jenni korhonen antibiotic resistance of lactic acid bacteria
TRANSCRIPT

Publications of the University of Eastern FinlandDissertations in Forestry and Natural Sciences
Publications of the University of Eastern Finland
Dissertations in Forestry and Natural Sciences
isbn 978-952-61-0096-8
Jenni Korhonen
Antibiotic Resistance ofLactic Acid Bacteria
Resistance to antibiotics is a common
characteristic in the world of bacteria.
Hitherto, only little attention has
been paid to the antimicrobial
susceptibilities of beneficial lactic acid
bacteria, on the opposite to several
pathogenic bacterial species. However,
there is some concern that antibiotic
resistance in lactic acid bacteria could
be transferred to pathogenic bacterial
species, complicating the treatment of
a diesease or infection and lead to the
spread of antibiotic-resistant bacteria.
Studies were conducted to isolate and
identify lactic acid bacteria of animal
and human origins, to evaluate their
antimicrobial resistance patterns
and to propose minimum inhibitory
concentrations for Lactobacillus species.
dissertatio
ns | 0
07 | Jenn
i Ko
rh
on
en | A
ntib
iotic R
esistan
ce of La
ctic Acid
Ba
cteria
Jenni Korhonen
Antibiotic Resistance ofLactic Acid Bacteria

JENNI KORHONEN
Ant ibiot ic Resistance of Lact ic Acid Bacteria
Publications of the University of Eastern Finland Dissertations in Forestry and Natural Sciences
7
To be presented by permission of the Facu lty of Forestry and Natural Sciences
Department of Biosciences
University of Eastern Finland , Kuopio, on Saturday 24th April 2010, at 12 noon. University of Eastern Finland for public examination in the Auditorium L22, Snellmania Build ing,
,
University of Eastern Finland Kuopio 2010

Kopijyvä Kuopio, 2010
Editors: Prof. Pertti Pasanen Prof. Tarja Lehto, Prof. Kai Peiponen
Distribution: Eastern Finland University Library / Sales of publications
P.O.Box 107, FI-80101 Joensuu, Finland tel. +358-50-3058396
http:/ / www.uef.fi/ kirjasto
Print: ISBN: 978-952-61-0096-8
ISSNL: 1798-5668 ISSN: 1798-5668
Online:
ISBN: 978-952-61-0097-5 (PDF) ISSN: 1798-5676 (PDF)

Author´s address: University of Eastern Finland
Department of Biosciences P.O.Box 1627 70211 KUOPIO
FINLAND email: [email protected] Supervisors: Professor Atte von Wright, Ph.D. University of Eastern Finland
Department of Biosciences P.O.Box 1627 70211 KUOPIO
FINLAND email: [email protected]
Professor Sinikka Pelkonen, Ph.D. Research Department of Veterinary Bacteriology Finnish Food Safety Authority Evira
Neulaniementie 4 70150 KUOPIO FINLAND email: [email protected]
Reviewers: Professor Wolfgang Kneifel
BOKU-University of Natural Resources and Applied Life Sciences Department of Food Science and Technology
VIENNA AUSTRIA email: [email protected]
Associate Professor Arthur Ouwehand University of Turku
Functional Foods Forum TURKU FINLAND email: [email protected]
Opponent: Professor Sven Lindgren National Food Administration
UPPSALA SWEDEN email: [email protected]


ABSTRACT: Lactic acid bacteria (LAB) are a heterogeneous group of bacteria found widely in nature . They colonize the gastrointestinal and urogenital tracts of humans and animals, and are present in foods such as dairy products, fermented meats, fruits and vegetables. LAB are also intentionally added to several probiotic products because of their potential health benefits. Many LAB species are generally recognized as safe (GRAS), and several LAB species have received a Qualified Presumption of Safety (QPS) status given by European Food Safety Authority (EFSA). Resistance to antimicrobial d rugs (antibiotics) is a common character istic in the world of bacteria. In the interaction between bacteria, genetic material is transferred from one bacterium to another, and also genes coding for resistance to a certain antibiotic may be passed on to other bacterial species. Since LAB are natural and profitable inhabitants in many environments (gastrointestinal tract, several foods), strains with resistance to antibiotics would not be detrimental to the wellbeing of humans or animals. However, there is some concern that antibiotic resistance in LAB could then be transferred to possibly pathogenic bacterial species, complicating the treatment of a d isease or infection and lead to the spread of antibiotic-resistant bacteria. Hitherto, only little attention has been paid to the antimicrobial susceptibilities of LAB (European Commission. 2005, European Commission 2008). In order to illustrate the current situation of antibiotic resistance patterns in beneficial LAB, stud ies were first conducted to isolate and identify lactic acid bacteria of animal and human origins, and secondly to evaluate their antimicrobial resistance patterns. Moreover, tentative cut-off values d ivid ing the populations into susceptible and resistant were also proposed based on minimum inhibitory concentrations (MIC) for a total of fourteen Lactobacillus species. Most of the LAB strains were found to be suscep tible to all antimicrobial agents used in the stud ies. The most frequently found resistance was against tetracycline, followed by resistance against aminoglycosides. The horizontal transferability of antibiotic resistance between LAB (Lactococcus garvieae species) and pathogenic Listeria monocytogenes was also achieved , demonstrating that a gene coding for tetracycline resistance tet(S) can be transferred from a fish pathogen to a human pathogen by conjugation in vitro. Universal Decimal Classification:579.864, 615.015.8, 615.33, 577.18 National Library of Medicine Classification: QW 142.5.A8, QW 45 CAB Thesaurus: lactic acid bacteria; Lactobacillus; drug resistance; antibiotics; tetracycline; aminoglycoside antibiotics; horizontal transmission; Lactococcus garvieae, Listeria monocytogenes TIIVISTELMÄ: Maitohappobakteerien vastustuskykyä antibiootteja kohtaan selvitettiin porsaiden, vasikoiden ja ihmisten suolistosta sekä elintarvikkeista eristetyistä kannoista. Tulosten perusteella ehdotettiin neljälletoista laktobasillilajille raja-arvoja, joita voitaisiin käyttää jaettaessa kantoja herkkiin ja vastustuskykyisiin. Eniten vastustuskykyisyyttä havaittiin tetrasykliinille sekä aminoglykosideille. Lisäksi havaittiin, että antibioottivastustuskyky voi siirtyä lajien välillä laboratorio-olosuhteissa Lactococcus garvieae – bakteerin ja ihmiselle tautia aiheuttavan Listeria monocytogenes –bakteerin välillä. Yleinen suomalainen asiasanasto: maitohappobakteerit; resistenssi -- antibiootit


In nature we never see anything isolated, but everything in connection with something else---
Johann Wolfgang von Goethe


ACKNOWLEDGEMENTS This work was carried out in the Deparment of Forestry and Natural Sciences, University of Eastern Finland (formerly Department of Biosciences, Nutrition and Biotechnology, University of Kuop io). My work was financially supported by European Union 6th Framework Program project “Assessment and Critical Evaluation of Antibiotic Resistance Transferability in Food Chain, ACE-ART”, and also by Finnish Cultural Found ation. In particular, I wish to express my sincere gratitude to my principal supervisor, Professor Atte von Wright, for giving me the opportunity to work comparatively independently and still having his full support when needed . Thank you for sharing your time and thoughts with me during these years. Sincere thanks go also to my other supervisor, Professor Sinikka Pelkonen . Sincere thanks to Paula Hyvönen, for her comments on the thesis, and your gu idance and infinite support during my stud ies and work at the department. I owe my thanks to the official referees, Professor Wolfgang Kneifel and Associate Professor Arthur Ouwehand , for agreeing to review my thesis. Thank you for your constructive criticisms. Thanks also to Owen MacDonald for language consu lting. I wish to thank all the former “ACE-ARTists”, especially all co-authors around Europe and also in the department. Thank you for the pleasant collaboration and your efforts in scientific research. Many of your comments have essentially improved the published papers. Special thanks for thorough review ing to Angela van Hoek, Geert Huys and Sigrid Mayrhofer, and also to Mia Egervärn for nice collaboration. Particular thanks to members of our “subunit”, Nutrition and Food Biotechnology , for the pleasant working atmosphere. Special thanks to Carme Plumed Ferrer, Kristiina Kinnunen and Jouni Heikkinen, for sharing your expertise on lactic acid bacteria. For their technical assistance, I would like to thank Mirja Rekola, Elvi Mäkirinne, Riitta Venäläinen and Eeva-Liisa Palkispää. I would like to thank my mother and father, Marjatta and Sakari, and my mother - and father-in-law, Eeva and Veikko, for their care and supp ort during my stud ies and work. A clean house, d inner read y and child ren happy, after sometimes long hours at work. What more could you ask! Finally, I w ish to thank our child ren, Aino, Lauri and Elsa, for bringing me the joy, and giving me a perspective of life in general. To my husband Petri, a special thank you for those constructive arguments that we both so much enjoy, no mat ter the subject! Thank you for your unconditional love and support during our years together. Kuopio, April 2010 Jenni Korhonen


ABBREVIATIONS
ACE-ART Assessment and critical evaluation of antibiotic resistance
transferability in food chain
ATCC American Type Culture Collection
BLAST Basic local alignment search tool
CAMBH cation ad justed Müller-Hinton agar
CFU Colony forming unit
CLSI Clinical and Laboratorty Stand ards Institute
EFSA European Food Safety Authority
EMBL European Molecu lar Biology Laboratory
FEEDAP Panel on Additives and Products or Substances used in Animal Feed
GIT Gastrointestinal tract
GRAS Generally recognized as safe
LAB Lactic acid bacteria
LAMVAB Lactobacillus Anaerobic MRS with Vancomycin and Bromocresol green
L. Listeria
Lb. Lactobacillus
Lc. Lactococcus
LSM Lactic acid bacteria susceptibility medium
MIC Minimum inhibitory concentration
MRS de Man, Rogosa and Sharpe
MTT Agrifood Research Finland
NCCLS National Committee on Clinical Laboratory Stand ards
OD Optical density
PCR Polymerase chain reaction
QPS Qualified presumption of safety
RAPD Randomly amplified polymorphism DNA
rpm Revolutions per minute
TC transconjugant
TF transformant
W. Weissella


LIST OF ORIGINAL PUBLICATIONS
This thesis in based on data presented in the following articles, referred to by
their Roman numerals.
I Korhonen, J.M., Sclivagnotis, Y. and von Wright, A. (2007)
Characterization of dominant cultivable lactobacilli and their antibiotic
resistance profiles from faecal samples of weaning piglets. Journal of
Applied Microbiology 103: 2496-2503.
II Korhonen, J.M., Hassel, A., Komulainen, H. and von Wright, A.
Antimicrobial susceptibility of lactic acid bacteria from fecal samples of
weaned calves. Manuscript.
III Korhonen, J.M., van Hoek, A.H.A.M., Saarela, M., Huys, G., Tosi, L.,
Mayrhofer, S. and von Wright, A. (2010) Antimicrobial susceptibility of
Lactobacillus rhamnosus. Beneficial Microbes 1: 75-80.
IV Korhonen, J.M., Danielsen, M., Mayo, B., Egervärn, M., Axelsson, L.,
Huys, G. and von Wright, A. (2008) Antimicrobial susceptibility and
proposed microbiological cut-off values of lactobacilli by phenotypic
determination. International Journal of Probiotics and Prebiotics 3: 257-268.
V Guglielmetti, E., Korhonen, J.M., Heikkinen, J., Morelli, L. and von
Wright, A. (2008) Transfer of plasmid -mediated resistance to
tetracycline in pathogenic bacteria from fish and aquaculture
environments. FEMS Microbiology Letters 293: 28-34.


Contents 1. Introduction .............................................................................. 15
2. Review of the literature ......................................................... 17
2.1 ANTIBIOTIC RESISTANCE IN THE FOOD CHAIN ................................... 17
2.1.1 Mechanisms of antibiotic resistance ......................................................... 19
2.1.2 Antibiotic resistance determinations ........................................................ 20
2.2 LACTIC ACID BACTERIA ..................................................................... 21
2.2.1 Identification of LAB ............................................................................. 23
2.2.2 LAB of human and animal origins ........................................................... 24
2.2.3 Safety of LAB ....................................................................................... 25
2.3 ANTIBIOTIC RESISTANCE OF LAB ...................................................... 27
2.3.1 Phenotypic antibiotic resistance of LAB .................................................... 27
2.3.2 Genotypic antibiotic resistance of LAB ..................................................... 29
2.3.3 Horizontal transferability of antibiotic resistance from LAB in the food chain . 29
3. Aims of the study .................................................................... 31
4. Materials and methods .......................................................... 32
4.1 BIOLOGICAL ORIGINS OF LAB STRAINS (I-V) ..................................... 32
4.2 ISOLATION AND IDENTIFICATION OF LAB (I-IV) ............................... 33
4.2.1 Sampling and culturing of LAB from faecal samples (I-III) .......................... 33
4.2.2 Identification with biochemical methods (I-III) ........................................... 33
4.2.3 Identification with molecular biology methods (I-III, V) .............................. 33
4.2.4 LAB strain characterization (I-II) ............................................................ 34
4.3 MICROARRAY ASSAY FOR LB. RHAMNOSUS (III) ................................ 34

4.4 PLASMID TRANSFORMATION AND MATING PROTOCOL OF LACTOCOCCUS
SPECIES (V) ............................................................................................... 35
4.5 ANTIMICROBIAL RESISTANCE DETERMINATIONS (I-IV) ..................... 36
4.5.1 Agar dilution method (I-III) .................................................................... 36
4.5.2 Broth microdilution method (III-V) .......................................................... 37
4.5.3 Etest method (III-IV) ............................................................................. 37
5. Results and discussion ........................................................... 38
5.1 SPECIES DISTRIBUTION AMONG THE FAECAL SAMPLES OF PIGLETS AND
CALVES (I-II) ............................................................................................ 38
5.2 RAPD FINGERPRINTS OF PIGLET AND CALF ISOLATES (I-II) .............. 39
5.3 ANTIMICROBIAL SUSCEPTIBILITY PROFILES OF LACTOBACILLUS
SPECIES OF PIGLET ORIGIN (I) ................................................................. 44
5.4 ANTIMICROBIAL SUSCEPTIBILITY PROFILES OF LAB SPECIES OF CALF
ORIGIN (II) ............................................................................................... 46
5.5 ANTIMICROBIAL SUSCEPTIBILITY PROFILES OF LACTOBACILLUS
RHAMNOSUS AND PROPOSED MICROBIOLOGICAL CUT-OFF VALUES (III) 52
5.6 ANTIMICROBIAL SUSCEPTIBILITY PROFILES OF LACTOBACILLUS
SPECIES AND PROPOSED MICROBIOLOGICAL CUT-OFF VALUES (IV) ........ 53
5.7 ANTAGONISTIC ACTIVITY AND TRANSFORMATION OF LACTOCOCCUS
SPECIES (V) ............................................................................................... 58
5.8 TRANSFER OF THE TET(S) GENE FROM LACTOCOCCUS TO L.
MONOCYTOGENES (V) ................................................................................ 59
6. Conclusions ............................................................................... 60
7. References ............................................................................. 62

15
1. Introduction Lactic acid bacteria are a heterogeneous group of bacteria , many of them having received a generally recognized as safe (GRAS) or qualified presumption of safety (QPS) –status. These bacteria are widely found in nature, including the gastrointestinal and urogenital tracts of humans and animals, and are present in many fermented foods like salted gherkins, marinated olives, capers and salami, and d ifferent milk based products such as cheeses and yoghurts. Lactic acid bacteria have trad itionally been associated with these dairy products and with cereal-, vegetable- and meat-based fermented foods, either as intentionally added starters or due to their natural presence leading to spontaneous fermentation. Certain lactic acid bacteria are also used as probiotics added to confer health benefits to consumers or to improve animal production. In this respect, lactic acid bacteria species are economically very important to the food and feed industry. During the recent decades, there has been concern about the possibility of the spread of antibiotic resistance in the environment. According to the European Commission (2005), it has been estimated that somewhere from one to ten million tons of antibiotics have been released into the biosphere over the last 60 years. This has lead to a very strong selective pressure for the appearance of resistant bacterial strains. Much of the concern has been about pathogenic bacteria and their antibiotic resistances, since infections caused by these resistant micro-organisms are not only more complicated to treat, but the treatment is much more costly due to the more intensive and time consuming care needed in these cases. Since lactic acid bacteria are present in the gastrointestinal tract in large amounts and are also intentionally added to our d iet, concerns have been raised about the antibiotic resistance in these beneficial bacterial species. For example, lactic acid bacteria resistant to certain antibiotics could benefit the host (human or animal) by helping to maintain balance in the gastrointestinal tract in cases of d iarrhea caused by antibiotic treatment. However, there is a risk associated with the ability of these resistant strains to transmit the resistance factor (gene) to other, possibly pathogenic bacteria. This could complicate the treatment of a patient with an antibiotic resistant bacterial infection or d isease. Hence, the possibility of the circulation of genes coding for antibiotic resistance also from beneficial lactic acid bacteria, in the food chain via animals to humans, has been investigated .

