membrane–cytoskeleton interactions
TRANSCRIPT

Vincent Das Membrane–cytoskeleton interactionsBeatrice Nal during the formation of theAnne Roumier
immunological synapse andVannary Meas-YedidChristophe Zimmer subsequent T-cell activationJean-Christophe Olivo-MarinPascal RouxPierre FerrierAlice Dautry-VarsatAndres Alcover
Authors’ address
Vincent Das1, Beatrice Nal1, Anne Roumier1, VannaryMeas-Yedid2, Christophe Zimmer2, Jean-Christophe Olivo-Marin2, Pascal Roux3, Pierre Ferrier4, Alice Dautry-Varsat1 and Andres Alcover1,1Unite de Biologie des Interactions Cellulaires,CNRS URA 1960, 2Unite d’Analyse d’ImagesQuantitative, CNRS URA 1947, 3Centred’Imagerie Dynamique, Institut Pasteur,Paris, France, 4Centre d’Immunologie deMarseille-Luminy, INSERM-CNRS-Universitede la Mediterranee, Marseilles France
Correspondence to:
Andres Alcover,Institut PasteurUnite de Biologie des Interactions Cellulaires28, rue du Dr Roux75724 Paris Cedex 15FranceTel: π33 1 40 61 30 64Fax: π33 1 40 61 32 38e-mail: [email protected]
AcknowledgmentsThe work described here was supported by aProgramme Transversal de Recherche from theInstitut Pasteur. V. Das is supported by anAllocation de Recherche du Ministere del’Education Nationale, de la Recherche et de laTechnologie. A. Roumier was supported by aBourse from La Ligue Contre le Cancer. B. Nal issupported by a Bourse from the Associationpour la Recherche sur le Cancer. C. Zimmer issupported by the Programme Transversal deRecherche from the Institut Pasteur. The experttechnical help of Annick Dujeancourt is thankfullyacknowledged.
Immunological Reviews 2002Vol 189: 123–135Printed in Denmark. All rights reserved
Copyright c Blackwell Munksgaard 2002
Immunological Reviews0105-2896
123
Summary: Upon antigen recognition, T cells undergo substantial mem-brane and cytoskeletal rearrangements that lead to the formation of theimmunological synapse and are necessary for subsequent T-cell activation.However, little is known about how membrane and cytoskeletal mol-ecules interact during these processes. Here we discuss the involvementof the membrane-microfilament linker ezrin. We propose that ezrin is acomponent of the cytoskeleton-mediated architecture of the immunolog-ical synapse that plays a role in T-cell receptor clustering, protein kinaseC q translocation and intracellular signaling.
Introduction
Antigen recognition and subsequent T-cell activation depend
on the appropriate interaction between T cells and antigen
presenting cells (APC). This interaction involves a series of
events that depend on membrane and cytoskeleton dynamics,
namely cell motility, cell–cell adhesion, polarization and re-
ceptor relocalization. Initial T-cell receptor (TCR) signaling
induces actin cytoskeleton rearrangements, which in turn are
necessary for the stability of T cell–APC interactions, for the
maturation of the immunological synapse and for sustained
T-cell signaling. During maturation of the immunological
synapse, a precise molecular reorganization occurs at the con-
tact zone between T cells and APCs. Thus, TCR, coreceptors,
intracellular signaling molecules and adhesion receptors po-
larize to the T cell–APC interface and segregate into distinct
supramolecular clusters that redistribute following a precise
relative topology. This molecular patterning needs a func-
tional actin and myosin cytoskeleton and agonistic TCR stimu-
lation (reviewed in 1–3).
An important question in the field is how plasma mem-
brane–cytoskeleton interactions take place and facilitate the
formation of the immunological synapse. The first molecule
found to be likely involved in these membrane–cytoskeleton

Das et al ¡ Membrane-cytoskeleton links in the IS
interactions was talin (4). This protein can interact with ad-
hesion molecules of the integrin family and with cytoskeletal
components (5), and was found to accumulate in the periph-
eral zone of the immunological synapse (6). Moreover,
CD2AP could also be involved in membrane–cytoskeleton in-
teractions by linking the CD2 molecule with the cytoskeleton
(7). More recently, several laboratories including ours showed
that the membrane-microfilament linkers ezrin and moesin
were involved in the organization of membrane components
at the T cell–APC contact zone (8–12). Previously, ezrin and
moesin were shown to be substrates of protein kinases in T
lymphocytes (13–15), suggesting that they are effectors of
TCR signaling. Interestingly, talin, ezrin and moesin belong
to the same large family of proteins, whose prototype is the
band 4.1 of red blood cells.
Ezrin, radixin and moesin are highly homologous polypep-
tides that form, together with merlin/schwannomin, the
ERM (Ezrin/Radixin/Moesin) family of proteins. These pro-
teins control cell shape, cytokinesis and cell adhesion in vari-
ous cell types including lymphocytes. ERMs can mediate the
anchoring of some transmembrane proteins to the actin cyto-
skeleton, either directly or through adapter molecules, such
as EBP50. In addition, ERMs are involved in intracellular sig-
naling (reviewed in 16, 17). As all members of the band 4.1
family, ERMs share a common homologous domain of about
300 amino acids, the FERM domain, which is located at the
N-terminal part of the protein. ERMs interact with the plasma
membrane through their FERM domain, and with the actin
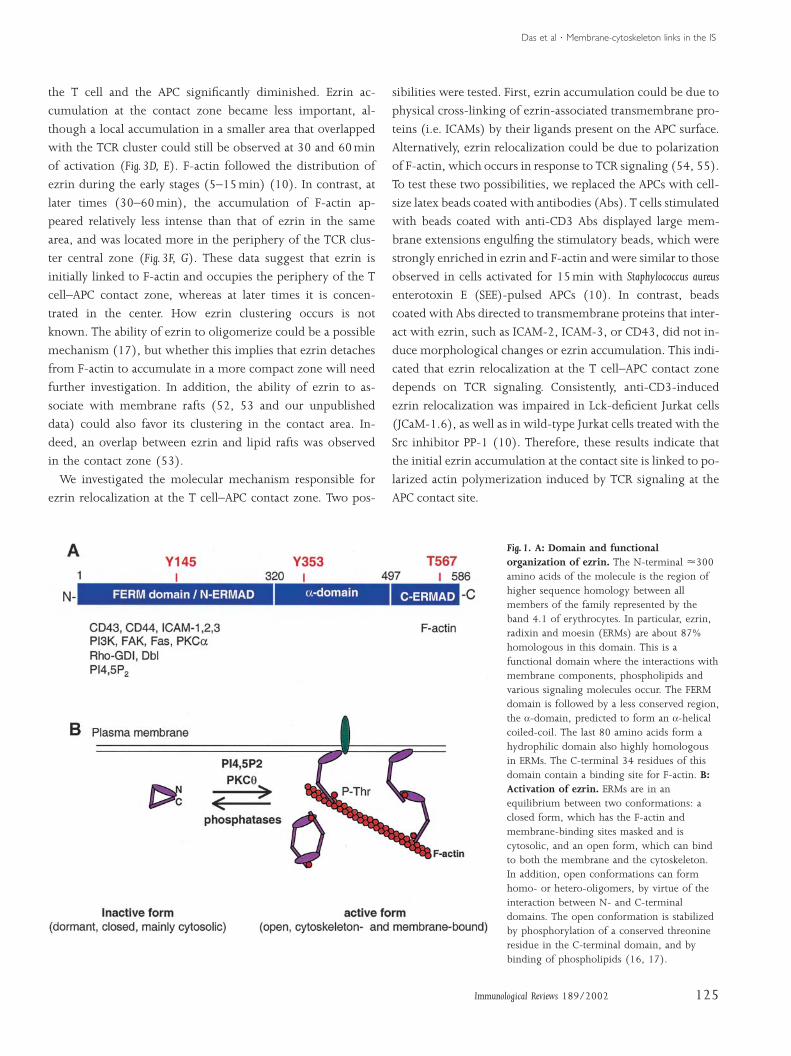
cytoskeleton through their C-terminal domain (18) (Fig. 1A).
These proteins exist under two conformations: a dormant
state, which is folded due to the interaction between the N-
terminal domain and the C-terminal domain (19), and an
active state, which is unfolded and able to interact with the
plasma membrane and the actin-based cytoskeleton (Fig. 1B).
