microbial mineralization of atrazine and 2,4-dichlorophenoxyacetic acid in riparian pasture and...
TRANSCRIPT

Biol Fertil Soils (1994) 18:89-94 © Springer-Verlag 1994
James A. Entry • P.K. Donelly • W.H. Emmingham
Microbial mineralization of atrazine and 2,4-dichlorophenoxyacetic acid in riparian pasture and forest soils
Received: 19 May 1993
Abstract Microbial biomass and mineralization of atra- zine [2-chloro-4(ethylamino)-6(isopropylamino)s-tria- zine] and 2,4-D (2,4-dichlorphenoxyacetic acid) were ex- amined in the top 10 cm of riparian pasture soils and in the litter layer and top 10 cm of mineral soils of riparian forest ecosystems. The riparian forest litter had higher levels of active and total fungal biomass than forest or pasture mineral soils in winter, spring, and fall. Active bacterial biomass was higher in forest litter than in forest and pasture mineral soils in spring and autumn, and high- er in forest mineral soils than in pasture soils in summer. Total bacterial biomass was higher in forest mineral soils than in pasture soils during all seasons. In spring, it was also higher in forest litter than in pasture soils. Atrazie and 2,4-D mineralization in pasture soils was exceeded by that in forest litter in spring and au tumn and by that in forest mineral soils in summer and autumn. There was no correlation between either active or total fungal and bac- terial biomass with pesticide degradation.
Key words Microbial biomass • Atrazine • 2,4D Riparian soils • Iodo-nitrotetrazolium chloride stain • Epifluorescence microscopy
Paper 2837 of the Forest Research Laboratory, Oregon State University
J.A. Entry (~c~) ~ . W.H. Emmingham Department of Forest Science, Oregon State University, Corvallis, OR 97331, USA
R K. Donelly Department of Bacteriology and Biochemistry, University of Idaho, Moscow, ID 83843, USA
Current address: t Department of Agronomy and Soils, 202 Funchess Hall, Auburn University, Auburn, AL 36849-5412, USA
Introduction
The extensive use of atrazine and 2,4-D in agricultural re- gions of the United States and Canada has been well doc- umented (McEwen and Stephenson 1979). The persis- tence of these two herbicides in the environment has gen- erated much concern because of their high mobility, their tendency to accumulate in both terrestrial and marine ecosystems, and their potential effects on human health (Hamilton et al. 199). Agriculture has contributed an in- creasing amount of pollutants to lakes and streams as farm management practices have been intensified (Peter- john and Correll 1984).
Previous studies have shown that certain fungi (Aust 1990) and bacteria (Behki and Khan 1986; Cook and Hurter 1984; Kilpi et al. 1990) can degrade atrazine and 2,4-D to CO2; however, this can be an extremely slow process when environmental conditions are not favorable (Chaudhry and Chapalamagdugu 1991), and pollutants may be deposited into lakes and streams via ground water flow and runoff (Wu et al. 1983; Pickett et al. 1992). Pre- vious studies also have demonstrated that forest riparian filter strips can protect stream waters f rom input of non- point-source nutrients (Omernik et al. 1981; Schtosser and Karr 1981; Lowrance et al. 1984). Lowrance et al. (1984) discovered that riparian forests provide an excel- lent nutrient sink for surrounding agricultural lands.
No studies have investigated the influence of forest filter strips on herbicide degradation. The objective of the present study, therefore, was to determine whether forest riparian buffer strips have higher bacterial and fungal biomass and atrazine and 2,4-D mineralization than do riparian pasture ecosystems.
Materials and methods
Soil sampling procedures
To ensure that results would not be specific to a given site, the sam- pling scheme was arranged in a random block design (Kirk 1982)

90
with three sites as blocks. Three soil types, forest litter, forest miner- al soil, and pasture mineral soil (pasture land had no litter layer), were considered treatments. Soil samples were taken in the Wilamette Valley near Corvallis, Oregon, from three riparian sites along Oak Creek, Jackson Creek, and Soap Creek. Soil types, pre- cipitation, temperature, and vegetation characteristics of those sites are given in Table 1.
Moisture and temperature readings in the different soil types at each site were taken at each sampling (Table 2). On each site, each soil type was randomly sampled three times around each of three sampling centers (2-m diameter) on January 12, April 23, July 29, and November 19, 1991 (n = 324; three sites xthree soil types xthree sampling centers x three samples x four seasons). The sampling pe- riod was chosen to coincide with the peak season of active bacterial and fungal biomass reported previously for a nearby site (Ingham et al. 1991).
