lectin histochemistry of the esophagus in several mammalian species
TRANSCRIPT

Abstract The mucosa of the esophagus consists of strat-ified squamous epithelium that has a considerable resis-tance to injury. Intercellular glycoconjugates appear toconstitute a major permeability barrier in the superficialportion of the esophageal mucosa. In the present study,we used a panel of lectins to investigate the differencesin glycoconjugate production among different mammali-an species. A battery of 12 lectins was used to studybinding in sections from the esophagus of 6 mammalianspecies, including man. In general, the strongest stainingwas obtained in the stratum superficiale and the weakeststaining in the stratum germinativum. In rabbit esopha-gus, exposure to pepsin/HCl produced a superficial dam-age to the epithelium, a considerable decrease in electri-cal resistance and a decreased staining of the esophagealepithelium with selected lectins. Pretreatment of theesophageal mucosa with sucrose octasulfate, a com-pound with protective properties, prevented, to some ex-tent, the decrease in resistance and lectin staining.
Key words Lectin · Esophagus · Histochemistry ·Sucrose octasulfate · Mammals
Introduction
The esophagus has normally a mechanically robust andtight mucosa that endures the feeding-induced mechani-cal, chemical and thermal stresses that are imposed uponit. This endurance is provided by a stratified squamousepithelium that in some species, such as the rat, is kera-tinized. However, in contrast to several other epithelia,extracellular glycoconjugates may contribute to the mu-cosal integrity by providing a barrier that restricts accessto epithelial cells and subepithelial spaces (Orlando et al.1992). In several species, including man, pig, ferret and
dog, but not rabbit or cat, intra- and/or submucosalglands are also present. Those glands produce mucus andbicarbonate (Hamilton and Orlando 1989), which mayfurther increase the resistance of the mucosa by provid-ing cleansing and neutralizing functions.
In the present study we wished to characterize thestructure and the location of the glycoconjugates in thenormal esophageal mucosa from several mammalianspecies, including man. For this purpose, we utilized lec-tin histochemical methods to survey the distribution ofsix major classes of carbohydrate moieties: fucose, ga-lactose, galactosamine, glucosamine, mannose and sialicacid. We also wanted to get some insight into the func-tion of these carbohydrates and therefore we investigatedacute damage in the rabbit esophagus, a model that hasbeen extensively studied by Orlando and coworkers (Or-lando et al. 1984, 1987, 1992). By exposure of the epi-thelium to pepsin and hydrochloric acid in acute experi-ments we mimicked, to some extent, the stress caused byreflux of gastric contents. By using sucrose octasulfate(SOS), to which protective properties have been attribut-ed (Schweitzer et al. 1985), we then tried to preventchanges that might be induced by exposure of the epithe-lium to pepsin and hydrochloric acid.
Materials and methods
The following species were studied: human, rabbit, pig, cat, dogand ferret. The human biopsies were obtained by endoscopy fromtwo patients at Sahlgren’s University Hospital, Gothenburg, Swe-den. The patients were being diagnosed for unrelated disorders ofthe digestive tract. For all sampling and studies, appropriate per-mission was granted by the local Ethical Committee. The princi-ples of laboratory animal care (NIH, 1985) were followed.
Specimens from the distal esophagus were obtained from threeadult New Zealand White rabbits, three cats, three ferrets, threebeagle dogs (all species sacrificed by an injection of 1 ml/kg sodi-um pentobarbital) and three pigs from a slaughterhouse. The spec-imens were immediately placed in 4% buffered formaldehyde forup to 48 h. Following dehydration in ethanol and clearing in xy-lene, the tissues were embedded in paraffin. Sections, 4 µm thick,were deparaffinized and rehydrated in water. In order to block en-dogenous peroxidase, the sections were exposed to 0.3% H2O2 for
N. Poorkhalkali () · I. Jacobson · H.F. HelanderPreclinical Research and Development, Astra Hässle AB,S-43183 Mölndal, SwedenTel. +46-31776602, Fax: +46-317763827
Anat Embryol (1999) 200:541–549 © Springer-Verlag 1999
O R I G I N A L A RT I C L E
Nadereh Poorkhalkali · Ingemar JacobsonHerbert F. Helander
Lectin histochemistry of the esophagus in several mammalian species
Accepted 12. April 1999

30 min in methanol. After rinsing in distilled water, the sectionswere treated with 0.1% trypsin and 1% CaCl2 in 0.05M TRIS buf-fered saline (TBS)for 30 min at room temperature, followed byrinsing in TBS. Sections were then incubated with lectin (5–20 µg/ml in TBS) for 30–90 min at room temperature. Control sec-tions were incubated with the lectin in the presence of a 0.2–0.5 Msolution of the appropriate blocking monosaccharide (able 1),which resulted in the abolition or pronounced reduction of thestaining. Staining with biotin-labeled lectins was visualized withthe ABC-peroxidase technique, utilizing diaminobenzidine (VectorLaboratories, Burlingame, Calif., USA) as a chromogen. Mayer’shematoxylin staining was carried out before dehydration andmounting. Fluorescein- and rhodamine-labeled lectins were dem-onstrated by the direct labeling method.