The open conformation is stabilized by the phosphorylation
of a conserved threonine residue in the C-terminal domain
and the presence of phosphatidylinositides (20–25). Protein
kinase C (PKC) q can phosphorylate this threonine residue in
T cells (15), suggesting that ERMs can be activated during T-
cell activation.
The FERM domain can interact with transmembrane pro-
teins, such as intercellular adhesion molecules (ICAMs),
CD43, CD44, or P-selectin glycoprotein ligand 1 (PSGL-1)
in various cell types including lymphocytes (26–30). This is
particularly striking in T cells polarized by chemokines, in
which ERMs and these adhesion receptors strongly accumu-
late in membrane protrusions called uropods. These are
membrane protrusions, budding at the rear end of migrating
124 Immunological Reviews 189/2002
cells, which are involved in cell–cell adhesion. The localiz-
ation of these transmembrane proteins in the uropod depend
on their interaction with ERMs and ensure the cell adhesion
properties of this structure (28–32).
The N-terminal domain of ERMs can also interact with
phosphatidylinositol 4,5-bisphosphate (PIP2). This enhances
the capacity of these proteins to interact with the plasma
membrane, as well as with transmembrane proteins (26–28,
33, 34). Moreover, the N-terminal domain of ERMs can inter-
act with components of various intracellular signaling cas-
cades, such as RhoGDI (35), the regulatory subunit of phos-
phatidylinositol 3-kinase (36), the receptor Fas (37), PKCa
(38), or focal adhesion kinase (FAK) (39). ERM proteins may
also play a role in actin assembly on phagosomal membranes
(40). Therefore, ERMs are multifunctional proteins involved
in cellular architecture, intracellular signaling and membrane
trafficking.
Polarization of the actin cytoskeleton upon T-cell antigenrecognition: involvement of ezrin
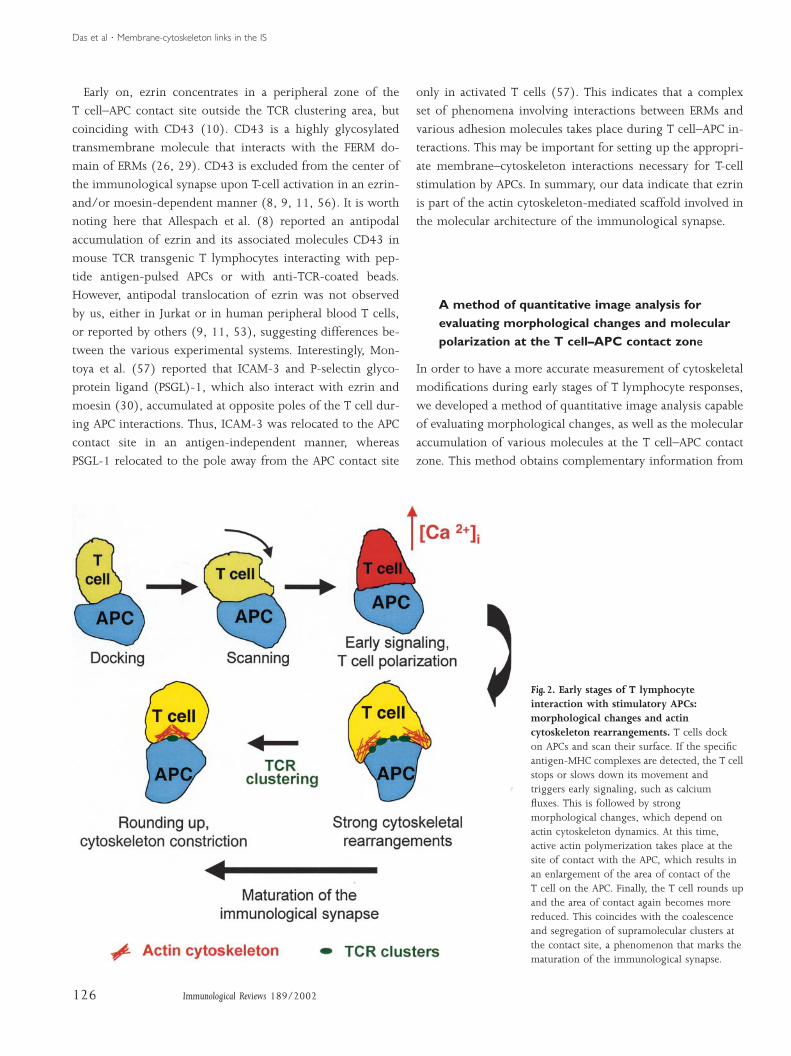
Antigen recognition requires an intimate contact between T
cells and APCs. Imaging T-cell antigen recognition in vitro and
in situ revealed that the interaction of T cells with APCs occurs
in a polarized manner. After docking on the APC, T cells ac-
tively crawl on the surface of the APC, and can move from one
APC to another. At the contact site, T cells develop membrane
extensions that contact and scan the APC surface. If a specific
antigen is detected, T cells stop or slow down their move-
ment, trigger intracellular calcium fluxes and progressively
change their shape to increase the area of contact with the
APC. Later on, the cell progressively rounds up again and the
contact surface between both cells diminishes. This coincides
with molecular segregation at the contact zone, which marks
the maturation of the immunological synapse (Fig. 2). Polar-
ization, motility, morphological changes, and molecular seg-
regation taking place in T cells interacting with APCs are sen-
sitive to early signaling and depend on actin and myosin cyto-
skeleton dynamics (6, 7, 41–51).
We investigated the involvement of the ERM family of
membrane-microfilament linkers in actin cytoskeleton re-
arrangements occurring at the T cell–APC contact zone
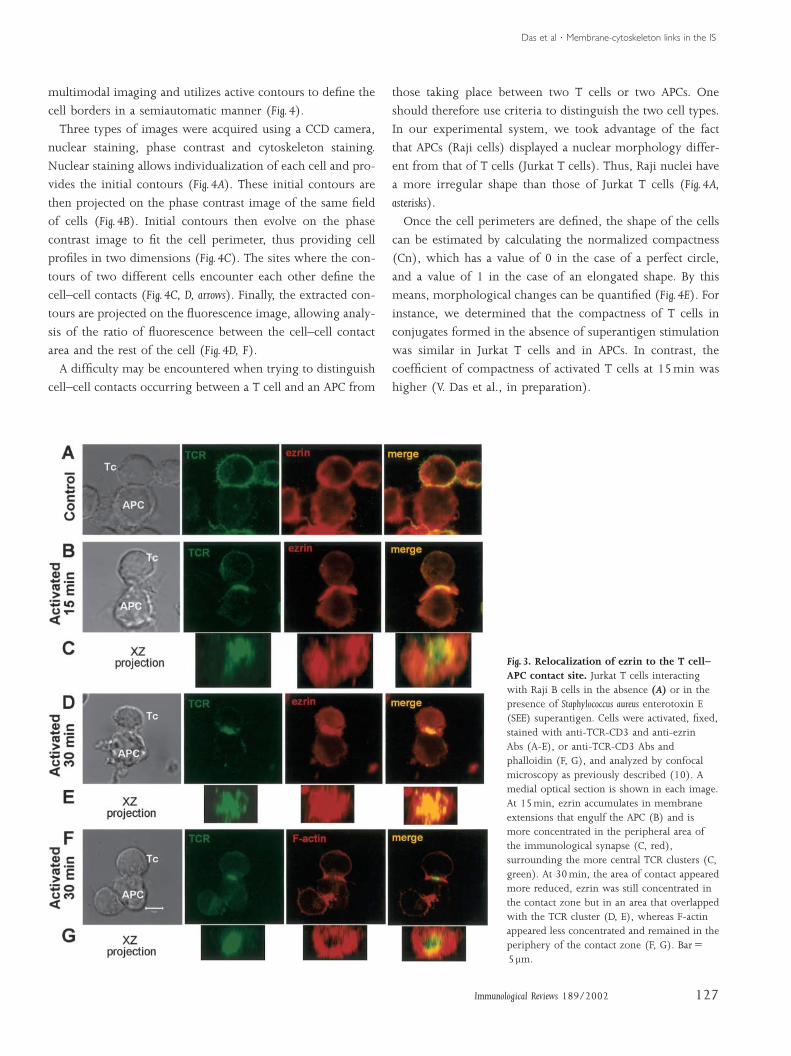
(Fig. 3A). We observed ezrin accumulated at the T cell–APC
contact zone and concentrated in F-actin-rich membrane ex-
tensions that engulfed the APC (Fig. 3B, C) (10). Moesin was
also enriched in these membrane extensions (A. Alcover, un-
published data). Between 15 and 30min of activation, these
membrane extensions retracted and the contact area between
Das et al ¡ Membrane-cytoskeleton links in the IS
the T cell and the APC significantly diminished. Ezrin ac-
cumulation at the contact zone became less important, al-
though a local accumulation in a smaller area that overlapped
with the TCR cluster could still be observed at 30 and 60min
of activation (Fig. 3D, E). F-actin followed the distribution of
ezrin during the early stages (5–15min) (10). In contrast, at
later times (30–60min), the accumulation of F-actin ap-
peared relatively less intense than that of ezrin in the same
area, and was located more in the periphery of the TCR clus-
ter central zone (Fig. 3F, G). These data suggest that ezrin is
initially linked to F-actin and occupies the periphery of the T
cell–APC contact zone, whereas at later times it is concen-
trated in the center. How ezrin clustering occurs is not
known. The ability of ezrin to oligomerize could be a possible
mechanism (17), but whether this implies that ezrin detaches
from F-actin to accumulate in a more compact zone will need
further investigation. In addition, the ability of ezrin to as-
sociate with membrane rafts (52, 53 and our unpublished
data) could also favor its clustering in the contact area. In-
deed, an overlap between ezrin and lipid rafts was observed
in the contact zone (53).
