metabolic action of prolactin in regressing prostate: independent of androgen action
TRANSCRIPT

The Prostate 6:49-59 (1985)
Metabolic Action of Prolactin in Regressing Prostate: Independent of Androgen Action Cornelius Smith, Dean Assimos, Chung Lee, and John T. Grayhack
Department of Urology, Northwestern University Medical School, Chicago
The mechanism of the observed synergistic effect of prolactin and androgen on the lateral lobe of the rat prostate is not established. The observation that prolactin alone delayed the rate of loss of weight, protein, and DNA of the lateral lobe in castrated rats has led us to question the assumption that the effect of prolactin is produced by a modification of recognized androgen-induced intracellular changes. The present study was conducted to explore whether or not the sites of prolactin action in the rat prostate coincided with those recognized as the androgen effect. Two anterior pituitaries from female donors were grafted under the right renal capsule of adult male Sprague-Dawley rats. Seven days later, bilateral orchiectomy and unilateral nephrectomy were performed in these rats. In one half of the animals, the kidney bearing the pituitary grafts was removed. In the other half, the contralateral kidney was removed. Seven days following the orchiectomy-nephrectorny, animals bearing the pituitary grafts had a higher level of serum prolactin (93 f 7 nglml, mean f SE) than in those without the graft (26 f 3 ng/ml). This condition of hyperprolac- tinemia was associated with the delay of castration-induced regression in the lateral prostate. The rate of protein degradation, as judged by the amount of radioactivity remaining in the tissue following a single i.v. pulse of 3H-leucine 24 hr before orchiectomy-nephrectomy, was significantly slower in the lateral prostate in graft-bearing animals than in those without grafts. However, the rate of protein synthesis, as judged by both in vitro and in vivo incorporation of 3H-leucine into the protein fractions of the prostate, was not significantly different between the two groups. Nor was the total activity of cathepsin D in the lateral prostate different. These results were not consistent with parameters known as the effect mediated through androgen-controlled mechanisms. We therefore conclude that the action of prolactin in the regressing rat prostate is probably not mediated through the action of androgen.
Key words: prostate, prolactin, protein metabolism, cathepsin D, androgen, castration
INTRODUCTION
Hyperprolactinemia, produced by transplanting pituitary grafts under the renal capsule, retards the rate of loss of weight and protein following castration in the prostate of rats [ 13. This effect of prolactin is most pronounced in the lateral lobe of the prostate. Although the mechanism of this prolactin action in the prostate remains unclear, the probability that modifications of androgen-dependent anabolic or cata- bolic events are responsible seems likely to us and to other investigators [ 2 4 . The possibility that a minute amount of residual endogenous androgen present in the prostate at the time of castration acts synergistically with the elevated levels of prolactin to cause a delay in prostatic regression seems to warrant primary consider- ation. However, a previous study indicating that a potent nonsteriodal antiandrogen,
Presented at the Workshop on Prostate Cancer: A Decade of Progress and New Horizons held in Bethesda, Maryland, January 9-11, 1984.
Address reprint requests to Dr. Chung Lee, Department of Urology, Northwestern University Medical School, Chicago, IL 60611.
0 1985 Alan R. Liss, Inc.

50 Smith et al
Flutamide, failed to inhibit the effect of prolactin on this delay of castration-induced regression in the rat prostate [5] led us to consider the possibility that prolactin may act independent of testosterone or, at least, at a site distal to the binding of dihydro- testosterone (DHT) with androgen receptors. Further investigation to delineate the effect of prolactin on events following the formation of DHT-receptor complex in the rat prostate seems desirable.
If the action of prolactin in the regressing prostate is to enhance the effect of the minute amount of residual androgen in the tissue, many metabolic features that are characteristic of androgen stimulation in the rat prostate should be evident. These androgen-induced features in protein metabolism include (a) an increase in the rate of protein synthesis [6-81, (b) a decrease in the rate of protein degradation [8], and (c) a decrease in the activity of proteolytic enzyme, cathepsin D [9]. The present study was conducted to determine the effect of prolactin on these androgen-controlled parame- ters in the rat prostate.