All lectins were obtained from Vector Laboratories, exceptMaackia amurensis lectin, which was purchased from EY Labora-tories, San Mateo, Calif., USA. Table 1 shows the different lectinsused in the present study, their specificities and the inhibitors usedto prevent lectin binding in control sections.
In vivo experiments
For the exposure to pepsin/HCl and sucrose octasulfate, part ofthe distal esophagus was isolated and perfused in situ through sil-icone tubes. The transmucosal electrical resistance was used as anindex of epithelial integrity (Kidder et al. 1983). New ZealandWhite rabbits (2.1–3 kg body weight) were anesthetized (for in-duction: Hypnorm Vet (Janssen Cilag, Saunderton, UK) 0.3 ml/kgin combination with Dormicum (Hoffmann-La Roche, Basel,Switzerland) 0.4 ml/kg; for maintenance: a continuous infusion ofHypnorm Vet diluted 1:10 1–3 ml/h.kg) and tracheotomized. Theabdomen was opened and a silicon drain tube (5 mm o.d.) was in-serted through the stomach into the lower region of the esopha-gus. The tube was positioned 2–3 cm above the gastro-esophagealjunction. A second silicon tube (3 mm o.d.) was then passedthrough the mouth of the animal into the esophagus and posi-tioned above the drain catheter. The segment of the esophagus be-tween the catheters measured 2–3 cm and was perfused at a rateof 1 ml/min by the use of a peristaltic pump connected to the oralcatheter. Catheters in the major ear veins were used for infusionof anesthetics and for connection to an electrical reference elec-trode.
By connecting the silicon tubes via salt bridges to amplifiers(IsoDam, WPI Instruments, USA) it became possible to deliversquare wave current injections between 5 and 10 µA in strength,10 s in duration, and to monitor the corresponding voltage deflec-tions. This enabled the estimation of the transmural electrical re-sistance (Kidder et al. 1983). Estimates of transmucosal electricalresistances are given as the percentage of the value obtained im-mediately after changing to pepsin/HCl solution. Control values(input resistances) averaged 1.7 kΩ.
The rabbits were divided into four groups with three animals ineach. The rabbits in groups 1–3 were perfused through the esopha-gus with 140 mM saline, 1 ml/min, for 20 min. In group 1 therewas an additional perfusion for 20 min with saline and then expo-sure to pepsin/HCl, (10 mM HCl, 130 mM NaCl, pH 2, pepsin0.9 mg/ml (pepsin A, Sigma, St Louis, Mo., USA)) for 60 min.Group 2 animals were perfused with 10 mM SOS (Katsura Chemi-cals, Tokyo, Japan) in 140 mM saline for 20 min and SOS in pep-sin/Hcl for 60 min. Control animals in group 3 were perfused withsaline for 80 min and group 4 animals were intact animals withoutany treatment.
Results
The normal eosphageal mucosa showed wide-spread lec-tin binding in all species investigated (Figs. 1–3, Table2). In man and dog, 10 of the 12, rabbit 9 of 12, in pigand cat 6 of 12, and in ferret 11 of 12 lectins used result-ed in staining. UEA I, UEA II, Jacalin and PSA stainedsome or all layers of the epithelium of all species thatwere examined. While the similarities in the location ofthe lectin staining between the species thus were marked,there were frequently differences in the intensity of thestaining. In general, the most superficial layer of theesophageal mucosa, stratum superficiale, stained withmore lectin species than did the deeper layers. The stain-ing was usually seen along the cell borders. MAA – spe-cific for sialic acid – produced no staining in any of themucosae.
Certain lectins produced intense cytoplasmic stainingof the cells of the esophageal glands (Fig. 3b, d, f, g);however, glands were observed only in the specimensfrom the pigs, ferrets and dogs.