The litter layer (depth 0 - 7 cm) and the top 10 cm of each min- eral soil were sampled separately and estimates of microbial bio- mass and atrazine and 2,4-D degradation were made. Soil moisture was determined gravimetrically; soil temperatures were taken at the time of sampling; pH measurements were made on a l-to-1 paste
mixture with a model 901 ion analyzer (Orion Instruments). All soil samples were held in the laboratory at 4 °C under moisture condi- tions the same as those in the field. Samples were prepared for her- bicide mineralization experiments and microbial testing within 24 h so that microbial activity would not be significantly altered (West et al. 1986).
Concentrations of total N were determined with methods de- scribed by Bremner and Mulvaney (1982), extractable concentra- tions of N H [ and NO~- by a microdiffusion method (Keeney and Nelson 1982), mineralizable NH~ with methods of Giest (1977), and extractable P with Bray extraction techniques (Olsen and Som- mers 1982). Total C was estimated by dry-ashing (Nelson and Som- mers 1982). The C : N ratio was calculated by dividing total C by to- tal N in each sample.
Microbial biomass measurements
Total and active bacterial and fungal biomass values were measured with methods described by Ingham and Klein (1984). A suspension
Table 1 Soil, precipitation, temperature, and vegetation characteristics of Jackson Creek, Oak Creek, and Soap Creek riparian sites
Site Soil type a Soil Slope Average Temperature Forest riparian vegetation Pasture riparian pH annual (° C) vegetation
precipitation Overstory Understory
Jackson Vertic Haploxeroll 6.0 5% 100- 150 mm 1 0 - 12 Quercus Rosa woodsii Festuca Creek Fine, montmorillic kelloggii Lindl. arundinacea
mesic Newberry Schreb.
Oak Fluvaquentic 6.3 3 % 100 mm Creek Haplaquoll
Fine, mixed mesic
Soap Cumulic Ultic 5.8 8% 100- 150 mm Creek Haploxeroll
Fine, mixed mesic
1 0 - 1 2
1 0 - 12
Rubus parviflorus Nutt.
Pseudotsuga Polystichum Trifolium menziesii munitum pratense L. (Mirb.) Roth. Franco
Quercus Rosa woodsii kelloggii Lindl. Newberry Rubus
parviflorus Nutt.
Pseudotsuga menziesili (Mirb.) Franco
Symphocarpus albus R.
Rubus parviflorus Nutt.
Polystiehum munitum Roth.
Loloim perenne L.
Festuca arundinacea Schreb.
Trifolium pratense L.
Lolium perenne L.
Festuca arundinacea Schreb.
Trifolium subterraneum L.
Lolium perenne L.
A lopecurus pratensis L.
a Data obtained from Knezevich (1975)
91
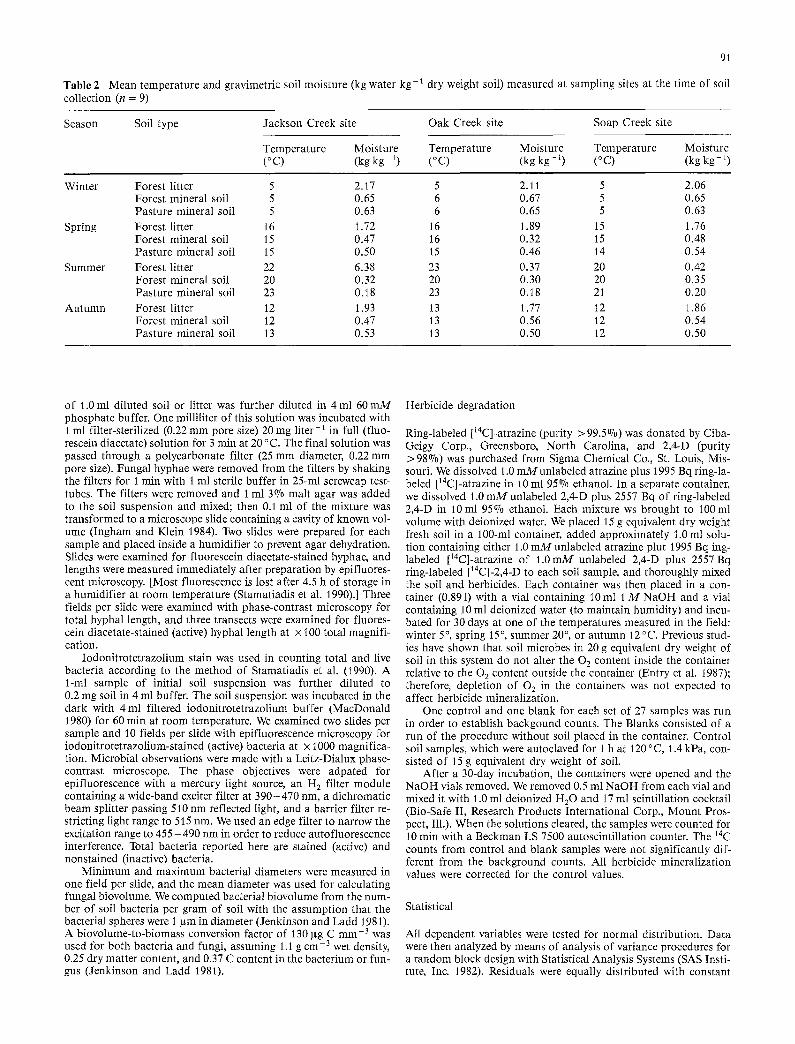
Table 2 Mean temperature and gravimetric soil moisture (kg water kg- ~ dry weight soil) measured at sampling sites at the time of soil collection (n = 9)
Season Soil type Jackson Creek site Oak Creek site Soap Creek site
Temperature Moisture Temperature Moisture Temperature Moisture (°C) (kgkg ~) (°C) (kgkg -1) (°C) (kgkg -1)
