metabolic fate of myo-inositol in soybean callus cells
TRANSCRIPT

Plant Science. 75 (1991) 203--213 203 Elsevier Scientific Publishers Ireland Ltd.
Metabolic fate of myo-inositol in soybean callus cells
M a r k Biffen a n d D a v i d E. H a n k e
Botany School. University of Cambridge. Downing Street, Cambridge. CB2 3EA. England ( U.K.)
(Received September 4th, 1990; revision received November 14th, 1990; accepted January 7th, 1991)
For cell division, soybean callus cells in suspension culture require an intracellular level of no'o-inositol (ins) in excess of that re- quired to maintain viability and growth (M. Biffen and D.E. Hanke, Biochem. J., 265 (1990) 809--814). In pursuit of the cause of this requirement we have investigated Ins metabolism at different intracellular levels of Ins generated by incubating cells with 2-deoxyglucose or varying concentrations of Ins. incorporation of Ins into phospholipid was constant over the range of intracellular Ins concentrations, whereas when intracellular Ins levels were raised there was a proportionately higher incorporation of Ins into pectin, the result of an increase in the relative contribution of Ins to pectin monomers. Oxidation of Ins to uronic acid residues of pectin was inhibited by the Ins-antagonist 2-O,C-methylene myo-inositol (MMO). The inhibition was prevented by the addition of Ins. MMO treatment did not affect cell division, showing that a high rate of Ins oxidation is not necessary for cell division. MMO treatment reduced cell fresh mass and the incorporation of [3H]lns into phospholipid and both of these effects were not prevented by exogenous Ins. It follows that where effects of MMO are detected they may not be due specifically to inhibition of Ins oxidation. In conclusion, we have largely eliminated the possibility that the requirement for Ins in cell division is to sustain Ins incorporation into pectin, instead our results suggest that the function of Ins oxidation and incorporation into pectin is the irreversible elimination of Ins in excess of the cells immediate requirements.
Key words: myo-inositol; pectin; polyphosphoinositide; soybean (cell-suspension)
Introduction
Myo-inositol (Ins) is a standard constituent of plant media. It is readily taken up by growing cells and once in the cell can be metabolised via a number of diverse pathways. The flux through these pathways is dependent upon the growth phase and developmental state of the plant. Incor- poration of Ins into phytate predominates in seed formation [1], whereas young dividing tissues ox- idise Ins, producing glucuronic acid which is utilis- ed for pectin in the cell wall [2,3]. Myo-inositol is incorporated into phosphatidylinositol (PI) which, in soybean callus cells, accounts for 10% of cell phospholipid [4] and it is also incorporated into
Correspondence to: Mark Biffen, Botany School, University of Cambridge, Downing Street, Cambridge, CB2 3EA, U.K. Abbreviations: dGIc, 2-deoxyglucose; Ins, myo-inositol; MMO, 2-O,C-methylene myo-inositol; Pi, phosphatidylinositol; PIP, phosphatidylinositol monophosphate, PIP 2, phos- phatidylinositol bisphosphate.
phosphorylated species of PI, namely PIP and PIP2 [5]. A distinct group of inositol-containing phosphosphingolipids also occurs in plants [6]. Myo-inositol can be conjugated to indoleacetic acid [7] , methylated for form mono-ether derivatives [ 8 ], or Ins may be metabolised to form galactinol, a compound which acts as a galactose donor in the formation of the raffinose family of carbohydrates [9].
Although Ins is oxidised and the products incor- porated into pectin, the production of pectin is not solely from Ins. Glucose is also incorporated into the uronic acid fraction of pectin [10] by oxidation of UDP-glucose by UDP-glucose dehydrogenase [11] followed by isomerisation of the product, UDP-glucuronic acid, to UDP-galacturonic acid, a major precursor of pectin. The oxidation of Ins to glucuronic acid also leads to production of UDP-glucuronic acid, by way of phosphorylation of glucuronate by glucuronokinase [12] and uridylyl transfer by UDP-glucuronic acid
0168-9452/91/$03.50 © 1991 Elsevier Scientific Publishers Ireland Ltd. Printed and Published in Ireland

204
pyrophosphorylase [ 13]. Therefore, both pathways result in formation of UDP-glucuronic acid. Why plants possess both pathways is not known, neither is the compartmentation of these pathways.
To gain an understanding of the metabolism of Ins we made use of the finding that dGlc treatment of soybean cells reduced levels of intracellular Ins [14]. These lowered levels of intracellular Ins coin- cided with reduced cell division but did not affect cell viability. By altering the intracellular level of lns we hoped to detect changes in either the incor- poration of Ins into phospholipids, or oxidation of Ins to pectin, to gain an insight into which of these pathways was important in the process of cell division.
We proposed to dissect out the 2 pathways using the compound 2-O,C-methylene myo-inositol (MMO) a putative inhibitor of Ins-oxygenase [15]. In vitro inhibition has been demonstrated for the enzyme from rat kidney [16] and there is evidence for inhibition by MMO of the incorporation of [3H]Ins into pectin in Lilium pollen [17] and in germinating wheat [3].
Materials and Methods
Cell culture Soybean (Glycine max [L.] Merr. cv. Acme) cells
were cultured in the dark in 500-ml Ehrlenmeyer flasks using medium and conditions previously described [18]. The plant growth substances naphth-l-ylacetic acid (11 #M) and kinetin (2.3 /~M) were present in all culture medium used. The level of myo-inositol, when present, was 0.5 mM in the fresh medium at the time of subculturing. Subsequent incubations were set up by subcultur- ing 4 ml of the 7-day-old stock culture cells into 20 ml medium in 100-ml Ehrlenmeyer flasks under aseptic conditions. Each flask was stoppered with a cotton wool bung, covered in aluminium foil and incubated at 25°C on an orbital shaker set at 100 rev/min.
Cell counts, yield and viabilio, Cell suspension, 0.5 ml, was added to 1 ml of
15% (w/v) chromium trioxide solution and heated at 70°C for 30 rain. Water, 3.5 ml, was added and
the cells passed twice through a hypodermic needle (int. diam., 0.8 mm) and counted on a modified Fuchs-Rosenthal haemocytometer with a clearance of 0.2 mm. The density of stock cells at 7 days prior to subculturing was 2.5--3.0 x l0 6
cells ml- I. The fresh cell mass was determined by pipetting
a known volume of cells onto glass fibre paper (Whatman GF/A) in a Buchner flask under vacu- um. The cell pellet on the filter paper was weighed and the yield presented in units of mg fresh mass per ml of suspension.
Cell viability was based on exclusion of the dye phenosafranin [19]. By this method cell viability in all the experiments described was between 90 and 95%.
