methylseleninic acid super-activates p53-senescence...
TRANSCRIPT

1
Methylseleninic acid super-activates p53-senescence cancer progression
barrier in prostate lesions of Pten-knockout mouse
Lei Wang1, Xiaolan Guo1, Ji Wang1, Cheng Jiang2, Maarten C. Bosland3, Junxuan Lü2* and
Yibin Deng1*
1Hormel Institute, University of Minnesota, 801 16th Ave NE, Austin, MN 55912
2Department of Biomedical Sciences, Texas Tech University Health Sciences Center School of
Pharmacy, 1300 S. Coulter, Amarillo, TX 79106
3Department of Pathology, University of Illinois at Chicago (UIC), College of Medicine,
Chicago, IL 60612
*Corresponding authors: Dr. Yibin Deng, University of Minnesota Hormel Institute, 801 16th
Ave NE, Austin, MN 55912. E-mail: [email protected]; Dr. Junxuan Lü, Department of
Biomedical Sciences, Texas Tech University Health Sciences Center School of Pharmacy, 1300
S. Coulter St, Amarillo, TX 79106. E-mail: [email protected];
Running title: Next-gen selenium strengthens a cancer barrier
Key words: senescence, monomethylated selenium, cancer chemoprevention
Word count (2394). Figures (5) and Supplementary Figure (1). Abstract word count (248)
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236

2
Funding supports: Grants R21 CA155522 (Y. Deng, J. Lü) and R01 CA172169 (J. Lü, Y.
Deng, M. Bosland) from the National Cancer Institute, National Institutes of Health.
Disclosure of Potential Conflicts of Interest: All authors have no personal or financial
conflict of interest and have not entered into any agreement that could interfere with our access
to the data on the research or on our ability to analyze the data independently, to prepare articles,
and to publish them.
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236

3
ABSTRACT
Monomethylated selenium (MM-Se) forms that are precursors of methylselenol such as
methylseleninic acid (MSeA) differ in metabolism and anti-cancer activities in preclinical cell
and animal models from seleno-methionine that had failed to exert preventive efficacy against
prostate cancer (PCa) in North American men. Given that human PCa arises from precancerous
lesions such as high-grade prostatic intraepithelial neoplasia (HG-PIN) which frequently have
lost PTEN tumor suppressor permitting AKT oncogenic signaling, we tested the efficacy of
MSeA to inhibit HG-PIN progression in Pten prostate specific knockout (KO) mice and assessed
the mechanistic involvement of p53-mediated cellular senescence and of the androgen receptor
(AR). We observed that short-term (4 weeks) oral MSeA treatment significantly increased
expression of P53 and P21Cip1 proteins and senescence-associated-β-galactosidase staining, and
reduced Ki-67 cell proliferation index in Pten KO prostate epithelium. Long-term (25 weeks)
MSeA administration significantly suppressed HG-PIN phenotype, tumor weight, and prevented
emergence of invasive carcinoma in Pten KO mice. Mechanistically, the long-term MSeA
treatment not only sustained P53-mediated senescence, but also markedly reduced AKT
phosphorylation and AR abundance in the Pten KO prostate. Importantly, these cellular and
molecular changes were not observed in the prostate of wild type littermates which were
similarly treated with MSeA. Since p53 signaling is likely to be intact in HG-PIN compared to
advanced PCa, the selective super-activation of p53-mediated senescence by MSeA suggests a
new paradigm of cancer chemoprevention by strengthening a cancer progression barrier through
induction of irreversible senescence with additional suppression of AR and AKT oncogenic
signaling.
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236

4
INTRODUCTION
Selenium (Se) compounds have been studied for their chemopreventive potential in
various animal models of carcinogenesis, notably mammary, colon, lung, and prostate cancer.
Selenized yeast (Se-yeast) and its principal Se form Se-methionine (SeMet) have been tested in
several human trials in North America for the prevention of prostate cancer (PCa) (1-4). The
outcomes of the Selenium and vitamin E Cancer Trial (SELECT) and other trials failed to
demonstrate a preventive efficacy of these Se forms in the Se-adequate North American cohorts
(2-4). Many, including us, have opined on the possible reasons for such failures (5-7) (8). One
key factor was the selection of ineffective Se agents. In fact, the scarce animal efficacy data that
existed prior to the initiation of these trials did not support PCa preventive efficacy of SeMet and
these negative data were not published in full-length until after SELECT was terminated (9, 10).
Preclinical and mechanistic research has demonstrated that SeMet has little in common
with the mono-methylated methylselenol precursor Se forms (MM-Se), such as methylseleninic
acid (MSeA), in terms of metabolism and anti-cancer activities (8, 11). We have posited that the
failure of SeMet should not be taken to indicate that other Se forms are ineffective for PCa
chemoprevention (12). Indeed, we have shown that daily orally-administered MSeA inhibited
the growth of DU145 and PC-3 human PCa xenografts in athymic nude mice whereas an equal
Se dose of SeMet was inactive, in spite of SeMet leading to much higher retention of Se in the
xenograft tumors (13). We also reported the efficacy of MSeA to inhibit prostate carcinogenesis
in the transgenic adenocarcinoma mouse prostate (TRAMP) model which improved survival
with no observable long-term adverse effect (12). More efficacy and biomarker assessments in
clinically-relevant prostate carcinogenesis models with MM-Se will be essential to evaluate their
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236

