looking inside mechanistic models of carcinogenesis
TRANSCRIPT

l
Looking inside mechanistic models ofcarcinogenesis
Sascha Zöllner
Helmholtz Zentrum München (Germany)Institute of Radiation Protection

l
Motivation
• Qualitative behavior?• Why do we needmechanistic model?

l
Outline
1 Basic ingredients of multi-stage models
2 Stochastic modelsStochastic processesSimplest case: 1-stage process2-stage process (w/o clonal growth)
3 2-stage model (and beyond)Qualitative features: HazardBeyond the 2-stage modelTime-dependent parameters

l
1 Basic ingredients of multi-stage models
2 Stochastic modelsStochastic processesSimplest case: 1-stage process2-stage process (w/o clonal growth)
3 2-stage model (and beyond)Qualitative features: HazardBeyond the 2-stage modelTime-dependent parameters

l
Modeling cancer: Key ingredients
(epi)genetic transitions: X0µ→ X1
µ→ ··· µ→ Xk
dXk
dt= µXk−1 =⇒ Xk(t) =
∫ t
0dt ′Xk−1µ
(...)= X0(µt)k/k!
[Armitage/Doll (1957)] polynomial growth
clonal expansion of pre-malignant cells: X1γ X1 +1
dX1
dt= γX1 =⇒ X1(t) = X1(0)eγt
exponential growth

l
Modeling cancer: Key ingredients
(epi)genetic transitions: X0µ→ X1
µ→ ··· µ→ Xk
dXk
dt= µXk−1 =⇒ Xk(t) =
∫ t
0dt ′Xk−1µ
(...)= X0(µt)k/k!
[Armitage/Doll (1957)] polynomial growth
clonal expansion of pre-malignant cells: X1γ X1 +1
dX1
dt= γX1 =⇒ X1(t) = X1(0)eγt
exponential growth

l
k = 2 stages
X1 = Nµ0 +X1γ (γ ≡ α−β )
X2 = X1µ1
Link to medical data• Hazard (↔Survival S)
h =probability of new case in (t, t + ∆t]
time step ∆t=−d lnS
dt
• Deterministic approximation:
h ≈ ddt
X2 = µ1X1 =⇒ h ≈ Nµ0µ1 + γh
l
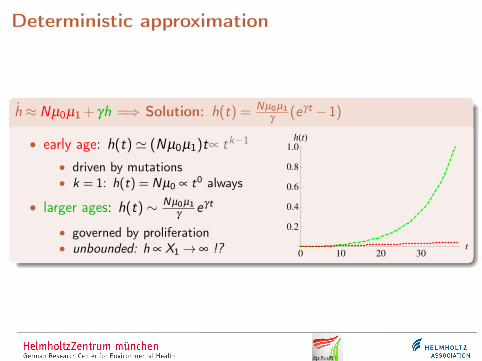
Deterministic approximation
h ≈ Nµ0µ1 + γh =⇒ Solution: h(t) = Nµ0µ1γ
(eγt −1)
• early age: h(t)' (Nµ0µ1)t∝ tk−1
• driven by mutations• k = 1: h(t) = Nµ0 ∝ t0 always
• larger ages: h(t)∼ Nµ0µ1γ
eγt
• governed by proliferation• unbounded: h ∝ X1→ ∞ !?
0 10 20 30t
0.2
0.4
0.6
0.8
1.0
hHtL

l
What’s the problem with h→ ∞?
1 Accuracy• typical data: slowing incidence at t & t∗, w/ t∗ ∼ 60−90• good approximation for t� t∗
2 Conceptually...?• Problem with cancer probability? Consider density ρ:
ρ(t) = h(t)× S(t)︸︷︷︸=exp(−
∫h)
→ 0 OK!
• ...but this implies near-total extinction of population:
S = O(e−eγt)≪ 1

l
What’s the problem with h→ ∞?
1 Accuracy• typical data: slowing incidence at t & t∗, w/ t∗ ∼ 60−90• good approximation for t� t∗
2 Conceptually...?• Problem with cancer probability? Consider density ρ:
ρ(t) = h(t)× S(t)︸︷︷︸=exp(−
∫h)
→ 0 OK!
• ...but this implies near-total extinction of population:
S = O(e−eγt)≪ 1

