mechanism for inhibition of deoxyribonuclease activity by antisera
TRANSCRIPT

Journal of Protein Chemistry, Vol. 16, No. 2, 1997
Mechanism for Inhibition of Deoxyribonuclease Activityby Antisera
Yuh-Fan Liu1 and Ta-Hsiu Liao^
Received November 1, 1996
The activity of bovine DNase, but not that of porcine DNase, is inhibited by antiseraagainst bovine DNase, and vice versa. Inhibition of DNase is found in the immunoglobulinG-containing fractions, as shown by ion exchange chromatography. Inactive DNase,carboxymethylated specifically at the active site His134, competes with active DNase andreverses the antisera inhibition of DNase, suggesting that the epitode responsible forinhibition does not contain the active site His134. Alignment of the sequences of DNase ofthese two species shows that the greatest variation occurs between residues 153 and 163,within which are three consecutive peptide bonds, Lys-Trp-His-Leu, that are readilycleaved by trypsin, chymotrypsin, or thermolysin. The 8-hr digest of DNase by each ofthese three proteases has lost the ability to reverse antisera inhibition. The degree ofantisera inhibition varies with the metal ion used as the activator for DNase-catalyzedreactions. When Mn2+, Co2+, or Mg2"1' plus Ca2+ are used as activators, inhibition isapproximately 50%. When pBR322 plasmid is used as substrate, gel electrophoresis showsthat the DNase-catalyzed DNA hydrolysis produces a significant amount of double-strandcuts with Mn2"1", Co2+, or Mg2* plus Ca2* as activators and antisera inhibit DNase actiononly on double-strand cuts. With only Mg2* as the activator no double-strand cuts areobserved, either in the presence or absence of antisera, and the DNase activity is notsignificantly inhibited. We conclude that antisera inhibition is due to antibody binding ofthe DNase polypeptide chain within residues 153 and 163. These residues are not crucialfor catalysis, but are required for DNA binding, which results in double-strand cuts.
KEY WORDS: DNase; antisera inhibition; divalent metal ions; DNA scission.
1. INTRODUCTION
Divalent metal ions are required for activation ofbovine pancreatic DNase (Kunitz, 1950) and theseions may have more than one function (Shack andBynum, 1964; Melgar and Goldthwait, 1968;Wiberg, 1958). Several binding sites on the enzymeand/or on DNA have been suggested (Price, 1972;1975; Douvas and Price, 1975; Jouve and Jouve,1975; Price et al., 1969). The metal ions satisfyingthe requirement are Mg2+, Ca2+, Co2+, and Mn2+
1 Institute of Biochemistry, College of Medicine, NationalTaiwan University, Taipei, Taiwan.
2 To whom correspondence should be addressed; e-mail:[email protected]. tw.
(Junowicz and Spencer, 1973). However, there arelarge differences in their efficiencies of activation.The mode of action of DNase on duplex DNA alsodepends on which metal ion is used (Campbell andJackson, 1980). The DNase-catalyzed hydrolysis ofduplex DNA makes a significant number ofdouble-strand cuts in the presence of Mn2+, Co2+,or Mg2"1" plus Ca2+, but only single-strand cuts whenMg2"1" is the activator. A previous study (Abe andLiao, 1983) showed that antisera against bovineDNase inhibits bovine, but not porcine DNase.Since the amino acid sequences of the two DNasesare highly conserved (Paudel and Liao, 1986),particularly around the active site His134, the resultof this study suggested that antisera act on sites
750277-8033/97/0200-0075S12.50/0 © 1997 Plenum Publishing Corporation

76
other than the catalytic site. Herein we presentattempts to correlate the antisera inhibition ofDNase with DNase activation by metal ions andwith the mode of DNase action on duplex DNA.
2. MATERIALS AND METHODS
2.1. Materials
Calf thymus DNA and peroxidase-labeled goatantibody against rabbit IgG were obtained fromSigma. The pBR322 plasmid was from Promega.Antisera were prepared essentially as describedpreviously (Abe and Liao, 1983). Porcine DNasewas purified from crude extracts of porcinepancreas according to the procedure of Paudel andLiao (1986). Bovine pancreatic DNase was purifiedfrom the semipurified preparation of Worthington(Code DP) as described previously (Liao et al.,1991).