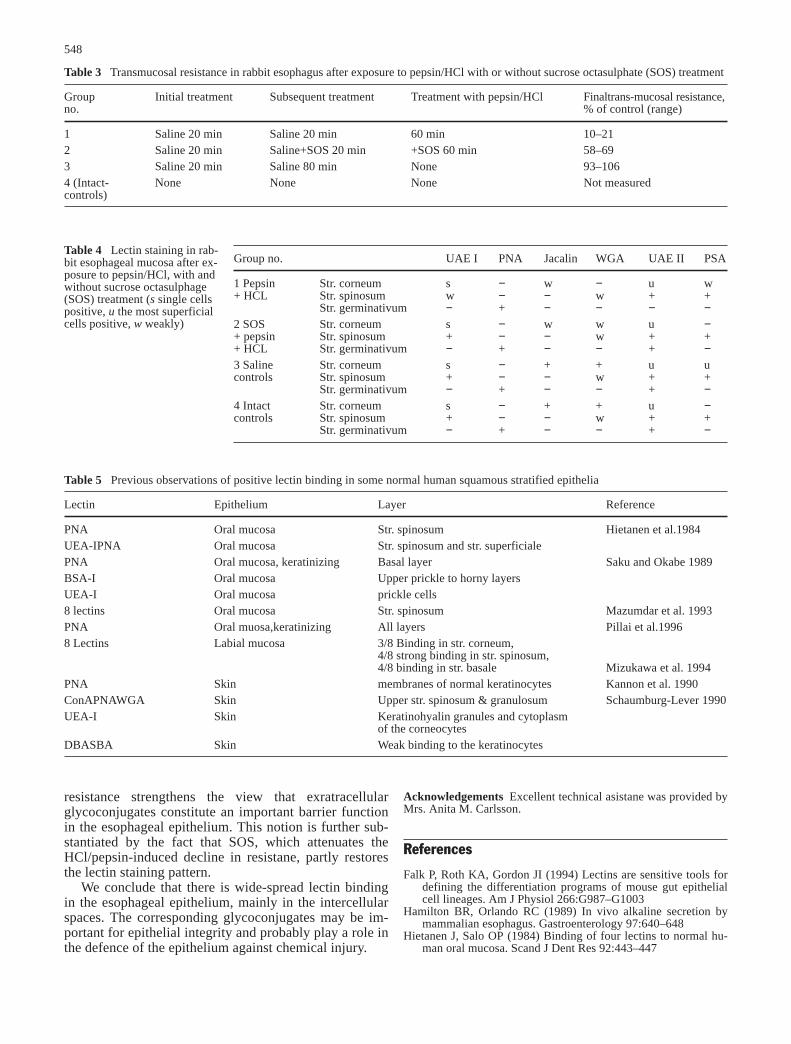
The transmucosal electrical resistance is an index ofthe epithelial permeability (Kidder et al. 1983) and expo-sure to pepsin/HCl reduced the electrical transmucosalresistance of 10–21% of the baseline value. In contrast,addition of the protective agent SOS to the pepsin/HClinfusion attenuated the acid/pepsin-induced decline inresistance, which then levelled off at 58–69% of thebaseline value (Table 3). Exposure of the esophageal epi-thelium to pepsin/HCl reduced UEA I staining of thestratum spinosum and UEA II staining in the stratum
542
Table 1 Carbohydrates, their corresponding lectins and their specific inhibitors
Carbohydrate Lectin Lectin abbreviation Inhibitor
Fucose Ulex europaeus agglutinin UEA I 50 mM L-fucoseGalactose Peanut agglutinin PNA 200 mM Galactose