l
Can we fix it?
1 Go beyond deterministic approximation h 6= µ1X1 Stochastic model (Sec. 2)
2 Within deterministic model? Start from
h(t) = µ1(t)X1(t); X1 = Nµ0 + γX1
Phenomenological modifications:• Compensate X1 ∼ eγt? Age-dependent µ1(t)∼ e−γt (as t→ ∞)
• Keep X1 bounded? X1!
= 0• Age-dependent γ(t), Nµ0(t)...?• ad-hoc cell-cell interaction term: X1 = Nµ0 + γX1−εX 2
1[Sachs, Rad. Research 164 (2005)]

l
Can we fix it?
1 Go beyond deterministic approximation h 6= µ1X1 Stochastic model (Sec. 2)
2 Within deterministic model? Start from
h(t) = µ1(t)X1(t); X1 = Nµ0 + γX1
Phenomenological modifications:• Compensate X1 ∼ eγt? Age-dependent µ1(t)∼ e−γt (as t→ ∞)
• Keep X1 bounded? X1!
= 0• Age-dependent γ(t), Nµ0(t)...?• ad-hoc cell-cell interaction term: X1 = Nµ0 + γX1−εX 2
1[Sachs, Rad. Research 164 (2005)]

l
1 Basic ingredients of multi-stage models
2 Stochastic modelsStochastic processesSimplest case: 1-stage process2-stage process (w/o clonal growth)
3 2-stage model (and beyond)Qualitative features: HazardBeyond the 2-stage modelTime-dependent parameters

l
1 Basic ingredients of multi-stage models
2 Stochastic modelsStochastic processesSimplest case: 1-stage process2-stage process (w/o clonal growth)
3 2-stage model (and beyond)Qualitative features: HazardBeyond the 2-stage modelTime-dependent parameters

l
Stochastic process
• So far: “Sharp” # of cells Xi (t)
• But cancer evolution is stochastic process:Xi (t) are random, w/ probability
Prob{X1(t) = x1, . . . ,Xk(t) = xk}=: Px1...xk (t)≡ Px(t)
Goal• Find Px(t) with initial condition Px(0) = δx ,0 (healthy cells only)• More precisely, we want to model time evolution
P(t0) 7→ P(t)

l
Markov process
• Markov’s condition: “Short-memory” time evolution,i.e., {Px(t)} completely determines Px(t + ∆t)
• leads to Chapman-Kolmogorov eq.
Px(t + ∆t) = ∑x ′Px ′(t)px ′→x
• Transition probabilities• normalized: ∑x px ′→x = 1• completely define the time evolution (i.e., parametrize our model!)

l
Markov processContinuous-time process
Px(t + ∆t) = ∑x ′(6=x)
Px ′(t)px ′→x +Px(t)
(1− ∑
x ′(6=x)
px→x ′
)= Px(t) + ∑
x ′( 6=x)
(Px ′(t)px ′→x −Px(t)px→x ′)
Master equation• Take ∆t→ 0, assuming px ′→x( 6=x ′) ' Ax ′,x∆t:
ddt
Px(t) = ∑x ′
(Px ′(t)Ax ′,x −Px(t)Ax ,x ′
)• Formal solution: P(t)≡A P(t) =⇒ P(t) = eA tP(0)

l
1 Basic ingredients of multi-stage models
2 Stochastic modelsStochastic processesSimplest case: 1-stage process2-stage process (w/o clonal growth)
3 2-stage model (and beyond)Qualitative features: HazardBeyond the 2-stage modelTime-dependent parameters

l
1-stage (Poisson) process
• States: x ≡ (x1) – #cells in stage 1• Assume only 1 transition (from “healthy” → “malignant”)
px ′→x = Nµ0∆t if x ′ = x−1
Px(t + ∆t) = Px−1(t)Nµ0∆t +Px(t)(1−Nµ0∆t)
• Transitions between states:
(x1 = 0)µ0−→ (1)
µ0−→ (2)µ0−→ ·· ·
continuous transfer from (x1 = 0) toward (x1→ ∞)

l
1-stage process: Master equation
Px(t) = Nµ0
Px−1(t)︸ ︷︷ ︸transfer from x−1→x
− Px(t)︸ ︷︷ ︸transfer x→x+1
; Px(0) = δx ,0
Solve:
Px=0 =−Nµ0P0 ⇒ P0(t) = e−Nµ0t
P1 = Nµ0(P0−P1) ⇒ P1(t) = e−Nµ0tNµ0t...
...