16
At the beginning of 2004, the European Union launched a 6th framework project “Assessment and Critical Evaluation of Antibiotic Resistance Transferability in Food Chain” (ACE-ART), in which most of the work included in this thesis, was conducted . The major objective of this project was to critically evaluate the impact of antibiotic use in animal and plant production and in the prophylaxis and treatment of d isease in humans using non -pathogenic lactic acid bacteria as model organisms. In order to obtain a wide perspective of the current situation of antibiotic resistance patterns in lactic acid bacteria, a large array of these beneficial bacterial strains belonging to Lactobacillus, Lactococcus and Streptococcus thermophilus, together with Bifidobacterium, were included in the studies of ACE-ART -project. In this thesis, the antibiotic resistance profiles from faecal samples of weaning piglets and calves were determined. In addition, the antibiotic resistance of Lactobacillus rhamnosus strains from various sources (mostly human and food origins) w as studied . In summary, antimicrobial susceptibility and proposed microbiological cut-off values of lactobacilli by phenotypic determination were performed , containing both published and unpublished data of fourteen Lactobacillus species with their phenotypic antibiotic resistance phenotypes. Moreover, in order to detect the horizontal transfer of antibiotic resistance genes in the food chain, in vitro transfer of a plasmid derived tetracycline resistance gene, tet(S), was achieved from Lactococcus garvieae to the pathogenic Listeria monocytogenes.

17
2. Review of the literature
2.1 ANTIBIOTIC RESISTANCE IN THE FOOD CHAIN
There is a close association between the quantities of antimicrobials being used and the rate of development of resistance to these substances and thus the misuse of antibiotics in human medicine is believed to be the principal cause of the antibiotic resistance problem (Singer et al. 2003). Another aspect, however, is the selection of resistant bacteria in the food chain due to the heavy utilization of antimicrobial agents in animal husbandry (Teale 2002).
As early as 1969, the Swann report (Anonym. 1969) drew attention to the potential transfer of antibiotic resistan t bacteria from animals to the food chain and to humans. In animals, antibiotics are used for three d ifferent purposes: to treat sick animals (therapeutic use), to prevent infection in animals (prophylactic use) and to improve feed utilization and production, in other words, to convert feed into more body mass (growth promoters) ( Barton 2000, Singer et al. 2003). The therapeutic use of antibiotics is not questioned in farm animals, but the views of experts d iffer on the use of antibiotics as animal growth promoters, and there is debate whether the banning of these substances would have beneficial effects on human health. In the European Union, avoparcin (a glycopeptide, which produces cross-resistance to vancomycin), virginiamycin (a streptogramin), bacitracin (also used in human medicine), tylosin and spiramycin (both macrolides) have been banned as feed additives (Teale 2002, Nousiainen et al. 2004), and the EU is moving towards a total restriction on growth promoters, thus fulfilling the recommendation of the Swann report (1969) where it was stated that antibiotics used in human medicine should not be used as growth promoters. Nonetheless, the recommendations of the Swann report have not been adopted in many countries outside the EU (Teale 2002). For example, in the USA a total of 19 d ifferent antibiotics are allowed to be used for growth promotion , including several antibiotics, like penicillin and streptomycin, which are used also in human medicine (Singer et al. 2003).
If an antibiotic resistance is to cause a disease in humans via the food chain, certain events have to take place (Figure 1). First, there has to be an antibiotic resistant bacterium and a selection pressure present in a particular group of animals. The bacterium has to remain in the food process, through, for example, faecal contamination in the process or via a recontamination by improper handling, or lack of an adequate temperature treatment in the process. After consumption, the resistant bacterium has to colonize the GI-tract. (Singer et al. 2003) In conclusion, the transm ission of antibiotic resistance via the food chain is the same as for food borne pathogens. The risk of transmission

18
of antibiotic resistance from animals to humans is considerably reduced with proper food handling and good food preparation practices. (Singer et al. 2003) It should also be noted that the use of antimicrobials is not restricted to animal husbandry but also occurs in horticulture, for example the use of aminoglycosides in apple growing (Teale 2002).
Figure 1. Possible transmission routes of antibiotic resistance bacteria from animals to humans. Modified from Khachatourians 1998, Anonym 2004 and Claycamp and Hooberman 2004.

19
In Finland, the use of antimicrobial d rugs for animals has remained rather stable for several years; although a slight increase may have occurred . According to National Agency for Medicines, the consumption of veterinary antimicrobials was 15 100 kg year 2007, calculated as weight of active substance. In comparison, in the year 2001 the consumption was 13 800 kg. The possible reason for the increment remains to be clarified , i.e. is it because of an actual increase in antimicrobial consumption or simply due to statistical variation. The overall good resistance situation can be traced to the strict antimicrobial policy. However, pressure for increasing the use of antibiotics in Finland will probably rise in the future. This is mainly because of increased herd size and the trend towards more inten sive livestock production. (Myllyniemi et al. 2007)
2.1.1 Mechanisms of ant ibiot ic resistance The genetic basis for the development of antibiotic resistance in bacteria in the food chain is based on two facts. Firstly, the bacteria must come into contact with the antimicrobial agent in concern, and secondly, resistance against the antibiotic must develop (Khachatourians 1998, Levy and Marshall 2004). Resistance against a certain antimicrobial agent can be inherent in a bacterial species, this being referred to as intrinsic resistance, or “natural resistance”. In this case, the resistance is typical for all of the strains of that particular species. In contrast, the resistance is considered as acquired , when a strain of a normally susceptible species becomes resistant to an antimicrobial drug. (European Commission 2008)
The antibiotic resistance genes can be spread from one bacterium to another through several mechanisms. Intrinsic resistance is estimated to present a minimal potential for horizontal spread (between different bacterial species), as has been demonstrated for example with the chromosomal vancomycin resistance determinant of the Lactobacillus rhamnosus strain GG (Tynkkynen et al. 1998). Similar to intrinsic resistance, acquired resistance usually possesses a low risk of horizontal d issemination, when the resistance is a result of a chromosomal mutation. In contrast, acquired resistance is considered as having a higher potential for horizontal dissemination of antibiotic resistance, when the resistance genes are present on mobile genetic elements (plasmids and transposons). (Khachatourians 1998, European Commission 2008)
Antimicrobial drugs can be d ivided into d ifferent groups based on their mechanisms of action. Today there are more than 250 antibiotics available for therapeutic use (more than 100 of those are β-lactams), but these drugs act only against a few different bacterial target sites (van den Bogaard and Stobberingh 1996). These sites of action include cell wall synthesis, protein synthesis (targeting for 30S, 50S or tRNA), DNA gyrase or folic acid metabolism (Neu 1992). The major antibiotic families and their target of action relevant to this thesis are shown in Table 1.

20
Table 1. Major antibiotic families and their target of action. Sites of inhibition Group Antibiotic Cell wall synthesis β-lactams Amoxicillin (AMO) Ampicillin (AMP) Penicillin C (PEN) Glycopeptides Vancomycin (VAN)
Protein synthesis Aminoglycosides Gentamicin (GEN) Kanamycin (KAN) Neomycin (NEO) Streptomycin (STR)
Chloramphenicols Chloramphenicol (CHL) Tetracyclines Tetracycline (TET) Macrolides Erythromycin (ERY) Tylosin (TYL) Lincosamides Clindamycin (CLI) Lincomycin (LIN)
DNA replication/transcription Quinolones Enrofloxacin (ENR)
Folate synthesis Sulphonamides Trimethoprim (TMP)
2.1.2 Ant ibiot ic resistance determinat ions Antimicrobial susceptibility testing may be performed using d ifferent phenotypic test methods. In CLSI (Clinical and Laboratory Standards Institute, formerly NCCLS, National Committee on Clinical Laboratory Standards) the approved standards state that the methods of choice are agar dilution and broth microdilution (Anonym. 2007). Other widely used methods include the agar gradient method and commercial methods, such as Etest, which consists of a predefined gradient of antibiotic concentrations on a plastic strip (AbBiomerieux, Sweden).
In addition to phenotypic antibiotic resistance determinations, also genotypic detection of particular genes causing resistance may be performed. These genotypic methods include d ifferent PCR –based methods, southern hybrid ization, plasmid profiling and microarray (Aquilanti et al. 2007, Ammor et al. 2008).
The situation is clearest when the phenotypic and genotypic resistance patterns are in agreement. However, a phenotypically resistant bacterium strain may be genotypically “susceptible”. This is usually due to the fact that appropriate genes are not included in the test patterns, or th ere exist unknown resistance genes. Tetracycline, for example, has more than 40 d ifferent genes conferring antibiotic resistance d iscovered at the moment, and the number of tetracycline resistance genes continues to increase (Roberts 2005). With tetracycline, also new mosaic genes have recently been d iscovered (Patterson et al. 2007, van Hoek et al. 2008a). In contrast, a susceptible phenotype may also carry silent genes, which are observed with genotyping. The silence of antibiotic resistance may be caused by down -regulation in a promoter region or

21
by other mechanisms. Despite of their “silence”, they still could be a potential concern since they could be transferred to other species where they would be activated.
2.2 LACTIC ACID BACTERIA
Lactic acid bacteria (LAB) are gram -positive, acid -tolerant and non-spore forming cocci and rods. They are a heterogeneous group of bacteria comprising about 20 genera within the phylum Firmicutes. From a practical point of view the genera Aerococcus, Carnobacterium, Enterococcus, Lactobacillus, Lactococcus, Leuconostoc, Oenococcus, Pediococcus, Streptococcus, Tetragenococcus, Vagococcus and Weissella have been consid ered as the principal LAB. (Holzapfel et al. 2001, Axelsson 2004)
One common feature of the LAB is their ability to produce lactic acid as a major end product of their fermentation of hexoses. As fermenting organisms, LAB lack electron transport systems and cytochromes, and they do not have a functional Krebs cycle (Batt 2000). Based on end products of glucose metabolism (Figure 2), LAB can be d ivided into two groups, namely homofermentative and heterofermentative (Jay 2000).
LAB are widespread organisms and they may be found in many environments rich in carbohydrates. In addition to carbohydrates, LAB have complex nutritional requirements for aminoacids, peptides, fatty acid esters, salts, nucleic acid derivatives and vitamins (Tannock 2004). In short, they have complex nutritional requirements due to their lack of many biosynthetic pathways. On the other hand, they are found in a wide range of different environmental niches due to their good capacity for adaptation. In food products, they are found in dairy products, such as yoghurt and cheese, in fermented vegetables (olives, sauerkraut), in fermented meats (salami) and in sourdough bread (Tannock 2004). They are also associated with both terrestrial and marine animals (see chapter 2.2.2, LAB of human and animal origins).
Figure 2. Generalized pathways for the production of fermentation products from glucose by A: homofermentative LAB and B: heterofermentative LAB (Kandler 1983).

22
Lactobacillus species The genus Lactobacillus is the largest LAB group comprising at the moment around 140 species and 30 subspecies (Bernardeau et al. 2008, Claesson et al. 2008). These numbers are constantly being revaluated on the basis of modern molecular biology methods and whole genome-based techniques (Makarova et al. 2006, Felis and Dellaglio 2007).
Based on their fermentation characters, Lactobacillus species can be d ivided into three groups; obligately homofermentative, facu ltatively heterofermentative and obligately heterofermentative (Hammes and Vogel 1995, Axelsson 2004). Moreover, if one uses 16S phylogeny, then the Lactobacillus species can be d ivided into three groups: the L. casei-Pediococcus group, the Leuconostoc group and the Lactobacillus acidophilus/delbrueckii group (Collins et al. 1991, Hammes and Vogel 1995, Stiles and Holzapfel 1997). Since then, based again on 16S rDNA sequences, it was proposed to d ivide, the Lactobacillus species into five groups, namely Lb. acidophilus, Lb. salivarius, Lb. reuteri, Lb. buchneri and Lb. plantarum (Schleifer and Ludwig 1995). However, these classifications have generally been considered as unsatisfactory and also the use of 16S rRNA genes as phylogenetic markers has been criticized (Claesson et al. 2008). New proposals for the classification of the lactobacilli species claim that the genus could be d ivided into seven or eight groups (Dellaglio and Felis 2005, Hammes and Hertel 2006). As complete genome sequences become available, the high d iversity of Lactobacillus has also been suggested to require the creation of new, subgeneric d ivisions (Canchaya et al. 2006, Claesson et al. 2008). Lactococcus species In contrast to Lactobacillus species, the genus Lactococcus comprises at the moment only five species, namely Lactococcus garvieae, Lc. lactis (subspecies cremoris, ssp. hordniae and ssp. lactis), Lc. piscium, Lc. plantarum, and Lc. raffinolactis (http:/ / www .ncbi.nlm.nih.gov/ Taxonomy/ ). They can initially be d istinguished from each other by their possible ability to grow at temperatures above 40 °C and in >4 % sodium chloride (Batt 2000).
Plasmids are common components of Lc. lactis genomes, these being d iverse in size, copy number and d istribution. Plasmids often carry some significant characteristics, such as carbohydrate fermentation, proteolysis, polysaccharide production or bacteriosine production. Occasionally some plasmids also encode the determinants necessary for conjugation, and transfer of conjugal plasmids may be a significant factor in horizontal gene transfer, likewise other transposable elements (Courtney 2000). Weissella species Weissella species have been classified as Lactobacillus or Leuconostoc species until the reclassification propose by Collins et al. 1993. Weissella species are heterofermentative lactics (Batt 2000, Jay 2000), and comprise at the moment of fourteen species (http:/ / www.ncbi.nlm.nih.gov/ Taxonomy/ ). Weissella ssp. have been found in different fermented vegetable based foods, such as

23
fermenting cassava (Kostinek et al. 2007) and sauerkraut (Plengvidhya et al. 2007), in sourdough (Di Cagno et al. 2006) and in blood sausage, “Morcilla de Burgos” (Santos et al. 2005) and in “Shochu”, a trad itional Japanese liquor (Endo and Okada 2005). Weissella ssp. have also been isolated in human samples from vaginal microbiota (Nam et al. 2007) and saliva (Kang et al. 2006).
2.2.1 Ident ificat ion of LAB Reliable identification of LAB is crucial in many tasks, e.g. applications in industrial processes (technological properties), but also safety and quality control problems (Temmerman et al. 2004). Previously identification of bacterial species was based on the phenotypic characterization, but molecular biology (genotypic) methods, largely DNA-based techniques, offer much greater d iscriminatory power, all the way to d ifferentiation of individual strains. Usually a combination of both phenotypic and genotypic identification techniques (polyphasic approach) is preferred (Temmerman et al. 2004, Aquilanti et al. 2007).
Phenotypic methods include morphological analysis, growth characteristics and sugar fermentation profiles (e.g. with commercial API-tests, BioMerieux, France). However, these methods have a low taxonomic resolution and often allow differentiation only at the genus level (Temmerman et al. 2004). On the other hand, no special laboratory equipment is required to perform these tests. Cultivation, microscopy and sugar fermentation profiles are important, even though in most of the cases, these tests are insufficient for accurate species identification due to the great number of d ifferent LAB species with similar phenotypic characteristics.
The use of molecular tools has revolutionized the identification of many bacterial species, including LAB. Many of these techniques are based on the polymerase chain reaction (PCR) using oligonucleotide primers to amplify targeted DNA fragments. These PCR primers may be designed to different taxonomical levels, from genus-specific (for example d ifferentiation between Weissella and Leuconostoc species, Schillinger et al. 2008) to the species-specific level (for example d ifferentiation between Lb. brevis, Lb. fermentum and Lb. parabuchneri, (Coton et al. 2008)), to sub-species level (for example between Lc. lactis subsp. lactis and Lc. lactis subsp. cremoris (Pu et al. 2002)), and also further to the strain-level (for example identification of Lb. reuteri strain 35 (Coudeyras et al. 2008)). This approach is suitable when the bacterial species has been preliminary characterized with other, usually phenotypic, methods.
Another genotypic method which can be used to identify unknown bacterial isolates is ribosomal DNA sequencing, usually targeted to 16S or 23S ribosomal subunits. The obtained sequence is compared with DNA sequences in online databases, such as EMBL (European Molecular Biology Laboratory) or Genbank using a sequence search engine like BLAST or FASTA (http:/ / www.ebi.ac.uk/ embl/ ; http:/ / blast.ncbi.nlm.nih.gov/ Blast.cgi/ , (Altschul et al. 1990).