5
PCa chemoprevention potential in the post-SELECT era to support future translation of these
data to humans.
The PTEN (phosphatase and tensin homolog) protein, antagonizes the
phosphatidylinositol-3-OH kinase (PI3K)-protein kinase B (AKT) signaling pathway that
stimulates cancer cell metabolism, proliferation and survival (14). Human PTEN loss has been
identified in 45% of high-grade prostatic intraepithelial neoplasia (HG-PIN) and 70% of
advanced PCa [(14) and reference therein]. Mouse genetic studies have demonstrated that loss
of Pten in prostate epithelium rapidly causes HG-PIN that ultimately progresses to invasive
adenocarcinoma and metastatic disease (15). As men diagnosed with HG-PIN are at increased
risk of developing PCa, this prostate specific conditional Pten KO mouse model recapitulates
essential characteristics of human prostate carcinogenesis and is considered clinically relevant
for studies of PCa chemoprevention. In the Pten KO model, the sustained activation of AKT not
only initiates and perpetuates oncogenic signaling and progression pathways, but at the same
time induces cellular senescence (known as Pten-deficiency Induced Cellular Senescence PICS),
which acts as a formidable barrier to restrain oncogenic progression to invasive and metastatic
disease (16, 17). Mechanistic studies suggest that PICS primarily depends on P53 protein
overexpression, which is induced through AKT/mTOR-mediated protein synthesis and p19ARF
sequestration of MDM-2 resulting in inhibition of proteasome-mediated P53 degradation (16).
The critical role of androgen receptor (AR) signaling in PCa, even at the advanced
metastatic castration-resistant stage, is well established and therapeutically exploited (18).
Unfortunately, recent studies have shown that inhibition of AR signaling by castration or
antagonist drugs inadvertently promotes the progression of stable HG-PIN to invasive
carcinomas in Pten KO model (19), raising concerns for utilization of these androgen
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236

6
deprivation strategies for chemoprevention in high risk men and PCa patients with PTEN
deficiency or mutations.
In cell culture studies, we and others have shown MSeA suppression of AR abundance
and signaling in PCa cells (20, 21), and the phosphorylative activation of AKT Ser473 (pAKT)
(22, 23). MSeA was recently shown to induce cellular senescence in human primary lung
fibroblasts and this cellular effect was likely mediated by ATM/P53 signaling (24)(25).
Therefore in the current study we tested the hypothesis that MSeA would inhibit Pten-deficient
carcinogenesis and PCa progression and assessed the mechanistic involvement of p53, PICS,
AKT, and AR in conferring the hypothesized in vivo efficacy.
MATERIALS AND METHODS
Generation of Pten KO mice
The conditional Ptenflox/flox mouse was generated as previously described (26). PB-Cre4
transgenic mice were obtained from the NCI Mouse Repository. Female mice carrying Ptenflox/+
were crossed with male mice harboring PB-Cre4+Ptenflox/+ to generate mutant mice with prostate
epithelium-specific deletion of Pten. Tail DNA was used for PCR-based genotyping as described
(26). All animal protocols were approved by the University of Minnesota Institutional Animal
Care and Use Committee.
Intervention experiments with MSeA
In the short-term experiment, 12-weeks old Pten KO mice (Cre+;PtenFlox/Flox, hereafter
indicated as Pten∆/∆mice) were randomly assigned, 5 mice per group, to receive water or MSeA
for 4 weeks by daily (5 days per week) oral application at the base of the tongue as before (12,
13). Wild type (WT) littermates (Cre-;PtenFlox/Flox; hereafter as Pten+/+; n=3) were treated
identically with water or MSeA to provide comparison control tissues. AIN93G semi-purified
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236

7
diet and water were provided ad libitum. Mouse body weight was monitored weekly. At
necropsy, total prostate was dissected, photographed and weighed. One portion of prostate
tissues from each mouse was fixed in formalin for H&E and immunohistochemistry staining.
The reminder was stored frozen at -80oC for senescence-associated β-galactosidase (SA-gal)
staining and Western blot analyses.
The long-term experiment was carried out with same design, except that the mice were
10-weeks old at the start of the MSeA and water treatments (5 days per week), lasting for 25
weeks (8 mice per group). At necropsy, the genitourinary (GU) tract was collected and weighed.
Then the different prostate lobes were dissected and weighed. The prostate lobes were saved and
processed individually for histopathology and biochemical analyses.
Histopathology analysis
Tissue processing and staining were as performed previously (12, 26). H&E stained lesions
were verified by a pathologist (Maarten Bosland). The pathological changes of all lobes of
prostate were classified according to Shapell et al. (27).
Senescence-associated β-galactosidase staining
Fresh prostate tissues were embedded with OCT and cut into 4 μm sections. The sections were
stained for SA-gal and counterstained with eosin, as described previously (26). The integrated
optical density (IOD) in the prostate epithelium/lesions was quantified by ImagePro-Plus 6.3
software.
Immunohistochemistry (IHC) and immunoblot analysis
IHC and immunoblot (Western) were performed as previously described (26). Briefly, Ki67
(NeoMarker), AR (Millipore), cleaved caspase 3, and p-AKT Ser473 (Cell Signaling, Beverly,
MA) were diluted at 1:100 for IHC. Images were captured and analyzed by ImagePro-Plus 6.3
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236

