long-term supplementation of honokiol and magnolol ameliorates body fat accumulation, insulin...
TRANSCRIPT

Mol. Nutr. Food Res. 2013, 00, 1–11 1DOI 10.1002/mnfr.201300113
RESEARCH ARTICLE
Long-term supplementation of honokiol and magnolol
ameliorates body fat accumulation, insulin resistance,
and adipose inflammation in high-fat fed mice
Young-Je Kim1∗, Myung-Sook Choi1,2∗, Byung Yoon Cha3∗, Je Tae Woo4, Yong Bok Park5,Sang Ryong Kim5,6 and Un Ju Jung2
1 Department of Food Science and Nutrition, Kyungpook National University, Daegu, Republic of Korea2 Center for Food and Nutritional Genomics Research, Kyungpook National University, Daegu, Republic of Korea3 Research Institute for Biological Functions, Chubu University, Kasugai, Japan4 Yuto Teruya, Oknawa Research Center Co. Ltd., Suzaki, Uruma, Okinawa, Japan5 School of Life Sciences and Biotechnology, Kyungpook National University, Daegu, Republic of Korea6 Brain Science and Engineering Institute, Kyungpook National University, Daegu, Republic of Korea
Scope: This study investigated the effect of honokiol (HON) and magnolol (MAG), phenoliccompounds in Magnolia plants, on adiposity and adiposity-related metabolic disturbances inmice fed high-fat diet (HFD), and the potential underlying mechanisms focusing on the lipidmetabolism and inflammatory response.Method and results: C57BL/6J mice were fed HFD (45 kcal% fat) with or without HON (0.02%,w/w) or MAG (0.02%, w/w) for 16 wk. Despite no changes in body weight, food intake, andhepatic fat accumulation, HON and MAG significantly lowered the weight of white adiposetissue (WAT) as well as adipocyte size and protected against insulin resistance induced byHFD. These effects were associated with increases in energy expenditure and adipose fatty acidoxidation and decreases in fatty acid synthase activity and expression of genes related to fattyacid synthesis, desaturation, and uptake, as well as adipocyte differentiation in WAT. Moreover,HON and MAG significantly lowered the expression of proinflammatory genes in WAT andelevated the plasma IL-10 level. Particularly, HON significantly decreased the plasma resistinlevel and increased the plasma adiponectin level compared to the control group.Conclusion: HON and MAG have potential as novel agents for amelioration of adiposity andassociated insulin resistance and inflammation.
Keywords:
Adipogenesis / Antiobesity / Honokiol / Inflammation / Magnolol
Received: February 8, 2013Revised: May 6, 2013
Accepted: May 7, 2013
� Additional supporting information may be found in the online version of this article atthe publisher’s web-site
Correspondence: Dr. Un Ju Jung, Center for Food and NutritionalGenomics Research, Kyungpook National University, 1370 San-Kyuk Dong, Puk-Ku, Daegu 702-701, Republic of KoreaE-mail: [email protected]: +82-53-958-1230
Abbreviations: FAS, fatty acid synthase; FAT/CD36, fatty acidtranslocase/cluster of differentiation 36; HFD, high-fat diet;HOMA-IR, homeostatic index of insulin resistance; HON, hon-okiol; IPGTT, intraperitoneal glucose tolerance test; IRS2, in-sulin receptor substrate 2; MAG, magnolol; MCP-1, monocytechemoattractant protein-1; NF-�B, nuclear factor kappa-light-chain-enhancer of activated B cells; PPAR-�, peroxisome pro-
1 Introduction
Obesity is defined as excessive fat accumulation that mayhave adverse effects on health [1]. It is well known as a potentrisk factor for metabolic diseases including type 2 diabetes,dyslipidemia, atherosclerosis, hepatic steatosis, and cancer,and insulin resistance in obesity is the underlying cause formetabolic syndrome [2]. Obesity-induced inflammation, a key
liferator activated receptor-�; SCD1, stearoyl-CoA desaturase;SREBP1c, sterol-regulatory-element-binding protein 1c; TLR2,toll-like receptor 2; TNF-�, tumor necrosis factor-�; WAT, whiteadipose tissue∗These authors contributed equally to this work.
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

2 Y.-J. Kim et al. Mol. Nutr. Food Res. 2013, 00, 1–11
feature of adipose tissue dysfunction, has been proposed to bean important link between obesity and insulin resistance [3].Adipose tissue can respond rapidly and dynamically to ex-cess energy intake through adipocyte hypertrophy and hy-perplasia [4], and enlarged adipose tissue results in alteredproduction of various adipokines (e.g. adiponectin, resistin,and leptin) and inflammatory chemokines and cytokines (e.g.monocyte chemoattractant protein-1 (MCP-1), tumor necro-sis factor-� (TNF-�), IL-6, and IL-10), which impacts insulinsensitivity [5]. The level of circulating resistin and mRNA ex-pression of proinflammatory chemokines and cytokines, in-cluding MCP-1 and TNF-�, in adipose tissue were increasedin mice on a high-fat diet (HFD) that developed obesity andinsulin resistance [6, 7]. In contrast, plasma adiponectin andanti-inflammatory IL-10 levels were low in subjects with obe-sity and type 2 diabetes, and they are positively correlated withinsulin sensitivity [8, 9].
Consumption of a diet high in fat is considered a majorfactor in promoting obesity, insulin resistance, and othermetabolic diseases in humans as well as in various animalmodels [4, 10, 11]. Conversely, high-level consumption ofvegetables, fruits, and whole grains is associated with lowerinflammation [10] and reduced risk for obesity and metabolicdisease [11]. Magnolia plants have been used as Korean andBrazilian complementary and alternative medicines for thetreatment of diabetes and diabetic complications [12] and asJapanese and Chinese traditional herb medicine for the treat-ment of various ailments, including fever, headache, anxiety,stroke, diarrhea, and anemia, due to their antithrombotic,antidepressant, and antibacterial properties [13]. Honokiol(HON) and magnolol (MAG) (Supporting InformationFig. 1), biphenolic structural isomers isolated from the stembark of Magnolia plants, are considered as their major bioac-tive constituents [14]. It is reported that HON and MAG exertanti-inflammatory effects by inhibiting the downstream path-way of MEKK-1 in nuclear factor kappa-light-chain-enhancerof activated B cells (NF-�B) activation signaling [15]. MAGreduced fasting blood glucose and plasma insulin levelsin type 2 diabetic rats without altering body weight [16],and induced glucose uptake in 3T3-L1 adipocytes [17].HON also enhanced insulin signaling pathways such as theRas/ERK1/2 and phosphoinositide-3-kinase/Akt signalingpathways [18], and ameliorated alcoholic steatosis by blockingfatty acid synthesis regulated by sterol-regulatory-element-binding protein 1c (SREBP1c) [19]. Moreover, a recent studyshowed that HON and MAG possessed antioxidant andanti-inflammatory activities, along with low cytotoxicity [20].However, the effects of HON and MAG on diet-inducedobesity have not been explored.
In the present study, we investigated whether HON andMAG attenuate adiposity, insulin resistance, hepatic steato-sis, and inflammation in C57BL/6J mice fed HFD, a widelyused animal model for obesity [21]. To elucidate their poten-tial underlying mechanisms, we evaluated not only plasmaadipokines and cytokines levels, but also the activity of lipid-regulating enzymes and/or the expression of genes involved
in lipid metabolism and inflammation in visceral adiposetissue and liver.