Winter Forest litter 5 2.17 5 2.11 5 2.06 Forest mineral soil 5 0.65 6 0.67 5 0.65 Pasture mineral soil 5 0.63 6 0.65 5 0.63
Spring Forest litter 16 1.72 16 1.89 15 1.76 Forest mineral soil 15 0.47 16 0.32 15 0.48 Pasture mineral soil 15 0.50 15 0.46 14 0.54
Summer Forest litter 22 6.38 23 0.37 20 0.42 Forest mineral soil 20 0.32 20 0.30 20 0.35 Pasture mineral soil 23 0.18 23 0.18 21 0.20
Autumn Forest litter 12 1.93 13 1.77 12 1.86 Forest mineral soil 12 0.47 13 0.56 12 0.54 Pasture mineral soil 13 0.53 13 0.50 12 0.50
of 1.0 ml diluted soil or litter was further diluted in 4 ml 60 m M phosphate buffer. One milliliter of this solution was incubated with 1 ml filter-sterilized (0.22 mm pore size) 20 mg liter i in full (fluo- rescein diacetate) solution for 3 min at 20 °C. The final solution was passed through a polycarbonate filter (25 mm diameter, 0.22 mm pore size). Fungal hyphae were removed from the filters by shaking the filters for 1 rain with 1 ml sterile buffer in 25-ml screwcap test- tubes. The filters were removed and 1 ml 3% malt agar was added to the soil suspension and mixed; then 0.1 ml of the mixture was transformed to a microscope slide containing a cavity of known vol- ume (Ingham and Klein 1984). Two slides were prepared for each sample and placed inside a humidifier to prevent agar dehydration. Slides were examined for fluorescein diacetate-stained hyphae, and lengths were measured immediately after preparation by epifluores- cent microscopy. [Most fluorescence is lost after 4.5 h of storage in a humidifier at room temperature (Stamatiadis et al. 1990).] Three fields per slide were examined with phase-contrast microscopy for total hyphal length, and three transects were examined for fluores- cein diacetate-stained (active) hyphal length at × 100 total magnifi- cation.
Iodonitrotetrazolium stain was used in counting total and live bacteria according to the method of Stamatiadis et al. (1990). A 1-ml sample of initial soil suspension was further diluted to 0.2 mg soil in 4 ml buffer. The soil suspension was incubated in the dark with 4 ml filtered iodonitrotetrazolium buffer (MacDonald 1980) for 60 min at room temperature. We examined two slides per sample and 10 fields per slide with epifluorescence microscopy for iodonitrotetrazolium-stained (active) bacteria at x 1000 magnifica- tion. Microbial observations were made with a Leitz-Dialux phase- contrast microscope. The phase objectives were adpated for epifluorescence with a mercury light source, an H z filter module containing a wide-band exciter filter at 390 -470 nm, a dichromatic beam splitter passing 510 nm reflected light, and a barrier filter re- stricting light range to 515 nm. We used an edge filter to narrow the excitation range to 455 - 4 9 0 nm in order to reduce autofluorescence interference. Total bacteria reported here are stained (active) and nonstained (inactive) bacteria.
