lecture 2 phylum cnidaria - boston...
TRANSCRIPT

Lecture 2
Phylum CnidariaAnemones, corals, jellyfishes,
hydras and relativesby John R. Finnerty
Animal Phylogeny
PROTOSTOMIA
ChordataHemichordataEchinodermata
Arthropoda OnychophoraNematoda
Deuterostomia
AnnelidaMolluscaPlatyhelminthes
Ecdysozoa Lophotrochozoa
Sili
cisp
ongi
ae
Cal
cisp
ongi
a
Acoelomorpha
Porifera Ctenophora Cnidaria

Phylum Cnidaria Cnidaria (Greek: “stinging thread”) distinguished by the possession of
cnidae 10,000 described species—sea
anemones, corals, jellyfishes and hydras
diploblast = 2 germ layers (ectoderm & endoderm)
blind gut (single opening) nerve net & muscle cells radial symmetry??? sexual & asexual reproduction
Cnidocytes & Cnidae Diagnostic of
cnidarians. However,
both ctenophores (Haeckelia [=Euchlora]) and aeolid nudibranchs may re-deploy cnidae that they have obtained from their cnidarian prey.
(Barnes, Invertebrate Zoology, 1987)

The cnidocyte is a sensory-effector cell containing a cnida. Each cnida is a rounded proteinaceous capsule, with an
opening on the apical surface that is often covered by a hinged operculum.
At the surface, where the cnida opens, there are generally found a number of modified cilia, called cnidocil, which assist in the perception of tactile stimulation and chemical stimulation.
In this sac, there is a long hollow thread. Upon mechanical contact and receipt of appropriate chemical stimuli, this thread is explosively everted from the sac.
The cnidae may serve to deliver venom, like a hypodermic needle. Many “nematocysts” function like this.
The cnidae may also serve to anchor the animal to a substrate or to adhere to a prey item. This adherent role can be performed by various subtypes of nematocysts but also by spirocysts and ptychocysts.
Cnidocytes & Cnidae
(Barnes, Invertebrate Zoology, 1987)
Cnidae
(Pechenik, Biology of the Invertebates, 2000)

Cnidae Discharge
Ca2+
macromolecule(e.g., protein)
cytoplasm of cnidocyte
cnida
stimulationcalcium releaseincrease in osmotic pressurewater rushes in by osmosis
cnida
H2O
cnida
eversion of tubule
Diploblasty Cnidaria diverged from Bilateria prior to the evolution
of Mesoderm. So, Cnidaria lack mesoderm. Cnidaria are diploblasts, having only two germ layers,
the primary germ layers: ectoderm and endoderm. Outer ectodermal epithelium (ectoderm) Inner gastrodermal epithelium (endoderm) Central layer of mesoglea of varying thickness.
Mesoglea is a gelatinous, largely acellular substance. It may have a few living cells within it—often mobile amoeboid cells. However, the cells are not organized into a tissue like true mesoderm. The mesoglea can act as a hydrostatic skeleton,
providing support to the rest of the cnidarian body which is really just two thin layers of epithelium

gastrodermis(endoderm)
epidermis(ectoderm)
mesoglea
mouth
pharynx
basal disc
enteron
Diploblasty
“For those contemplating reincarnation, a major drawback to life as a cnidarian would seem to be the absence of an anus. All undigested food material passes through the same opening through which the food enters: the mouth. This is not particularly appetizing from the human point of view, but the shortcomings of life without an anus are not merely aesthetic. The sequential disassembly of particulate food material that occurs in an open-ended tubular gut is not possible in the cnidarian digestive system and, indeed, the animal must expel the undigested remains of one meal before it can ingest more food.”
— Pechenik, 2001

In the through gut, different functions are localized to different sections of a linear tube.
For example, consider your own digestive tract.
Mouth Stomach Small Bowel
Large Bowel
Mechanical processing
Protein hydrolysis
Protein hydrolysis
Water resorption
Initial carbohydrate digestion
Pepsin(pH < 6)
Trypsin(pH 7-9)
Carbohydrate digestion
In the one way gut, it is widely thought that you cannot have specialized regions of extracellular digestion.
In other words, all extracellular digestion would have to occur in the same physio-chemical environment.
In animals with one-way guts, there tends to be a greater emphasis on intracellular digestion, where undigested food particles are phagocytosed into the cells lining the gut.
However, in both Cnidaria and Ctenophora, there is evidence for distinct gut regions with distinct extracellular environments
One-way gut