Jacalin JacGalNAc Sophora japonica agglutinin SJA 200 mM N-acetyl galactosamine
Dolichos biflourus agglutinin DBAGlcNAc Solanum tuberosum agglutinin STA 500 mM N-acetyl-glucosamine
Wheat germ agglutinin WGAUlexeuropaeus agglutinin UEA II
Mannose Pisum sativum agglutinin PSA 200 mM α-Methyl mannosideConcanavalinA Con A
Sialic acid Maackia amuresis MAA 75 mM Sialyllactose α2,3Complex (unspecified) Euonymus europaeus EEA 500 mM Lactose
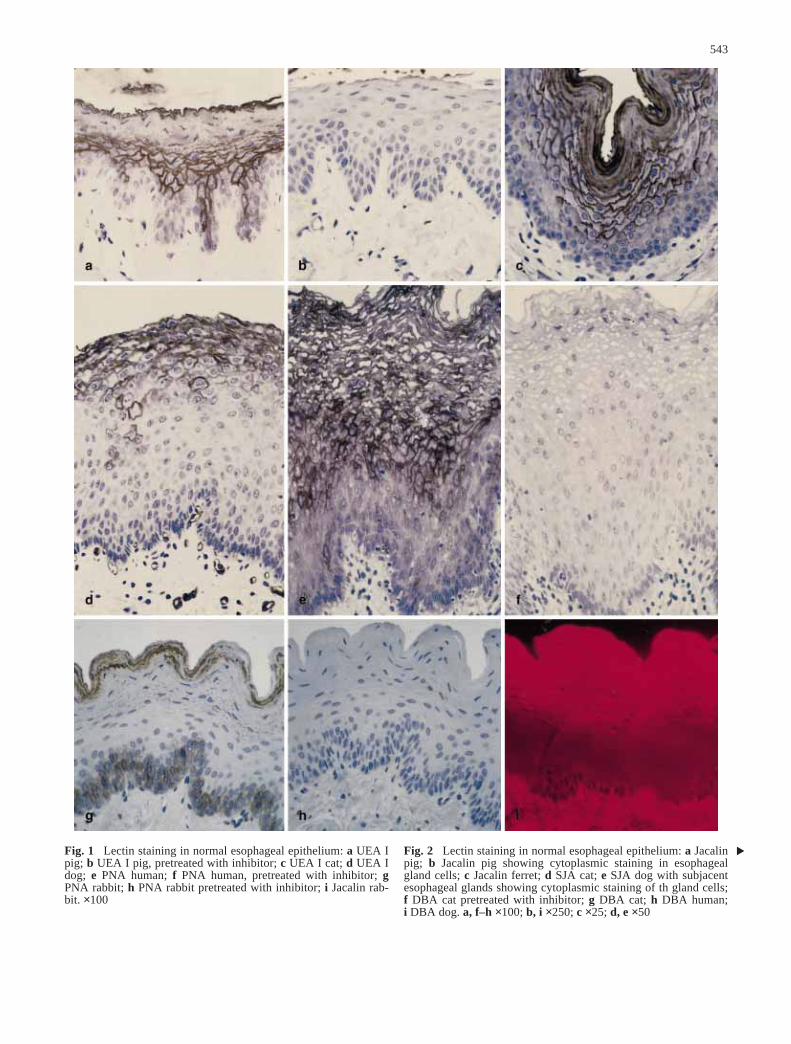
543
Fig. 1 Lectin staining in normal esophageal epithelium: a UEA Ipig; b UEA I pig, pretreated with inhibitor; c UEA I cat; d UEA Idog; e PNA human; f PNA human, pretreated with inhibitor; gPNA rabbit; h PNA rabbit pretreated with inhibitor; i Jacalin rab-bit. ×100