Minimum and maximum bacterial diameters were measured in one field per slide, and the mean diameter was used for calculating fungal biovolume. We computed bacterial biovolume from the num- ber of soil bacteria per gram of soil with the assumption that the bacterial spheres were 1 ~m in diameter (Jenkinson and Ladd 1981). A biovolume-to-biomass conversion factor of 130 gg C mm -3 was used for both bacteria and fungi, assuming 1.1 g cm 3 wet density, 0.25 dry matter content, and 0.37 C content in the bacterium or fun- gus (Jenkinson and Ladd 1981).
Herbicide degradation
Ring-labeled [~4C]-atrazine (purity > 99.5%) was donated by Ciba- Geigy Corp., Greensboro, North Carolina, and 2,4-D (purity > 9 8 % ) was purchased from Sigma Chemical Co., St. Louis, Mis- souri. We dissolved 1.0 mMunlabe led atrazine plus 1995 Bq ring-la- beled [14C]-atrazine in 10 ml 95% ethanol. In a separate container, we dissolved 1.0 m M unlabeled 2,4-D plus 2557 Bq of ring-labeled 2,4-D in 10 ml 95% ethanol. Each mixture ws brought to 100 ml volume with deionized water. We placed 15 g equivalent dry weight fresh soil in a 100-ml container, added approximately 1.0 ml solu- t ion containing either 1.0 m2V/unlabeled atrazine plut 1995 Bq ing- labeled [14C]-atrazine of 1 .0mM unlabeled 2,4-D plus 2557 Bq ring-labeled [14C]-2,4-D to each soil sample, and thoroughly mixed the soil and herbicides. Each container was then placed in a con- tainer (0.891) with a vial containing 10 ml 1 M NaOH and a vial containing 10 ml deionized water (to maintain humidity) and incu- bated for 30 days at one of the temperatures measured in the field: winter 5 °, spring 15 °, summer 20 °, or autumn 12 °C. Previous stud- ies have shown that soil microbes in 20 g equivalent dry weight of soil in this system do not alter the 02 content inside the container relative to the 02 content outside the container (Entry et al. 1987); therefore, depletion of 02 in the containers was not expected to affect herbicide mineralization.
One control and one blank for each set of 27 samples was run in order to establish backgound counts. The Blanks consisted of a run of the procedure without soil placed in the container. Control soil samples, which were autoclaved for 1 h at 120 °C, 1.4 kPa, con- sisted of 15 g equivalent dry weight of soil.
After a 30-day incubation, the containers were opened and the NaOH vials removed. We removed 0.5 ml NaOH from each vial and mixed it with 1.0 ml deionized HzO and 17 ml scintillation cocktail (Bio-Safe II, Research Products International Corp., Mount Pros- pect, Ill.). When the solutions cleared, the samples were counted for 10 min with a Beckman LS 7500 autoscintillation counter. The 14C counts from control and blank samples were not significantly dif- ferent from the background counts. All herbicide mineralization values were corrected for the control values.
Statistical
All dependent variables were tested for normal distribution. Data were then analyzed by means of analysis of variance procedures for a random block design with Statistical Analysis Systems (SAS Insti- tute, Inc. 1982). Residuals were equally distributed with constant
92
variances. All values reported are sample values minus control val- ues. Differences reported throughout are significant at P_< 0.05, as determined by Fisher's protected least significant difference test. Because analysis of variance for soil chemicals, active and total fungal and bacterial biomasses, and atrazine and 2,4-D mineraliza- tion did not indicate significant differences among sites, only differ- ences among soil types and season are reported (Kirk 1982).
Results
The forest organic layers on these riparian sites had sig- nificantly greater concentrations of all measured chemi- cals and a higher C : N ratio than the mineral soils in both forest and pasture. The pasture sites had significantly lower values than the mineral forest soils for all except NH2 and total N (Table 3).