8
software for integrated optical density (IOD) semi-quantitation. For immunoblot, the prostate
tissues were homogenized in non-denaturing lysis buffer and subjected to SDS-PAGE and
blotted with antibodies against P53, P21Cip1, p-AKT Ser473, AR, and tubulin. Pooling of
prostate tissues from the short-term experiment was necessary due to limited amount of material
available.
Statistical analyses
For parametric data, the mean and SEM were calculated for each experimental group.
Differences among groups were analyzed by ANOVA for more than 2 groups. For comparison
of only 2 groups, student t-test was used. Significant differences were accepted at P<0.05.
RESULTS
Short-term MSeA treatment of Pten KO mice led to super-activation of p53-p21 and
cellular senescence in prostate epithelium
In the first experiment, we evaluated the effect of 4-week MSeA treatment to identify
early biochemical and cellular changes that might correlate and predict its long-term preventive
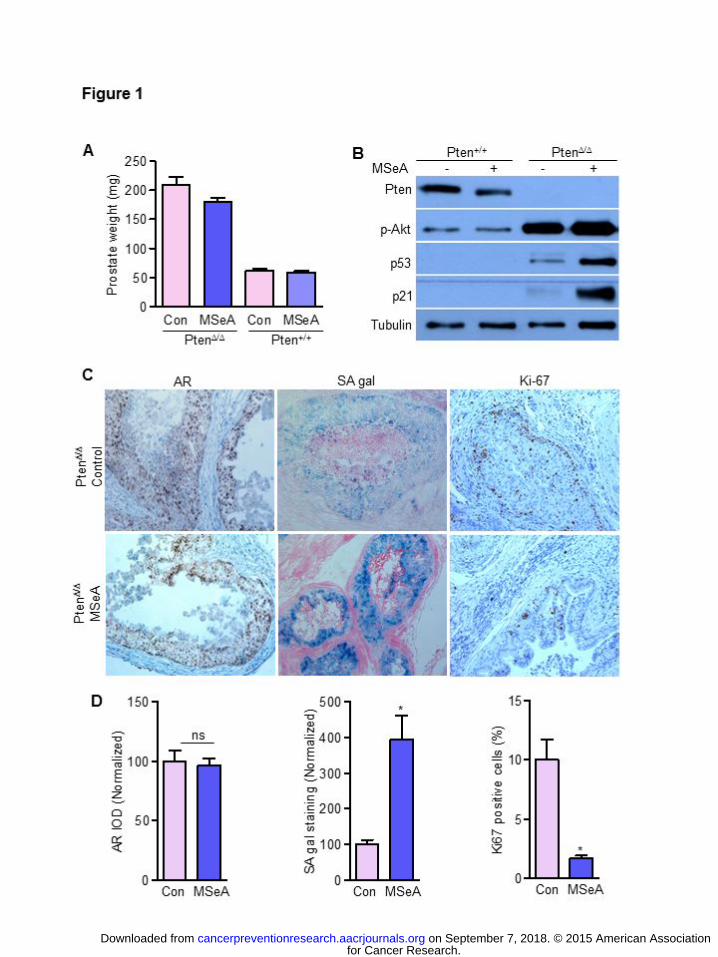
efficacy against HG-PIN growth and tumor progression in Pten KO mice. As shown in Fig. 1A,
MSeA treatment led to a reduction (~20%) of prostate weight in Pten KO mice compared to the
prostate weight in wild-type mice, which was not affected by MSeA. The protein levels of Pten
and phospho-Akt Ser473 (p-Akt) were analyzed in pooled prostate samples of WT mice and Pten
KO mice. As expected, Pten KO mice lacked Pten protein in the prostate and had greatly
increased p-Akt expression (Fig. 1B, lanes 3, 4 vs. 1, 2). Consistent with previous results (16, 17,
26), Pten KO mouse prostate showed increased basal expression of P53 and P21Cip1 over the
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236