We investigated the molecular mechanism responsible for
ezrin relocalization at the T cell–APC contact zone. Two pos-
Fig. 1. A: Domain and functionalorganization of ezrin. The N-terminal 300amino acids of the molecule is the region ofhigher sequence homology between allmembers of the family represented by theband 4.1 of erythrocytes. In particular, ezrin,radixin and moesin (ERMs) are about 87%homologous in this domain. This is afunctional domain where the interactions withmembrane components, phospholipids andvarious signaling molecules occur. The FERMdomain is followed by a less conserved region,the a-domain, predicted to form an a-helicalcoiled-coil. The last 80 amino acids form ahydrophilic domain also highly homologousin ERMs. The C-terminal 34 residues of thisdomain contain a binding site for F-actin. B:Activation of ezrin. ERMs are in anequilibrium between two conformations: aclosed form, which has the F-actin andmembrane-binding sites masked and iscytosolic, and an open form, which can bindto both the membrane and the cytoskeleton.In addition, open conformations can formhomo- or hetero-oligomers, by virtue of theinteraction between N- and C-terminaldomains. The open conformation is stabilizedby phosphorylation of a conserved threonineresidue in the C-terminal domain, and bybinding of phospholipids (16, 17).
125Immunological Reviews 189/2002
sibilities were tested. First, ezrin accumulation could be due to
physical cross-linking of ezrin-associated transmembrane pro-
teins (i.e. ICAMs) by their ligands present on the APC surface.
Alternatively, ezrin relocalization could be due to polarization
of F-actin, which occurs in response to TCR signaling (54, 55).
To test these two possibilities, we replaced the APCs with cell-
size latex beads coated with antibodies (Abs). T cells stimulated
with beads coated with anti-CD3 Abs displayed large mem-
brane extensions engulfing the stimulatory beads, which were
strongly enriched in ezrin and F-actin and were similar to those
observed in cells activated for 15min with Staphylococcus aureus
enterotoxin E (SEE)-pulsed APCs (10). In contrast, beads
coated with Abs directed to transmembrane proteins that inter-
act with ezrin, such as ICAM-2, ICAM-3, or CD43, did not in-
duce morphological changes or ezrin accumulation. This indi-
cated that ezrin relocalization at the T cell–APC contact zone
depends on TCR signaling. Consistently, anti-CD3-induced
ezrin relocalization was impaired in Lck-deficient Jurkat cells
(JCaM-1.6), as well as in wild-type Jurkat cells treated with the
Src inhibitor PP-1 (10). Therefore, these results indicate that
the initial ezrin accumulation at the contact site is linked to po-
larized actin polymerization induced by TCR signaling at the
APC contact site.
Das et al ¡ Membrane-cytoskeleton links in the IS
Early on, ezrin concentrates in a peripheral zone of the
T cell–APC contact site outside the TCR clustering area, but
coinciding with CD43 (10). CD43 is a highly glycosylated
transmembrane molecule that interacts with the FERM do-
main of ERMs (26, 29). CD43 is excluded from the center of
the immunological synapse upon T-cell activation in an ezrin-
and/or moesin-dependent manner (8, 9, 11, 56). It is worth
noting here that Allespach et al. (8) reported an antipodal
accumulation of ezrin and its associated molecules CD43 in
mouse TCR transgenic T lymphocytes interacting with pep-
tide antigen-pulsed APCs or with anti-TCR-coated beads.
However, antipodal translocation of ezrin was not observed
by us, either in Jurkat or in human peripheral blood T cells,
or reported by others (9, 11, 53), suggesting differences be-
tween the various experimental systems. Interestingly, Mon-
toya et al. (57) reported that ICAM-3 and P-selectin glyco-
protein ligand (PSGL)-1, which also interact with ezrin and
moesin (30), accumulated at opposite poles of the T cell dur-
ing APC interactions. Thus, ICAM-3 was relocated to the APC
contact site in an antigen-independent manner, whereas
PSGL-1 relocated to the pole away from the APC contact site
Fig. 2. Early stages of T lymphocyteinteraction with stimulatory APCs:morphological changes and actincytoskeleton rearrangements. T cells dockon APCs and scan their surface. If the specificantigen-MHC complexes are detected, the T cellstops or slows down its movement andtriggers early signaling, such as calciumfluxes. This is followed by strongmorphological changes, which depend onactin cytoskeleton dynamics. At this time,active actin polymerization takes place at thesite of contact with the APC, which results inan enlargement of the area of contact of theT cell on the APC. Finally, the T cell rounds upand the area of contact again becomes morereduced. This coincides with the coalescenceand segregation of supramolecular clusters atthe contact site, a phenomenon that marks thematuration of the immunological synapse.
126 Immunological Reviews 189/2002
only in activated T cells (57). This indicates that a complex
set of phenomena involving interactions between ERMs and
various adhesion molecules takes place during T cell–APC in-
teractions. This may be important for setting up the appropri-
ate membrane–cytoskeleton interactions necessary for T-cell
stimulation by APCs. In summary, our data indicate that ezrin
is part of the actin cytoskeleton-mediated scaffold involved in
the molecular architecture of the immunological synapse.
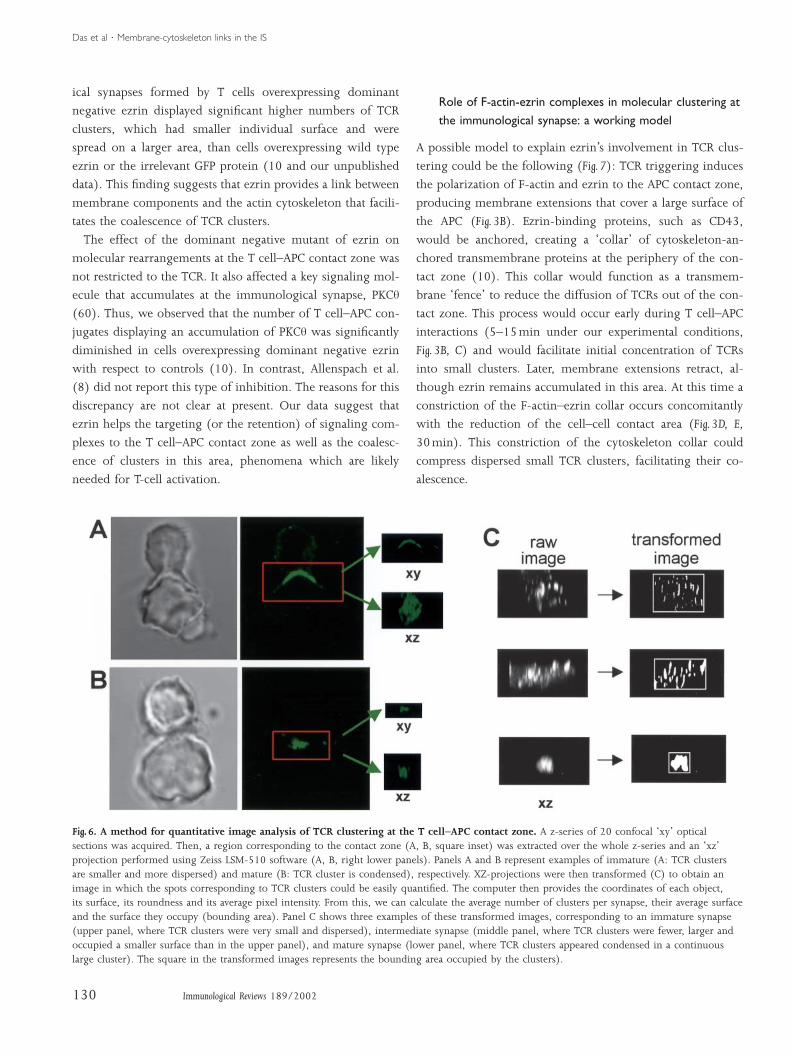
A method of quantitative image analysis forevaluating morphological changes and molecularpolarization at the T cell–APC contact zone
In order to have a more accurate measurement of cytoskeletal
modifications during early stages of T lymphocyte responses,
we developed a method of quantitative image analysis capable
of evaluating morphological changes, as well as the molecular
accumulation of various molecules at the T cell–APC contact
zone. This method obtains complementary information from
Das et al ¡ Membrane-cytoskeleton links in the IS
multimodal imaging and utilizes active contours to define the
cell borders in a semiautomatic manner (Fig. 4).