Radiolabelling with [3H]Ins Total uptake was measured by harvesting cells
(0.5-ml samples) as for fresh mass determinations. The cell pellet was washed with ice-cold 20 g 1-t sucrose solution to remove radiolabel not taken up into the cells. Cells were then scraped into vials, scintillation fluid (Optiphase 'Hisafe' II) added and the radioactivity quantified in a Packard 2000 Tri-Carb liquid scintillation analyser.
Extraction and separation of phospholipids The method was based on that of Schacht [20].
Cells were harvested on filter paper and added to 3 ml chloroform/methanol (1:2, v/v). Further addi- tions were chloroform, 1.0 ml, 2.4 M HCI, 1.0 ml and water to a total of 0.8 ml (the amount of water added was adjusted to take account of the tissue content, approx. 95% water by weight). After mix- ing vigorously the sample was centrifuged in a MSE Centaur for 5 min at gm~x = 1120 to aid phase separation. The lower phase was pipetted off and the upper phase re-extracted with 1.6 ml chloroform. The combined lower phases were dried under a stream of nitrogen. The phospho- lipids were resuspended in chloroform/methanol (2: l v/v) and spotted onto Silica G thin-layer chro- matography plates, previously sprayed with 10 g l-1 potassium oxalate in water/methanol (3:2, v/v) and air dried. The plates were developed by ascen- ding chromatography in chloroform/acetone/ methanol/acetic acid/water (40:15:13:12:8, by vol.),

[21] and the phospholipids visualised by molybdenum blue spray. The TLC plate was cut up into 5-mm strips, scintillation fluid added and radioactivity determined after 24 h.
Phosphate analysis o['phospholipids An equal volume of redissolved phospholipid
was either added to a glass vial and dried or ap- plied to a TLC plate and phospholipids separated as above. The area on the TLC plate correspon- ding to PI was scraped into a glass vial and to these samples and those of dried phospholipid (for total phosphate) 200 ~1 11.6 M perchloric acid: 2.5 M H2804 (1:1, v/v) was added and the samples heated at 200°C in a well-vented fume chamber for 1 h. When cool, 2.5 ml water, was added and the silica sedimented by centrifugation. Two miililitres were removed, and the phosphate quantified by the method of Fiske and Subbarow [22].
lnositol determination The method is described in detail elsewhere [14].
Cells were harvested on glass fibre paper under vacuum and the intracellular Ins extracted by boil- ing the cells in 80% (v/v) ethanol. The supernatant was collected following centrifugation and the sucrose hydrolysed by boiling the samples in 0.1 M H2SO 4. The sample was then made alkali by add- ing excess Ba(OH) 2 solution and boiled for a fur- ther 30 rain. The sample was neutralised and passed through Dowex-50 (H + form) followed by Dowex-1 (OH- form) and the eluate dried. The sample was dissolved in anhydrous pyridine and the Ins converted to the trimethylsilyl derivative. Ins concentrations were determined by gas chro- matography and quantified against a standard
curve.
Pectin extraction and determination Cell suspension, 16 ml, was collected on glass
fibre paper and boiled in 80% (v/v) ethanol for 2 min. The cell debris was pelleted by centrifugation, washed with 80% (v/v) ethanol and re-pelleted. Five millilitres of sodium hexametaphosphate so- lution (20 g 1-1), brought to pH 3.5 with HC1, was added and the sample boiled for 4 h [23]. When cooled, the sample was filtered by passing through glass wool in a 5-ml syringe and the clear filtrate
205
collected in dialysis tubing (26 mm flat width) and dialysed for 16 h against 1 1 deionised water with 2 changes of the washing solution. The sample was made 80% (v/v) with ice-cold ethanol, left until a precipitate formed and the precipitate collected by centrifugation. The precipitate was dissolved with gentle warming in 2.5 ml water. Samples, 0.5 ml, were taken for scintillation counting to determine incorporation of radiolabel from [3H]Ins. The re- maining sample was diluted to 10 ml and the amount of uronic acid in the pectin determined.
Uronic acid determination was based on the method of Blumenkrantz and Asboe-Hansen [24]. Determinations were in triplicate. Sample, 100/zl, was mixed with 0.15 M sodium tetraborate, 100 IA, and the tubes put on ice. Concentrated H2804, 1.2 ml, was added and the tubes whirlimixed vigorous- ly before being placed in a boiling water bath for 5 min. The tubes were cooled on ice, 20/xl 1.5 g 1-1 3-phenylphenol in 5 g I-1 NaOH added and mixed. The tubes were left for 10 min for the col- our to develop and A520 determined. A linear standard curve was constructed with up to 20 tzg glucuronic acid, galacturonic acid or polygalac- turonic acid. The actual form of uronic acid used did not affect the absorbance.
[2-3H]Ins was from Amersham International, Aylesbury, Buckinghamshire, U.K. All chemicals were obtained from Sigma, Poole, Dorset, U.K. Dialysis tubing and Optiphase HiSafe II were from FSA Laboratory Supplies, Leicester, U.K.
Results
Previous work of ours established a link be- tween the level of intracellular Ins in soybean cal- lus cells and growth by cell division [14]. Lowering the level of Ins in the cells resulted in a decreased ability of the cells to divide. Ins is metabolised by several different pathways in plant cells. We decid- ed to study the 2 major pathways common to all dividing plant cells, incorporation into phospho- lipids and oxidation to pectins, to determine whether altering the intracellular concentration of Ins changed the utilisation of Ins by these pathways.
