichs, 1995
Micrognathozoa, new class
Diagnosis. Acoelomate metazoans with bilateralsymmetry and epidermal dorsal and lateral intra-cellular plates. Epidermis cellular, not syncytial.Body divided into a head with the pharyngeal appa-ratus, an accordion-like thorax, and an abdomenwith a dorsal anus, which may be functional onlyperiodically. Ventral epidermis with thick glycoca-lyx and two rows of multiciliated ciliophores. Ven-tral mouth opening surrounded by a cuticular oralplate. Epidermis of the mouth cavity and the pha-ryngeal apparatus also with a cuticle. Jaw appara-tus with one unpaired and nine paired major scle-rites. Females with one pair of ovaries. One egg per
clutch. Two pairs of protonephridia with monocili-ated terminal cells. Direct development.
Etymology. Micro, gnathos and zoa are Greek for“small,” “jaws,” and “animal,” referring to the smallanimal with complex jaws.
Limnognathida, new orderDiagnosis. Same as the class and with a life-cycle
with free-living individuals in freshwaters.
Limnognathiidae, new familyDiagnosis. Same as the class.
Limnognathia gen. nov.Diagnosis. Same as the class.Type species. Limnognathia maerski, new spe-
cies by designation (Figs. 1, 2).Etymology. Limnos and gnathos are Greek for
“freshwater” and “jaws,” referring to the habitat be-ing freshwater; feminine gender.
Limnognathia maerski sp. nov.Diagnosis. Mature females 105–152 mm long, ju-
veniles 85–107 mm long, ovoid sculptured egg 40 330 mm. Stiff sensoria consisting of: 1) one pair ofapicalia, 2) one pair of frontalia, 3) five pairs oflateralia, 4) three pairs of dorsalia, and 5) two pairsof caudalia, each with a ring-shaped epidermalsocket. Second dorsalia (Fig. 1, do2) on the thoraxare double and may lack cilia.
Etymology. To honor Maersk McKinney Møller,who sponsored the new research vessel Porsild forthe Arctic Station. The new animal was discoveredduring the maiden trip with the new research vesselin 1994.
Type material. The holotype (MIC 0001, ZMUC)is an adult female with two unsculptured oocytes.This wholemount slide, together with 19 paratypes(MIC 0002-MIC 0020, ZMUC), is deposited in theZoological Museum of Copenhagen, Denmark. Sevensodium hypochlorite-treated jaws are also depositedon microslides (MIC 0021-MIC 0023, ZMUC). Threeparatypes and two jaw apparatuses are located onSEM stubs. Six paratypes are ultrasectioned andlocated on 105 grids. The type material is placed inZMUC and a single paratype is in the National
™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™3Fig. 1. Holotype of Limnognathia maerski nov. gen. et nov. sp.
Dorsal view of the slightly squeezed living female from the springat Isunngua, Disko Island, W. Greenland. A few details fromstudy after the animal was fixed are included. ac, apicalia; an,anus; ap, apical plate; at, apical cilia tuft; ca, cauda of the mainjaws (ja); cd1-cd2, caudalia; do1-do3, dorsalia; dp, dorsal plate; fg,flagellar head structure; fr, frontalia; la1-la5, lateralia; lp, lateralplate; mg, midgut; oo1-oo2, oocytes; ph, pharyngeal apparatus; sg,salivary gland; ta, tail (pygidium).
4 R. M. KRISTENSEN AND P. FUNCH
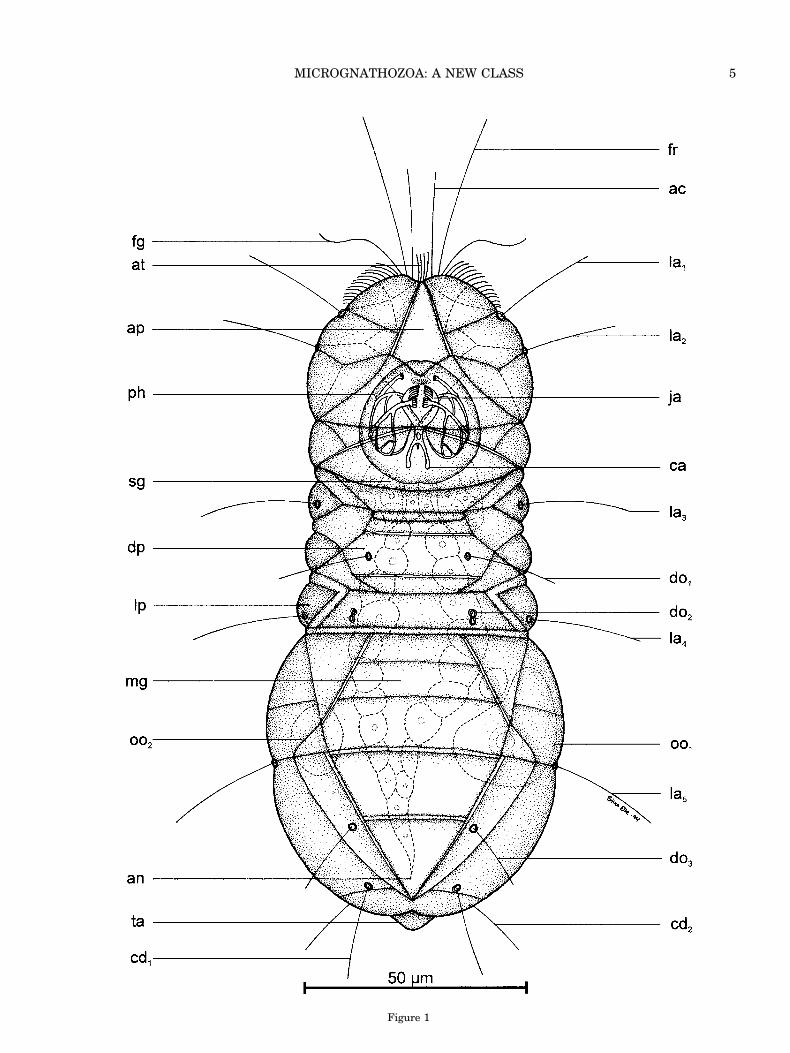
Figure 1
5MICROGNATHOZOA: A NEW CLASS

Museum of Natural History, Smithsonian Institu-tion (USNM), Washington DC, USA.
Type locality. The type material was extractedfrom living water mosses from the cold Isunnguaspring (69°43.799’N, 51°56.549’W), located at theeastern corner of Disko Island, West Greenland(Figs. 34–39). The living material was collected bythe authors on 22 July 1994, 25 July 1994, 5 August1994, and 20 May 1995. The holotype was from the5 August 1994 collection.
Additional material. Three strongly retractedanimals had been mounted on microslides in 1979.This material is from a cold homothermic springclose to Lymnaea lake (69°42.2979N, 52°11.4359W)in the valley of Sullorsuaq/Kvandalen (Fig. 36). Thespecimens were collected during a survey for tardi-grades in July 1979 (Kristensen, 1982).
Description of the Holotype (Adult Female)
The holotype was observed alive and after fixationwith 1% osmium-tetroxide and preparation of a gly-cerol wholemount. The total length of the living an-imal was 142 mm and the maximum width of theabdomen was 55 mm. The drawing (Fig. 1) is indorsal view. The body seems segmented or dividedinto a two-parted head, accordion-like thorax, andovoid abdomen with a small retractile pygidium (tail).
External anatomy. The whole dorsal part (dor-sum) of the animal is covered with plates. The ani-mal bears sensoria on all parts of the body. As inmany marine interstitial animals, e.g., gnathosto-mulids, gastrotrichs, and polychaetes, these tactilebristles (stiff, adjoined cilia) consist of more than onecilium. In each sensorium the cilia seem to emergefrom one cell. However, this observation is not yetconfirmed with TEM. In the holotype, two to threeadjoined stiff cilia arise from a circular reinforce-ment, the socket. Still, when the bristle disappearsunder fixation the socket can be recognized as a ringwith a pore in the middle. The tactile sensoria arealways found in pairs, serially arranged on the body.The arrangement is consistent in all investigatedspecimens of Limnognathia maerski, including theholotype. On the anterior part of the head are fourpairs of bristles. A pair of long frontalia (fr, 34 mmlong) and a pair of shorter apicalia (ac, 23 mm long)are located on the frontal margin of the head. Thesesensoria are directed forward. Two pairs of laterallyoriented sensoria (la1 and la2, 22 mm long) are lo-cated between the sutures of the first two lateralheadplates. The posterior part of the head lacksbristles. The thorax has two pairs of lateralia (la3and la4, 25 mm long) and two pairs of dorsalia (do1,18 mm long and do2, without ciliary structure). Theyare located on middorsal plates. The posterior pair(do2) lacks the external ciliary structures in the ho-lotype, and the socket for the sensory structure is adouble structure (8-shaped) and has a pore in theanterior part of the structure.
The abdomen has a single pair of lateralia (la5, 24mm long) located in a constriction that divides theabdomen into two parts. Furthermore, a shorter pairof dorsalia (do3, 15 mm long), which is orienteddorso-caudally, is located on two thin dorso-lateralplates close to the triangular anal plate. Dorso-caudally, a pair of caudalia (cd1, 19 mm long) andventro-caudally another pair of caudalia (cd2, 22 mmlong) are located. Both pairs are oriented caudallyand the sockets of the ventro-caudal bristles arelocated on the two caudal plates, which can be seenonly in the ventral view (Fig. 2), and therefore arenot seen in the holotype. All sensoria break off easilyand in the wholemount preparation of the holotypeonly the sockets of the sensoria can be observed.
Three other ciliary structures could be observedon the head of the holotype when it was alive: 1) Onesmall apical cilia tuft (at) was observed between thetwo apicalia. It consists of four short, stiff cilia,although eight cilia were seen in some paratypes.The cilia are about 8 mm long and are not adjoined,but it seems that they arise from the same epider-mal cell. 2) Close to the base of each frontalium, along single flagellum-like structure (30 mm long) ispresent (fg). This structure has the typical stroke ofa flagellum. 3) Between the frontalia and the firstpair of lateralia, a broom of short cilia is present.These cilia are perhaps a part of the ventral preoralcilia field (see Fig. 2, pc). These cilia persist in thewholemount preparation.
The characteristic dorsal plates can best be inves-tigated in the head. Here it is clearly seen that eachplate comprises 3–4 epidermal cells. The bordersbetween cells are shown as broken lines in Figure 1.An exception seems to be the thick apical plate (ap),which strongly reflects light in DIC microscopy andseems to be formed by a single giant epidermal cell.The forehead is separated from the posterior part bya constriction. The forehead consists of eight platesand the posterior part consists of five plates.
The thorax has thick dorsal plates (dp) and moreflexible lateral plates (lp). The lateral plates worklike an accordion and the thorax can change shaperapidly from a broad and solid structure to a thin-ner, longer, and flexible structure. Between the dor-sal and lateral plates, up to five smaller rod-like torhomboid plates are located.
The abdomen is subdivided dorsally with a sulcus(incomplete transverse furrow). The plates are largeand there are no distinctions between dorsal and
™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™3Fig. 2. Limnognathia maerski nov. gen. et nov. sp. Ventral
view of living paratype (slightly squeezed) from the spring atIsunngua, Disko Island. ab, abdomen; ad, adhesive ciliated pad;cd1-cd2, caudalia; cp, caudal plate; rb1-rb2, refractive bodies; ey,eye structure ?; fg, flagellar head structure; hc, head ciliophore;he, head; me, mid-ventral sensorium; mo, mouth opening, oo1-oo2,oocytes; pc, preoral cilia field; ph, pharyngeal apparatus; pr1-pr2,protonephridia; tc, trunk ciliophore; th, thorax (neck).
6 R. M. KRISTENSEN AND P. FUNCH
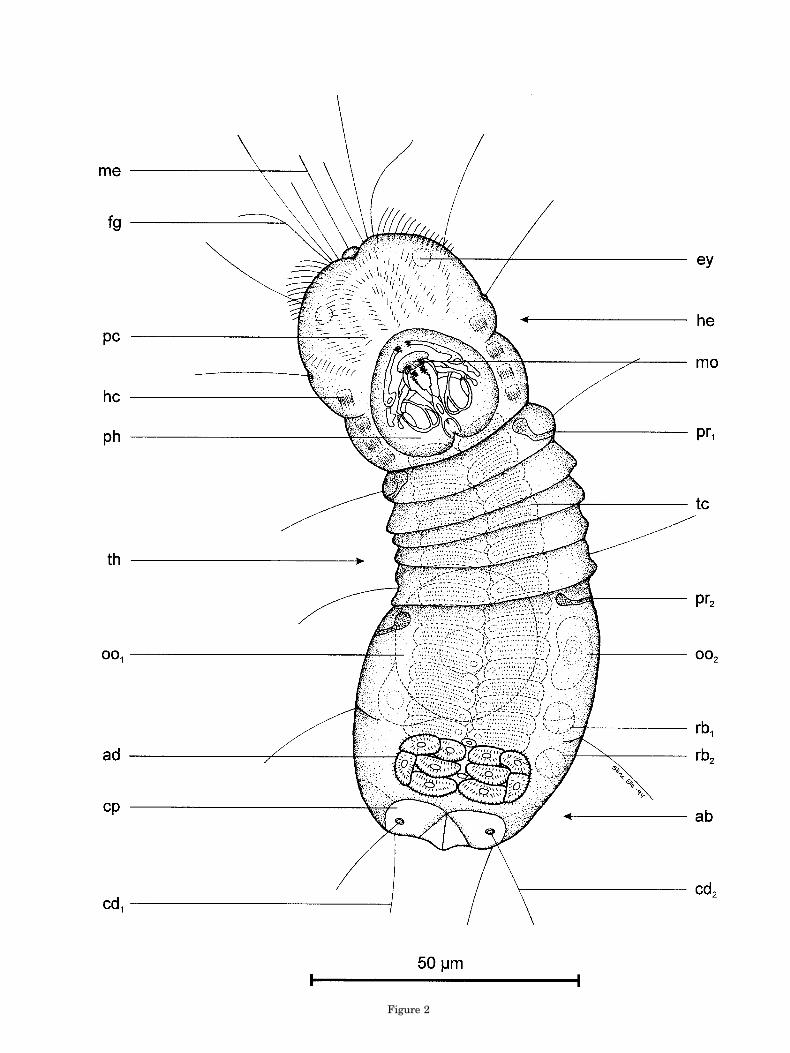
Figure 2

lateral plates. The dorsal triangular anal plate isthick and seems to lack an anal opening.
Internal structures. The thick dorsal plates ob-scure many of the internal structures in the holo-type. Some of these structures could be seen in theallotype from the ventral view (Fig. 2).
The most conspicuous structure in the holotypeand all the paratypes is the large pharyngeal appa-ratus (25 3 23 mm) located in the head. From thedorsal view all three pairs of jaw-like structures (ja)can be seen, but the subunits in these complex struc-tures can only be distinguished in sodium-hypochlorite-treated animals. The cross-striatedmuscles are not drawn in Figure 1 to avoid compli-cating the figure. The muscles are best analyzedwith TEM. The ovoid mouth opening is surroundedby a true cuticle and therefore it can be seen fromthe dorsal view through the transparent tissue. Thepharyngeal apparatus will be described in detailbelow. The large salivary glands (sg) seem to openinside the midgut.
The digestive system continues with a shortesophagus, which leads to a large midgut, consistingof large transparent endoderm cells. The midguttotally lacks ciliary structures. The anus has beendifficult to locate. It seems that Limnognathiamaerski has only a temporary opening. In the holo-type, the anal plate covers the rectum and anus.
The ovary is paired and two large oocytes domi-nate the abdomen. A third right oocyte is seen closeto the large oocyte (oo1). A gonopore was not ob-served.
Description of Allotype
The animal was observed for several hours at 4°Cby DIC optics in a cooling room at Arctic Station.Later, observations were continued on the whole-mount glycerol preparation fixed by 1% osmium-tetroxide vapor. The drawing was made with cameralucida in Greenland of the ventral view (Fig. 2).Details of the pharyngeal apparatus were addedlater. The total length of the living animal was 127mm; maximum width of the abdomen was 47 mm.
External gross anatomy. The entire ventralpart (ventrum) of the animal lacks intracellularplates, except for the two caudal plates (cp), which inthe living animal can be seen ventrally; the pair ofcaudo-ventral caudalia (cd2) is located on theseplates. Furthermore, a large cuticular oral plate ispresent, but it was only observed after the animalwas fixed.
It is clearly seen from a ventral view that the headis divided into two parts. A sulcus separates theforehead from the posterior part. The accordion-shaped thorax is divided by five annulations (trans-verse furrows), which are very flexible and thereforedisappear after fixation. The abdomen has a smallsulcus, but it does not continue as a transverse fur-row to the midventral part side. In a few paratypes,
a very thin furrow was seen in the anterior part ofthe abdomen. This furrow was not as distinct as theannulations on the thorax.
The oval mouth opening (Fig. 2, mo) is locatedmidventrally on the anterior edge of a spade-shapedoral plate. The large oral plate is not drawn in Fig-ure 2, but is seen clearly on the SEM micrograph ofa paratype (Fig. 6, op). The width of the oral plate inthe allotype is 21 mm, and the length is 24 mm. Theoral plate consists of true cuticle and lacks ciliation(Figs. 25, 27, op). In retracted animals the oral platereaches far posterior to the edge of the ventral tho-rax. In the allotype preparation the shape of themouth is a small oval opening, as in all wholemountpreparations of Limnognathia maerski. Therefore itwas very interesting to see the behavior of the livingallotype. In a relaxed swimming position, the mouthopening has the typical oval form, but when theanimal feeds the two ventral jaw elements (ja1) canbe protruded out through the mouth as two smallarms to grasp the substrate. Furthermore, if theanimal swallows some unwanted items the largecross-striated muscles in the head retract the dorsalpart of the forehead, which is lifted upward andbackward. Consequently, most of the pharyngealapparatus with the whole jaw apparatus could beseen to stick out of the mouth. Several fast move-ments of all the jaws elements often accompany thisbehavior. The movements consist both of a snappingreaction and of turning the cauda of the main jaws(Fig. 1, ca) forward and backward. We called thisaction of the jaws a “vomit” behavior. One of theparatypes sectioned for TEM is fixed with the fore-head lifted upward and the jaws protruded.
Ciliophores and ventral ciliation. The ventralciliation is well developed and all body parts have acomplex ciliation, unlike ciliation in taxa such asGastrotricha, Rotifera, and Gnathostomulida. Theforehead is covered ventrally by rows of single cilia.The rows of cilia are formed as arcs, leading parti-cles directly to the mouth opening. From the mouthopening itself several stiff sensory cilia stick out likea broom.
Lateral to the oral plate a quite different ciliationexists. Four pairs of specialized ciliated areas wereseen in the allotype. One pair is located at the fore-head and three pairs are located at the posteriorpart of the head. The cilia in each area are stiff andthey arise from a single rectangular cell. The ciliaare not bounded by a common membrane, as in thegastrotrichs, but they move in unison nevertheless.Similar multiciliated cells in other invertebrates arecalled ciliophores (Kristensen and Niilonen, 1982;Eibye-Jacobsen and Kristensen, 1994). The cilio-phores are large epidermal cells with numerous ciliawhose basal bodies are ordered in regular rows. Thehead ciliophores in Limnognathia maerski are lo-cated in the same position as the metastomial cilio-phores in the two interstitial polychaetes Diurodri-lus westheidei and Neotenotrocha sterreri.
8 R. M. KRISTENSEN AND P. FUNCH