Microbial biomass
The active and total fungal biomass was significantly greater in forest litter than in the forest and pasture min- eral soils in winter, spring, and autumn samples (Table 4). The active fungal biomass was greater in the litter collect- ed in spring and fall than in that collected in summer and winter.
The active bacterial biomass did not vary among soil types collected during winter, but it was significantly greater in forest litter than in the forest and pasture min- eral soils in samples collected in spring and autumn. The total bacterial biomass was greater in forest litter than in the pasture soils in spring only. In the forest mineral soils, the total bacterial biomass exceeded than in the pasture mineral soils in all seasons.
Soil herbicide degradation
Atrazine mineralization did not differ among soil types collected in winter and incubated at 5 °C, but it was sig- nificantly greater in forest litter than in the forest and pasture mineral soils in spring-collected samples incubat- ed at 15°C (Table 4). When soils were collected during summer and incubated at 20 °C, atrazine mineralization was greater in forest mineral soil than in pasture soil, and it was greater in both forest soils than in pasture soils in autumn samples incubated at 12°C. Significantly more atrazine was mineralized in the summer and autumn sam- ples incubated at 20 ° and 12°C, respectively, than in spring or winter samples incubated at 15 ° and 5 °C.
Mineralization of 2,4-D, like that of atrazine, did not differ among soils collected during winter and incubated at 5 °C. In soils collected in spring and incubated at
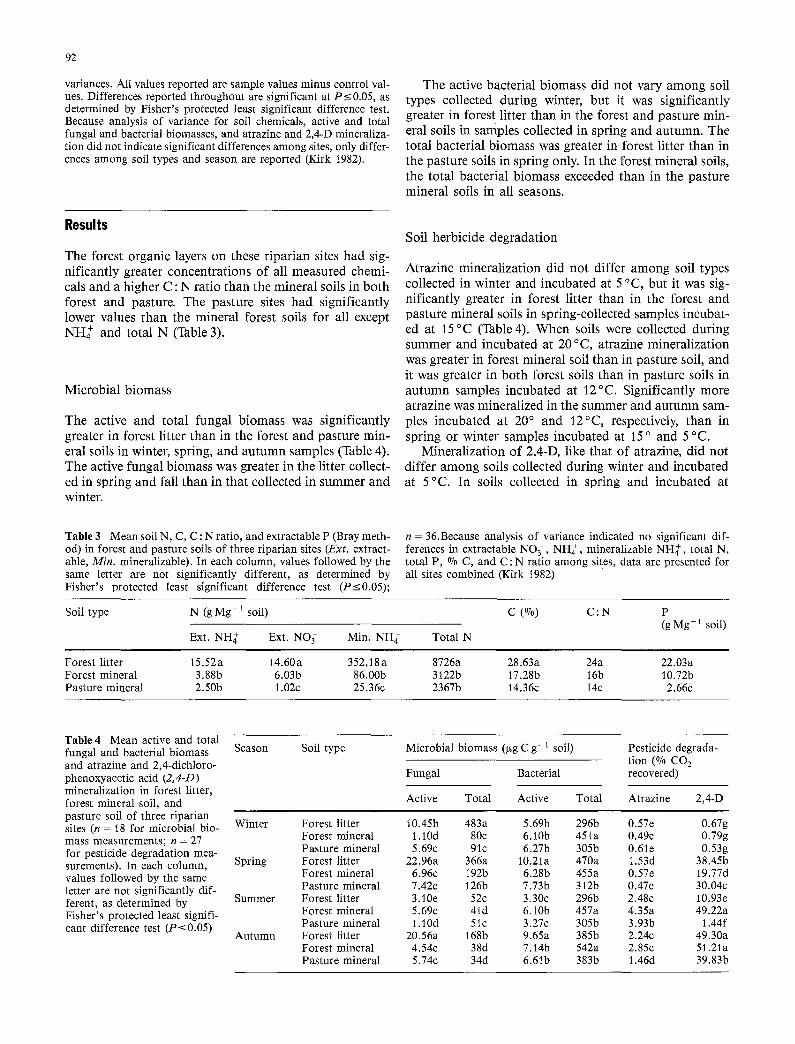
Table 3 Mean soil N, C, C : N ratio, and extractable P (Bray meth- od) in forest and pasture soils of three riparian sites (Ext. extract- able, Min. mineralizable). In each column, values followed by the same letter are not significantly different, as determined by Fisher's protected least significant difference test (P_<0.05);
n = 36.Because analysis of variance indicated no significant dif- ferences in extractable N O r , NH~, mineralizable NH~-, total N, total P, % C, and C : N ratio among sites, data are presented for all sites combined (Kirk t982)
Soil type N (g M g - I soil)
Ext. NH~ Ext. NO 3 Min. NH~ Total N
C (%) C : N P (g M g - ~ soil)
Forest litter 15.52a 14.60a 352,18a Forest mineral 3.88b 6.03b 86.00b Pasture mineral 2.50b 1.02c 25.36c
8726a 28.63a 24a 22.03a 3122b 17.28b 16b 10.72b 2367b 14.36c 14c 2.66c
Table 4 Mean active and total fungal and bacterial biomass Season and atrazine and 2,4-dichloro- phenoxyacetic acid (2,4-£)) mineralization in forest litter, forest mineral soil, and pasture soil of three riparian sites (n = 18 for microbial bio- Winter mass measurements; n = 27 for pesticide degradation mea- surements). In each column, Spring values followed by the same letter are not significantly dif- ferent, as determined by Summer Fisher's protected least signifi- cant difference test (P_<0.05)
Autumn
Soil type Microbial biomass (gg C g- 1 soil)
Fungal Bacterial
Pesticide degrada- tion (% CO 2 recovered)
Active Total Active Total Atrazine 2,4-D