Three types of images were acquired using a CCD camera,
nuclear staining, phase contrast and cytoskeleton staining.
Nuclear staining allows individualization of each cell and pro-
vides the initial contours (Fig. 4A). These initial contours are
then projected on the phase contrast image of the same field
of cells (Fig. 4B). Initial contours then evolve on the phase
contrast image to fit the cell perimeter, thus providing cell
profiles in two dimensions (Fig. 4C). The sites where the con-
tours of two different cells encounter each other define the
cell–cell contacts (Fig. 4C, D, arrows). Finally, the extracted con-
tours are projected on the fluorescence image, allowing analy-
sis of the ratio of fluorescence between the cell–cell contact
area and the rest of the cell (Fig. 4D, F).
A difficulty may be encountered when trying to distinguish
cell–cell contacts occurring between a T cell and an APC from
Fig. 3. Relocalization of ezrin to the T cell–APC contact site. Jurkat T cells interactingwith Raji B cells in the absence (A) or in thepresence of Staphylococcus aureus enterotoxin E(SEE) superantigen. Cells were activated, fixed,stained with anti-TCR-CD3 and anti-ezrinAbs (A-E), or anti-TCR-CD3 Abs andphalloidin (F, G), and analyzed by confocalmicroscopy as previously described (10). Amedial optical section is shown in each image.At 15min, ezrin accumulates in membraneextensions that engulf the APC (B) and ismore concentrated in the peripheral area ofthe immunological synapse (C, red),surrounding the more central TCR clusters (C,green). At 30min, the area of contact appearedmore reduced, ezrin was still concentrated inthe contact zone but in an area that overlappedwith the TCR cluster (D, E), whereas F-actinappeared less concentrated and remained in theperiphery of the contact zone (F, G). BarΩ5 mm.
127Immunological Reviews 189/2002
those taking place between two T cells or two APCs. One
should therefore use criteria to distinguish the two cell types.
In our experimental system, we took advantage of the fact
that APCs (Raji cells) displayed a nuclear morphology differ-
ent from that of T cells (Jurkat T cells). Thus, Raji nuclei have
a more irregular shape than those of Jurkat T cells (Fig. 4A,
asterisks).
Once the cell perimeters are defined, the shape of the cells
can be estimated by calculating the normalized compactness
(Cn), which has a value of 0 in the case of a perfect circle,
and a value of 1 in the case of an elongated shape. By this
means, morphological changes can be quantified (Fig. 4E). For
instance, we determined that the compactness of T cells in
conjugates formed in the absence of superantigen stimulation
was similar in Jurkat T cells and in APCs. In contrast, the
coefficient of compactness of activated T cells at 15min was
higher (V. Das et al., in preparation).

Das et al ¡ Membrane-cytoskeleton links in the IS
After determining the intersection of cell contours, as de-
scribed above, we quantified the number of pixels having
values above background levels within the contact zone, or
outside this area. We could then calculate a ‘relocalization
index’ R (Fig. 4F). For instance, when we measured the ac-
cumulation of F-actin at the T cell–APC junctions, we calcu-
lated that activated T cells at 15min had a value of R 0.9,
whereas for non-activated cells, its value was R 0.4 (V. Das
et al., in preparation). Although this method still requires sys-
tematization to analyze large numbers of samples, it already
provides more precise data on cell shape and cytoskeletal re-
modeling during T cell–APC interactions.
Fig. 4. New method for quantitative analysisof morphological changes and molecularpolarization at the T cell–APC contact zone.This method obtains complementaryinformation from multimodal imaging andutilizes active contours to define the cellborders in a semiautomatic manner. Threetypes of images acquired using a CCD camerawere used in this example: nuclear staining,phase contrast and cytoskeleton staining.Nuclear staining allows individualization ofeach cell and provides the initial contours (A),which are then projected on the phase contrastimage of the same field of cells (B). Initialcontours then evolve on the phase contrastimage to fit the cell perimeter, thus providingcell profiles in two dimensions (C). The siteswhere the contours of two different cellsencounter each other define the cell–cellcontacts (C, D, arrows). Finally, the extractedcontours are projected on the fluorescenceimage (D, F). From the contours, the shapeof the cell can be estimated by calculating thenormalized compactness (Cn), according to theformula, where A is the area and P theperimeter. Cn has a value of 0 in the case ofa perfect circle, and a value of 1 in the case ofan elongated shape. By these means,morphological changes can be quantified. Anexample of Cn calculated for an activated T cell(E, left), vs. a control T cell (E, right) isshown. After determining the intersection ofcell contours, the molecular polarization couldbe estimated. The number of pixels havingvalues above background were quantifiedwithin the contact zone (Njunction), or outsidethis area (NC
junction). A ‘relocalization index’ Rwas calculated according to the formula (F,bottom).
128 Immunological Reviews 189/2002
A role for ezrin in TCR-CD3 clustering at theimmunological synapse: quantitative imaging analysis
Molecular polarization and subsequent segregation into dis-
tinct clusters in the immunological synapse require an intact
actin-myosin cytoskeleton (6, 45, 46). Ezrin is known to link
the membrane with the actin cytoskeleton and could there-
fore facilitate molecular polarization and/or clustering in the
T cell–APC contact zone. To investigate this possibility, we
studied the effect of a dominant negative mutant of ezrin.
This mutant contains only the FERM domain, which interacts
with the membrane but lacks the main F-actin-binding re-
Das et al ¡ Membrane-cytoskeleton links in the IS
gion (18). It has previously been shown to inhibit morpho-
genesis and migration of epithelial cells (58). We observed
that overexpression of this ezrin mutant inhibited the coalesc-
ence of dispersed TCR clusters into more compact ones, a
characteristic of the immunological synapse in mature
lymphocytes (6, 46).