24
A widely used DNA fingerprint technique based on the PCR –method is randomly amplified polymorphic DNA (RAPD), where short arbitrary primers are used to randomly amplify DNA fragments. RAPD-PCR has been used with LAB in several studies and has successfully typed LAB strains (Urso et al. 2006, Aquilanti et al. 2007). Despite the usefulness of RAPD-PCR for strain typing it also has been criticized . Most concerns are related to the reproducibility of the results. However, with careful optimization, including ensuring the quality of the template DNA, polymerase enzyme and PCR conditions, the RAPD-PCR method has been found reliable and reproducible (Ravelo et al. 2003, Foschino et al. 2008).
According to EFSA, an indispensable pre-requisite for determining antibiotic resistance with accuracy is the correct identification of the strain of concern at the species level by means of molecular taxonomy methods (European Commission, 2008).
2.2.2 LAB of human and animal origins A composition of LAB species d iffers between environments. Especially in humans, detailed reports about LAB species and strains, as useful bacteria, have received enormous attention in recent decades. According to Reuter et al. (2001), the major autochthonous Lactobacillus species found both in infants and adults are Lb. ruminis, Lb. salivarius, Lb. reuteri and Lb. gasseri (Reuter 2001). Dal Bello and co-workers (2002) found also food -associated LAB, namely Lb. sakei and Ln. mesenteroides as intestinal inhabitants, when alternative incubation conditions (30 ºC, 2 % O2) were used (Dal Bello et al. 2003).
The list of d ifferent lactobacilli species found in humans is inevitably much longer. However, most of the LAB species found in human intestinal samples are transient over time, and colonize the intestinal tract for only a short period of time (Walter 2005). In addition, variation between individuals most probably influences the species composition, due to antibiotic treatments, d ifferences in the d iet consumed or other environmental factors (Donohue 2004, Dicksved et al. 2007). Taking into account the recent development in taxonomy of LAB, the complexity of the species composite on of LAB will most probably increase, not least when culture independent methods are used to identify the strain . For example, the PCR-DGGE (denaturing gradient gel electrophoresis) system described by Walter et al. (2001) permitted not only the detection of Lactobacillus species considered to be normal intestinal inhabitants but also several LAB commonly associated with food and often used as starter organisms. These non-cultivable LAB species included lactobacilli (Lb. sakei and Lb. curvatus), pediococci (P. pentosaceus), leuconostocs (Ln. mesenteroides) and weissellas (W. confusa). (Walter et al. 2001).
In piglets as in humans, the most dominant LAB species found are Lb. ruminis, Lb. reuteri/fermentum , Lb. salivarius and Lb. acidophilus/delbrueckii group lactobacilli (Naito et al. 1995, Leser et al. 2002, Yin and Zheng 2005, Yun et al. 2009). This is not surprising, since both humans and pigs are monogastrics. The LAB species composition varies in piglets depending to some extent on which methods are being used for analysis (culture-dependent versus culture-

25
independent methods), and between d ifferent gastrointestinal locations. In addition, the diet and the age of the animals have an influence on the LAB species composition. Lb. reuteri has been observed to be a major component of Lactobacillus species in intestine of pigs, but also in chickens, cattle, dogs, mice and rats (Mitsuoka 1992).
The identification of lactobacilli species composition in calves has not received as much attention as in humans or pigs when considering LAB isolated from faecal samples. Instead , much of the work on calves and adult cows has focused on which LAB species can be found in the rumen. The most frequently found lactobacilli species from faecal samples in calves are Lb. mucosae and Lb. reuteri, followed with Lb. acidophilus/delbrueckii group lactobacilli (Busconi et al. 2008). Streptococci were other important LAB together in addition to the lactobacilli found in the study of Busconi et al. (2008). A novel finding in faecal samples of calves has been Weissella species, also Lactococcus species have been found (see publication II, Korhonen et al.). Interestingly, Lc. garvieae has been associated with mastitis in cow s (Devriese et al. 1999).
In fish, the dominant LAB species are carnobacteria, for example Carnobacterium divergens and C. piscicola. Lactobacilli and lactococci have also been isolated from fish as well as several other LAB like aerococci and streptococci. (Gatesoupe 2008) However, LAB normally account for only a trivial percentage of the intestinal microbiota of fish, which may partly be due to the inappropriate cultivation methods used in these studies. The main reason for not isolating LAB from aquatic animals might be that the LAB from fish are generally slow -growing microorganisms and for this reason, a longer incubation time of up to four weeks and a lower incubation temp erature (4-12 ºC) have been recommended, as well as resorting to non-culturable methods (Ringo and Gatesoupe 1998).
2.2.3 Safety of LAB Lactic acid bacteria have been used all over the world in various trad itional and industrial food fermentations. Moreover, LAB have intentionally been added as probiotics in order to achieve beneficial effects on health of humans and animals. Members of Lactobacillus and Bifidobacteria species are generally recognized as safe (GRAS-status), since they have a long history of use (Donohue 2004).
However, the huge variability of LAB and also other microorganisms require a more detailed assessment of safety. Due to the apparent need to develop a tool for setting priorities associated with the risk of using these bacteria in food or feed production, EFSA has been considering how best to conduct a formal assessment of safety. Thus, a system was proposed for a pre-market safety assessment of selected grou ps of microorganisms leading to granting a “Qualified Presumption of Safety (QPS)”. Therefore, EFSA proposed that a safety assessment of a defined taxonomic group, such as a genus or group of related species could be made based on establishing identity, body of

26
knowledge, possible pathogenicity and end use (European Commission 2007). Thus EFSA has stated (2007) that today a total of 33 Lactobacillus species can be considered to have QPS-status (European Commission 2007); these species are shown in Table 2. Table 2. Lactobacillus species with QPS- status according to EFSA. Lactobacillus acidophilus Lactobacillus amylolyticus Lactobacillus amylovorus Lactobacillus alimentarius Lactobacillus aviaries Lactobacillus brevis Lactobacillus buchneri Lactobacillus casei Lactobacillus crispatus Lactobacillus curvatus Lactobacillus delbrueckii
Lactobacillus farciminis Lactobacillus fermentum Lactobacillus gallinarum Lactobacillus gasseri Lactobacillus helveticus Lactobacillus hilgardii Lactobacillus johnsonii Lactobacillus kefiranofaciens Lactobacillus kefiri Lactobacillus mucosae Lactobacillus panis
Lactobacillus paracasei Lactobacillus paraplantarum Lactobacillus pentosus Lactobacillus plantarum Lactobacillus pontis Lactobacillus reuteri Lactobacillus rhamnosus Lactobacillus sakei Lactobacillus salivarius Lactobacillus sanfranciscensis Lactobacillus zeae
In addition to Lactobacillus species, also other LAB species have been granted QPS –status. They include three leuconostocs, (Ln. citreum, Ln. lactis and Ln. mesenteroides), three pediococci (P. acidilactici, P. dextrinicus and P. pentosaceus), Lc. lactis and Streptococcus thermophilus (European Commission 2007).
One of the difficulties in the QPS approach is the difficulty to apply a strain-to-strain approach to undefined microbial cultures, as are found in many fermented foods, such as trad itional dried sausages and cheeses; where the fermentation is based on the use of these undefined cultures and/ or backslopping (Rossetti et al. 2009). Since systematic investigations have rarely been undertaken, there is no convincing body of knowledge available for a given microorganism. Nonetheless, in many instances there exists a long and documented history of safe use of LAB in foods.
Some LAB have also been associated with d isease, although this occurs in very rare cases, where they can cause opportunistic infections in people with severe underlying illnesses. LAB have been isolated from endocarditis, bacteremia, bloodstream and local infections (Ishibashi and Yamazaki 2001, Cannon et al. 2005). In most cases, the infective bacterium has been shown to be of host origin. However, there are a few cases where the infection has also been associated with the consumption of probiotics.
One of the most important safety aspects of LAB is their resistance towards antimicrobial drugs that might be transferred to other, possib ly pathogenic, bacterial species, see chapter 2.3.3, Horizontal transferability of antibiotic resistance in the food chain.

27
2.3 ANTIBIOTIC RESISTANCE OF LAB
Studies of antibiotic resistance of LAB have not been extensivelly investigated until recently, in contrast to the situation with pathogenic species and their antibiotic resistance. However, interest in LAB and their antibiotic resistances has recently gained strength since the resistant determinants are known to be able to be transferred between bacterial species, also from beneficial bacteria to pathogens (see below).
2.3.1 Phenotypic ant ibiot ic resistance of LAB The first step in characterizing LAB species as being either susceptible or resistant to antibiotics is to determine the susceptible/ resistant patterns with phenotypic methods. This is easier said than done, due to the various methods available. These include factors like size of the inocula and incubation time (Egervärn et al. 2007), different test methods (broth versus agar applications), including microdilution (Kushiro et al. 2009), Etest (Danielsen and Wind 2003), agar d ilution (Florez et al. 2005) and d isc d iffusion (Gevers et al. 2000). Moreover, d ifferent growth media have been used for actual testing, including Iso Sensitest, MRS, M17 and Müller-Hinton (Huys et al. 2002, Hummel et al. 2007).
In the study of Klare and co-workers (2005), a new growth medium, LSM (LAB susceptibility test medium) was developed particularly to support the growth of Lactobacillus, Lactococcus, Pediococcus and Bifidobacteria but which would not have any antagonistic interactions between components of the medium and specific antimicrobial agents (Klare et al. 2005). Subsequently, several studies on phenotypic antibiotic resistance of LAB have been conducted using LSM broth and agar (Klare et al. 2007, Devirgiliis et al. 2008, Huys et al. 2008, Devirgiliis et al. 2009, Haakensen et al. 2009, Kushiro et al. 2009) assisting in the comparison of results obtained in different laboratories and on the other hand easing the determination of comparable MICs and the safety evaluation.
The European Food Safety Authority has recently (European Commission 2008) updated the microbiological breakpoints that categorize the LAB as either resistant or susceptible, see Table 3. The major difference to the previous microbiological breakpoints given by EFSA (European Commission 2005) is that the categories have been d ivided more specifically according to species. This means that the correct taxonomical identification is even more crucial, but on the other hand it does provide tools for assessing more precise MIC values of the LAB species under study. When an adequate number of strains of a particular species have been studied (in the ACE-ART project this was set as 50 strains) and the MIC values determined , the microbiological breakpoints (or epidemiological cut-off values) can be estimated . In some cases, however, the d istribution of MIC values is not “bell-shaped” and the MIC distributions cover more than optimal five dilutions. This causes uncertainties with the proposed cut-off values, and thus one has to apply the cut-off value only as a conservative estimation.

28
Table 3. Microbiological breakpoints for selected LAB (European Commission. 2008). Species/group AMP VAN GEN KAN STR ERY CLI Q+D TET CHL Lb. oblig. homoferm. 1 2 16 16 16 1 1 4 4 4 Lb. helveticus 1 2 16 16 16 1 1 4 4 4 Lb. acidoph./delb. 1 2 16 16 16 1 1 4 4 4 Lb. oblig. heteroferm. 2 n.r. 16 16 64 1 1 4 8 4 Lb. reuteri 2 n.r. 8 16 64 1 1 4 16 4 Lb. fermentum 1 n.r. 16 32 64 1 1 4 8 4 Lb. facult. heteroferm. 4 n.r. 16 64 64 1 1 4 8 4 Lb. plantarum 2 n.r. 16 64 n.r. 1 1 4 32 8 Lb. rhamnosus 4 n.r. 16 64 32 1 1 4 8 4 Lb. paracasei 2 n.r. 32 64 n.r. 1 1 4 4 4 Enterococcus 4 4 32 512 128 4 4 4 8 8 Pediococcus 4 n.r. 16 64 64 1 1 4 2 4 Leuconostoc 2 n.r. 16 16 64 1 1 4 8 4 Lc. lactis 2 4 32 64 64 2 4 4 8 8 AMP: ampicillin; VAN: vancomycin; GEN: gentamicin; KAN: kanamycin; STR; streptomycin; ERY: erythromycin; CLI: clindamycin; Q+D: Quinupristin + dalfopristin; TET: tetracycline; CHL: chloramphenicol; n.r., not required.
The Lactobacillus species have been found susceptible to many cell wall synthesis inhibitors, like penicillins and ampicillin (Danielsen and Wind 2003, Coppola et al. 2005), in contrast to glycopeptides such as vancomycin, most Lactobacillus species, excluding obligate heterofermentative species, have been found to be resistant to these types of antibiotics. However, the resistance towards vancomycin has been demonstrated being as intrinsic (Tynkkynen et al. 1998) and should not be compared with transmissible, plasmid -mediated resistance found in enterococci (Leclercq et al. 1992).
Lactobacillus species are usually susceptible to chloramphenicol, erythromycin and clindamycin, antibiotics that inhibit protein synthesis, (Coppola et al. 2005, Klare et al. 2007). In addition, resistance against inhibitors of nucleic acid synthesis, such as trimethoprim, seems to be intrinsic, although further characterizations are required on this topic (Ammor et al. 2007).
Resistance to tetracycline has been observed more often among Lactobacillus species, and it has been shown to have a wide range of MICs (Korhonen et al. 2008), also with a multimodal distribution of MICs, probably due to the extensive variability of tetracycline resistance mechanisms conferring d iverse levels of susceptibility (Roberts 2005). Especially with tetracycline, molecular methods should be applied in order to revea l the nature of resistance, i.e. is it due to intrinsic mechanisms, mutation or added, mobile genes.
Resistance against aminoglycosides, such as neomycin, kanamycin, streptomycin and gentamicin has been observed more frequently among lactobacilli (Danielsen and Wind 2003, Coppola et al. 2005, Zhou et al. 2005). The medium of choice has been demonstrated to significantly affect to the classification of LAB as being either susceptible or resistant (Huys et al. 2002), especially with aminoglycosides and MRS medium.

29
2.3.2 Genotypic ant ibiot ic resistance of LAB When a strain clearly deviates from other strains of that particular species , as determined with phenotypic methods, the antibiotic resistance needs to be further studied with genotypic methods. The potentially transferable genes in LAB have been described in multiple studies and have been reviewed in Ammor et al. 2007. Two of the most commonly observed resistance genes in LAB found so far are tet(M) for tetracycline resistance and erm(B) for erythromycin, followed with cat genes coding for chloramphenicol resistance (Lin et al. 1996, Danielsen 2002, Gevers et al. 2003, Cataloluk and Gogebakan 2004).
2.3.3 Horizontal t ransferabilit y of ant ibiot ic resistance from LAB in the food chain Horizontal transfer of genetic material may happen via three d ifferent mechanisms: transduction, transformation or conjugation. In transduction, the DNA is transferred from one bacterium to another via bacteriophages. The importance of bacteriophages in d isseminating antibiotic determinants is, however, questionable, because the phages are often highly species -specific. In transformation, the DNA is released from a bacterium and taken up by another. Similar to the situation with transduction, transformation is not believed to be a very important mechanism of transfer of antibiotic resistance (Ammor et al. 2007). In contrast, conjugation, i.e. the d irect cell-to-cell contact, potentially can achieve horizontal gene transfer, as it has been shown to be a mechanism of transfer of genetic information with a broad host range (Courvalin 1994).
The possible transfer of antibiotic resistance genes betw een bacterial species have been studied mostly in harmful or pathogenic species , but also recently with LAB. The vast majority of the experiments have been made in vitro, using methods such as filter-mating (Gevers et al. 2003, Klare et al. 2007, Ouoba et al. 2008), although these in vitro methods do not mimic the circumstances in nature, and results obtained cannot be compared with the results achieved or expected using in vivo methods. The transferability of antibiotic resistance genes in the gastrointestinal tract (GIT) from LAB is not straightforward , since the GIT is a hostile environment to many allochthonous bacteria. Moreover, studies made in vivo usually are based on “worst-case scenario”, simulating very high daily intake of food products containing the resistant bacteria (Jacobsen et al. 2007).
One well-characterized resistance gene in LAB, originally detected in Enterococcus faecalis is the broad-host range plasmid pAMβ1, which has been transferred from Streptococcus lactis (Lactococcus lactis at present) to Lb. reuteri and between lactobacilli (Tannock 1987), and also from Lc. lactis to Enterococccus and Lactobacillus species (both in vitro and in vivo in the mouse gastrointestinal tract) (Morelli et al. 1988, Gruzza et al. 1993, Igimi et al. 1996). In the recent study of Devirgiliis et al. (2009), tetracycline-resistant Lb. paracasei strains were

30
identified in samples of milk and natural whey starter cultures . A transposon Tn916 including tet(M) was transferred to E. faecalis strain JH2-2 in mating experiments, but only at a low conjugation frequency (Devirgiliis et al. 2009). Recently, experiments of antibiotic resistance transferability in vivo were also conducted from Lb. plantarum to E. faecalis (Jacobsen et al. 2007). The transfer frequencies have been observed to increase when the animals have received the antibiotic in question at subtherapeutic levels (Igimi et al. 1996, Salyers and Shoemaker 1996, Licht et al. 2003) in their drinking water or feed , suggesting that increasing the antibiotic pressure can amplify the transfer of antibiotic resistance between bacterial species.
All of these above studies indicate that antibiotic resistant factors may be transferred from food related bacterium species to other, potentially pathogenic species, such as enterococci. Another interesting point of view is that intestinal bacteria might also interact with bacteria that are just passing through the colon (commensal bacteria), allowing these bacteria to acquire and transmit antibiotic resistance genes (Salyers et al. 2004).