l
Solution: Poisson distribution
Px(t) = e−Nµ0t (Nµ0t)x
x!
• E (X1)≡ X1(t) = Nµ0t – probability “travels” w/ speed Nµ0
• Var(X1)≡∆X 21 = Nµ0t – spreads out
• Steady state as t→ ∞? Px(t) = Nµ0 [Px−1(t)−Px(t)]?= 0
0 2 4 6 8 10x
0.2
0.4
0.6
0.8
1.0
PHx;tL

l
A toy model
Imagine the #cells, x , were continuous: Px(t) =: P(x ; t)
P(x ; t) =−Nµ0 [P(x ; t)−P(x−1; t)]→−Nµ0∂
∂xP(x ; t)
Solution: Any “traveling wave” with P(x ; t) = f (x−Nµ0t)
• Proof: ∂t f (x−Nµ0t) = f ′(x−Nµ0t)︸ ︷︷ ︸=∂x f
×[−Nµ0]
• Gives “central” dynamics, but no diffusion

l
Back to “deterministic” model
So far, solved whole problem, Px(t)What is specific dynamics of mean cell #, X (t)?
ddt
X (t) = ∑xx Px(t)
= ∑xx Nµ0 (Px−1−Px)
= Nµ0(X +1−X
)= Nµ0
∴ Heuristic model in Sec. 1 ⇐⇒ Exact dynamics of X (t)

l
Link to risk model
What is the hazard / survival probability for this model?
S(t) = Prob(Tcancer > t) =?
Simplest model: Interpret person as healthy:⇔ X1(t) = 0
S(t) = Prob{X1(t) = 0}= P0(t)
• Survival: S(t) = e−Nµ0t
• Hazard: h(t) =− ddt lnS(t) = +Nµ0
• Same as deterministic model!• Age-independence not realistic for cancer data/biology – let’s move on!

l
Link to risk model
What is the hazard / survival probability for this model?
S(t) = Prob(Tcancer > t) =?
Simplest model: Interpret person as healthy:⇔ X1(t) = 0
S(t) = Prob{X1(t) = 0}= P0(t)
• Survival: S(t) = e−Nµ0t
• Hazard: h(t) =− ddt lnS(t) = +Nµ0
• Same as deterministic model!• Age-independence not realistic for cancer data/biology – let’s move on!

l
1 Basic ingredients of multi-stage models
2 Stochastic modelsStochastic processesSimplest case: 1-stage process2-stage process (w/o clonal growth)
3 2-stage model (and beyond)Qualitative features: HazardBeyond the 2-stage modelTime-dependent parameters

l
Two mutation steps
• States: x ≡ (x1,x2) – #cells in stage 1 (pre-) and 2 (malignant)• 2 possible transitions:
px ′→x =
{Nµ0∆t if x ′ = (x1−1,x2)
x1µ1∆t if x ′ = (x1,x2−1)
• Now 2-D transition chain:
(x1 = 0,x2 = 0)µ0−→ (1,0)
µ0−→ (2,0)µ0−→ ·· ·
↓µ1 ↓µ1
(1,1)µ0−→ (2,1)
µ0−→ ·· ·↓µ1 ↓µ1

l
2-stage process: Master equation
Px1x2(t) = Nµ0 (Px1−1,x2(t)−Px1,x2(t))
+ x1µ1 (Px1,x2−1(t)−Px1,x2(t))
If we are only interested in the hazard, h =−S/S , with
S(t) = Prob{X2(t) = 0}= ∑x1
Px1,0
then we only need the x2 = 0 entries:
Px1,0(t) = Nµ0 [Px1−1,0(t)−Px1,0(t)]− x1µ1Px1,0(t)
Same as 1-stage process, but w/ loss term for high #1-cells (1→2)

l
Hazard
S = ∑x1
Px1,0 =−µ1 ∑x1
x1Px1,0
h =− SS
= +µ1 ∑x1
x1Px1,0
∑x Px ,0= µ1E (X1|X2 = 0)
• Looks like deterministic approximation,but w/ mean X1|0 conditional on X2 = 0!
• Relevant probability distribution for X1:Px1|0 ≡ Px1,0/∑x Px ,0 (normalized)

l
Conditional X1 distribution
Px1|0 obeys the Master-like equation
Px1|0(t) = Nµ0(Px1−1|0(t)−Px1|0(t)
)−[x1− X1|0(t)
]µ1Px1|0(t)
• 1st term: push toward x1→ ∞ (µ0)• 2nd term: redistribute to x1 < X1|0 (µ1)
• Approaches steady state, Px1|0 = 0: Balance between x1-input (fromhealthy cells) and output (to malign cells)
• Explicit solution (const. parameters): Poisson
Px |0(t) = e−X1|0(t) X1|0(t)x
x!, with X1|0(t) =
Nµ0
µ1
(1− e−µ1t
)