Forest litter Forest mineral Pasture mineral Forest litter Forest mineral Pasture mineral Forest litter Forest mineral Pasture mineral Forest litter Forest mineral Pasture mineral
10.45b 483a 5.69b 296b 0.57e 0.67g 1.10d 80c 6.10b 451a 0.49e 0.79g 5.69c 91c 6.27b 305b 0.61e 0.53g
22.96a 366a 10.21a 470a 1.53d 38.45b 6.96c 192b 6.28b 455a 0.57e 19.77d 7.42c 126b 7.73b 312b 0.47e 30.04c 3.10e 52c 3.30c 296b 2.48c 10.93e 5.69c 41d 6.10b 457a 4.35a 49.22a 1.10d 51c 3.27c 305b 3.93b 1.44f
20.56a 168b 9.65a 385b 2.24c 49.30a 4.54c 38d 7.14b 542a 2.85c 51.21a 5.74c 34d 6.61b 383b 1.46d 39.83b

93
15 °C, it was significantly greater in forest litter than in forest or pasture mineral soils (Table 4). In samples col- lected during summer and incubated at 20 °C, 2,4-D min- eralization was far greater in forest mineral soil than in the other soil types. Mineralization in the two types of forest soil did not differ in soils collected in autumn and incubated at 12 °C, both soils having significantly more 2,4-D degradation than pasture soils. More mineraliza- tion of 2,4-D occurred in soils collected during autumn and incubated at 12 ° than in soils collected during the other three seasons and incubated at 5 °, 15 °, or 20°C.
No significant relationship between fungal or bacterial biomass and herbicide degradation was found.
Discussion
Microbial biomass was present in the winter but enzyme production was apparently not great enough to affect atrazine and 2,4-D mineralization in this study. Soil abiotic variables that affect the microbial community are moisture, temperature, aeration, and pH. Soil tempera- ture and moisture, as further verified by this study, are particularly important regulators of microbial activity (Entry et al. 1986), and therefore, of pesticide degrada- tion (Parker and Doxtader 1983). Previous studies of 2,4-D degradation rates in soils indicate that the optimum temperature is 27 °C and the optimum moisture 0.01 MPa (Parker and Doxtader 1983). It follows that there will be seasonal variation in degradation. In this experiment, pasture soils averaged 45+4% moisture, and the forest mineral soil averaged 34_+3°70 moisture at 0 .01MPa (n = 12); field capacity was 20% moisture at 0.1 MPa for all soils.
The organic horizon layer on the forested sites was 7 + 2 cm thick and was composed of a humus layer that averaged 1.5cm, a fermentation layer that averaged 2.0 cm, and a litter layer 3.5 cm thick. In spring and au- tumn, forest vegetation provides a litter layer that sup- ports a greater amount of active bacterial and fungal bio- mass than found in pasture soil, often resulting in a high rate of atrazine and 2,4-D mineralization. In summer, the organic layer helps protect the forest mineral soil from moisture loss, again resulting in greater active fungal and bacterial biomass and a larger increase in atrazine and 2,4-D degradation in forest mineral soil than in pasture soil. In winter, atrazine and 2,4-D mineralization dramati- cally decreases in all soils as a result of low soil tempera- tures, presumably because the enzymes involved in the degradation of phenolic compounds and halogenated aromatics are inhibited.