2 Materials and methods
2.1 Isolation and identification of HON and MAG
Commercially available air-dried bark of Magnolia obovatawas purchased from Daikoshoyaku Co. Ltd. (Nagoya, Japan,lot 7G25M). M. obovata bark (dry weight, 3 kg) was extractedin methanol (8 L) for 2 wk. The evaporated methanol extract(362.5 g) was dissolved in distilled water, and successivelyfractionated with ethyl acetate (220.2 g), hexane (90.17 g),and 90% methanol (125 g). The 90% methanol-eluted fractionwas fractionated using a silica gel open column (BW silica gel;BW-820M, Fuji Silysia Chemical Ltd., Japan; methanol andchloroform (0:1, 1:20, 1:10, 1:5, 1:1, and 1:0, v/v). Part of theeluted fractions was further fractioned using an octadecylsilylsilica gel (Cosmosil 75C18-OPN, 60% aqueous methanol tomethanol). The eluted factions were subsequently establishedas HON and MAG by the 1H NMR (400 MHz, CDCl3) and13C NMR (100 MHz, CDCl3) spectra obtained on an FT NMRsystem (JEOL, Japan) (Supporting Information Figs. 2 and3) [22].
2.2 Animals and diets
Obesity-prone C57BL/6J mice (4 wk old, male) were pur-chased from Jackson Laboratories (Bar Harbor, ME, USA).All mice were individually housed under a constant tempera-ture (24�C) with a 12-h light/dark cycle, and fed a normal chowdiet for acclimation for 1 wk after delivery. At 5 wk of age,they were randomly divided into three groups of ten mice eachand fed an HFD (D12451, Research Diets, New Brunswick,NJ, USA) with or without HON (0.02%, w/w, approximately17 mg/kg body weight per day) or MAG (0.02%, w/w, approx-imately 17 mg/kg body weight per day) for 16 wk. The HFDcontains 45 kcal% fat, 20 kcal% protein, and 35 kcal% carbo-hydrate. In the HFD, 87.7% w/w of total fat was from lard,which contains high amounts of saturated fat, and 12.3% w/wof total fat was from soybean oil, an unsaturated fat source.They were given free access to food and distilled water, andfood consumption and body weight were measured daily andweekly, respectively. At the end of the experimental period,all the mice were anesthetized with isoflurane (5 mg/kg bodyweight, Baxter, USA) after a 12-h fast, and blood samples werecollected from the inferior vena cava into heparin-coated tubefor determination of their plasma lipids, adipokines, and cy-tokines levels. The blood was centrifuged at 1000 × g for15 min at 4�C, and the plasma was separated. After bloodcollection, epididymal white adipose tissue (WAT), perirenalWAT, retroperitoneal WAT, mesentery WAT, subcutaneousWAT, and liver were promptly removed, rinsed with physio-logical saline, and weighed. Among them, epididymal WAT
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

Mol. Nutr. Food Res. 2013, 00, 1–11 3
and liver were immediately frozen in liquid nitrogen andstored at –70�C until enzyme activity and/or RNA analyses.Studies were performed under protocols for animal studiesapproved by the Kyungpook National University Ethics Com-mittee (Approval No. KNU-2011–49).
2.3 Energy expenditure
Energy expenditure was measured using an indirect calorime-ter (Oxylet; Panlab, Cornella, Spain). The mice were placedinto individual metabolic chambers at 25�C, with free accessto food and water. O2 and CO2 analyzers were calibratedwith highly purified gas. Oxygen consumption and carbondioxide production were recorded at 3-min intervals using acomputer-assisted data acquisition program (Chart 5.2; ADInstrument, Sydney, Australia) over a 24-h period, and thedata were averaged for each mouse. Energy expenditure wascalculated according to the following formula:
EE(kcal/day/bodyweight0.75) = Vo2× 1.44 × [3.815
+ (1.232 × Vo2/Vco2)]
2.4 Fasting blood glucose, intraperitoneal glucose
tolerance test (IPGTT), and homeostatic index of
insulin resistance (HOMA-IR)
The blood glucose concentration was measured with wholeblood obtained from the tail veins after withholding foodfor 12 h using a glucose analyzer (Glucocard, Arkray, Japan)based on the glucose oxidase method. The IPGTT was per-formed on the 15th wk. After a 12-h fast, the mice were in-jected intraperitoneally with glucose (0.5 g/kg body weight).The blood glucose level was determined from the tail veinat 0, 30, 60, and 120 min after the glucose injection. Thecumulative changes in blood glucose responses were quan-tified by the incremental area under the curve. Homeo-static index of insulin resistance (HOMA-IR) was calcu-lated according to the homeostasis of the assessment asfollows:
HOMA − IR = [fasting glucose(mmol/L) × fasting insulin
(�LU/mL)]/22.51
2.5 Plasma biomarkers
Plasma insulin, adipokines (adiponectin, resistin, and leptin),and cytokines (IL-10, IL-6, TNF-�, and MCP-1) were deter-mined with a multiplex detection kit from Bio-Rad (Hercules,CA, USA). All samples were assayed in duplicate and ana-lyzed with a Luminex 200 Labmap system (Luminex, Austin,TX, USA). Data analyses were done with Bio-Plex Managersoftware version 4.1.1 (Bio-Rad).
Plasma lipid concentrations were determined with com-mercially available kits: Plasma free fatty acid and phospho-lipid levels were measured using the Wako enzymatic kit(Wako Chemicals, Richmond, VA, USA), and triglyceride, to-tal cholesterol, and HDL-cholesterol levels were determinedusing the Asan enzymatic kits (Asan, Seoul, Republic ofKorea).
2.6 Hepatic lipids contents
Hepatic lipids were extracted [23], and then the dried lipidresidues were dissolved in 1 mL of ethanol for triglycerideand cholesterol assays. Triton X-100 and a sodium cholate so-lution in distilled water were added to 200 �L of the dissolvedlipid solution for emulsification. The hepatic triglyceride andcholesterol contents were analyzed with the same enzymatickit used for the plasma analysis.
2.7 Lipid-regulating enzyme activity
To measure the lipid-regulating enzymes activities in the epi-didymal WAT, samples were prepared and analyzed as previ-ously described [24]. Briefly, fatty acid synthase (FAS) activitywas determined with a spectrophotometric assay accordingto the method by Carl et al. [25]; one unit of FAS activity rep-resented the oxidation of 1 nmol of NADPH per minute at30�C. Carnitine palmitoyltransferase activity was determinedaccording to the method by Markwell et al. [26] and the re-sults were expressed as nmol/min/mg protein. Fatty acid�-oxidation was measured spectrophotometrically by moni-toring the reduction of NAD to NADH in the presence ofpalmitoyl-CoA as described by Lazarow [27], with slight modi-fication. Protein concentration was measured by the Bradfordmethod using BSA as the standard [28].