31
3. Aims of the study The main objective of th is thesis was to address the antimicrobial resistance patterns of LAB. The work started with the isolation and characterization of LAB from animal and human origins, and continued with the accurate species identification followed by the determination of antimicrobial susceptibility profiles of antimicrobial substances from different classes. Horizontal transferability of antibiotic resistance gene tet(S) coding for tetracycline was also examined between Lactococcus and Listeria species. The specific aims of the individual studies were:
To isolate, identify and define the distribution of the Lactobacillus species in weaning piglets (I);
To determine the susceptibility phenotype to antibiotics in lactobacilli
species isolated from piglet faecal samples (I);
To isolate, identify and define the d istribution of LAB species in young calves (II);
To determine the susceptibility phenotype to antibiotics in LAB isolated
from calf faecal samples (II);
To determine the MICs of Lactobacillus rhamnosus strains mostly from human and food isolates and to study the prevalence of antimicrobial resistance genes with microarray analysis (III);
To summarize phenotypic data of antimicrobial resistances and to
devise tentative microbiological cut-off values for fourteen Lactobacillus species (IV);
To study the transfer of plasmid derived antibiotic resistance to
tetracycline from Lactococcus garvieae to Listeria monocytogenes (V).

32
4. Materials and methods
4.1 BIOLOGICAL ORIGINS OF LAB STRAINS (I-V)
The biological origins of LAB strains used in the antibiotic susceptibility tests in studies I-IV are shown in Table 4. Table 4. Biological origins of the LAB species.
Biological origin
Number of strains
Study I Study II Study III Study IV
Animals Pig 129 45
Calf 104 1
Poultry 18
Other 29
Humans 57 148
Food Dairy 9 220
Meat 66
Plant 111
Other 2
Other/unknown 6 38
Total 129 104 75 675
The biological origins of the strains used in study V were: � intestinal sample of salmonid fish (Oncorhynchus mykiss), Lc. lactis ssp. lactis
strain KYA7; � kidney of yellowtail, (Seriola lalandi), Lc. lactis ssp. garvieae strain DSMZ
6783; � human, Listeria monocytogenes strain ATCC 7644; � water in aquaculture, Listeria monocytogenes strains LMK8 and LMK16.

33
4.2 ISOLATION AND IDENTIFICATION OF LAB (I-IV)
The LAB isolates were identified with conventional, cu lture based methods (morphology and sugar fermentation profiles) and further with molecular biology based methods (16S rDNA based PCR, species-specific PCR, RAPD).
4.2.1 Sampling and culturing of LAB from faecal samples (I-III) Fresh faecal samples were collected from piglets, calves and human volunteers and stored refrigerated until analysis (Studies I, II and III, respectively). The piglet faecal samples (n=78) were received from MTT (Agrifood Research Finland), Hyvinkää, Finland and the calf faecal samples (n= 36) were collected from the area of Northern Savo, Finland. Some of the human samples (n = 19, total n = 75) were from adult volunteers living in Kuopio, Finland , while other human samples had been gathered in earlier studies. The fresh samples were serially d iluted in 0.1% peptone saline suspension to a final d ilution of 10-8. Appropriate d ilutions were then plated onto MRS and/ or LAMVAB (Hartemink et al. 1997) agar plates and incubated anaerobically at 37°C for 48 h.
4.2.2 Ident ificat ion w ith biochemical methods (I-III) The isolated LAB were selected from the MRS plates on the basis of their morphology, gram-staining and sugar fermentation profiling, using commercial API CHL (Lactobacillus species) and API Strep (Lactococcus and Streptococcus species) –kits with APIWEB database, bioMerieux SA, France.
4.2.3 Ident ificat ion w ith molecular biology methods (I-III, V) Biochemically identified LAB isolates were further character ized with molecular biology methods. The DNAs from the pure isolates were extracted using lysozyme, proteinase K, chloroform extraction and ethanol precipitation. The DNA was used as a template for partial 16S rDNA sequencing targeting for a 676-bp fragment of the 16S rDNA gene (studies (I-II). The primers used were 9f (5´GAGTTTGATCCTGGCTCAGGA´3) (Chagnaud et al. 2001) and 685r (5´TCTACGCATTTCACCGCTAC´3) (E. coli numbering system). The purified amplification products were subjected to sequencing, and the results achieved compared with the EMBL nucleotide database using the BLAST database sequence search engine. With Lb. rhamnosus strains (III), a partial 16S rDNA sequence (fragment size 863 bp) was applied in verifying the species as Lb. rhamnosus.
In study II, species-specific PCR methods were used for Lb. rhamnosus (Alander et al. 1999) (used also in study III), Lb. brevis (Guarneri et al. 2001), Lb. curvatus (Lee et al. 2004), Lb. farciminis (Rachman et al. 2003), and Lb. plantarum group, namely Lb. pentosus, Lb. plantarum, and Lb. paraplantarum (multiplex-PCR, Torriani et al. 2001).
In study V, total genomic DNA from Lactococcus and Listeria strains was extracted using the NucleoSpin Tissue kit (Macherey-Nagel, Germany) and the plasmid DNA using the Wizard Plus SV Minipreps DNA Purification System

34
kit (Promega, Madison, U.S.A.) enhanced with the lysozyme technique. For Southern hybrid ization, primers tetS-FW (5´GGAGTACAGTCACAAACTCG´3) and tetS-RW (5´GGATATAAGGAGCAACTTTG´3) targeting for a PCR product of 335 bp in size were used .
4.2.4 LAB st rain characterizat ion (I-II) The LAB isolates of piglet and calf origin were further analyzed with RAPD method using primer P1 (5´ACGCGCCCT´3) in order to d ifferentiate the strains and to avoid duplicates of isolates within a certain species in the antimicrobial susceptibility tests. The RAPD fingerprints were analyzed using the GELCOMPAR II software from Applied Maths, Belgium.
4.3 MICROARRAY ASSAY FOR LB. RHAMNOSUS (III)
L. rhamnosus strains were studied with the microarray method in order to identify the potential erythromycin, streptomycin and tetracycline resistance genes in more detail. Genes coding for chloramphenicol resistance were also included in the microarray (not tested by phenotypic methods). The antibiotic resistance genes represented on the oligonucleotides microarray are given below (also presented in Mättö et al. 2007 and van Hoek and Aarts 2008b).
Aminoglycoside resistance genes: aac(3)-Ia; aac(3)-Ib; aac(3)-IIa; aac(3)-IIIb; aac(3)-IIIc; aac(3)-VII; aac(6' )-aph(2' ); aac(6' )-Ib; aac(6' )-Il; aac(6' )-Iq; aac(6' )-Iy; aac(6)-II; aac(6)-IIb; aac(6)-IIc; aacA1; aacA4; aacA5; aacA7; aacC1; aacC2; aacC3; aacC9; aadA1; aadA2; aadA6; aadA10; aadA11; aadA13; aadB; aadD; aadE; aph(2' )-Ib; aph(2' )-Ic; aph(2' )-Id; aph(3' )-Id; aph(3' )-IIa; aphA1; aphA1-IAB; aphA2; aphA3; aphA6; aphA7; aphE; nptII; sat2; sat3; sat4; strA ; strB.
β-lactam resistance genes (for example blaACC-01_03 means that blaACC-01, blaACC-02 and blaACC-03 are represented on the microarray): blaACC-01_03; blaACT-
01,02; blaCARB-04_09; blaCMY-01,09_11,19; blaCTX-M-01 group; blaCTX-M-02 group; blaCTX-M-09 group; blaDHA-01_02; blaFOX; blaIMP; blaKPC; blaMIR; blaMOR; blaMOX-01; blaOXA-
01_07,10,13_17,19_35,37,40,47_49,56,72_74,101,102; blaPER-01_03; blaPSE-01; blaROB-01; blaTEM; blaVIM-01_11,14; blaZEG-
01.
Chloramphenicol resistance genes: cat; cat(pC194); cat-TC; catII; catIII; catA1; catA3; catB; catB2; catB3; catB6; catB8; catB9; catD; catP; catQ; cmlA ; cmlA1; cmlA4; cmlA5; cmlA6; cmlA7; cmlB; floR.
Macrolides lincosamide and streptogramin resistance genes: ere(A);
ere(A2); ere(B); erm(A); erm(B); erm(C); erm(D); erm(F); erm(FS); erm(FU); erm(G;) erm(GM); erm(GT); erm(J); erm(K); erm(Q); erm(T); mef(A); mef(E);

35
mph(A); mph(BM); mph(C); mph(K); msr(A); msr(B); msr(SA); sat(A); sat(G); vat; vat(A); vat(B); vat(C); vat(D); vat(E); vat(E3_E8); vga; vga(A); vga(A)LC; vgb; vgb(B).
Sulfonamide resistance genes: sul1; sul2; sulA .
Tetracycline resistance genes: otr(A); otr(B); tet(30); tet(31); tet(32); tet(33);
tet(34); tet(35); tet(36); tet(37); tet(A); tet(A(P)); tet(B); tet(B(P)); tet(C); tet(D); tet(E); tet(G); tet(H); tet(J); tet(K); tet(L); tet(M); tet(O); tet(Q); tet(S); tet(T); tet(U); tet(V); tet(W); tet(X); tet(Y); tet(Z).
Trimethoprim resistance genes: dfrA ; dfrA1; dfrA2; dfrA3; dfrA5; dfrA6;
dfrA7; dfrA8; dfrA9; dfrA10; dfrA12; dfrA14; dfrA15; dfrA16; dfrA17; dfrA19; dfrA21; dfrA22; dfrA23; dfrB2; dfrB3; dfrC; dfrD.
Vancomycin resistance genes: vanA ; vanB; vanC1; vanC2/ C3; vanD; vanE.
4.4 PLASMID TRANSFORMATION AND MATING PROTOCOL OF LACTOCOCCUS SPECIES (V)
In study V, Lc. garvieae strain DSMZ 6783 was used as a recipient in electroporation for plasmid located tet(S) gene derived from L. lactis ssp. lactis strain KYA-7. Plasmid located tet(S) gene from Lc. lactis ssp. lactis KYA7 strain was transformed by electroporation to Lc. garvieae DSMZ 6783 by the method described in Holo and Nes 1989. Pure culture of strain DSMZ 6783 was grown on M17 medium with 0.5 M sucrose and 1 % glycine, and washed twice with electroporation buffer. This suspension was mixed with purified plasmid DNA and exposed to an electrical pulse (Bio-Rad Laboratories, CA, USA). Transformants (TF) were counted from tetracycline –selective (5 μg/ ml) plates after incubation at 30 ºC for 72 h, and verified using species-specific primers (Pu et al. 2002).
Furthermore, the ability of transformed Lc. garvieae DSMZ 6783 (TF-α) to transfer the tet(S) resistance gene was examined by filter mating. Recipients in these conjugation studies for tet(S) gene were Listeria monocytogenes strains ATCC 7644, LMK8 and LMK16. Overnight grown bacterial cells were mixed with a 1:10 ratio (donor:recipient), collected on a filter, washed with 0,9 % saline suspension (2 ml) and incubated on BHI agar plate with 0,2 μg/ ml of tetracycline at 37 ºC overnight. After incubation , serial d ilutions were made on PALCAM Listeria selective agar with 10μg/ ml of tetracycline and incubated at 37 ºC for 72 h. Transconjugants (TCs) were identified by the API Listeria test (bioMerieux, SA, France).

36
4.5 ANTIMICROBIAL RESISTANCE DETERMINATIONS (I-IV)
Antimicrobial resistance determinations were mad e with three d ifferent methods. In studies I and II, the determinations were made by agar d ilution, in study III by agar dilution, broth microdilution and Etest , in study IV by broth microdilution and Etest methods and in study V by broth microdilution . The medium used was LSM (Lactic acid bacteria susceptibility medium, Klare et al. 2005), which has been especially designed to detect antibiotic resistances in LAB, except for experiments with Listeria monocytogenes (study V), where the cation adjusted Müller-Hinton (CAMBH, Difco) was used . Antibiotics used in studies I-IV are listed in Table 5.
Table 5. Antimicrobial agents for phenotypic susceptibility determination used in studies I - IV. Antibiotic Study I
(Piglets) Study II (Calves)
Study III (Lb. rhamnosus)
Study IV (Lactobacillus ssp.)
Ampicillin � � � �
Clindamycin � � � �
Erythromycin � � � �
Gentamicin � � � �
Streptomycin � � � �
Tetracycline � � � �
Chloramphenicol � �
Kanamycin � �
Trimethoprim � �
Vancomycin � �
Ciprofloxacin �
Amoxicillin �
Enrofloxacin �
Lincomycin �
Neomycin �
Penicillin �
Tylosin �
4.5.1 Agar dilut ion method (I-III) Single pure colonies were suspended in sterile 0.9 % saline and the density was adjusted with spectrophotometry to an OD625 of 0.16 – 0.20, which is estimated to correspond to McFarland standard 1 or 3 × 108 CFU/ml. Subsequently, 1 μl aliquots of bacterial suspensions were spotted onto the LSM agar along with the antibiotics. The appropriate dilutions (see table 6, Ericsson and Sherris 1971) of the antibiotic solutions were prepared in a twofold series in LSM agar and plates were used within a week of their preparation to avoid any loss of antibiotic activity. With certain antibiotics, solvents (methanol, ethanol, DMSO or NaOH) were used to dilute the substances. The

37
effect of solvent agents was controlled in susceptibility tests. The growth of the bacteria was compared to the growth on LSM plates without any antibiotic agent. The LSM plates were then allowed to dry for 15 – 30 min and incubated in anaerobic atmosphere at 37 ºC for 24 and 48 hours. Table 6. Antibiotic dilution procedure. Modified from Ericsson and Sherris, 1971. Step Concentration
(μg/ml) Source Vol.
(ml) Dist. H2O (ml)
Intermediate concentration
(μg/ml)
Final concentration
(μg/ml) 1 5120 stock - - 5120 512 2 5120 step1 2 2 2560 256 3 5120 step 1 1 3 1280 128 4 1280 step 3 2 2 640 64 5 1280 step 3 1 3 320 32 6 1280 step 3 1 7 160 16 7 160 step 6 2 2 80 8 8 160 step 6 1 3 40 4 9 160 step 6 1 7 20 2 10 20 step 9 2 2 10 1 11 20 step 9 1 3 5 0,5 12 20 step 9 1 7 2,5 0,25 13 2,5 step 12 2 2 1,25 0,125
4.5.2 Broth microdilut ion method (III-V) The bacterial cell suspension described in the agar d ilution method was diluted 1:1000 to obtain a concentration of 3 × 105 CFU/ ml. An aliquot of 100 μl of the inoculums was added to each well of tailor-made VetMIC™ ACE-ART microdilution plates (National Veterinary Institute, Uppsala, Sweden). The microdilution plates contained the following six antibiotics: tetracycline (0.5-128 μg/ ml), erythromycin (0.12-16 μg/ ml), streptomycin (2-256 μg/ ml), gentamicin (0.5-32 μg/ ml), clindamycin (0.12-8 μg/ ml) and ampicillin (0.12-8 μg/ ml) in twofold d ilutions. The 96-well plates were incubated as described in the agar dilution method, except for the Lactococcus species and L. monocytogenes (study V), which were incubated aerobically at 30 ºC or 37 ºC for 24 h, respectively.
4.5.3 Etest method (III-IV) The same bacterial cell suspension density as in agar d ilution method (3 × 108 CFU/ ml) was used in Etests (Ab Biomerieux, Solna, Sweden). A sterile cotton swab was d ipped in the inoculum and swabbed using a plate turntable or by swabbing the surface of agar in three d irections. The LSM plates with bacteria were allowed to dry, after which the Etest strip was applied . All strips had a concentration range 0.016 -256 μg/ ml. The plates were incubated as described for the agar d ilution method.