9
WT counterpart (Fig. 1B, lane 3 vs. 1). MSeA treatment of the Pten KO mice dramatically
increased P53 and P21Cip1, but this did not occur in WT mice, while p-Akt expression was not
affected in MSeA-treated KO or WT mice (Fig. 1B). In addition, there was no observable change
in AR expression determined by IHC in Pten KO mice after 4 weeks of MSeA treatment (Fig.
1C, 1D).
Since increased P53 protein abundance causes PICS in the Pten KO mice (16, 17), we
examined SA-gal expression in situ in frozen prostate sections (Fig. 1C). Pten KO prostate
showed low but detectable SA-gal staining (Fig. 1C). However, in the prostate of MSeA-treated
Pten KO mice, the staining intensity was remarkably elevated in the epithelial cells by as much
as 4 fold, estimated by ImagePro-Plus software (Fig. 1C, 1D). Because senescence is an
irreversible terminal proliferative arrest, we examined Ki67 as a proliferation indicator and
detected significant suppression of Ki67 labeling index (%) in prostate of the MSeA-treated Pten
KO mice compared to water-treated mice (Fig. 1C, 1D).
Prolonged MSeA treatment of Pten KO mice prevented prostate adenocarcinoma
The promising biochemical and cellular responses to the short-term MSeA intervention
prompted us to evaluate its chemopreventive efficacy on Pten KO HG-PIN growth and
progression in the second experiment with 25-week administration. Consistent with long-term
safety of MSeA supplement in our previous study with the TRAMP model (12), no significant
effect of MSeA was observed on the body weight gain of the mice of each genotype (Supplement
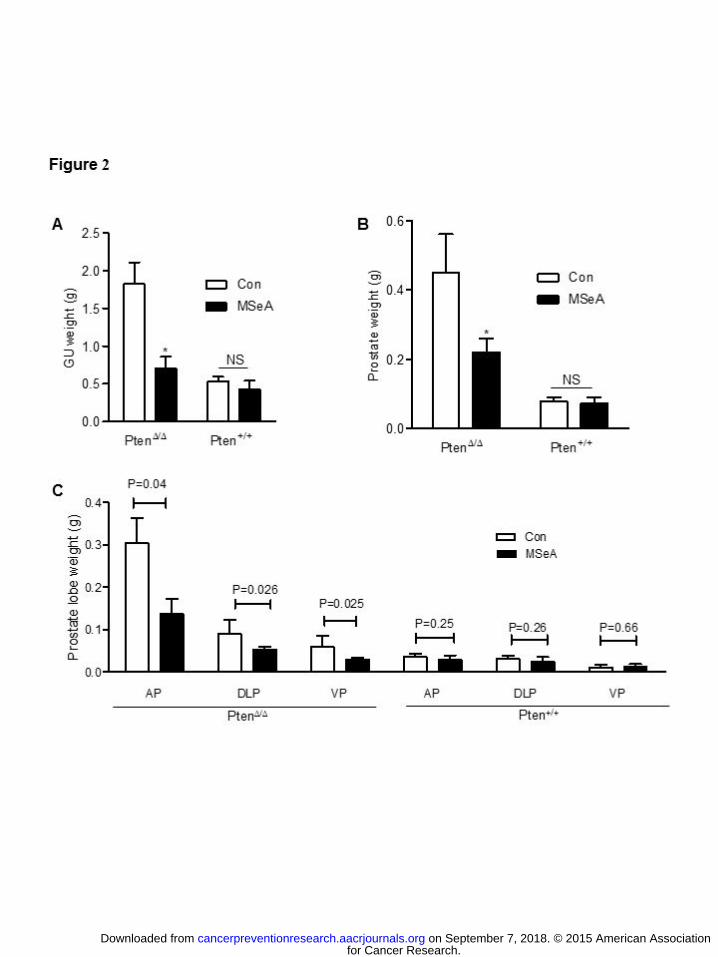
Fig. S1A). As shown in Fig. 2A, long-term MSeA daily treatment did not significantly affect the
genito-urinary tract (GU) weight of the WT mice, but decreased Pten KO-driven expansion of
GU over the WT baseline by more than 70% (Fig. 2A). Likewise, the prostate weight was not
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236