The dominating ventral ciliation in Limnognathiamaerski is located on the thorax and the abdomen.Two rows of trunk ciliophores exist. The length ofeach ciliophore is about 5 mm and the width is about15 mm. The cilia are ordered in four rows. There areabout 20 cilia in each row. In the allotype there exist18 pairs of trunk ciliophores, ten pairs on the thoraxand eight on the abdomen. There is no differencebetween the ciliophores of the thorax and those ofthe abdomen.
Just behind the last abdominal ciliophores is alarge adhesive ciliated pad (Fig. 2, ad). The pad
consists of two paired groups of five cells. The cilia inthe pad are also stiff, but they are longer than thecilia in the ciliophores and they do not beat in uni-son. In the strongly squeezed paratype (Fig. 4, gs), asecretion is squeezed out from a pore midventralbetween the two clusters of cells. We assume thesecretion is adhesive. This assumption was sup-ported by live observations of the allotype. The ani-mal stuck to the substrate with the posterior ventralpart when we tried to remove it with a Pasteur pipette.
The action of the trunk ciliophores was also ob-served in the living allotype. The trunk ciliophores
Figs. 3, 4. Limnognathia maerski nov. sp. et nov. sp. Micrographs (DIC) of live specimens. Fig. 3: Habitus photo of the whole animaldivided into head (he), thorax (th), and abdomen (ab). Note the different size of the oocytes (oo1 and oo2). br, brain; gc, midgut cells;ja1, ventral jaw; ph, pharyngeal apparatus. Fig. 4: Ventral view of strongly squeezed caudal part. ad, adhesive ciliated pad consistingof ten ciliated cells (ce); bb, basal bodies in rows; ep, unciliated epidermal cells; gs, glue secretion; oo1, oocyte; tc; trunk ciliophore.
9MICROGNATHOZOA: A NEW CLASS
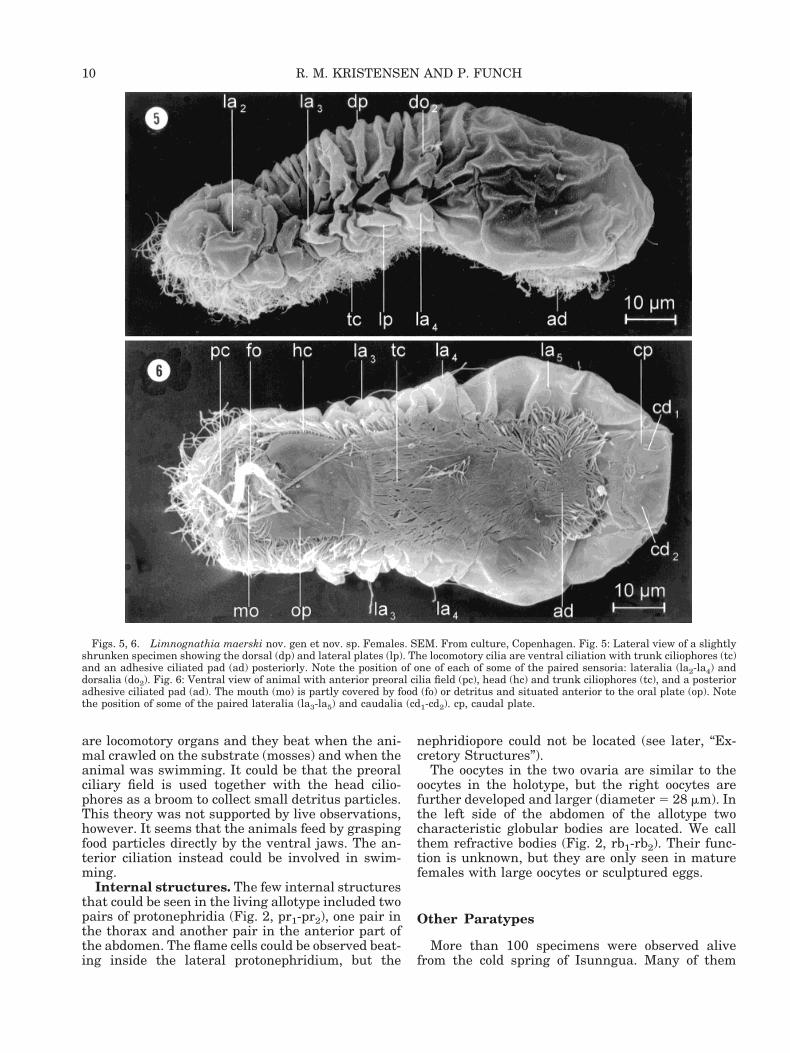
are locomotory organs and they beat when the ani-mal crawled on the substrate (mosses) and when theanimal was swimming. It could be that the preoralciliary field is used together with the head cilio-phores as a broom to collect small detritus particles.This theory was not supported by live observations,however. It seems that the animals feed by graspingfood particles directly by the ventral jaws. The an-terior ciliation instead could be involved in swim-ming.
Internal structures. The few internal structuresthat could be seen in the living allotype included twopairs of protonephridia (Fig. 2, pr1-pr2), one pair inthe thorax and another pair in the anterior part ofthe abdomen. The flame cells could be observed beat-ing inside the lateral protonephridium, but the
nephridiopore could not be located (see later, “Ex-cretory Structures”).
The oocytes in the two ovaria are similar to theoocytes in the holotype, but the right oocytes arefurther developed and larger (diameter 5 28 mm). Inthe left side of the abdomen of the allotype twocharacteristic globular bodies are located. We callthem refractive bodies (Fig. 2, rb1-rb2). Their func-tion is unknown, but they are only seen in maturefemales with large oocytes or sculptured eggs.
Other Paratypes
More than 100 specimens were observed alivefrom the cold spring of Isunngua. Many of them
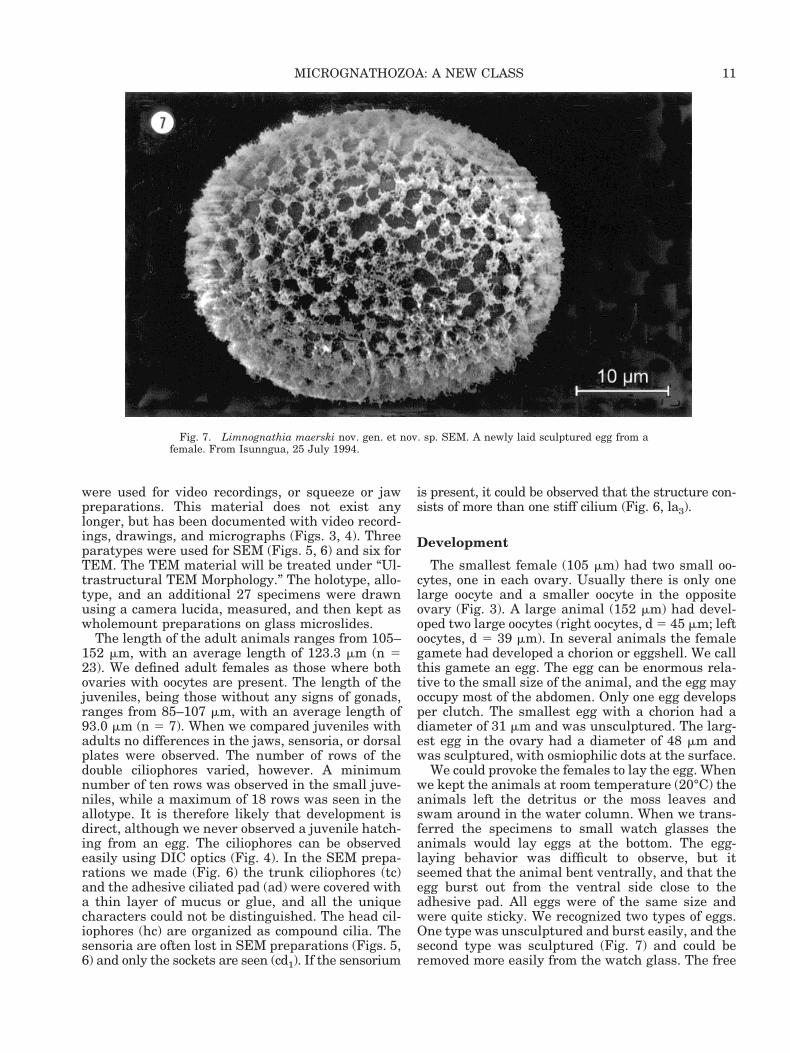
Figs. 5, 6. Limnognathia maerski nov. gen et nov. sp. Females. SEM. From culture, Copenhagen. Fig. 5: Lateral view of a slightlyshrunken specimen showing the dorsal (dp) and lateral plates (lp). The locomotory cilia are ventral ciliation with trunk ciliophores (tc)and an adhesive ciliated pad (ad) posteriorly. Note the position of one of each of some of the paired sensoria: lateralia (la2-la4) anddorsalia (do2). Fig. 6: Ventral view of animal with anterior preoral cilia field (pc), head (hc) and trunk ciliophores (tc), and a posterioradhesive ciliated pad (ad). The mouth (mo) is partly covered by food (fo) or detritus and situated anterior to the oral plate (op). Notethe position of some of the paired lateralia (la3-la5) and caudalia (cd1-cd2). cp, caudal plate.
10 R. M. KRISTENSEN AND P. FUNCH
were used for video recordings, or squeeze or jawpreparations. This material does not exist anylonger, but has been documented with video record-ings, drawings, and micrographs (Figs. 3, 4). Threeparatypes were used for SEM (Figs. 5, 6) and six forTEM. The TEM material will be treated under “Ul-trastructural TEM Morphology.” The holotype, allo-type, and an additional 27 specimens were drawnusing a camera lucida, measured, and then kept aswholemount preparations on glass microslides.
The length of the adult animals ranges from 105–152 mm, with an average length of 123.3 mm (n 523). We defined adult females as those where bothovaries with oocytes are present. The length of thejuveniles, being those without any signs of gonads,ranges from 85–107 mm, with an average length of93.0 mm (n 5 7). When we compared juveniles withadults no differences in the jaws, sensoria, or dorsalplates were observed. The number of rows of thedouble ciliophores varied, however. A minimumnumber of ten rows was observed in the small juve-niles, while a maximum of 18 rows was seen in theallotype. It is therefore likely that development isdirect, although we never observed a juvenile hatch-ing from an egg. The ciliophores can be observedeasily using DIC optics (Fig. 4). In the SEM prepa-rations we made (Fig. 6) the trunk ciliophores (tc)and the adhesive ciliated pad (ad) were covered witha thin layer of mucus or glue, and all the uniquecharacters could not be distinguished. The head cil-iophores (hc) are organized as compound cilia. Thesensoria are often lost in SEM preparations (Figs. 5,6) and only the sockets are seen (cd1). If the sensorium
is present, it could be observed that the structure con-sists of more than one stiff cilium (Fig. 6, la3).
Development
The smallest female (105 mm) had two small oo-cytes, one in each ovary. Usually there is only onelarge oocyte and a smaller oocyte in the oppositeovary (Fig. 3). A large animal (152 mm) had devel-oped two large oocytes (right oocytes, d 5 45 mm; leftoocytes, d 5 39 mm). In several animals the femalegamete had developed a chorion or eggshell. We callthis gamete an egg. The egg can be enormous rela-tive to the small size of the animal, and the egg mayoccupy most of the abdomen. Only one egg developsper clutch. The smallest egg with a chorion had adiameter of 31 mm and was unsculptured. The larg-est egg in the ovary had a diameter of 48 mm andwas sculptured, with osmiophilic dots at the surface.
We could provoke the females to lay the egg. Whenwe kept the animals at room temperature (20°C) theanimals left the detritus or the moss leaves andswam around in the water column. When we trans-ferred the specimens to small watch glasses theanimals would lay eggs at the bottom. The egg-laying behavior was difficult to observe, but itseemed that the animal bent ventrally, and that theegg burst out from the ventral side close to theadhesive pad. All eggs were of the same size andwere quite sticky. We recognized two types of eggs.One type was unsculptured and burst easily, and thesecond type was sculptured (Fig. 7) and could beremoved more easily from the watch glass. The free
Fig. 7. Limnognathia maerski nov. gen. et nov. sp. SEM. A newly laid sculptured egg from afemale. From Isunngua, 25 July 1994.
11MICROGNATHOZOA: A NEW CLASS

egg is oval (30 3 40 mm) and other observed eggsranged in size from 40–60 mm at the longest axis.The unsculptured eggs could be abortive, while thesculptured eggs were capable of further develop-ment. The justification for this assumption is thatthe whole population of adult females would deliverthe oocytes and eggs when they were kept at hightemperature. We did not observe any cleavage in theeggs. Furthermore, we never found eggs with jaws ofjuveniles inside.
We looked very intensively for smaller males bothin Greenland and in the culture in Copenhagen.Unfortunately, we never observed any, and there-fore, we expect that the species reproduces by par-thenogenesis, at least during the summer. We can-not rule out that the animal is hermaphroditic. Thetwo refractive bodies (Fig. 2, rb) may be a part of themale reproductive system. The smallest juvenile (85mm) was observed alive. The head was relativelylarge (35 mm) and the thorax weakly developed. Asmentioned above, it had only ten rows of ciliophoresand it may be the ciliophores of the thorax that arelacking. It moved exactly like the adults and it al-ready had a full set of jaws in the pharyngeal appa-ratus. The animals were kept in a moss culture in a4°C refrigerator for more than 2 years. During thisperiod only seven juveniles were observed and mea-sured.
Fine Structure of the Jaw Apparatus
The fine structure of the “true” cuticularized partsof the pharyngeal apparatus is important for a dis-cussion of the phylogenetic position of Limnogna-thia. We therefore describe the isolated cuticular-ized parts (sclerites) of the pharyngeal apparatus inthis section.
A complete investigation of the ultrastructure ofthe whole pharyngeal apparatus, e.g., cuticular ele-ments, epidermal cells, and the mesodermal cross-striated muscles, is needed. The only way to do thatis to combine SEM and TEM techniques (Fiege,1990). A less sophisticated technique is to serial-section the entire pharyngeal apparatus and makethree-dimensional reconstructions of the location ofall cuticular elements, the nuclei of the epidermalcells, and the cross-striated muscles. This techniquehas been used with great success in the eutardi-grades (Eibye-Jacobsen, 1997). Unfortunately, thistechnique is time-consuming and requires single-hole grids for serial sectioning. We sectioned sixpharyngeal apparatuses, but we used the more se-cure 200-mesh grids. The additional information ob-tained from the TEM study was crucial to under-standing the general organization of the pharyngealapparatus; for example, that the cuticularized partsare extracellular and that the jaw apparatuses arebuilt in the same way as the jaws of gnathostomu-lids. The results from our TEM study of the pharyn-
geal apparatus will be treated under the section onultrastructure.
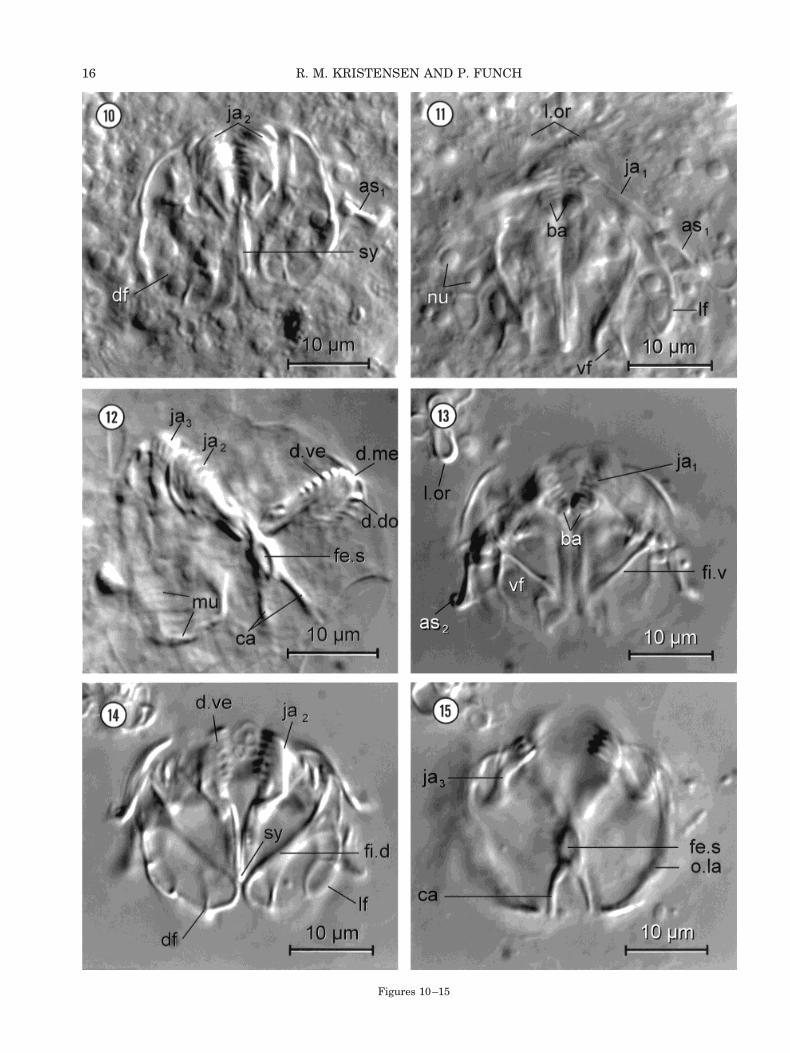
In this section, we use the technique described byRiedl and Rieger (1972) for gnathostomulids andthereafter combine light microscopical observationswith SEM micrographs. We first observed living,squeezed animals by DIC optics (Figs. 10,11). A fewanimals were treated with OsO4-vapor and werestrongly squeezed (Fig. 12). This allowed us to seethe dentarium and articularium of the main jaws(ja2). Soon it became clear that some parts of thecuticular jaw apparatus were hidden in the epider-mal tissues and the muscles. In order to reveal thehidden parts, whole animals were treated with 2%sodium hypochlorite (Figs. 13–15). This bleachingprocess allowed us to see the main cuticular parts,the basal plates, the three pairs of jaws, and thecomplex structure of the fibularium with its manyfenestrae and fibulae. These structures are stronglyresistant to the bleaching process. Unfortunately,all ligaments, the oral plate, and the lamellae oralesdissolved immediately. Consequently, the ventraljaws (ja1) and the accessory sclerites (as1, as2) werelost in all sodium hypochlorite preparations. It soonbecame obvious that the fine structure of the cutic-ularized pieces was too small and complex to be fullyresolved with light microscopy. A modified SEMtechnique of Koehler and Hayes (1969) was there-fore introduced (Figs. 16, 17). In this technique thesoft tissue was also removed with sodium hypochlo-rite. The three-dimensional configurations wereadded to the two schematic drawings (Figs. 8, 9),which were made with the camera lucida technique.When the DIC micrographs (Figs. 10–15) were com-pared with the SEM micrographs (Figs. 16, 17), itwas obvious that both techniques have their advan-tages. When we used DIC it was possible to observethe living animals first and later to treat the ani-mals with sodium hypochlorite. This affords the op-portunity to observe the cuticular elements in situattached to epidermal cells, ligaments, and muscles.We never observed all cuticular parts together usingSEM and the thin lamellae of the fibularium weretwisted, so it gave the impression that the structureis much more flattened compared to similar struc-tures in living specimens.
On the other hand, many details were seen onlywith SEM and these structures are still not wellunderstood, e.g., the free teeth between the mainjaws (ja2) and the dorsal jaws (ja3), the pseudodentes(p.de), the trochanter (tr), and the spinula (sp) of thedorsal jaws (see Figs. 16, 17). We have to admit thatour interpretation is preliminary and we may havemisinterpreted several structures, but otherwise wehope the readers will understand that the complex-ity of the jaw apparatus of Limnognathia maerskiis far beyond what is seen in other invertebratessuch as gnathostomulids, rotifers, and dorvilleidpolychaetes. For a comparison of the fine structure
12 R. M. KRISTENSEN AND P. FUNCH

of their jaw apparatuses and that of Limnognathia,see below.
We used a new terminology for the more impor-tant cuticular parts of the jaw apparatus. We em-ployed the Latin nomenclature in many generalstructures, such as apophysis, fenestra, trochanter,etc. Unfortunately, this nomenclature has been usedin gnathostomulids, and to some extent in rotifers aswell. We are still not sure if these structures arehomologous in the three groups, but the ultrastruc-ture of some of the cuticular elements (Rieger andTyler, 1995) indicates that this is the case. Thefollowing description of the main elements 1) thebasal plates, 2) the lamellae orales, 3) the ventraljaws, 4) the main jaws, 5) the two fibularia, and 6)the dorsal jaws is given from the ventral perspectiveto the dorsal view (Figs. 8–17).
Basal Plates (ba)
A pair of molar-like structures (Figs. 9, 16, ba) islocated on the ventro-caudal part of the mouth open-ing. These two structures can be extruded out fromthe lower lips when the animal forages. The molarstructure is triangular in shape, with five cusps ofheavily sclerotized material. The two molars are thefirst elements to be seen in optical sections with DICtechniques (Figs. 11, 13) in ventral view. Each molaris 2 mm wide and 1.5 mm high. The outer cuticularpart of the basal plates is fixed to thin jointed la-mella plates, which continue inside the lower part ofthe mouth cavity. The thin plates were not observedwith DIC optics, but were clearly seen in specimenstreated with sodium hypochlorite (Figs. 8, 9, 16, 17).The thin lamellar parts are 4 mm long and are fusedrostrally with a 1.5 mm-long suture. Two large cross-striated muscles seem to attach close to the molarportion of the basal part. In connection with thebasal plates, five additional dentes oralis (de.o) areseen in the SEM micrographs (Figs. 16, 17). Thesestructures were not observed by DIC optics and wereconsequently omitted from Figure 9. They couldhave been hidden by the outer cuticular rim of themouth opening itself.
Lamellae Orales
Lamellae orales were observed only in living ani-mals and in the TEM micrographs. They consist oftwo halfcone-shaped structures on the rostral part ofthe mouth cavity. In ventral optical sections thelamellae orales are seen as two arched upper lips(Fig. 11). These structures may constitute a contin-uous, folded cuticular membrane covering the upperpart of the mouth cavity. By DIC techniques (Fig.11) the lamellae orales are seen as two arcs withdelicate striation. At least 13 folds are observed ineach arc. The lamellae orales support the dorsalmouth cavity and prevent cavity collapse duringfeeding. The lamellae orales quickly disappear in
the sodium hypochlorite bleaching process and thecollapsed lamellae can be seen as an extra,membrane-like structure outside the cleaned jawapparatus (Figs. 13–15).
Ventral Jaws (Pseudophalangia)
The ventral jaws consist of two pairs of stronglycuticularized elements, the pseudophalangia (ja1)and the accessory sclerites (as1). These two elementsare joined with a ligament and a ball-and-socketjoint. Furthermore, a ligament connects the pseu-dophalangium to the fibula ventralis of the fibu-larium. Both ligaments and the large cross-striatedmuscle disappeared when sodium hypochlorite wasadded, and the pseudophalangium was found inde-pendent from the rest of the jaw apparatus. Theaccessory sclerite, on the other hand, stays attachedto the lateral part of the fibularium (Figs. 13, 14, 16,17), even after the pseudophalangium disappears.The link between the pseudophalangium, the fibu-larium, and the accessory sclerite is much more com-plicated than we have drawn in the squeezed prep-aration of the jaw apparatus (Figs. 8, 9).
The pseudophalangium is a large sclerite and itcan be quite moveable, facilitated by the large cross-striated muscles. The muscle attachment is formedas a fenestra pseudophalangialis (fe.p) at the swol-len base of the sclerite. Both pseudophalangia can beprotruded through the mouth opening under forag-ing. In relaxed swimming behavior the two pseu-dophalangia would be located latero-rostral of themouth opening (Figs. 8, 9). The tip of each pseu-dophalangium consists of four large digits; in a fewspecimens a fifth smaller digit also was observed asa dorsal thumb. The length of the pseudophalan-gium is 12 mm and the length of the accessory scle-rite is 5 mm. Both structures are a hollow tube-likestructure in cross section (see below).
Main Jaws
The main jaws are pincer-like. The pincer consistsof the paired dentarium and the unpaired articu-larium with the symphysis. Dorso-caudally the mainstructure again is split into a symmetrical cauda(Figs. 8, 9, 12, 15–17, ca), which is the apodeme forthe posterior pharyngeal muscle sac. Ventro-caudally the symphysis (Figs. 9, 17, sy) continues asan unpaired element. The fibularium, the largestelement in the jaw apparatus, attaches to the mainjaws laterally. The fibularium could be regarded as apart of the main jaws, but will be treated separatelybecause of its complexity.
The dentarium is the teeth-bearing distal partof the pincer. Each branch of the pincer has anarc-shaped arrangement of teeth which can beobserved from a lateral view (Fig. 12). The ventralarc (arcus ventralis) is prominent and possessesfive large teeth, the dentes ventrales (Fig. 9, d.ve).
13MICROGNATHOZOA: A NEW CLASS
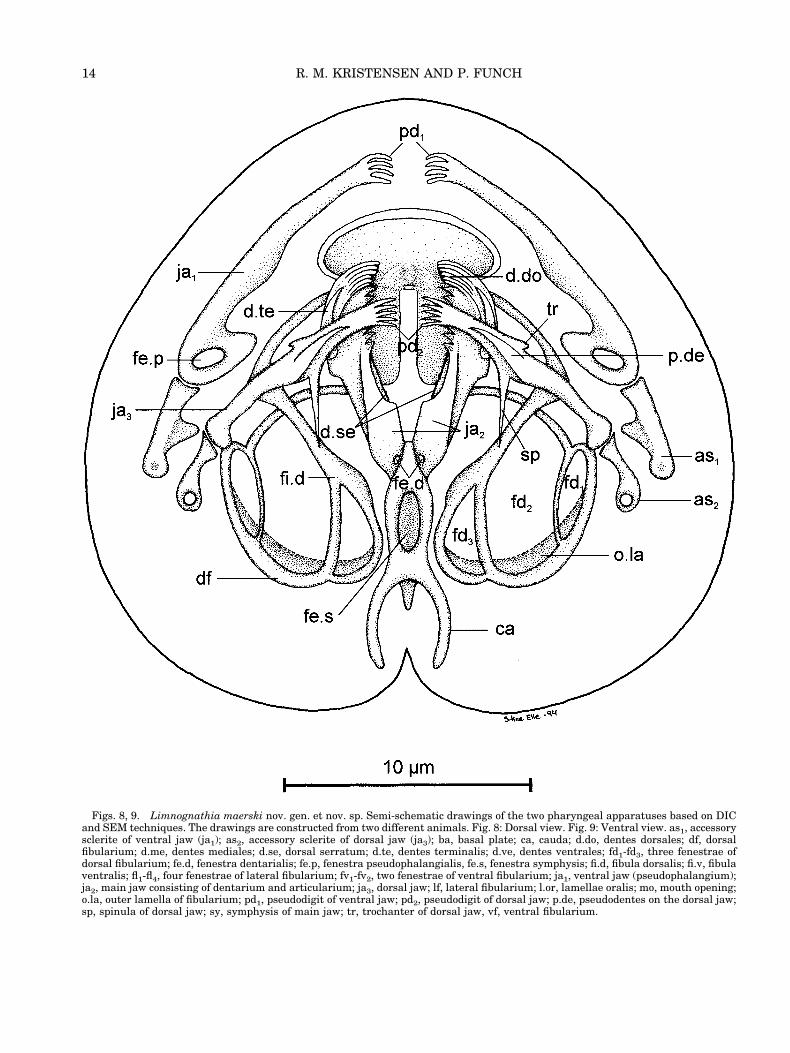
Figs. 8, 9. Limnognathia maerski nov. gen. et nov. sp. Semi-schematic drawings of the two pharyngeal apparatuses based on DICand SEM techniques. The drawings are constructed from two different animals. Fig. 8: Dorsal view. Fig. 9: Ventral view. as1, accessorysclerite of ventral jaw (ja1); as2, accessory sclerite of dorsal jaw (ja3); ba, basal plate; ca, cauda; d.do, dentes dorsales; df, dorsalfibularium; d.me, dentes mediales; d.se, dorsal serratum; d.te, dentes terminalis; d.ve, dentes ventrales; fd1-fd3, three fenestrae ofdorsal fibularium; fe.d, fenestra dentarialis; fe.p, fenestra pseudophalangialis, fe.s, fenestra symphysis; fi.d, fibula dorsalis; fi.v, fibulaventralis; fl1-fl4, four fenestrae of lateral fibularium; fv1-fv2, two fenestrae of ventral fibularium; ja1, ventral jaw (pseudophalangium);ja2, main jaw consisting of dentarium and articularium; ja3, dorsal jaw; lf, lateral fibularium; l.or, lamellae oralis; mo, mouth opening;o.la, outer lamella of fibularium; pd1, pseudodigit of ventral jaw; pd2, pseudodigit of dorsal jaw; p.de, pseudodentes on the dorsal jaw;sp, spinula of dorsal jaw; sy, symphysis of main jaw; tr, trochanter of dorsal jaw, vf, ventral fibularium.
14 R. M. KRISTENSEN AND P. FUNCH
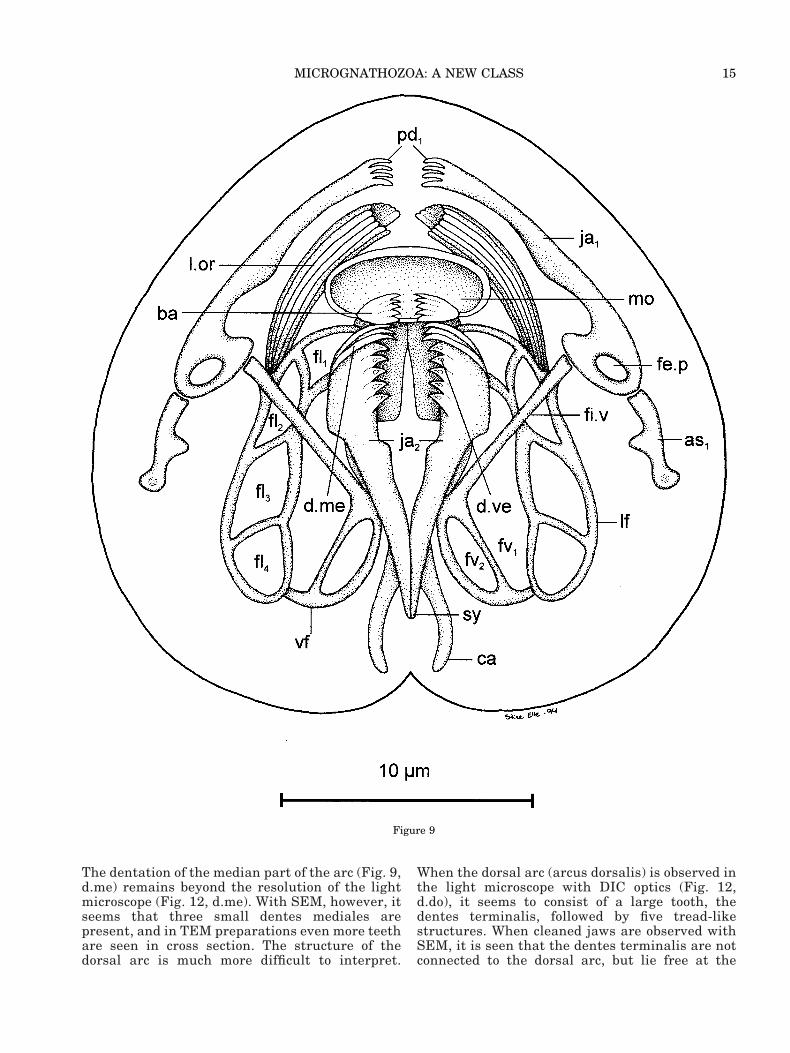
The dentation of the median part of the arc (Fig. 9,d.me) remains beyond the resolution of the lightmicroscope (Fig. 12, d.me). With SEM, however, itseems that three small dentes mediales arepresent, and in TEM preparations even more teethare seen in cross section. The structure of thedorsal arc is much more difficult to interpret.
When the dorsal arc (arcus dorsalis) is observed inthe light microscope with DIC optics (Fig. 12,d.do), it seems to consist of a large tooth, thedentes terminalis, followed by five tread-likestructures. When cleaned jaws are observed withSEM, it is seen that the dentes terminalis are notconnected to the dorsal arc, but lie free at the
Figure 9
15MICROGNATHOZOA: A NEW CLASS
Figures 10–15
16 R. M. KRISTENSEN AND P. FUNCH