2.2. Preparation of CM-His134-DNase
Bovine CM-His134-DNase was prepared asdescribed by Price et al. (1969). Purified bovinepancreatic DNase (1 mg/ml) was equilibrated at25°C in 0-05 M Tris-HCl, pH7-2, 4mM CuCl2, forat least 15 min. lodoacetate was then added and thesolution was kept in the dark. At suitable intervalsaliquots were removed and assayed. DNase activitywas determined by the hyperchromicity assay ofKunitz (1950) as modified by Liao (1974). TheDNase activity of CM-His134-DNase was less than1 % of that of unmodified DNase.
2.3. Assay of Single and Double Cuts of DNA
In a typical assay, 9-2 mg/ml of pBR322plasmid DNA in 0-1 M HEPES, 0-1 M NaCl, pH7-2, and 3-6 mM divalent metal ions was incubatedat 25°C with 7-lmU/ml DNase. At various times,14-ju.l aliquots were removed and added to 3 /A! of50 mM EDTA, pH 7-2, 25% glycerol, and 0-01%bromophenol blue on ice. All samples wereresolved into their component species by el-ectrophoresis on 1% agarose slab gels using a TAErunning buffer containing 2-5 mg/ml of ethidiumbromide (Manitis et al., 1982). Total electrophoresistime was 60 min.
Liu and Liao
2.4. Enzyme-Linked Inununosorbent Assay(ELISA)
The procedure for ELISA was essentiallyaccording to Engvall (1980). In wells of themicrotiter plates, bovine or porcine DNase was firstcoated for 2 hr at 37°C in the coating buffer (3 mMNaN3, 0.035 M NaHCO3, 0.014 M Na2CO3, pH 9.6)and then blocked with 0.5% skim milk powder inphosphate-buffered saline, pH 7.2, overnight at4°C. The coated antigen was incubated at 37°C withserial diluted antisera for 1 hr and then washed fourtimes with 0.05% Tween-20 in phosphate-bufferedsaline, pH 7.2. Antibodies bound to antigens weredetermined by 1 hr incubation with the peroxidase-labeled goat