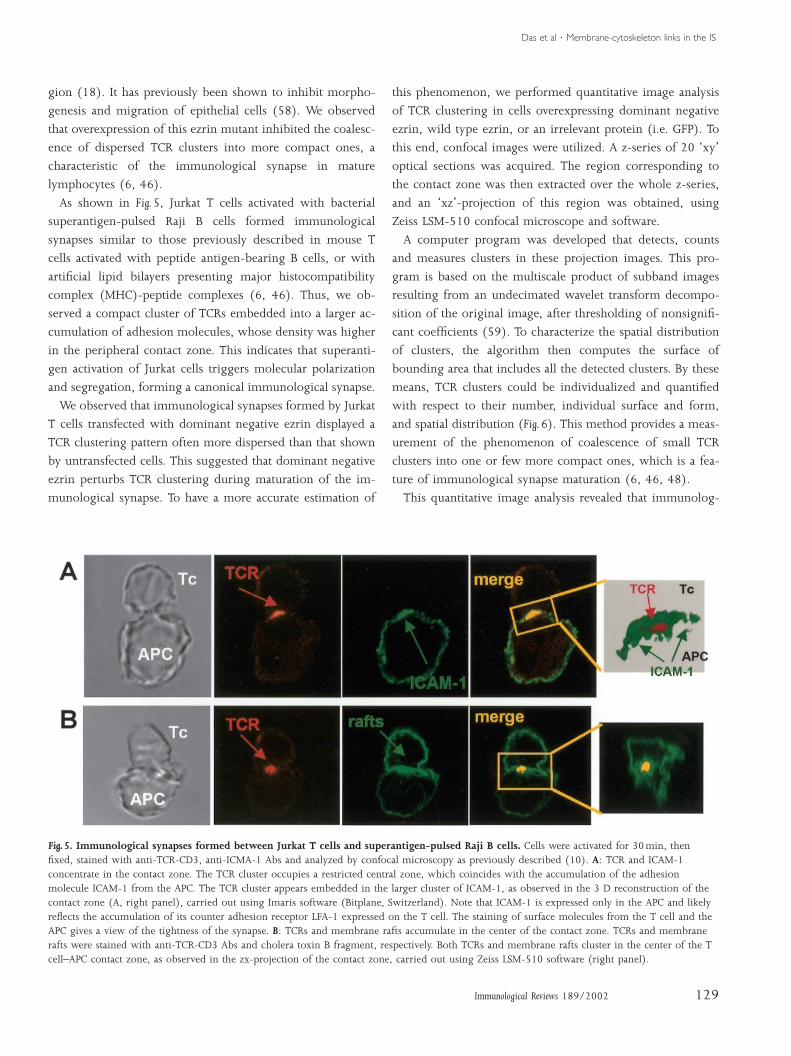
As shown in Fig. 5, Jurkat T cells activated with bacterial
superantigen-pulsed Raji B cells formed immunological
synapses similar to those previously described in mouse T
cells activated with peptide antigen-bearing B cells, or with
artificial lipid bilayers presenting major histocompatibility
complex (MHC)-peptide complexes (6, 46). Thus, we ob-
served a compact cluster of TCRs embedded into a larger ac-
cumulation of adhesion molecules, whose density was higher
in the peripheral contact zone. This indicates that superanti-
gen activation of Jurkat cells triggers molecular polarization
and segregation, forming a canonical immunological synapse.
We observed that immunological synapses formed by Jurkat
T cells transfected with dominant negative ezrin displayed a
TCR clustering pattern often more dispersed than that shown
by untransfected cells. This suggested that dominant negative
ezrin perturbs TCR clustering during maturation of the im-
munological synapse. To have a more accurate estimation of
Fig. 5. Immunological synapses formed between Jurkat T cells and superantigen-pulsed Raji B cells. Cells were activated for 30min, thenfixed, stained with anti-TCR-CD3, anti-ICMA-1 Abs and analyzed by confocal microscopy as previously described (10). A: TCR and ICAM-1concentrate in the contact zone. The TCR cluster occupies a restricted central zone, which coincides with the accumulation of the adhesionmolecule ICAM-1 from the APC. The TCR cluster appears embedded in the larger cluster of ICAM-1, as observed in the 3 D reconstruction of thecontact zone (A, right panel), carried out using Imaris software (Bitplane, Switzerland). Note that ICAM-1 is expressed only in the APC and likelyreflects the accumulation of its counter adhesion receptor LFA-1 expressed on the T cell. The staining of surface molecules from the T cell and theAPC gives a view of the tightness of the synapse. B: TCRs and membrane rafts accumulate in the center of the contact zone. TCRs and membranerafts were stained with anti-TCR-CD3 Abs and cholera toxin B fragment, respectively. Both TCRs and membrane rafts cluster in the center of the Tcell–APC contact zone, as observed in the zx-projection of the contact zone, carried out using Zeiss LSM-510 software (right panel).
129Immunological Reviews 189/2002
this phenomenon, we performed quantitative image analysis
of TCR clustering in cells overexpressing dominant negative
ezrin, wild type ezrin, or an irrelevant protein (i.e. GFP). To
this end, confocal images were utilized. A z-series of 20 ‘xy’
optical sections was acquired. The region corresponding to
the contact zone was then extracted over the whole z-series,
and an ‘xz’-projection of this region was obtained, using
Zeiss LSM-510 confocal microscope and software.
A computer program was developed that detects, counts
and measures clusters in these projection images. This pro-
gram is based on the multiscale product of subband images
resulting from an undecimated wavelet transform decompo-
sition of the original image, after thresholding of nonsignifi-
cant coefficients (59). To characterize the spatial distribution
of clusters, the algorithm then computes the surface of
bounding area that includes all the detected clusters. By these
means, TCR clusters could be individualized and quantified
with respect to their number, individual surface and form,
and spatial distribution (Fig. 6). This method provides a meas-
urement of the phenomenon of coalescence of small TCR
clusters into one or few more compact ones, which is a fea-
ture of immunological synapse maturation (6, 46, 48).
This quantitative image analysis revealed that immunolog-
Das et al ¡ Membrane-cytoskeleton links in the IS
ical synapses formed by T cells overexpressing dominant
negative ezrin displayed significant higher numbers of TCR
clusters, which had smaller individual surface and were
spread on a larger area, than cells overexpressing wild type
ezrin or the irrelevant GFP protein (10 and our unpublished
data). This finding suggests that ezrin provides a link between
membrane components and the actin cytoskeleton that facili-
tates the coalescence of TCR clusters.
The effect of the dominant negative mutant of ezrin on
molecular rearrangements at the T cell–APC contact zone was
not restricted to the TCR. It also affected a key signaling mol-
ecule that accumulates at the immunological synapse, PKCq
(60). Thus, we observed that the number of T cell–APC con-
jugates displaying an accumulation of PKCq was significantly
diminished in cells overexpressing dominant negative ezrin
with respect to controls (10). In contrast, Allenspach et al.
(8) did not report this type of inhibition. The reasons for this
discrepancy are not clear at present. Our data suggest that
ezrin helps the targeting (or the retention) of signaling com-
plexes to the T cell–APC contact zone as well as the coalesc-
ence of clusters in this area, phenomena which are likely
needed for T-cell activation.
Fig. 6. A method for quantitative image analysis of TCR clustering at the T cell–APC contact zone. A z-series of 20 confocal ‘xy’ opticalsections was acquired. Then, a region corresponding to the contact zone (A, B, square inset) was extracted over the whole z-series and an ‘xz’projection performed using Zeiss LSM-510 software (A, B, right lower panels). Panels A and B represent examples of immature (A: TCR clustersare smaller and more dispersed) and mature (B: TCR cluster is condensed), respectively. XZ-projections were then transformed (C) to obtain animage in which the spots corresponding to TCR clusters could be easily quantified. The computer then provides the coordinates of each object,its surface, its roundness and its average pixel intensity. From this, we can calculate the average number of clusters per synapse, their average surfaceand the surface they occupy (bounding area). Panel C shows three examples of these transformed images, corresponding to an immature synapse(upper panel, where TCR clusters were very small and dispersed), intermediate synapse (middle panel, where TCR clusters were fewer, larger andoccupied a smaller surface than in the upper panel), and mature synapse (lower panel, where TCR clusters appeared condensed in a continuouslarge cluster). The square in the transformed images represents the bounding area occupied by the clusters).
130 Immunological Reviews 189/2002
Role of F-actin-ezrin complexes in molecular clustering atthe immunological synapse: a working model
A possible model to explain ezrin’s involvement in TCR clus-
tering could be the following (Fig. 7): TCR triggering induces
the polarization of F-actin and ezrin to the APC contact zone,
producing membrane extensions that cover a large surface of
the APC (Fig. 3B). Ezrin-binding proteins, such as CD43,
would be anchored, creating a ‘collar’ of cytoskeleton-an-
chored transmembrane proteins at the periphery of the con-
tact zone (10). This collar would function as a transmem-
brane ‘fence’ to reduce the diffusion of TCRs out of the con-
tact zone. This process would occur early during T cell–APC
interactions (5–15min under our experimental conditions,
Fig. 3B, C) and would facilitate initial concentration of TCRs
into small clusters. Later, membrane extensions retract, al-
though ezrin remains accumulated in this area. At this time a
constriction of the F-actin–ezrin collar occurs concomitantly
with the reduction of the cell–cell contact area (Fig. 3D, E,
30min). This constriction of the cytoskeleton collar could
compress dispersed small TCR clusters, facilitating their co-
alescence.

Das et al ¡ Membrane-cytoskeleton links in the IS
The TCR clustering observed may be part of a more general
phenomenon involving cholesterol and sphingolipid-enrich-
ed membrane microdomains, also called membrane rafts
(61). In T cells, membrane rafts contain TCRs and signaling
molecules (62, 63), and concentrate at the T cell–APC contact
zone (64) (Fig. 5B) upon activation. Interestingly, ERMs can
interact with membrane rafts, as assessed by sucrose gradient
buoyancy (53 and our unpublished data). This may facilitate
raft coalescence in this zone.