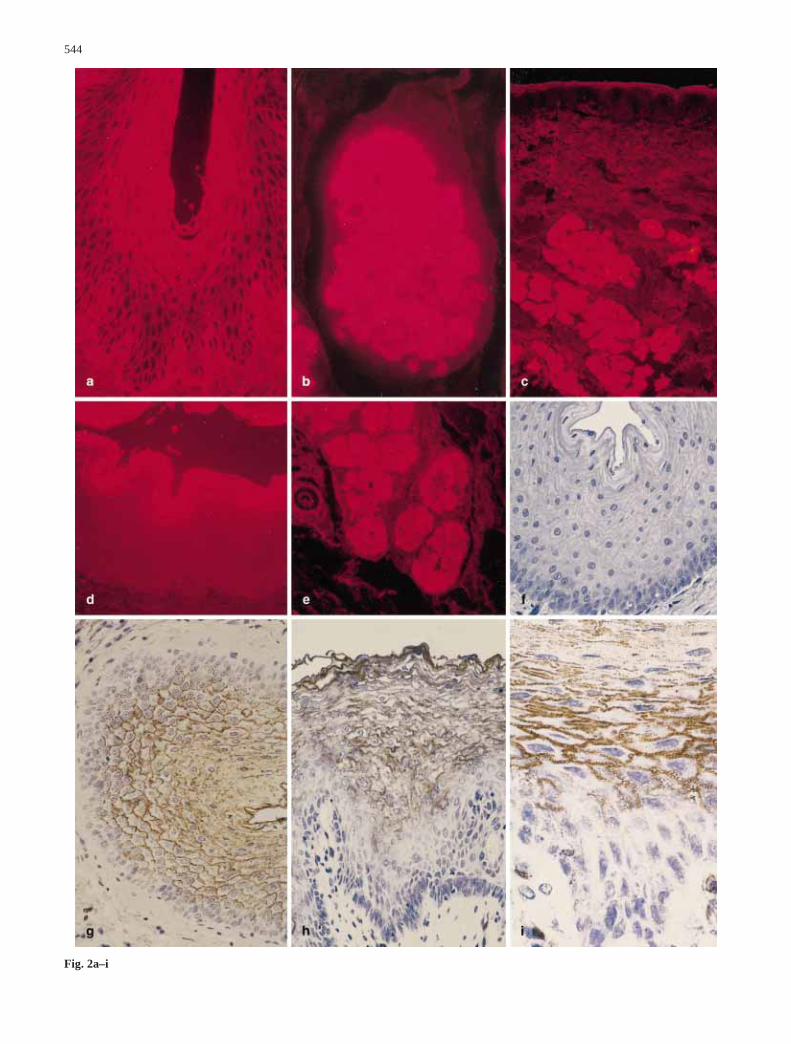
Fig. 2 Lectin staining in normal esophageal epithelium: a Jacalinpig; b Jacalin pig showing cytoplasmic staining in esophagealgland cells; c Jacalin ferret; d SJA cat; e SJA dog with subjacentesophageal glands showing cytoplasmic staining of th gland cells;f DBA cat pretreated with inhibitor; g DBA cat; h DBA human;i DBA dog. a, f–h ×100; b, i ×250; c ×25; d, e ×50
544
Fig. 2a–i
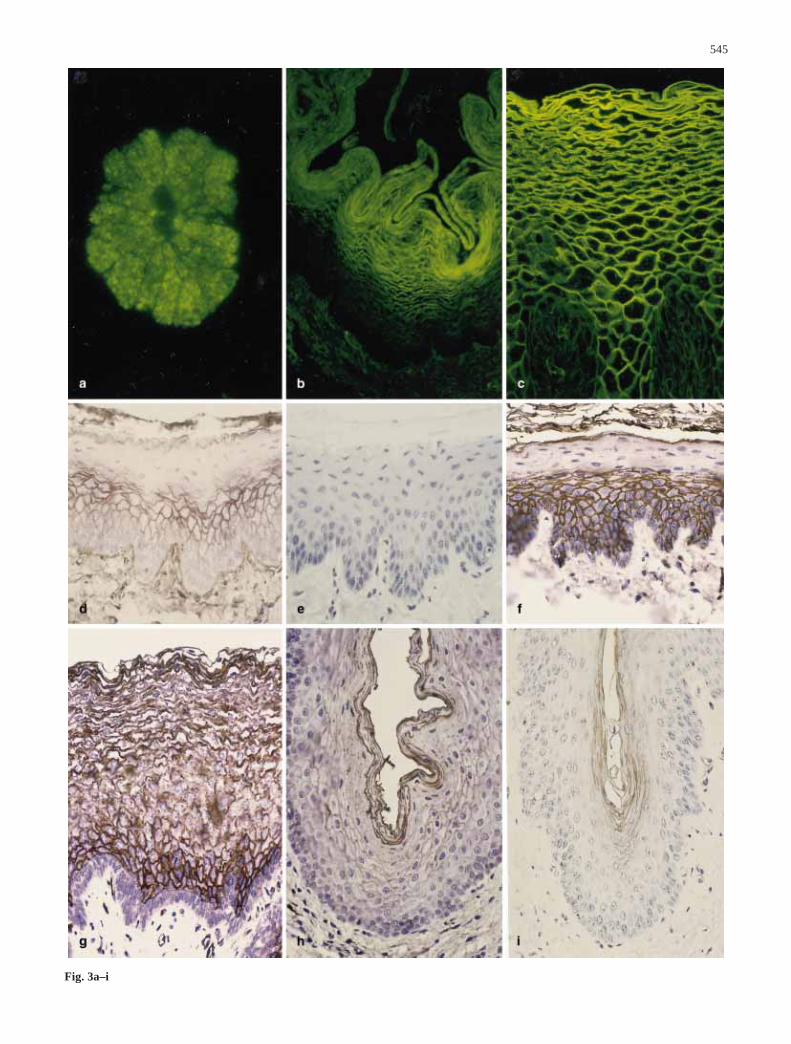
545
Fig. 3a–i
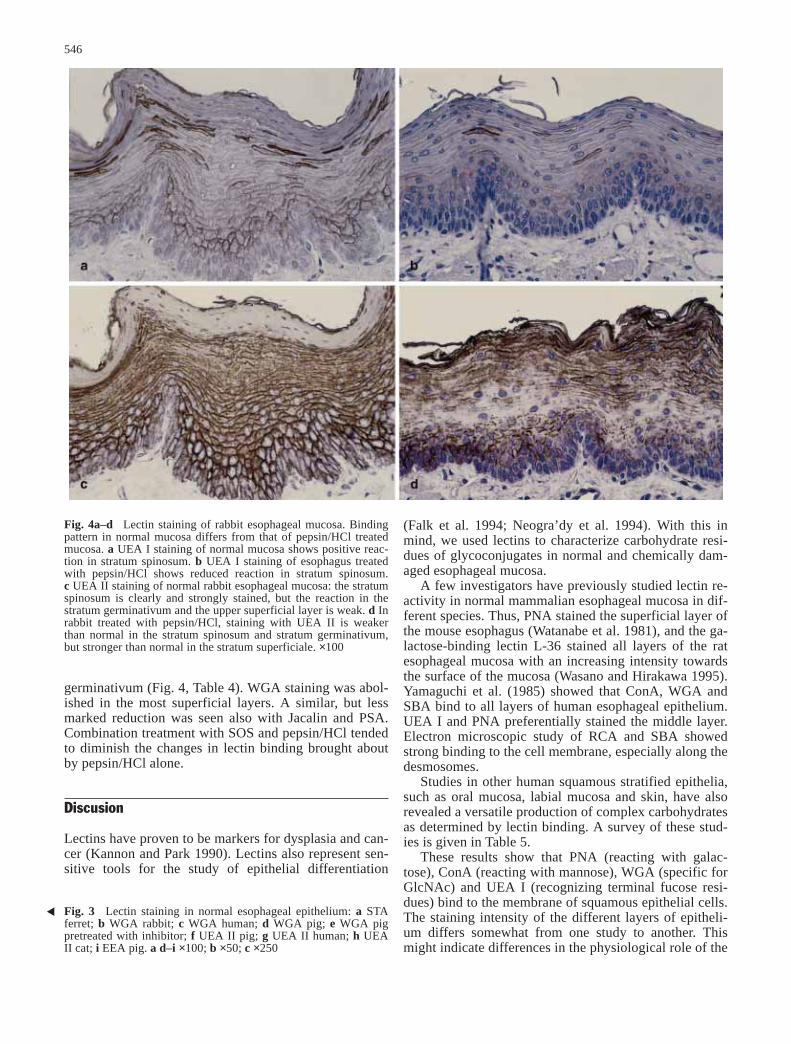
germinativum (Fig. 4, Table 4). WGA staining was abol-ished in the most superficial layers. A similar, but lessmarked reduction was seen also with Jacalin and PSA.Combination treatment with SOS and pepsin/HCl tendedto diminish the changes in lectin binding brought aboutby pepsin/HCl alone.