antibody against rabbit IgG. Afteranother wash step, the wells were covered with thesubstrate solution, which was 2,2'-azinobis(3-ethlbenothiazoline-6-sulfonic acid) (0.4 mg/ml) in0.1 M phosphate/citrate buffer, pH 4.0, containing0.03% H2O2. The absorbance at 405 nm after 5 minwas measured on the ELISA plate reader (Bio-TekEL312).
3. RESULTS
3.1. Cross-Reaction Between Antisera AgainstBovine and Porcine DNase
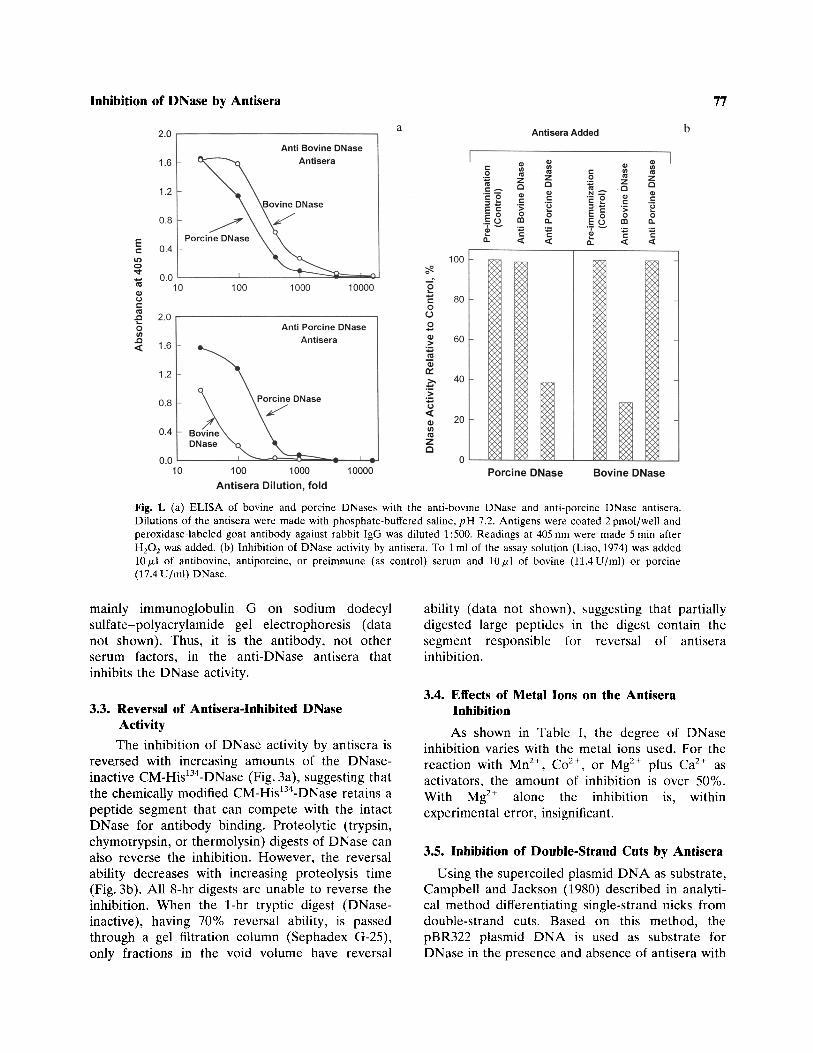
In the immuno-cross-reaction, as analyzed byELISA (Fig. la), both the anti-bovine and theanti-porcine DNase sera have a higher titer for itsown antigen than for its counterpart, suggestingthat the two DNase molecules of different speciesshare some common antigenic determinants.However, other epitope sites, not common to bothDNase molecules, may also exist. As for DNaseactivity, the anti-bovine DNase antisera inhibit 72%of the bovine DNase, but not the porcine DNaseactivity (Fig. Ib, right panel), whereas antiseraagainst porcine DNase inhibit 61% of the porcineDNase, but not the bovine DNase (Fig. Ib, leftpanel).
3.2. Nature of the Inhibition Factor
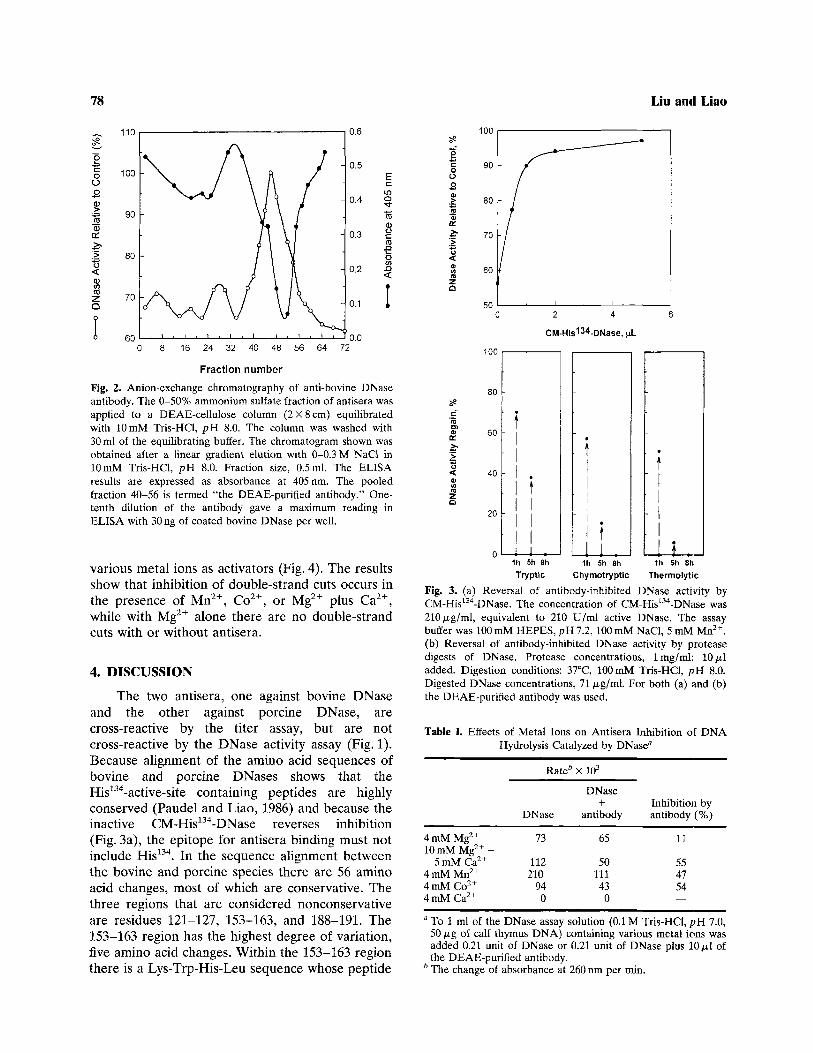
When the 0-50% ammonium sulfate fractionof the anti-bovine DNase antisera is applied to aDEAE-cellulose column, the elution profile (Fig. 2)shows that the inhibition of DNase activity occursin fractions 40-56, which contain antibodies againstDNase as shown by ELISA (absorbance at405 nm). The proteins in these fractions show
Inhibition of DNase by Antisera 77