MATERIALS AND METHODS Animals and Surgical Procedures
Sprague-Dawley rats purchased from Harlan Industries (Indianapolis, IN) were kept in large plastic cages with five animals per cage. They were housed in an air- conditioned room (23°C) with 14 hours of illumination per day (05:OO-19:OO) and fed Purina Rat Chow and tap water ad libitum. The basic surgical scheme was camed out according to the procedure described by Kolbusz and associates [ 11. Briefly, two pituitaries from female rats weightng 175-200 g were transplanted under the right renal capsule of male rats weighing 300-325 g. Seven days later, all animals were castrated and a unilateral nephrectomy was done. In half of the animals the left kidney (non-graft-bearing) was removed; in the other half the right kidney (graft-bearing) was removed. Since animals in the former group retained the pituitary grafts, they should have higher levels of circulating prolactin and their prostates should regress at a slower rate than those in the latter group [l].
Experiment 1: In vivo retention of 'H-leucine from the rat prostate: Estimating the apparent rate of protein degradation based on the rate of in vivo release of 3H-leucine by the single pulse technique has been characterized previously for the rat prostate [S]. Following a single pulse of 3H-leucine in male rat, the radioactivity present in the protein fraction of the prostate could be accounted for by 3H-leucine. Reutilization of incorporated 3H-leucine was found to be insignificant in the rat prostate. The same approach was used in the present study. A total of 35 male rats were used in this experiment. Six days after the pituitary grafting, each rat received a single intravenous injection of a sterile saline solution containing 3H-leucine with a concentration of 100 pCi per ml at a dose of 15 pCi per 100 g body weight (3H4,5- L-leucine, 56.5 Ci per m o l e , New England Nuclear, Boston).
Twenty-four hours later, bilateral orchiectomy and unilateral nephrectomy were camed out. At the same time, a group of 13 3H-leucine-injected animals were sacrificed to establish the initial activity in the tissue. The three lobes of the prostate (ventral, lateral, and dorsal) were dissected separately, according to the procedure of Jesik et a1 [lo]. The prostates were weighed and placed in 10% trichloroacetic acid (TCA) for protein purification. Eight days after the 3H-leucine pulse (or 7 days after orchiectomy and unilateral nephrectomy), the remaining 22 animals were sacrificed

Prolactin Action in Regressing Prostate 51
and three lobes of the prostate were collected for protein extraction. The amount of radioactivity remaining in the protein fraction of the prostate, expressed as cpm/ prostatic lobe, served as an indication of the rate of protein degradation. The greater amount of radioactivity remaining in the prostate the slower being the rate of degradation.
Experiment 2: In vitro incorporation of 3H-leucine by the rat prostate: The technique of in vitro incorporation of 3H-leucine by prostatic tissue pieces was employed as a means to assess apparent protein-synthesizing ability [8]. One group of six animals were sacrificed by decapitation on the day of castration (or 7 days after pituitary grafting) and two groups of four each were sacrificed 7 days postcastration. Three lobes of the prostate were dissected separately. Each lobe was placed in medium 199 (GIBCO, Grand Island, NY), which was prewarmed to 37"C, and was cut with a scalpel into 2 mm3 pieces. The tissue pieces were then incubated in 4 ml of Medium 199 containing 3H-leucine (0.5 pCi per ml) for 2 h under 95 % oxygen and 5 % carbon dioxide at 37°C in a shaker water bath as described by Lee [ 111 and Lee et al [ 121. The reaction was stopped by placing the tissue in 10% TCA. The tissue was processed for protein extraction and the amount of radioactivity present in the protein fraction, expressed as cpm/mg protein/hr, served as an indication of the rate of protein synthesis.
Experiment 3: In vivo incorporation of 3H-leucine by the rat pros- tate: Results of our preliminary study indicated that, following a single i.v. pulse of 3H-leucine in rats, the amount of radioactivity incorporated into the protein fraction of the prostate reached a peak within 24 h. The radioactivity thus incorporated by the prostate during this 24-h pulse period was used as another reference to indicate the protein-synthesizing ability of the tissue. The 13 animals used at 1 day before castration were the same animals used at the initial stage of Experiment 1. Six days following castration two additional groups of 13 rats each received a single i.v.injection of the same sterile saline solution containing 3H-leucine as described earlier. They were sacrificed 24 h after the 3H-leucine pulse and their prostates were removed for process as described. The amount of radioactivity present in the protein fraction was expressed as c p d m g protein.