38
5. Result s and discussion
5.1 SPECIES DISTRIBUTION AMONG THE FAECAL SAMPLES OF PIGLETS AND CALVES (I-II)
From the 129 isolates of piglet origin (Study I), 67 isolates were chosen for further study based on their API-profiles and partial 16S rDNA sequencing. According to 16S rDNA sequence, the major Lactobacillus groups in piglets were Lb. reuteri (n = 45), Lb. salivarius (n = 15) and Lb. acidophilus (n = 7). In piglets, Lb. reuteri has been found as a common Lactobacillus species in colonizing intestine also in previous studies (Axelsson and Lindgren 1987, Leser 2002). However, the role of this bacterium species has remained debatable, and the clarification of its possible adhesive or probiotic properties demands further study. Interestingly, Lb. salivarius is rarely detected in the faeces of adult pigs, so this bacterium species might have an age-dependent occurrence, as also suggested in some human studies (Heilig et al. 2002). A similar d istribution of Lactobacillus species has been reported from different geographical locations (Japan, United Kingdom and Finland) and with d ifferent breeds of piglets, so these factors apparently have no major effect on the Lactobacillus species composition in piglet faecal samples (Tannock et al. 1990, Naito et al. 1995, Korhonen et al. 2007).
In faecal samples of calves (Study II), the LAB species distribution was found to be considerably larger (Figure 3). On average, the number of colony forming units (CFU) in calf faecal samples was 2.5 ± 7.6 × 106 CFU/ g. The amounts of total Lactobacillus populations have been shown to vary remarkably depending on the age of the animal (Karney et al. 1986, Agarwal et al. 2002). In addition to Lactobacillus species, the other species found in calf faecal samples were Weissella ssp. (n = 15), Streptococcus bovis/infantarius (n = 13), Enterococcus ssp. (n = 13), Lactococcus lactis ssp. cremoris (n = 2), Lactococcus garvieae (n = 1), Pediococcus pentosaceus (n = 2) and Leuconostoc mesenteroides (n = 1). A novel finding in our study with calves was the identification of Weissella ssp (n = 15), which were present in ten samples. As far as we are aware, this is the first study demonstrating this LAB genus to be present in faecal samples of calves. In addition to weissellas, another interesting finding was the identification of a single Lc. garvieae isolate, since this bacterium species has been associated with mastitis in cows (Devriese 1999), although in relatively rare cases.

39
Figure 3. The Lactobacillus species distribution of calf faecal samples.
5.2 RAPD FINGERPRINTS OF PIGLET AND CALF ISOLATES (I-II)
In piglets, the Lactobacillus isolates (n = 67) were grouped into eight major clusters (Figure 4, next page). All the isolates identified as Lb. reuteri were in four clusters and no other species were detected in these clusters. The only Lb. vaginalis isolate, which belongs to the Lb. reuteri group, d id not cluster with Lb. reuteri isolates, but with Lb. johnsonii group. Lb. reuteri and Lb. salivarius isolates were clearly d istinct from each other in the RAPD-PCR used in the piglet study.
In calves, the LAB isolates (n = 104) were also studied with RAPD-PCR, but no clear clusters could be created according to the species. However, the RAPD-PCR was found to be useful in distinguishing the bacterial strains from each other. In figures 5a-e, the LAB isolates are d ivided in to six d ifferent analyses based on their morphology, sugar fermentation pathways and 16S rDNA analysis. The strain characterization is crucial in antimicrobial susceptibility determinations; hence the use of duplicate isolates could skew the susceptibility data.
28
15
8 74
2
0
5
10
15
20
25
30
Lb. reuteri Lb. plantarum Lb. buchneri Lb. salivarius Lb. acidophilus Lb. casei

40
Figure 4. RAPD-PCR profiles of Lactobacillus group bacteria from piglets. (A) Lb. agilis, (G) Lb. gallinarum, (J) Lb. johnsonii, (M) Lb. mucosae, (R) Lb. reuteri, (S) Lb. salivarius and (V) Lb. vaginalis; (P) Lactobacillus isolate number.

41
Figure 5a. Cluster analysis of RAPD patterns of obligately heterofermentative Lactobacillus species.

42
Figure 5b. Cluster analysis of RAPD patterns of facultatively heterofermentative Lactobacillus species.
The obligately heterofermentative Lactobacillus species (Fig. 5a) were clustered according to species, with the exception of one Lb. mucosae strain. With respect to the closely related Lb. plantarum and Lb. pentosus (Fig 6b), RAPD-PCR was also shown to be d iscriminatory, although only two Lb. pentosus strains were used . To verify the d iscriminatory power for Lb. plantarum and Lb. pentosus, more strains need to be tested with this method.
Figure 5c. Cluster analysis of RAPD patterns of homofermentative Lactobacillus species.

43
Figure 5d. Cluster analysis of RAPD patterns of Weissella ssp. species.
Figure 5e. Cluster analysis of RAPD patterns of cocciform species.

44
5.3 ANTIMICROBIAL SUSCEPTIBILITY PROFILES OF LACTOBACILLUS SPECIES OF PIGLET ORIGIN (I) The d istribution of the minimum inhibitory concentrations (MICs) is shown in Figure 6a-k.
chloramphenicol
1
42
2 12
1 2 3 4 0
10 20 30 40 50
<2 2 - 4 4 - 8 8 - 16 16 - 32 32 - 64 >64 concentration (μg/ml)
f tetracycline
1 1 11
3 2 1 1 3 1 3 3 3 1 4
25
4 0
10 20 30 40 50
<2 2 - 4 4 - 8 8 - 16 16 - 32 32 - 64 >64 concentration (μg/ml)
e
erythromycin
12 2 1 1
45
6 0
10 20 30 40 50
<0.5 0.5 - 1 1 - 2 2 - 4 4 - 8 8 - 16 > 16 concentration (μg/ml)
d ciprofloxacin*
1 1
17 24
2 1 1 2 2 2 1 2 6 5 0
10 20 30 40 50
<0.5 0.5 - 1 1 - 2 2 - 4 4 - 8 8 - 16 > 16 concentration (μg/ml)
c
trimethoprim
7 12 12 3 1 1 1 1 2 5 5 12
5 0
10 20 30 40 50
<4 4 - 8 8 - 16 16 - 32 32 - 64 64 - 128 >128 concentration (μg/ml)
b vancomycin
3
45
15 4
0 10 20 30 40 50
<0.5 0.5 - 1 1 - 2 2 - 4 4 - 8 8 - 16 > 16 concentration (μg/ml)
a

45
Figure 6a-k. The distribution of MICs of Lactobacillus species according to groups. (■) Lb. reuteri group; (□) Lb. salivarius group; (■) Lb. acidophilus group; (↓) MICs for obligate homofermentative and heterofermentative Lactobacillus determined by FEEDAP (European Commission 2005); * MIC not determined by FEEDAP. All the Lb. reuteri, Lb. salivarius and Lb. agilis isolates were highly resistant to vancomycin (MIC >16 μg/ ml), while both Lb. johnsonii and Lb. gallinarum were sensitive with MIC <0.5 μg/ ml (Fig 6a). In all, 31 of the total 67 isolates were observed to be resistant to trimethoprim (Fig. 6b) according to FEEDAP (2005) breakpoint (8μg/ ml). The two trimethoprim-resistant Lb. acidophilus group bacteria were both Lb. johnsonii. With respect to ciprofloxacin (Fig 6c), MICs >16 μg/ ml were detected with Lb. reuteri, Lb. mucosae, Lb. johnsonii and Lb. gallinarum. The vast majority of the isolates were resistant to tetracycline (Fig.
gentamycin
1
42
2 10
7 5 0
10 20 30 40 50
<1 1 - 2 2 - 4 4 - 8 8 - 16 16 - 32 >32 concentration (μg/ml)
k
streptomycin
9 1 2 1
12 2
33
4 3 0
10 20 30 40 50
<4 4 - 8 8 - 16 16 - 32 32 - 64 64 - 128 >128 concentration (μg/ml)
j kanamycin
1 1 2 2 1 2 7 2 6
33
5 5 0
10 20 30 40 50
<4 4 - 8 8 - 16 16 - 32 32 - 64 64 - 128 >128 concentration (μg/ml)
i
clindamycin
42
2 1 12
1 1 1 3 1 3 0
10 20 30 40 50
<0.25 0.25 - 0.5
0.5 - 1 1 - 2 2 - 4 4 - 8 >8
concentration (μg/ml)
h ampicillin
1
27
2 8 1 1
15 3 3 3 3
0 10 20 30 40 50
<0.25 0.25 -0.5
0.5 - 1 1 - 2 2 - 4 4 - 8 >8
concentration (μg/ml)
g

46
6e), but were grouped in the vicinity of the FEEDAP (European Commission 2005) breakpoint (8 μg/ ml).
When ampicillin and gentamicin were examined (Fig 6g and 6k, respectively), all the 67 isolates were found to be susceptible. Only sporadic resistances were observed with erythromycin, clindamycin and streptomycin (Fig 6d , 6h and 6j, respectively). A very high level of resistance was typical of erythromycin and streptomycin resistant isolates. In the Lb. salivarius group, most of the isolates (n = 13) were resistant to kanamycin, with only two (P57 and P62) being susceptible (Fig 6i). The same two Lb. salivarius isolates were multiresistant, showing simultaneous resistance to vancomycin, streptomycin, clindamycin and tetracycline; P57 was additionally resistant to erythromycin and P62 to trimethoprim and chloramphenicol.
In conclusion, exceptional antibiotic resistances were rare among the isolates studied , and the vast majority of the MIC values were below the FEEDAP (European Comission 2005) breakpoints. Based on these phenotypic tests, the situation of resistance/ susceptibility of lactobacilli from piglet faecal samples is relatively good. However, in order to scientifically guarantee the absence of acquired resistance genes molecular tools should also be applied due to the d iscrepancies observed between phenotypic and genotypic test methods (Hummel et al. 2007, van Hoek et al. 2008c)
5.4 ANTIMICROBIAL SUSCEPTIBILITY PROFILES OF LAB SPECIES OF CALF ORIGIN (II)
The MICs against d ifferent antibiotics tested are shown in Tables 7a-7d. All the strains had a MIC value ≤4 μg/ ml for amoxicillin, while with penicillin and ampicillin the highest MIC observed was 8 μg/ ml. As expected, the majority of the Lactobacillus strains were highly vancomycin resistant, with the exception of all four Lb. johnsonii strains. One Lb. ruminis isolate (C216) was highly susceptible to vancomycin. All the Weissella ssp. species were vancomycin resistant. On the other hand, cocciform bacteria were sensitive, with the exception of one Str. bovis/ infantarius (C112) and the only Ln. mesenteroides (C187) isolate (Table 7a).
The highest MICs for aminoglycosides were observed with kanamycin. One strain of Str. bovis/infantarius (C118) and one strain of Enterococcus spp. (C113) were found to be highly resistant outliers from the other members of the respective groups of bacteria (MIC 128 μg/ ml for kanamycin) (Table 7b).
High resistances towards erythromycin (MIC 64 μg/ ml) and tylosin (MIC ≥ 256 μg/ ml) were observed with one strain of Lb. farciminis (C234). Only one Str. bovis/infantarius (C112, MIC 64 μg/ ml) and one Lc. garvieae (C176, MIC 128 μg/ ml) isolate were found resistant to tetracycline (Table 7c). Clindamycin resistances among the strains were rarely observed (Table 7d). The two clindamycin resistant Lb. plantarum group strains were both Lb. farciminis (C257, MIC 8 μg/ ml and C234, MIC ≥256 μg/ ml). Other strains resistant to clindamycin were P. pentosaceus (C110 and C228), Enterococcus ssp. (C223 and C225), and Lc. garvieae (C176).

47
With trimethoprim (similar to the situation in feacal samples from piglets) and lincomycin, the determinations of meaningful MICs were impractical, due to the broad range of MICs. No multiresistant (resistance to more than two antibiotics) strains were found according to the microbiological breakpoints given by FEEDAP (European Commission 2008) in LAB from calves.

48
Table 7a. Distribution of MICs of penicillins and vancomycin from calf samples. n.g., not given; E., Enterococcus; Lc., Lactococcus; Ln., Leuconostoc; P., Pediococcus. Microbiological breakpoint for 1enterococci, 2lactococci, 3leuconostoc, and 4pediococci, according to FEEDAP Panel (European Commission 2008). Strains with MICs higher than the breakpoints are considered resistant. Test agent
Bacterial group
FEEDAP
breakpoint
μg/ml
≤0.125 0.25 0.5 1 2 4 8 16 32 64 128 ≥256
Amoxicillin Lb. reuteri (n = 28) n.g. 4 10 2 6 4 2 Lb. plantarum (n = 15) n.g. 1 10 3 1 Lb. buchneri (n = 8) n.g. 2 5 1 Lb. salivarius (n = 7) n.g. 5 1 1 Lb. acidophilus (n = 4) n.g. 1 1 2 Lb. rhamnosus (n = 2) n.g. 1 1 Weissella ssp. (n = 15) n.g. 14 1 Str. bovis/infantarius (n = 13) n.g. 12 1 E., Lc., Ln., and P. (n = 12) n.g. 10 1 1 Ampicillin Lb. reuteri (n = 28) 2 8 4 4 6 5 1 Lb. plantarum (n = 15) 2 11 1 3 Lb. buchneri (n = 8) 2 5 2 1 Lb. salivarius (n = 7) 4 5 1 1 Lb. acidophilus (n = 4) 1 1 1 1 1 Lb. rhamnosus (n = 2) 4 2 Weissella ssp. (n = 15) 1 13 2 Str. bovis/infantarius (n = 13) 1 12 1 E., Lc., Ln., and P. (n = 12) 22,3, 41,4 10 1 1 Penicillin C Lb. reuteri (n = 28) n.g. 1 7 4 6 7 2 1 Lb. plantarum (n = 15) n.g. 1 7 2 4 1 Lb. buchneri (n = 8) n.g. 6 1 1 Lb. salivarius (n = 7) n.g. 2 2 2 1 Lb. acidophilus (n = 4) n.g. 3 1 Lb. rhamnosus (n = 2) n.g. 2 Weissella ssp. (n = 15) n.g. 12 1 2 Str. bovis/infantarius (n = 13) n.g. 12 1 E., Lc., Ln., and P. (n = 12) n.g. 8 3 1 Vancomycin Lb. reuteri (n = 28) n.g. 1 1 26 Lb. plantarum (n = 15) n.g. 15 Lb. buchneri (n = 8) n.g. 8 Lb. salivarius (n = 7) n.g. 1 6 Lb. acidophilus (n = 4) 2 3 1 Lb. rhamnosus (n = 2) n.g. 2 Weissella ssp. (n = 15) 2 15 Str. bovis/infantarius (n = 13) 2 9 2 1 1 E., Lc., Ln., and P. (n = 12) 41,2, n.g.3,4 1 2 6 2 1

49
Table 7b. Distribution of MICs of aminoglycosides from calf samples. n.g., not given; E., Enterococcus; Lc., Lactococcus; Ln., Leuconostoc; P., Pediococcus. Microbiological breakpoint for 1enterococci, 2lactococci, 3leuconostoc, and 4pediococci, according to FEEDAP Panel (European Commission 2008). Strains with MICs higher than the breakpoints are considered resistant. Test agent
Bacterial group
FEEDAP
breakpoint
μg/ml
≤0.125 0.25 0.5 1 2 4 8 16 32 64 128 ≥256
Gentamicin Lb. reuteri (n = 28) 8 4 12 8 3 1 Lb. plantarum (n = 15) 16 14 1 Lb. buchneri (n = 8) 16 4 4 Lb. salivarius (n = 7) 16 1 4 1 1 Lb. acidophilus (n = 4) 16 2 2 Lb. rhamnosus (n = 2) 16 1 1 Weissella ssp. (n = 15) 4 3 6 6 Str. bovis/infantarius (n = 13) 4 1 3 5 4 E., Lc., Ln., and P. (n = 12) 163,4, 321,2 1 1 2 3 4 1
Kanamycin Lb. reuteri (n = 28) 16 1 1 13 8 5 Lb. plantarum (n = 15) 64 1 1 2 9 2 Lb. buchneri (n = 8) 16 1 3 1 3 Lb. salivarius (n = 7) 64 1 3 2 1 Lb. acidophilus (n = 4) 16 1 2 1 Lb. rhamnosus (n = 2) 64 1 1 Weissella ssp. (n = 15) 16 4 3 1 2 3 2 Str. bovis/infantarius (n = 13) 16 2 4 6 1 E., Lc., Ln., and P. (n = 12) 163, 642,4, 5121 3 6 2 1
Neomycin Lb. reuteri (n = 28) n.g. 3 6 6 10 3 Lb. plantarum (n = 15) n.g. 2 12 1 Lb. buchneri (n = 8) n.g. 4 3 1 Lb. salivarius (n = 7) n.g. 1 3 2 1 Lb. acidophilus (n = 4) n.g. 2 2 Lb. rhamnosus (n = 2) n.g. 1 1 Weissella ssp. (n = 15) n.g. 1 6 3 3 2 Str. bovis/infantarius (n = 13) n.g. 1 1 1 1 9 E., Lc., Ln., and P. (n = 12) n.g. 1 1 2 2 5 1
Streptomycin Lb. reuteri (n = 28) 64 1 2 15 7 3 Lb. plantarum (n = 15) n.g. 1 8 6 Lb. buchneri (n = 8) 64 2 3 3 Lb. salivarius (n = 7) 64 1 3 2 1 Lb. acidophilus (n = 4) 16 2 2 Lb. rhamnosus (n = 2) 32 1 1 Weissella ssp. (n = 15) 8 3 1 3 8 Str. bovis/infantarius (n = 13) 8 4 3 6 E., Lc., Ln., and P. (n = 12) 642,3,4, 1281 6 4 1 1