Fig. 1. (a) ELISA of bovine and porcine DNases with the anti-bovine DNase and anti-porcine DNase antisera.Dilutions of the antisera were made with phosphate-buffered saline, pH 7.2. Antigens were coated 2pmol/well andperoxidase-labeled goat antibody against rabbit IgG was diluted 1:500. Readings at 405 nm were made 5 min afterH2O2 was added, (b) Inhibition of DNase activity by antisera. To 1 ml of the assay solution (Liao, 1974) was added10/j.1 of antibovine, antiporcine, or preimmune (as control) serum and 10/zl of bovine (11.4 U/ml) or porcine(17.4 U/ml) DNase.
mainly immunoglobulin G on sodium dodecylsulfate-polyacrylamide gel electrophoresis (datanot shown). Thus, it is the antibody, not otherserum factors, in the anti-DNase antisera thatinhibits the DNase activity.
3.3. Reversal of Antisera-Inhibited DNaseActivityThe inhibition of DNase activity by antisera is
reversed with increasing amounts of the DNase-inactive CM-His134-DNase (Fig. 3a), suggesting thatthe chemically modified CM-His134-DNase retains apeptide segment that can compete with the intactDNase for antibody binding. Proteolytic (trypsin,chymotrypsin, or thermolysin) digests of DNase canalso reverse the inhibition. However, the reversalability decreases with increasing proteolysis time(Fig. 3b). All 8-hr digests are unable to reverse theinhibition. When the 1-hr tryptic digest (DNase-inactive), having 70% reversal ability, is passedthrough a gel filtration column (Sephadex G-25),only fractions in the void volume have reversal
ability (data not shown), suggesting that partiallydigested large peptides in the digest contain thesegment responsible for reversal of antiserainhibition.
3.4. Effects of Metal Ions on the AntiseraInhibition
As shown in Table I, the degree of DNaseinhibition varies with the metal ions used. For thereaction with Mn2+, Co2+, or Mg2+ plus Ca2+ asactivators, the amount of inhibition is over 50%.With Mg2+ alone the inhibition is, withinexperimental error, insignificant.