Although microbial degradation of herbicides in soil is a function of microoganisms and the activity of their en- zymes (Anderson 1984), we found no correlation between either active and total bacterial biomass or active and to- tal fungal biomass and atrazine and 2,4-D mineralization. Estimates of fungal and bacterial biomass are inherently variable and are only general indices. Also, the activity and production of enzymes in soil microorganisms in-
volved in atrazine and 2,4-D degradation are not yet fully known. Herbicide degradation may be predicted in the fu- ture by improved indices of microbial activity, but other factors will need to be considered. Estimates of key en- zymes involved in atrazine and 2,4-D degradation made with various combinations of soil parameters may pro- vide accurate indicators of degradation rates. The degra- dation pathways and enzymology of atrazine and 2,4-D degradation are well known (Giardina et al. 1982; Chaudhry and Chapalamagdugu 1991).
In the present experiment we measured herbicide min- eralization (i.e., CO2 evolution) and microbial biomass, as opposed to measuring the disappearance of the com- pounds in the soil. Measurements of disappearance ac- count for herbicide taken up by plants or bound to clays or organic compounds over the course of an experiment.
Soil factors such as the quantity and quality of soil organic matter and bioavailability of nutrients, and there- fore base saturation and cation exchange capacity, may affect atrazine and 2,4-D degradation. Morgan et al. (1993) found that degradation of polyaromatic hydrocar- bons by Phanerochaete chrysosporium, Chrysosporium lignorum, and Trametes versicolor was enhanced when C in the form of glucose hay, wood chips, pine bark, and peat were added to the soil. The addition of large amounts of N decreased atrazine and 2,4-D degradation in cultures (Donnelly et al. 1993) and in grassland soils (Entry et al. 1993). Improved knowledge of the influence of forest and pasture vegetation on soils is necessary if we are to optimize the effectiveness of riparian strips for fil- tering herbicide input to lakes and streams.
References
Anderson JPE (1984) Herbicide degradation in soil: Influence of microbial biomass. Soil Biol Biochem 16:483-489
Aust SD (1990) Degradation of environmental pollutants by Phanerochaete chrysosporium. Microb Ecol 20:197 - 209
Behki RM, Khan SU (1986) Degradation of atrazine by Pseudomo- nas: N-dealkylation and dehalogenation of atrazine and its me- tabolites. J Agric Food Chem 34:746-749
Bremner HM, Mulvaney CS (1982) Nitrogen-total. In: Page PA, Miller RH, Keeney DR (eds) Methods of soil analysis. Part 2. Chemical and microbiological properties. Agronomy 9, Am Soc Agron, Madison, Wis, pp 595-622
Chaudhry GR, Chapalamagdugu S (1991) Biodegradation of halogenated organic compounds. Microb Rev 55:59-79
Cook AM, Hutter RJ (i984) Diethylsimazine: Bacterial dechlorina- tion, deamination and complete degradation. J Agric Food Chem 32:581-585
Donnelly PK, Entry JA, Crawford DL (1993) Degradation of atrazine and 2,4-dichlorophenoxyacetic acid by mycorrhizal fungi at three nitrogen concentrations in vitro. Appl Environ Microbiol 59:2642-2647
Enry JA, Stark NM, Loewenstein H (1986) Effect of timber harvest- ing on microbial biomass fluxes in a northern Rocky Mountain forest soil. Can J For Res 16:1076-1081
Entry JA, Stark NM, Loewenstein H (1987) Timber harvesting: Ef- fects on degradation of cellulose and lignin. For Ecol Manage 22:79- 88
Entry JA, Mattson KG, Emmingham WH (1993) The influence of nitrogen on atrazine and 2,4-dichlorophenoxyacetic acid miner- alization in grassland soils. Biol Fertil Soils 16:179-182

94
Giest JM (1977) Nitrogen response relationships of some volanic ash soils. Soil Sci Soc Am J 41:996-1000