50
Table 7c. Distribution of MICs of chloramphenicol, tetracycline and macrolides from calf samples. n.g., not given; E., Enterococcus; Lc., Lactococcus; Ln., Leuconostoc; P., Pediococcus. Microbiological breakpoint for 1enterococci, 2lactococci, 3leuconostoc, and 4pediococci, according to FEEDAP Panel (European Commission 2008). Strains with MICs higher than the breakpoints are considered resistant. Test agent
Bacterial group
FEEDA
P
μg/ml
≤0.125 0.25 0.5 1 2 4 8 16 32 64 128 ≥256
Chloramphenicol Lb. reuteri (n = 28) 4 2 19 7 Lb. plantarum (n = 15) 8 1 14 Lb. buchneri (n = 8) 4 8 Lb. salivarius (n = 7) 4 1 6 Lb. acidophilus (n = 4) 4 2 2 Lb. rhamnosus (n = 8) 4 1 1 Weissella ssp. (n = 15) 2 8 6 1 Str. bovis/infantarius (n = 13) 2 2 3 2 4 1 1 E., Lc., Ln., and P. (n = 12) 43,4, 81,2 1 3 7 1
Erythromycin Lb. reuteri (n = 28) 1 17 11 Lb. plantarum (n = 15) 1 13 1 1 Lb. buchneri (n = 8) 1 1 7 Lb. salivarius (n = 7) 1 1 5 1 Lb. acidophilus (n = 4) 1 3 1 Lb. rhamnosus (n = 2) 1 2 Weissella ssp. (n = 15) 0.5 5 9 1 Str. bovis/infantarius (n = 13) 0.5 12 1 E., Lc., Ln., and P. (n = 12) 13,4, 22, 5 3 1 1 1 1
Tetracycline Lb. reuteri (n = 28) 16 1 4 14 4 5 Lb. plantarum (n = 15) 32 2 2 11 Lb. buchneri (n = 8) 8 5 1 2 Lb. salivarius (n = 7) 8 1 1 1 3 1 Lb. acidophilus (n = 4) 4 2 1 1 Lb. rhamnosus (n = 2) 8 2 Weissella ssp. (n = 15) 2 2 8 4 1 Str. bovis/infantarius (n = 13) 2 8 4 1 E., Lc., Ln., and P. (n = 12) 21, 42, 9 1 1 1
Tylosin Lb. reuteri (n = 28) n.g. 1 9 10 8 Lb. plantarum (n = 15) n.g. 1 1 12 1 Lb. buchneri (n = 8) n.g. 2 1 5 Lb. salivarius (n = 7) n.g. 1 2 3 1 Lb. acidophilus (n = 4) n.g. 1 2 1 Lb. rhamnosus (n = 2) n.g. 1 1 Weissella ssp. (n = 15) n.g. 1 8 5 1 Str. bovis/infantarius (n = 13) n.g. 11 1 1 E., Lc., Ln., and P. (n = 12) n.g. 1 4 1 5 1

51
Table 7d. Distribution of MICs of lincosamides, quinolone and sulphonamide from calf samples. n.g., not given; E., Enterococcus; Lc., Lactococcus; Ln., Leuconostoc; P., Pediococcus. Microbiological breakpoint for 1enterococci, 2lactococci, 3leuconostoc, and 4pediococci, according to FEEDAP Panel (European Commission 2008). Strains with MICs higher than the breakpoints are considered resistant. Test agent
Bacterial group
FEEDAP
breakpoint
μg/ml
≤0.125 0.25 0.5 1 2 4 8 16 32 64 128 ≥256
Clindamycin Lb. reuteri (n = 28) 1 24 4 Lb. plantarum (n = 15) 1 6 3 3 1 1 1 Lb. buchneri (n = 8) 1 2 4 1 1 Lb. salivarius (n = 7) 1 6 1 Lb. acidophilus (n = 4) 1 3 1 Lb. rhamnosus (n = 2) 1 1 1 Weissella ssp. (n = 15) 0.25 9 4 2 Str. bovis/infantarius (n = 13) 0.25 13 E., Lc., Ln., and P. (n = 12) 13,4, 41,2 4 2 1 4 1
Enrofloxacin Lb. reuteri (n = 28) n.g. 5 10 9 4 Lb. plantarum (n = 15) n.g. 1 1 1 7 5 Lb. buchneri (n = 8) n.g. 1 7 Lb. salivarius (n = 7) n.g. 1 3 3 Lb. acidophilus (n = 4) n.g. 1 1 1 1 Lb. rhamnosus (n = 2) n.g. 1 1 Weissella ssp. (n = 15) n.g. 1 9 2 3 Str. bovis/infantarius (n = 13) n.g. 8 3 1 1 E., Lc., Ln., and P. (n = 12) n.g. 4 2 2 3 1
Lincomycin Lb. reuteri (n = 28) n.g. 13 9 1 1 4 Lb. plantarum (n = 15) n.g. 1 1 2 3 1 6 1 Lb. buchneri (n = 8) n.g. 1 1 4 2 Lb. salivarius (n = 7) n.g. 2 1 2 1 1 Lb. acidophilus (n = 4) n.g. 4 Lb. rhamnosus (n = 2) n.g. 1 1 Weissella ssp. (n = 15) n.g. 7 1 2 4 1 Str. bovis/infantarius (n = 13) n.g. 7 3 1 2 E., Lc., Ln., and P. (n = 12) n.g. 2 1 2 2 1 4
Trimethoprim Lb. reuteri (n = 28) n.g. 2 3 2 8 2 5 1 3 1 1 Lb. plantarum (n = 15) n.g. 2 4 5 1 2 1 Lb. buchneri (n = 8) n.g. 3 1 3 1 Lb. salivarius (n = 7) n.g. 1 1 2 1 1 1 Lb. acidophilus (n = 4) n.g. 1 1 2 Lb. rhamnosus (n = 2) n.g. 1 1 Weissella ssp. (n = 15) n.g. 1 1 1 4 6 1 1 Str. bovis/infantarius (n = 13) n.g. 2 1 1 8 1 E., Lc., Ln., and P. (n = 12) n.g. 5 2 1 3 1

52
5.5 ANTIMICROBIAL SUSCEPTIBILITY PROFILES OF LACTOBACILLUS RHAMNOSUS AND PROPOSED MICROBIOLOGICAL CUT-OFF VALUES (III)
The results of the susceptibility tests are shown in Table 8. For all six antibiotics, histograms of the corresp onding MIC data showed a unimodal, bell-shaped curve, with a few clearly resistant outliers (resistant strains). The only exception was tetracycline with Etest, which d isplayed a bimodal curve with intermediate resistant strains (four strains with MIC 8 -16 μg/ ml).
Four multi-resistant strains (resistant to more than one antimicrobial substance) were also detected . According to the microbiological breakpoints given by FEEDAP panel (European Commission 2008), two of the strains were resistant to clindamycin, erythromycin and streptomycin (strains L0544 and L0545 from starter culture and feed , respectively), one strain to ampicillin and tetracycline (L0543, probiotic strain) determined with all three methods, and one strain to streptomycin and tetracycline (L0905 from human vagina) as determined with agar d ilution and broth microdilution methods (intermediate resistance with Etest). Table 8. Distribution of MICs and proposed microbiological cutoff values. Test ranges are shaded in grey. Strains with MICs higher than the cutoff values are considered as resistant, the MICs of the wild type population are ≤ the indicated value. Resistant strains according to FEEDAP breakpoints (European Commission 2008) in bold. AD: agar dilution; BMD: broth microdilution. a ≥32 μg/ml.
Test agent Method
Test range (μg/ml)
Proposed cutoff
≤0.125 0.25 0.5 1 2 4 8 16 32 64 128 ≥256 (μg/ml)
Ampicillin AD 1 32 37 3 2 8 BMD 3 46 22 2 2 8 Etest 1 1 3 33 35 2 4 Clindamycin AD 32 41 2 0.25 BMD 2 1 28 42 2 1 Etest 18 32 22 1 2 1 Erythromycin AD 73 2a 0.125 BMD 59 12 2 2a 0.5 Etest 71 2 2 0.25 Gentamicin AD 2 5 56 12 8 BMD 1 23 40 8 3 16 Etest 1 8 50 16 8 Streptomycin AD 7 41 18 5 4 16 BMD 19 30 15 7 4 32 Etest 15 43 9 4 4 16 Tetracycline AD 1 47 22 3 1 1 2 BMD 2 63 5 4 1 4 Etest 44 26 3 1 1 1 The microarray screening d id not detect resistance determinants for erythromycin, streptomycin and tetracycline among the phenotypically resistant strains. It is possible that the responsible genes were missing on the microarray, although over 70 antibiotic resistance genes were represented . This highlights the need to include both phenotypic and molecular biology methods

53
in the safety evaluation of antibiotic resistances in bacteria intended for human consumption.
5.6 ANTIMICROBIAL SUSCEPTIBILITY PROFILES OF LACTOBACILLUS SPECIES AND PROPOSED MICROBIOLOGICAL CUT-OFF VALUES (IV)
The d istribution of MIC for ampicillin, tetracycline, erythromycin, clindamycin, streptomycin and gentamicin are shown in Tables 9a-f. Table 9a. Distribution of MICs for ampicillin. Fields shaded in grey indicate range of MICs tested for each antibiotic. MICs above the range are given as the concentration closest to the range. MICs below the range are given as the lowest concentration. Lb. paracasei includes one Lb. casei strain. Number of strains tested in parentheses. MDIL: Broth microdilution. a ≥32 μg/ml.
Generally, the Etest and broth microdilution methods were in good agreement in d ifferentiating the susceptible strains from their resistant counterparts. For several species-antibiotic combinations, however, MICs determined with the microdilution method were higher than those estimated with Etest, especially when Lb. reuteri, Lb. paracasei, Lb. rhamnosus and Lb. plantarum were tested with tetracycline. However, the opposite results were also observed, for example Lb. paracasei with erythromycin. Certain Lactobacillus strains have been observed to
Species (total n) Method Distribution of MICs (μg/ml) with ampicillin
≤0.12 0.25 0.5 1 2 4 8 16 32 64 128 ≥256
L. acidophilus (11) Etest 1 2 6 1
L. amylovorus (31) Etest 3 15 11 1 1
L. crispatus (7) Etest 5 2
L. delbrueckii (44) Etest 41 3
L. gallinarum (8) Etest 1 5 1 1
L. gasseri (36) Etest 4 14 18
L. helveticus (27) Etest 21 6
L. johnsonii (26) Etest 2 10 13 1
L. fermentum (21) Etest 14 7
L. reuteri (56) Etest 1 9 22 7 1 2 9 5
L. reuteri (56) MDIL 3 24 10 2 1 16a
L. paracasei (66) Etest 1 3 5 23 32 2
L. paracasei (66) MDIL 1 9 44 11 1
L. rhamnosus (75) Etest 1 1 3 33 35 2
L. rhamnosus (75) MDIL 3 46 22 2 2
L. plantarum (66) Etest 3 39 19 3 2
L. plantarum (54) MDIL 6 12 27 7 2
L. sakei (83) MDIL 6 39 35 3

54
grow better in broth than in agar, and also the d iffusion rate of the antibiotic substance in the Etest strips may vary from compound to compound. Table 9b. Distribution of MICs for tetracycline. Fields shaded in grey indicate range of MICs tested for each antibiotic. MICs above the range are given as the concentration closest to the range. MICs below the range are given as the lowest concentration. Lb. paracasei includes one Lb. casei strain. Number of strains tested in parentheses. MDIL: Broth microdilution.
Species (total n) Method Distribution of MICs (μg/ml) with tetracycline
≤0.12 0.25 0.5 1 2 4 8 16 32 64 128 ≥256
L. acidophilus (11) Etest 1 4 2 3 1
L. amylovorus (31) Etest 3 7 2 1 3 15
L. crispatus (7) Etest 1 2 3 1
L. delbrueckii (44) Etest 8 5 10 16 4 1
L. gallinarum (8) Etest 1 4 3
L. gasseri (36) Etest 1 1 10 21 3
L. helveticus (27) Etest 6 12 7 2
L. johnsonii (26) Etest 2 2 4 1 2 1 1 1 1 11
L. fermentum (56) Etest 1 38 17
L. reuteri (56) Etest 1 6 14 6 4 25
L. reuteri (56) MDIL 1 5 20 1 1 28
L. paracasei (66) Etest 15 45 2 1 1 1 1
L. paracasei (66 MDIL 43 18 1 1 1 2
L. rhamnosus (75) Etest 44 26 3 1 1
L. rhamnosus (75) MDIL 2 63 5 4 1
L. plantarum (121) Etest 1 13 44 51 8 1 3
L. plantarum (81) MDIL 54 24 1 2
L. sakei (83) MDIL 4 27 29 16 3 1 3
A wide range of high MICs w as obtained with tetracycline, which may be explained by the high variability of tetracycline resistance mechanisms conferring d iverse levels of susceptibility (reviewed in Roberts 1996). Tetracyclines have been used extensively in humans and animals for more than 50 years for clinical purposes, as prophylactics and for promoting growth in food animals. Interestingly, the closely related Lb. reuteri and Lb. fermentum displayed high and low MICs, respectively. The majority of the Lb. fermentum strains were of dairy origin, while Lb. reuteri strains were mainly animal isolates.
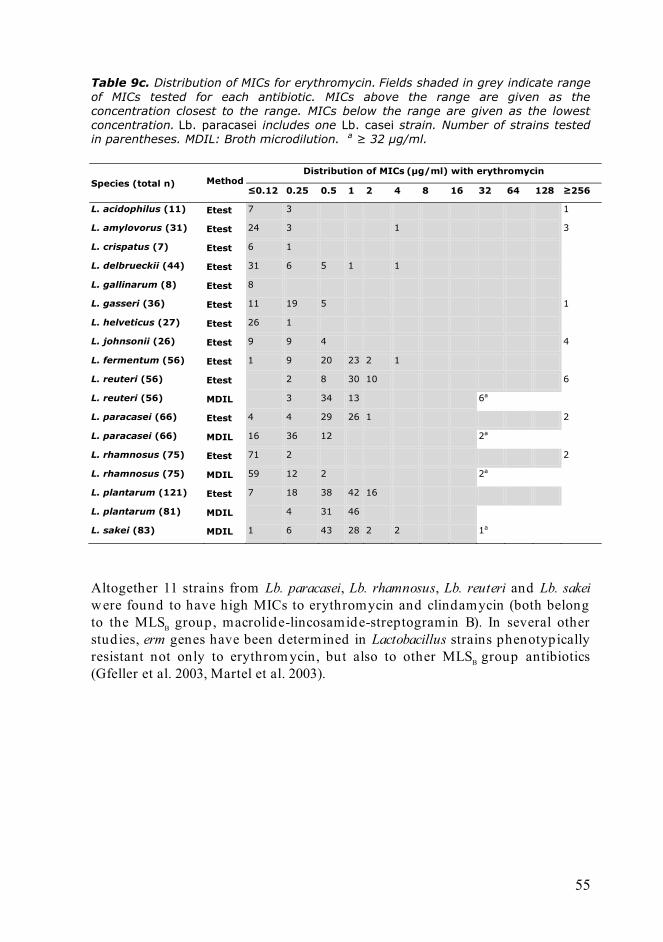
55
Table 9c. Distribution of MICs for erythromycin. Fields shaded in grey indicate range of MICs tested for each antibiotic. MICs above the range are given as the concentration closest to the range. MICs below the range are given as the lowest concentration. Lb. paracasei includes one Lb. casei strain. Number of strains tested in parentheses. MDIL: Broth microdilution. a ≥ 32 μg/ml.
Species (total n) Method Distribution of MICs (μg/ml) with erythromycin
≤0.12 0.25 0.5 1 2 4 8 16 32 64 128 ≥256
L. acidophilus (11) Etest 7 3 1
L. amylovorus (31) Etest 24 3 1 3
L. crispatus (7) Etest 6 1
L. delbrueckii (44) Etest 31 6 5 1 1
L. gallinarum (8) Etest 8
L. gasseri (36) Etest 11 19 5 1
L. helveticus (27) Etest 26 1
L. johnsonii (26) Etest 9 9 4 4
L. fermentum (56) Etest 1 9 20 23 2 1
L. reuteri (56) Etest 2 8 30 10 6
L. reuteri (56) MDIL 3 34 13 6a
L. paracasei (66) Etest 4 4 29 26 1 2
L. paracasei (66) MDIL 16 36 12 2a
L. rhamnosus (75) Etest 71 2 2
L. rhamnosus (75) MDIL 59 12 2 2a
L. plantarum (121) Etest 7 18 38 42 16
L. plantarum (81) MDIL 4 31 46
L. sakei (83) MDIL 1 6 43 28 2 2 1a
Altogether 11 strains from Lb. paracasei, Lb. rhamnosus, Lb. reuteri and Lb. sakei were found to have high MICs to erythromycin and clindamycin (both belong to the MLSB group, macrolide-lincosamide-streptogramin B). In several other studies, erm genes have been determined in Lactobacillus strains phenotypically resistant not only to erythrom ycin, but also to other MLSB group antibiotics (Gfeller et al. 2003, Martel et al. 2003).