3.5. Inhibition of Double-Strand Cuts by Antisera
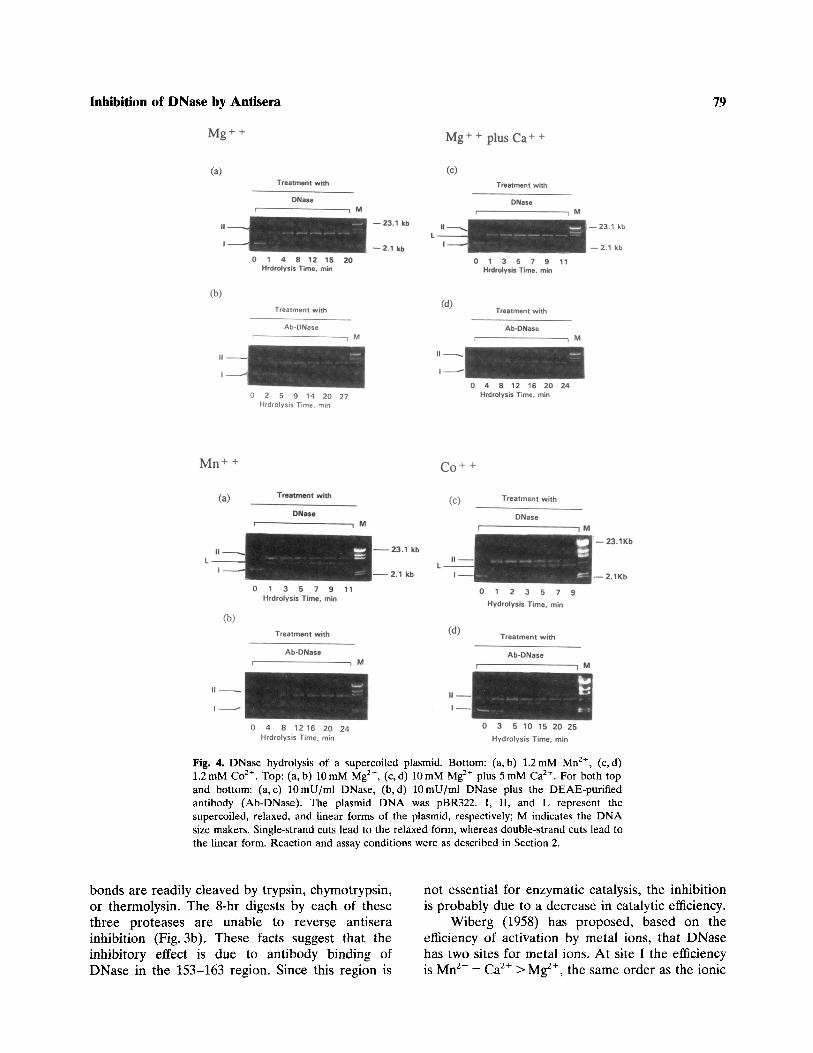
Using the supercoiled plasmid DNA as substrate,Campbell and Jackson (1980) described in analyti-cal method differentiating single-strand nicks fromdouble-strand cuts. Based on this method, thepBR322 plasmid DNA is used as substrate forDNase in the presence and absence of antisera with
78 Liu and Liao
Fig. 2. Anion-exchange chromatography of anti-bovine DNaseantibody. The 0-50% ammonium sulfate fraction of antisera wasapplied to a DEAE-cellulose column (2X8 cm) equilibratedwith 10 mM Tris-HCl, pH 8.0. The column was washed with30 ml of the equilibrating buffer. The chromatogram shown wasobtained after a linear gradient elution with 0-0.3 M NaCl in10 mM Tris-HCl, pH 8.0. Fraction size, 0.5ml. The ELISAresults are expressed as absorbance at 405 nm. The pooledfraction 40-56 is termed "the DEAE-purifled antibody." One-tenth dilution of the antibody gave a maximum reading inELISA with 30 ng of coated bovine DNase per well.
various metal ions as activators (Fig. 4). The resultsshow that inhibition of double-strand cuts occurs inthe presence of Mn2+, Co2+, or Mg2+ plus Ca2+,while with Mg2+ alone there are no double-strandcuts with or without antisera.