Giardina MC, Giardi MT, Filucchioni G (1982) Atrazine metab- olism by Nocurdia. Elucidation of initial pathway and initial synthesis of potential metabolites. Agric Biol Chem 46:1439-1445
Hamilton PB, Lean DRS, Jackson GS, Kaushik NK, Solomon KR (1989) Effect of two applications of atrazine on the water quali- ty of freshwater enclosures. Environ Pollut 60:291-304
Ingham ER, Klein DA (1984) Soil fungi relationships between hy- phal activity and staining with fiuorescein diacetate. Soil Biol Biochem 16:273-278
Ingham ER, Griffiths RP, Cromack K Jr, Entry JA (1991) Compar- ison of direct versus fumigation-flush microbial biomass estimates from ectomycorrhizal mat and non-mat soils. Soil Biol Biochem 23:465-472
Jenkinson DS, Ladd JM (1981) Microbial biomass in soil: Measure- ment and turnover. Soil Biochem 5:415-471
Keeney DR, Nelson DW (1982) Nitrogen-inorganic forms. In: Page PA (eds) Methods of soil analysis. Part 2. Chemical and micro- biological properties. Agronomy 9, Am Soc Agron, Madison, Wis, pp 643-693
Kilpi S, Backstrom V, Korhola M (1980) Degradation of 2-meth- yl-4-chlorophenoxyacetic acid (MCPA), 2,4 dichlorophenoxy- acetic acid (2,4-D), benzoic and salicylic acid by Pseudomonas sp. HV3 FEMS. Microbiol Lett 8:177-182
Kirk RE (1982) Experimental design: Procedures for the behavioral sciences, 2nd edn. Brooks Cole Publishing Co, Belmont, Calif
Knezevich CA (1975) Soil survey of Benton County area, Oregon. USDA Soil Conservation Service, US Government Printing Of- rice, Washington, DC
Lowrance R, Todd R, Fail J Jr, Hendrickson O Jr, Leonard R, Asmussen L (1984) Riparian forests as nutrient filters in agricul- tural watersheds. BioScience 34:374-377
MacDonald RM (1980) Cytochemical demonstration of catabolism in soil-microorganisms. Sol Biol Biochem 12:419-423
McEwen LF, Stephenson GR (1979) The use and significance of pesticides in the environment. John Wiley and Sons, New York, NY
Morgan P, Lee SA, Lewis ST, Sheppard AN, Watkinson RJ (1993) Growth and biodegradation by white rot fungi inoculated into soil. Soil Biol Biochem 25:279-287
Nelson DW, Sommers LE (1982) Total carbon, organic carbon and organic matter. In: Page PA, Miller RH, Keeney DR (eds) Methods of soil analysis. Part 2. Chemical and microbiological properties. Agronomy 9, Am Soc of Agron, Madison, Wis, pp 539-577
Olsen SR, Sommers LE (1982) Phosphorus. In: Page PA, Miller RH, Keeney DR (eds) Methods of soil analysis. Part 2. Chemical and microbiological properties. Agronomy 9, Am Soc Agron, Madison, Wis, pp 403-430
Omernik JM, Abernathy AR, Male LM (1981) Stream nutrient levels and proximity of agricultural and forest land to streams: Some relationships. J Soil Water Conserv 36:227-231
Parker LW, Doxtader KG (1983) Kinetics of the microbial degrada- tion of 2,4-D in soil: Effects of temperature and moisture. J En- viron Qual 12:553-558
Peterjohn WT, Correll DL (1984) Nutrient dynamics in an agricul- tural watershed: Observations of the role of a riparian forest. Ecology 65:1466-1475
Pickett CH, Hawkins LS, Pehrson JE, O'Connell NVO (1992) Irri- gation practices, herbicide use and groundwater contamination in citrus production: A case study in California. Agric Ecoyst Environ 41:1 - 17
SAS Institute, Inc (1982) SAS user's guide to statistics. SAS Insti- tute Inc, Cary, NC
Schlosser I J, Karr JR (1981) Water quality in agricultural water- sheds: Impact of riparian vegetation during base flow. Water Resour Bull 17:233-240
Stamatiadis S, Doran JW, Ingham ER (1990) Use of staining inhibi- tors to separate fungal and bacterial activity in soil. Soil Biol Biochem 22:81-88
West AW, Ross DJ, Cowling JC (1986) Changes in microbial C, N, P and ATP contents, numbers and respiration of storage of soil. Soil Biol Biochem 18:141-148
Wu TL, Correll DL, Remenapp HEH (1983) Herbicide runoff from experimental watersheds. J Environ Qual 12:330-336