Polyp and Medusa
mouthmouth mouth
gastrodermis(endoderm)
pharynx
basal disc (Oliver & Coates, in The Fossil Invertebrates, 1992)
epidermis(ectoderm)
enteron
enteronenteron mesoglea
Medusa = pelagic drifter

Jellyfish are mixing the oceans?
A theoretical model for the relative contributions of Darwinian mixingand turbulent wake mixing is created and validated by in situfield measurements of swimming jellyfish using a newly developedscuba-based laser velocimetry device. Extrapolation of theseresults to other animals is straightforward given knowledge ofthe animal shape and orientation during vertical migration. Onthe basis of calculations of a broad range of aquatic animal species,we conclude that biogenic mixing via Darwin’s mechanism can be asignificant contributor to ocean mixing and nutrient transport.
Polyp = sessile benthic(but capable of some movement)

Cnidarian Nerve Net
Cnidarian Nerve Net

Spread of Excitation in Cnidarian Nerve Net
Cellular Composition

Cnidarian Muscle Histology
(Blanquet and Riordan, 1981)
Scale bar = 1.0 µm
Cnidarian Diversity & Evolution
Class Anthozoa: sea anemones, corals, sea pens, etc.
Class Cubozoa: box jellyfishesClass Scyphozoa: true jellyfishes
ANTHOZOA — polyp body form only; simpler life histories; bilateral and biradial symmetry
Class Hydrozoa: hydras, hydroids, hydromedusae
MEDUSOZOA — most have both polyp and medusa; radial and tetraradial symmetry

Hydrozoan Life-History & Bodyplan Diversity
Polyp Planula Medusa Colony Worm
_Anthomedusae yes yes yes yes no
_Leptomedusae yes yes yes yes no
_Limnomedusae reduced yes yes no no
_Trachymedusae
no yes yes no no
_Hydra yes no no no no
_Siphonophora yes yes yes yes no
_Buddenbrockia no yes no yes yes
_Myxobolus no yes no yes yes
Cnidarian Phylogeny
+
Anthozoa
-
Cubozoa ScyphozoaTrachylinehydrozoa
Otherhydrozoa
(Bridge et al. 1997)
hydras
-
MEDUSOZOA