According to this model, a crucial step for cluster coalescence
would be the constriction of the F-actin–ezrin network and the
concomitant reduction of the T cell–APC contact area, which
we observed between 15 and 30min (Figs 3 and 7). This con-
striction could occur by shifting from a state of high actin poly-
merization dynamics, which generates the membrane exten-
sions observed at early times, to a state of lower actin poly-
merization and higher cross-linking of actin fibers. In addition,
ezrin could also oligomerize in this area. The cytoskeleton col-
lar would then become a more compact and stable structure,
which could restrict molecular diffusion and stabilize supra-
molecular clusters. These phenomena would be perturbed by
the overexpression of the dominant negative mutant of ezrin,
Fig. 7. Working model to explain the role of F-actin-ezrincomplexes in molecular clustering at the immunologicalsynapse. TCR triggering induces the polarization of F-actin andezrin to the APC contact zone producing membrane extensionsthat cover a large surface of the APC. We propose that this anchorsezrin-binding proteins, such as CD43, creating a ‘collar’ ofcytoskeleton-anchored transmembrane proteins at the periphery ofthe contact zone. This collar would function as a transmembrane‘fence’ reducing the diffusion of TCRs out of the contact zone andfacilitating the initial concentration of TCRs into small clusters (2).Later on, membrane extensions retract although ezrin remainsaccumulated in this area. At this time a constriction of the F-actin-ezrin collar occurs which is concomitant to the reduction ofthe cell–cell contact area. This constriction of the cytoskeletoncollar could compress dispersed small TCR clusters facilitating theircoalescence into a single cluster (3).
131Immunological Reviews 189/2002
which lacks the C-terminal half of the molecule, and is there-
fore unable to bind to actin fibers and to oligomerize.
The actin-cytoskeleton associated protein coronin-1 istransiently recruited to the T cell–APC contact area
First described in Distyostelium discoideum, coronin and coronin-
like proteins are expressed in many eukaryotic species from
yeast to mammals. They have been described as actin-binding
proteins implicated in cellular processes involving actin cyto-
skeleton dynamics, such as cell migration, cytokinesis, phago-
cytosis, or morphogenesis (65). Coronins from different spe-
cies display a conserved structural organization. Common fea-
tures include 5 WD repeats, a coiled-coil domain at the C-
terminus, and a linker sequence in between of variable length
referred to as the unique region. Similar to WD-containing Gb
proteins (66), WD repeats of coronins are thought to be organ-
ized in a b-propeller domain, likely involved in protein interac-
tions. Coronin-1 is the mammalian paralogue specifically en-
riched in hematopoietic tissues. Depending on the cell lineage,
coronin-1 appears to have different molecular interactions and,
presumably, different functional activities. For instance, in neu-
trophils, coronin-1 was found associated with components of
the NADPH oxidase complex, which produces superoxide in
phagocytic cells (67). Moreover, it was found as a component
of the phagosome coat in macrophages (68, 69).
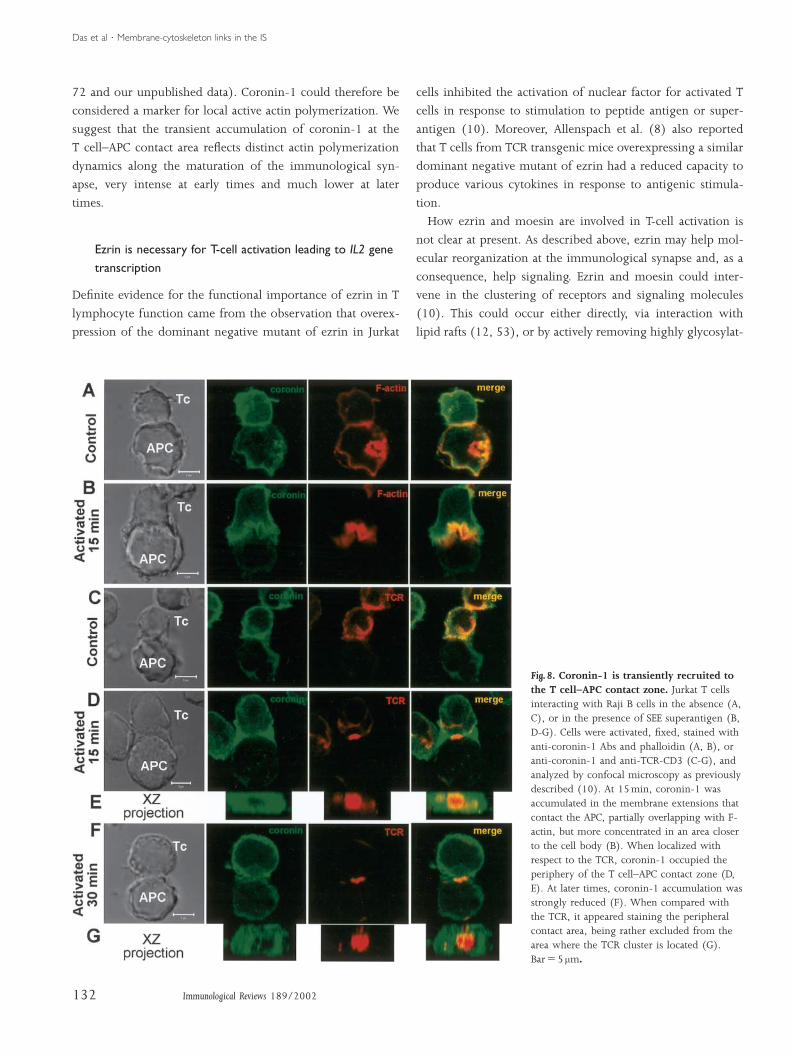
We found that the coronin-1 gene is expressed in lymphoid
precursor cells within mice embryos as well as in differentiat-
ing thymocytes. Coronin-1 is enriched in subcellular struc-
tures rich in F-actin, such as lamellipodia and filopodia, in
both mouse and human T lymphocytes (Nal et al., submit-
ted). In T cells encountering superantigen-pulsed APCs, co-
ronin-1 transiently accumulates at the cell–cell contact area
(Fig. 8). The maximal accumulation occurs within the large
membrane protrusions that engulf the APC at early stages of
the interaction between T cells and APCs. Coronin-1, F-actin
and ezrin are enriched in these membrane structures; they
partially overlap, although they have distinct relative spatial
localizations. Coronin-1 is more enriched in the area closer
to the cell body, F-actin in the middle and ezrin in the more
cortical area (10) (Fig. 8B). In contrast, at later times, coronin-
1 accumulation in the contact area mostly disappeared,
whereas ezrin remained accumulated (Figs. 3D, E, 8F, G).
Interestingly, coronins were shown to be related to actin
filament assembly rather than to stability. For instance, co-
ronins were found enriched in actin tails induced by Listeria
monocytogenes in infected cells, at phagocytic cups, in lamellipo-
dia and in membrane ruffles, but less in stress fibers (68, 70–
Das et al ¡ Membrane-cytoskeleton links in the IS
72 and our unpublished data). Coronin-1 could therefore be
considered a marker for local active actin polymerization. We
suggest that the transient accumulation of coronin-1 at the
T cell–APC contact area reflects distinct actin polymerization
dynamics along the maturation of the immunological syn-
apse, very intense at early times and much lower at later
times.
Ezrin is necessary for T-cell activation leading to IL2 genetranscription
Definite evidence for the functional importance of ezrin in T
lymphocyte function came from the observation that overex-
pression of the dominant negative mutant of ezrin in Jurkat
Fig. 8. Coronin-1 is transiently recruited tothe T cell–APC contact zone. Jurkat T cellsinteracting with Raji B cells in the absence (A,C), or in the presence of SEE superantigen (B,D-G). Cells were activated, fixed, stained withanti-coronin-1 Abs and phalloidin (A, B), oranti-coronin-1 and anti-TCR-CD3 (C-G), andanalyzed by confocal microscopy as previouslydescribed (10). At 15min, coronin-1 wasaccumulated in the membrane extensions thatcontact the APC, partially overlapping with F-actin, but more concentrated in an area closerto the cell body (B). When localized withrespect to the TCR, coronin-1 occupied theperiphery of the T cell–APC contact zone (D,E). At later times, coronin-1 accumulation wasstrongly reduced (F). When compared withthe TCR, it appeared staining the peripheralcontact area, being rather excluded from thearea where the TCR cluster is located (G).BarΩ5 mm.