surface (Fig. 16, d.te). We still believe it is thesame structure as seen by DIC optics, because thedentes dorsalis (d.do) are also located superficiallyas thin tread-like teeth on the dorsal arc.
Caudal to the arc-structure, the dorsal edge of thedentarium has a serrated appearance. This dorsalserratum (Figs. 8, 16, d.se) consists of 20 small sawteeth on each side of the pincer-arms. Just before thesymphysis the dentarium has a pair of small holes(Figs. 8, 16, fe.d). The symphysis continues ventrallyas an unpaired caudal part (Figs. 9, 14, 17, sy). Thedorsal part of the articularium is more complicated.Behind the articulation of the two arms in the den-tarium, is a 4 mm-long single piece of the articu-larium. This lamella structure is penetrated by alarge hole, the fenestra symphysis (Figs. 8, 12, 15,16, fe.s). The articularium has two symmetricalcauda (ca). We believe that this structure is theapodeme for the large caudal muscle-sac.
The Two Fibularia
The fibularium consists of cuticularized fibulaeand fenestrae. Each fenestra is a window with cel-lular tissue surrounded by cuticle. The cellular partis involved in forming the extracellular fibulae. Theunique fibularium is perhaps only a part of the ap-ophysis of the main jaws as seen in some gnatho-stomulids and rotifers, but in Limnognathia thestructure has developed to an extreme degree. Thefibularium in Limnognathia is a three-dimensionalstructure that totally surrounds the main jaws asthree compartments: the ventral, lateral, and dorsalfibularia.
The ventral fibularium is characterized by astrong fibula ventralis (Figs. 9, 13, fi.v). This fibularuns across the ventral fibularium as a straight barand it connects the basal parts of the pseudophalan-gium. The epidermal cells in the ventral fibulariumhave large nuclei and abundant cytoplasm (Figs. 10,11). There are at least five ventral fenestrae withlarge cells. The lateral fibularium has four fenestraewhose cells have large nuclei. Each nucleus nearlyfills the lateral fenestra (Fig. 11). The fibulae of thelateral fibularium are thin. We could only recognizethree fibulae of the dorsal fibularium. The fibuladorsalis (Figs. 8, 14, 16, 17, fi.d) is thick and stronglysclerotized.
The outer limit of the fibularium consists of athick lamella that can be seen as a nearly circulardorsal structure (o.la) by DIC optics (Fig. 15) or as arobust structure by SEM (Figs. 16, 17). The fibu-larium is attached to the dentarium rostrally but isembedded in the cellular tissue as an apophysialstructure in its caudal part.
Dorsal Jaw System
The pharyngeal channel passes through the twoarcs of the main jaws and continues dorso-caudallyto a short esophagus. Before entering the esophagusthe food particles must pass through the third jawsystem (ja3). The dorsal jaws are in many wayssimilar to the pseudophalangia. These dorsal cutic-ular structures consist of a pair of arm-shaped rods(Figs. 8, 15–17, ja3). Rostrally, each arm has fourlarge digits, a smaller dorsal thumb-like digit, andseveral small teeth, which could not be seen by lightmicroscopy. The caudal part of the dorsal jaw alsoextends a ligament to an accessory sclerite (Fig. 8,as2). The ligament disappears with sodium hypo-chlorite treatment and the pair of accessory scleritesis therefore missing in the SEM preparation (Figs.16, 17).
The dorsal jaws are surely serially homologouswith the ventral jaws, but have a more complicatedstructure. The jaws have a dorsal trochanter (tr) forthe attachment of a large striated muscle and amidventral spinula (sp) attaching the fibula dorsalis(fi.d). Furthermore, the caudal part is attached tothe outer lamina of the fibularium (Fig. 16, o.la). Thepaired dorsal jaw system is also connected to thefibularium in the sodium-hypochlorite-treated ani-mals (Figs. 15–17).
Ultrastructural TEM Morphology
The ultrastructural observations by TEM arebased on trialdehyde-fixed materials (Figs. 18–33).A few artifacts were clearly seen by TEM. The spa-cious body cavity between the digestive system andthe epidermal cells (see Figs. 18, 22) is an artifact ofinappropriate fixation. The same osmotic problemmay be seen in the dorsal epidermal cells with the
Figs. 10–15. Limnognathia maerski nov. gen. et nov. sp. DICmicrographs of jaws. Figs. 13–15. The jaw apparatus of the sameanimal as Figure 11 treated with sodium hypochlorite. The dis-solved lamellae oralis (l.or) is seen at the upper left corner in allthree photos. Fig. 10: Living animal, middorsal optical section,strongly squeezed preparation. The dorsal fibularium (df) is at-tached to the main jaws (ja2). Note the symphysis (sy) of thearticularium in the middle and the accessory sclerite (as1) of theventral jaw to the right. Fig. 11: Living animal, ventral opticalsection focused on ventral jaws (ja1) with the accessory sclerite(as1), lamellae oralis (l.or), and basal plates (ba). The fenestrae ofboth the ventral (vf) and lateral fibularium (lf) contain severalnuclei (nu). Fig. 12: Strongly squeezed, osmium-fixed animal. Tothe left a large cross-striated muscle (mu) seems to attach to thedorsal jaw (ja3). The main jaws (ja2) are twisted. The arcs of thedentarium with teeth (d.do, d.me, d.ve) are seen to the right. Thearticularium with a large fenestra symphysis (fe.s) and the twosymmetrical arms of the cauda (ca) are recognized. Fig. 13: Ven-tral optical section focused on the basal plates (ba), ventral jaw(ja1), accessory sclerite (as2), and the strong fibula ventralis (fi.v)in the ventral fibularium (vf). Fig. 14: Midventral optical sectionfocused on the ventral arc of dentarium with teeth (d.ve), thesymphysis (sy) of the articularium, the lateral (lf) and the dorsalfibularium (df) with the strong cutilarized fibula (fi.d). Fig. 15:Dorsal optical section focused on the dorsal jaw (ja3), the outerlamella (o.la) of the fibularium and the symmetrical arms of thecauda (ca). fe.s, fenestra symphysis.
17MICROGNATHOZOA: A NEW CLASS

Figs. 16, 17. Limnognathia maerski nov. gen. et nov. sp. SEM. Jaw apparatus treated with sodium hypochlorite. The dorsalfibularium (df) is twisted ventrally compared to the view in Figure 14. Note also: The ventral jaws (pseudophalangia), the accessorysclerite, and lamellae oralis are lacking. Fig. 16: Dorsal view. Fig. 17: Lateral view. ar, articularium of main jaws (ja2); as1, accessorysclerite to ventral jaw (which is lacking); ba, basal plates; ca, cauda; de, dentarium of main jaws (ja2); de.o, dentes oralis; d.se, dorsalserratum; d.te, dentes terminales; fd1-fd3, fenestrae of dorsal fibularium; fe.d, fenestra dentarialis; fe.s, fenestra symphysis; fi.d, fibuladorsalis; ja2, main jaws consisting of articularium (ar) and dentarium (de); ja3, dorsal jaw with pseudodentes (p.de), trochanter (tr) andspinula (sp); o.la, outer lamella; sy, symphysis.
18 R. M. KRISTENSEN AND P. FUNCH

plates and the nearly mature egg (Fig. 22). The thickglycocalyx may also become separated from the ven-tral ciliated cells, but otherwise the trialdehyde fix-ation was excellent, especially for low magnifica-tions of different cell structures.
Integumentary Structures
The main integumentary structures in all adultfemales can be divided into the nonciliated dorsalepidermis (dorsal and lateral plates) with an intra-cellular matrix layer and into the ventrallocomotory/feeding organ with multiciliated epider-mal cells covered with a glycocalyx. The glycocalyxmay be thin and simple or, as on the adhesive cili-ated pad, thick and complex. Furthermore, the epi-dermal sensory cells (sensoria) consist of monocili-ary to multiciliary cells without a cuticle. The onlyexternal cuticular structure is the ventral oral plate,which is formed by nonciliated epidermal cells.Other epidermal cells in the pharyngeal apparatusproduce hard cuticular parts (Fig. 23), but the ultra-structure of these is treated in the section “DigestiveSystem,” below. All epidermal structures are non-syncytial. Syncytia were not observed at all in Lim-nognathia maerski.
Dorsal and Lateral Intracellular Plates
The dorsal epidermis in Limnognathia maerski iscellular, unciliated, and generally 2–5 mm thick (Fig.18, ep). In the sutures between the plates the epi-dermis is much thinner, being less than 0.2 mmthick. A very thin glycocalyx (120 nm) covers the cellmembranes of the epidermis (Fig. 21). Cell con-stancy (eutely) might be present in the epidermis.Two to four cells form each dorsal plate. The pro-nounced apical plate seems to be formed by a singlelarge epidermal cell. The cell borders can be seen byDIC optics through the plate structure as dottedlines (Fig. 1). The dotted structure can be inter-preted by TEM observations. It consists of two junc-tions (Fig. 26, dj) with a distance of about 0.5 mm,where the two cells form some interdigitations. Thecell processes from one cell extend deeply into theapposing cell. Two other junctional complexes arepresent between neighboring epidermal cells. Be-tween the two middorsal epidermal cells, whichform the first dorsal plate in the thoracic region (Fig.18, dp), a very characteristic cell junction complex ispresent (Fig. 21). Distally, the two cells form a largeintercellular space (ic) distal to a gap junction,which can be up to 0.5 mm long. A unique type ofseptate junction is present between several dorso-lateral epidermal cells (Fig. 19, zj) on the bordersbetween plates (see review, Green and Bergquist,1982). The junction consists of three to sevenbridges, which open up like a zipper when the ani-mal is treated with sodium hypochlorite, and theplates are separating. We named this new type of
junction the zipper junction. The nucleus of the dor-sal epidermal cells is round to oval and has hetero-chromatin close to the nuclear membrane. The cyto-plasm in these epidermal cells is osmiophobic, withfew vesicles and mitochondria (Figs. 18, 19).
The dorsal and lateral plates are situated insidethe epidermal cells. A plate consists of a conspicuousintracellular matrix layer (Figs. 19, 21, im). Thematrix layer can be differentiated into a stronglyosmiophilic outer layer and a less osmiophilic innerlayer. The intracellular-matrix layer may range inthickness from 0.1–0.3 mm. It has been very difficultto observe the outer cell membrane of the epidermalcell (Figs. 19, 26), but with high magnification thecell membrane is seen as a typical unit membraneconsisting of two osmiophilic layers with an osmio-phobic layer in between (Fig. 21, cm). The platestructure is clearly located beneath the cell mem-brane, and is therefore intracellular.
The lateral plates are formed like the dorsalplates (Figs. 18, 20, lp) but they seem to be moreflexible than the thick dorsal plates. The lateralplate continues to the ventral side, where the borderbetween the plate and the ciliated ventrum is easilyobserved (Fig. 20). Usually, the lateral epidermisforms one to two folds in the junction between theplate structure and the stiff, flat ventrum.
Ventral Ciliated Epidermis
The ventrum has four quite different ciliatedstructures: the preoral cilia field, the head cilio-phores, the trunk ciliophores, and the adhesive cili-ated pad (Figs. 2, 5, 6, 18, 22). The preoral ciliaryfield consists of five rows of single cilia on both sidesof the head and two rows of fronto-lateral rows ofshorter cilia. The ciliary roots are relatively short.These cilia are associated with a labyrinth of extra-cellular membranes. Similar membrane structuresare seen in the pharyngeal apparatus associatedwith the pharyngeal cilia. The cilia may work as abroom during foraging, but they are clearly alsoinvolved in swimming behavior. The four pairs ofhead ciliophores are located lateral to the oral plate.These ciliophores consist of rectangular cells withstiff compound cilia. The ciliary roots are relativelylong.
The pairs of 18 trunk ciliophores are a uniquecharacter for Limnognathia maerski. They are thelocomotory organ when the animal is creeping on thesubstrate. Each cell has four rows of cilia, each withabout 20 stiff compound cilia. The beat of one row ofcompound cilia is synchronized. Each cilium is sur-rounded at its base by an extracellular ring of cuti-cle. The same feature is seen in the adhesive ciliatedpad (Fig. 32, cu). Often, four ciliophores are seen incross section. There exist only two rows of cells, butthey overlap slightly, so the distal part of the ante-rior row is also cut. The ciliophores are all coveredwith a thick glycocalyx (Fig. 18, gx).
19MICROGNATHOZOA: A NEW CLASS

Figures 18–21
20 R. M. KRISTENSEN AND P. FUNCH

The adhesive ciliated pad consists of ten cells (Fig.4). The cilia are ordered in register like those of thetrunk ciliophores. The cross-striated ciliary rootpasses through the entire cell. The ciliary root mayattach to the nucleus, which lies proximally in thecell. The glycocalyx is thick and is ordered in acharacteristic network of osmiophilic granules withthin fibers (Fig. 32, gx).
Oral Plate
The oral plate was not visible by DIC optics in theliving animals observed in Greenland. The jawstructure and the pharyngeal bulb completely over-shadowed the outer cuticular plate. Later, the thinbut large oral plate was discovered using SEM (Fig.6, op) and TEM (Fig. 25, op). The oral plate is veryflexible and can be transformed from a flat plate intoa cone-shaped structure when the ventral jaws areprotruded. The internal part of the oral plate isosmium-negative, as are the main elements in thejaws. The tubular subunits are seen between twounit membranes similar to plasma membrane (Fig.27, tu).
Sensory Structures
The tactile bristles consist of stiff cilia arisingfrom a ring-shaped socket (Fig. 5, do2, la3). Thisreinforcement consists of a thickening in the intra-cellular matrix layer. The pore in the middle wherethe cilia arise is naked, covered only with the cellmembrane. The cilium may be surrounded by veryshort microvilli. It seems likely that all stiff cilialack the central doublet of microtubules. There maybe only one stiff cilium or up to three long, stiff, andadjoined cilia, as in the lateralia. Other sensorystructures comprising cilia are located around themouth opening. The ciliary structures in the phar-
ynx (Figs. 22, 23, ci) may also have a sensory func-tion.
A pair of round vesicular structures in front of thehead was observed both in the allotype (Fig. 2) andin several paratypes (Fig. 40). They are osmiophilicand may be lipoid eye granules. The granules disap-pear in wholemount preparation, but an eye struc-ture similar to a phaosome was observed with TEMjust in front of the pharyngeal structure.
Glands and Secretion
Limnognathia maerski is poor in epidermalglands. The only epidermal gland complex is associ-ated with the adhesive ciliated pad. It is located inthe tail (Fig. 29, ag) and consists of two cells filledwith large vesicles. These cells are large, up to 10mm. The substructure in the glands does not resem-ble the duo-gland systems in other meiofaunal ani-mals; it looks more like mucous glands as found inkinorhynchs. The glands seem to open just behindthe adhesive ciliated pad. In a strongly compressedanimal the glandular material is extruded (Fig. 4,gs). The glands related to the digestive system willbe described in that section.
Nervous SystemBrain. The large cerebral ganglion occupies most
of the head in front of the pharyngeal apparatus(Fig. 3, br). It is slightly bilobate. The neuronal cellbodies form the outer rim of the brain and surroundthe central neuropile (Fig. 29, br). The basal laminaeare poorly developed in the head, so it is unclearwhether the brain is intra- or subepithelial in posi-tion. The perikarya of the outer rim consist of about175 round cell bodies, each with a large nucleus. Theheterochromatin forms several clusters inside thenucleus. The commissural connections between thetwo brain lobes are prominent. The commissure con-sists of neuropile fibers, which contain vesicles rang-ing in diameter from 40–100 nm. A ventral longitu-dinal nerve cord extends from each brain lobe. Othernerve cords were not observed, but they may bepresent because nerves were observed in the pha-ryngeal apparatus.
Ventral nerve cords. A pair of ventral longitu-dinal nerve cords extends lateroventrally to the pha-ryngeal apparatus. Each consists of about 15–17nerve fibers filled with neurovesicles (Fig. 20). Theventral nerve cords are deeply submerged and theyseem to be subepidermal, but again the basal laminais hard to locate. It is therefore very difficult to statethe position of the whole nervous system. The twonerve cords may be associated with a double gan-glion in the thorax and a caudal ganglion close to thetail (pygidium).
Figs. 18–21. Limnognathia maerski nov. gen. et nov. sp.TEM. Fig. 18: Cross section of the animal just behind the pha-ryngeal apparatus. Note the large vesicle (ve) and the Golgiapparatus (go) inside the midgut cell (gc). The midgut (mg) lackscilia. The dorsum is covered with epidermal cells (ep) forming thedorsal plates (dp) and lateral plates (lp), while the ventrum con-sists of trunk ciliophores (tc) that are only covered with glycocalyx(gx). The two ventral nerve cords (vn) are located close to thelateral protonephridia (pr). Fig. 19: High magnification of a newtype of septate junction, the zipper junction (zj), between twodorsolateral epidermal cells. The plate, or intracellular matrixlayer (im), is formed inside the epidermal cell (ep). ic, intercellu-lar space. Fig. 20: A close-up of Figure 18 showing a midgut cell(gc), the right protonephridium (pr), and the right ventral nervecord (vn). The arrow indicates the junction between the lateralplate (lp) and the ciliated ventrum that is only covered withglycocalyx (gx). The muscle (mu) is obliquely cross-striated. Fig.21: High magnification of junction between two middorsal epider-mal cells showing the intracellular matrix layer (im) inside theepidermal cell (ep). The two epidermal cells form a gap junction(gj) after a distal intercellular space (ic). The cell membrane (cm)is only covered with a very thin glycocalyx (gx).
21MICROGNATHOZOA: A NEW CLASS

Figs. 22–24. Limnognathia maerski nov. gen. et nov. sp. TEM. Fig. 22: Latero-longitudinal section of the whole animal. Note thepharyngeal apparatus (ph), the salivary gland (sg), the midgut (mg), and nearly mature oocyte of the right side (oo1). The buccal gland(bg) is located dorsal of the pharyngeal lumen. Only a few trunk ciliophores (tc) are cut. Fig. 23: Close-up of the pharyngeal apparatusseen in Figure 22. Note the true cross-striated pharyngeal muscles (pm), the negative staining of the main jaws (ja2), and the cilia (ci)in the pharynx. The epithelial cells (ec) lie in clusters surrounded by the fibularium (fi). Fig. 24: The lumen of the midgut withmicrovilli (mv) from the endodermal cells.