Extraction of prostatic protein and determination of radioactivity: The prostatic tissue was homogenized in 10% TCA with a Polytron (model PT-10, Brinkman Instrument, Westbury, NY). The homogenate was heated to 90°C for 30 min and the resulting precipitate was washed twice with 5 ml of a mixture of ethano1:ether (2:l by volume) and thereafter once with 5 ml of ether. The washed precipitate was then dissolved in 2 ml of 0.4 N NaOH. Duplicated aliquots of 0.5 ml of this alkaline protein solution were counted with 10 ml of scintillation fluid (Dimil- ume, Packard Instrument Co, Downers Grove, IL) in a Beckman model 8100 liquid scintillation counter with a counting efficiency for tritium of 48% which was estab- lished by using a 3H-water standard under the above described conditions. Protein determination was performed on a portion of the remaining solution according to the procedure of Lowry et al [13]. Results were expressed either as cpm per mg protein or cpm per prostate lobe.
Experiment 4 Assay for cathepsin D activity in the prostate: One group of five rats was sacrificed on the day of castration-nephrectomy and two groups of ten rats each (one hyperprolactinemic group and one control group) were sacrificed 7 days later. Three lobes of the prostate were removed and stored in liquid nitrogen

52 Smithetal
freezer. Cathepsin D activity in the prostatic tissue was determined by the method of Anson [ 141 and of Ferguson and associates [ 151. The procedure was carried out with minor modifications [9,16]. Each prostatic lobe was allowed to thaw in the refrigera- tor and was homogenized in 3 ml of an ice-cold solution containing 1 % NaCl and 2% n-butanol using a glass-glass homogenizer. A volume of 10% Triton-X100 was then added to this suspension to bring the final concentration of Triton-X100 to 0.5%. This preparation was placed in the refrigerator overnight and was then centrifuged for 20 min at 10,OOO rpm (Sorvall RC-SB, with SE-12 Rotor). An aliquot of 0.2 ml of the Supernatant fraction was added to 2 ml of a hemoglobin buffer substrate which was prewarmed for 5 min in a 40°C water bath. The substrate consisted of a 4: 1 mixture of 0.83% bovine hemoglobin solution (twice crystallized and lyophilized, Sigma Chemical Co, St. Louis) and 1.35 M acetate buffer, pH 3.7. The incubation was carried out for 30 min at 40°C and the reaction was stopped by adding 2 ml of cold 10% TCA. For the zero-time tubes, the aliquot of 0.2 ml of the supernatant, instead of at the beginning of the incubation, was added to the hemoglobin substrate at the end of incubation after the reaction was stopped by 10% TCA. The incubated samples were placed in the refrigerator overnight and were then centrifuged at 3,000 rpm for 10 min. The clear supernatant was filtered through a 0.45-pm Millipore filter (type HA). The filtrate was measured in a spectrophotometer at the wavelength of 280 nm. A sample having its filtrate to exhibit an increase in absorbance of 0.01 above that of the zero-time tubes was considered of having one unit of cathepsin D. The protein content in the prostatic homogenate was measured following TCA precipitation and the activity of cathepsin D was expressed as units/mg protein.
Prolactin dererminurions: The levels of serum prolactin were measured by radioimmunoassay using the NIADDK rat prolactin kit, with NIADDK rat PRL-RP- 3 as the standard [ 171.
Data analysis: All numerical values were expresed as mean f standard error. The Student’s t-test was employed to detect differences in means of different param- eters between castrated animals bearing the pituitary grafts and those without the grafts. For the study of in vivo decay of the labeled protein in the prostate, the amount of radioactivity per prostate lobe (cpdprostate) was adjected for the prostatic wet weight in each group by a covariance analysis prior to statistical analysis [18].