Discusion
Lectins have proven to be markers for dysplasia and can-cer (Kannon and Park 1990). Lectins also represent sen-sitive tools for the study of epithelial differentiation
(Falk et al. 1994; Neogra’dy et al. 1994). With this inmind, we used lectins to characterize carbohydrate resi-dues of glycoconjugates in normal and chemically dam-aged esophageal mucosa.
A few investigators have previously studied lectin re-activity in normal mammalian esophageal mucosa in dif-ferent species. Thus, PNA stained the superficial layer ofthe mouse esophagus (Watanabe et al. 1981), and the ga-lactose-binding lectin L-36 stained all layers of the ratesophageal mucosa with an increasing intensity towardsthe surface of the mucosa (Wasano and Hirakawa 1995).Yamaguchi et al. (1985) showed that ConA, WGA andSBA bind to all layers of human esophageal epithelium.UEA I and PNA preferentially stained the middle layer.Electron microscopic study of RCA and SBA showedstrong binding to the cell membrane, especially along thedesmosomes.
Studies in other human squamous stratified epithelia,such as oral mucosa, labial mucosa and skin, have alsorevealed a versatile production of complex carbohydratesas determined by lectin binding. A survey of these stud-ies is given in Table 5.
These results show that PNA (reacting with galac-tose), ConA (reacting with mannose), WGA (specific forGlcNAc) and UEA I (recognizing terminal fucose resi-dues) bind to the membrane of squamous epithelial cells.The staining intensity of the different layers of epitheli-um differs somewhat from one study to another. Thismight indicate differences in the physiological role of the
546
Fig. 3 Lectin staining in normal esophageal epithelium: a STAferret; b WGA rabbit; c WGA human; d WGA pig; e WGA pigpretreated with inhibitor; f UEA II pig; g UEA II human; h UEAII cat; i EEA pig. a d–i ×100; b ×50; c ×250
Fig. 4a–d Lectin staining of rabbit esophageal mucosa. Bindingpattern in normal mucosa differs from that of pepsin/HCl treatedmucosa. a UEA I staining of normal mucosa shows positive reac-tion in stratum spinosum. b UEA I staining of esophagus treatedwith pepsin/HCl shows reduced reaction in stratum spinosum.c UEA II staining of normal rabbit esophageal mucosa: the stratumspinosum is clearly and strongly stained, but the reaction in thestratum germinativum and the upper superficial layer is weak. d Inrabbit treated with pepsin/HCl, staining with UEA II is weakerthan normal in the stratum spinosum and stratum germinativum,but stronger than normal in the stratum superficiale. ×100

glycoconjugates, but may also depend on differences inthe methods used for staining.
In the present investigation, MAA, with a high speci-ficity for α 2,3-linked neuraminic acids, did not stain theesophageal mucosa or the submucosal glands in any ofthe species that we investigated. These results are in con-trast to the strong MAA staining of the mouse intestinalmucosa demonstrated by Falk et al. (1994). Still, in theesophagus, alcian blue at pH 2.5 – a general marker forsialic acids – provided a strong staining of the submu-cosal glands and in some species, it also stained the stra-tum superficiale. We conclude that the sialic acids of theesophagus have a low concentration of the α 2,3-linkedneuraminic acids that characteristically bind MAA.
Several authors have utilized lectin histochemistry asa tool for studies of gastrointestinal epithelial differentia-tion (Falk et al. 1994; Neogra’dy et al. 1994). In thepresent study, the staining intensity for most lectins waslow in the young cells of the stratum germinativum, butincreased as the cells become older and transferred to thesuperficial layers.
The esophagus is exposed not only to food and liq-uids, but also to gastric contents, including acid and pep-sin (and possibly intestinal juices), which infrequentlyregurgitate into the esophagus, even in healthy subjects.In gastro-intestinal epithelia, the first line of defense is alayer of mucus that is more or less firmly attached to thesurface cells and reaches a thickness of up to a few hun-dred micrometers. This layer is almost totally lacking inthe esophagus.
The next line of defense is therefore the epithelium it-self. Previous studies have shown that the main pathwayfor movement of protons across the epithelium is via thetranscellular route and that the high resistance of thispathway is broken down by excessive exposure to acid(Orlando et al. 1984). In the study by Orlando et al.(1992), the intercellular glycoconjugates, vizualized byPAS and alcian blue staining in the rabbit esophagealmucosa, were observed in the stratum superficiale and –to a much lesser extent – in the stratum spinosum. Ourresults show lectin binding in the intercellular spaces ofall layers of the esophageal mucosa, although for mostlectins the intensity of staining was highest in the stra-tum superficiale. This suggests that the lectins are moresensitive tools for studying the glycoconjugates than thetraditional histochemical methods.