Sexual Reproduction (Anthozoa)
Planula larvaBroadcast spawning
Polyps
Sexual Reproduction (Medusozoa)
Planula larva
Broadcast spawning
Medusae
Polyps
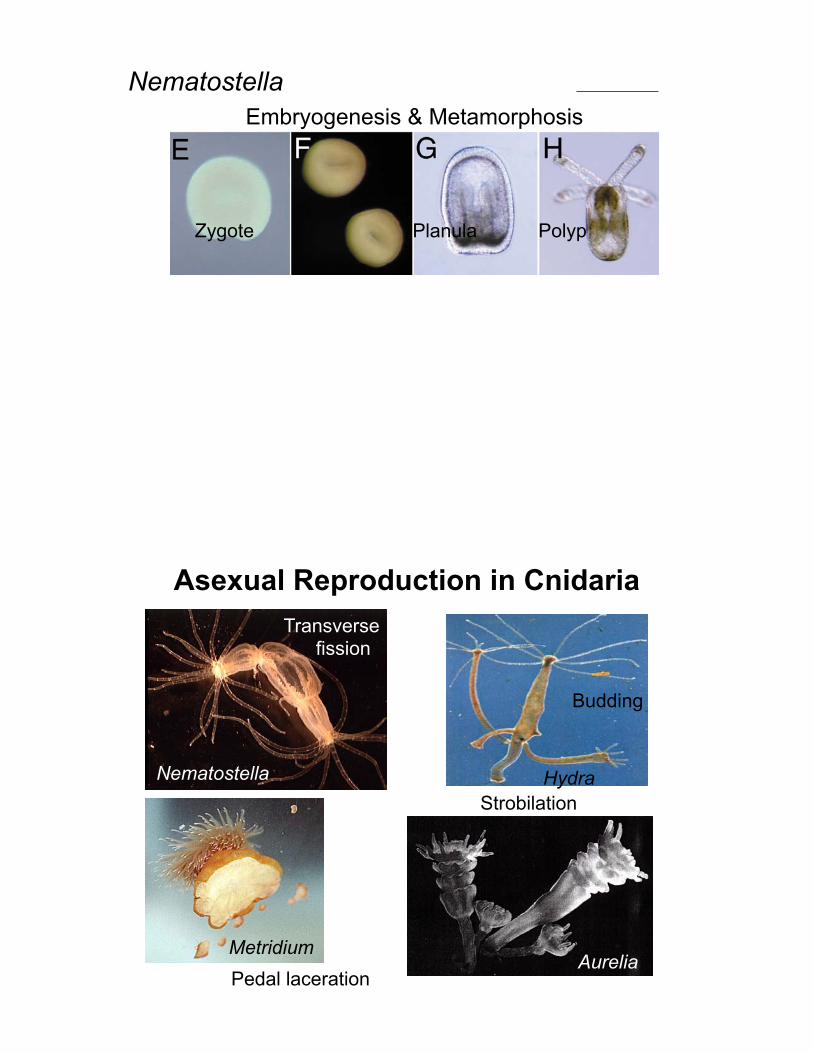
Zygote Planula Polyp
NematostellaEmbryogenesis & Metamorphosis
Nematostella
Asexual Reproduction in Cnidaria
Hydra
MetridiumAurelia
Strobilation
Budding
Transverse fission
Pedal laceration

“Cnidarians are radially symmetrical animals.”
-Audesirk et al., 2001-Barnes et al., 2001
-Brusca & Brusca, 1990-Campbell et al., 2002-Enger & Ross, 2003
-Lewis et al., 2004-Mader, 2004
Hydra is radially symmetrical
ectoderm
endoderm
mesoglea
colenteron(gut)

BILATERIACBA
oral
aboral
ant. post.
dorsal
ventral
BILATERAL SYMMETRYprimary body axis (A-P)& secondary body axis (D-V)
RADIAL SYMMETRYprimary body axis (oral-aboral)
Head
Foot
Column
oral
aboral
Nematostella — the starlet sea anemone
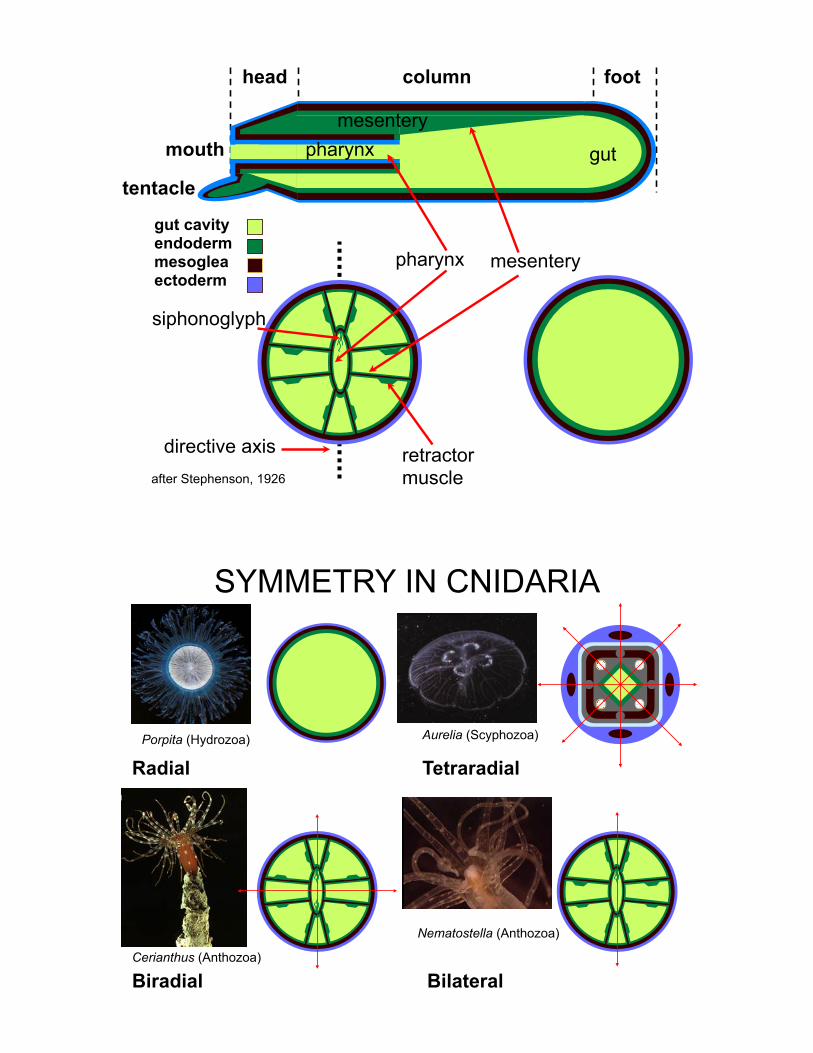
footcolumnhead
tentacle
mouthmesentery
gutpharynx
retractormuscle
pharynx
*
after Stephenson, 1926
directive axis
siphonoglyph
mesentery
gut cavityendodermmesogleaectoderm
SYMMETRY IN CNIDARIA
Radial TetraradialPorpita (Hydrozoa) Aurelia (Scyphozoa)
BiradialCerianthus (Anthozoa)
Bilateral
Nematostella (Anthozoa)