RESULTS Serum Prolactin Levels
A total of 24 animals were sacrificed at 1 week following pituitary grafting. The average concentration of serum prolactin in these animals was 92.7 f 7.6 ngl ml. The remainder of the animals received orchiectomy and unilateral nephrectomy at that point. One week later, 38 animals bearing the pituitary grafts continued to have an elevated level of serum prolactin (93.8 f 6.8 ng/ml). The other 38 animals were without the grafts. The average serum prolactin at a level normally observed in nongrafted animals (26.2 f 3.4 ng/ml) was significantly lower (P < 0.01) than that of the graft-bearing animals.
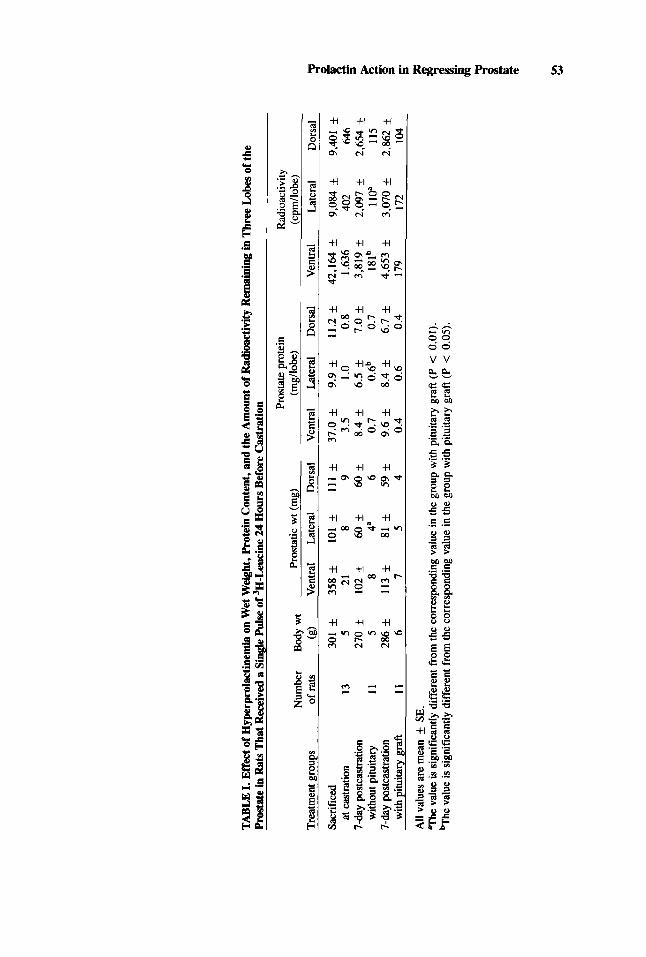
In Vivo Retention of 3H-Leucine in the Rat Prostate Table I shows that, at 7 days postcastration, rats with pituitary grafts had
significantly (P < 0.01) larger lateral lobes (both wet weight and protein content)
TABL
E I. Effect o
f Hyp
erpr
olac
tinem
ia on
Wet
Wei
ght,
Prot
ein
Con
tent
, and
the
Am
ount
of R
adio
activ
ity R
emai
ning
in T
hree
Lob
es of
the
Pros
tate
in Rats
Tha
t Rec
eive
d a
Sing
le P
ulse
of 3
H-L
euci
ne 24
Hou
rs B
efor
e Cas
trat
ion Pr
osta
te p
rote
in
Rad
ioac
tivity
N
umbe
r B
odyw
t Pr
osta
tic w
t (m
g)
(mgl
lobe
) (c
pm/lo
be)
7 Tr
eatm
ent g
roup
s of
rat
s (g
) V
entra
l La
tera
l D
orsa
l V
entra
l La
tera
l D
orsa
l V
entra
l La
tera
l D
orsa
l
Sacr
ifice
d 30
1 f
358 f
101 f
111 f
37.0
f
9.9 f
11.2
f
42,1
64 f
9,08
4 9,
401 *
at c
astra
tion
13
5 21
8
9 3.
5 1 .o
0.