132 Immunological Reviews 189/2002
cells inhibited the activation of nuclear factor for activated T
cells in response to stimulation to peptide antigen or super-
antigen (10). Moreover, Allenspach et al. (8) also reported
that T cells from TCR transgenic mice overexpressing a similar
dominant negative mutant of ezrin had a reduced capacity to
produce various cytokines in response to antigenic stimula-
tion.
How ezrin and moesin are involved in T-cell activation is
not clear at present. As described above, ezrin may help mol-
ecular reorganization at the immunological synapse and, as a
consequence, help signaling. Ezrin and moesin could inter-
vene in the clustering of receptors and signaling molecules
(10). This could occur either directly, via interaction with
lipid rafts (12, 53), or by actively removing highly glycosylat-

Das et al ¡ Membrane-cytoskeleton links in the IS
ed molecules, such as CD43 (8, 9, 11), or PSGL-1 (57),
whose presence could destabilize T cell–APC interactions and
therefore activation. It might also act by facilitating initial in-
teractions between T cells and APC (57). Ezrin and moesin
might also intervene in one or various T-cell signaling path-
ways, by directly interacting with signal transduction mol-
ecules, as shown in other experimental systems (17). An in-
volvement of ERM molecules at various stages of the T-cell
activation process could explain the strong effect of dominant
References
1. 11. 20.Dustin ML, Cooper JA. The immunological Savage NDG, Kimzey SL, Bromley SK, John- Simons PC, Pietromonaco SF, Reczec D,synapse and the actin cytoskeleton: molecular son KG, Dustin ML, Green JM. Polar redistri- Bretscher A, Elias L. C-terminal threoninehardware for T cell signaling. Nat Immunol bution of the sialoglycoprotein CD43: im- phosphorylation activates ERM proteins to2000;1:23–29. plications for T cell function. J Immunol link the cell’s cortical lipid bilayer to the cyto-
2. 2002;168:3740–3746.Krummel MF, Davis MM. Dynamics of the skeleton. Biochem Biophys Res Communimmunological synapse: finding, establishing 12. Itoh K, et al. Negative regulation of immune 1998;253:561–565.and solidifying a connection. Curr Opin Im- synapse formation by anchoring lipid raft to 21. Matsui T, et al. Rho-kinase phosphorylatesmunol 2002;14:66–74. cytoskeleton through Cbp-EBP50-ERM as- COOH-terminal threonines of ezrin/radixin/
3. sembly. J Immunol 2002;168:541–544.van den Merwe PA. Formation and function moesin (ERM) proteins and regulates theirof the immunological synapse. Curr Opin 13. Egerton M, Burgess WH, Chen D, Druker BJ, head to tail association. J Cell BiolImmunol 2002;14:293–298. Bretscher A, Samelson LE. Identification of 1998;3:647–657.
4. ezrin as an 81-kDa tyrosine-phosphorylatedKupfer A, Singer SJ. Cell biology of cytotoxic 22. Nakamura F, Amieva MR, Furthmayr H.and helper T-cell functions: immunofluor- protein in T cells. J Immunol Phosphorylation of threonine 558 in theescence microscopic studies of single cells 1992;149:1847–1852. carboxyl-terminal actin-binding domain ofand cell couples. Annu Rev Immunol 14. Thuillier L, Hivroz C, Fagard R, Andreoli C, moesin by thrombin activation of human1989;7:309–337. Mangeat P. Ligation of CD4 surface antigen platelets. J Biol Chem 1995;270:31377–
5. induces rapid tyrosine phosphorylation ofCritchley DR. Focal adhesions ª the cyto- 31385.skeletal connection. Curr Opin Cell Biol the cytoskeletal protein ezrin. Cell Immunol 23. Huang L, Wong TYW, Lin RCC, Furthmayr2000;12:133–139. 1994;156:322–331. H. Replacement of threonine 558, a critical
6. 15. site of phosphorylation of moesin in vivo,Monks CRF, Freiberg BA, Kupfer H, Sciaky Pietromonaco SF, Simons PC, Altman A, Eli-N, Kupfer A. Three-dimensional segregation as L. Protein kinase C-q phosphorylation of with aspartate activates F-actin binding ofof supramolecular activation clusters in T moesin in the actin-binding sequence. J Biol moesin. Regulation by conformationalcells. Nature 1998;395:82–86. Chem 1998;273:7594–7603. change. J Biol Chem 1999;274:12803–
7. 16. 12810.Dustin ML, et al. A novel adaptor protein Bretscher A, Chambers D, Nguyen R, Reczekorchestrates receptor patterning and cyto- D. ERM-Merlin and EBP50 protein families 24. Nakamura F, Huang L, Pestonjamasp K,skeletal polarity in T cell contacts. Cell in plasma membrane organization and func- Luna EJ, Furthmayr H. Regulation of F-actin1998;94:667–677. tion. Ann Rev Cell Dev Biol 2000;16:113– binding to platelet moesin in vitro by both
8. 143.Allenspach EJ, et al. ERM-dependent move- phosphorylation of threonine 558 and phos-ment of CD43 defines a novel protein com- 17. Gautreau A, Louvard D, Arpin M. ERM pro- phatidylinositides. Mol Biol Cellplex distal to the immunological synapse. teins and NF2 tumor suppressor: the yin 1999;10:2669–2685.Immunity 2001;15:739–750. and yang of cortical actin organization and 25. Gautreau A, Louvard D, Arpin M. Morphog-
9. cell growth signaling. Curr Opin Cell BiolDelon J, Kaibuchi K, Germain RN. Exclusion enic effects of ezrin require a phosphoryla-of CD43 from the immunological synapse is 2002;14:104–109. tion-induced transition from oligomers tomediated by phosphorylation-regulated re- 18. Algrain M, Turunen O, Vaheri A, Louvard D, monomers at the plasma membrane. J Celllocation of the cytoskeletal adaptor moesin. Arpin M. Ezrin contains cytoskeleton and Biol 2000;150:193–203.Immunity 2001;15:691–701. membrane binding domains accounting for 26. Yonemura S, et al. Ezrin/Radixin/Moesin
10. its proposed role as a membrane-cyto-Roumier A, et al. The membrane-micro- (ERM) proteins bind to a positively chargedfilament linker ezrin is involved in the for- skeletal linker. J Cell Biol 1993;120:129– aminoacid cluster in the juxta-membranemation of the immunological synapse and 139. cytoplasmic domain of CD44, CD43 andin T cell activation. Immunity 2001;15:715– 19. Pearson MA, Recezk D, Bretscher A, Karplus ICAM-2. J Cell Biol 1998;140:885–895.728. PA. Structure of the ERM protein moesin re-
veals the FERM domain fold masked by anextended actin binding tail domain. Cell2000;101:259–270.
133Immunological Reviews 189/2002
negative ezrin in late activation events, such as cytokine gene
activation (10).
Clearly, more work is necessary to elucidate the role of the
actin cytoskeleton, and in particular the role of ezrin and co-
ronin-1 in early T-cell functions. The role of these proteins in
the molecular reorganization of the T cell–APC contact zone
and in promoting T-cell activation will be of particular inter-
est.

Das et al ¡ Membrane-cytoskeleton links in the IS
27. 39. 53.Heiska L, Alfthan A, Gronholm M, Vilja P, Poullet P, Gautreau A, Kadare G, Girault JA, Tomas EM, Chau TA, Madrenas J. ClusteringVaheri A, Carpen O. Association of ezrin Louvard D, Arpin M. Ezrin interacts with of a lipid-raft associated pool of ERM pro-with intercellular adhesion molecule-1 focal adhesion kinase and induces its acti- teins at the immunological synapse upon Tand -2 (ICAM-1 and ICAM-2). Regulation by vation independently of cell-matrix adhesion. cell receptor or CD28 ligation. Immunol Lettphosphatidylinositol 4,5-bisphosphate. J J Biol Chem 2001;276:37686–37691. 2002;83:143–147.Biol Chem 1998;273:21893–21900. 40. Defacque H, et al. Involvement of ezrin/ 54. Lowin-Kropf B, Smith-Shapiro V, Weiss A.