Musculature
The musculature consists of fiber-form musclecells. Myosyncytia are not present. The somaticmuscles consist of several longitudinal fiber cellsattaching to various parts of the trunk and fibersrunning through the entire trunk, attaching in thecaudal end close to the tail. The dorsoventral mus-cles attach through the epidermal cells on the edgeof lateral plates. Circular muscles were not ob-served. Minute muscle fibers attaching one epider-mal plate to another contract the thorax. Such con-tractions were observed in many living animals. Themuscles of the pharyngeal apparatus are classicallycross-striated (Fig. 23, pm), while many of the so-matic muscles in the trunk are obliquely striated(Fig. 20, mu). The number of sarcomeres varies fromone to five in the pharyngeal muscles. The length ofeach sarcomere is about 2 mm. A continuous Z-line isabsent. Instead, there are five to seven Z-discs atsites where the Z-components would be expected.
All the pharyngeal muscles observed attach toepidermal cells (Fig. 25, mu). We never observedmuscles attaching directly to the cuticle. The epider-mal cells may have both microtubules and thread-like fibers in the muscle attachment (Fig. 28, mt).Furthermore, the somatic muscles never attach di-rectly on the dorsal and lateral plates. The connec-tion is always through an epidermal cell (Fig. 26,ep), which contains the intracellular plate structure.The somatic muscles (Fig. 26, mu) may either lack abasal lamina or the lamina is very diffuse and thin.Since embryological data are missing, ideas on theorigin of the musculature are based solely on thelocation of the musculature, proximally to the epi-dermal cells and the basement membrane (two basallaminae). When the basal lamina is lacking or isdiffuse, as in Limnognathia maerski, the origin ofthe musculature remains uncertain. Myoepithelialmuscle cells were not observed either in the bodywall or associated with the sclerites of the pharyn-geal apparatus. We therefore suggest that all mus-cles of L. maerski are of mesodermal origin.
Digestive System
Mouth opening. The oval mouth opening (Figs.2, 9, mo) is located ventrally on the frontal edge ofthe oral plate (Fig. 6, mo). We observed living ani-mals with a slightly protruded mouth cone, whichsuddenly protruded most of the pharyngeal appara-tus out of the mouth (see Discussion). To be capableof that the whole rim of the mouth must be flexible.The flexible part is the upper lip, which can beexpanded enormously. The lower lip is attached tothe basal plate complex and cannot be everted. Amulticellular, ciliary organ surrounds the anteriormargin of the mouth and may, in fact, constitutepart of the buccal cavity (Figs. 25, 27, 29, bc). Thesecilia stick out of the mouth when the animal is
foraging. They do not beat like locomotory cilia, butonly move rigidly. They may be chemoreceptors.
Pharyngeal apparatus. The fine structure ofthe hard part of the jaw apparatus has been de-scribed under “Description.” Here we will deal withthe cellular parts and the ultrastructure of the solidparts of the pharyngeal apparatus.
The pharyngeal apparatus of Limnognathiamaerski consists of the following structures: epithe-lial cells forming the cuticular structures, gland cellscovering the buccal cavity, cuticular jaw elements(sclerites), the buccal nervous system, sensory orciliated cells, and the pharynx musculature.
The epithelial cells in the pharyngeal bulb secretea true cuticle that covers most of the mouth cavityand the pharyngeal lumen itself; furthermore, thesecells also form the cuticular sclerites (the jaw sys-tem). Some epithelial cells lie inside the sclerites(Fig. 27, ja1) and connect to other epithelial cells ormuscle cells through fenestrae. Other epithelial cellslie in clusters surrounded with thin cuticle, as in thefibularium (Fig. 23, fi). In adult specimens, the epi-thelial cells lack any indication of secretory activity,suggesting that generation of the cuticle only hap-pened once (already in the egg?) and that the cuticleis not molted. The shape of cuticle-generating cellsranges from round in the fibularium to elongatedinside the jaw elements. There are only a few or-ganelles and their cytoplasm is osmiophobic. Thenucleus shows greater areas of condensed chromatinthan in cells with a secretory activity. All featuressuggest that these cells are resting cells.
Unicellular buccal glands seem to be indistinct,but several glandular cells are scattered around theanterior part of the mouth cavity. One large dorsalgland of about 20 cells covers the upper lumen of thepharynx (Fig. 22, bg). These cells have enlargedperinuclear and cytoplasmic cisternae, several mito-chondria, and numerous secretory vesicles—features indicating secretory activity.
The buccal nervous system is not well investi-gated but two large buccal nerves (each with 17nerve fibers) run laterally between the two halves ofthe fibularium and the main jaws. A buccal ganglionmay be present but was not observed.
Limnognathia maerski has overwhelming num-bers of pharyngeal solid parts, the sclerites. Themain parts consist of two ventral basal plates fusedposteriorly, two dorsal lamellae orales, two fibularia,and three sets of jaws, where the main jaws arefused and consist of a dentarium and an articu-larium. Furthermore, most of the lumen of the phar-ynx is covered with a thin cuticle or glycocalyx.
By DIC optics we observed that each half of thethin lamellae orales (Figs. 9, 11, l.or) consists ofvertical rows of 12 longitudinal cuticular rods ortubes. Therefore, it was not surprising to discover byTEM that all other sclerites have the same features,e.g., the fused part of the basal plate (Fig. 27, tu).The tubes consist of lucent, osmiophobic material
23MICROGNATHOZOA: A NEW CLASS
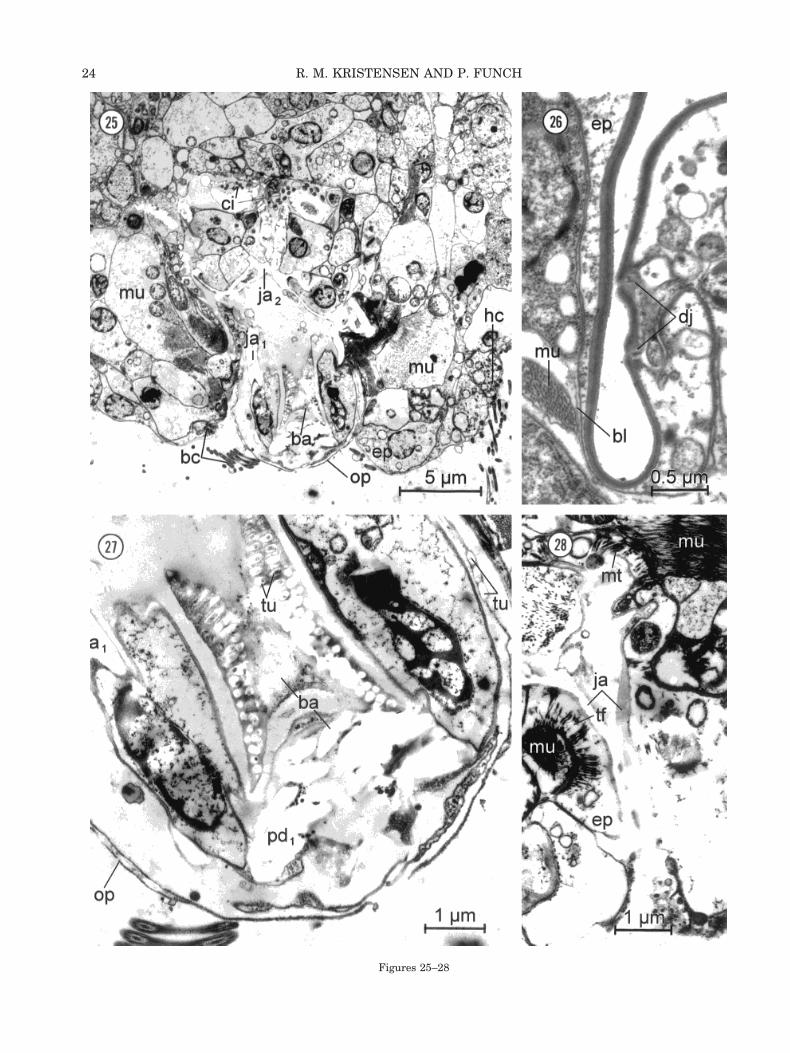
Figures 25–28
24 R. M. KRISTENSEN AND P. FUNCH

surrounding a dense osmiophilic core. The tubularsubstructures are embedded in true cuticle, so incross section the sclerite appears porous and in lon-gitudinal section the sclerite is striated due to themany vertical rows of cuticular tubes. How thesestructures are generated is still uncertain, but inone sclerite we observed microvilli in the tubes. Itseems that the epithelial cell microvilli secrete thetubes. Later, when the microvilli are retracted theyleave a dense core in the middle. In fact, we observedthe osmiophilic microfibers in the middle of thesemicrovilli.
The tubular substructure may be hidden in thesclerotized part of the sclerite, e.g., the pseudodigitsof the ventral jaws (pseudophalangia) and in molarparts of the basal plates. The “arm” of the pseu-dophalangium is hollow in the middle. This lumenstill contains the cells that generated the sclerite.The tubular substructure can be observed as a stri-ation in the lamellae orales and the symphysis byDIC optics. While the diameter of the tubes in thesetwo sclerites is up to 0.4 mm, the diameter of thetubes in all other cuticular elements is about 0.2 mm.
Ciliated epithelial cells (Fig. 23, ci) without cuticleare located dorsal of the two jaw halves of the den-tarium (Fig. 23, ja2). Microvilli are rarely observedin the pharyngeal lumen. Only one type of microvil-lus is present, short with an electron-dense core,being similar to the microvilli of the gut epithelium(Fig. 24, mv).
The muscular part of the pharyngeal system isobserved as three-lobed by light microscopy. Onemedian, caudal lobe attaches to the two arms of thecauda and two lateral lobes attach to the three setsof jaws (Fig. 12, mu). By TEM we observed manysmaller muscle bundles attaching to the accessorysclerites of both the ventral and the dorsal jaws. Thefine structure of all these muscles is beyond thescope of this article, and serial sections of the wholepharyngeal bulb are needed to determine the truelocation of each muscle bundle. A capsular muscle
observed in several gnathostomulids seems not to bepresent in Limnognathia maerski. The whole pha-ryngeal bulb is surrounded only by a weakly devel-oped basement membrane.
Esophagus. The esophagus is dorsal of the pha-ryngeal apparatus. It is very short and lacks cilia ormicrovilli. The esophageal lumen is usually closed,but a sphincter muscle was not observed. The esoph-agus is surrounded laterally by a pair of large sali-vary glands. It seems that the salivary glands do notopen into the pharyngeal apparatus or into theesophagus. They open into the lumen of the midgut.Further investigations are needed before it can bedecided if the salivary glands are homologous withstomach glands in other Gnathifera.
Salivary glands. The so-called salivary glandsare located latero-posteriorly of the pharyngeal ap-paratus (Fig. 1, sg) and they interdigitate with themidgut cells. The cytoplasm of the salivary glands isosmiophilic and numerous granules and vesicles arepresent (Fig. 22, sg). The large nuclei are lobed, witha centrally positioned nucleolus. The salivary glandcells have the ultrastructure of highly active cellsthat export enzymes to the digestive system.
Midgut. The midgut fills up most of the thoracicregion and the anterior part of the abdomen (Figs. 1,3, 18). The midgut consists of cells with large vesi-cles and a Golgi apparatus (Fig. 18, ve, go) thatsurround the lumen of the midgut and peripheralcells with smaller vesicles and giant nuclei. Thesenuclei can reach a diameter of 4 mm and are thesecond-largest nuclei in the entire animal. Only thenucleus of the oocyte may be larger, at up to 10 mmin diameter. In well-fed animals the lumen of themidgut is a labyrinth of crevices (Fig. 18, mg), but inother animals an oval lumen is present in crosssection. All midgut cells have a brush border ofsmall microvilli, only 0.2 mm long (Fig. 24, mv). Eachmicrovillus has an osmiophilic core of two to threefibers, and from the surface of the microvillus nu-merous fine osmiophilic fibers extend out into thelumen of the midgut.
Rectum and anal system. Limnognathia maer-ski lacks a true cuticle-lined hindgut, as seen inmany other invertebrates with a cuticle, except forGnathostomulida and some Gastrotricha that alsolack the permanent anus. Instead, the gut cells de-crease in size posteriorly and end on the edge of thetriangular anal plate (Fig. 1, an). The six most cau-dal cells, the rectal cells (Fig. 33, rc), differ from themidgut cells. They lack the microvilli and onlyslightly interdigitate with the epidermal cell. Thelumen in the gut disappears and only a lacunarsystem exists between the cells. Two small musclesterminate at the edge of the dorsal anal plate, andmay be involved in opening what we interpret as aperiodically functioning anus. There is no basementmembrane between the rectal cells, the two muscles,and the epidermal cells. Because basal laminae arepoorly developed in any case in Limnognathia
Figs. 25–28. Limnognathia maerski nov. gen. et nov. sp. TEM.Pharyngeal apparatus and epidermal cells. Fig. 25: Cross sectionthrough the cuticular oral plate (op) and the anterior of thepharyngeal apparatus with basal plates (ba), the ventral jaw (ja1)and the main jaws (ja2); bc, buccal cilia; ci, cilia in the pharynx;ep; epidermal cell; hc, head ciliophores; mu, muscle. Fig. 26: Theinterdigitated junction (dj) between two latero-dorsal epidermalcells. Note that the matrix layer is intracellular in the epidermalcell (ep). The muscle (mu) is surrounded by a diffuse basal lamina(bl). Fig. 27: Close-up of the pharyngeal apparatus seen in Figure25. The jaw structures (ba, basal plate) and (pd1, pseudodigit) areextracellular, as is the oral plate (op) surrounding the mouthopening. The tubular elements (tu) with the dense core are seenboth in the basal plate and the oral plate. The left ventral jaw(ja1) has an epidermal cell inside the cuticular structure. Fig. 28:Muscle-epidermal junction attached to the jaw elements (ja).Note that the epidermal cells (ep) have both microtubuli (mt) andthread-like (tf) fibers (tonofilaments) in the muscle-attachment.The muscle (mu) is cross-striated.
25MICROGNATHOZOA: A NEW CLASS

Figs. 29–31. Limnognathia maerski nov. gen. et nov. sp. TEM. Brain, pharyngeal apparatus, ventral trunk ciliophores, and theexcretory system. Fig. 29: Latero-longitudinal section of the whole animal. ag, adhesive gland; bc, buccal cilia; br, brain; mo, mouth;oo1, oocyte; ph, pharyngeal apparatus; pr1-pr2, protonephridia; tc, trunk ciliophores. Fig. 30: Close-up of the anterior protonephridiumseen in Figure 29. Note the terminal cell (tl) with flagellum (ft), ciliary root (cr) and microvilli (mv). The canal cell (cc) forms the outerrods (or) of the weir. The nucleus (nu) of the terminal cell is located laterally on the canal. mi, mitochondria. Fig. 31: Canal cell (cc)with nucleus (nu), autodesmosome (au), and two flagella (ft), each surrounded by 9–10 microvilli (mv).
26 R. M. KRISTENSEN AND P. FUNCH

maerski, it is unclear whether these rectal cells areectodermal or mesodermal in origin, but they differfrom the epidermal cells in having many vesiclesand mitochondria. In this way they look more likethe midgut cells. Defecation was never observed.
Body Cavity
There is no coelom in the context of an extracel-lular fluid-filled space (pseudocoel) or of mesodermallining cells (eucoelom). In nearly all cross and lon-gitudinal sections the animal is totally compact(Figs. 29, 33), but in a few cross and longitudinalsections a cavity is observed between the epidermisand the midgut (Figs. 18, 22). This may be an arti-
fact of fixation and the shrinking it causes. Theextracellular matrix between the epidermis and me-sodermal muscles consists of a very thin to diffusebasal lamina (Fig. 26). A distinct basement mem-brane was never observed between mesodermal cellsand the endodermal part of the digestive system orthe ectodermal ventral nerve cord (Fig. 20, vn). Allmesodermal cells are fixed and amebocytes are notpresent. We therefore believe that the micrognatho-zoans are fundamentally acoelomate.
Excretory Structures
The excretory system of Micrognathozoa consistsof two pairs of protonephridia lying ventrolateral to
Fig. 32. Limnognathia maerski nov. gen. et nov. sp. TEM. The adhesive ciliated pad, where two(ce1 and ce2) of the ten adhesive cells are shown. The nucleus (nu) is located proximally in the cell.The cells are multiciliated; each cilium (ci) has a long ciliary root (cr). The cell surface is coveredby a thick glycocalyx (gx). Each cilium is surrounded proximally with a tube-shaped cuticle (cu).
27MICROGNATHOZOA: A NEW CLASS

the gut, one pair in the thoracic region and the otheranterior in the abdominal region (Figs. 2, 29,pr1-pr2). In Limnognathia maerski, the proto-nephridium consists of four terminal cells, two canalcells, and a single nephridiopore cell. The nephrid-iopore cell opens laterally between two lateralplates. All cells in the protonephridium are mono-ciliated, so in a cross section in the distal part of theprotonephridium, seven cilia are seen—four flagella-like cilia from the four terminal cells, two short ciliafrom the two canal cells, and one cilium close to thewall of the nephridiopore cell (Fig. 20).
The terminal cell forms a cylindrical weir appara-tus or ciliary pit (Fig. 30), which consists of nine toten stiff microvilli (inner rods of the weir) and thesingle flagellum in the middle. Between the mi-crovilli the weir is filled with an extracellular stri-ated material (Fig. 31). This material is formed in-side large, coated vesicles in the cytoplasm of theterminal cell and is released by the vesicles into theweir apparatus. The striated material may be ho-mologous with the fenestration lamina in other in-vertebrates. The terminal cell contains a few mito-chondria and an abortive ciliary root (Fig. 30, cr).The cell body and nucleus of the terminal cell arelocated laterally on the weir, as is the nucleus of thecanal cell, a cell that engulfs the weirs from twodifferent terminal cells. The weirs lie in pairs in theproximal part of the canal cell (Fig. 31). The canal
cell also has its own flagellar structure, which entersthe lumen just after the weir. The canal cell sur-rounds the two weirs and closes itself with an au-todesmosome (Fig. 31, au). In a few cases the canalcell also is involved in formation of the weir. Thecanal forms the outer rods (Fig. 30, or) that sur-round two flagella and the two sets of inner rods(Figs. 30, 31, mv). This interpretation could bewrong. The terminal cell may have two flagella, as insome Turbellaria and all known Kinorhyncha, but inthe six animals investigated there were always fourterminal cells and only two canal cells. The nephrid-iopore cell is not well investigated (Fig. 20). It looksvery similar to the canal cell and it also forms acanal by closing on itself with an autodesmosome.The two weirs do not reach to the nephridiopore cell,but four of the six cilia are the flagella from the fourterminal cells; two are shorter cilia from the canalcell and one is the cilium from the nephridioporecell. In the wall of the nephridiopore these sevenciliary structures often may be seen (Fig. 20, pr). Ifour interpretation is correct, all cells in the proto-nephridium are monociliated.
Reproductive Components
After five years of investigation, we found onlymature females of Limnognathia maerski and we
Fig. 33. Limnognathia maerski nov. gen. et nov. sp. TEM. Cross section of the animal in theanal region. an, temporary anus; D, dorsal; mu, muscle; nu, nucleus; oo1, mature oocyte; oo2,secondary oocyte; rc, rectal cells; tc, trunk ciliophores; V, ventral.
28 R. M. KRISTENSEN AND P. FUNCH

assume that the species reproduces by parthenogen-esis, as do many other freshwater meiofaunal ani-mals, such as bdelloid rotifers and echiniscid hetero-tardigrades. There is a small chance the animalcould be a hermaphrodite (see below, “refractivebodies”) or we may have overlooked dwarf males inour culture.
The investigated part of the reproductive systemis anatomically simple. There always exist paired,compact ovaries in mature animals (Figs. 1–3). Theovary seems to consist only of oocytes; nongerminalsheath cells and oviducal cells were not observed.Usually only one egg of the two oocytes matures perclutch; just before oviposition this egg may fill one-third of the entire abdomen. The mature ovum islocated ventro-posteriorly (Fig. 22). In the finalstage of vitellogenesis, when a thin eggshell is de-veloped, it lies in direct contact with the rectal cells(Fig. 33). Oviposition has not been thoroughly exam-ined. Video documentation failed just when the eggswere laid, but we suggest that oviposition may occurthrough a permanent ventral pore just behind theadhesive pad, and not through the temporary dorsalanus as we first thought.
Vitellogenesis may be autosynthetic, i.e., yolk issynthesized within the oocyte from low-weight mol-ecules that diffuse across the thin oolemma directlyfrom the midgut. Evidence for that is seen at earlierstages of vitellogenesis before the eggshell is formed(Fig. 22). Here the oocyte is in direct contact with themidgut. The midgut contains large vesicles in theregion where it contacts the oocyte. The oocyte hasan enlarged nucleus with a nucleolus, many mito-chondria, and prominent endoplasmic reticulum.This clearly indicates that the oocyte itself is veryactive in the formation of the osmiophilic yolk gran-ules.
The mature egg itself also forms the eggshell. Twotypes of eggshell surfaces were observed on newlylaid eggs. One type of eggshell is smooth and sticky;the other is strongly sculptured (Fig. 7). We firstthought that the unsculptured, sticky egg was anabortive egg, considering that we had introducedthem into the depression slides at too high temper-atures. Later, both types of eggs were found in theovary of different specimens. It seems that smallerfemales lay sticky eggs, while larger females laysculptured eggs. Thus, there is a possibility thatthere exist two types of eggs, as in gastrotrichs androtifers. The sticky eggs may be quick-developingsummer eggs (tachyblastic) and the sculptured eggsmay be resting winter eggs (opsiblastic).
Two refractive bodies found in close contact withthe mature egg (Fig. 2, rb1-rb2) may be accessoryorgans. The function of these two structures is notunderstood, and more TEM investigations areneeded. We believe that it is unlikely that these arepart of a male reproductive system, as found re-cently in freshwater chaetonotid gastrotrichs. There
is no evidence for modified spermatozoa inside thesetwo bodies.
Live Observations
More than 100 living specimens of Limnognathiamaerski have been observed, both in Greenland andin the moss culture in Copenhagen. Usually theanimals were collected by an Irwin loop when theywere swimming. If we tried to collect animals sittingon the substrate, they adhered very strongly to thesubstrate with their adhesive pad. Color photos(Figs. 40, 41) were taken immediately after the an-imals were removed from the culture in Copenha-gen. These photos showed that the body shape wasflexible; the thorax especially could contract like anaccordion, but it could also constrict to a narrow,neck-like shape, and then the animal looked nearlylike a figure eight.
Video technique was used to observe swimmingand crawling behaviors. The swimming behaviorwas very characteristic, because the animal movesin a spiral. It is a slow movement, different fromthat of monogonont rotifers, which are common inthe cold spring from Isunngua. It seems that thetrunk ciliophores are used both in swimming and incrawling on the substrate. The preoral ciliary field isnot the main locomotory organ, as we first expected.Specimens were never observed moving backwards,as nearly all gnathostomulids do, when they reversetheir long cilia. Kept cool at 4°C and mounted on adepression microslide with a cover slip, the animalscould be observed alive for about 5 h. When theanimal crawls on the substrate, e.g., pieces of mossleaf or detritus, it moves by slow gliding, using theciliophores in unison. The gliding is caused by thetrunk ciliophores in exactly the same way as theciliophores in Diurodrilus (Kristensen and Niilonen,1982), or as the two rows of ventral ciliation in someof the chaetonotid gastrotrichs (Ruppert andBarnes, 1994). The gastrotrich genus Chaetonotuswas also represented in the same moss samples asLimnognathia, but these animals were easily distin-guished because Chaetonotus has scales and lacksjaws in the pharyngeal apparatus.
The foraging behavior of Limnognathia maerskiwas also observed several times. It consists of arhythmic oscillation of the head from side to side. Itseems like food items, which could be blue-greenbacteria or diatoms, are collected with the preoralciliary field (Fig. 2, pc) and picked up by the pseu-dophalangia. The pseudophalangia may be extrudedout of the mouth opening, and it was observed thatthey could pick up a single diatom, which later wasmacerated by the main jaws. Whole diatoms werenever observed in the midgut in any of the typespecimens. The vomit behavior, which was describedunder “the allotype,” is very similar to the samebehavior in one species of gnathostomulids, Problo-gnathia minima (see Sterrer and Farris, 1975). Most
29MICROGNATHOZOA: A NEW CLASS

of the pharyngeal apparatus is protruded outthrough the mouth, followed by a fast snapping re-action of the main jaws. Defecation was never ob-served in any of the animals, and it was first thoughtthat the anus was located ventrally. Later, when wehad located the dorsal anal tissue by TEM investi-gations, we could also see by DIC optics that themidgut continues dorsally, directly into a closed rec-tum, which ends on the edge of the most posteriortriangular dorsal plate. There is no permanent func-tional anal pore. Such a pore would only open to theexterior, when defecation is happening. A closedanal pore is found in several other meiofaunalgroups. The anus may be secondarily closed, as insome loriciferans and in all Heterotardigrada. AllEutardigrada have a permanent open cloaca anddefecation behavior is very easy to observe. Thechance of observing defecation behavior in livingLimnognathia is small, and we did not succeed inmaking that observation, although we videoed theanimals for hours.
DISCUSSIONComparison Between Micrognathozoaand Gastrotricha
The phylum Gastrotricha consists of only about500 species of microscopical acoelomate worms. Thephylum is divided into two orders, the marine Mac-rodasyida and the marine and freshwaterChaetonotida. The phylum is one of the best TEM-investigated groups in the world (see the excellentreview of Ruppert, 1991b). The so-called primitivegroups of gastrotrichs may have monociliated epi-dermal cells, a plesiomorphic character shared withall gnathostomulids (Rieger, 1976). Superficially,Micrognathozoa look like chaetonotid gastrotrichs.Like all gastrotrichs, the chaetonotids have onlyventral locomotory cilia, with which they glidesmoothly over the substrate. In some chaetonotidsthe locomotory cilia are arranged in two longitudinalbands, as in Limnognathia maerski. Furthermore,the ciliary bands of chaetonotids are grouped in cirrilike the ciliophores of Limnognathia. The cilia (in-cluding the locomotory cilia) of gastrotrichs are al-ways surrounded by the exocuticle (Ruppert, 1991b)and this has been considered a good autapomorphyfor the phylum. The cilia of the adhesive pad ofLimnognathia clearly have a cuticular membranemost proximally, but this membrane does not con-tinue all the way out on the cilia (Fig. 32). Thesimilarities between the Micrognathozoa and Gas-trotricha seem to be only superficial, and are notsupported by ultrastructural evidence. The differ-ences in the pharyngeal apparatus and the integu-mental structures are so large that the other simi-larities must be considered convergence. Allgastrotrichs have a myoepithelial pharyngeal bulbwith a triangular lumen that is lined by a layer ofcuticle. This type of pharyngeal system with a pha-
ryngeal channel running through the center of thebulb is very different from the jaw system and theventrally placed pharyngeal bulb seen in both Gna-thostomulida and Micrognathozoa. Furthermore, inthe two last-mentioned groups the bulb consists ofan epithelial and a muscle layer. The epidermis ofgastrotrichs is covered with a true cuticle, which insome gastrotrichs may be specialized to form scalesor spines. There is no evidence for an intracellularplate structure, as in Micrognathozoa and Rotifera.We therefore consider gastrotrichs to be members ofthe Cycloneuralia clade (Nielsen, 1995) and Gastro-tricha is not the sister-group to Gnathostomulida(Wallace et al., 1996; Cavalier-Smith, 1998; Zrzavyet al., 1998), but to the Introverta clade, which in-cludes Nematoda, Nematomorpha, Priapulida, Ki-norhyncha, and Loricifera (Nielsen et al., 1996; Sø-rensen et al., 2000).
Comparison Between Micrognathozoaand Gnathostomulida
The phylum Gnathostomulida consists of micro-scopical, acoelomate worms that live exclusively inmarine sediments, especially those with a high con-tent of hydrogen sulfide and a lack of oxygen. Theapproximately 80 described species may be found incryptic habitats such as stromatolites, cold seeps, ordeep in the sediments, avoiding oxygen (for review,see Sterrer, 1998). The monociliated epidermis andprotonephridia, the blind tubular gut system andacoelomate body condition (Lammert, 1991) maysupport the theory of an ancient origin of Gnatho-stomulida and they may have survived from thePrecambrian, when oxygen was nearly lacking(Fenchel and Riedl, 1970; Boaden 1975, 1989). How-ever, the complex cuticular jaws and extremely com-plex hermaphroditic gonads do not support the the-ory that gnathostomulids should be the evolutionarybridge between a cnidarian ancestor and the Platy-helminthes (Ruppert and Barnes, 1994). The phylo-genetic position of Gnathostomulida has been veryturbulent. Haszprunar (1996a) regarded them to bethe “most primitive” of all spiralian Metazoa, whileNielsen (1995) considered them to be specializedpolychaetes. Before presenting a comparison be-tween the new group, Micrognathozoa, and Gnatho-stomulida, we therefore outline a short historicalreview of the “enigmatic” Gnathostomulida (seeSterrer et al., 1985).
Gnathostomulida was described by Ax (1956) asan order of Turbellaria, but the group was knownsince the 1920s by Remane (unpublished). Riedl(1969) established a new phylum for the gnathosto-mulids and Sterrer (1972) supported this, but healso mentioned that gnathostomulids share charac-teristics with both Platyhelminthes and Aschel-minthes. Ax (1985, 1989, 1996) could not give up hisfirst impression and placed the Platyhelminthes(flatworms) and Gnathostomulida as sister taxa in
30 R. M. KRISTENSEN AND P. FUNCH