10
affected in the WT mice by MSeA, but was decreased by more than 50% in the Pten KO mice
over the WT baseline (Fig. 2B). At the gross anatomical level, blood-rich prostate tumors were
visible in some water-treated control Pten KO mice (Supplement Fig. S1B). Among the different
lobes, the AP exhibited the most Pten KO-driven growth (Fig. 2C; Supplement Fig. S1C),
whereas in TRAMP mice the DLP undergo the most growth (12). The prostate weight
suppressing effect of MSeA in the Pten KO mice was uniform across AP, DLP and VP lobes
(Fig. 2C; Supplement Fig. S1C). In sharp contrast, the weight of prostate lobes in WT mice of
the MSeA and control groups was not different (Fig. 2C).
Histologically, the prostate lesions from the control Pten KO mice showed HG-PIN
phenotypes in all three lobes (Fig. 3A). Notably 38% (3 out of 8 mice) of Pten KO mice
progressed from HG-PIN to invasive adenocarcinomas at termination of the experiment at 35
weeks of age (Supplement Fig. S1D). In contrast, MSeA-treated mice showed dramatic
histopathological modification, many approaching near normal appearance of the prostate of the
WT mice and none of them with detectable invasive adenocarcinoma features (Fig. 3A, 3C).
Consistent with selectivity of targeting oncogenic growth, MSeA treatment of WT mice did not
affect their typical normal glandular structures (Fig. 3B, 3C). These findings indicate that long-
term MSeA treatment significantly inhibited HG-PIN growth and progression to carcinoma in
vivo.
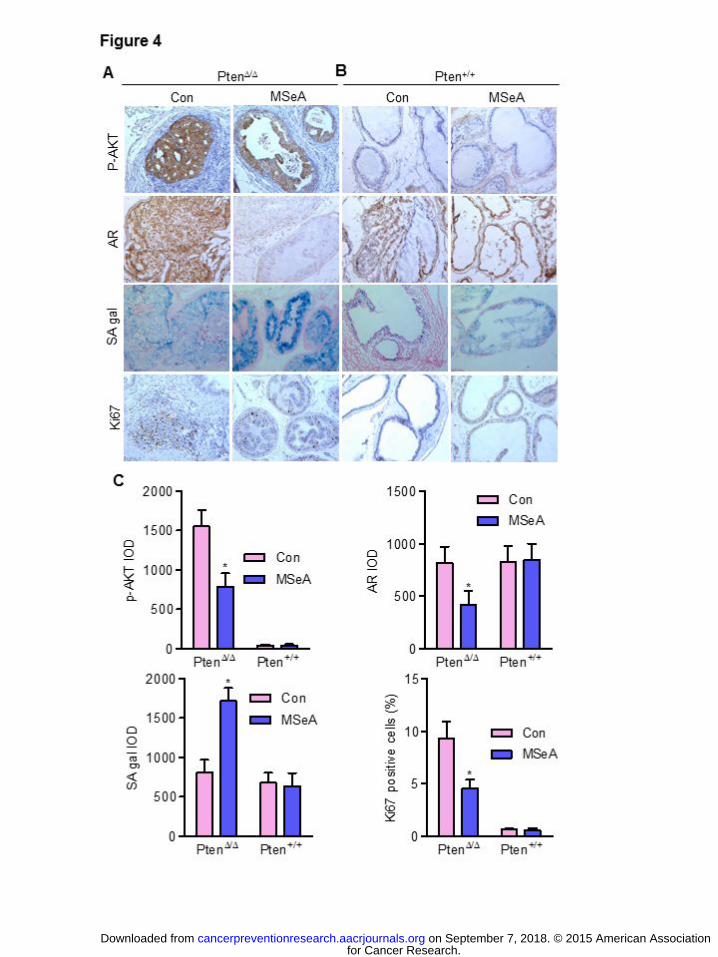
Long-term MSeA treatment decreased p-Akt and AR abundance in Pten KO prostate
epithelium
Since short-term MSeA super-activated P53/P21Cip1 and increased senescence in the Pten-
KO epithelium in vivo, we examined whether long-term MSeA was able to sustain the cellular
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236

11
senescence phenotype. SA-gal staining of AP lobes showed intense senescence in the MSeA-
treated Pten KO mice (Fig. 4A) with little effect in the WT mice (Fig. 4B, 4C). Ki67 staining
confirmed the paucity of proliferating cells in the MSeA-treated Pten KO prostate epithelium
(Fig. 4A, 4C). No appreciable apoptosis, indicated by cleaved caspase-3, was induced by MSeA
treatment in Pten KO mice (Data not shown).
It is noteworthy that IHC staining intensity of p-Akt and AR proteins in the AP lobe of
MSeA-treated Pten KO mice was noticeably decreased (Fig. 4A, 4C) without corresponding
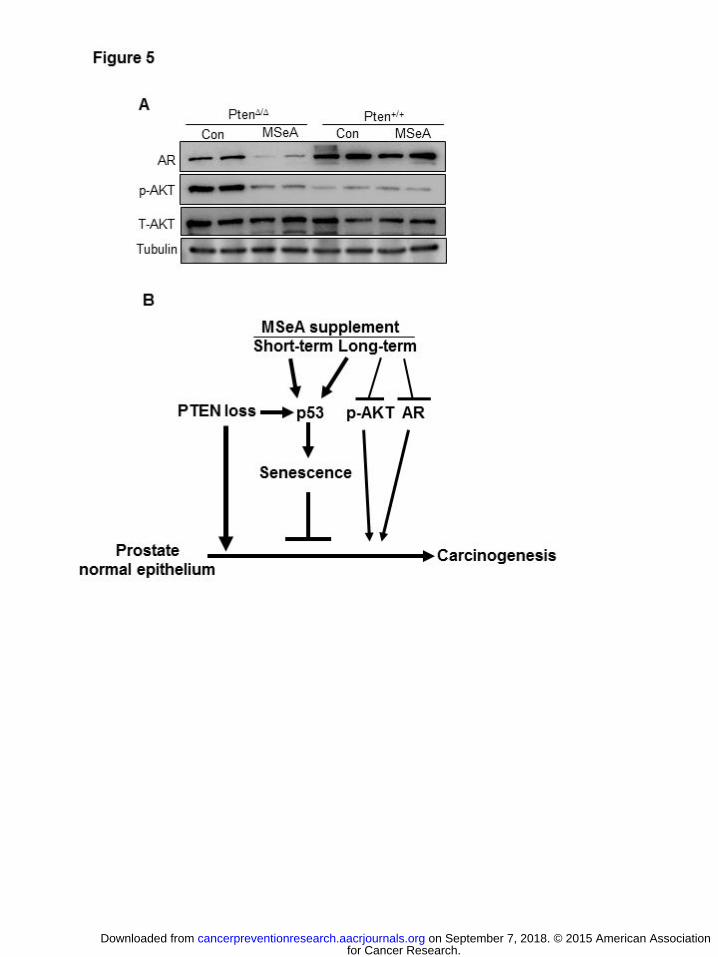
observable changes in the WT mice (Fig. 4B, 4C). Immunoblot confirmed the IHC results for p-
AKT and AR protein abundance suppression by MSeA in the Pten-KO prostate (Fig. 5A).
DISCUSSION
To our best knowledge, this study is the first in which any form of Se has been tested in the
Pten KO PCa mouse model for chemopreventive efficacy. It is also the first time that in vivo
senescence was measurably increased by MSeA treatment selectively in the HG-PIN epithelium
of Pten KO mice without any detectable impact on the prostate of the WT mice. The observed
concomitant increase of P53 and P21Cip1 in the prostate of MSeA-treated Pten KO mice was
not evident in the prostate of WT mice (Fig. 1 B), consistent with the selective super-activation
of this crucial senescence signaling axis. In addition to boosting and sustaining P53-P21Cip1
senescence as a cell proliferation barrier, long-term treatment with MSeA led to considerably
reduced tumor burden (Fig. 2) with decreased AR abundance and phosphorylation of Akt (Fig.
4,5), together contributing to effective suppression of the progression of HG-PIN to carcinoma
(shown schematically in Fig. 5B). Given that current Akt/mTOR inhibitor drugs activate AR
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236