l
Back to hazard
Since h = µ1X1|0, all we need is effective equation for X1|0:
ddt
X1|0 = ∑x1
x1[Nµ0
(Px1−1|0−Px1|0
)−(x1− X1|0
)µ1Px1|0
]= Nµ0
(X1 +1−X1
)|0−
(X 2
1|0− X 21|0
)µ1
ddt
X1|0 = Nµ0−∆X 21|0µ1
• “Deterministic” term (from 1st step) – describes mean X1
• “Stochastic” fluctuation term (2nd step)• leads to steady state!• similar effect as phenomenological term inhibiting cell growth (Sec. 1)

l
Hazard: Constant parameters
• For const. parameters: Px1|0 Poissonian ⇒ ∆X 21|0 = X1|0
ddt
X1|0 = Nµ0− X1|0µ1
• strong “damping” if many pre-malign cells (likely already malign –discount, since no longer cause new cancer)
• Rewrite in terms of h = X1|0×µ1:
h = µ1(Nµ0−h) h(t) = Nµ0(1− e−µ1t
)

l
Summary so far
Deterministic model
• Exact equations for mean #cells X1 = µ0X0 + γ1X1
• Approximation for hazard: h ≈ µ1X1
• works well for earlier ages: polynomial / exp. growth• growth unbounded!
Stochastic model• Exact hazard: h = µ1E (X1|X2 = 0)≡ µ1X1|0
• X1|0 obeys similar equation as X1:• same deterministic term (more 1-cells due to µ0)• extra fluctuation term (fewer 1-cells due to µ1)
• Equilibrium at older age: Hazard saturates

l
1 Basic ingredients of multi-stage models
2 Stochastic modelsStochastic processesSimplest case: 1-stage process2-stage process (w/o clonal growth)
3 2-stage model (and beyond)Qualitative features: HazardBeyond the 2-stage modelTime-dependent parameters

l
1 Basic ingredients of multi-stage models
2 Stochastic modelsStochastic processesSimplest case: 1-stage process2-stage process (w/o clonal growth)
3 2-stage model (and beyond)Qualitative features: HazardBeyond the 2-stage modelTime-dependent parameters

l
2-stage model w/ clonal expansion
• Deterministic model: h = Nµ0µ1 + (α−β )h• Include stochastic term −∆X 2
1|0 (constant parameters):
h = Nµ0µ1 + (α−β −µ1︸ ︷︷ ︸=:γ
)h− α
Nµ0h2
• 1st-order term −µ1h (“Poisson contribution ∆X 2 = X ”)• 2nd-order term ∝−αh2 (“high α X1 ↑ increased loss to 2-cells”)
[Moolgavkar (1979-81)]

l
Phases
h
h'deterministic
Nµ0µ1/q
Nµ0µ1
exact
0 20 40 60 80t
0.2
0.4
0.6
0.8
1.0
hHtL
What can we learn from h = Nµ0µ1 + γh− α
Nµ0h2?
• initially: h ' Nµ0µ1 + γh =⇒ h(t)' Nµ0µ1γ
[eγt −1]
• max. growth: h(t∗) = 0 =⇒ h(t)' h(t∗) + h(t∗)(t− t∗)• steady state: h→ 0 =⇒ h(t)→ Nµ0µ1/q
w/ q(q + γ)≡ αµ1; γt∗ ≡ ln γ+qq

l
Effective parameters
h = Nµ0µ1 + γh− α
Nµ0h2
• Scaling invariance only 3 parameters “ identifiable” from h:• Nµ0µ1; γ (deterministic – early age)• αµ1 (stochastic)
• One interpretation:• time scale: t ′ = γt• hazard scale: h′ = hγ/Nµ0µ1
• functional shape: ε ≡ αµ1/γ2 h′ = 1+h′− εh′2

l
1 Basic ingredients of multi-stage models
2 Stochastic modelsStochastic processesSimplest case: 1-stage process2-stage process (w/o clonal growth)
3 2-stage model (and beyond)Qualitative features: HazardBeyond the 2-stage modelTime-dependent parameters