8 1,
636
402
646
7-da
y po
stca
stra
tion
270 f
102 f
60 f
60 f
8.4 f
6.5 f
7.0 f
3,81
9 f
2,09
7 f
2,65
4 f
z * w
ithou
t pitu
itary
11
5
8 4a
6
0.7
0.6b
0.
7 18
Ib
110”
11
5 .,
7-day p
ostc
astra
tion
286 f
113 f
81 k
59 f
9.6 f
8.4
+ 6.
7 f
4,65
3 f
3,07
0 f
2,86
2 f
g w
ith p
ituita
ry g
raft
II
6 7
5 4
0.4
0.6
0.4
179
172
104
5 E 1 8
All
valu
es a
re m
ean f SE.
Th
e va
lue
is si
gnifi
cant
ly d
iffer
ent f
rom
the
corr
espo
ndin
g val
ue in
the
grou
p w
ith p
ituita
ry g
raft
(P <
0.0
1).
-he
valu
e is
sign
ifica
ntly
diff
eren
t fro
m th
e co
rres
pond
ing v
alue
in th
e gr
oup
with
pitu
itary
gra
ft (P
< 0
.05)
.
rp s:

54 Smithet al
than those without the grafts. The rate of decay of labeled protein as assessed by the radioactivity remaining in the lateral lobe following a single i.v. pulse of 3H-leucine given on the day before castration was slower in castrated rats bearing pituitary grafts than that of castrated rats without the grafts (P < 0.01; Table I). The radioactivity was significantly greater in the ventral lobes of rats with the pituitary grafts than in those without the grafts (P < 0.05), despite the absence of a significant difference in the wet weight and protein content of the ventral lobes of these two groups of rats. The wet weight, protein content, and radioactivity of the dorsal lobe were not significantly different in the graft-bearing and nongraft animals.
In Vitro and In Vivo Incorporation of 3H-Leucine by the Rat Prostate
Tables II and III show the in vitro and in vivo incorporation of 3H-leucine in the three lobes of the rat: prostate, respectively. Although the wet weight and protein content were significantly greater for the lateral lobe of the graft-bearing rats than those of the nongrafted rats, the amount of radioactivity incorporated by respective lobes was not significantly different (P > 0.05) in these two groups of animals. The parameters for the ventral and the dorsal lobes were not significantly different between the two groups (P > 0.05).
Cathepsin D Activity
Table IV indicates that, although the wet weight and the protein content in the lateral lobe in rats with pituitary graft were significantly greater (P < 0.01) than in those without the graft, no significant difference (P > 0.05) was noted in the activity of cathepsin D in these tissues between the two groups of rats. Parameters in the ventral and dorsal lobes were not significantly different (P > 0.05) between the two groups.
DISCUSSION
Results of the present study have confirmed and added to the observation that hyperprolactinemia achieved by pituitary grafts under the renal capsule causes a delay in the rate of castration-induced regression in the rat prostate, particularly in the lateral lobe [ 11. The demonstration that the delay in regression in the lateral prostate is characterized by a rate of protein synthesis normally observed in 7-day castrated rats and a lower than normal rate of protein degradation indicates that the catabolic phase of cellular metabolism is primarily affected by hyperprolactinemia. Since testosterone stimulation in the prostate is characterized both by an increase in protein synthesis and a decrease in protein degradation [8,19], the present observations demonstrate that the metabolic action of prolactin and testosterone in the rat prostate is different.