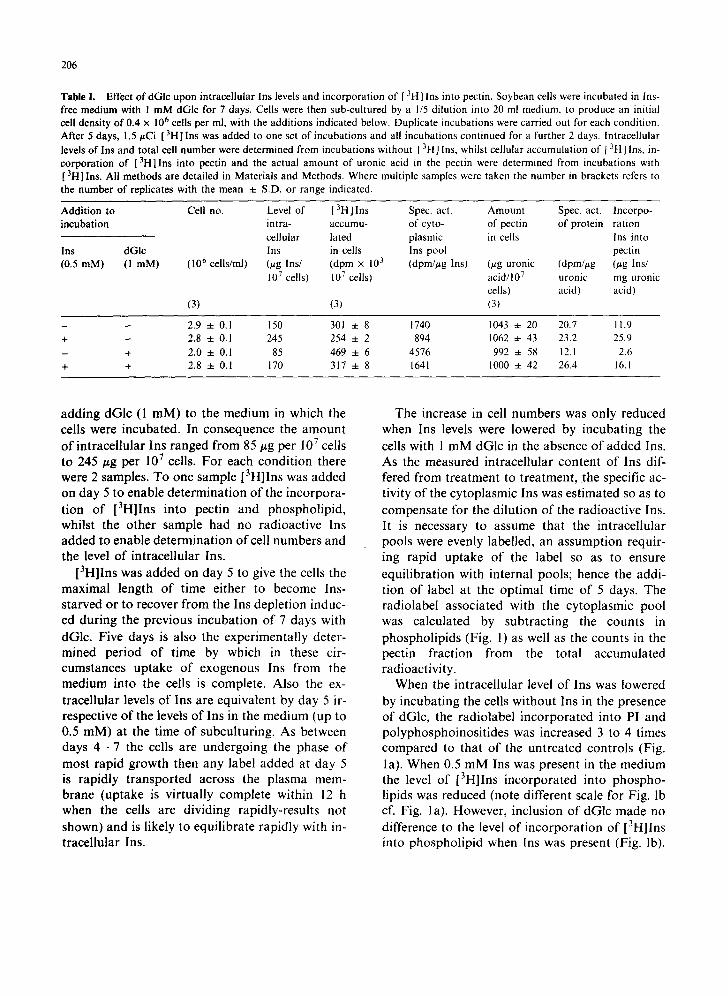
Table I shows the results of an experiment in which the intracellular level of Ins was varied by
206
Table I. Effect of dGlc upon intracellular Ins levels and incorporation of [ 31-1] Ins into pectin. Soybean cells were incubated in Ins- free medium with ! m M dGlc for 7 days. Cells were then sub-cultured by a 1/5 dilution into 20 ml medium, to produce an initial cell density of 0.4 x 106 cells per ml, with the additions indicated below. Duplicate incubations were carried out for each condition. After 5 days, 1.5/zCi [ 3HI Ins was added to one set of incubations and all incubations continued for a further 2 days. lntracellular levels o f Ins and total cell number were determined from incubations without [ 3H ] Ins, whilst cellular accumulation of [ 3H] Ins, in- corporation of [3H]Ins into pectin and the actual amount of uronic acid in the pectin were determined from incubations with [ 3H] Ins. All methods are detailed in Materials and Methods. Where multiple samples were taken the number in brackets refers to the number of replicates with the mean ± S.D. or range indicated.
Addition to Cell no. Level of [3H]Ins Spec. act. Amount Spec. act. Incorpo- incubation intra- accumu- of cyto- of pectin of protein ration
cellular lated plasmic in cells Ins into Ins dGIc Ins in ceils Ins pool pectin (0.5 mM) (1 raM) (106 cells/ml) (~tg Ins/ (dpm × 103 (dprn//~g Ins) (#g uronic (dpm/#g (/~g Ins/
107 cells) 107 cells) acid/107 uronic mg uronic ceils) acid) acid)
(3) (3) (3)
- - 2.9 ± 0.1 150 301 4- 8 1740 1043 ± 20 20.7 11.9 + - 2.8 ± 0.1 245 254 -4- 2 894 1062 ± 43 23.2 25.9 - + 2.0 ± 0.1 85 469 ± 6 4576 992 ± 58 12.1 2.6 + + 2.8 ± 0.1 170 317 + 8 1641 1000 ± 42 26.4 16.1
adding dGlc (1 mM) to the medium in which the cells were incubated. In consequence the amount of intracellular Ins ranged from 85 #g per 107 cells to 245/~g per 107 cells. For each condition there were 2 samples. To one sample [3H]Ins was added on day 5 to enable determination of the incorpora- tion of [3H]Ins into pectin and phospholipid, whilst the other sample had no radioactive Ins added to enable determination of cell numbers and the level of intracellular Ins.
[3H]Ins was added on day 5 to give the cells the maximal length of time either to become Ins- starved or to recover from the Ins depletion induc- ed during the previous incubation of 7 days with dGlc. Five days is also the experimentally deter- mined period of time by which in these cir- cumstances uptake of exogenous Ins from the medium into the cells is complete. Also the ex- tracellular levels of Ins are equivalent by day 5 ir- respective of the levels of Ins in the medium (up to 0.5 raM) at the time of subculturing. As between days 4--7 the cells are undergoing the phase of most rapid growth then any label added at day 5 is rapidly transported across the plasma mem- brane (uptake is virtually complete within 12 h when the cells are dividing rapidly-results not shown) and is likely to equilibrate rapidly with in- tracellular Ins.
The increase in cell numbers was only reduced when Ins levels were lowered by incubating the cells with 1 mM dGlc in the absence of added Ins. As the measured intracellular content of Ins dif- fered from treatment to treatment, the specific ac- tivity of the cytoplasmic Ins was estimated so as to compensate for the dilution of the radioactive Ins. It is necessary to assume that the intracellular pools were evenly labelled, an assumption requir- ing rapid uptake of the label so as to ensure equilibration with internal pools; hence the addi- tion of label at the optimal time of 5 days. The radiolabel associated with the cytoplasmic pool was calculated by subtracting the counts in phospholipids (Fig. 1) as well as the counts in the pectin fraction from the total accumulated radioactivity.
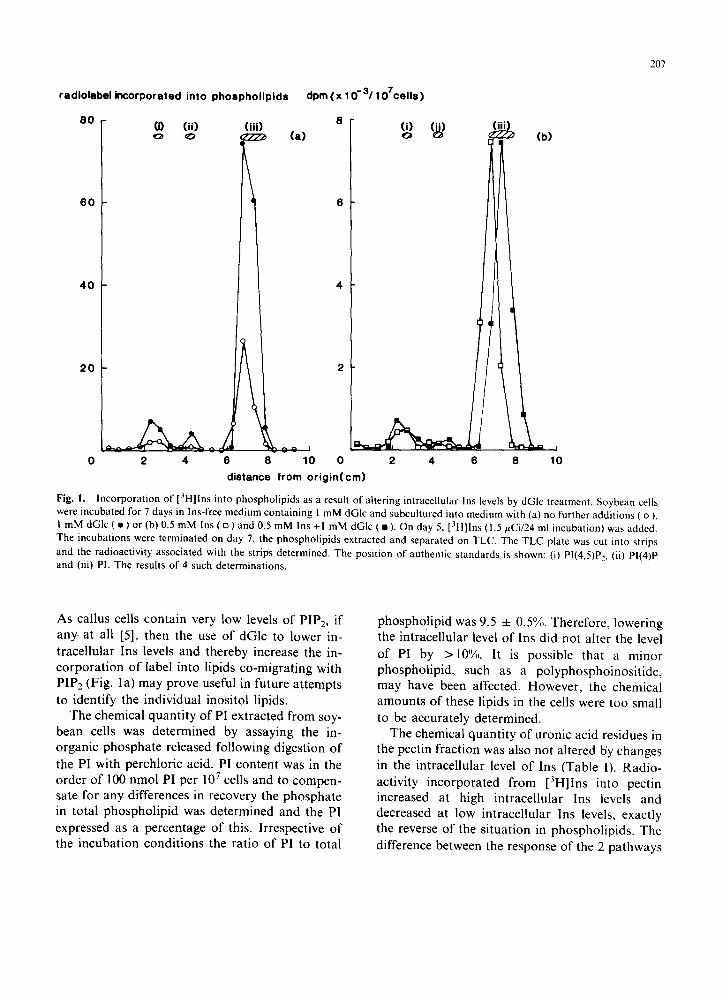
When the intracellular level of Ins was lowered by incubating the cells without Ins in the presence of dGlc, the radiolabel incorporated into PI and polyphosphoinositides was increased 3 to 4 times compared to that of the untreated controls (Fig. la). When 0.5 mM Ins was present in the medium the level of [3H]Ins incorporated into phospho- lipids was reduced (note different scale for Fig. lb cf. Fig. la). However, inclusion of dGlc made no difference to the level of incorporation of [3H]Ins into phospholipid when Ins was present (Fig. lb).