12
signaling while androgen ablation and AR antagonist drugs cross-induce Akt pathway through a
reciprocal feedback regulatory loop (28), the inhibition of both Akt and AR signaling pathways
by prolonged MSeA supplement suggests that combined use of MSeA with these drugs may
mitigate their undesirable side effects and result in greater PCa risk reduction.
Human PCa arises from precancerous HG-PIN lesions with a prolonged clinical course,
affording unique windows of opportunity for chemoprevention/intervention. Since p53 signaling
is more likely to be intact in precancerous lesions than advanced PCa, the super-activation of
p53-senescence by MSeA offers a new paradigm for PCa chemoprevention through
strengthening a cancer progression barrier in the precursor lesions. Our data support that MSeA
super-activated and sustained P53-mediated cellular senescence and subsequently inhibited both
Akt and AR signaling pathways to suppress Pten-deficient HG-PIN progression to
adenocarcinoma (Fig. 5B). The in vivo mechanisms mediating these cellular and molecular
actions of MSeA are currently be elucidated. The efficacy for chemoprevention of Pten-
deficient HG-PIN progression by MSeA documented in the current work and the previously
demonstrated efficacy and safety of MSeA in other prostate cancer mouse models (12) (13)
provide strong justification for further development of MM-Se toward human translational
studies.
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236

13
REFERENCES
1. Clark LC, Combs GF, Jr., Turnbull BW, Slate EH, Chalker DK, Chow J, et al. Effects of
selenium supplementation for cancer prevention in patients with carcinoma of the skin. A
randomized controlled trial. Nutritional Prevention of Cancer Study Group. JAMA : the journal
of the American Medical Association. 1996;276:1957-63.
2. Algotar AM, Stratton MS, Ahmann FR, Ranger-Moore J, Nagle RB, Thompson PA, et al.
Phase 3 clinical trial investigating the effect of selenium supplementation in men at high-risk for
prostate cancer. Prostate. 2013;73:328-35.
3. Lippman SM, Klein EA, Goodman PJ, Lucia MS, Thompson IM, Ford LG, et al. Effect
of selenium and vitamin E on risk of prostate cancer and other cancers: the Selenium and
Vitamin E Cancer Prevention Trial (SELECT). JAMA : the journal of the American Medical
Association. 2009;301:39-51.
4. Marshall JR, Tangen CM, Sakr WA, Wood DP, Jr., Berry DL, Klein EA, et al. Phase III
Trial of Selenium to Prevent Prostate Cancer in Men with High-grade Prostatic Intraepithelial
Neoplasia: SWOG S9917. Cancer Prev Res (Phila). 2011;4:1761-9.
5. El-Bayoumy K. The negative results of the SELECT study do not necessarily discredit
the selenium-cancer prevention hypothesis. Nutr Cancer. 2009;61:285-6.
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236