Results of the present study also indicate that, unlike the action of testosterone stimulation which leads to a decrease in the activity of cathepsin D in the prostate [9, 191, the action of prolactin has failed to alter this enzyme activity from a level normally seen in the prostate of 7-day castrated rats. Since both hormones cause a reduction in degradation of prostatic protein, the difference in enzyme activity in the tissue may reflect different sites of action. Our previous findings have indicated that administering exogenous testosterone to castrated rats resulted in a dose-related decrease in prostatic cathepsin D activity which bears an inverse relationship with the

TABL
E 11
. Effe
ct o
f Hyp
erpr
olac
tinem
ia o
n In
Vitr
o 3H
-Leu
cine
Inco
rpor
atio
n by
Thr
ee Lo
bes
of th
e R
at P
rost
ate
3H-le
ucin
e inc
orpo
ratio
n N
umbe
r w
t Pr
osta
tic w
t (m
g)
Prot
ein
cont
ent (
mg/
lobe
) (c
pmlm
g pr
otei
nlhr
) Tr
eatm
ent g
roup
s of
rat
s (g
) V
entra
l La
tera
l D
orsa
l V
entra
l La
tera
l D
orsa
l V
entra
l La
tera
l D
orsa
l
Sacr
ifice
d 35
1 f
412 f
131
f 13
1 f
40.2
k
15.0
f
14.1
f
6,87
7 f
4,22
3 f
3,59
1 *
3
7-da
y-po
stca
stra
tion
329 f
186 f
97
83 f
15.7
f
11.0
f
10.3
f
3,03
5 k
3,23
9 f
2,55
5 f
8 at
cas
tratio
n 6
7 13
8
14
1.1
0.8
0.7
263
175
487
E- 'I-
day
-pos
tcas
tratio
n 32
8 rt
143 f
53 f
75 f
10.9
f
6.6 i-
9.4 f
3,40
9 f
3,17
7 +
2,18
6 f
E w
ithou
t pitu
itary
4
I1
10
3=
9 I .
6 0.
3a
1.1
80
1 I9
279
* w
ith p
ituita
ry g
raft
4 7
11
11
7 I .
3 1.
7 0.
6 14
3 68
23
1 1
All
valu
es a
re m
ean f S
E.
Th
e va
lue
is si
gnifi
cant
ly d
iffer
ent f
rom
the
corr
espo
ndin
g va
lue
in th
e gr
oup
with
pitu
itary
gra
ft (P
< 0
.05)
.
01
01

TA
BL
E 11
1. Ef
fect
of
Hyp
erpr
olac
tine
mia
on
In V
ivo
'H-L
euci
ne
Inco
rpor
atio
n by
Thr
ee L
obes
of
the
Rat
Pro
stat
e
'H-le
ucin
e in
corp
orat
ion
Num
ber
Bod
ywt
Pros
tatic
wt
(mg)
Pr
otei
n co
nten
t (rn
g/lo
be)
(cpm
lmg
prot
ein)
Tr
eatm
ent g
roup
s of
rats
(g
) V
entra
l La
tera
l D
orsa
l V
entra
l La
tera
l D
orsa
l V
entra
l La
tera
l D
orsa
l
Sacr
ifice
d 30
1 f
358 f
101 f
Ill f
37.0
f
9.9 f
11.2
f
1.19
4 f
963 f
863 f
at c
astra
tion
13
5 21
8
9 3.
5 I .o
0.
8 69
59
60
7-
day
-pos
tcas
tratio
n 28
7 f
95 f
54 f
60 f
7.4 f
5.3 f
6.1 f
369 f
490 f
358 f
with
out p
ituita
ry
13
6 4
2"
5 0.
5 0.
4"
0.5
15
15
18
with
oitu
itarv
era
ft
13
7 9
3 4
0.7
0.3
0.5
23
20
29
7-da
y-po
stca
stra
tion
277 f
114 f
81 f
63 f
8.5 f
8.0 f
6.5 f
364 f
534 f
394 *
All
valu
es a
re m
ean f S
E.
The
grou
p of
ani
mal
s sac
rific
ed a
t cas
trat
ion
are
the
sam
e an
imal
s lis
ted
in T
able
11.
"The
val
ue is
sig
nific
antly
diff
eren
t fr
om c
orre
spon
ding
val
ue in
the
grou
p w
ith p
ituita
ry g
raft
(P
< 0
.01)
.

TABL
E IV.