56
Table 9d. Distribution of MICs for clindamycin. Fields shaded in grey indicate range of MICs tested for each antibiotic. MICs above the range are given as the concentration closest to the range. MICs below the range are given as the lowest concentration. Lb. paracasei includes one Lb. casei strain. Number of strains tested in parentheses. MDIL: Broth microdilution. a ≥ 16 μg/ml.
Species (total n) Method Distribution of MICsa (μg/ml) with clindamycin
≤0.12 0.25 0.5 1 2 4 8 16 32 64 128 ≥256
L. acidophilus (11) Etest 2 3 2 3 1
L. amylovorus (31) Etest 17 4 2 2 2 4
L. crispatus (7) Etest 4 2 1
L. delbrueckii (44) Etest 28 3 9 1 1
L. gallinarum (8) Etest 7 1
L. gasseri (36) Etest 1 4 5 5 7 3 11
L. helveticus (27) Etest 7 6 3 4 3 4
L. johnsonii (26) Etest 4 2 4 2 3 4 1 1 5
L. fermentum (21) Etest 21
L. reuteri (56) Etest 43 5 1 1 6
L. reuteri (56) MDIL 21 26 1 1 1 6a
L. paracasei (66) Etest 1 5 14 35 11
L. paracasei (66) MDIL 4 31 22 6 1 2a
L. rhamnosus (75) Etest 18 32 22 1 2
L. rhamnosus (75) MDIL 2 1 28 42 2
L. plantarum (73) Etest 15 15 7 13 11 4 8
L. plantarum (54) MDIL 10 3 13 9 19
L. sakei (83) MDIL 34 22 6 7 5 6 2 1a

57
Table 9e. Distribution of MICs for streptomycin. Fields shaded in grey indicate range of MICs tested for each antibiotic. MICs above the range are given as the concentration closest to the range. MICs below the range are given as the lowest concentration. Lb. paracasei includes one Lb. casei strain. Number of strains tested in parentheses. MDIL: Broth microdilution.
Species (total n) Method Distribution of MICsa (μg/ml) with streptomycin
≤0.12 0.25 0.5 1 2 4 8 16 32 64 128 ≥256
L. acidophilus (11) Etest 3 4 3 1
L. amylovorus (31) Etest 3 9 6 3 1 1 8
L. crispatus (7) Etest 1 1 4 1
L. delbrueckii (44) Etest 1 2 8 15 12 4 1 1
L. gallinarum (8) Etest 3 1 1 3
L. gasseri (36) Etest 1 1 11 18 2 3
L. helveticus (27) Etest 2 1 15 7 2
L. johnsonii (26) Etest 3 8 8 3 1 2 1
L. fermentum (56) Etest 1 9 28 16 2
L. reuteri (56) Etest 3 9 35 9
L. reuteri (56) MDIL 1 12 26 17
L. paracasei (66) Etest 1 3 10 52
L. paracasei (66) MDIL 1 10 43 12
L. rhamnosus (75) Etest 15 43 9 4 4
L. rhamnosus (75) MDIL 19 30 15 7 4
L. plantarum (121) Etest 1 2 7 20 18 27 31 6
L. plantarum (81) MDIL 3 13 24 31 6
L. sakei (83) MDIL 1 2 27 53
Since d iscrepancies were demonstrated between phenotypic and genotypic characterization in the study with Lb. rhamnosus (study III), it is recommended to include also genotypic approaches in all studies of antibiotic resistance, when their safety is being assessed .

58
Table 9f. Distribution of MICs for gentamicin. Fields shaded in grey indicate range of MICs tested for each antibiotic. MICs above the range are given as the concentration closest to the range. MICs below the range are given as the lowest concentration. Lb. paracasei includes one Lb. casei strain. Number of strains tested in parentheses. MDIL: Broth microdilution. a ≥ 64 μg/ml.
Species (total n) Method Distribution of MICs (μg/ml) with gentamicin
≤0.12 0.25 0.5 1 2 4 8 16 32 64 128 ≥256
L. acidophilus (11) Etest 1 1 4 3 1 1
L. amylovorus (31) Etest 1 4 13 12 1
L. crispatus (7) Etest 2 3 1 1
L. delbrueckii (44) Etest 5 1 9 17 12
L. gallinarum (8) Etest 2 1 4 1
L. gasseri (36) Etest 5 7 18 6
L. helveticus (27) Etest 1 4 11 9 2
L. johnsonii (26) Etest 4 11 5 6
L. fermentum (21) Etest 15 6
L. reuteri (56) Etest 1 8 35 12
L. reuteri (56) MDIL 1 12 17 26
L. paracasei (66) Etest 6 7 19 26 8
L. paracasei (66) MDIL 3 13 31 19
L. rhamnosus (75) Etest 1 8 50 16
L. rhamnosus (75) MDIL 1 23 40 8 3
L. plantarum (66) Etest 3 3 2 13 31 14
L. plantarum (54) MDIL 5 9 15 19 6
L. sakei (83) MDIL 3 3 31 42 4a
5.7 ANTAGONISTIC ACTIVITY AND TRANSFORMATION OF LACTOCOCCUS SPECIES (V)
In the mating experiments, the cell densities were severely reduced when Lc. garvieae DSMZ 6783 and L. monocytogenes were grown along with Lc. lactis ssp. lactis KYA7 –strain. This was indicative of antagonistic activity, which is widely noted among LAB (Abriouel et al. 2005). This phenomenon could have represented a methodological limitation during the conjugation experiment in vitro, so the tet(S) gene was electroporated from KYA7 –strain to Lc. garvieae DSMZ 6783. The transfer of the tet(S) gene was detected with Southern hybrid ization, showing a positive signal of approximately 30 kbp in size (Figure 7). The transfer of the plasmid was also confirmed with an increase of MIC for tetracycline from 0.25 μg/ ml before the transformation to 256 μg/ ml in the transformants.

59
Figure 7. Agarose gel electrophoresis analyses of plasmids isolated from Lc. lactis ssp. lactis KYA-7, Lc. garvieae DSM 6783 strains before and after tet(S) plasmid transformation by (a) electroporation, and (b) Southern hybridization analysis with tet(S) probe. Lanes: M, DNA Molecular Weight Marker II, DIG- labeled (Roche); 1, supercoiled DNA ladder (Promega); 2, plasmid DNA of Lc. lactis ssp. cremoris DSM 4645 as plasmid DNA size marker, 3, Lc. lactis ssp. lactis KYA-7; 4, Lc. garvieae DSM 6783 before electroporation; 5, Lc. garvieae DSM 6783 TF-a after electroporation.
5.8 TRANSFER OF THE TET(S) GENE FROM LACTOCOCCUS TO L. MONOCYTOGENES (V)
Although there are many d ifferent theoretical ways for transferring of the genes in laboratory conditions e.g. genes coding for antibiotic resistance (transformation, transduction, conjugation), only conjugation appears to be significant in vivo if one considers transfer between d ifferent bacterium species and genera (van Elsas 1992). The filter mating procedure was applied to detect the transferability of the tet(S) gene from Lactococcus to tetracycline susceptible L. monocytogenes strains (Table 10). Table 10. Conjugal transfer of tetracycline resistance from Lc. garvieae to L. monocytogenes. Donor Recipients Transfer
frequency (TC per recipient)
Tetracycline MIC (μg/ml) for tet(S) positive TCs
L. garvieae DSMZ 6783 TF-α
L. monocytogenes LMK8 7 × 10-7 64 L. monocytogenes LMK16 < 10-10 - L. monocytogenes ATCC 7644
4 × 10-7 128
The results indicate that the transfer of resistance from Lactococcus to Listeria might occur in the aquatic environment e.g. in fish especially in situations when subinhibitory levels of tetracycline are present due to use of medicated feed .
M 1 2 3 4 5 M 1 2 3 4 5
a b

60
6. Conclusions LAB SPECIES DISTRIBUTION IN PIGLETS AND CALVES
Lactobacillus species d istribution was found to be more variable in faecal samples of calves than in piglets. Our study with piglets confirmed that Lb. reuteri is the dominant lactobacilli in piglets after weaning.
The moderately high proportion of Lb. salivarius isolates in piglet faecal samples may be evidence for an age-dependent occurrence, since this species is rarely detected in faeces of adult pigs.
A novel finding was the identification of Weissella ssp. in faecal samples
from calves. Previously Weissella have been isolated from field grass, which most likely is the origin of the isolated weissellas in our samples.
Another interesting finding was the identification of Lactococcus garvieae
in faecal samples taken from calves, since this species has previously been associated with mastitis in cows.
SUITABILITY OF RAPD-PCR METHOD IN STRAIN CHARACTERIZATION
RAPD-PCR method was found to be a useful tool in separating isolates from each other. In piglet isolates, the RAPD-PCR was able to cluster Lb. reuteri from Lb. salivarius isolates.
With LAB from calves, the RAPD-PCR failed to cluster d ifferent bacterial species into separate clusters. However, the RAPD-PCR was able to d iscriminate strains from each other, which was the main target for this application.
ANTIMICROBIAL SUSCEPTIBILITY OF LAB
Most of the Lactobacillus isolates were susceptible to all of the antibiotics examined in this thesis.
All the strains isolated from piglets were susceptible to chloramphenicol, ampicillin and gentamicin. Two Lb. salivarius strains

61
were multiresistant to vancomycin, streptomycin, clindamycin and tetracycline, and furthermore the first strain was also resistant to erythromycin and the second additionally to trimethoprim and chloramphenicol.
In specimens from calves, the most often encountered resistance was
against tetracycline together with kanamycin in Lactobacillus species, but no multidrug resistances were found.
In Lb. rhamnosus, two of the 75 examined strains were phenotypically resistant to clindamycin, erythromycin and streptomycin, one strain to ampicillin and tetracycline, and one strain to streptomycin and tetracycline.
The genotypic susceptibility tests conducted with the microarray method gave contradictory results to those obtained with the phenotypic patterns of the tested species. This indicates that phenotypically susceptible strains may harbor silent antibiotic resistant genes, and their transferability between bacterial species needs to be clarified .
PROPOSED MICROBIOLOGICAL CUT-OFF VALUES
Several factors can influence the determination of minimum inhibitory concentrations, e.g. the inoculum volume, incubation time and temperature, method for cultivation (agar vs. broth applications) and not least, the growth medium. If one wants to be able to compare the results obtained in different laboratories, the microbiological cut-off values should be given according to the species (not genera) and method used , with standardized medium and test procedures.
The LSM agar and broth was found to be suitable in determining the MIC values of LAB, provid ing a sufficient growth environment for all LAB examined in this thesis.
TRANSFER OF ANTIBIOTIC RESISTANCE BETWEEN BACTERIAL SPECIES
A gene coding for tetracycline resistance tet(S) was able to be transferred from a fish pathogen Lc. garvieae to a human pathogen, L. monocytogenes, by conjugation in vitro.

62
7. References Abriouel, H ., Lucas, R., Ben Omar, N., Vald ivia, E., Maqueda, M., Martinez-
Canamero, M. and Galvez, A. (2005) Enterocin AS-48RJ: a variant of enterocin AS-48 chromosomally encoded by Enterococcus faecium RJ16 isolated from food. Syst Appl Microbiol 28, 383-397.
Agarwal, N., Kamra, D.N., Chaudhary, L.C., Agarwal, I., Sahoo, A. and Pathak, N.N. (2002) Microbial status and rumen enzyme profile of crossbred calves fed on d ifferent microbial feed additives. Lett Appl Microbiol 34, 329-336.
Alander, M., Satokari, R., Korpela, R., Saxelin, M., Vilpponen -Salmela, T., Mattila-Sandholm, T. and von Wright, A. (1999) Persistence of colonization of human colonic mucosa by a probiotic strain, Lactobacillus rhamnosus GG, after oral consumption. Appl Environ Microbiol 65, 351-354.
Altschul, S.F., Gish, W., Miller, W., Myers, E.W. and Lipman, D.J. (1990) Basic local alignment search tool. J Mol Biol 215, 403-410.
Ammor, M.S., Florez, A.B. and Mayo, B. (2007) Antibiotic resistance in non-enterococcal lactic acid bacteria and bifidobacteria. Food Microbiol 24, 559-570.
Ammor, M.S., Florez, A.B., van Hoek, A.H., de Los Reyes-Gavilan, C.G., Aarts, H .J., Margolles, A. and Mayo, B. (2008) Molecular characterization of intrinsic and acquired antibiotic resistance in lactic acid bacteria and bifidobacteria. J Mol Microbiol Biotechnol 14, 6-15.
Anonym. (1969) Report of the Joint Committee on the use of Antibiotics in Animal Husbandry and Veterinary Medicine. Cmnd 4190, Nov. 1969.
Anonym. (2004) Antibiotic Resistance. GAO-04-490 Antibiotic Use in Animals. Anonym. (2007) Methods for Antimicrobial Susceptibility Testing of Anaerobic
Bacteria. Approved Standard . CLSI document M11-A7. Aquilanti, L., Silvestri, G., Zannini, E., Osimani, A., Santarelli, S. and Clementi,
F. (2007) Phenotypic, genotypic and technological characterization of predominant lactic acid bacteria in Pecorino cheese from central Italy. J Appl Microbiol 103, 948-960.
Axelsson, L. (2004) Lactic Acid Bacteria: Classification and Physiology. In Lactic Acid Bacteria. Microbiological and Functional Aspects ed . Salminen, S., von Wright, A. and Ouwehand, A. pp. 1-66. New York: Marcel Dekker Inc.
Axelsson, L. and Lindgren, S. (1987) Characterization and DNA homology of Lactobacillus strains isolated from pig intestine. J Appl Bacteriol 62, 433-440.
Barton, M.D. (2000) Antibiotic use in animal feed and its impact on human healt. Nutr Res Rev 13, 279-299.