207
radiolabel incorporated into phospholipids dpm (x 10-3/107cells )
80 8
6 0
4 0
2 0
(0 (ii) (iii) o ~ ~ (a)
4
(i) (i~) o (b)
2 4 6 10 8
ti 0 0 2 4 6 8 10
dis tance from o r i g i n ( c m )
Fig. I. Incorporation of [3H]Ins into phospholipids as a result of altering intracellular Ins levels by dGIc treatment. Soybean cells were incubated for 7 days in Ins-free medium containing I mM dGlc and subcultured into medium with (a) no further additions ( o ), 1 mM dGIc ( • ) or (b) 0.5 mM Ins ( o ) and 0.5 mM Ins + I mM dGlc ( • ). On day 5, [3H]lns ( 1.5 #Ci/24 ml incubation) was added. The incubations were terminated on day 7, the phospholipids extracted and separated on TLC. The TLC plate was cut into strips and the radioactivity associated with the strips determined. The position of authentic standards is shown: (i) PI(4,5)P~, (ii) PI(4)P and (iii) PI. The results of 4 such determinations.
As cal lus cells con ta in very low levels o f PIP2, if
any. at all [5], then the use o f dGlc to lower in- t race l lu la r Ins levels and thereby increase the in- c o r p o r a t i o n o f label into l ipids co-migra t ing with PIP2 (Fig. la ) may prove useful in future a t t empts to ident i fy the ind iv idua l inosi tol l ipids.
The chemical quan t i ty o f PI ex t rac ted f rom soy- bean cells was de te rmined by assaying the in- o rganic p h o s p h a t e released fo l lowing digest ion o f
the PI with perch lor ic acid. PI con ten t was in the o rde r o f 100 nmol PI per 10 7 cells and to compen- sate for any differences in recovery the phospha t e in to ta l phospho l i p id was de t e rmined and the PI
expressed as a percentage o f this. I r respect ive o f the incuba t ion cond i t ions the ra t io o f PI to total
phospho! ip id was 9.5 -4- 0.5%. Therefore , lower ing the in t race l lu lar level o f Ins d id not a l ter the level
o f PI by >10%. It is poss ible that a minor phospho l ip id , such as a po lyphospho inos i t i de , may have been affected. However , the chemical amoun t s o f these l ipids in the cells were too small to be accura te ly de te rmined .
The chemical quan t i ty o f uronic acid residues in the pect in f ract ion was also not a l tered by changes
in the in t race l lu la r level o f Ins (Table I). Rad io - act ivi ty i nco rpo ra t ed f rom [3H]Ins into pectin increased at high in t race l lu la r Ins levels and decreased at low in t race l lu la r Ins levels, exact ly the reverse o f the s i tua t ion in phosphol ip ids . The
difference between the response o f the 2 pa thways
208
to altered intracellular levels of Ins is very clear and dramatic. When Ins levels were lowered the in- crease in the specific activity of free Ins was match- ed by increased incorporation of [3H]Ins into phospholipid (Fig. 1) showing that the rate of this pathway was maintained, while the specific radioactivity of pectin declined, evidence that this pathway was being suppressed. From the specific activity of the intracellular Ins and the specific ac- tivity of the pectin extracted it was possible to estimate how much Ins was incorporated into the uronic acid fraction. In cells incubated with ex- ogenous Ins an estimated 26 #g Ins were incor- porated into 1 mg of uronic acid, whereas when Ins was omitted from the medium and dGlc in- cluded only 3 t~g Ins were incorporated per mg of uronic acid. These figures show that in soybean callus cells inositol oxidation products are a minor component of the pectin fraction, accounting for < 5% by mass of the uronic acid residues of the pectin. This value is likely to be an underestimate as the determination of the mass of pectin includes any unlabelled pectin made during the initial 5 days of incubation. As cell division and growth is delayed up to day 4, the contribution of unlabelled pectin, at the time of addition of label, to total pec- tin on day 7 is likely to be less than 50%.
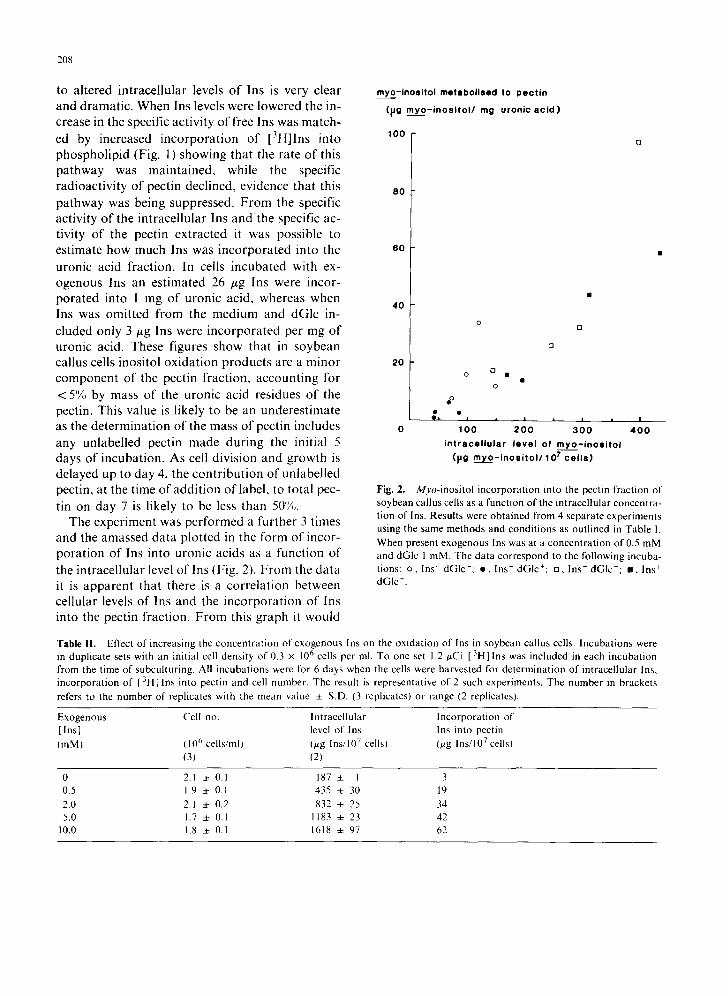
The experiment was performed a further 3 times and the amassed data plotted in the form of incor- poration of Ins into uronic acids as a function of the intracellular level of Ins (Fig. 2). From the data it is apparent that there is a correlation between cellular levels of Ins and the incorporation of Ins into the pectin fraction. From this graph it would
myo-ino$itol metebolised to pectin
(iJg myo- inoei to l / mg uronicacid)
100
80
60
40
20
0 [3
o o •
o
eo
a 8 a I i |
0 tO0 200 3 0 0 4 0 0 intracel luler level of m y o - i n o s i t o l
([Jg m ¥ o - i n o s i t o l / l O 7 cel ls)
Fig. 2. Myo-inositol incorporation into the pectin fraction of soybean callus cells as a function of the intracellular concentra- tion of Ins. Results were obtained from 4 separate experiments using the same methods and conditions as outlined in Table 1. When present exogenous Ins was at a concentration of 0.5 mM and dGlc 1 mM. The data correspond to the following incuba- tions: o, Ins- dGIc-: e , Ins- dGIc+: ,7 Ins + dGIc-; m, Ins + dGIc +.