l
Multi-stage models
...X0=N
healthy cells
Xk-1
initiatedcells
µ0 μk-1
αk-1
division
βk-1inactivation/differentiation
Xk
malignantcells
X1
initiatedcells
µ1µ1µ1µ1 µk-2
α1
division
β1inactivation/differentiation
k-stage model w/ clonal expansion (k = 2 ⇐⇒ TSCE)• Parameters
• transitions (µ0,...,k−1): k parameters• proliferation (α1...k−1;β1...k−1): 2(k−1) – too many!
• Qualitative behavior?[Little, Int. J. Rad. Biol. 78 (2002)]

l
Example: 3-stage “pre-initiation” model
Analytic solution
h(t) = Nµ0
1−(qe(γ+q)t + (γ +q)e−qt
γ +2q
)−µ1/α
• 4 (out of 5) identifiable parameters: Nµ0, γ , q, plus µ1/α. . .
• Phases: polynomial ∝t2, exponential, linear, saturation to Nµ0
[Luebeck, PNAS 99 (2002); Meza, PNAS 105 (2008)]

l
What can we really learn?
Can we identify the # of stages from data?• Cancer biology: Many different stages (pathways) involved (k � 1)— why use 2-stage model?
• Modeling: Consider 2 models• simple 2-stage: h2(t) = Nµ0(1− e−µ1t)• 1-stage: h1(t) = Nν(t) w/ ν(t)≡ µ0(1− e−µ1t)
Multistage dynamics not unambiguously observable• Plausible assumptions (rates constant unless motivated, #stages)• Only slow enough (“rate-limiting”) steps leave imprint• Need large enough data set to detect qualitative difference

l
What can we really learn?
Can we identify the # of stages from data?• Cancer biology: Many different stages (pathways) involved (k � 1)— why use 2-stage model?
• Modeling: Consider 2 models• simple 2-stage: h2(t) = Nµ0(1− e−µ1t)• 1-stage: h1(t) = Nν(t) w/ ν(t)≡ µ0(1− e−µ1t)
Multistage dynamics not unambiguously observable• Plausible assumptions (rates constant unless motivated, #stages)• Only slow enough (“rate-limiting”) steps leave imprint• Need large enough data set to detect qualitative difference

l
1 Basic ingredients of multi-stage models
2 Stochastic modelsStochastic processesSimplest case: 1-stage process2-stage process (w/o clonal growth)
3 2-stage model (and beyond)Qualitative features: HazardBeyond the 2-stage modelTime-dependent parameters

l
What about radiation. . .?
• Radiation-induced (epi-)genetic effects• non-repair: apoptosis β ↑• misrepair: additional mutations µi ↑, (α−β ) ↑• . . .
• Toy model to understand effects:
µi (t) = µ(0)i [1+ f (t)] , etc.
f (t) =
{const. t ∈ [t1, t2]
0 else
How could we solve that?

l
What about radiation. . .?
• Radiation-induced (epi-)genetic effects• non-repair: apoptosis β ↑• misrepair: additional mutations µi ↑, (α−β ) ↑• . . .
• Toy model to understand effects:
µi (t) = µ(0)i [1+ f (t)] , etc.
f (t) =
{const. t ∈ [t1, t2]
0 else
How could we solve that?

l
µ1(t): Deterministic model
h(t) = µ1(t)X1(t); X1 = Nµ0 + γX1
µ1 = µ(0)1 (1+ f ) =⇒ h = h(0)(1+ f )
• Jump at t1,2: ∆h = ∆µ1X1 — instant effect!• Excess relative risk = f
l
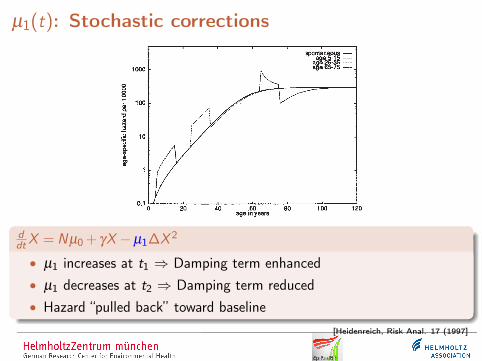
µ1(t): Stochastic corrections
ddtX = Nµ0 + γX −µ1∆X 2
• µ1 increases at t1 ⇒ Damping term enhanced• µ1 decreases at t2 ⇒ Damping term reduced• Hazard “pulled back” toward baseline
[Heidenreich, Risk Anal. 17 (1997]

l
Now for µ0(t):

l
µ0(t)
ddtX = Nµ0 + γX −µ1∆X 2
• Kinks (no jumps): ∆h = µ1×N∆µ0 – extra stage delays effect• Slow return to baseline:Kink at t2 neglible compared to accumulated clonal growth

l
Excercise: α(t)