4. DISCUSSION
The two antisera, one against bovine DNaseand the other against porcine DNase, arecross-reactive by the titer assay, but are notcross-reactive by the DNase activity assay (Fig. 1).Because alignment of the amino acid sequences ofbovine and porcine DNases shows that theHis134-active-site containing peptides are highlyconserved (Paudel and Liao, 1986) and because theinactive CM-His134-DNase reverses inhibition(Fig. 3a), the epitope for antisera binding must notinclude His134. In the sequence alignment betweenthe bovine and porcine species there are 56 aminoacid changes, most of which are conservative. Thethree regions that are considered nonconservativeare residues 121-127, 153-163, and 188-191. The153-163 region has the highest degree of variation,five amino acid changes. Within the 153-163 regionthere is a Lys-Trp-His-Leu sequence whose peptide
Fig. 3. (a) Reversal of antibody-inhibited DNase activity byCM-His134-DNase. The concentration of CM-His134-DNase was210/j.g/ml, equivalent to 210 U/ml active DNase. The assaybuffer was 100 mM HEPES, pH 7.2, 100 mM NaCl, 5 mM Mn2+.(b) Reversal of antibody-inhibited DNase activity by proteasedigests of DNase. Protease concentrations, Img/ml; 10/x.ladded. Digestion conditions: 37°C, 100mM Tris-HCl, pH 8.0.Digested DNase concentrations, 71 /xg/ml. For both (a) and (b)the DEAE-purified antibody was used.
Table I. Effects of Metal Ions on Antisera Inhibition of DNAHydrolysis Catalyzed by DNase"
Rate6 X 103
4mMMg2+
10mMMg2+ +5 mM Ca2+
4 mM Mn2+
4mMCo2+
4mMCa2+
DNase
73
112210
940
DNase+
antibody
65
50111430
Inhibition byantibody (%)
11
554754
" To 1 ml of the DNase assay solution (0.1 M Tris-HCl, pH 7.0,50 /j.g of calf thymus DNA) containing various metal ions wasadded 0.21 unit of DNase or 0.21 unit of DNase plus 10 ^1 ofthe DEAE-purified antibody.
* The change of absorbance at 260 nm per min.
Inhibition of DNase by Antisera 79
Fig. 4. DNase hydrolysis of a supercoiled plasmid. Bottom: (a, b) 1.2 mM Mn2+, (c, d)1.2 mM Co2+. Top: (a,b) 10 mM Mg2^, (c,d) 10 mM Mg2+ plus 5mM Ca2+. For both topand bottom: (a,c) lOmU/ml DNase, (b, d) lOmU/ml DNase plus the DEAE-purifiedantibody (Ab-DNase). The plasmid DNA was pBR322. I, II, and L represent thesupercoiled, relaxed, and linear forms of the plasmid, respectively; M indicates the DNAsize makers. Single-strand cuts lead to the relaxed form, whereas double-strand cuts lead tothe linear form. Reaction and assay conditions were as described in Section 2.