Table I1. Effect of increasing the concentration of exogenous Ins on the oxidation of Ins in soybean callus cells. Incubations were in duplicate sets with an initial cell density of 0.3 × 10 6 cells per ml. To one set 1.2 ~Ci [3H] Ins was included in each incubation from the time of subculturing. All incubations were for 6 days when the cells were harvested for determination of intracellular Ins, incorporation of [ 3H]lns into pectin and cell number. The result is representative of 2 such experiments. The number in brackets
refers to the number of replicates with the mean value 4- S.D. (3 replicates) or range (2 replicates).
Exogenous Cell no. Intracellular Incorporation of [Ins] level of Ins Ins into pectin (mM) (10 6 cells/ml) (~g Ins/10 7 cells) (~g Ins/10 7 cells)
(3) (2)
0 2.1 4. 0.1 187 + 1 3 0.5 1.9 :~ 0.1 435 4. 30 19
2.0 2.1 :~ 0.2 832 + 25 34 5.0 1.7 ~ 0.1 1183 ± 23 42
10.0 1.8 :~ 0.1 1618 ± 97 62

be expected that at an intracellular level of Ins of about 30 t~g Ins/107 cells the oxidation of Ins would cease. As the incorporation of Ins into the uronic acid component of the pectin was related to the level of intracellular Ins irrespective of how that level of Ins was generated this suggests that dGlc was not directly inhibiting Ins oxidation. In- stead dGlc will be expected to inhibit Ins- oxidation as a consequence of lowering the cellular level of Ins.
For further examination of the effect of in- tracellular Ins concentration upon Ins oxidation, soybean callus cells were incubated in medium with concentrations of exogenous Ins ranging from 0 to 10 mM (Table II). To a parallel set of in- cubations [3H]Ins was also added for the estima- tion of Ins oxidation to pectins. Over the 6 days incubation the cells were unable to take up all the extracellular Ins at the higher concentrations so the accumulation of label varied. To calculate the specific activity of the Ins in each incubation the initial total content of Ins in each flask was calculated, including 300 ~g Ins which was carried over in the inoculum of cells. [3H]Ins was present at t = 0 and the initial specific activity determined. After 6 days the counts incorporated into pectin were converted to an amount of Ins using this specific activity. The largest errors are likely to be in the case of the incubations in which little or no unlabelled exogenous Ins was added, as Ins- synthesis would have occurred and diluted the radioactivity. However, at higher levels of ex- ogenous Ins the errors are likely to be reduced as L-l-myo-inositol phosphate synthase would be suppressed [25].
Increasing exogenous Ins to 10 mM had no detectable effect upon cell division. When cells were incubated in 10 mM Ins the intracellular level of Ins was 1.6 mg per 10 7 cells. As the fresh mass of 10 7 cells was 320 mg, without compensating for dry mass this gives a mean intracellular concentra- tion of 28 mM. Clearly cells are able to store large quantities of Ins. Whether the vacuole plays a part in this is not known. When the amount of lns con- verted to uronic acid was calculated it was ap- parent that increased availability of Ins had resulted in increased incorporation of Ins into pec- tin (Table II).
209
To try to assess the importance of Ins oxidation in cell growth and division an inhibitor of Ins- oxidation, MMO, was used. The results are presented as Table 11I. Incubation of soybean cal- lus cells with 0.4 mM MMO in Ins-free medium for 7 days had no detectable effect on cell division, though the cell wet-mass was reduced to nearly 50% of that of cells incubated without MMO. The effect upon cell mass was not reversed when 0.5 mM Ins was included in the incubation. MMO was used at 0.4 mM as at this concentration cell viabili- ty was not affected. At 0.2 mM MMO or less no effect on cell mass was detected and at higher con- centrations browning of the callus cells was
observed. When cells were incubated for up to 5 days with
MMO no change in the level of pectin labelling from [3H]Ins was observed, but a reduction in the incorporation of [3H]lns into pectin was observed if incubations were continued beyond 5 days (results not shown). Therefore, cells were pre- incubated with MMO for 5 days before [3H]lns was added. The cells were then harvested on day 7. MMO had no effect upon the intracellular level of Ins when Ins was present in the medium. The detection and quantification of Ins by gas chroma- tography was not affected by MMO as MMO in standards which had been through the procedure used to extract Ins from soybean callus cells eluted from the column 1.3 min after Ins.