2.8 Analysis of gene expression
Epididymal WAT and liver were homogenized in TRIzolreagent (Invitrogen Life Technologies, Grand Island, NY,USA) and total RNA was isolated according to the manufac-turer’s instructions. The total RNA was converted to cDNA us-ing the QuantiTect Reverse Transcription Kit (Qiagen Gmbh,Hilden, Germany). The RNA expression was quantified byquantitative real-time PCR using the QuantiTect SYBR greenPCR kit (Qiagen Gmbh) and the SDS7000 sequence-detectionsystem (Applied Biosystems, CA, USA). Each cDNA samplewas amplified using primers labeled with SYBR Green dyefor glyceraldehyde-3-phosphate dehydrogenase. The amplifi-cation was performed as follows: 10 min at 90�C, 15 s at 95�C,and 60 s at 60�C for a total of 40 cycles. The cycle threshold val-ues obtained were those cycles at which a statistically signifi-cant increase in the SYBR green emission intensity occurred.Ct data were normalized using glyceraldehyde-3-phosphate
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com
4 Y.-J. Kim et al. Mol. Nutr. Food Res. 2013, 00, 1–11
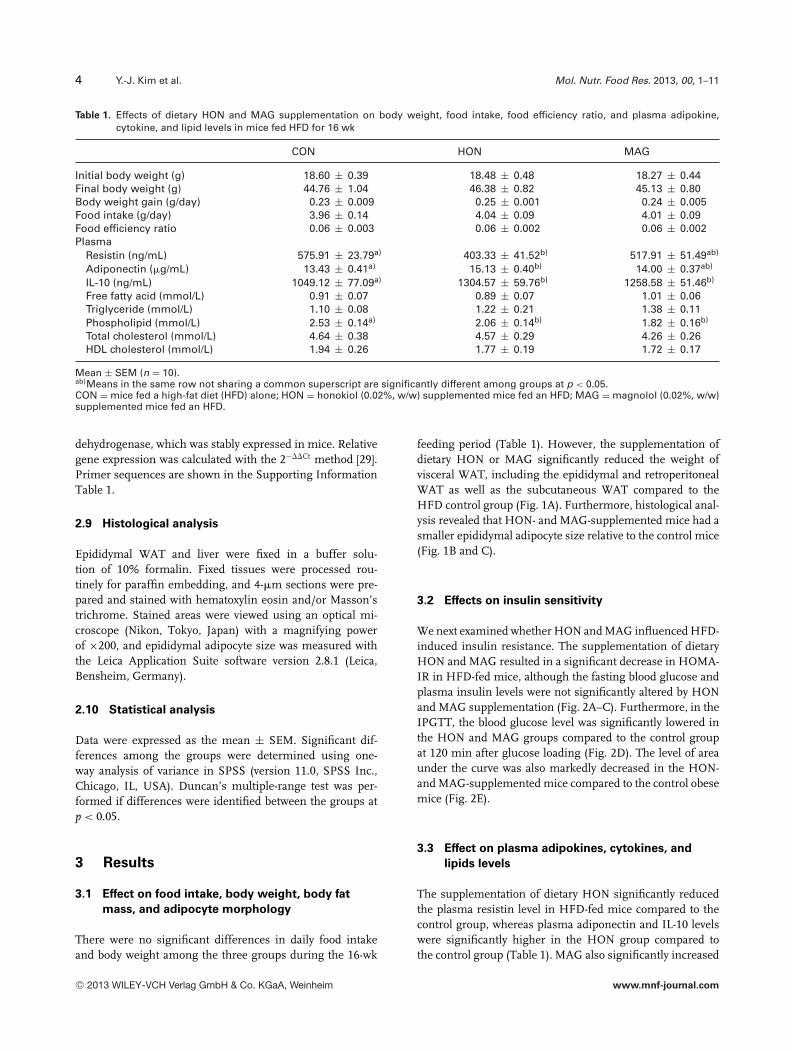
Table 1. Effects of dietary HON and MAG supplementation on body weight, food intake, food efficiency ratio, and plasma adipokine,cytokine, and lipid levels in mice fed HFD for 16 wk
CON HON MAG
Initial body weight (g) 18.60 ± 0.39 18.48 ± 0.48 18.27 ± 0.44Final body weight (g) 44.76 ± 1.04 46.38 ± 0.82 45.13 ± 0.80Body weight gain (g/day) 0.23 ± 0.009 0.25 ± 0.001 0.24 ± 0.005Food intake (g/day) 3.96 ± 0.14 4.04 ± 0.09 4.01 ± 0.09Food efficiency ratio 0.06 ± 0.003 0.06 ± 0.002 0.06 ± 0.002Plasma
Resistin (ng/mL) 575.91 ± 23.79a) 403.33 ± 41.52b) 517.91 ± 51.49ab)
Adiponectin (�g/mL) 13.43 ± 0.41a) 15.13 ± 0.40b) 14.00 ± 0.37ab)
IL-10 (ng/mL) 1049.12 ± 77.09a) 1304.57 ± 59.76b) 1258.58 ± 51.46b)
Free fatty acid (mmol/L) 0.91 ± 0.07 0.89 ± 0.07 1.01 ± 0.06Triglyceride (mmol/L) 1.10 ± 0.08 1.22 ± 0.21 1.38 ± 0.11Phospholipid (mmol/L) 2.53 ± 0.14a) 2.06 ± 0.14b) 1.82 ± 0.16b)
Total cholesterol (mmol/L) 4.64 ± 0.38 4.57 ± 0.29 4.26 ± 0.26HDL cholesterol (mmol/L) 1.94 ± 0.26 1.77 ± 0.19 1.72 ± 0.17
Mean ± SEM (n = 10).ab)Means in the same row not sharing a common superscript are significantly different among groups at p < 0.05.CON = mice fed a high-fat diet (HFD) alone; HON = honokiol (0.02%, w/w) supplemented mice fed an HFD; MAG = magnolol (0.02%, w/w)supplemented mice fed an HFD.
dehydrogenase, which was stably expressed in mice. Relativegene expression was calculated with the 2−��Ct method [29].Primer sequences are shown in the Supporting InformationTable 1.
2.9 Histological analysis
Epididymal WAT and liver were fixed in a buffer solu-tion of 10% formalin. Fixed tissues were processed rou-tinely for paraffin embedding, and 4-�m sections were pre-pared and stained with hematoxylin eosin and/or Masson’strichrome. Stained areas were viewed using an optical mi-croscope (Nikon, Tokyo, Japan) with a magnifying powerof ×200, and epididymal adipocyte size was measured withthe Leica Application Suite software version 2.8.1 (Leica,Bensheim, Germany).
2.10 Statistical analysis
Data were expressed as the mean ± SEM. Significant dif-ferences among the groups were determined using one-way analysis of variance in SPSS (version 11.0, SPSS Inc.,Chicago, IL, USA). Duncan’s multiple-range test was per-formed if differences were identified between the groups atp < 0.05.
3 Results
3.1 Effect on food intake, body weight, body fat
mass, and adipocyte morphology
There were no significant differences in daily food intakeand body weight among the three groups during the 16-wk
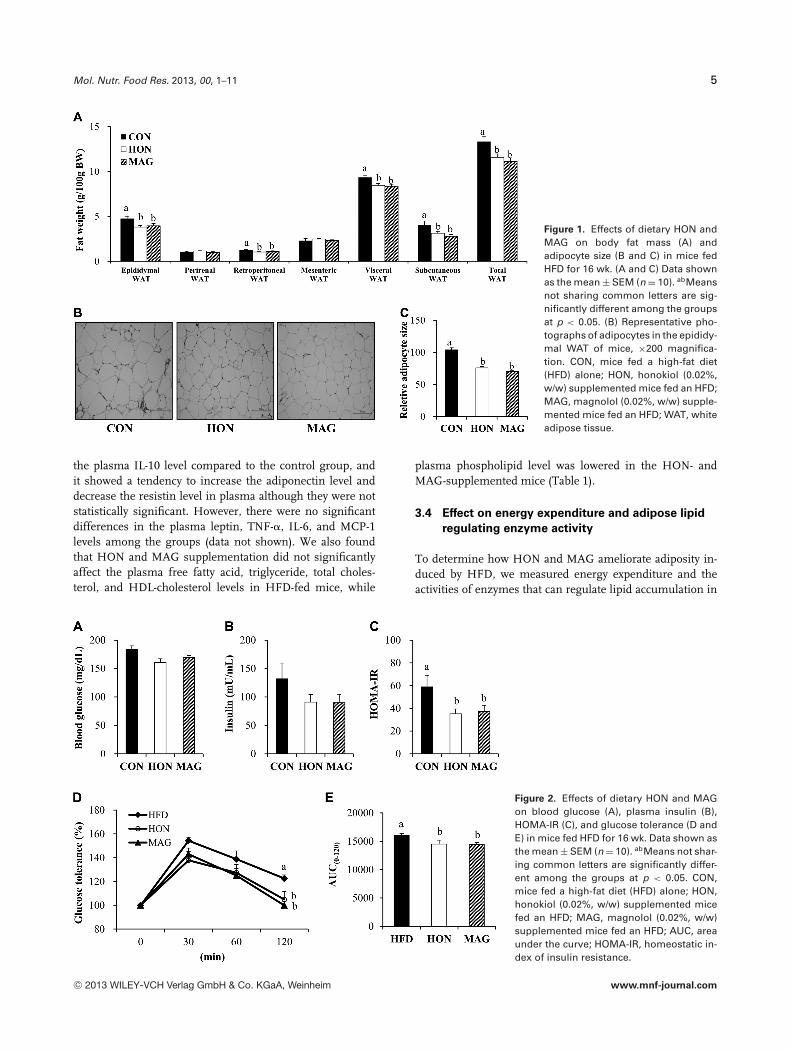
feeding period (Table 1). However, the supplementation ofdietary HON or MAG significantly reduced the weight ofvisceral WAT, including the epididymal and retroperitonealWAT as well as the subcutaneous WAT compared to theHFD control group (Fig. 1A). Furthermore, histological anal-ysis revealed that HON- and MAG-supplemented mice had asmaller epididymal adipocyte size relative to the control mice(Fig. 1B and C).