bonds are readily cleaved by trypsin, chymotrypsin,or thermolysin. The 8-hr digests by each of thesethree proteases are unable to reverse antiserainhibition (Fig. 3b). These facts suggest that theinhibitory effect is due to antibody binding ofDNase in the 153-163 region. Since this region is
not essential for enzymatic catalysis, the inhibitionis probably due to a decrease in catalytic efficiency.
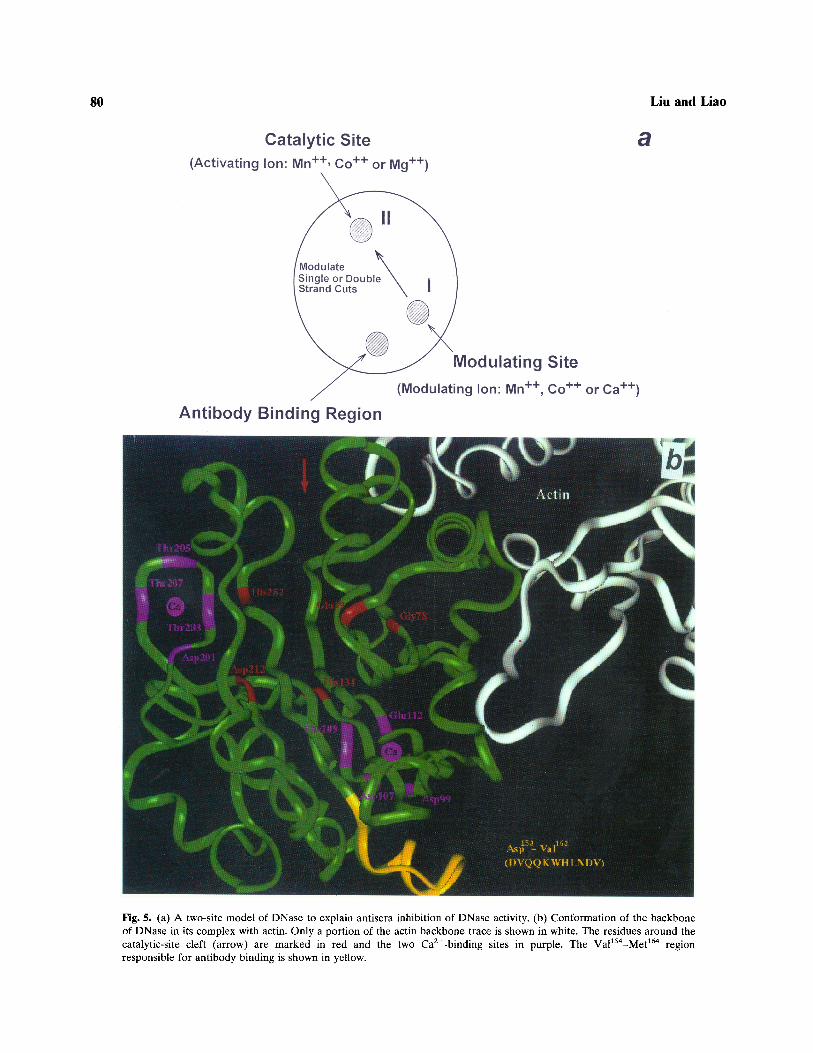
Wiberg (1958) has proposed, based on theefficiency of activation by metal ions, that DNasehas two sites for metal ions. At site I the efficiencyis Mn2+ = Ca2+ > Mg2+, the same order as the ionic
80 Liu and Liao
Fig. 5. (a) A two-site model of DNase to explain antisera inhibition of DNase activity, (b) Conformation of the backboneof DNase in its complex with actin. Only a portion of the actin backbone trace is shown in white. The residues around thecatalytic-site cleft (arrow) are marked in red and the two Ca2+-binding sites in purple. The Val154-Met164 regionresponsible for antibody binding is shown in yellow.

Inhibition of DNase by Antisera
radii, Mn2+ > Ca2+ > Mg2+. At site II the efficiencyis Mn2+>Mg2+»Ca2+, an order similar to theirtendency to form coordinate complexes. More thanone site for metal ions has also been suggested byphysical data (Price, 1972, 1975; Douvas and Price,1975; Jouve and Jouve, 1975; Price et al., 1969). Wepropose for the metal-ion binding a model (Fig. 5a)in which site II, the catalytic site, accommodatesmetal ions such as Mn2+, Co2+, or Mg2+, while siteI, the modulating site, binds Mn2+, Co2+, or Ca2+.The effect of antisera, either on hydrolysis rates oron the mode of action of double-strand cuts, is inaccord with the binding efficiency for site I. On thethree-dimensional structure (Fig. 5b), the catalytic-site cleft formed by the side chains of His252, His134,Glu39, Gly78, and Asp212 is away from site I andfrom the actin-binding site (Kabsch et al., 1990).However, the 153-163 region is close to site I,which is at or near one of the two Ca2+-bindingsites involving the side chains of Glu112, Asp107,Phe109, and Asp" (Fig. 5b). Thus, the binding ofCa2+ or other metal ions at site I may be preventedby antibody binding to region 153-163. As a result,the DNase molecule loses its ability to modulateand to perform double-strand cuts.