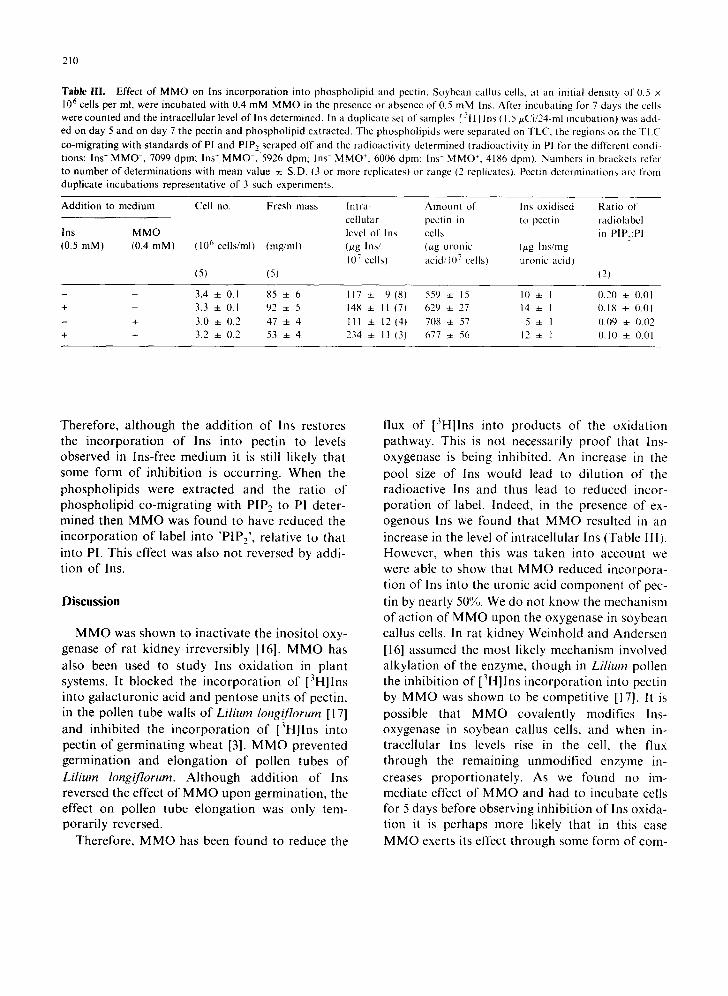
Addition of MMO to the medium resulted in a 17% increase in the mass of polyuronate per cell. Since as a result of this treatment the total fresh mass per cell was decreased by some 37%, the con- tribution of the uronic acid residues of pectin to cell mass appears to have doubled, approximately (1.9 x ). Using the specific radioactivity of the in- tracellular Ins to calculate the contribution from Ins to uronyl monomers in pectin (as in Table I), it appears that MMO in the absence of exogenous Ins reduced the level of incorporation of Ins into pectin by nearly 50%. When Ins was included in the medium with MMO, the level of Ins incor- porated into pectin was comparable to that estimated for cells in Ins-free medium without MMO. However, with an inositol content of 234 tzg per 10 7 cells higher levels of incorporation of Ins into pectin would be expected (from Fig. 2).
210
Table III , Effect o f M M O on Ins inco rpora t ion into phospho l ip id and pectin. Soybean cal lus cells, at an init ial densi ty of 0.5 x 106 cells per ml, were incuba ted with 0.4 mM M M O in the presence or absence of 0.5 mM Ins. After incuba t ing for 7 days the cells were counted and the in t race l lu la r level of Ins determined. In a dupl ica te set of samples [ ~H] Ins ( 1.5 p~Ci/24-ml incubat ion) was add-
ed on day 5 and on day 7 the pectin and phospho l ip id extracted. The phospho l ip ids were separa ted on TLC, the regions on the TLC
co-migra t ing with s tandards of PI and P IP , scraped off and the rad ioac t iv i ty de termined ( radioact iv i ty in PI for the different condi- t ions: Ins- M M O - , 7099 dpm; Ins + M M O , 5926 dpm; Ins M M O +, 6006 dpm: Ins + M M O +, 4186 dpm). Numbers in brackets refer to number of de te rmina t ions with mean value 4- S.D. (3 or more replicates) or range (2 replicatest . Pectin de te rmina t ions are from dupl ica te incuba t ions representa t ive of 3 such exper iments .
Add i t i on to med ium Cell no. Fresh mass ln t ra- A m o u n t of Ins oxidised Ra t io of
cel lular pectin in to pectin radiolabel Ins M M O level of Ins cells in P[P,:PI (0.5 raM) (0.4 mM) (106 cells/ml) (mg/ml) (#g Ins/ (~g uronic (/xg Ins/rag
107 cells) acid/107 cells) uronic acid)
(5) (5) (2)
- - 3.4 4- 0.1 85 4- 6 117 4- 9 (81 559 4- 15 10 4- I 0.20 4- 0.01 + - 3.3 + 0.1 92 + 5 148 ± I1 (7) 629 4- 27 14 ± 1 0.18 4- 0.01
- + 3.0 4- 0.2 47 + 4 111 4- 12(4! 708 ~: 57 5 + I 0.09 4- 0.02
+ + 3.2 4- 0.2 53 4- 4 234 + 11 (3) 677 ± 56 12 4- I 0.10 + 0.01
Therefore, although the addition of Ins restores the incorporation of Ins into pectin to levels observed in Ins-free medium it is still likely that some form of inhibition is occurring. When the phospholipids were extracted and the ratio of phospholipid co-migrating with PIP2 to Pl deter- mined then MMO was found to have reduced the incorporation of label into 'PIP2' , relative to that into PI. This effect was also not reversed by addi- tion of Ins.
Discussion
MMO was shown to inactivate the inositol oxy- genase of rat kidney irreversibly [161, MMO has also been used to study Ins oxidation in plant systems. It blocked the incorporation of [3H]Ins into galacturonic acid and pentose units of pectin. in the pollen tube walls of Lilium longi[lorum [17] and inhibited the incorporation of [3H]Ins into pectin of germinating wheat [3]. MMO prevented germination and elongation of pollen tubes of Lilium longiflorum. Although addition of Ins reversed the effect of MMO upon germination, the effect on pollen tube elongation was only tem- porarily reversed.
Therefore, MMO has been found to reduce the
flux of [3Hllns into products of the oxidation pathway. This is not necessarily proof that Ins- oxygenase is being inhibited. An increase in the pool size of Ins would lead to dilution of the radioactive Ins and thus lead to reduced incor- poration of label. Indeed, in the presence of ex- ogenous Ins we found that MMO resulted in an increase in the level of intracellular Ins (Table 1II). However, when this was taken into account we were able to show that MMO reduced incorpora- tion of lns into the uronic acid component of pec- tin by nearly 50%. We do not know the mechanism of action of MMO upon the oxygenase in soybean callus cells. In rat kidney Weinhold and Andersen [161 assumed the most likely mechanism involved alkylation of the enzyme, though in Lilium pollen the inhibition of [3H]lns incorporation into pectin by MMO was shown to be competitive [17]. It is possible that MMO covalently modifies Ins- oxygenase in soybean callus cells, and when in- tracellular Ins levels rise in the cell, the flux through the remaining unmodified enzyme in- creases proportionately. As we found no im- mediate effect of MMO and had to incubate cells for 5 days before observing inhibition of Ins oxida- tion it is perhaps more likely that in this case MMO exerts its effect through some form of corn-

petitive inhibition of the Ins-oxygenase. Nucleophilic attack on the methyl-group in MMO could result in methylation of the attacking group and formation of inos-2-ose. Such a compound has been shown to be an inhibitor of Ins- oxygenase in rat kidney [26].