3.2 Effects on insulin sensitivity
We next examined whether HON and MAG influenced HFD-induced insulin resistance. The supplementation of dietaryHON and MAG resulted in a significant decrease in HOMA-IR in HFD-fed mice, although the fasting blood glucose andplasma insulin levels were not significantly altered by HONand MAG supplementation (Fig. 2A–C). Furthermore, in theIPGTT, the blood glucose level was significantly lowered inthe HON and MAG groups compared to the control groupat 120 min after glucose loading (Fig. 2D). The level of areaunder the curve was also markedly decreased in the HON-and MAG-supplemented mice compared to the control obesemice (Fig. 2E).
3.3 Effect on plasma adipokines, cytokines, and
lipids levels
The supplementation of dietary HON significantly reducedthe plasma resistin level in HFD-fed mice compared to thecontrol group, whereas plasma adiponectin and IL-10 levelswere significantly higher in the HON group compared tothe control group (Table 1). MAG also significantly increased
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com
Mol. Nutr. Food Res. 2013, 00, 1–11 5
Figure 1. Effects of dietary HON andMAG on body fat mass (A) andadipocyte size (B and C) in mice fedHFD for 16 wk. (A and C) Data shownas the mean ± SEM (n = 10). abMeansnot sharing common letters are sig-nificantly different among the groupsat p < 0.05. (B) Representative pho-tographs of adipocytes in the epididy-mal WAT of mice, ×200 magnifica-tion. CON, mice fed a high-fat diet(HFD) alone; HON, honokiol (0.02%,w/w) supplemented mice fed an HFD;MAG, magnolol (0.02%, w/w) supple-mented mice fed an HFD; WAT, whiteadipose tissue.
the plasma IL-10 level compared to the control group, andit showed a tendency to increase the adiponectin level anddecrease the resistin level in plasma although they were notstatistically significant. However, there were no significantdifferences in the plasma leptin, TNF-�, IL-6, and MCP-1levels among the groups (data not shown). We also foundthat HON and MAG supplementation did not significantlyaffect the plasma free fatty acid, triglyceride, total choles-terol, and HDL-cholesterol levels in HFD-fed mice, while
plasma phospholipid level was lowered in the HON- andMAG-supplemented mice (Table 1).
3.4 Effect on energy expenditure and adipose lipid
regulating enzyme activity
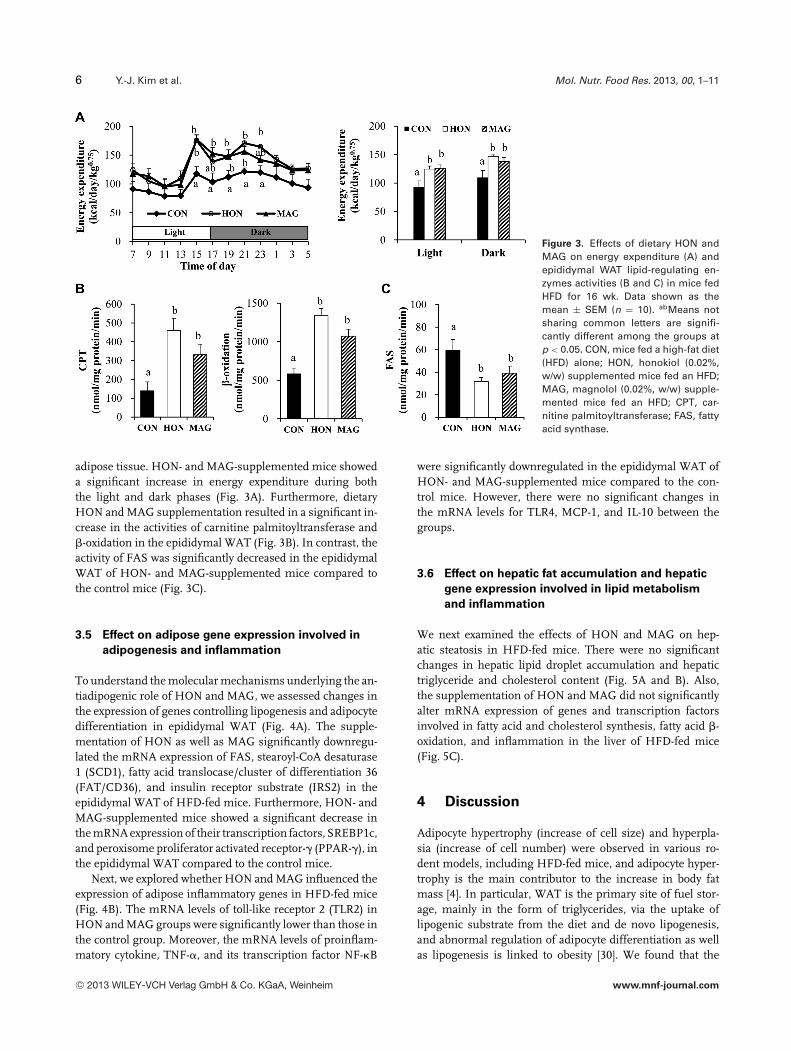
To determine how HON and MAG ameliorate adiposity in-duced by HFD, we measured energy expenditure and theactivities of enzymes that can regulate lipid accumulation in
Figure 2. Effects of dietary HON and MAGon blood glucose (A), plasma insulin (B),HOMA-IR (C), and glucose tolerance (D andE) in mice fed HFD for 16 wk. Data shown asthe mean ± SEM (n = 10). abMeans not shar-ing common letters are significantly differ-ent among the groups at p < 0.05. CON,mice fed a high-fat diet (HFD) alone; HON,honokiol (0.02%, w/w) supplemented micefed an HFD; MAG, magnolol (0.02%, w/w)supplemented mice fed an HFD; AUC, areaunder the curve; HOMA-IR, homeostatic in-dex of insulin resistance.
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com
6 Y.-J. Kim et al. Mol. Nutr. Food Res. 2013, 00, 1–11
Figure 3. Effects of dietary HON andMAG on energy expenditure (A) andepididymal WAT lipid-regulating en-zymes activities (B and C) in mice fedHFD for 16 wk. Data shown as themean ± SEM (n = 10). abMeans notsharing common letters are signifi-cantly different among the groups atp < 0.05. CON, mice fed a high-fat diet(HFD) alone; HON, honokiol (0.02%,w/w) supplemented mice fed an HFD;MAG, magnolol (0.02%, w/w) supple-mented mice fed an HFD; CPT, car-nitine palmitoyltransferase; FAS, fattyacid synthase.
adipose tissue. HON- and MAG-supplemented mice showeda significant increase in energy expenditure during boththe light and dark phases (Fig. 3A). Furthermore, dietaryHON and MAG supplementation resulted in a significant in-crease in the activities of carnitine palmitoyltransferase and�-oxidation in the epididymal WAT (Fig. 3B). In contrast, theactivity of FAS was significantly decreased in the epididymalWAT of HON- and MAG-supplemented mice compared tothe control mice (Fig. 3C).