the clade Plathelminthomorpha. This theory wassupported by four synapomorphies: 1) hermaphro-ditism, 2) direct transfer of sperm and internal fer-tilization, 3) thread-like sperm, and 4) lack of mito-sis in somatic cells. The three first-mentionedautapomorphies are very weak characters becausethey are observed in many other lines of Metazoa,and the fourth character may be a plesiomorphiccharacter. The monociliated epidermal cells (Rieger,1976; Rieger and Mainitz, 1977) may also be a ple-siomorphic character present in all gnathostomulidsand some gastrotrichs, e.g., Chordodasys and Xeno-dasys (Rieger, 1976; Ruppert, 1991b).
Sterrer et al. (1985) questioned the link betweenthe Platyhelminthes and Gnathostomulida on thebasis of the ventral pharyngeal bulb, the true cross-striated muscles, and the presence of a periodicallyfunctioning anus (Knauss, 1979). Kristensen andNørrevang (1977, 1978) indicated similarities be-tween polychaetes (Dorveillidae) and gnathostomu-lids in jaw formation and structure. They also statedthat the similarities between the mastax of Rotifera(Koehler and Hayes, 1969) and Gnathostomulidaseem rather more inconclusive (see below, “Compar-ison Between Micrognathozoa and Annelida”). Themolecular data (18S ribosomal DNA) are not reallyhelping to clarify the phylogenetic position ofGnathostomulida. Littlewood et al. (1998) indicate arelationship with the Gnathostomulida and a Nem-atoda 1 Chaetognatha clade. Both Zrzavy et al.(1998) and Giribet et al. (2000) offer phylogenetichypotheses based on combined morphological andmolecular data. Although the datasets in the twoanalyses are quite similar, their analyses differ andZrzavy et al. (1998) suggest a close relationship be-tween the Gnathostomulida and Gastrotricha(Neotrichozoa), while Giribet et al. (2000) suggestthat Gnathostomulida is a sister-group to Cyclio-phora 1 Syndermata. Recently, Rieger and Tyler(1995) and Ahlrichs (1995, 1997) published ultra-structural evidence indicating a sister-group rela-tionship of Gnathostomulida with Rotifera-Acanthocephala. Ahlrichs (1995) established a newmonophylum Gnathifera for the three mentionedgroups. Sørensen (2000) used a new SEM techniqueto clean both the rotifer trophi and the jaws of gna-thostomulids. His observations support a close rela-tionship between Gnathostomulida and Rotifera.The TEM investigation of pharyngeal epithelium,pharyngeal musculature, and the pharyngeal jawshas already shown the same evidence (Herlyn andEhlers, 1997). In the same study, Herlyn and Ehlers(1997) suggest that the absence of pharyngeal cilia-tion is an autapomorphy for the Gnathifera. Wedisagree with that interpretation. Pharyngeal ciliain Rotifera are clearly present (see fig. 26, Clementand Wurdak, 1991), and cilia are present in thepharyngeal epithelium of Micrognathozoa as well(Fig. 23). Gnathostomulida, however, seems to lackthe ciliation in the pharyngeal epithelium; instead,
they have thick microvilli (fig. 13, Kristensen andNørrevang, 1977). It is therefore interesting thatLimnognathia maerski has both cilia and microvilli(Figs. 23, 29) in the pharyngeal epithelium.
In this research, we used the same SEM techniqueas described by Sørensen (2000) for Gnathostomu-lida and also used in Rotifera (De Smet, 1996, 1997)to clean the cuticular elements of Limnognathia-pharyngeal apparatus. It should therefore be easy tocompare the solid parts of the pharyngeal system inthe three groups. Unfortunately the comparison isnot easy. The largest dissimilarity is between the“compact” type of gnathostomulid jaws found in theorder Filospermoidea. Several similarities arepresent between the jaws of Micrognathozoa and the“basket type” of gnathostomulid jaws found in theorder Bursovaginoidea. Especially, basket jaws ofthe suborder Scleroperalia are very similar to themain jaws of Limnognathia. The basket type is alsomore similar to the trophi (jaws) of Eurotatoria thanthe compact jaws. These findings are very confusingbecause the compact type of jaws has always beenconsidered as the plesiomorphic condition within theGnathostomulida (Sterrer, 1972). If the compacttype is the plesiomorphic condition, several ele-ments of rotiferan trophi and micrognathozoan jawsmay have been formed by convergence, so that theylook similar to the basket jaws, or the ground pat-tern for both rotiferan trophi and micrognathozoanjaws was also present in the stem group of Gnatho-stomulida. Then the compact jaws in filospermoidgnathostomulids are a secondary reduction. Withinthe Rotifera we have the same problem. The trophiof the marine genus Seison (Order Seisonidea) arevery aberrant and are not easy to compare with thetrophi of the Eurotatoria (Ahlrichs, 1995; Segers andMelone, 1998).
The similarities between the isolated jaws of thegenus Gnathostomula and the main jaws of Limnogna-thia are really astonishing (Riedl and Rieger, 1972).There are similarities, which are so convincing thatthey may be homologies. Similarities are seen, notonly in the dentarium, where three arcs of teeth arepresent in both genera, but also in the articularium.The articularium consists of two symphysis lamellaethat are joined by a symphysis. The cuticularizedpart of the main jaws in Limnognathia ends in apaired cauda system, as in Gnathostomula. In Lim-nognathia the caudae are symmetrical, as in, e.g.,Gnathostomula microstyla (see Riedl and Rieger,1972). The fibularium of Limnognathia may be bet-ter understood if we follow the theory (Riedl andRieger, 1972) of fibularization of the involucrum inthe family Gnathostomulidae. The fibularium maybe homologous to the apophysis and the involucrumsystem of Gnathostomulida. If so, then the fibu-larium is developed to an extreme degree in Limnog-nathia. As in gnathostomulids, each fenestra con-tains a single cell, but instead of three fenestrae (ineach jaw) in Gnathostomulida there exist at least
31MICROGNATHOZOA: A NEW CLASS

eight fenestrae in Limnognathia on both sides ofeach main jaw element (Figs. 8, 9). It is very difficultto find counterparts in the gnathostomulid jaws tothe other sclerites in Limnognathia jaws. The basalplates of Limnognathia are paired but partly fused.The basal plate of gnathostomulids consists of asingle plate, but may be secondarily fused of severalelements. The two lamellae orales in Limnognathiahave the same substructure as the lamellae adden-talis in Valvognathia pogonostoma. They may in factconstitute a continuous, folded cuticle, covering alarge part of the pharyngeal lumen. The ventraljaws and dorsal jaw elements are not observed inany Gnathostomulida. The complexity of the jaw ofLimnognathia is greater than that of jaws of anyinvertebrate group and far beyond the complexity ofGnathostomulida. Unfortunately, they are alsoamong the smallest jaws, so subunits in the scleritescan only be observed by high-resolution SEM.
The new group Micrognathozoa supports the the-ory that the jaws of gnathostomulids are homolo-gous with rotiferan trophi. The jaws of Limno-gnathia have several elements that are similar tothose of gnathostomulids; other elements may becloser to some of the sclerites of rotiferan trophi.
A closer relationship (sister-group relationship)between Micrognathozoa and Gnathostomulida wassupposed when we discovered Limnognathia in 1994(Kristensen and Funch, 1995). This was based onthe SEM investigations of the main jaws of Limnogna-thia and the jaws of scleropleralian gnathostomulidsand also on behavioral observations of Problo-gnathia minima and Limnognathia maerski.
Problognathia minima was described from CastleHarbour of Bermuda (Sterrer and Farris, 1975). It isa very small, plump gnathostomulid that lacks theability to swim backwards. Furthermore, the man-ner in which Problognathia protrudes the pharyn-geal apparatus (Sterrer and Farris, 1975) closelyresembles the “vomit” behavior we observed in Lim-nognathia. The two refractive bodies of the allotype(Fig. 2, rb) were thought to be bursae or prebursae,like the round bursa of Problognathia. Unfortu-nately, we still do not have any TEM photographs ofrefractive bodies, but after investigations of morethan 50 living specimens of Limnognathia it seemsthat the bodies are glandular structures and there isno evidence that L. maerski should be a herma-phrodite. The new SEM investigations of the trophiof planktonic Rotifera (Sørensen and Kristensen,2000) and the TEM investigations of L. maerski mayindicate that the Rotifera is the sister-group to Mi-crognathozoa, and not the Gnathostomulida.
Comparison Between Micrognathozoaand Rotifera
The rotifers were discovered just after the inven-tion of the microscope, but Leeuwenhoek could notdistinguish them from Protozoa and they were in-
cluded in the group Infusoria (for historical review,see Hyman, 1951). Today more than 1,500 specieshave been described, the majority from freshwaterenvironments, e.g., zooplankton, algae, and mosses(Nogrady et al., 1993). Several species of the classBdelloidea may enter into cryptobiosis and can thenbe found in extremely dry habitats such as theNamib desert, where they form anhydrobiotic tunsunder quartz stone (unpublished observations), oron the surface of glaciers, where they may havesurvived for many years in a cryobiotic stage in thecryoconite holes (De Smet and Van Rompu, 1994).Still, the active stage always needs water and allrotifers are aquatic animals. The marine species ofrotifers have generally been neglected. In manytextbooks rotifers are only treated as freshwateranimals, although planktonic rotifers (e.g., the ge-nus Synchaeta) are very important in nutrient recy-cling in all marine systems. Recently, marine roti-fers of the class Seisonida have been placed centrallyin phylogenetic discussions of the origin of theRotifera (Ahlrichs, 1993a, 1997; Ricci et al., 1993;Segers and Melone, 1998). The monophyly of theRotifera has been questioned and the class Sei-sonida (only one genus Seison) has been placed as asister-group to the exclusively parasitic phylum Ac-anthocephala (Ahlrichs, 1995, 1997; Herlyn andEhlers, 1997). The new theory to include the Acan-thocephala as an ingroup inside the Rotifera isbased primarily on ultrastructural observations ofthe integumentary structures. Storch and Welsch(1969) first discovered several ultrastructural simi-larities between the rotiferan and acanthocephalaninteguments. Later, the monophylum Syndermatawas proposed for Rotifera and Acanthocephala (seeAhlrichs, 1995), based on several other ultrastruc-tural characters. Molecular data strongly supportsuch a monophyletic group (Winnepenninckx et al.,1995, 1998) but the interrelationship inside theclade remains unclear (Garey et al., 1998; Welsch,2000). Unfortunately, we cannot compare importantmorphological characters present in Micrognatho-zoa with potential homolog characters in Acantho-cephala because Acanthocephala totally lacks an al-imentary tract, including the pharyngeal apparatus(Dunagan and Miller, 1991). We can only state thatMicrognathozoa lacks a syncytial epidermis, one ofthe proposed autapomorphies for the Syndermataclade.
One of the autapomorphies for all Rotifera is thecomplicated jaw apparatus (the mastax) that alwayscontains cuticular parts elements, the trophi. Theplesiomorphic condition for the trophi is the condi-tion in which the trophi (singular: trophus) comprisefour different kinds of cuticular elements called scle-rites (Markevich, 1989). Before we compare the tro-phi of Rotifera and the jaw apparatus of Micro-gnathozoa, we should briefly define the four primarycuticular elements of the rotiferan trophi.
32 R. M. KRISTENSEN AND P. FUNCH

The central parts of the trophi consist of twopaired sclerites, the rami. The rami are held to-gether posteriorly by a flexible cuticular ligament, sothe two sclerites can be moved independently bymuscles. Each ramus is formed by three epithelialcells (Markevich, 1989). The epithelial cells formchambers inside the sclerites, and each of theseopens to the surface of the sclerite by a small win-dow, or fenestra. At least two windows, basifenestraand sub-basifenestra, are used today in systematicstudies of monogonont rotifers (Markevich, 1989; DeSmet, 1997). According to Markevich (1989), a thirdwindow, the fenestrulum, might be present apicallyon the ramus. Each ramus may have a strong den-tition going all the way down to the symphysis, butoften the distal part of the ramus is smooth.
The unpaired fulcrum is positioned caudally to thesymphysis of the rami. By light microscopy and SEM(Sørensen, 2000) the fulcrum appears striated,which is caused by longitudinally arranged tubes.This striation may be present in other sclerites, butoften the microtubes are embedded in a homoge-neous matrix layer and the subunits cannot be ob-served by SEM. The fulcrum serves as the cuticularmuscle attachment for the large abductor muscles,which are responsible for moving the rami like for-ceps.
The paired unci are positioned apically or ven-troapically to the paired rami. Each uncus may be asingle large tooth or a large plate with several teeth.The uncus seems to lack windows or fenestra. Theunci and the rami are responsible for handling thefood and may crush food particles in different ways.
The last pair of primary sclerites is the manubria.They are located lateral to the rami. Each manu-brium is attached to the uncus by a thin ligament.An extensor muscle attaches to the posterior part ofthe uncus and the caudal part of the manubrium isresponsible for the movement of these two sclerites.The manubrium is formed by three epithelial cells,each lying in a chamber or on the surface of themanubrium. If all three chambers with fenestra arepresent the openings are referred to as the anterior,the median, and the posterior fenestra.
Secondary sclerites may be present in some mono-gonont trophi as well in the Seison trophi. The sec-ondary sclerites form the epipharynx (De Smet,1997; Segers and Melone, 1998) or pseudoepiphar-ynx (Seison; Ahlrichs, 1995). At least in the genusDicranophorus, the paired epipharynx consists ofstriated plates. The location of the epipharynx mayvary from one species to another (De Smet, 1997),but often it is found anterior to the unci. Two pairsof accessory sclerites may be present between theuncus and the manubrium. These sclerites are thesupramanubrium and the intramalleus.
In principle, all cuticular elements of Rotifera areformed by tubular rods with a dense osmiophilic core(fig. 27, Clement and Wurdak, 1991; fig. 1, Riegerand Tyler, 1995), as in Gnathostomulida and Mi-
crognathozoa, but in different species the scleritesmay be reduced or highly modified, so the tubularsubunit can only be observed in the unpaired ful-crum (Segers and Melone, 1998; Sørensen and Kris-tensen, 2000). When the fulcrum is reduced, as inthe class Bdelloidea, the tubular subunits are diffi-cult to recognize, but we still believe that the tubu-lar subunits were present in all sclerites in the stemspecies of Rotifera, and they are an autapomorphyfor Gnathostomulida, Rotifera, and Micrognathozoa.There is no evidence that the cuticular sclerites maymolt or that the juveniles of rotifers have fewersclerites than the adults. The embryo inside therotiferan egg already has all elements. In the sameway, the cuticular elements of the gnathostomulidjaw may degenerate (Sterrer, 1968), but molting ofthe cuticular elements is not observed in gnathosto-mulids or in Limnognathia.
It is not easy to homologize the jaw elements ofLimnognathia with the sclerites of rotiferan trophi.The complexity of the Limnognathia jaw is muchgreater than that of any known rotiferan trophi. Atleast seven paired elements and one unpaired cutic-ular element are present in Limnognathia jaws. Thegreatest similarity is found in the trophi of the fam-ily Lindiidae, which have the cardate type of trophi(Nogrady et al., 1993). Cardate trophi are similar tothe virgate trophi that are common in marine mono-gonont rotifers (Synchaeta; Sørensen and Kris-tensen, 2000). The virgate type may be the plesi-omorphic type of trophi with Eurotatoria. Like thetrophi of Synchaeta, where the manubria, unci, andfulcrum curve (Koste, 1978), the jaw apparatus ofMicrognathozoa has a globular form.
Using the genus Lindia trophi as a model may bedangerous if the cardate trophi are an advancedtype within the Rotifera. We had the same problemwith Gnathostomula jaws within the Gnathostomu-lida. The comparison between the trophi of Lindiaand Limnognathia is as follows: 1) The unci and theventral jaws (pseudophalangia) are homologous; 2)The manubria and the accessory sclerites attachedto the ventral jaws are homologous; 3) The rami andthe dentarium of the main jaws are homologous; 4)The fulcrum and the articularium of the main jawsare homologous; 5) The epipharynx and the lamellaeorales are homologous; 6) The fibularia are elementsof rami, which have developed to an extreme degreein Limnognathia; 7) The dorsal jaws with accessorysclerites are an autapomorphy for Micrognathozoa;and 8) The basal plates are absent in Rotifera(present in Gnathostomulida).
There are several problems with these eight state-ments. The major problem is the main jaws. Theylook much more like the jaws of gnathostomulids.The fulcrum of the rotiferan trophi is never split intotwo symmetrical arms, like the cauda of Gnathosto-mula and Limnognathia. Furthermore, the dentesterminales (Fig. 8, d.te) may be homologous with theuncinal teeth, and then the fibularium may be the
33MICROGNATHOZOA: A NEW CLASS

manubrium (Ahlrichs, pers.com.). More extensiveSEM investigations of the jaws using higher magni-fications may clarify these questions. The synapo-mophies for Rotifera, Gnathostomulida, and Microgna-thozoa are very limited, but a possible uniquecharacter is that the epithelial cells lie inside thesclerites and form openings to the outside via thewindows or fenestra. Recently Sørensen (2000) hasshown that the fenestra dorsalis in Rastrognathiamacrostoma also contains a cell. All fenestrae ofLimnognathia maerski have the same feature (Figs.10, 11).
Except for the possible homology of the cuticularjaws of Limnognathia and the trophi of rotifers,there are very few similarities in the ultrastructureof the two groups. In fact, the micrognathozoans lackstructures that could be interpreted as homologousto the corona, the germovitellarium, the retrocere-bral complex, and the syncytial origin of many tis-sues, as is seen in rotifers. The intracellular skeletallamina found in Syndermata and in Micrognathozoais probably homologous, in spite of the lack of thisstructure on the ciliated ventrum of Limnognathia.
The integument of Limnognathia exhibits a dis-tinctive intracellular dense matrix layer (Fig. 21,im) that resembles the intracytoplasmic skeletallamina of rotifers (Clement and Wurdak, 1991). Theskeletal lamina in rotifers has been considered ho-mologous to a similar structure in the integument ofAcanthocephala, and this shared feature has beenused to suggest a sister-group relationship betweenRotifera and Acanthocephala (Storch and Welsch,1969). But in Rotifera and Acanthocephala the in-tracellular lamina are in a syncytial epidermis,while in Micrognathozoa the dense matrix is withina cellular epidermis. The cellular epidermis of theMicrognathozoa is likely to represent the plesiomor-phic condition as it is found in most invertebrates.
Otherwise, the apical position of an electron-dense, intracytoplasmic lamina in the epidermis,just beneath the cell membrane, is shared in allthree groups. The lamina could be considered a spe-cialized terminal web, with a skeletal function forsomatic muscle attachment in the absence of a trueextracellular cuticle. Such a molting cuticle is miss-ing in all three groups. Other integumental struc-tures in Micrognathozoa are not similar to those ofRotifera. Micrognathozoa has multiciliated epider-mal cells, as in Rotifera, and, typically, rotifers pos-sess a corona consisting of a circumapical band anda buccal field. Micrognathozoa has no structure thatcould be considered homologous to the rotiferan co-rona. The locomotory cilia in Micrognathozoa arewithout exception ventral, and they extend to theposterior trunk. A ciliated band that encircles theanterior is not present. Furthermore, the cilia of thecorona in Rotifera are restricted to the anterior end,although some rotifers, such as species belonging tothe family Dicranophoridae, have a ventral anteriorciliated field of some extent.
An organ similar to the characteristic germovitel-larium of rotifers is not present in Micrognathozoa.Both Rotifera and Micrognathozoa have proto-nephridia, but their ultrastructure differs. The ro-tiferan terminal cell is multiciliated (Ahlrichs1993a,b), while the micrognathozoan terminal organis monociliated. The stomach of Eurotatoria is typi-cally ciliated; in some species it is syncytial. Themicrognathozoan gut is neither; it consists of anepithelium with a brush border of microvilli extend-ing into the gut lumen. The stomach of Seisonidaalso exhibits a brush border of microvilli (Ricci et al.,1993; Ahlrichs, 1995). Most rotifers have a dorsalcloaca, although members of the genus Asplanchnahave no anus. The condition with a temporary anusin Micrognathozoa is not known in rotifers.
Comparison Between Gnathifera Ahlrichs,1995 and Annelida
The gross anatomy of the body of Limnognathiamaerski appears similar to that of some interstitialpolychaetes (cf. Westheide, 1990). As in many ma-rine interstitial animals, e.g., turbellarians, gnatho-stomulids, gastrotrichs, and interstitial polychaetes,tactile bristles or sensoria are present on the body ofMicrognathozoa. It is well known that these sensorystructures have developed by convergence in thedifferent phyla as an adaptation to living in theinterstitium between sand grains (Swedmark, 1964;Ax, 1966). The problem with L. maerski is that it isneither marine nor interstitial. It has been over-looked that many freshwater and epiphytic meiofau-nal animals also have tactile bristles, e.g.,chaetonotid gastrotrichs and aelosomatid annelids.The tactile bristles always consist of compound cilia,but they may have a different origin. The tactilebristles of gnathostomulids are all formed from sev-eral monociliary cells. In annelids, however, thebristle may be formed by either a single multiciliarycell or by several monociliary cells. The long stiffbristles (apicalia, frontalia, dorsalia, lateralia, andcaudalia) of L. maerski consist of one to three longcilia. The lateralia, dorsalia, and caudalia have aring-shaped to oval opening, the aperture, throughthe epidermal cell. Such an aperture often is foundin the dorsal antenna of loricated rotifers (Notholcaikaitophila; Sørensen and Kristensen, 2000) but un-til now has not been observed in annelids. However,the repetition of the tactile bristles on the body is notfound in rotifers, but is present in several intersti-tial dorvilleid polychaetes (Neotenotrocha sterreri;Eibye-Jacobsen and Kristensen, 1994) and in theexclusively interstitial annelid order Diurodrilida(see Diurodrilus westheideri; Kristensen and Ni-ilonen, 1982), and in L. maerski. Furthermore, thelateral tactile bristles (lateralia) are similar in pat-tern in both annelids and L. maerski. Three trunkdorsalia are present serially in L. maerski, and theyare always paired. In contrast, the dorsalia in Diu-
34 R. M. KRISTENSEN AND P. FUNCH