The in vivo effects of MMO are diverse, sug- gesting that it may interact with more than one component of the cell. It has been reported to in- hibit malic acid synthesis from glucose in Lilium longiflorum [27] and to reduce Ins biosynthesis in the kidney of rats [28], and we found that it reduc- ed the increase in fresh mass of soybean callus cells in suspension culture, an effect not reversed by ex- ogenous Ins. The failure of exogenous Ins to pre- vent the effect of MMO on fresh mass increase contrasts with the success of exogenous Ins in preventing the effect of MMO on Ins conversion to pectin. From this observation we assume that MMO probably acts on at least 2 different target processes in vivo. It follows that in this tissue, MMO action in vivo is not specific enough for it to be regarded as inhibiting the Ins-oxidation pathway only. This point does not invalidate the results of our investigation of the effect of MMO on cell division since MMO was found to be without any effect on division at the same time as it could be shown to inhibit Ins-oxidation in an Ins-reversible manner.
The effect of MMO on the incorporation of Ins into putative polyphosphoinositide relative to PI is of interest as Kaul and Lester [6] established that one of the components of the phospho- sphingolipids is glucuronic acid. If the phos- pholipid migrating with authentic PIP2 was poly- phosphoinositide then the ratio of label in 'PIP2', to that in PI would be expected to stay constant. For example, when exogenous Ins is added and reduces the specific activity of the in- tracellular Ins pool the ratio (0.18) is comparable with that obtained when Ins is omitted from the in- cubation (0.20), Table IIl. However, MMO reduces this ratio to 0.10 which suggests that the majority of this lipid is not PIP2, but possibly phosphosphingolipid. Thus MMO may be useful in establishing if PIP 2 is present by adding it to cells to reduce levels of phosphosphingolipid. As MMO decreased the fresh mass of soybean cells in
211
a seemingly Ins-dependent manner, and the ratio of labelling of 'PIP2', to PI in the presence of MMO was similarly Ins-dependent this raises the possibility that the lipid co-migrating with PIP, may be important in the regulation of cell ex- pansion.
From our experiments in which intracellular levels of Ins were manipulated either by addition of dGlc (Fig. 1) or by increasing the level of ex- ogenous Ins (Table 1I) it was apparent that the in- corporation of Ins into the pectin fraction was a function of the intracellular level of Ins. Whereas the level of PI in the membranes was maintained at a constant level irrespective of the cellular level of Ins, the incorporation of Ins into pectins was reduced as the level of cellular Ins declined. Although the amount of Ins oxidised to form the pectin fraction decreased when intracellular Ins was lowered, the actual level of pectic uronic acid in the cells was unchanged (Table I).
Estimates of the contribution of Ins oxidation to pectin vary. In corn (Zea mays) roots 14% of the [3H]lns taken up by the cells was converted to an ethanol-insoluble fraction [29], whereas in ger- minating Lilium Iongiflorum pollen 20% of the sup- plied Ins was converted to pectic wall substances [2]. However, in a callus tissue culture of Fraxinus pennsylvanica < 1% of the galacturonic acid incor- porated into pectin was derived from Ins [30], and Verma and Dougal [31] concluded that Ins- oxidation provided only a minor portion of the pentoses and uronic acids for cell wall biosynthesis in suspension cultures of wild carrot cells. Assum- ing that endogenous synthesis of Ins was minimal when the extracellular level of Ins was 10 mM we similarly observed low incorporation of Ins ( < 7% of the uronic acid residues) into pectin in soybean callus cells. What, then, is the importance of the pectic fraction derived from Ins? In a study of the activities of UDP-glucuronic acid pyrophos- phorylase, necessary for formation of UDP-glucu- ronic acid from Ins-oxidation, and UDP-glucose dehydrogenase, the enzyme responsible for UDP- glucuronic acid formation from glucose, the dehy- drogenase was found in both cambial tissue and the xylem tissue produced by it, whereas UDP- glucuronic acid pyrophosphorylase was only pres- ent in the cambium [32]. From the results of this

212
and studies of other tissues the authors concluded that UDP-glucuronic acid pyrophosphorylase was associated only with younger tissues synthesising primary wall. A separate issue is whether the pec- tin synthesised from Ins differs from the pectin made from glucose.
Although M M O did not specifically inhibit Ins- oxidation, it did reveal that when incorporation of Ins into pectin was reduced to levels similar to those obtained following dGlc treatment, cell divi- sion was not inhibited. Some inhibition might have been expected if less of a crucial pectin fraction (for example an oligosaccharin precursor) was syn- thesised when dGlc was present and as a result division was slowed or stopped. Therefore, cell division appears not to be linked to the level of Ins-oxidation products incorporated into the pec- tin. Instead we propose that Ins-oxidation is a means whereby a cell can eliminate excess Ins. When the level of Ins rises at the site of the enzyme then Ins is oxidised and removed by conversion to cell wall, thus preventing re-uptake. From our model, hexose, via UDP-glucose, normally sup- plies the bulk of the uronic acid residues of pectin and only excess Ins is oxidised and contributes to pectin synthesis.
Experiments to determine the relative contribu- tion of glucose and Ins to uronic residues of pectin have been reported. Roberts and Loewus [29] using Zea mays roots showed that [14C]GIc was incorporated into cell wall galacturonic acid. If unlabelled Ins was added then the incorporation of [14C]Glc was reduced. A similar result with cultured cells from Acer pseudoplatanus has also been reported [33]. From this it was concluded that glucose was converted to Ins and thence into pectin, as by adding Ins the [14C]lns pool made from [14C]Glc was being diluted. However, other work had shown that adding exogenous Ins sup- pressed L- 1-myo-inositol l -phosphate synthase ac- tivity in Acer [25], so adding Ins might be expected to suppress conversion of glucose to lns. We would predict that [14C]GIc would feed into galacturonosyl and arabinosyl units of pectic substances via UDP-glucose dehydrogenase, then when Ins was added this would provide an alter- native substrate and as a result the incorporation of glucose would decrease. As the authors later comment [17] 'the effect of Ins is best interpreted
as due to inhibition of UDP-glucose dehydroge- nase by products of Ins-produced glucuronate me- tabolism'. It will be necessary to examine the regulation of the enzymes in these pathways to see if this is true. I f Ins oxidation is linked to levels of intracellular Ins then there are at least 2 possible ways by which the process may be regulated. The expression of Ins-oxygenase may be induced by Ins, or the K m of the enzyme for Ins could be such that a high level of Ins has to be reached before a significant formation of glucuronic acid occurs.
In conclusion, the results have thrown some light on the changing pattern of Ins metabolism at different levels of intracellular Ins. Because the synthesis of PI is unaffected by declining levels of Ins, it is unlikely that the associated cessation of cell division is due to the lack of an inositol lipid, though a decline in the level of a minor phosphoinositide would not have been detected. From the results of MMO-treatment , incorpora- tion of Ins oxidation products into pectin does not appear to be essential for cell division. The de- pendency of the conversion of Ins into pectin on the level of cellular Ins shows it may be a way of removing excess Ins. By making it less likely that the source of the Ins requirement for cell division lies in either phospholipid or pectin, this study points towards some other Ins metabolite as necessary to sustain growth by cell division.