EcoRI restriction endonuclease has a homodi-meric structure that enables the enzyme to makedouble-strand cuts on duplex DNA (McClarin etal., 1986). Fungal DNase also has a homodimericstructure (Chen et al., 1993) which makes asignificant number of double-strand cuts, regardlessof the metal ions used. Probably this dimericstructure accounts for the fact that fungal DNasehydrolyzes DNA almost seven times faster thandoes bovine DNase. In the case of bovine DNase,because CM-His134-DNase does not inhibit double-strand cuts when it is mixed with active DNase(date not shown), the heterodimer of CM-His134-DNase/DNase must not occur. In an experiment inwhich the DNase amount was reduced 100-fold tominimize possible dimerization, the number ofdouble-strand cuts remained unchanged (data notshown). Thus, double-strand cutting is not causedby dimerization. Since DNA hydrolysis is due tomonomeric action, the cutting of the second strandmay occur in a processive way. That is, the enzymemay remain bound to DNA during many reactioncycles. This view is supported by the finding(Campbell and Jackson, 1980) that at the end ofeach double-strand cut there is a single-strandregion of 11 nucleotide that is about equal to oneturn of duplex DNA. Perhaps metal-ion binding at
81
site I converts DNase into a conformation with amuch higher affinity with DNA, thus enabling theenzyme to cut the duplex DNA at almost the sameposition along the duplex ladder.
Endogeneous Ca2+, Mg2+-DNase may beinvolved in nuclear DNA degradation duringapoptosis (Peitsch et al., 1993; Polzar et al., 1993).This DNase was identified as DNase type I, thesame type as the DNases used in this investigation.Thus, it is possible that site I in DNase, whichregulates the double cuts through Ca2+ binding,may be one of the factors in apoptosis.
ACKNOWLEDGMENTS
We thank Dr. Roger E. Koeppe of OklahomaState University for critical reading of themanuscript. This work was supported in part by agrant from the National Science Council of ROC(#NSC83-0412-B002-208).
REFERENCES
Abe, A., and Liao, T.-H. (1983). J. Biol. Chem. 258, 10283-10288.
Campbell, V. W., and Jackson, D. A. (1980). /. Biol. Chem. 255,3726-3735.
Chen, L.-Y., Ho, H.-C, Tsai, Y.-C, and Liao, T.-H. (1993).Arch. Biochem. Biophys. 302, 51-56.
Douvas, A., and Price, P. A. (1975). Biochim. Biophys. Ada395, 201-212.
Engvall, E. (1980). Meth. Enzymol. 70, 419-439.Jouve, H., and Jouve, H. (1975). J. Biol. Chem. 250, 6631-
6635.Junowicz, E., and Spencer, J. H. (1973). Biochim. Biophys. Acta
312, 72-84.Kabsch, W., Mannherz, H. G., Suck, D., Pai, E. F., and Holmes,
K. C. (1990). Nature 347, 37-44.Kunitz, M. (1950). /. Gen. Physiol. 33, 349-362.Liao, T.-H. (1974). /. Biol. Chem. 249, 2354-2356.Liao, T.-H., Ho, H.-C., and Abe, A. (1991). Biochim. Biophys.
Acta 1079, 335-342.Maniatis, T., Fritsch, E. F., and Sambrook, J. (1982). In
Molecular Cloning: A Laboratory Manual (Maniatis, T.,Fritsch, E. F., and Sambrook, J., eds.), Cold Spring HarborLaboratories, Cold Spring Harbor, New York, pp. 150-172.
McClarin, J. A., Frederick, C. A., Wang, B. C., Greene, P.,Boyer, H. W., Grable, J., and Rosenberg, J. M. (1986).Science 234,1526-1541.
Melgar, E., and Goldthwait, D. A. (1968). /. Biol. Chem. 243,4409-4416.
Paudel, K., and Liao, T.-H. (1986). J. Biol. Chem. 261, 16012-16017.
Peitsch, M. C., Polzar, B., Stephan, H., Crompton, T.,MacDonald, H. R., Mannherz, H. G., and Tschopp, J.(1993). EMBO J. 12,371-377.

82
Polzar, B., Peitsch, M. C, Loos, R., Tschopp, J., and Mannherz,H. G. (1993). Eur. J. Cell Biol. 62, 397-405.
Price, P. A. (1972). J. Biol. Chem. 247, 2895-2899.Price, P. A. (1975). J. Biol. Chem. 250, 1981-1986.Price, P. A, Moore, S., and Stein, W. H. (1969). J. Biol. Chem.
Liu and Liao
244, 924-928.Shack, J., and Bynum, B. S. (1964). J. Biol. Chem. 239,
3843-3848.Wilberg, J. S. (1958). Arch. Biochem. Biophys. 73, 337-
385.