rodrilus are located only on the protostomium (head)and the dorsalia may be paired (two pairs) or un-paired (two single middorsal structures). We believethat tactile bristles have developed by convergencein several meiofaunal groups, including the Anne-lida and Micrognathozoa, but the similarities aresurprising.
More surprising is the presence of ciliophores inboth Diurodrilus and Limnognathia. Ciliophores arelarge, ventral epidermal cells with numerous cilia towhich basal bodies are attached in regular rows. Thebeat of the cilia in one row is synchronized and theciliary roots are very long in both Diurodrilus (fig.14, Kristensen and Niilonen, 1982) and Limnognathia(Fig. 20). The cilia are not bounded by a commonmembrane, as in gastrotrichs (Ruppert, 1991b).
The prostomial ciliophores of Diurodrilus have avery specific pattern and have been used in speciesdetermination. These ciliophores may be used as abroom to collect small detritus particles. In Diuro-drilus, three rows of prostomial ciliophores exist: ananterior, a central, and two lateral groups. The an-terior rows of prostomial ciliophores have rectangu-lar bases, while the large central and the smalllateral groups vary considerably. In addition, threeto six pairs of metastomial ciliophores usually flankthe pharyngeal apparatus. Kristensen and Niilonen(1982) stated that these prostomial and metastomial(5 peristomial) ciliophores may be an autapomorphyfor the genus Diurodrilus. However, head cilio-phores have also been found recently in dorvilleidpolychaetes (Eibye-Jacobsen and Kristensen, 1994)and now in Limnognathia. Both protostomial andperistomial ciliophores are present in the small dor-villeid, Neotenotrocha sterreri. Here the bases of theciliophores are rectangular. The rows of both theprotostomial and peristomial ciliophores are notonly located on the ventral side, as in Diurodrilusand Limnognathia, but the two rows of ciliophoresin N. sterreri continue onto the dorsal side as well. Itis too early to state whether the head ciliophores arehomologous in the two annelid species.
The preoral ciliary field (Fig. 2, pc) in Limnognathiasuperficially looks like ciliophores, but the cilia arenot placed in regular rows or groups. The most sur-prising findings in Limnognathia are the lateral cil-iophores that flank the pharyngeal apparatus (Fig.2, hc), exactly in the same way as in Diurodrilus (fig.2 in Kristensen and Niilonen, 1982). The head cilio-phores of both Limnognathia and Neotenotrochahave a rectangular base, while only the anteriorgroup of ciliophores of Diurodrilus has this verycharacteristic configuration. This presumed homol-ogy between the head ciliophores of Micrognathozoaand Annelida probably represents a plesiomorphiccondition, because it is very difficult to believe thatthe head ciliophores of Micrognathozoa and Anne-lida could be convergent structures. Consequently,head ciliophores have been secondarily reduced sev-
eral times independently in various taxa, and that isnot a very parsimonious hypothesis.
Trunk ciliophores are only found in Diurodrilusand Limnognathia. The 18 trunk ciliophores of Lim-nognathia consist of double cells (Figs. 2, 4). Allciliophores are very similar, with four rows of cilia ineach cell. The first rows of trunk ciliophores arelocated just behind the oral plate (Fig. 6). In livingand slightly contracted animals (Fig. 2), the first rowmay overlap the oral plate, but the trunk ciliophoresare always located on the thorax and the abdomen,and not on the head. Just behind the last row oftrunk ciliophores is located the adhesive ciliated pad(Figs. 2, 4, 32). The cilia of the pad may be modifiedtrunk ciliophores, which have changed from a loco-motory function to being adhesive. The adhesiveglands seem to open in a midventral position be-tween the two clusters of five large ciliated cells. InDiurodrilus the number of trunk ciliophores is con-stant within the species, but shows marked differ-ences between the species. The plesiomorphic condi-tion seems to be 15 trunk ciliophores, which meansthree ciliophores on each trunk segment. Each cilio-phore has ten rows of cilia ordered in a very strictway. In Diurodrilus westheidei there exist an ante-rior postpharyngeal (ppc) and a posterior preanalciliary field (see fig. 2, Kristensen and Niilonen,1982). The preanal ciliary field closely resembles theadhesive ciliated pad in Limnognathia, but in Diu-rodrilus the ciliary pad is not adhesive. The adhe-sive glands (duo glands) are located in the so-calledtoes. By DIC optics it looks as though the trunkciliophores of Diurodrilus consist of only one ele-ment (not double, as in Limnognathia). However, byTEM technique it is seen that there may be up tofour ciliophores forming the specialized ciliated ar-eas, which form the locomotory organs (“Mem-branellen,” Ax, 1967). In an as-yet undescribed Diu-rodrilus species from Australia with dorsal plates,the first two rows of trunk ciliophores are still notfused. These ciliophores consist of four ciliary fields.The three to nine rows of ciliophores comprise twocells, and the last (posterior) row nine is reduced toa single ciliophore. The rows of trunk ciliophores inthe genus Diurodrilus may vary considerably, andciliophore pattern is used to determine the differentspecies (Mock, 1981; Villora-Moreno, 1996). Thenumber of rows of trunk ciliophores may be relatedto the dorsal reinforcements or to the so-called dor-sal cuticular plates. These structures were describedfor the first time by Ax (1967) in D. ankeli. They areabsent in D. westheidei and in D. minimus, and arepresent as very delicate structures (Mock, 1981) inD. subterraneus. Species with strong dorsal rein-forcements seem to have fewer rows of ciliophores.The rows of ciliophores may be reduced to nine andit is always the posterior rows that are reduced. Inthe new species of Diurodrilus from Queensland(Australia), the dorsal reinforcements as well as thefine structure of the trunk ciliophores can only be
35MICROGNATHOZOA: A NEW CLASS

observed when the animal is alive. The dorsal platesare not only present on the prostomium and meta-stomium (5 peristomium) as described in D. ankeli,but are also present on all five trunk segments. Inwholemount preparations, the so-called cuticularplates disappear, as in Limnognathia. New TEMinvestigations are greatly needed to determinewhether the dorsal reinforcements in Diurodrilusmay also be intracellular matrix plates, as in Lim-nognathia.
In the classic work of Ax (1956), in which he de-scribed the gnathostomulids, he stated that “Ar-chiannelida” and Gnathostomulida could not be re-lated because the “Archiannelida” lack jaws (p. 553,Ax, 1956). Many new forms with jaws in the so-called “Archiannelida” have been found since 1956(for review, see Westheide, 1982; Westheide and vonNordheim, 1985). Today the group “Archiannelida”has been abandoned by nearly all scientists and thedifferent families are included in the classPolychaeta. The “Archiannelida” with jaws are allincluded in the polychaete family Dorvilleidae(Eibye-Jacobsen and Kristensen, 1994).
Ax (1956) cited Remane (unpubl.) as the first per-son to have stated that gnathostomulids were re-lated to annelids (“Archiannelida”). Later, Kris-tensen and Nørrevang (1977, 1978) came to thesame conclusion when they described the pharyn-geal apparatus of the gnathostomulids Rastro-gnathia macrostoma and Valvognathia pogonos-toma. The muscular part of the pharyngealapparatus is similar to that found in various anne-lids, being ventral, with the pharyngeal tract pass-ing dorsally to the muscular bulb. In Turbellaria, towhich gnathostomulids were first assigned (Ax,1956), the pharyngeal muscles surround the phar-ynx that passes through the middle of the muscularpharyngeal bulb. As a consequence, Kristensen andNørrevang (1977, 1978) followed the idea of Riedl(1969) and regarded Gnathostomulida as a separatephylum, clearly unrelated to Platyhelminthes,thinking that gnathostomulids were closely relatedto annelids. They based their hypothesis on the for-mation and structure of the jaw apparatus of thedorvilleid polychaete, Ophryotrocha sp. These TEMinvestigations were never published because the fix-ation was of poor quality.
Later, Kristensen and Niilonen (1982) found sim-ilarities between the Diurodrilidae and Gnathosto-mulida. In some gnathostomulids the pharyngealarea is provided with glands, which, with respect tonumber and location, may be compared to the glandsof the Diurodrilidae. Some aberrant spermatozoa ofgnathostomulids (suborder Scleroperalia) have mi-crovillar feet that superficially look like themushroom-shaped bodies from the mature sperma-tozoa of Diurodrilus subterraneus. The similarity,however, is due to convergence. When Kristensenand Eibye-Jacobsen (1995) made a thorough TEMinvestigation of spermiogenesis of D. subterraneus,
they also noted important differences at the func-tional level. The spermatozoa of D. subterraneus areflagellar and the mushroom-shaped bodies are notinvolved in their movement. Those spermatozoa ofGnathostomula that have been investigated (Graeb-ner, 1969) are aflagellar, and movement is per-formed with the aid of microvilli, which look like themushroom-shaped bodies. The theory regarding theGnathostomulida–Annelida relationship proposedby Kristensen and Nørrevang (1977) was totally ne-glected by other scientists, except for Nielsen (1995),who denied gnathostomulids phylum status and in-corporated them in Annelida as “a highly specializedpolychaete group.”
A few years ago the finding of Remane’s hypothet-ical “archiannelids with jaws” (Neotenotrocha ster-reri) would have supported Nielsen’s theory aboutthe gnathostomulids as polychaetes. Today we knowthat the jaws of dorvilleids (Purschke, 1987) arebuilt quite differently from those of the gnathosto-mulids (Herlyn and Ehlers, 1997). When Eibye-Jacobsen and Kristensen (1994) described N. ster-reri, they abandoned the sister-group relationshipbetween gnathostomulids and annelids. Instead,they included the new taxon in the polychaete fam-ily Dorvilleidae. They followed the new concepts (seeWestheide, 1987) that neoteny and progenesis mayhave a major role in the adaptive radiation of inter-stitial annelids and the characteristics of the inter-stitial dorvilleids may be included, although Neote-notrocha did not “fit” into any of the hypotheticalreduction series for interstitial polychaetes (West-heide and Riser, 1983; Westheide, 1984, 1987). Ithas been thought that the loss of setae and parapo-dia happened after the loss of jaws and other ele-ments of the pharyngeal apparatus. Suddenly, Neo-tenotrocha turned up with a full set of jaw elements,but with reduced parapodia and without polychaetesetae. It appears that the archiannelid problem willnever be resolved by morphological data. That allso-called archiannelids are only very advancedpolychaetes is very easy to show in a hypotheticalreduction series, but the “true” evolutionary eventsof archiannelids may be more complex.
Unfortunately, it is possible that some “archian-nelids” may be primarily adapted to interstitial lifevery early in animal evolution, e.g., Diurodrilidaeand Lobatocerebridae (Rieger, 1980), whereas Pro-todrilidae, Saccocirridae, Protodriloidae, Nerillidae,and interstitial Dorvilleidae are secondarily modi-fied polychaetes. It is not possible for morphologicaldata alone to show the difference between primaryadaptation to interstitial life or secondary adapta-tion happening several times in several phyla with-out any close relationship. In fact, a modern cladisticanalysis would have difficulties helping us with anew animal such as Limnognathia maerski. In grossanatomy it clearly exhibits characters of an intersti-tial annelid, e.g., tactile bristles, ciliophores, and thebody plan.
36 R. M. KRISTENSEN AND P. FUNCH

The hypothesis that Micrognathozoa could be aninterstitial polychaete is not supported by our TEMand SEM observations, but this theory should beexamined in the light of the old theory of Semper(1872), who proposed that Rotifera are only simpleneotenic annelids. When Semper discovered the ab-errant rotifer Trochosphaera in 1859, he stated thatthis rotifer was of the primitive ancestral type, andall rotifers developed by neoteny from a trochophorelarva of an annelid. This theory was abandoned af-ter it was clearly shown that Trochosphaera is avery specialized floscularicean rotifer (Hyman,1951). Still, the Annelida–Rotifera relationshipshould not be ruled out in any discussion. Today,most scientists believe that rotifers evolved from asmall, ciliated, creeping ancestor (Ruppert andBarnes, 1994) and not from a trochophore-like an-cestor. The well-developed buccal field seen in someDicranophorus species may represent a vestige ofthe ancestral ciliation, the neurotroch. Many vermi-form phyla have ventral ciliation, but the similari-ties between the neotenous polychaetes, e.g., Neote-notrocha, are very convincing. The behavior of N.sterreri is rotifer-like, especially swimming and for-aging behavior. The jaw apparatus consists superfi-cially of a trophi-like structure, or perhaps it is moresimilar to the jaws of Limnognathia maerski. Thepincer-like mandibles of Neotenotrocha superficiallyresemble the main jaws (ja2) of Limnognathia, thefree denticles of the maxillary apparatus resemblethe pseudophalangia (ja1), and the basal plates re-semble the molar structure (also called the basalplates) in Limnognathia. Without our TEM knowledgeof dorvilleid-jaws (Purschke, 1987), the rotiferan tro-phi (Clement and Wurdak, 1991; Rieger and Tyler,1995; Kleinow and Wratil, 1996) and gnathostomu-lid jaws (Herlyn and Ehlers, 1997), we could notresolve the type of Limnognathia jaws. The tubularsubstructures of the Limnognathia jaw are seenclearly in Figure 27, and such substructures arepresent in the fulcrum of all Rotifera and in thearticularium (lamellae symphyses) of Gnathostomu-lida, but never in dorvilleid Polychaeta. The expla-nation for the similarity in gross morphology of thejaws of dorvilleid polychaetes (Neotenotrocha) andmicrognathozoans (Limnognathia) is convergence.Furthermore, Neotenotrocha is an advanced neo-tenic polychaete, and its jaws can very easily beplaced into a reduction series from complex dorvil-leid jaws (Westheide, 1982) to the situation in whichjaws are totally lost, as in dinophilids (Eibye-Jacobsen and Kristensen, 1994).
When Ahlrichs (1995) established the monophy-lum Gnathifera (Gnathostomulida 1 Rotifera 1 Sei-son 1 Acanthocephala), he indicated Platyhel-minthes as the sister-group. The monophyly ofPlatyhelminthes and Gnathifera may be seen in anew light after the discovery of the Micrognathozoa.Limnognathia maerski has several characters incommon with both Gnathostomulida/Rotifera and
interstitial Annelida. Except for the tactile bristles(compound cilia), Limnognathia shows no similari-ties with the Platyhelminthes. The tactile bristlesare seen in many meiofaunal groups, and these sen-sory structures may be an adaptation to small sizeor to an interstitial way of life (Ax, 1966). Platyhel-minthes lack all the characters found in the pharyn-geal apparatus of Gnathifera, and today several au-thors (Nielsen, 1995; Sørensen et al., 2000) placePlatyhelminthes as an ingroup in Euspiralia underthe monophylum Parenchymia (Platyhelminthes 1Nemertini). To remove the Platyhelminthes as thesister-group to Gnathifera and place them as aningroup in Euspiralia causes a new problem: What isthe sister-group to the monophylum Gnathifera?There are two possibilities: 1) the Euspiralia is thesister-group to Gnathifera or 2) the Aschelminthesis a monophylum and Cycloneuralia (Gastrotricha 1Nematoda 1 Nematomorpha 1 Priapulida 1 Kino-rhyncha 1 Loricifera) is the sister-group to Gnathifera.
Limnognathia shares some characters in commonwith Euspiralia, especially Annelida, which are notseen in the other members of Gnathifera. Thesecharacters are the ciliation of the pharyngeal epithe-lium and ventral ciliation (ciliophores). It is too earlyto state that these characters are plesiomorphic.
Monophyletic, Paraphyletic, or PolyphyleticOrigin of Aschelminthes
The Aschelminthes currently consist of twoclades: Cycloneuralia (Gastrotricha, Nematoda;Nematomorpha, Priapulida, Kinorhyncha, andLoricifera) and Gnathifera (Gnathostomulida, Euro-tatoria, Seisonida, Acanthocephala, and the newgroup, Micrognathozoa). Other groups have beensuggested to belong to Aschelminthes, such as Tar-digrada (Ruppert and Barnes, 1994), Entoprocta(Hyman, 1951; Bartolomaeus, 1993), and Chaeto-gnatha (Nielsen, 1995). Although we find these ideasinteresting, we will not discuss them further in thissection.
The phylum Aschelminthes was proposed byGrobben in 1910 (see Hyman, 1951) to cover anassemblage of animal groups with the so-calledpseudocoel or pseudocoelom. The five groups wereRotifera, Gastrotricha, Kinorhyncha, Nematoda,and Nematomorpha. Later, Hyman (1951) includedthe Acanthocephala and Entoprocta as “thepseudocoelomate Bilateria.” Still, in many textbooksthe name “Pseudocoelomata” is used for Aschel-minthes, inspired by Hyman’s coelom theory: “prim-itive” Acoelomata without coelom, Pseudocoelomatawith a persisting blastocoel, and “advanced” Eucoe-lomata with “true coelom.” Unfortunately, the coe-lom characters alone have no value in the interpre-tation of metazoan phylogeny, at least not at thehigher levels such as classes or phyla (Haszprunar,1996b). Bartolomaeus (1993) has excellently dis-cussed the coelom concept on the basis of his new
37MICROGNATHOZOA: A NEW CLASS

ultrastructural studies. Rotifera, Acanthocephala,large Nematoda, and one family of Loricifera (Pli-ciloricidae) are pseudocoelomate. In Priapulida, allthree body cavity conditions exist: acoelomate,pseudocoelomate, and a body cavity with an incom-plete epithelial lining (eucoelomate) in Meiopriapu-lus fijiensis (see Storch et al., 1989). Other Aschel-minthes are acoelomate, e.g., Gnathostomulida,Gastrotricha, many small Nematoda, and someLoricifera and Kinorhyncha (Kristensen, 1995).
Several authors have been aware of the pitfall ofrecognizing the Pseudocoelomata (Aschelminthes)as a monophylum because they require special con-sideration in terms of convergent evolution (Rup-pert, 1991a). The so-called pseudocoelom is the em-bryonic blastocoel that persists in the adultorganism, regardless of the time at which it firstappeared during ontogeny. Rieger (1976) regardedthe pseudocoelom or pseudocoel as a neotenic condi-tion secondarily derived from coelomate ancestorsrather than a plesiomorphic character in metazoanevolution (see Brusca and Brusca, 1990; Haszpru-nar, 1996b; Valentine, 1997). Others still believethat Aschelminthes constitute a monophyletic group(Nielsen, 1995), but they now avoid the termPseudocoelomata. When the phylum Loricifera wasdescribed (Kristensen, 1983), it was assigned to As-chelminthes on the basis of at least one synapomor-phic character shared with each other phylum ofAschelminthes, except for the Gnathostomulida.Some of these characters have later been recognizedas not synapomorphic but clearly convergent, e.g.,the similarities of the lorica of rotifers with dorsalantennae and the lorica of Nanaloricidae withflosculae are not homologous but convergent. TheLoricifera clearly belongs to the Cycloneuralia-cladeand the Rotifera to the Gnathifera-clade.
Likewise, it can be difficult to answer the questionabout the phylogenetic position of Micrognathozoa:is the new animal Limnognathia maerski an aschel-minth? The ultrastructure of the jaw clearly indi-cates affinities with the Gnathifera. It lacks thesyncytia in the epidermis and therefore cannot beincluded in Syndermata (Rotifera and Acanthoceph-ala), but it has an intracellular matrix in the dorsalplates, as in many Rotifera. The ventrum ofL. maerski has multiciliated cells and cannot beincluded in Gnathostomulida (epidermal cells arealways monociliated), but L. maerski has a tempo-rary dorsal anus like Gnathostomula. The monocili-ation of the terminal cell in the protonephridia andthe ciliated pharynx may be plesiomorphic charac-ters shared with other Euspiralia.
The question about monophyly, paraphyly, orpolyphyly of Aschelminthes is very difficult to an-swer (Winnepenninckx et al., 1995; Wallace et al.,1996) and there exists considerable disagreementbetween morphological and molecular data. Whenwe described Cycliophora (Funch and Kristensen,1995, 1997), we placed the new group as a sister-
group to the Entoprocta/Ectoprocta clade in Euspi-ralia (Protostomia). We argued that the chordoidlarva of Symbion pandora is a modified trochophore(Funch, 1996) in our conclusion. Later, Winnepen-ninckx et al. (1998) used the 18S ribosomal DNAdataset to include Cycliophora in the Rotifera/Acanthocephala clade. In a new cladistic approachbuilt only on morphological data, Sørensen et al.(2000) placed Entoprocta as a sister-group to Cyclio-phora in the Euspiralia. Ectoprocta is placed in theDeuterostomia by these authors.
While nearly all morphologists discussed themonophyletic or paraphyletic nature of Aschel-minthes (Lorenzen, 1985; Ehlers et al., 1996;Nielsen et al., 1996; Neuhaus et al., 1996), newmolecular data may indicate a polyphyletic origin ofAschelminthes (Aguinaldo et al., 1997). The newphylogenetic analysis of 18S ribosomal DNA se-quences indicates a close relationship between Pan-arthropoda (Tardigrada, Onychophora, Arthropoda)and molting Aschelminthes (Nematoda, Nemato-morpha, Priapulida, Kinorhyncha [and Loricifera?]).Ecdysis (the molting cycle) has only arisen once inthe Animal Kingdom. The new clade is called Ec-dysozoa and contains all molting animals (Gareyand Schmidt-Rhaesa, 1998). The Aschelminthesshould then be a polyphyletic group. This new the-ory could explain why Tardigrada may have charac-ters from both Aschelminthes and Euarthropoda.The theory is also supported by fossil evidence fromthe Cambrian (Conway Morris, 1989; Kristensen,1991). Some segmental worms, e.g., Opabinia, couldbe aschelminths with a segmental cuticle, e.g., asthe kinorhynchs. Unfortunately, the clade Ecdyso-zoa is not supported by many morphological data,e.g., the Gastrotricha, belonging to the Cycloneura-lia (Nielsen et al., 1996), do not molt. Schmidt-Rhaesa et al. (1998) suggest Gastrotricha as a sister-group to Ecdysozoa based on the synapomorphiccharacters: triradiate muscular sucking pharynx,terminal mouth opening, and bilayered cuticle witha trilaminate epicuticle. Most morphologists haveplaced Gastrotricha as a sister-group to theNematoda/Nematomorpha clade. The new animalLimnognathia maerski does not resolve the problemabout the Aschelminthes phylogenetic relationship,which still remains unclear.
Additional Meiofauna of theIsunngua Spring
The usually characteristic fauna of warm homo-thermic springs (Kristensen, 1987) was not found inthe cold spring of Isunngua (Figs. 34–36). Instead, acold water fauna seen in some mud volcanoes andcold homothermic springs was found. There is a richfauna of turbellarians in the spring, including thekalyptorhynch genus Geoplana. The gastrotrichfauna is relatively poor; only one species, Chaetono-tus sp., was found. The rotifer fauna in the sediment
38 R. M. KRISTENSEN AND P. FUNCH

Figs. 34–36. Maps of the type locality for Limnognathia maerski. Fig. 34: Map of Greenland indicating Disko Island. Fig. 35: Mapof Disko Island showing Kvandalen. Fig. 36: Close-up of Mudderbugten (The Bay of Mud) and Kvandalen (The Valley of Angelika)showing the type locality, the cold spring at Isunngua, the Lymnaea lake where Limnognathia maerski was collected in 1979, and allthe known homothermic springs in Kvandalen.