3.5 Effect on adipose gene expression involved in
adipogenesis and inflammation
To understand the molecular mechanisms underlying the an-tiadipogenic role of HON and MAG, we assessed changes inthe expression of genes controlling lipogenesis and adipocytedifferentiation in epididymal WAT (Fig. 4A). The supple-mentation of HON as well as MAG significantly downregu-lated the mRNA expression of FAS, stearoyl-CoA desaturase1 (SCD1), fatty acid translocase/cluster of differentiation 36(FAT/CD36), and insulin receptor substrate (IRS2) in theepididymal WAT of HFD-fed mice. Furthermore, HON- andMAG-supplemented mice showed a significant decrease inthe mRNA expression of their transcription factors, SREBP1c,and peroxisome proliferator activated receptor-� (PPAR-�), inthe epididymal WAT compared to the control mice.
Next, we explored whether HON and MAG influenced theexpression of adipose inflammatory genes in HFD-fed mice(Fig. 4B). The mRNA levels of toll-like receptor 2 (TLR2) inHON and MAG groups were significantly lower than those inthe control group. Moreover, the mRNA levels of proinflam-matory cytokine, TNF-�, and its transcription factor NF-�B
were significantly downregulated in the epididymal WAT ofHON- and MAG-supplemented mice compared to the con-trol mice. However, there were no significant changes inthe mRNA levels for TLR4, MCP-1, and IL-10 between thegroups.
3.6 Effect on hepatic fat accumulation and hepatic
gene expression involved in lipid metabolism
and inflammation
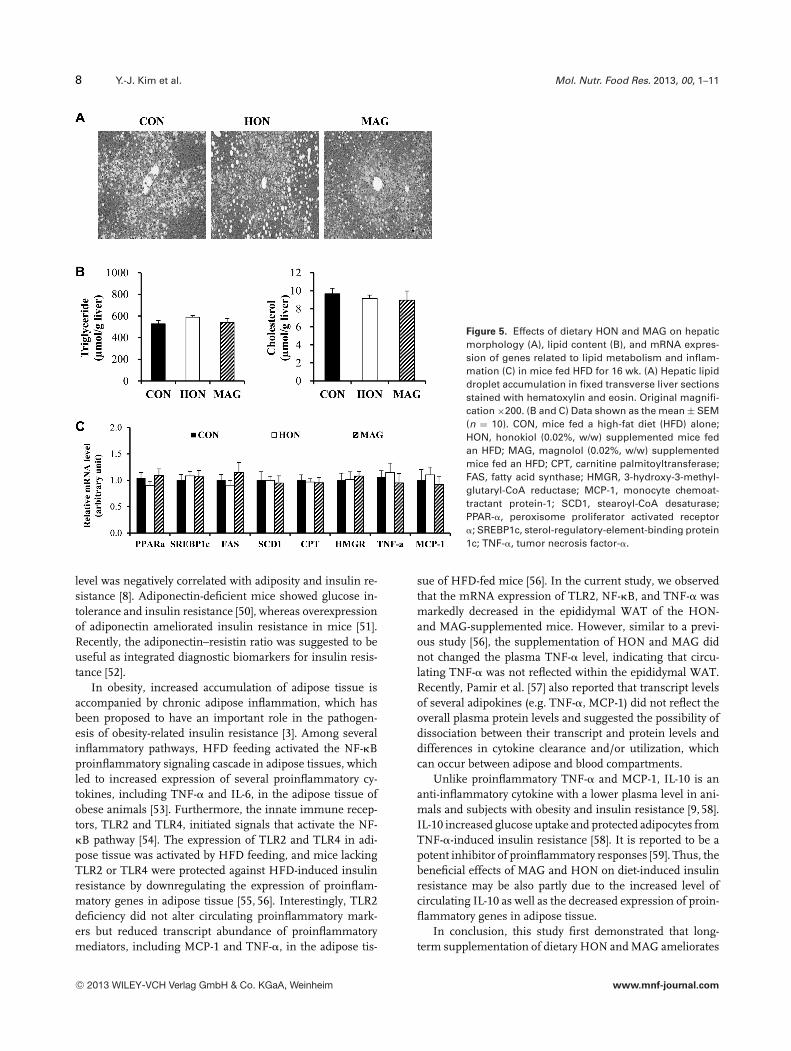
We next examined the effects of HON and MAG on hep-atic steatosis in HFD-fed mice. There were no significantchanges in hepatic lipid droplet accumulation and hepatictriglyceride and cholesterol content (Fig. 5A and B). Also,the supplementation of HON and MAG did not significantlyalter mRNA expression of genes and transcription factorsinvolved in fatty acid and cholesterol synthesis, fatty acid �-oxidation, and inflammation in the liver of HFD-fed mice(Fig. 5C).
4 Discussion
Adipocyte hypertrophy (increase of cell size) and hyperpla-sia (increase of cell number) were observed in various ro-dent models, including HFD-fed mice, and adipocyte hyper-trophy is the main contributor to the increase in body fatmass [4]. In particular, WAT is the primary site of fuel stor-age, mainly in the form of triglycerides, via the uptake oflipogenic substrate from the diet and de novo lipogenesis,and abnormal regulation of adipocyte differentiation as wellas lipogenesis is linked to obesity [30]. We found that the
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

Mol. Nutr. Food Res. 2013, 00, 1–11 7
Figure 4. Effects of dietary HON and MAG on mRNAexpression of genes related to adipogenesis (A) andinflammatory response (B) in epididymal WAT of micefed HFD for 16 wk. Data shown as the mean ± SEM(n = 10). abMeans not sharing common letters aresignificantly different among the groups at p < 0.05.CON, mice fed a high-fat diet (HFD) alone; HON, hon-okiol (0.02%, w/w) supplemented mice fed an HFD;MAG, magnolol (0.02%, w/w) supplemented mice fedan HFD; FAS, fatty acid synthase; FAT/CD36, fattyacid translocase/cluster of differentiation 36; IRS2,insulin receptor substrate 2; MCP-1, monocytechemoattractant protein-1; NF-�B, nuclear factorkappa-light-chain-enhancer of activated B cells; SCD1,stearoyl-CoA desaturase; PPAR-�, peroxisome pro-liferator activated receptor �; SREBP1c, sterol-regulatory-element-binding protein 1c; TLR2, toll-likereceptor 2; TNF-�, tumor necrosis factor-�.
decrease of adipocyte size and body fat mass in HON- andMAG-supplemented mice was associated with a significantdecrease in activity of FAS and in the mRNA expression of keygenes involved in fatty acid synthesis (FAS, SCD1) and up-take (FAT/CD36) as well as adipocyte differentiation (IRS2)in epididymal WAT. Interestingly, HON and MAG did not al-ter body weight, which may be due to an increase in lean bodymass or bone mass by these phytonutrients [31,32], althoughwe did not determine it.