Our finding that exposure to pepsin/HCl reduces thetransmural electrical resistance is in agreement with ear-lier reports showing that treatment with pepsin/HCl leadsto an increased epithelial permeability and epithelialdamage (Kidder et al. 1983; Schweitzer et al. 1985). Ourresults also corroborate previous findings that acute pep-sin/HCl-induced damage of the esophageal epithelium isattenuated by SOS (Schweitzer et al. 1985). The protec-tive properties of the drug has been suggested to dependon binding with the pepsin-substrate (Schweitzer et al.1985).
The moderate los of lectin staining in the outer celllayers in parallel with a drastic reduction in the electrical
547
Tab
le 2
R
esul
ts o
f le
ctin
sta
inin
g in
nor
mal
eso
phag
eal m
ucos
a (p
patc
hy p
osit
ive,
ssi
ngle
cel
ls p
osit
ive,
uth
e m
ost s
uper
ific
ial c
ells
pos
itiv
e, w
wea
kly
posi
tive
)
Hum
anR
abbi
tPi
gC
atFe
rret
Dog
Str.
-St
r.St
r. ge
r-St
r. St
r.St
r. ge
r-St
r. St
r.St
r. ge
r-E
soph
a-St
r. -
Str.
Str.
ger-
Str.
Str.
Str.
ger-
Eso
pha-
Str.
Str.
Str.
ger-
Eso
pha-
supe
r-sp
ino-
min
a-su
per-
spin
o-m
ina-
supe
r-sp
ino-
min
a-ge
al
supe
r-sp
ino-
min
a-su
per-
spin
o-m
ina-
geal
supe
r-sp
ino-
min
a-ge
alfi
cial
esu
mtiv
umfi
cial
esu
mtiv
umfi
cial
esu
mtiv
umgl
ands
fici
ale
sum
tivum
fici
ale
sum
tivum
glan
dsfi
cial
esu
mtiv
umgl
ands
UE
A I
++
−s
+−
u+
−+
++
−u
−−
++
−−
+PN
A+
+−
−−
+−
−−
−−
−−
−+
+−
−−
−−
Jaca
lin+
++
+−
−u
+w
++
−−
+w
−+
−+
−−
SJA
+−
−w
p−
−−
−−
−+
−−
w−
−+
w−
−+
DB
A+
−−
++
−−
−−
−+
+−
wp
wp
−−
++
−+
STA
ww
−−
−−
−−
−−
−−
−+
−−
++
−−
+W
GA
++
++
w−
−+
+−
−−
−−
−+
−+
+w
−U
EA
II+
+−
u+
+w
++
ww
+−
w+
−+
+−
+PS
A+
+−
+w
−u
+w
−+
ws
+w
−−
w+
−−
Con
A+
−−
−−
−−
−−
−−
−−
w−
−−
ww
w−
MA
A−
−−
−−
−−
−−
−−
−−
−−
−−
−−
−−
EE
A−
−−
wp
−−
+w
−+
−−
−p
−−
+uw
−−
w
resistance strengthens the view that exratracellularglycoconjugates constitute an important barrier functionin the esophageal epithelium. This notion is further sub-stantiated by the fact that SOS, which attenuates theHCl/pepsin-induced decline in resistane, partly restoresthe lectin staining pattern.
We conclude that there is wide-spread lectin bindingin the esophageal epithelium, mainly in the intercellularspaces. The corresponding glycoconjugates may be im-portant for epithelial integrity and probably play a role inthe defence of the epithelium against chemical injury.
Acknowledgements Excellent technical asistane was provided byMrs. Anita M. Carlsson.