Why did bilateral symmetry originate?
What was its original selective advantage?
The Standard Explanation:Directed Locomotion

An alternate scenario….In the Cnidaria, locomotion is not correlated with symmetry.
Modern Cnidaria are either sessile, or they locomote in a manner that is random with respect to their secondary axis.
The bilaterally symmetrical corals and anemones are essentially sessile.
The ancestral Cnidarian was a sessile polypoid animal.(Bridge et al., 1992, 1995, Collins 2003, and others)
Therefore, bilateral symmetry did not evolve under selection for directed locomotion in the Cnidaria.
In the Cnidaria, symmetry IS correlated with internal ciliary circulation…...
pharynxsiphonoglyph
mesentery
coelenteron
ciliary filaments on asulcal septum
Modified from Kaestner, 1984
Alcyonaria polyp
Why is a sessile organism bilateral?

An alternate scenario….The correlation of symmetry with internal circulation holds for bilaterally symmetrical forms, bi-radially symmetrical forms, and tetradially symmetrical forms.
Among polyps, true radiality characterizes the smallest hydrozoan polyps.
Size interacts with symmetry and the location of ciliary tracts to affect the efficiency of internal circulation.
Size dependency of internal polyp anatomy
Anthozoa
Scyphozoa
Hydrozoa
biradial or bilateral
tetraradial
radial biradial

Can this selective explanation be extrapolated back to the Cnidarian-Bilaterian Ancestor?
If we assume:
Homology of bilateral symmetry in Bilateria and Cnidaria (Finnerty et al., 2004)
That the Cnidarian-Bilaterian ancestor was a sessile benthic animal (e.g., Collins, 2004).
Hox genes, dpp
Ancestral Bilaterian Benthic Crawling Bilateral symmetry
(manifest internally & externally)
Cnidarian-Bilaterian Ancestor
An alternate scenario….
gut
Bilateral symmetry (developmentally plastic?)
gutgut
Ancestral Cnidarian Benthic Sessile Bilateral symmetry
(manifest primarily internally)
Finnerty, BioEssays 2005
! Pronounced external manifestations of bilateral symmetry! Centralized nervous system! Directed locomotion.

Predictions and Implications…The location of ciliary tracts was under the control of “dorsal-ventral patterning genes” in the Cnidarian-Bilaterian ancestor. Perhaps this aspect of developmental gene regulation is conserved among modern Cnidaria and Bilateria.
Variation in the arrangement of ciliary tracts within Cnidaria may be attributable to variation in the expression of dpp and other genes that pattern the “directive” axis.