14
6. Hatfield DL, Gladyshev VN. The Outcome of Selenium and Vitamin E Cancer
Prevention Trial (SELECT) reveals the need for better understanding of selenium biology.
Molecular interventions. 2009;9:18-21.
7. Christensen MJ. Selenium and prostate cancer prevention: what next-if anything? Cancer
Prev Res (Phila). 2014;7:781-5.
8. Lu J, Jiang C, Zhang J. Cancer Prevention with Selenium: costly lessons and difficult but
bright future prospects. In: Kong A-NT, editor. Inflammation, Oxidative Stress and Cancer: CRC
Press Taylor Francis; 2014. p. 477-94.
9. Ozten N, Horton L, Lasano S, Bosland MC. Selenomethionine and alpha-tocopherol do
not inhibit prostate carcinogenesis in the testosterone plus estradiol-treated NBL rat model.
Cancer Prev Res (Phila). 2010;3:371-80.
10. McCormick DL, Rao KV, Johnson WD, Bosland MC, Lubet RA, Steele VE. Null
activity of selenium and vitamin e as cancer chemopreventive agents in the rat prostate. Cancer
Prev Res (Phila). 2010;3:381-92.
11. Lu J, Jiang C. Selenium and cancer chemoprevention: hypotheses integrating the actions
of selenoproteins and selenium metabolites in epithelial and non-epithelial target cells. Antioxid
Redox Signal. 2005;7:1715-27.
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236

15
12. Wang L, Bonorden MJ, Li GX, Lee HJ, Hu H, Zhang Y, et al. Methyl-selenium
compounds inhibit prostate carcinogenesis in the transgenic adenocarcinoma of mouse prostate
model with survival benefit. Cancer Prev Res (Phila). 2009;2:484-95.
13. Li GX, Lee HJ, Wang Z, Hu H, Liao JD, Watts JC, et al. Superior in vivo inhibitory
efficacy of methylseleninic acid against human prostate cancer over selenomethionine or selenite.
Carcinogenesis. 2008;29:1005-12.
14. Song MS, Salmena L, Pandolfi PP. The functions and regulation of the PTEN tumour
suppressor. Nature reviews Molecular cell biology. 2012;13:283-96.
15. Wang S, Gao J, Lei Q, Rozengurt N, Pritchard C, Jiao J, et al. Prostate-specific deletion
of the murine Pten tumor suppressor gene leads to metastatic prostate cancer. Cancer Cell.
2003;4:209-21.
6. Alimonti A, Nardella C, Chen Z, Clohessy JG, Carracedo A, Trotman LC, et al. A novel
type of cellular senescence that can be enhanced in mouse models and human tumor xenografts
to suppress prostate tumorigenesis. The Journal of clinical investigation. 2010;120:681-93.
17. Chen Z, Trotman LC, Shaffer D, Lin HK, Dotan ZA, Niki M, et al. Crucial role of p53-
dependent cellular senescence in suppression of Pten-deficient tumorigenesis. Nature.
2005;436:725-30.
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236

16
18. Mills IG. Maintaining and reprogramming genomic androgen receptor activity in prostate
cancer. Nature reviews Cancer. 2014;14:187-98.
19. Jia S, Gao X, Lee SH, Maira SM, Wu X, Stack EC, et al. Opposing effects of androgen
deprivation and targeted therapy on prostate cancer prevention. Cancer discovery. 2013;3:44-51.
20. Cho SD, Jiang C, Malewicz B, Dong Y, Young CY, Kang KS, et al. Methyl selenium
metabolites decrease prostate-specific antigen expression by inducing protein degradation and
suppressing androgen-stimulated transcription. Mol Cancer Ther. 2004;3:605-11.
21. Dong Y, Lee SO, Zhang H, Marshall J, Gao AC, Ip C. Prostate specific antigen
expression is down-regulated by selenium through disruption of androgen receptor signaling.
Cancer Res. 2004;64:19-22.
22. Jiang C, Wang Z, Ganther H, Lu J. Distinct effects of methylseleninic acid versus selenite
on apoptosis, cell cycle, and protein kinase pathways in DU145 human prostate cancer cells. Mol
Cancer Ther. 2002;1:1059-66.
23. Wang Z, Jiang C, Ganther H, Lu J. Antimitogenic and proapoptotic activities of
methylseleninic acid in vascular endothelial cells and associated effects on PI3K-AKT, ERK,
JNK and p38 MAPK signaling. Cancer Res. 2001;61:7171-8.
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236

17
24. Wu M, Wu RT, Wang TT, Cheng WH. Role for p53 in selenium-induced senescence.
Journal of agricultural and food chemistry. 2011;59:11882-7.
25. Wu M, Kang MM, Schoene NW, Cheng WH. Selenium compounds activate early
barriers of tumorigenesis. J Biol Chem. 2010;285:12055-62.
26. Wang L, Xiong H, Wu F, Zhang Y, Wang J, Zhao L, et al. Hexokinase 2-mediated
Warburg effect is required for PTEN- and p53-deficiency-driven prostate cancer growth. Cell
reports. 2014;8:1461-74.
27. Shappell SB, Thomas GV, Roberts RL, Herbert R, Ittmann MM, Rubin MA, et al.
Prostate pathology of genetically engineered mice: definitions and classification. The consensus
report from the Bar Harbor meeting of the Mouse Models of Human Cancer Consortium Prostate
Pathology Committee. Cancer Res. 2004;64:2270-305.
28. Carver BS, Chapinski C, Wongvipat J, Hieronymus H, Chen Y, Chandarlapaty S, et al.
Reciprocal feedback regulation of PI3K and androgen receptor signaling in PTEN-deficient
prostate cancer. Cancer Cell. 2011;19:575-86.
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236