Effe
ct o
f Hyp
erpr
olac
tinem
ia on
the
Act
ivity
of C
athe
psin
D in
Thr
ee L
obes
of
the
Rat
Pro
stat
e
Cat
heps
in D
leve
ls
Num
ber
Body w
t Pr
osta
tic w
t (m
g)
Prot
ein
cont
ent (
mgl
lobe
) (u
nits
hg pr
otei
n)
7 Tr
eatm
ent g
roup
s of
rats
(g
) V
entra
l La
tera
l D
orsa
l V
entra
l La
tera
l D
orsa
l V
entra
l La
tera
l D
orsa
l
Sacr
ifice
d 32
6 f
411 f
140 f
125 f
39.8
f
15.8
f
14.1
f
1.3 f
1.7 f
23 f
* 7d
ay-p
ostc
astr
atio
n 32
1 +
145 f
62 f
74 f
12.4
f
7.0
8.6 f
7.6
+ 17
.2 k
27 k
2
7-da
y-po
stca
stra
tion
325 f
163 f
100 f
96 f
13.9
f
11.5
f
10.9
f
6.9
15.6
f
26 f
5’ ,
5’ A
ll va
lues
are
mea
n SE.
aThe
val
ue is
sig
nific
antly
dif
fere
nt fr
om th
e co
rres
pond
ing
valu
e in
the
grou
p w
ith p
ituita
ry g
raft
(P
< 0
.01)
. %
he v
alue
is s
igni
fican
tly d
iffe
rent
from
the
corr
espo
ndin
g va
lue
in th
e gr
oup
with
pitu
itary
gra
ft (P
< 0
.05)
.
e at
cas
tratio
n 5
10
10
5 10
0.
5 1 .o
0.
9 0.
4 0.
6 3.
0 2
with
out p
ituita
ry
10
9 8
3”
4b
I .o
f 0
.5“
0.5a
1.
3 1.
8 3.
5
with
pitu
itary
gra
ft
10
6 13
5
6 1 .o
I .o
0.
6 1.
3 2.
9 1.
9
x i! 5 aa

58 Smith et al
magnitude of prostatic growth [9]. Furthermore, an inhibitor to protein synthesis, actinomycin D, can reduce the rate of prostatic regression in castrated rats [20], while at the same time it blocks the castration-induced increase in cathepsin D activity in the prostate [9]. These observations provide support to indicate that the reduction in prostatic regression and protein degradation by testosterone is, at least in part, due to its action to reduce the synthesis of prostatic cathepsin D. Our present findings demonstrate that the activity of cathepsin D is stable in the presence of hyperprolac- tinemia. This important difference in proiactin and testosterone effects on the rat prostate suggests differing mechanisms are involved.
The reduction in protein degradation in the prostate in the hyperprolactinemic rats in the presence of an unsuppressed activity of this proteolytic enzyme seems paradoxal. A similar partial delay of prostatic regression and an elevated activity of cathepsin D has been noted in castrated rats treated with chloroquine [16]. We have hypothesized that, being a lysosomal stabilizing agent, chloroquine somehow inter- fered with the normal interaction between the proteolytic enzymes and their cellular substrates in the prostate [19]. Although the effect of prolactin does not necessarily duplicate that of chloroquine, the possibility that hyperprolactinemia alters the inter- action between cathepsin D (and other proteolytic enzymes) and cellular proteins in the prostate is high. Under this circumstance, the synthesis of the proteolytic enzyme in the prostate would be increased as usual in 7day castrated rats but the synthesized enzyme somehow could not interact with its cellular substrate. As a consequence, the rate of protein degradation would be decreased.
Based on the above consideration, we conclude that, the action of prolactin on protein metabolism in the regressing prostate is likely to be independent of the action of testosterone. Testosterone stimulates protein synthesis and reduces protein degra- dation. Prolactin does not stimulate protein synthesis; it only reduces the rate of protein degradation. Furthermore, the manner in which the two hormones act on the prostate to reduce the rate of protein degradation is different. The testosterone- stimulated reduction in protein degradation is due to a reduction in synthesis of proteolytic enzymes; while prolactin interferes with the interaction of the existing degradative enzymes with their respective substrates.
ACKNOWLEDGMENTS
The authors wish to thank Julia Sensibar and Anthony Emerson for technical assistance; Brigittee G. Mann from the laboratory of Dr Neena B. Schwartz, North- western University, Evanston, Illinois, for her help on radioimmunoassay for prolac- tin; and the National Institute of Arthritis, Diabetes, Digestive Disease and Kidney for the supply of the kit for rat prolactin radioimmunoassay (rat-rat: RIA-NIADDK- rPRL-W-3).