Figs. 37–41. The type locality and living Limnognathia maerski nov. gen. et sp. Fig. 37: The first sledge expedition from ArcticStation to Isunngua (5 April 1978). Fig. 38: The type locality for Limnognathia maerski. Note the moss cushions where more than 100specimens were collected 25 July 1994. Fig. 39: The base camp for the Arctic Field Course in 1994. The small cold Isunngua spring wasused for drinking water and the spring can be seen just behind the camp (22 July 1994). Fig. 40: Limnognathia maerski nov. gen. etnov. sp. Ventral view of living Limnognathia maerski. The so-called eye spots can be seen as round vesicles just in front of thepharyngeal bulb. The animal is 115 mm long. Fig. 41: Limnognathia maerski nov. gen. et nov. sp. Dorsal view of a slightly squeezedanimal. The sensoria located on the head and the nucleus in the large oocyte are clearly seen. The animal is 110 mm long.
40 R. M. KRISTENSEN AND P. FUNCH

Figs. 42–46. Search for Limnognathia maerski year-round. Fig. 42: Isunngua spring in autumn (27 August 1996). Fig. 43: Diggingdown to the type locality (5 May 1998). Fig. 44: The spring was totally frozen beneath 1.5 m snow cover (5 May 1998). Note the researchvessel Porsild in the Vaigat Strait and Arveprinsens Ejland more than 50 km away. Fig. 45: The third moor and Isunngua spring earlyin the spring (20 May 1995). Only one juvenile was collected. Fig. 46: Breaking the sea ice to reach Isunngua (20 May 1995). Note theCretaceous sand on the beach of Isunngua and the mountain Pingu in the background.

consists of Nothololca squamula f. lapponica, Lep-adella patella, and Encentrum unicinatum. The ro-tifer fauna in the water mosses in the spring con-sists of Colurella obtusa, Lepadella acuminata,Lecane lunaris, Trichocerca weberi, T. longiseta, andT. rattus rattus.
The tardigrade fauna is very interesting becausetwo of the species were originally described from amud volcano (Eohypsibius nadjae) and a cold homo-thermic spring near Isunngua (Microhypsibiusminimus). The other members of the tardigradefauna are the following species: Amphibolus nebulo-sus, A. weglarskae, Dactobiotus ambiguus, Dactobio-tus nov. sp., Diphascon scoticum, Hypsibius arcticus,H. dujardini, Isohypsibius elegans, Macrobiotusechinogenitus, M. richtersi, and Murryon pullari. Allthe mentioned species are very common in freshwa-ter (lakes, ponds, or rivers) or in wet soil at DiskoIsland. The copepod Bryocampus arcticus was rare,as well as other members of the meiofaunal crusta-ceans. The marine fauna element found in the ra-dioactive salt springs at Disko Island (Kristensen,1977, 1982, 1987) was not present in the Isunnguaspring.
Ecological Remarks
The senior author visited the Isunngua area thefirst time by a sledge expedition from Arctic Stationin the winter of 1978 (Fig. 37). The temperature atnoon was measured as 229°C on 5 April 1978. Thespring of Isunngua has abiotic parameters similar toother small springs dominated by melt water atDisko Island: the temperature varies throughout theyear and the springs have a very low conductivity(43–63 mmho). The pH (6.4) is low because thespring is running through three moor systems. Afterthe last moor, the spring runs in a deep channel ofwell-sorted Cretaceous sand (Figs. 38, 39, 42). Thereexists a rich interstitial meiofauna (e.g., the rotiferNotholca squamula) in this former tropical sand, butthe taxon Limnognathia maerski was only foundassociated with the spring mosses. The radioactivityin the spring is very low (about 40–60 counts persecond). In the salt springs at Disko Island the ra-dioactivity may be up to 1,500 cps (Kristensen,1987). The cold spring of Isunngua is totally frozenduring the long Arctic winter. At Disko Island thismeans that all springs, which are not homothermal,are frozen from about early November to early May.The type locality was visited twice in May (Figs.43–46). Both in 1995 and 1998 spring came early toDisko Island and we could reach the remote place bythe research vessel Porsild without any problemwith sea ice. On 21 May 1995, the spring was notcovered with snow and the water temperature wasalready 4°C, while the air temperature was 1.5°C.The meiofauna in the spring was already activated.Seven species of monogonont rotifers and five spe-cies of tardigrades were found in the spring mosses,
but only a single specimen of Limnognathia maerskiwas found. The length of the specimen measuredonly 84 mm. It is absolutely the smallest specimenwe have ever found. The animal was transparentand it could be a newly hatched juvenile. It couldindicate that Limnognathia survives the Arctic win-ter as eggs, a different life strategy than some of theother meiofaunal animals in the spring, whichclearly overwinter in all different stages of the lifecycle, e.g., the large tardigrade Amphibolus nebulo-sus, which was present in the spring as eggs, juve-niles, and adults.
The spring was revisited on 5 May 1998. The airtemperature was 22.2°C. The spring was coveredwith nearly two meters of snow (Figs. 43, 44). Thecrew of Porsild dug down to the Isunngua spring;the spring was totally frozen and the ice tempera-ture in the spring was 20.1°C. Frozen moss sampleswere carried to the Arctic Station at Qeqertarsuaq.Several species of rotifers, including two species ofbdelloids and four species of monogononts, were ob-served. The very rare tardigrade Eohypsibius nadjae(see Bertolani and Kristensen, 1987) and a species ofthe gastrotrich genus Chaetonotus were also presentin the frozen samples. Not a single specimen of Lim-nognathia was found. This finding supports the the-ory that with Limnognathia overwintering as eggs,the eggs hatch early in the summer and the maxi-mum of the population will be in July–August. Latein August the population begins to decline. The twodifferent eggs (unsculptured and sculptured) foundin August 1994 may be summer eggs and overwin-tering eggs (Fig. 7). The so-called unsculptured,abortive eggs laid by several females may not beinduced by high temperature in the culture, butcould instead be true summer eggs. The sculpturedeggs could then be winter eggs.
Temperature data recorders placed in the sub-merged mosses at the type locality in the winter of1996/97 from 26 October to 1 May showed that Lim-nognathia maerski endures minimum temperatures(22.7°C) at the end of October. Typical winter tem-peratures in the frozen mosses are remarkably sta-ble and range between 21.0 to 20.5°C beneath theprotective snow cover, with an average for the wholeperiod of 0.8°C. Summer temperatures were mea-sured between 2 July and 16 August 1997, each for4.5 h. The average temperature in this period was8.7°C. The highest temperatures are in the after-noon and evening (maximum temperature: 14.2°Cwas measured on 6 July 1997, 6 PM). Low summertemperatures are typically in the morning (mini-mum temperature: 4.6°C was measured on 16 Au-gust 1997, 6 AM). We attempted to record tempera-tures for an entire year, but the recorder was lost.Probably the powerful melting water flushed therecorder out into the sea. Although Limnognathiamaerski lives in Arctic springs, the temperaturesencountered are moderate. Adaptations to cope withlow temperatures do not seem to be important for
42 R. M. KRISTENSEN AND P. FUNCH

the adults. However, adaptation to attach firmly tothe substrate is important. Limnognathia probablyendures powerful currents caused by melting waterin the spring. Eggs are probably the stages thatdisperse over longer distances and they might sur-vive low temperatures.
Zoogeography
If rather similar vermiform phyla had divergedalready in the deep Precambrian (Wray et al., 1996;Sterrer, 1998), as some molecular data indicate, thezoogeographic data cannot help us in the phyloge-netic discussion, e.g., the cosmopolitan distributionof some low-water species of Gnathostomulida andTardigrada without obvious mechanisms for long-range dispersal only tell us that they are groups thatgo back to the supercontinent Pangaea in early Me-sozoic time. The earlier supercontinent Rodinia (seeMcMenamin, 1998) in the Precambrian would nothave any influence on worldwide distribution of re-cent species. Recently, what seems to be a memberof Micrognathozoa was found in Antarctica (DeSmet, pers. com.). The bipolar distribution of Mi-crognathozoa may indicate that the group is veryancient. The problem is that the spring at Isunnguais young compared with other springs throughoutthe world. In the last glaciation the Isunngua springwas, as were many of the homothermic springs atDisko Island, about 100 meters below sea level.When glaciation ended in the area about 6,900 yearsago, the land masses rose and subsequently thespring was left above sea level. This uplifting of theland masses happened so slowly that the marineforms of animals could adapt to freshwater in someof the homothermic springs (Kristensen, 1977). Thenew taxon, Limnognathia maerski, is found in a coldspring that is frozen for nearly half of the year. Itcannot go into anhydrobiosis, as do many of theother members of the meiofauna in the spring. Thus,it has a very low dispersal range. The animal may bea marine relict in the spring, but there are no othermarine animals in the Isunngua spring. More than20 years research for marine interstitial meiofaunajust outside the spring in Disko Bay (Kristensen andNiilonen, 1982; Kristensen and Nørrevang, 1982)did not produce any Micrognathozoa. The discoveryof three specimens of Limnognathia from 1979 inthe valley of Sullorsuaq/Kvandalen may indicatethat the new taxon belongs to an ancient true fresh-water fauna. The source of the cold homothermicspring at Sullorsuaq (Fig. 36) is about 100 metersabove sea level, and there is no evidence to assumethat the springs were under sea level during the lastglaciation. The finding of a “missing link” betweenthe marine gnathostomulids and the marine/freshwater rotifers in a remote Arctic spring is veryenigmatic, and we have no explanation as to why theanimal was not discovered before.
Phylogenetic Position of Micrognathozoa
The discovery of the new taxon, Limnognathiamaerski, gave the impetus to the proposal of thehomology of gnathostomulid and rotifer jaws (Riegerand Tyler, 1995) and the phylogenetic discussion ofthe validity of the monophylum Gnathifera (Ahl-richs, 1997; Herlyn and Ehlers, 1997). When weobserved the first living specimens of L. maerski in1994, it became clear to us that it was a uniqueanimal (Kristensen, 1995; Kristensen and Funch,1995). We have therefore discussed these new find-ings of morphological characters with several of ourcolleagues and a new computer-assisted cladisticanalysis of all Metazoa will soon be published (Sø-rensen et al., 2000). In this cladistic analysis boththe phylogenetic position of Cycliophora and Micro-gnathozoa will be treated. Here, we discuss only thephylogenetic position of Micrognathozoa within themonophylum Gnathifera (see Fig. 47).
We consider the possession of cuticular jaws withtubes (rods) as a synapomorphy for all Gnathifera(Fig. 47, apomorphy 1). Jaw-like structures arefound in other taxa as well, for instance, in theprobosces of kalyptorhynch turbellarians, in dorvil-leid polychaetes, and aplacophoran mollusks, butstudies of their ultrastructure show that none ofthese jaws are homologous with the jaws in Gnatho-stomulida, Micrognathozoa, and Rotifera (Riegerand Tyler, 1995; this article). We are uncertain ofthe origin of the basal plates in Gnathifera. A singlecuticular basal plate could belong to the ground pat-tern of Gnathifera; consequently, basal plates werelost in the stem lineage of Syndermata and becamepaired and modified in the stem lineage of Micro-gnathozoa. A second alternative is that paired basalplates were present in the ancestor of Gnathifera,and were completely lost in Syndermata. A thirdalternative is that the basal plates in Gnathostomu-lida and Micrognathozoa are not homologous. Weconsider the loss of the digestive system, includingthe pharyngeal apparatus, as an autapomorphiccharacter for Acanthocephala.
The presence of a monociliated cellular epidermishas been postulated to be present in the stem speciesof the Bilateria (Ehlers, 1985; Ax, 1985), but Sø-rensen et al. (2000) regard that a multiciliated epi-dermis evolved in the ancestor of Bilateria. Mono-ciliation in Gnathifera is only found on the overallsurface of the body in Gnathostomulida and in theprotonephridia of both Gnathostomulida and Mi-crognathozoa. Rieger (1976) interpreted completemonociliation with microvilli surrounding the ciliumas a plesiomorphic character. However, the longflagellar-like ciliation present in all gnathostomu-lids may be interpreted as an autapomorphic char-acter for gnathostomulids. Absence of motile cilia onthe dorsal and lateral sides and ventral multicili-ated epidermal cells may be interpreted as a syna-pomorphy of Micrognathozoa and Syndermata (Ro-
43MICROGNATHOZOA: A NEW CLASS
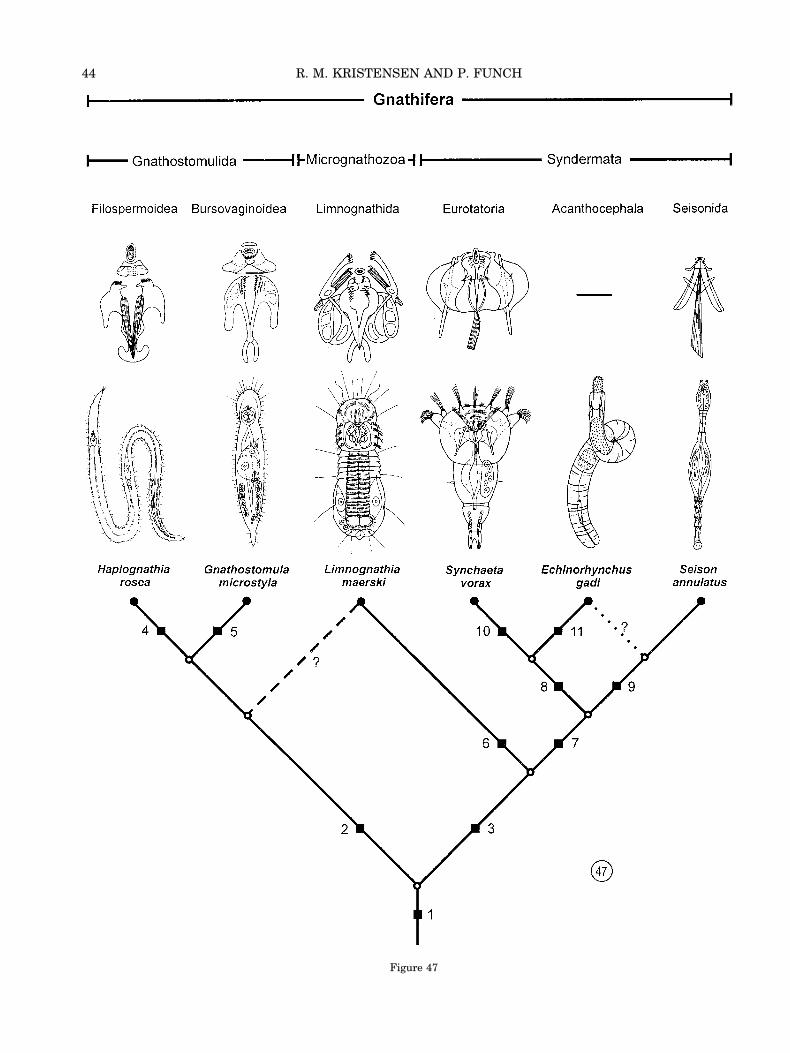
Figure 47
44 R. M. KRISTENSEN AND P. FUNCH

tifera 1 Acanthocephala). We consider the reductionof ventral ciliation in Eurotatoria 1 Seisonida andthe total absence of epidermal ciliation in Acantho-cephala (Dunagan and Miller, 1991) as apomorphiccharacters. Ventral ciliation may be a plesiomorphiccharacter for Limnognathia, while the presence ofventral ciliophores is an autapomorphy for Microgna-thozoa. The thick dorsal and lateral epidermalplates in Micrognathozoa are not cuticular but areformed by an intracellular matrix layer, as in Syn-dermata. We consider this character as a synapo-morphy for Micrognathozoa and Syndermata (Fig.47, apomorphy 3). The syncytial epidermis is anautapomorphy for the clade Syndermata, which in-cludes the Eurotatoria, Acanthocephala, and Sei-sonida. The new taxon, Micrognathozoa, lacks syn-cytia.
We also consider the presence of separate sexes asa plesiomorphic character in Gnathifera, and thepresence of hermaphroditism and a complex repro-ductive system in Gnathostomulida as autapomor-phic characters. Furthermore, we follow the idea ofSterrer (1972) that the presence of a filiform sper-matozoon with one flagellum and an acrosome is aplesiomorphic character, and that all other types ofspermatozoa are derived from this type.
Figure 47 shows our phylogeny of the main groupsof Gnathifera. It differs somewhat from Ahlrichs(1997) and Herlyn and Ehlers (1997). The two ques-tion marks indicate two other possibilities for rela-tionships in Gnathifera. The ultrastructure of themain jaws of Limnognathia maerski is similar to thejaws of advanced scleroperalian gnathostomulids,especially in the jaw apophyses and the pairedcauda. We have therefore previously suggested thatGnathostomulida and Micrognathozoa were closelyrelated (Kristensen and Funch, 1995). New SEMdata on the jaws of Haplognathia rosea and Rastro-gnathia macrostoma (see Sørensen, 2000) made us
consider that some of these jaw characters couldbelong to the ground pattern of Gnathifera, e.g., thelongitudinally arranged rods in the lamellae sym-physis of gnathostomulids might be homologouswith the striation present in the fulcrum of Eurota-toria (for review, see Markevich, 1989) and Sei-sonida (Segers and Melone, 1998). Other characters,such as the presence of a paired cauda, might beconvergently developed in some species of Gnatho-stomula and Limnognathia. But it is too early toconclude that the paired fibularia of Limnognathiaand the fibularization of the jaw apophyses ofGnathostomula belong to the ground pattern ofGnathifera. A consequence of fibularization belong-ing to the ground pattern of Gnathifera is that theso-called solid type (arc type) of jaws in somegnathostomulids represents a derived type. How-ever, this contradicts the most accepted classifica-tion of Gnathostomulida (Sterrer, 1972), where thecompact arc type is considered to have the plesi-omorphic characters and the row type with fibular-ization has the apomorphic characters. Sterrer’sclassification of the jaws is strongly supported by othercharacters, e.g., the sperm types and the fine structureof the reproductive organs. We therefore suggest thatMicrognathozoa is the sister-group to the taxon Syn-dermata, and not Gnathostomulida. An important sy-napomorphy for the clade Micrognathozoa 1 Synder-mata is the presence of an intracellular matrix withinthe epidermis. The micrognathozoans do not belong toSyndermata (Eurotatoria 1 Acanthocephala 1 Sei-sonida), because Limnognathia totally lacks the syn-cytia in the epidermis (Fig. 47, apomorphy 7).
Lorenzen (1985) argued that the characteristicsacs, lemnisci in acanthocephalans, were in factpresent in bdelloid rotifers but had been overlooked.Based on this and other morphological similaritiesLorenzen suggested that acanthocephalans weremerely aberrant bdelloid rotifers. The first phyloge-
Fig. 47. Phylogeny of the Gnathifera. The cladogram is modifiedfrom Ahlrichs (1997), Herlyn and Ehlers (1997), and Sørensen etal. (2000). Our suggestion is shown with solid lines. The brokenline with a question mark denotes the suggestion of Ahlrichs(1997) and the dotted line with a question mark is our alternativesuggestion (Kristensen and Funch, 1995). Ventral views of theanimals and their jaws. From left to right: Haplognathia rosea(Filospermoidea, Gnathostomulida); Gnathostomulida microstyla(Bursovaginoidea, Gnathostomulida); Limnognathia maerskinov. gen et nov. sp. (Micrognathozoa); Synchaeta vorax (Monogo-nonta, Eurotatoria); Echinorhynchus gadi (Palaeacanthocephala,Acanthocephala); Seison annulatus (Seisonida, Rotifera). Theblack squares 1–11 mark syn- and autapomorphies. 1. Gnathifera.Cuticular jaws with tubes composed of lucent material surround-ing an electron-dense core. 2. Gnathostomulida. Hermaphroditeswith complex reproductive system. Pharyngeal apparatus withmuscle capsule. 3. Micrognathozoa 1 Syndermata. Epidermiswith intracellular matrix, motile cilia absent on the surface ofdorsal and lateral trunk regions. 4. Filospermoidea. Elongation ofthe body with a long and slender rostrum. Compact jaws with8–12 longitudinally arranged rods in lamellae symphyses. 5.
Bursovaginoidea. Fibularization of the jaw apophyses. Myosyn-cytia in the pharyngeal apparatus. Complex female organs with abursa and often a vagina. Male opening with an injectory penisoften consisting of 8–12 intracellular rod-like structures. Sper-matozoon strongly modified, lacking flagellar or centriolar struc-tures. 6. Micrognathozoa. Ventral locomotory organ consists oftwo rows of multiciliated cells (trunk ciliophores). Ventral epider-mis covered only with a thin glycocalyx, except for the largecuticular oral plate. Dorsal and lateral intracellulary platespresent. The solid parts of the jaws are very complex, with pairedbasal plates, paired lamellae orales, paired fibularia, and threesets of paired jaws. 7. Syndermata. Syncytial epidermis; epider-mal cells with apical crypts. Eutelic epidermis. Spermatozoonwith anteriorly inserted flagellum. 8. Eurotatoria 1 Acantho-cephala. Spermatozoon without acrosome. 9. Epizoic. The headciliation consists of a few tufts of stiff bristles. The trophi form ananterior cuticular sheath. Presence of spermatophore. 10. Euro-tatoria. Corona, unpaired retrocerebral glands, female reproduc-tive organs with vitellarium. 11. Acanthocephala. Endoparasiticwith a two-host life cycle. Epidermal lacunar system. Probosciswith intraepidermal hooks. Females with a uterine bell.
45MICROGNATHOZOA: A NEW CLASS

netic analyses of the relationship of acanthocepha-lans to rotifers, using nuclear 18S ribosomal DNAand mitochondrial 16 ribosomal RNA genes, gavestrong support for a sister-group relationship be-tween Acanthocephala and Bdelloidea (Garey et al.,1998). Ahlrichs (1997) argued that ultrastructuralfeatures such as the presence of dense bodies in thespermatozoa and an epidermis with similar bundlesof filaments in both Acanthocephala and Seisonidagave support for a sister-group relationship (Fig. 47,dotted line), a point of view supported by Herlyn andEhlers (1997). We are in favor of a monophyletictaxon Eurotatoria consisting of the two classesMonogononta and Bdelloidea based on a number ofmorphological characters (Fig. 47). This point ofview is supported by a computer-generated cladisticanalysis of Animalia (Sørensen et al., 2000), wherethe sister-group to Eurotatoria is Acanthocephala.The loss of the acrosome in the spermatozoon is asynapomorphy supporting this relationship (Fig. 47,apomorphy 8). Thus, acanthocephalans are in factlarge parasitic rotifers. A recent phylogenetic studyby Welch (2000) using a nuclear protein-coding geneand comprising sequences for several species of ro-tifers actually supports our interpretation, and thesame conclusion was made by Garcıa-Varela et al.(2000) in their molecular study of Acanthocephalaand Eurotatoria.
In the classical taxonomy of rotifers the orderSeisonidea is placed in the class Digononta, alsocontaining Bdelloidea (Beauchamp, 1965; Nogradyet al., 1993). Segers and Melone (1998), on the otherhand, treat Seisonidea as an independent class ofrotifers. We consider the presence of paired gonadsas a plesiomorphic character for Gnathifera and sug-gest that Digononta is paraphyletic. Seisonidea isinstead the sister-group to Eurotatoria 1 Acantho-cephala. The two marine species of Seison are veryaberrant rotifers, especially in trophi. In fact, differ-ent studies disagree in labeling the sclerites consis-tently (see Segers and Melone, 1998).
CONCLUSION
The jaws of Gnathostomulida, Rotifera, and thenew group Micrognathozoa are homologous struc-tures. The ultrastructure of cuticular elements ofthe jaws in all three groups has a tubular substruc-ture. The tube-like rods consist of an osmiophiliccore surrounded by an osmiophobic layer of cuticle.The rods may be present only in special areas or thewhole jaw apparatus is built of parallel-layered rods.The three mentioned groups 1 Acanthocephala havebeen included in the monophylum group Gnathifera.The absence of pharyngeal ciliation cannot be used asan autapomorphy of the Gnathifera, as suggested byHerlyn and Ehlers (1997). The Micrognathozoa has aciliated pharyngeal epithelium, as do several intersti-tial polychaetes. Furthermore, the pharyngeal epithe-lium is covered with a thin cuticle, as in Rotifera.
The new animal, Limnognathia maerski nov. gen.et sp., is probably closely related to both Rotiferaand Gnathostomulida. Rotifera 1 Acanthocephalaand Micrognathozoa might be sister-groups, whileGnathostomulida may be the sister-group to thisassemblage. The monophyly of Plathelminthomor-pha (Platyhelminthes and Gnathostomulida) andPlatyhelminthes (see Ax, 1996; Haszprunar, 1996a)is doubtful. The discovery of Limnognathia maerskisupports the sister-group relationship of Gnathosto-mulida and Syndermata (Rotifera-Acanthocephalaclade), as first stated by Rieger and Tyler (1995) andlater supported by Ahlrichs (1995, 1997) when heestablished the monophylum Gnathifera. The newmolecular data of Gnathostomula based on 18S ri-bosomal DNA are interesting, but also confusing.The data of Littlewood et al. (1998) supported arelationship with Gnathostomulida and theNematoda-Chaetognatha clade, while the data ofZrzavy et al. (1998) supported an old theory about asister-group relationship between Gastrotricha andGnathostomulida (Neotrichozoa). The old theorywas based on morphological evidence, such as thepresence of monociliated epidermal cells in all Gna-thostomulida and some Gastrotricha (Rieger, 1976),but we consider the monociliation in the epidermalcells and the terminal cells of the protonephridiumas a plesiomorphic character in all Euspiralia, as didRieger (1976), and so not sufficient to support themonophyly of Gnathostomulida 1 Gastrotricha. Be-fore we can abandon the clade Neotrichozoa, how-ever, we need to have molecular data on Micro-gnathozoa. The ultrastructural similarities of thenew animal to certain gastrotrichs and interstitialpolychaetes may be convergent, or may show theplesiomorphic condition in Euspiralia.
Autapomorphies for Micrognathozoa are the fol-lowing: on the dorsum, the presence of dorsal platessclerotized by an intracellular matrix, serially ho-mologous tactile bristles consisting of one to threecilia, and zipper-junctions between all dorsal epider-mal cells; on the ventrum are 18 trunk ciliophoresand paired ventral head ciliophores flanking thepharyngeal apparatus, a posterior adhesive ciliatedpad, and sclerotization only in an oral plate; allother ventral cells lack cuticle and sclerotization.The complexity of the cuticular pharyngeal elementsis greater than in other members of Gnathifera andits special features constitute good autapomorphies.
Limnognathia maerski cannot be included in anyexisting group without difficulty. Therefore, we haveestablished a new class, Micrognathozoa, for thenew freshwater animal, indicating that it has thesame rank as Gnathostomulida and Syndermata inthe Gnathifera. We suppose a closer relationship(sister-group relationship) to the Syndermata thanto Gnathostomulida, and we support the theory ofmonophyly of Gnathifera.
46 R. M. KRISTENSEN AND P. FUNCH