18
Figure Legends
Figure 1. Oral supplement of MSeA (3 mg Se/kg body weight, 5 days per week) for 4 weeks
induced p53-p21-mediated senescence and inhibited prostate epithelial cell proliferation in
vivo. MSeA or water treatment commenced at 12 weeks of age. (A). Prostate weight of
Pten KO (Pten∆/∆) mice (n=5) and wild type littermates (Pten+/+) (n=3). (B) Immunoblot
analyses of Pten, p-Akt Ser473, P53 and P21Cip1 proteins in pooled samples of each
group. Tubulin served as loading control. (C) Representative photomicrographs of
immunohistochemical staining of androgen receptor (AR), Ki67 and SA-galactosidase
staining of Pten KO prostate from water-treated control mice and MSeA-treated
mice. Magnification 100x. (D) Quantitation results from individual mice for each group
are presented as mean + SEM, n=5 for Pten∆/∆ mice and n=3 for Pten+/+ mice. For AR and
SA-gal staining, the integrated optical density (IOD) was normalized to control mice as
100. For Ki-67, percentage of positive nuclei in the prostate epithelium was quantified.
Figure 2. Effect of oral supplement of MSeA (3 mg Se/kg body weight, 5 days per week) for
25 weeks on Pten-KO-driven growths of genitourinary organs and prostate tumors. MSeA
or water treatment commenced at 10 weeks of age. (A) Genitourinary (GU) tract weight of
Pten KO (Pten∆/∆) mice (n=8) and wild type littermates (Pten+/+) (n=8). (B) Whole prostate
weight of Pten∆/∆ mice (n=8) and Pten+/+ mice (n=8). (C) Individual lobe weights of
Pten∆/∆ mice (n=8) and Pten+/+ mice (n=8). AP, anterior prostate; DLP, dorsolateral
prostate; VP, ventral prostate. *P = 0.05 or 2-sided t-test p values were shown.
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236

19
Figure 3. Long-term oral MSeA supplementation inhibited prostate cancer progression in Pten
KO mice. Oral supplementation of MSeA (3 mg Se/kg body weight, 5 days per week) was
initiated at 10 weeks of age for 25 weeks. (A and B) Representative photomicrographs of
H&E stained prostate lobes of (A) Pten KO (Pten∆/∆) mice treated with water (Con) or
MSeA and (B) wild type (Pten+/+) mice treated with water (Con) or
MSeA. (C) Quantitation of anterior (AP), ventral (VP) and dorsolateral (DLP) prostate
pathological changes. Mean + SEM, Pten∆/∆ mice (n=8) and Pten+/+ mice (n=8). *Denotes
statistical difference from Pten KO control mice, p<0.05.
Figure 4. Effect of long-term MSeA supplementation on p-Akt and androgen receptor (AR),
senescence and cellular proliferative index in the anterior prostate of mice in Figure 3.
(A,B) Representative photomicrographs of IHC staining of pAkt Ser473, AR, Ki67 and
SA-galactosidase activity in anterior prostate of (A) Pten KO (Pten∆/∆) mice treated with
water (Con) or MSeA and (B) wild type (Pten+/+) mice treated with water (Con) or
MSeA. (C) Quantitation of changes from A and B. Mean + SEM, Pten∆/∆ mice (n=8) and
Pten+/+ mice (n=8). The integrated optical density (IOD) was estimated by ImagePro-Plus
software. *Denotes statistical difference from Pten KO control mice, p<0.05.
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236

20
Figure 5. Selective suppression effect of oral supplement of MSeA (3 mg Se/kg body weight, 5
days per week) for 25 weeks on androgen receptor (AR) and p-Akt Ser473 abundance in
the Pten KO prostate. (A) Immunoblot analyses of AR, p-Akt and total Akt in the anterior
prostate lobe of Pten KO (Pten∆/∆) mice and wild type littermates (Pten+/+) treated with
water (Con) or MSeA. Two mice per group were randomly chosen for analyses. Tubulin
served as loading control. (B) Schematic illustration of temporal sequence of cellular and
molecular events in Pten KO prostate carcinogenesis and the entry points of MSeA
actions.
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236

for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236

Published OnlineFirst October 28, 2015.Cancer Prev Res Lei Wang, Xiaolan Guo, Ji Wang, et al. progression barrier in prostate lesions of Pten-knockout mouseMethylseleninic acid super-activates P53-senescence cancer
Updated version
10.1158/1940-6207.CAPR-15-0236doi:
Access the most recent version of this article at:
Material
Supplementary
36.DC1
http://cancerpreventionresearch.aacrjournals.org/content/suppl/2016/01/09/1940-6207.CAPR-15-02Access the most recent supplemental material at:
Manuscript
Authoredited. Author manuscripts have been peer reviewed and accepted for publication but have not yet been
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.36http://cancerpreventionresearch.aacrjournals.org/content/early/2015/10/28/1940-6207.CAPR-15-02To request permission to re-use all or part of this article, use this link
for Cancer Research. on September 7, 2018. © 2015 American Associationcancerpreventionresearch.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on October 28, 2015; DOI: 10.1158/1940-6207.CAPR-15-0236