Supported by NM grant HD11611, Edwin and Lucy Kretschmer Fund of Northwestern University Medical School, and a grant from the Grainger Foundation.
REFERENCES
1 . Kolbusz WE, Lee C. Grayhack JT: Delay of castration-induced regression in rat prostate by pituitary
2. Holland JM, Lee C: Effects of pituitary grafts on testosterone stimulated growth of rat prostate. homografts. J Urol 127:581-584, 1982.
Biol Reprod 22:351-355, 1980.

Prolactin Action in Regressing Prostate 59
3. Manandhar MSP, Thomas JA: Effect of prolactin on the metabolism of androgens by the rat ventral
4. Johansson R: Effect of prolactin, growth hormone and insulin in uptake and binding of dihydrotes-
5 . Assimos D, Smith C, Lee C, Grayhack JT: Failure of Flutamide to block the effect of prolactin on
6. Liao S, Tymoczko JL, Castaneda E, Liang T: Androgen receptors and androgen-dependent initiation
7. Parker MG, Scrace GT, Mainwaring WIP: Testosterone regulates the synthesis of major proteins in
8. Bockrath JM, Lee C, Grayhack JT: Incorporation and release of tritiated leucine in rat prostate
9. Tanabe ET, Lee C, Grayhack JT: Activities of cathepsin D in rat prostate during castration-induced
10. Jesik CJ, Holland JM, Lee C: An anatomical and histologic study of the rat prostate. The Prostate
11. Lee C: Uterine responsiveness to estrogen in 4-day cycling and in estrogen primed ovariectomized rats as assessed by uptake and incorporation of 3H-leucine. Endocrinology 95: 1754-1758, 1974.
12. Lee C, Murphy JJ, Diamond CA, Rafferty NS, Oyasu R: Alternate fluctuations of leucine and thymidine incorporation by mammary tumors in rats during the estrous cycle. Cancer Res 37:3301- 3305, 1977.
13. Lowry OH, Rosebrough NJ, Farr AL, Randall RJ: Protein measurement with folin phenol reagent. J Biol Chem 193:265-275, 1951.
14. Anson ML: The estimation of pepsin, trypsin, papain, and cathepsin with hemoglobin. J Gen Physiol22:79-89, 1938.
15. Ferguson JB, Andrews JR, Voynick IM, Fruton JS: The specificity of cathepsin D. J Biol Chem
16. Sharer WC, Lee C, Grayhack JT: Partial inhibition of castration-induced involution in rat prostate
17. Schwartz NB, Just0 SN: Acute changes in serum gonadotropins and steroids following orchidectomy
18. Steel RED, Torrie JH: “Principles and Procedures of Statistics.” New York: McGraw-Hill, 1960. 19. Lee C: Physiology of castration-induced regression of the rat prostate. In Murphy G, Sandberg AA,
Karr JP (eds): “The Prostatic Cell: Structure and Function, Part A.” New York: Alan R. Liss,
20. Stanisic T, Sadlowski R, Lee C, Grayhack JT: Partial inhibition of castration induced ventral
prostate gland in vitro. Invest Urol 14:20-22, 1976.
tosterone in the cultured rat ventral prostate. Acta Endocrinol 81:854-864, 1976.
delay of castration-induced regression in rat prostate. The Prostate (submitted).
of protein synthesis in the prostate. Vitam Horm 33:297-317, 1975.
rat ventral prostate. Biochem J 170:115-121, 1978.
during castration-induced involution. Invest Urol 19: 169- 175, 1981.
involution. J Urol 127:826-828, 1982.
3:81-97, 1982.
248~670 1-6708, 1973.
by chloroquine. A preliminary observation. Invest Urol 17: 135-137, 1979.
in the rat: Role of the adrenal gland. Endocrinology 100: 1550-1556, 1977.
1981, pp 145-159.
prostate regression with actinomycin D and cycloheximide. Invest Urol 16: 19-22, 1978.