References
Falk P, Roth KA, Gordon JI (1994) Lectins are sensitive tools fordefining the differentiation programs of mouse gut epithelialcell lineages. Am J Physiol 266:G987–G1003
Hamilton BR, Orlando RC (1989) In vivo alkaline secretion bymammalian esophagus. Gastroenterology 97:640–648
Hietanen J, Salo OP (1984) Binding of four lectins to normal hu-man oral mucosa. Scand J Dent Res 92:443–447
548
Table 3 Transmucosal resistance in rabbit esophagus after exposure to pepsin/HCl with or without sucrose octasulphate (SOS) treatment
Group Initial treatment Subsequent treatment Treatment with pepsin/HCl Finaltrans-mucosal resistance,no. % of control (range)
1 Saline 20 min Saline 20 min 60 min 10–212 Saline 20 min Saline+SOS 20 min +SOS 60 min 58–693 Saline 20 min Saline 80 min None 93–1064 (Intact- None None None Not measuredcontrols)
Table 4 Lectin staining in rab-bit esophageal mucosa after ex-posure to pepsin/HCl, with andwithout sucrose octasulphage(SOS) treatment (s single cellspositive, u the most superficialcells positive, w weakly)
Group no. UAE I PNA Jacalin WGA UAE II PSA
1 Pepsin Str. corneum s − w − u w+ HCL Str. spinosum w − − w + +
Str. germinativum − + − − − −2 SOS Str. corneum s − w w u −+ pepsin Str. spinosum + − − w + ++ HCL Str. germinativum − + − − + −3 Saline Str. corneum s − + + u ucontrols Str. spinosum + − − w + +
Str. germinativum − + − − + −4 Intact Str. corneum s − + + u −controls Str. spinosum + − − w + +
Str. germinativum − + − − + −
Table 5 Previous observations of positive lectin binding in some normal human squamous stratified epithelia
Lectin Epithelium Layer Reference
PNA Oral mucosa Str. spinosum Hietanen et al.1984UEA-IPNA Oral mucosa Str. spinosum and str. superficialePNA Oral mucosa, keratinizing Basal layer Saku and Okabe 1989BSA-I Oral mucosa Upper prickle to horny layersUEA-I Oral mucosa prickle cells8 lectins Oral mucosa Str. spinosum Mazumdar et al. 1993PNA Oral muosa,keratinizing All layers Pillai et al.19968 Lectins Labial mucosa 3/8 Binding in str. corneum,
4/8 strong binding in str. spinosum,4/8 binding in str. basale Mizukawa et al. 1994
PNA Skin membranes of normal keratinocytes Kannon et al. 1990ConAPNAWGA Skin Upper str. spinosum & granulosum Schaumburg-Lever 1990UEA-I Skin Keratinohyalin granules and cytoplasm
of the corneocytesDBASBA Skin Weak binding to the keratinocytes

Kannon G, Park HK (1990) Utility of peanut agglutinin (PNA) inthe diagnosis of squamous cell carcinoma and keratoacantoma.Am J Dermatopathol 12:31–36
Kidder GW III, Lillemoe KD, Harmon JW, Maydonowitch CL,Bunte RM, Johnson L (1983) Evaluation of in vivo measure-ment of transesophageal electrical resistance as an indicator ofearly experimental esophageal mucosal injury. J Lab Clin Med102:477–486
Mazumdar S, SenGupta SK, Param R, Sinha SN (1993) Bindingpattern of eight different lectins in healthy subjects and pa-tients with dysplastic and malignant lesions of the oral cavity.Internat J Oral Maxillofac Surg 22:301–305
Mizukawa Y, Takata K, Ookusa Y, Nagashima M, Hirano H(1994) Lectin binding pattern in normal human labial mucosa.Histochem J 26:863–869
Neogra’dy S, Ga’lffi P, Veresgyha’zy T, Bardo’cz, Pusztai A(1994) Lectins as markers of rumen epithelia cell differenciat-ion. Histochem J 26:197–206
Orlando RC, Bryson JC, Powell DW (1984) Mechanisms of H+ injuryin rabbit esophageal epithelium. Am J Physiol 246:G718–G724
Orlando RC, Turjman NA, Tobey NA, Schreiner VJ, Powell DW(1987) Mucosal preotection by sucralfate and its components inacid-exposed rabbit esophagus. Gastroenterology 93:352–361
Orlando RC, Lacy ER, Tobey NA, Cowart K (1992) Barriers toparacellular permeability in rabbit esphageal epithelium. Gas-troenterology 102:910–923
Pillai KR, Remani P, Kannan S, Sujathan K, Mathew B, Vijayaku-mar T, Nair MK, Menon VP (1996) Lectin histochemistry oforal premalignant and malignant lesions: correlation of JFLand PNA binding pattern with tumour progression. Eur J Can-cer B Oral Oncol 32:32–37
Saku T, Okabe H (1989) Differential lectin-bindings in normal andprecancerous epithelium and squamous cell carcinoma of theoral mucosa. J Oral Pathol Med 18:438–445
Schaumburg-Lever G (1990) Ultrastructural localization of lectin-binding sites in normal skin. J Invest Dermatol 94:465–470
Schweitzer EJ, Bass BL, Johnson LF, Harmon JW (1985) Sucrai-fate prevents experiments peptic esopagitis in rabbits. Gastro-enterology 88:611–619
Wasano K, Hirakawa Y (1995) Rat intestinal galactoside-bindinglectin L-36 functions as a structural protein in the superficialsquamous cells of the esophageal epithelium. Cell Tissue Res281:77–83
Watanabe M, Muramatsu T, Shirane H, Ugai K (1981) Discretedistribution of binding sites for Dolichos biflorus agglutinin(DBA) and for peanut agglutinin (PNA) in mouse organ tis-sues. J Histochem Cytochem 29:779–780
Yamaguchi Y, Ito M, Saito S, Aoyagi T, Takata K, Hirano H(1985) Light and electron microscopic study of lectin bindingsites in human esophageal epithelium (Japanese). KaibogakuZasshi 60:104–108
549