28. moesin in de novo actin assembly on phagoso-Serrador JM, et al. Moesin interacts with the Cytoskeletal polarization of T cells is regu-cytoplasmic region of intercellular adhesion mal membranes. EMBO J 2000;19:199– lated by an immunoreceptor tyrosine-basedmolecule-3 and is redistributed to the uro- 212. mechanism. J Cell Biol 1998;140:861–871.pod of T lymphocytes during cell polariza- 41. Donnadieu E, Bismuth G, Trautman A. Anti- 55. Krause M, Sechi A, Konradt M, Monner D,tion. J Cell Biol 1997;138:1409–1423. gen recognition by helper T cells elicits a se- Gertler FB, Wehland J. Fyn binding protein
29. quence of distinct changes of their shapeSerrador JM, et al. CD43 interacts with moe- (Fyb) /SLP-76-associated protein (SLAP),sin and ezrin and regulates its distribution to and intracellular calcium. Curr Biol Ena/Vasodilator-stimulated phosphoproteinthe uropods of T lymphocytes at the cell– 1994;4:584–595. (VASP) proteins and the ARP2/3 complexcell contacts. Blood 1998;91:4632–4644. 42. Valitutti S, Dessing M, Aktories K, Gallati H, link T cell receptor (TCR) signaling to the
30. Lanzavecchia A. Sustained signaling leadingAlonso-Lebrero JL, et al. Polarization and in- actin cytoskeleton. J Cell Biolteraction of adhesion molecules P-selectin to T cell activation results from prolonged T 2000;149:181–194.glycoprotein ligand 1 and intercellular ad- cell receptor occupancy. Role of T cell actin 56. Sperling AI, Sedy JR, Manjunath N, Kupferhesion molecule 3 with moesin and ezrin in cytoskeleton. J Exp Med 1995;181:577– A, Ardam B, Burkhardt JK. TCR signalinglymphoid cells. Blood 2000;95:2413–2419. 584. induces selective exclusion of CD43 from
31. 43. the T cell-antigen presenting cell contact site.Helander TS, Carpen O, Turunen O, Kovanen Negulescu PA, Krasieva TB, Khan A, Kersch-P, Vaheri A, Timonen T. ICAM-2 redistributed baum HH, Cahalan MD. Polarity of T cell J Immunol 1998;161:6459–6462.by ezrin as a target for killer cells. Nature shape, motility, and sensitivity to antigen. 57. Montoya MC, et al. Role of ICAM-3 in the1996;382:265–268. Immunity 1996;4:421–430. initial interaction of T lymphocytes and
32. 44. APCs. Nat Immunol 2002;3:159–168.Sanchez-Madrid F, del Pozo MA. Leukocyte Delon J, Bercovici N, Liblau R, Trautmannpolarization in cell migration and immune A. Imaging antigen recognition by naive 58. Crepaldi T, Gautreau A, Comoglio PM, Lou-interactions. EMBO J 1999;18:501–511. CD4π T cells. compulsory cytoskeletal alter- vard D, Arpin M. Ezrin is an effector of he-
33. ations for the triggering of an intracellularBarret C, Roy C, Montcurrier P, Mangeat P, patocyte growth factor-mediated migrationNiggli V. Mutagenesis of the phosphatidyl- calcium response. Eur J Immunol and morphogenesis in epithelial cells. J Cellinositol 4,5-bisphosphate (PIP2) binding 1998;28:716–729. Biol 1997;138:423–434.site in the NH2-terminal domain of ezrin 45. Wülfing C, Davis MM. A receptor/cyto- 59. Olivo-Marin JC. Extraction of spots in bio-correlates with its altered cellular distri- skeletal movement triggered by costimula- logical images using multiscale products.bution. J Cell Biol 2000;151:1067–1079. tion during T cell activation. Science Pattern Recognition 2002;35:1989–1996.
34. 1998;282:2266–2269.Hirao M, et al. Regulation mechanisms of 60. Monks CRF, Kupfer H, Tamir I, Barlow A,ERM (ezrin, radixin, moesin) protein/ 46. Grakoui A, et al. The immunological syn- Kupfer A. Selective modulation of protein ki-plasma membrane association: possible in- apse: a molecular machine controlling T cell nase C-q during T-cell activation. Naturevolvement of phosphatidylinositol turnover activation. Science 1999;285:221–227. 1997;385:83–86.and Rho-dependent signaling pathway. J Cell 47. Gunzer M, et al. Antigen presentation in 61. Simons K, Toomre D. Lipid rafts and signalBiol 1996;135:37–51. extracellular matrix. interactions of T cells transduction. Nature Rev Mol Cell Biol
35. with dendritic cells are dynamic, shortTakahashi K, et al. Direct interaction of the 2000;1:31–39.Rho dissociation inhibitor with ezrin/ra- lived, and sequential. Immunity 62. Drevot P, et al. TCR signal initiation machin-dixin/moesin initiates the activation of the 2000;13:323–332. ery is pre-assembled and activated in a sub-Rho small G protein. J Biol Chem 48. Krummel MF, Sjaastad MD, Wülfing C, Davis set of membrane rafts. EMBO J1997;272:23371–23375. MM. Differential clustering of CD4 and CD3z 2002;21:1899–1908.
36. during T cell recognition. ScienceGautreau A, Poullet P, Louvard D, Arpin M. 63. Harder T. Raft membrane domains and im-Ezrin, a plasma membrane-microfilament 2000;289:1349–1352. munoreceptor functions. Adv Immunollinker, signals cell survival through the 49. Stoll S, Delon J, Brotz TM, Germain RN. Dy- 2001;77:45–92.phosphatidylinositol 3-kinase/Akt pathway. namic imaging of T cell–dendritic cell in- 64. Bi K, et al. Antigen-induced translocation ofProc Natl Acad Sci USA 1999;96:7300– teractions in lymph nodes. Science PKCq to membrane rafts is required for T cell7305. 2002;296:1873–1875. activation. Nat Immunol 2001;2:556–563.
37. 50. 65.Parlato S, et al. CD95 (APO1/Fas) linkage to Miller MJ, Wei SH, Parker I, Cahalan MD. de Hostos EL. The coronin family of actin-the actin cytoskeleton through ezrin in hu- Two-photon imaging of lymphocyte mo- associated proteins. Trends Cell Biolman T lymphocytes: a novel regulatory tility and antigen response in intact lymph 1999;9:345–350.mechanism of the CD95 apoptotic pathway. node. Science 2002;296:1869–1873. 66. Smith TF, Gaitatxes C, Saxena K, Neer EJ. TheEMBO J 2000;19:5123–5134. 51. Bousso P, Bhakta NR, Lewis RS, Robey E. Dy- WD repeat: a common architecture for di-
38. namics of thymocyte stromal cell interac-Ng T, et al. Ezrin is a downstream effector verse functions. Trends Biochem Sciof trafficking PKC-integrin complexes in- tions visualized by two photon microscopy. 1999;24:181–185.volved in the control of cell motility. EMBO Science 2002;296:1876–1880.J 2001;20:2723–2741. 52. Michaely PA, Mineo C, Ying YS, Anderson
RGW. Polarized distribution of endogenousRac1 and RhoA at the cell surface. J BiolChem 1999;274:21430–21436.
134 Immunological Reviews 189/2002

Das et al ¡ Membrane-cytoskeleton links in the IS
67. 69. 71.Grogan A, et al. Cytosolic phox proteins inter- Schüller S, Neefjes J, Ottenhoff T, Thole J, Maniak M, Rauchengerger R, Albrecht R,act with and regulate the assembly of coronin Young D. Coronin is involved in uptake of Murphy J, Gerisch G. Coronin involved inin neutrophils. J Cell Sci 1997;110:3071– Mycobacterium bovis BCG in human macro- phagocytosis. dynamics of particle-induced3081. phages but not in phagosome maintenance. relocalization visualized by a green fluo-
68. Cell Microbiol 2001;3:785–793.Ferrari G, Langen H, Naito M, Pieters J. A rescent protein tag. Cell 1995;15:915–924.coat protein on phagosomes involved in the 70. David V, et al. Identification of cofilin, co- 72. Mishima M, Nishida E. Coronin localizes tointracellular survival of Mycobacteria. Cell ronin, Rac and CapZ in actin tails using a leading edges and is involved in cell1999;97:435–447. Listeria affinity approach. J Cell Sci spreading and lamellipodium extension in
1998;111:2877–2884. vertevrate cells. J Cell Sci 1999;112:2833–2842.
135Immunological Reviews 189/2002