The SREBP1c is highly expressed in adipose tissue andplays a central role in the induction of lipogenic genes suchas FAS and SCD1 that catalyze the de novo synthesis of sat-urated and monounsaturated fatty acids, respectively [33,34].The expression of FAS was increased in the adipose tissueof both genetic and nutritional obese mice as well as pa-tients with obesity and type 2 diabetes, whereas mice lackingFAS or SCD1 prevented obesity induced by HFD [35, 36].PPAR-� is another important adipocyte transcription factorthat can regulate the expression of a number of genes in-volved in lipid metabolism and adipocyte differentiation. Thefatty acid transporter FAT/CD36, one of the target genes ofPPAR-�, controls circulating fatty acids uptake into adiposetissue [37]. The mRNA level of FAT/CD36 was increased inthe adipose tissue of animals with obesity and insulin resis-tance [38], whereas FAT/CD36 deficiency protected againstadiposity and body weight gain in HFD-fed mice [37]. IRS2gene expression was also increased in 3T3-L1 adipocytes andhuman primary adipocytes in response to the PPAR-� ac-tivator [39]. Miki et al. [40] have suggested that IRS2 playsa critical role in adipocyte differentiation through PPAR-�activation. PPAR-� activation was also related to the expres-sion of FAS and SCD1 in adipose tissue [35, 41]. Taken to-gether, our observation suggests that the downregulation ofSREBP1c and PPAR-� gene expression by HON and MAG
supplementation may have numerous effects on various adi-pogenic pathways regulated by these transcription factors,including those that support lipogenesis and adipose dif-ferentiation. Furthermore, the protection from adiposity ob-served in HON- and MAG-supplemented mice may also beexplained by the increased energy expenditure and fatty acidoxidation. In contrast to our in vivo findings, in vitro proadi-pogenic effects of HON and MAG have been reported [42–44],which may be due to the fact that the physiological con-ditions and complex interactions are absent in the in vitrosystem.
Excess adiposity, especially in visceral WAT, is consid-ered to cause insulin resistance [4]. Although PPAR-� ago-nists such as thiazolidinedione acted as insulin sensitizer bypotentiating insulin signaling in adipocytes [45], decreasingPPAR-� activity, either by PPAR-� antagonist treatment orby heterozygous PPAR-� deficiency, protected against HFD-induced adipocyte insulin resistance as well as hypertrophy inmice [46] as observed in the HON- and MAG-supplementedmice. Similarly, mice lacking FAS or SCD-1 either in all cellsor specifically in adipocytes were protected from the deteri-oration of glucose homeostasis induced by HFD [34, 36, 47].In addition, enhanced �-oxidation increased insulin sensitiv-ity [48]. Thus, these results indicate that the antiadipogeniceffects of HON and MAG partially account for overall im-provement in insulin resistance.
In particular, the increase in insulin sensitivity by HONsupplementation may also be related to the increased level ofadiponectin and decreased level of resistin in plasma, sinceadiponectin and resistin have been considered as a poten-tial link between obesity and insulin resistance [6, 8, 49–51].Serum resistin was elevated in obesity and insulin resis-tance [6], and resistin deficiency in mice led to improvedglucose homeostasis [49]. Conversely, circulating adiponectin
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com
8 Y.-J. Kim et al. Mol. Nutr. Food Res. 2013, 00, 1–11
Figure 5. Effects of dietary HON and MAG on hepaticmorphology (A), lipid content (B), and mRNA expres-sion of genes related to lipid metabolism and inflam-mation (C) in mice fed HFD for 16 wk. (A) Hepatic lipiddroplet accumulation in fixed transverse liver sectionsstained with hematoxylin and eosin. Original magnifi-cation ×200. (B and C) Data shown as the mean ± SEM(n = 10). CON, mice fed a high-fat diet (HFD) alone;HON, honokiol (0.02%, w/w) supplemented mice fedan HFD; MAG, magnolol (0.02%, w/w) supplementedmice fed an HFD; CPT, carnitine palmitoyltransferase;FAS, fatty acid synthase; HMGR, 3-hydroxy-3-methyl-glutaryl-CoA reductase; MCP-1, monocyte chemoat-tractant protein-1; SCD1, stearoyl-CoA desaturase;PPAR-�, peroxisome proliferator activated receptor�; SREBP1c, sterol-regulatory-element-binding protein1c; TNF-�, tumor necrosis factor-�.
level was negatively correlated with adiposity and insulin re-sistance [8]. Adiponectin-deficient mice showed glucose in-tolerance and insulin resistance [50], whereas overexpressionof adiponectin ameliorated insulin resistance in mice [51].Recently, the adiponectin–resistin ratio was suggested to beuseful as integrated diagnostic biomarkers for insulin resis-tance [52].
In obesity, increased accumulation of adipose tissue isaccompanied by chronic adipose inflammation, which hasbeen proposed to have an important role in the pathogen-esis of obesity-related insulin resistance [3]. Among severalinflammatory pathways, HFD feeding activated the NF-�Bproinflammatory signaling cascade in adipose tissues, whichled to increased expression of several proinflammatory cy-tokines, including TNF-� and IL-6, in the adipose tissue ofobese animals [53]. Furthermore, the innate immune recep-tors, TLR2 and TLR4, initiated signals that activate the NF-�B pathway [54]. The expression of TLR2 and TLR4 in adi-pose tissue was activated by HFD feeding, and mice lackingTLR2 or TLR4 were protected against HFD-induced insulinresistance by downregulating the expression of proinflam-matory genes in adipose tissue [55, 56]. Interestingly, TLR2deficiency did not alter circulating proinflammatory mark-ers but reduced transcript abundance of proinflammatorymediators, including MCP-1 and TNF-�, in the adipose tis-
sue of HFD-fed mice [56]. In the current study, we observedthat the mRNA expression of TLR2, NF-�B, and TNF-� wasmarkedly decreased in the epididymal WAT of the HON-and MAG-supplemented mice. However, similar to a previ-ous study [56], the supplementation of HON and MAG didnot changed the plasma TNF-� level, indicating that circu-lating TNF-� was not reflected within the epididymal WAT.Recently, Pamir et al. [57] also reported that transcript levelsof several adipokines (e.g. TNF-�, MCP-1) did not reflect theoverall plasma protein levels and suggested the possibility ofdissociation between their transcript and protein levels anddifferences in cytokine clearance and/or utilization, whichcan occur between adipose and blood compartments.
Unlike proinflammatory TNF-� and MCP-1, IL-10 is ananti-inflammatory cytokine with a lower plasma level in ani-mals and subjects with obesity and insulin resistance [9, 58].IL-10 increased glucose uptake and protected adipocytes fromTNF-�-induced insulin resistance [58]. It is reported to be apotent inhibitor of proinflammatory responses [59]. Thus, thebeneficial effects of MAG and HON on diet-induced insulinresistance may be also partly due to the increased level ofcirculating IL-10 as well as the decreased expression of proin-flammatory genes in adipose tissue.
In conclusion, this study first demonstrated that long-term supplementation of dietary HON and MAG ameliorates
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

Mol. Nutr. Food Res. 2013, 00, 1–11 9
Figure 6. Proposed mechanisms of HON and MAG for antiobesity. The supplementation of HON and MAG downregulates mRNA expressionof key adipogenic transcription factors, SREBP1c and PPAR-�, which in turn decreases mRNA expression of their target genes involved infatty acid synthesis, fatty acid uptake, and adipocyte differentiation, as well as FAS activity in the epididymal WAT, and simultaneouslyincreases energy expenditure along with the activities of CPT and �-oxidation in the epididymal WAT, indicating that these changes maybe potential mechanisms for improving body fat accumulation and insulin resistance induced by HFD. Furthermore, the antiadiposity andinsulin-sensitizing action of HON and MAG may be associated with the increased anti-inflammatory IL-10 level in plasma and downregulatedmRNA expression of proinflammatory transcription factor NF-�B and its target gene TNF-� as well as TLR2 in epididymal WAT. CPT,carnitine palmitoyltransferase; FAS, fatty acid synthase; FAT/CD36, fatty acid translocase/cluster of differentiation 36; IRS2, insulin receptorsubstrate 2; NF-�B, nuclear factor kappa-light-chain-enhancer of activated B cells; PPAR-�, peroxisome proliferator activated receptor �;SCD1, stearoyl-CoA desaturase; SREBP1c, sterol-regulatory-element-binding protein 1c; TLR2, toll-like receptor 2; TNF-�, tumor necrosisfactor-�.
adipogenesis in mice fed HFD by increasing energy expen-diture and visceral WAT fatty acid oxidation and a simulta-neous decrease in the expression of key genes involved inadipogenesis as well as enzyme activity for fatty acid synthe-sis in visceral WAT. Furthermore, HON and MAG exhibiteda protective role against insulin resistance induced by HFD.These changes were partly associated with the downregula-tion of proinflammatory gene expression in visceral WAT andthe increase of the anti-inflammatory IL-10 level in plasma.Figure 6 illustrates the possible mechanisms of the HON andMAG effects for antiobesity. Thus, our findings suggest thatHON and MAG may be useful novel agents for the control ofobesity and obesity-related insulin resistance.