ACKNOWLEDGMENTS
We gratefully acknowledge the collaboration withthe other two teachers, Dr. Jacob Larsen (BotanicalInstitute, University of Copenhagen) and Dr. HelgeA. Thomsen (Danish Institute for Fisheries Re-search), and all our students at the Arctic FieldCourse, Godhavn, 1994. We thank the crews of boththe old and the new research vessel Porsild, and wethank Ms. Stine Elle, Cand. Scient. Aslak Jør-gensen, Mr. Geert Brovad, and Mrs. Birgitte Rubæk,who made all the final drawings and illustrations;Ms. Eva von Linstow Roloff for excellent technicalsupport in ultrasectioning. The moss species werekindly determined by Dr. Gert Steen Mogensen (Bo-tanical Museum, University of Copenhagen). Wealso thank Cand. Scient. Martin V. Sørensen andDr. Wilko H. Ahlrichs for interesting discussions ona number of matters relevant to this article. Weespecially thank Professor Dr. Frederick W. Harri-son, who forced us after a 4-year delay to publish ournew findings, and the Dean of Science Faculty ofUniversity of Copenhagen, Dr. Henrik Jeppesen,who always supported our Arctic Research. Finan-cial assistance came from the Carlsberg Foundation(970345/30 - 488) and the Danish Research Agency(9701589) to R.M. Kristensen, and a grant to P.Funch from the Danish Research Agency (9801880).
LITERATURE CITED
Aguinaldo AMA, Turbeville JM, Lindford LS, Rivera MC, GareyJR, Raff RA. 1997. Evidence for a clade of nematodes, arthro-pods and other moulting animals. Nature 387:489–493.
Ahlrichs WH. 1993a. Ultrastructure of the protonephridia of Sei-son annulatus (Rotifera). Zoomorphology 113:245–251.
Ahlrichs WH. 1993b. On the protonephridial system of thebrackish-water rotifer Proales reinhardti (Rotifera, Monogo-nonta). Microfauna Mar 8:39–53.
Ahlrichs WH. 1995. Ultrastruktur und Phylogenie von Seisonnebaliae (Grube 1859) und Seison annulatus (Claus 1876). Got-tingen: Cuvillier Verlag. p 310.
Ahlrichs WH. 1997. Epidermal ultrastructure of Seison nebaliaeand Seison annulatus, and a comparison of epidermal struc-tures within the Gnathifera. Zoomorphology 117:41–48.
Ax P. 1956. Die Gnathostomulida, eine ratselhafte Wurmgruppeaus dem Meeressand. Abh math-naturw Kl Akad Wiss Mainz8:1–32.
Ax P. 1966. Die Bedeutung der interstiellen Sandfauna fur allge-meine Probleme der Systematik, Okologie und Biologie. Verof-fentl. d. Inst. f. Meeresforschung Bremerhaven, 2. Sonderband:15–66.
Ax P. 1967. Diurodrilus ankeli nov. spec. (Archiannelida) von dernordamerikanischen Pazifikkuste. Ein Beidrag zur Morpholo-gie, Systematik und Verbreitung der Gattung Diurodrilus. ZMorphol Oeko Tiere 60:5–16.
Ax P. 1985. The position of the Gnathostomulida and Plathel-minthes in the phylogenetic system of the Bilateria. In: ConwayMorris S, George JD, Gibson R, Platt HM, editors. The originsand relationships of lower invertebrates. Oxford: ClarendonPress. p 168–180.
Ax P. 1989. Basic phylogenetic systematization of the Metazoa.In: Fernholm B, Bremer K, Jornvall H, editors. The hierarchy oflife. Amsterdam: Elsevier Science Publishers. p 229–245.
Ax P. 1996. Multicellular animals: a new approach to the phylo-genetic order in nature, vol. 1. Berlin: Springer-Verlag. p 225.
Bartolomaeus T. 1993. Die Leibeshohlenverhaltnisse undNephridialorgane der Bilateria-Ultrastruktur, Entwicklungund Evolution. Habilitationsschrift. Gottingen: UniversitatGottingen. p 293.
Bertolani R, Kristensen RM. 1987. New records of Eohypsibiusnadjae Kristensen, 1982, and revision of the taxonomic position oftwo genera of Eutardigrada (Tardigrada). In: Bertolani R, editor.Biology of tardigrades. Modena: U.Z.I. 1, Mucci. p 359–372.
Boaden PJS. 1975. Anaerobiosis, meiofauna and early metazoanevolution. Cah Biol Mar 15:367–378.
Boaden PJS. 1989. Meiofauna and the origins of the Metazoa.Zool J Linn Soc 96:217–227.
Brusca RC, Brusca GJ. 1990. Invertebrates. Sunderland, MA:Sinauer Associates. p 992.
Cavalier-Smith T. 1998. A revised six-kingdom system of life. BiolRev Camb Philos Soc 73:203–266.
Clement P, Wurdak E. 1991. Rotifera. In: Harrison FW, RuppertEE, editors. Microscopic anatomy of invertebrates, vol. 4. As-chelminthes. New York: Wiley-Liss. p 219–297.
Conway Morris S. 1989. Burgess Shale faunas and the Cambrianexplosion. Science 246:339–346.
De Smet WH. 1996. Rotifera. The Proalidae (Monogononta), vol.4. In: Dumont HJF, editor. Guides to identification of microin-vertebrates of the continental waters of the world, vol. 9. Am-sterdam: SPB Academic. p 102.
De Smet WH. 1997. Rotifera. The Dicranophoridae (Monogo-nonta), vol. 5. In: Dumont HJF, editor. Guides to identificationof microinvertebrates of the continental waters of the world,vol. 12. Amsterdam: SPB Academic Publishing. p 325.
De Smet WH, Van Rompu EA. 1994. Rotifera and Tardigradafrom some cryoconite holes on a Spitsbergen (Svalbard) glacier.Belg J Zool 124:27–37.
Dunagan TT, Miller DM. 1991. Acanthocephala. In: Harrison FW,Ruppert EE, editors. Microscopic anatomy of invertebrates, vol. 4.Aschelminthes. New York: Wiley-Liss. p 299–332.
Ehlers U. 1985. Das phylogenetische System der Plathelminthes.Stuttgart: Gustav Fischer.
Ehlers U, Ahlrichs W, Lemburg C, Schmidt-Rhaesa A. 1996.Phylogenetic systematization of the Nemathelminthes (Aschel-minthes). Verh Dtsch Zool Ges 89:8.
Eibye-Jacobsen J. 1997. Development, ultrastructure and func-tion of the pharynx of Halobiotus crispae Kristensen, 1982(Eutardigrada). Acta Zool (Stockholm) 78:329–347.
Eibye-Jacobsen D, Kristensen RM. 1994. A new genus and spe-cies of Dorvilleidae (Annelida, Polychaeta) from Bermuda, witha phylogenetic analysis of Dorvilleidae, Iphitimidae and Di-nophilidae. Zool Scr 23:107–131.
Fenchel T, Riedl RJ. 1970. The sulfide system: a new bioticcommunity underneath the oxidized layer of marine sandy bot-toms. Mar Biol 7:225–268.
Fiege D. 1990. Morphology of the wings of euthecosomatouspteropods. Am Malac Bull 8:27–36.
Funch P. 1996. The chordoid larva of Symbion pandora (Cyclio-phora) is a modified trochophore. J Morphol 230:231–263.
Funch P, Kristensen RM. 1995. Cycliophora is a new phylum withaffinities to Entoprocta and Ectoprocta. Nature 378:711–714.
Funch P, Kristensen RM. 1997. Cycliophora. In: Harrison FW,Woollacott RM, editors. Microscopic anatomy of invertebrates,vol. 13. Lophophorates, Entoprocta, and Cycliophora. NewYork: Wiley-Liss. p 392–462.
Garcıa-Varela M, Perez-Ponce de Leon G, de la Torre P, Cum-mings MP, Sarma SSS, Laclette JP. 2000. Phylogenetic rela-tionships of Acanthocephala based on analysis of 18S ribosomalRNA gene sequences. J Mol Evol 50:532–540.
Garey JR, Schmidt-Rhaesa A. 1998. The essential role of “minor”phyla in molecular studies of animal evolution. Am Zool 38:907–917.
Garey JR, Schmidt-Rhaesa A, Near TJ, Nadler SA. 1998. Theevolutionary relationships of rotifers and acanthocephalans.Hydrobiologia 387/388:83–91.
Giribet G, Distel DL, Polz M, Sterrer W, Wheeler WC. Triploblas-tic relationships with emphasis on the acoelomates, and theposition of Gnathostomulida, Cycliophora, Plathelminthes and
47MICROGNATHOZOA: A NEW CLASS

Chaetognatha; a combined approach of 18S rDNA sequencesand morphology. Syst Biol (in press).
Graebner I. 1969. Vergleichende elektronenmikroskopische Un-tersuchung der Spermienmorphologie und Spermiogenese eini-ger Gnathostomula-Arten: Gnathostomula paradoxa (Ax 1956),Gnathostomula axi (Kirsteuer 1964), Gnathostomula jenneri(Riedl 1969). Microskopie 24:131–160.
Green CR, Bergquist PR. 1982. Phylogenetic relationships withinthe Invertebrata in relation to the structure of septate junctionsand the development of “occluding” junctional types. J Cell Sci53:279–305.
Haszprunar G. 1996a. Plathelminthes and Plathelminthomorpha— paraphyletic taxa. J Zoo Syst Evol Res 34:41–48.
Haszprunar G. 1996b The Mollusca: coelomate turbellarians ormesenchymate annelids? In: Taylor JD, editor. Origin and evo-lutionary radiation of the Mollusca. Oxford: Oxford UniversityPress. p 1–28.
Herlyn H, Ehlers U. 1997. Ultrastructure and function of thepharynx of Gnathostomula paradoxa (Gnathostomulida).Zoomorphology 117:135–145.
Hyman LH. 1951. The invertebrates: Acanthocephala, Aschel-minthes, and Entoprocta. The pseudocoelomate Bilateria. NewYork: McGraw-Hill. p 572.
Kalt MR, Tandler B. 1971. A study of fixation of early amphibianembryos for electron microscopy. J Ultrastruct Res 36:633–645.
Kleinow W, Wratil H. 1996. On the structure and function of themastax of Brachionus plicatilis (Rotifera), a scanning electronmicroscope analysis. Zoomorphology 116:169–177.
Knauss EB. 1979. Indication of an anal pore in Gnathostomulida.Zool Scr 8:181–186.
Koehler JK, Hayes TL. 1969. The rotifer jaw: a scanning andtransmission electron microscope study. I. The trophi of Philo-dina acuticornis odiosa. J Ultrastruct Res 27:402–418.
Koste W. 1978. Rotatoria, Die Radertiere Mitteleuropas. Text-band 1 Tafelband. Berlin, Stuttgart: Gebruder Borntraeger. p673.
Kristensen RM. 1977. On the marine genus Styraconyx (Tardi-grada, Heterotardigrada, Halechiniscidae) with description of anew species from a warm spring on Disko Island. Astarte 10:87–91.
Kristensen RM. 1982. New aberrant eutardigrades from homo-thermic springs on Disko Island, West Greenland. In: NelsonDR, editor. Proceedings of the Third International Symposiumon the Tardigrada. Johnson City, TN: East Tennessee StateUniversity Press. p 203–220.
Kristensen RM. 1983. Loricifera, a new phylum with Aschel-minthes characters from meiobenthos. Z Zool Syst Evolut For-sch 21:163–180.
Kristensen RM. 1987. The “southern” flora and the “marine”fauna elements in the homothermic springs on Disko Island,West Greenland. In: Petersen GH, editor. Gronland Exkursion,2.-25. August 1987. Kiel: Institut fur Polarokologie. p 202–226.
Kristensen RM. 1991. 6. New higher taxa — presented by theirdiscoverers. Loricifera — a general biological and phylogeneticoverview. Verh Dtsch Ges 84:231–246.
Kristensen RM. 1995. Are Aschelminthes pseudocoelomate oracoelomate? In: Lacivelli G, Valvassori R, Candia CarnevaliMD, editors. Body cavities function and phylogeny. Selectedsymposia and monographs. U.Z.I. 8. Modena: Mucci. p 41–43.
Kristensen RM, Eibye-Jacobsen D. 1995. Ultrastructure of sper-miogenesis and spermatozoa in Diurodrilus subterraneus(Polychaeta, Diurodrilidae). Zoomorphology 115:117–132.
Kristensen RM, Funch P. 1995. En ny aschelminth medgnathostomulid-lignende kæber fra en kold kilde ved Isunngua.In: Ehrhardt C, editor. Arktisk Biologisk Feltkursus,Qeqertarsuaq/ Godhavn 1994. Copenhagen: University ofCopenhagen. p 73–82.
Kristensen RM, Niilonen T. 1982. Structural studies on Diuro-drilus Remane (Diurodrilidae fam.n.), with description of Diu-rodrilus westheidei sp.n. from the Arctic interstitial meiob-enthos, W. Greenland. Zool Scr 11:1–12.
Kristensen RM, Nørrevang A. 1977. On the fine structure ofRastrognathia macrostoma gen. et sp. n. placed in Rastro-gnathiidae fam. n. (Gnathostomulida). Zool Scr 6:27–41.
Kristensen RM, Nørrevang A. 1978. On the fine structure ofValvognathia pogonostoma gen. et sp. n. (Gnathostomulida,Onychognathiidae) with special reference to the jaw apparatus.Zool Scr 7:179–186.
Kristensen RM, Nørrevang A. 1982. Description of Psammodrilusaeificator sp. n. (Polychaeta), with notes on the Arctic intersti-tial fauna of Disko Island, W. Greenland. Zool Scr 11:265–279.
Lake PS. 1973. Trialdehyde fixation of crustacean tissue for elec-tron microscopy. Crustaceana (Leiden) 24:244–246.
Lammert V. 1991. Gnathostomulida. In: Harrison FW, RuppertEE, editors. Microscopic anatomy of invertebrates, vol. 4. As-chelminthes. New York: Wiley-Liss. p 19–39.
Littlewood DTJ, Telford MJ, Clough KA, Rohde K. 1998. Gnatho-stomulida — an enigmatic metazoan phylum from both mor-phological and molecular perspectives. Mol Phylogenet Evol9:72–79.
Lorenzen S. 1985. Phylogenetic aspects of pseudocoelomate evo-lution. In: Conway Morris S, George JD, Gibson R, Platt HM,editors. The origins and relationships of lower invertebrates.Oxford: Clarendon Press. p 210–223.
Markevich GI. 1989. Morphology and principal organization ofthe sclerite system of the rotifer mastax. In: Biologiya, Sistema-tika i Funksionalnaya Morfologiya Presnovadnyk Zhivotnyk.Trudy Inst. Biol. Vnutrenney Vod. Acad. Nauk S.S.S.R:27-82(in Russian).
McMenamin MS. 1998. The garden of Ediacara. Discovery of thefirst complex life. New York: Columbia University Press. p 295.
Mock H. 1981. Zur Kenntnis von Diurodrilus subterraneus(Polychaeta, Dinophilidae) aus dem Sandhang der NordseeinselSylt. Helgol Wiss Meeresunters 34:329–335.
Neuhaus B, Kristensen RM, Lemburg C. 1996. Ultrastructure ofthe cuticle of Nemathelminthes and electron microscopical lo-calization of chitin. Verh Dtsch Zool Ges 89:221.
Nielsen C. 1995. Animal evolution: interrelationships of the liv-ing phyla. Oxford: Oxford University Press. p 467.
Nielsen C, Scharff N, Eibye-Jacobsen D. 1996. Cladistic analysesof the animal kingdom. Biol J Linn Soc Lond 57:385–410.
Nogrady T, Wallace RL, Snell TW. 1993. Rotifera. In: DumontHJF, editor. Guides to the identification of the microinverte-brates of the continental waters of the world, vol. 4. Rotifera.The Hague: SPB Academic Publishing. p 142.
Porsild MP. 1920. The flora of Disko Island and the adjacent coastof West Greenland from 66°-71° N Lat. with remarks on phy-togeography, ecology, flowering, fructation and hibernation.Meddr Grønland 58:1–156.
Purschke G. 1987. Anatomy and ultrastructure of ventral pha-ryngeal organs and their phylogenetic importance inPolychaeta (Annelida). IV. The pharynx and jaws of the Dorvil-leidae. Acta Zool (Stockholm) 68:83–105.
Reynolds ES. 1963. The use of lead citrate at high pH as anelectronopaque stain in electron microscopy. J Cell Biol 17:208–212.
Ricci C, Melone G, Sotgia C. 1993. Old and new data on Sei-sonidea (Rotifera). Hydrobiologia 255/256:495–511.
Riedl RJ. 1969. Gnathostomulida from America. Science 163:445–452.
Riedl RJ, Rieger RM. 1972. New characters observed on isolatedjaws and basal plates of the family Gnathostomulidae (Gnatho-stomulida). Z Morph Tiere 72:131–172.
Rieger RM. 1976. Monociliated epidermal cells in Gastrotricha:significance for concepts of early metazoan evolution. Z ZoolSyst Evolut Forsch 14:198–226.
Rieger RM. 1980. A new group of interstitial annelids (Lobato-cerebridae, nov. fam.) and its significance for metazoan phylog-eny. Zoomorphology 95:41–84.
Rieger RM, Mainitz M. 1977. Comparative fine structure study ofthe body wall in Gnathostomulida and their phylogenetic posi-tion between Platyhelminthes and Aschelminthes. Z Zool SystEvolut Forsch 15:9–35.
48 R. M. KRISTENSEN AND P. FUNCH

Rieger RM, Tyler S. 1995. Sister-group relationship of Gnatho-stomulida and Rotifera–Acanthocephala. Invert Biol 114:186-188.
Ruppert EE. 1991a. Introduction to the aschelminth phyla: aconsideration of mesoderm, body cavities and cuticle. In:Harrison FW, Ruppert EE, editors. Microscopic anatomy ofinvertebrates, vol. 4. Aschelminthes. New York: Wiley-Liss. p1–17.
Ruppert EE. 1991b. Gastrotricha. In: Harrison FW, Ruppert EE,editors. Microscopic anatomy of invertebrates, vol. 4. Aschel-minthes. New York: Wiley-Liss. p 41–109.
Ruppert EE, Barnes RD. 1994. Invertebrate zoology, 6th ed.Philadelphia: Saunders College Publishing. p 1056.
Schmidt-Rhaesa A, Bartolomaeus T, Lemburg C, Ehlers U, GareyJR. 1998. The position of the Arthropoda in the phylogeneticsystem. J Morphol 238:263–285.
Segers H, Melone G. 1998. A comparative study of trophi mor-phology in Seisonidea (Rotifera). J Zool Lond 244:201–207.
Semper C. 1872. Zoologische Aphorismen, III. Trochosphaera. ZWiss Zool 22:310–322.
Sterrer W. 1968. Beitrage zur Kenntniss der Gnathostomulida. I.Anatomie und Morphologie des Genus Pterognathia Sterrer.Ark F Zool 22:1–125.
Sterrer W. 1972. Systematics and evolution within the Gnatho-stomulida. Syst Zool 21:151–173.
Sterrer W. 1998. Gnathostomulida from the (sub)tropical north-western Atlantic. In: van der Steen LJ, editor. Studies on thenatural history of the Caribbean region, vol. 74. Amsterdam:Publications Foundation for Scientific Research in the Carib-bean Region: 142. p 1–186.
Sterrer W, Farris RA. 1975. Problognathia minima n. g., n. sp.,representative of a new family of Gnathostomulida, Problo-gnathiidae n. fam. from Bermuda. Trans Am Microsc Soc 94:357–367.
Sterrer W, Mainitz M, Rieger RM. 1985. Gnathostomulida: enig-matic as ever. In: Conway Morris S, George JD, Gibson R, PlattHM, editors. The origins and relationships of lower inverte-brates. Oxford: Clarendon Press. p 181–199.
Storch V, Welsch U. 1969. Uber den Aufbau des Rotatorienin-tegumentes. Z Zellforsch 95:405–414.
Storch V, Higgins RP, Morse MP. 1989. Internal anatomy ofMeiopriapulus fijiensis (Priapulida). Trans Am Microsc Soc 108:245–261.
Swedmark B. 1964. The interstitial fauna of marine sand. BiolRev 39:1–42.
Sørensen MV. 2000. An SEM study of the jaws of Haplognathiarosea and Rastrognathia macrostoma (Gnathostomulida), witha preliminary comparison with the rotiferan trophi. Acta Zool(Stockholm) 81:9–16.
Sørensen MV, Kristensen RM. 2000. Marine Rotifera from IkkaFjord, SW Greenland, with the description of a new speciesfrom the rare mineral ikaite. Meddr Grønland (Bioscience) 51:1–46.
Sørensen MV, Funch P, Hansen AJ, Willerslev E, Olesen J. 2000.On the phylogeny of Metazoa in light of Cycliophora and Mi-crognathozoa. Zool Anz (in press).
Valentine JW. 1997. Cleavage patterns and the topology of themetazoan tree of life. Proc Natl Acad Sci 94:8001–8007.
Villora-Moreno S. 1996. Ecology and distribution of the Diurodri-lidae (Polychaeta), with redescription of Diurodrilus benazzi.Cah Biol Mar 37:99–108.
Wallace RL, Ricci C, Melone G. 1996. A cladistic analysis ofpseudocoelomate (aschelminth) morphology. Invert Biol 115:104–112.
Welch DBM. 2000. Evidence from a protein-coding gene thatacanthocephalans are rotifers. Invert Biol 119:17–26.
Westheide W. 1982. Ikosipodus carolensis gen. et sp. n. an intersti-tial neotenic polychaete from North Carolina, U.S.A., and its phy-logenetic relations within Dorvilleidae. Zool Scr 11:117–126.
Westheide W. 1984. The concept of reproduction in polychaeteswith small body size: adaptation in interstitial species. In:Fischer A, Pfannenstiel H-D, editors. Polychaete Reproduction.Fortschr Zool 29:265–287.
Westheide W. 1987. Progenesis as a principle of meiofauna evo-lution. J Nat Hist 21:843–854.
Westheide W. 1990. Polychaetes: interstitial families. In: Ker-mack DM, Barnes RSK, editors. Synopsis of the British Fauna(new series). Oestgeest: Universal Book Services/Dr. W. Bach-huys. p 152.
Westheide W, Riser NW. 1983. Morphology and phylogeneticrelationships of the neotenic interstitial polychaete Apodotro-cha progenerans n. gen., n. spec. (Annelida). Zoomorphology103:67–87.
Westheide W, von Nordheim H. 1985. Interstitial Dorvilleidae(Annelida, Polychaeta) from Europe, Australia and New Zea-land. Zool Scr 14:183–199.
Winnepenninckx B, Backeljau T, Mackey LY, Brooks JM, DeWachter R, Kumar S, Garey JR. 1995. 18S rRNA data indicatethat Aschelminthes are polyphyletic in origin and consist of atleast three distinct clades. Mol Biol Evol 12:1132–1137.
Winnepenninckx B, Backeljau T, Kristensen RM. 1998. Relationsof the new phylum Cycliophora. Nature 393:636–638.
Wray GA, Levinton JS, Shapiro LH. 1996. Molecular evidence fordeep Precambrian divergences among metazoan phyla. Science274:568–573.
Zrzavy J, Stanislav S, Kepka P, Bezdek A. 1998. Phylogeny of theMetazoa based on morphological and 18S ribosomal DNA evi-dence. Cladistics 14:249–285.
49MICROGNATHOZOA: A NEW CLASS