Acknowledgements
This work was funded by a grant from the Gatsby Charitable Foundation.
References
1 B.L. O'Dell, A.R. de Boland and S.R. Koirtyohann, Distribution of phytate and nutritionally important elements among the morphological components of cereal grains. J. Agric. Food Chem., 20 (1972) 718--721.
2 M. Chen and F.A. Loewus, Myo-inositol metabolism in Lilium longi['lorum pollen-uptake and incorporation of myo-inositol-2-3H. Plant Physiol., 59 (1977)653~ 657.
3 I.B. Maiti and F.A. Loewus, Myo-inositol metabolism in germinating wheat. Planta, 142 (1978) 55--60,
4 R.J.A. Connen and D.E. Hanke, Changes in the pattern of phospholipid synthesis during the induction by cytokinin of cell division in soybean suspension cultures. Planta, 170 (1987) 161--167.
5 R.F. Irvine, A.J. Letcher, D.J. Lander, B.K. Dr~bbak, A.P. Dawson and A. Musgrave, Phosphatidylinositol

(4,5)bisphosphate and phosphatidylinositol(4) phosphate in plant tissues. Plant Physiol., 89 (1989) 888--892.
6 K. Kaul and R.L. Lester, Characterization of inositol- containing phosphosphingolipids from tobacco leaves. Plant Physiol., 55 (1975) 120~129.
7 R.S. Bandurski, Chemistry and physiology of conjugates of indole-3-acetic acid, in: N.B. Mandava (Ed.), Plant Growth Substances, American Chemical Society, Washington D.C., 1979, pp. 1--17.
8 H. Kindl and O. Hoffman-OstenhoL Cyclite: Biosynthese, Stoffwechsel und Vorkommen. Fortschr. Chemi. Organ. Naturst., 24 (1966) 149--205.
9 L. Lehle, W. Tanner and O. Kandler, Myo-inositol, a co- factor in the biosynthesis of raffinose. Hoppe-Seylers Z. Physiol. Chem., 351 (1970) 1494-~1498.
10 P. Albersheim, Hormonal control of myo-inositol incor- poration into pectin. J. Biol. Chem., 238 (1963) 1608--1610.
11 J.L. Strominger and L.W. Mapson, Uridine diphosphoglucose dehydrogenase of Pea seedlings. Biochem. J,, 66 (1957) 567--572.
12 D.B. Dickinson, Occurrence of glucurokinase in various plant tissues and comparison of enzyme activity of seed- lings and green plants. Phytochem, 21 (1982) 843--844.
13 R.M. Roberts, The formation of uridine diphosphate- glucuronic acid in plants. J. Biol. Chem., 246 (1971) 4995--5002.
14 M. Biffen and D.E. Hanke, Reduction in the level of in- tracellular myo-inositol in cultured soybean (Glycine max) cells inhibits cell division. Biochem, J., 265 (1990) 809--814.
15 1". Posternak, Studies on the cyclitol series. VII. On the cyclitol of mussels (mytilitol) and on some related substances. Helv. Chim. Acta, 27 (1944)457~68.
16 P.A. Weinhold and L. Anderson, Metabolism of myo- inositol in animals. III. Action of the antagonist 2-O.C- methylene-myo-inositol. Arch. Biochem. Biophys., 122 (1967) 529--536.
17 M. Chen, M.W. Loewus and F.A. Loewus, Effect of a myo-inositol antagonist, 2-O. C-methylene-,myo-inositol, on the metabolism of myo-inositol-2-3H and D- glucose- 1-14C in Lilium longiflorum pollen. Plant Physiol., 59 (1977) 658--663.
18 R.J.A. Connett and D.E. Hanke, Breakdown of phosphatidylinositol in soybean callus. Planta, 169 (1986) 216--221.
19 J.M. Widholm, The use of fluorescein diacetate and phenosafranine for determining viability of cultured plant cells. Stain Technol., 47 (1972) 189--194.
213
20 J. Schacht, Extraction and purification of polyphos- phoinositides. Methods Enzymol., 72 (1981) 626~631.
21 S. Heim and K.G. Wagner, Evidence of phosphorylated phosphatidylinositols in the growth cycle of suspension cultured plant cells. Biochem. Biophys. Res. Commun., 134 (1986) 1175--1181.
22 C.H. Fiske and Y. Subbarow, The colorimetric determi- nation of phosphorous. J. Biol. Chem., 66 (1925) 375~00.
23 R.W. Stoddart, A.J. Barrett and D.H, Northcote, Pectic polysaccharides of growing plant tissue. Biochem. J., 102 (1967) 194--204.
24 N. Blumenkrantz and G. Asboe-Hansen, New method for quantitative determination of uronic acids, Anal. Biochem., 54 (1973) 484 489.
25 M.W. Loewus and F. Loewus, D-Glucose-phosphate cycloaldolase: inhibition studies and aldolase function. Plant Physiol., 51 (1973) 263--266.
26 F.C. Charalampous, Biochemical studies on inositol. VI. Mechanism of cleavage of inositol to D-glucuronic acid. J. Biol. Chem., 235 (1960) 1286~1291.
27 I.B. Maiti and F.A. Loewus, Evidence for a functional myo-inositol oxidation pathway in Lilium hmgiflorum pollen. Plant Physiol., 62 (1978) 280~283.
28 G. Hauser, M. Arnold and V.N. Finelli, The effect of the injection of 2-O. C-methylene-myo-inositol into rats on in- ositol metabolism in brain and kidney slices. Biochim. Biophys. Acta, 116 (1966) 125--132.
29 R.M. Roberts and F. Loewus, The conversion of D- glucose-6-14C to cell wall polysaccharide material in Zea mays in presence of high endogenous levels of myo- inositol. Plant Physiol., 52 (1973) 646~650.
30 P. Jung, W. Tanner and K. Wolter, The fate of rnyo- inositol in Fraxinus tissue cultures. Phytochemistry, 11 (1972) 1655--1659.
31 D.C. Verma and D.K. Dougall, Biosynthesis of myo- inositol and its role as a precursor of cell-wall polysac- charide in suspension cultures of wild-carrot cells. Planta, 146 (1979) 55--62.
32 R.M. Roberts and J.J. Cetorelli, UDP-D-glucuronic acid pyrophosphorylase and the formation of UDP-D- glucuronic acid in plants, in: F. Loewus (Ed.). Biogenesis of Plant Cell Wall Polysaccharides, Academic Press, New York and London, 1973, pp. 49--68.
33 P.H. Rubery and D.H. Northcote, The effect of auxin (2,4-dichlorophenoxyacetic acid) on the synthesis of cell wall polysaccharides in cultured sycamore cells. Biochim. Biophys. Acta, 222 (1970) 95--108.