This research was supported by the Basic Science Research Pro-gram (number 2011-0022387) and the SRC program (Center forFood & Nutritional Genomics: number 2012-0000644) throughthe National Research Foundation of Korea (NRF) funded by theMinistry of Education, Science and Technology.
The authors have declared no conflict of interest.
5 References
[1] Rossner, S., Obesity: the disease of the twenty-first century.Int. J. Obes. 2002, 26, S2–S4.
[2] Gallagher, E.-J., Leroith, D., Karnieli, E., Insulin resistance inobesity as the underlying cause for the metabolic syndrome.Mt. Sinai. J. Med. 2010, 77, 511–523.
[3] Wellen, K.-E., Hotamisligil, G.-S., Inflammation, stress, anddiabetes. J. Clin. Invest. 2005, 5, 1111–1119.
[4] Jo, J., Gavrilova, O., Pack, S., Jou, W. et al., Hypertro-phy and/or hyperplasia: dynamics of adipose tissue growth.PLoS Comput. Biol. 2009, 5, e1000324.
[5] Antuna-Puente, B., Feve, B., Fellahi, S., Bastard, J.-P.,Adipokines: the missing link between insulin resistance andobesity. Diabetes Metab. 2008, 34, 2–11.
[6] Steppan, C.-M., Bailey, S.-T., Bhat, S., Brown, E.-J. et al., Thehormone resistin links obesity to diabetes. Nature 2001, 409,307–312.
[7] Chiang, S.-H., Bazuine, M., Lumeng, C.-N., Geletka, L.-M.et al., The protein kinase IKKepsilon regulates energy bal-ance in obese mice. Cell 2009, 138, 961–975.
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

10 Y.-J. Kim et al. Mol. Nutr. Food Res. 2013, 00, 1–11
[8] Weyer, C., Hypoadiponectinemia in obesity and type 2diabetes: close association with insulin resistance andhyperinsulinemia. J. Clin. Endocrinol. Metab. 2001, 86,1930–1935.
[9] van Exel, E., Gussekloo, J., de Craen, A.-J., Frolich, M. et al.,Low production capacity of interleukin-10 associates withthe metabolic syndrome and type 2 diabetes: the Leiden85-Plus Study. Diabetes 2002, 51, 1088–1092.
[10] Calder, P.-C., Ahluwalia, N., Brouns, F., Buetler, T., Dietary fac-tors and low-grade inflammation in relation to overweightand obesity. Br. J. Nutr. 2011, 106, S5–S78.
[11] Heidemann, C., Scheidt-Nave, C., Richter, A., Mensink, G.-B.,Dietary patterns are associated with cardiometabolic risk fac-tors in a representative study population of German adults.Br. J. Nutr. 2011, 106, 1253–1262.
[12] Mors, W.-B., Rizzini, C.-T., Pereira, N.-A. (Eds.), MedicinalPlants of Brazil. Reference Publication, Inc., Michigan, 2002.
[13] Tomas, S. C.-Li. (Ed.), Chinese and Related North AmericanHerbs: Phytopharmacology and Therapeutic Values. CRCPress, Boca Raton, FL 2002.
[14] Maruyama, Y., Kuribara, H., Overview of the pharmaco-logical features of honokiol. CNS Drug. Rev. 2000, 6,35–44.
[15] Lee, J., Jung, E., Park, J., Jung, K., Anti-inflammatory effectsof magnolol and honokiol are mediated through inhibitionof the downstream pathway of MEKK-1 in NF-kappaB acti-vation signaling. Planta. Med. 2005, 71, 338–343.
[16] Sohn, E.-J., Kim, C.-S., Kim, Y.-S., Jung, D.-H., Effects ofmagnolol (5,5_-diallyl-2,2_-dihydroxybiphenyl) on diabeticnephropathy in type 2 diabetic Goto-Kakizaki rats. Life Sci.2007, 80, 468–475.
[17] Choi, S.-S., Cha, B.-Y., Lee, Y.-S., Yonezawa, T. et al., Magnololenhances adipocyte differentiation and glucose uptake in3T3-L1 cells. Life Sci. 2009, 84, 908–914.
[18] Choi, S.-S., Cha, B.-Y., Iida, K., Sato, M. et al., Honokiolenhances adipocyte differentiation by potentiating insulinsignaling in 3T3-L1 preadipocytes. J. Nat. Med. 2011, 65,424–430.
[19] Yin, H.-Q., Kim, Y.-C., Chung, Y.-S., Kim, Y.-C. et al., Honokiolreverses alcoholic fatty liver by inhibiting the maturationof sterol regulatory element binding protein-1c and the ex-pression of its downstream lipogenesis genes. Toxicol. Appl.Pharmacol. 2009, 236, 124–130.
[20] Murakami, Y., Kawata, A., Seki, Y., Koh, T. et al., Comparativeinhibitory effects of magnolol, honokiol, eugenol and bis-eugenol on cyclooxygenase-2 expression and nuclear factor-kappa b activation in RAW264.7 macrophage-like cells stim-ulated with fimbriae of Porphyromonas gingivalis. In Vivo2012, 26, 941–950.
[21] Collins, S., Martin, T.-L., Surwit, R.-S., Robidoux, J., Geneticvulnerability to diet-induced obesity in the C57BL/6J mouse:physiological and molecular characteristics. Physiol. Behav.2004, 81, 243–248.
[22] Choi, S.-S., Cha, B.-Y., Iida, K., Sato, M. et al., Hon-okiol enhances adipocyte differentiation by potentiating in-sulin signaling in 3T3-L1 preadipocytes. J. Nat. Med. 2011,65, 424–430.
[23] Folch, J., Lees, M., Sloan-Stanley, G. H., A simple method forisolation and purification of total lipids from animal tissues.J. Biol. Chem. 1957, 226, 497–509.
[24] Kim, H.-J., Lee, K.-T., Park, Y.-B., Jeon, S.-M. et al., Dietary do-cosahexaenoic acid-rich diacylglycerols ameliorate hepaticsteatosis and alter hepatic gene expressions in C57BL/6J-Lep(ob/ob) mice. Mol. Nutr. Food Res. 2008, 52, 965–973.
[25] Carl, M.-N., Lakshmanan, M.-R., Porter, J.-W., Fatty acid syn-thase from rat liver. Meth. Enzymol. 1975, 35, 37–44.
[26] Markwell M, A.-K., McGroarty, E.-J., Bieber, L.-L., Tolbert, N.-E., The subcellular distribution of carnitine acyltransferasesin mammalian liver and kidney. J. Biol. Chem. 1973, 248,3426–3432.
[27] Lazarow, P.-B., Assay of peroxisomal �-oxidation of fattyacids. Meth. Enzymol. 1981, 72, 315–319.
[28] Bradford, M.-M., A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein-dye binding. Anal. Biochem. 1976, 72,248–254.
[29] Schmittgen, T.-D., Livak, K.-J., Analyzing real-time PCR databy the comparative C(T) method. Nat. Protoc. 2008, 3,1101–1108.
[30] Moller, D.-E., Flier, J., Insulin resistance-mechanism, syn-dromes, and implications. N. Engl. J. Med. 1991, 325,938–948.
[31] Anderson, R. A., Chromium and polyphenols from cinnamonimprove insulin sensitivity. Proc. Nutr. Soc. 2008, 67, 48–53.
[32] Sacco, S. M., Horcajada, M. N., Offord, E., Phytonutrients forbone health during ageing. Br. J. Clin. Pharmacol. 2013, 75,697–707.
[33] Kim, J.-B., Spiegelman, B.-M., ADD1/SREBP1 promotesadipocyte differentiation and gene expression linked to fattyacid metabolism. Genes Dev. 1996, 10, 1096–1107.
[34] Sampath, H., Miyazaki, M., Dobrzyn, A., Ntambi, J.-M.,Stearoyl-CoA desaturase-1 mediates the pro-lipogenic ef-fects of dietary saturated fat. J. Biol. Chem. 2007, 282,2483–2493.
[35] Lodhi, I.-J., Yin, L., Jensen-Urstad, A.-P., Funai, K. et al., In-hibiting adipose tissue lipogenesis reprograms thermogen-esis and PPAR-� activation to decrease diet-induced obesity.Cell Metab. 2012, 16, 189–201.
[36] Ntambi, J.-M., Miyazaki, M., Stoehr, J.-P., Lan, H. et al., Lossof stearoyl-CoA desaturase-1 function protects mice againstadiposity. Proc. Natl. Acad. Sci. USA 2002, 99, 11482–11486.
[37] Hajri, T., Hall, A.-M., Jensen, D.-R., Pietka, T.-A. et al.,CD36-facilitated fatty acid uptake inhibits leptin produc-tion and signaling in adipose tissue. Diabetes 2007, 56,1872–1880.
[38] Priego, T., Sanchez, J., Palou, A., Pico, C., Leptin intake dur-ing the suckling period improves the metabolic responseof adipose tissue to a high-fat diet. Int. J. Obes. 2010, 34,809–819.
[39] Smith, U., Gogg, S., Johansson, A., Olausson, T. et al., Thi-azolidinediones (PPARgamma agonists) but not PPARalphaagonists increase IRS-2 gene expression in 3T3–L1 and hu-man adipocytes. Faseb. J. 2001, 15, 215–220.
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

Mol. Nutr. Food Res. 2013, 00, 1–11 11
[40] Miki, H., Yamauchi, T., Suzuki, R., Komeda, K. et al., Es-sential role of insulin receptor substrate 1 (IRS-1) and IRS-2 in adipocyte differentiation. Mol. Cell. Biol. 2001, 21,2521–2532.
[41] Yao-Borengasser, A., Rassouli, N., Varma, V., Bodles, A.-M.et al., Stearoyl-coenzyme A desaturase 1 gene expression in-creases after pioglitazone treatment and is associated withperoxisomal proliferator-activated receptor-gamma respon-siveness. J. Clin. Endocrinol. Metab. 2008, 93, 4431–4439.
[42] Choi, S.-S., Cha, B.-Y., Lee, Y.-S., Yonezawa, T. et al., Magnololenhances adipocyte differentiation and glucose uptake in3T3-L1 cells. Life Sci. 2009, 84, 908–914.
[43] Alonso-Castro, A.J., Zapata-Bustos, R., Domınguez, F.,Garcıa-Carranca, A. et al., Magnolia dealbata Zucc and itsactive principles honokiol and magnolol stimulate glucoseuptake in murine and human adipocytes using the insulin-signaling pathway. Phytomedicine 2011, 18, 926–933.
[44] Choi, S.-S., Cha, B.-Y., Iida, K., Sato, M. et al., Honokiolenhances adipocyte differentiation by potentiating insulinsignaling in 3T3-L1 preadipocytes. J. Nat. Med. 2011, 65,424–430.
[45] Jiang, G., Dallas-Yang, Q., Li, Z., Szalkowski, D. et al., Potenti-ation of insulin signaling in tissues of Zucker obese rats afteracute and long-term treatment with PPARgamma agonists.Diabetes 2002, 51, 2412–2419.
[46] Rieusset, J., Touri, F., Michalik, L., Escher, P. et al., A newselective peroxisome proliferator-activated receptor gammaantagonist with antiobesity and antidiabetic activity. Mol.Endocrinol. 2002, 16, 2628–2644.
[47] Wueest, S., Rapold, R.-A., Schumann, D.-M., Rytka, J.-M.et al., Deletion of Fas in adipocytes relieves adipose tissueinflammation and hepatic manifestations of obesity in mice.J. Clin. Invest. 2010, 120, 191–202.
[48] Perdomo, G., Commerford, S.-R., Richard, A.-M., Adams,S.-H. et al., Increased beta-oxidation in muscle cells en-hances insulin-stimulated glucose metabolism and pro-tects against fatty acid-induced insulin resistance despite in-tramyocellular lipid accumulation. J. Biol. Chem. 2004, 279,27177–27186.
[49] Banerjee, R.-R., Rangwala, S.-M., Shapiro, J.-S., Rich, A.-S.et al., Regulation of fasted blood glucose by resistin. Science2004, 303, 1195–1198.
[50] Maeda, N., Diet-induced insulin resistance in mice lackingadiponectin/ACRP30. Nat. Med. 2002, 8, 731–737.
[51] Yamauchi, T., Kamon, J., Minokoshi, Y., Ito, Y. et al.,Adiponectin stimulates glucose utilization and fatty-acid oxi-dation by activating AMP-activated protein kinase. Nat. Med.2002, 8, 1288–1295.
[52] Lau, C.-H., Muniandy, S., Novel adiponectin-resistin (AR) andinsulin resistance (IRAR) indexes are useful integrated diag-nostic biomarkers for insulin resistance, type 2 diabetes andmetabolic syndrome: a case control study. Cardiovasc. Dia-betol. 2011, 10, 1–8.
[53] Cho, S., Choi, Y., Park, S., Park, T., Carvacrol prevents diet-induced obesity by modulating gene expressions involvedin adipogenesis and inflammation in mice fed with high-fatdiet. J. Nutri. Biochem. 2011, 23, 192–201.
[54] Medzhitov, R., Toll-like receptors and innate immunity. Nat.Rev. Immunol. 2001, 1, 135–145
[55] Shi, H., Kokoeva, M.-V., Inouye, K., Tzameli, I. et al., TLR4links innate immunity and fatty acid-induced insulin resis-tance. J. Clin. Invest. 2006, 116, 3015–3025.
[56] Davis, J.-E., Braucher, D.-R., Walker-Daniels, J., Spurlock, M.-E., Absence of TLR2 protects against high-fat diet-induced in-flammation and results in greater insulin-stimulated glucosetransport in cultured adipocytes. J. Nutr. Biochem. 2011, 22,136–141.
[57] Pamir, N., McMillen, T.-S., Edgel, K.-A., Kim, F. et al., Defi-ciency of lymphotoxin-� does not exacerbate high-fat diet-induced obesity but does enhance inflammation in mice.Am. J. Physiol. Endocrinol. Metab. 2012, 302, E961–E971.
[58] Lumeng, C.-N., Bodzin, J.-L., Saltiel, A.-R., Obesity inducesa phenotypic switch in adipose tissue macrophage polariza-tion. J. Clin. Invest. 2007, 117, 175–184.
[59] Ouyang, W., Rutz, S., Crellin, N.-K., Valdez, P.-A. et al., Regu-lation and functions of the IL-10 family of cytokines in inflam-mation and disease. Annu. Rev. Immunol. 2011, 29, 71–109.
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com