63
Batt, C.A. (2000) Lactococcus: Introduction. In Encyclopedia of Food Microbiology ed . Robinson, R.K., Batt, C.A. and Patel, P.D. pp. 1164-1166. London, U.K.: Academic Press.
Bernardeau, M., Vernoux, J.P., Henri-Dubernet, S. and Gueguen, M. (2008) Safety assessment of dairy microorganisms: the Lactobacillus genus. Int J Food Microbiol 126, 278-285.
Busconi, M., Reggi, S. and Fogher, C. (2008) Evaluation of biodiversity of lactic acid bacteria microbiota in the calf intestinal tracts. Antonie Van Leeuwenhoek 94, 145-155.
Canchaya, C., Claesson, M.J., Fitzgerald , G.F., van Sinderen, D. and O'Toole, P.W. (2006) Diversity of the genus Lactobacillus revealed by comparative genomics of five species. Microbiology 152, 3185-3196.
Cannon, J.P., Lee, T.A., Bolanos, J.T. and Danziger, L.H. (2005) Pathogenic relevance of Lactobacillus: a retrospective review of over 200 cases. Eur J Clin Microbiol Infect Dis 24, 31-40.
Cataloluk, O. and Gogebakan, B. (2004) Presence of drug resistance in intestinal lactobacilli of dairy and human origin in Turkey. FEMS Microbiol Lett 236, 7-12.
Chagnaud, P., Machinis, K., Coutte, L.A., Marecat, A. and Mercenier, A. (2001) Rapid PCR-based procedure to identify lactic acid bacteria: application to six common Lactobacillus species. J Microbiol Methods 44, 139-148.
Claesson, M.J., van Sinderen, D. and O'Toole, P.W. (2008) Lactobacillus phylogenomics--towards a reclassification of the genus. Int J Syst Evol Microbiol 58, 2945-2954.
Claycamp, H.G. and Hooberman, B.H. (2004) Antimicrobial resistance risk assessment in food safety. J Food Prot 67, 2063-2071.
Collins, M.D., Rodrigues, U., Ash, C., Aguirre, M., Farrow, J.A.E., Martinez-Murcia, A., Phillips, B.A., Williams, A.M. and Wallbanks, S. (1991) Phylogenetic analysis of the genus Lactobacillus and related lactic acid bacteria as determined by reverse transcriptase sequencing of 16S rRNA. FEMS Microbiol Lett 77, 5-12.
Coppola, R., Succi, M., Tremonte, P., Reale, A., Salzano, G. and Sorrentino, E. (2005) Antibiotic susceptibility of Lactobacillus rhamnosus strains isolated from Parmigiano Reggiano cheese. Lait 85, 193-204.
Coton, M., Berthier, F. and Coton, E. (2008) Rapid identification of the three major species of dairy obligate heterofermenters Lactobacillus brevis, Lactobacillus fermentum and Lactobacillus parabuchneri by species-specific duplex PCR. FEMS Microbiol Lett 284, 150-157.
Coudeyras, S., Marchandin, H ., Fajon, C. and Forestier, C. (2008) Taxonomic and strain-specific identification of the probiotic strain Lactobacillus rhamnosus 35 within the Lactobacillus casei group. Appl Environ Microbiol 74, 2679-2689.
Courtney, P.D. (2000) Lactococcus lactis Subspecies lactis and cremoris. In Encyclopedia of Food Microbiology ed . Robinson, R.K., Batt, C.A. and Patel, P.D. pp. 1166-1171. London, U.K.: Academic Press.

64
Courvalin, P. (1994) Transfer of antibiotic resistance genes between gram -positive and gram-negative bacteria. Antimicrob Agents Chemother 38, 1447-1451.
Dal Bello, F., Walter, J., Hammes, W.P. and Hertel, C. (2003) Increased complexity of the species composition of lact ic acid bacteria in human feces revealed by alternative incubation condition. Microb Ecol 45, 455-463.
Danielsen, M. (2002) Characterization of the tetracycline resistance plasmid pMD5057 from Lactobacillus plantarum 5057 reveals a composite structure. Plasmid 48, 98-103.
Danielsen, M. and Wind, A. (2003) Susceptibility of Lactobacillus spp. to antimicrobial agents. Int J Food Microbiol 82, 1-11.
Dellaglio, F. and Felis, G.E. (2005) Taxonomy of Lactobacilli and Bifidobacteria. In Probiotics and Prebiotics: Scientific Aspects. ed . Tannock, G.W. pp. 25-49. Norfolk, U.K.: Caister Academic Press.
Devirgiliis, C., Caravelli, A., Coppola, D., Barile, S. and Perozzi, G. (2008) Antibiotic resistance and microbial composition along the manufacturing process of Mozzarella d i Bufala Campana. Int J Food Microbiol 128, 378-384.
Devirgiliis, C., Coppola, D., Barile, S., Colonna, B. and Perozzi, G. (2009) Characterization of the Tn916 conjugative transposon in a food -borne strain of Lactobacillus paracasei. Appl Environ Microbiol 75, 3866-3871.
Devriese, L.A., Hommez, J., Laevens, H ., Pot, B., Vandamme, P. and Haesebrouck, F. (1999) Identification of aesculin -hydrolyzing streptococci, lactococci, aerococci and enterococci from subclinical intramammary infections in dairy cows. Vet Microbiol 70, 87-94.
Di Cagno, R., De Angelis, M., Limitone, A., Minervini, F., Carnevali, P., Corsetti, A., Gaenzle, M., Ciati, R. and Gobbetti, M. (2006) Glucan and fructan production by sourdough Weissella cibaria and Lactobacillus plantarum. J Agric Food Chem 54, 9873-9881.
Dicksved, J., Floistrup, H., Bergstrom, A., Rosenquist, M., Pershagen, G., Scheynius, A., Roos, S., Alm, J.S., Engstrand, L., Braun -Fahrlander, C., von Mutius, E. and Jansson, J.K. (2007) Molecular fingerprinting of th e fecal microbiota of children raised according to d ifferent lifestyles. Appl Environ Microbiol 73, 2284-2289.
Donohue, D.C. (2004) Safety of Novel Probiotic Bacteria. In Lactic Acid Bacteria. Microbiological and Functional Aspects ed . Salminen, S., von Wright, A. and Ouwehand, A. pp. 531-546. New York, U.S.A.: Marcel Dekker Inc.
Egervarn, M., Lindmark, H., Roos, S., Huys, G. and Lindgren, S. (2007) Effects of inoculum size and incubation time on broth microdilution susceptibility testing of lactic acid bacteria. Antimicrob Agents Chemother 51, 394-396.
Endo, A. and Okada, S. (2005) Monitoring the lactic acid bacterial d iversity during shochu fermentation by PCR-denaturing gradient gel electrophoresis. J Biosci Bioeng 99, 216-221.

65
Ericsson, H.M. and Sherris, J.C. (1971) Antibiotic sensitivity testing. Report of an international collaborative study. Acta Microbiol Scand 217, 1-90.
European Commission. (2005) Opinion of the scientific panel on additives and products or substances used in animal feed on the updating of the criteria used in the assessment of bacteria for resistance to antibiotics of human or veterinary importance. The EFSA J 223, 1-12.
European Commission. (2007) Opinion of the Scientific Committee on a request from EFSA on the introduction of a Qualified Presumption of Safety (QPS) approach for assessment of selected microorganisms referred to EFSA. The EFSA J 587, 1-16.
European Commission. (2008) Technical guidance prepared by the Panel on Additives and Products or Substances used in Animal Feed (FEEDAP) on the update of the criteria used in th e assessment of bacterial resistance to antibiotics of human or veterinary importance. The EFSA J, 1-15.
Felis, G.E. and Dellaglio, F. (2007) Taxonomy of lactobacilli and bifidobacteria. Curr Issues Intest Microbiol 8, 44-61.
Florez, A.B., Delgado, S. and Mayo, B. (2005) Antimicrobial susceptibility of lactic acid bacteria isolated from a cheese environment. Can J Microbiol 51, 51-58.
Foschino, R., Nucera, D., Volponi, G., Picozzi, C., Ortoffi, M. an d Bottero, M.T. (2008) Comparison of Lactococcus garvieae strains isolated in northern Italy from dairy products and fishes through molecular typing. J Appl Microbiol 105, 652-662.
Gatesoupe, F.J. (2008) Updating the importance of lactic acid bacteria in fish farming: natural occurrence and probiotic treatments. J Mol Microbiol Biotechnol 14, 107-114.
Gevers, D., Huys, G., Devlieghere, F., Uyttendaele, M., Debevere, J. and Swings, J. (2000) Isolation and identification of tetracycline resistant lactic acid bacteria from pre-packed sliced meat products. Syst Appl Microbiol 23, 279-284.
Gevers, D., Huys, G. and Swings, J. (2003) In vitro conjugal transfer of tetracycline resistance from Lactobacillus isolates to other Gram-positive bacteria. FEMS Microbiol Lett 225, 125-130.
Gfeller, K.Y., Roth, M., Meile, L. and Teuber, M. (2003) Sequence and genetic organization of the 19.3-kb erythromycin- and dalfopristin-resistance plasmid pLME300 from Lactobacillus fermentum ROT1. Plasmid 50, 190-201.
Gruzza, M., Langella, P., Duval-Iflah, Y. and Ducluzeau, R. (1993) Gene transfer from engineered Lactococcus lactis strains to Enterococcus faecalis in the d igestive tract of gnotobiotic mice. Microb Releases 2, 121-125.
Guarneri, T., Rossetti, L. and Giraffa, G. (2001) Rapid identification of Lactobacillus brevis using the polymerase chain reaction. Lett Appl Microbiol 33, 377-381.

66
Haakensen, M., Vickers, D.M. and Ziola, B. (2009) Susceptibility of Pediococcus isolates to antimicrobial compounds in relation to hop -resistance and beer-spoilage. BMC Microbiol 9, 190.
Hammes, W.P. and Hertel, C. (2006) The Genera Lactobacillus and Carnobacterium. In The Procaryotes. A handbook on the Biology of Bacteria. Bacteria: Firmicutes, Cyanobacteria. ed . Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.H. and Stackebrandt, E. pp. 320-403. New York: Springer.
Hammes, W.P. and Vogel, R.F. (1995) The genus Lactobacillus. In The Genera of Lactic Acid Bacteria ed . Wood, B.J.B. and Holzapfel, W.H. pp. 19-54. London, U.K.: Chapman & Hall.
Hartemink, R., Domenech, V.R. and Rombouts, F.M. (1997) LAMVAB -A new selective medium for the isolation of lactobacilli from feaces. J Microbiol Methods 29, 77-84.
Heilig, H .G., Zoetendal, E.G., Vaughan, E.E., Marteau, P., Akkermans, A.D. and de Vos, W.M. (2002) Molecular d iversity of Lactobacillus spp. and other lactic acid bacteria in the human intestine as determined by specific amplification of 16S ribosomal DNA. Appl Environ Microbiol 68, 114-123.
Holo, H. and Nes, I.F. (1989) High-Frequency Transformation, by Electroporation, of Lactococcus lactis subsp. cremoris Grown with Glycine in Osmotically Stabilized Media. Appl Environ Microbiol 55, 3119-3123.
Holzapfel, W.H., Haberer, P., Geisen, R., Björkroth, J. and Schillinger, U. (2001) Taxonomy and important features of probiotic microorganisms in food and nutrition. Am J Clin Nutr 73, 365S-73S.
Hummel, A.S., Hertel, C., Holzapfel, W.H. and Franz, C.M. (2007) Antibiotic resistances of starter and probiotic strains of lactic acid bacteria. Appl Environ Microbiol 73, 730-739.
Huys, G., D´Haene, K., Danielsen, M.M., J., Egervärn, M. and Vandamme, P. (2008) Phenotypic and molecular assessment of antimicrobial resistance in Lactobacillus paracasei strains of food origin. J Food Prot 71, 339-344.
Huys, G., D'Haene, K. and Swings, J. (2002) Influence of the culture medium on antibiotic susceptibility testing of food -associated lactic acid bacteria with the agar overlay d isc d iffusion method. Lett Appl Microbiol 34, 402-406.
Igimi, S., Ryu, C.H., Park, S.H., Sasaki, Y., Sasaki, T. and Kumagai, S. (1996) Transfer of conjugative plasmid pAM beta 1 from Lactococcus lactis to mouse intestinal bacteria. Lett Appl Microbiol 23, 31-35.
Ishibashi, N. and Yamazaki, S. (2001) Probiotics and safety. Am J Clin Nutr 73, 465S-470S.
Jacobsen, L., Wilcks, A., Hammer, K., Huys, G., Gevers, D. and Andersen, S.R. (2007) Horizontal transfer of tet(M) and erm(B) resistance plasmids from food strains of Lactobacillus plantarum to Enterococcus faecalis JH2-2 in the gastrointestinal tract of gnotobiotic rats. FEMS Microbiol Ecol 59, 158-166.

67
Jay, J.M. (2000) Fermentation and Fermented Dairy Products. In Modern Food Microbiology ed . Anonymous pp. 113-130. Gaithersburg, Maryland: Aspen Publishers, Inc.
Kandler, O. (1983) Carbohydrate metabolism in lactic acid bacteria. Antonie Van Leeuwenhoek 49, 209-224.
Kang, M.S., Chung, J., Kim, S.M., Yang, K.H. and Oh, J.S. (2006) Effect of Weissella cibaria isolates on the formation of Streptococcus mutans biofilm. Caries Res 40, 418-425.
Karney, T.L., Johnson, M.G. and Ray, B. (1986) Changes in the lactobacilli and coliform populations in the intestinal tract of calves from birth to weaning. J Sci Food Agric 73, 231-236.
Khachatourians, G.G. (1998) Agricultural use of antibiotics and the evolution and transfer of antibiotic-resistant bacteria. Can Medic Assoc J 159, 1129-1136.
Klare, I., Konstabel, C., Muller-Bertling, S., Reissbrodt, R., Huys, G., Vancanneyt, M., Swings, J., Goossens, H . and Witte, W. (2005) Evaluation of new broth media for microdilution antibiotic susceptibility testing of Lactobacilli, Pediococci, Lactococci, and Bifidobacteria. Appl Environ Microbiol 71, 8982-8986.
Klare, I., Konstabel, C., Werner, G., Huys, G., Vankerckhoven, V., Kahlmeter, G., Hildebrandt, B., Muller-Bertling, S., Witte, W. and Goossens, H. (2007) Antimicrobial susceptibilities of Lactobacillus, Pediococcus and Lactococcus human isolates and cultures intended for probiotic or nutritional use. J Antimicrob Chemother 59, 900-912.
Korhonen, J.M., Danielsen, M., Mayo, B., Egervärn, M., Axelsson, L., Huys, G. and von Wright, A. (2008) Antimicrobial susceptibility and proposed microbiological cut-off values of lactobacilli by phenotypic determination. Int J Prob Preb 3, 257-268.
Korhonen, J.M., Sclivagnotis, Y. and von Wright, A. (2007) Characterization of dominant cultivable lactobacilli and their antibiotic resistance profiles from faecal samples of weaning piglets. J Appl Microbiol 103, 2496-2503.
Kostinek, M., Specht, I., Edward , V.A., Pinto, C., Egounlety, M., Sossa, C., Mbugua, S., Dortu, C., Thonart, P., Taljaard , L., Mengu, M., Franz, C.M. and Holzapfel, W.H. (2007) Characterisation and biochemical properties of predominant lactic acid bacteria from fermenting cassava for selection as starter cultures. Int J Food Microbiol 114, 342-351.
Kushiro, A., Chervaux, C., Cools-Portier, S., Perony, A., Legrain -Raspaud, S., Obis, D., Onoue, M. and van de Moer, A. (2009) Antimicrobial susceptibility testing of lactic acid bacteria and bifidobacteria by br oth microdilution method and Etest. Int J Food Microbiol 132, 54-58.
Leclercq, R., Dutka-Malen, S., Brisson-Noel, A., Molinas, C., Derlot, E., Arthur, M., Duval, J. and Courvalin, P. (1992) Resistance of enterococci to aminoglycosides and glycopeptides. Clin Infect Dis 15, 495-501.
Lee, J., Jang, J., Kim, B., Kim, J., Jeong, G. and Han, H. (2004) Identification of Lactobacillus sakei and Lactobacillus curvatus by multiplex PCR-based restriction enzyme analysis. J Microbiol Methods 59, 1-6.

68
Leser, T.D., Amenuvor, J.Z., Jensen, T.K., Lindecrona, R.H., Boye, M. and Moller, K. (2002) Culture-independent analysis of gut bacteria: the pig gastrointestinal tract microbiota revisited . Appl Environ Microbiol 68, 673-690.
Levy, S.B. and Marshall, B. (2004) Antibacterial resistance worldwide: causes, challenges and responses. Nat Med 10, S122-9.
Licht, T.R., Struve, C., Christensen, B.B., Poulsen, R.L., Molin, S. and Krogfelt, K.A. (2003) Evidence of increased spread and establishment of plasmid RP4 in the intestine under sub-inhibitory tetracycline concentrations. FEMS Microbiol Ecol 44, 217-223.
Lin, C.F., Fung, Z.F., Wu, C.L. and Chung, T.C. (1996) Molecular characterization of a plasmid -borne (pTC82) chloramphenicol resistance determinant (cat-TC) from Lactobacillus reuteri G4. Plasmid 36, 116-124.
Makarova, K., Slesarev, A., Wolf, Y., Sorokin, A., Mirkin, B., Koonin, E., Pavlov, A., Pavlova, N., Karamychev, V., Polouchine, N., Shakhova, V., Grigoriev, I., Lou, Y., Rohksar, D., Lucas, S., Huang, K., Goodstein, D.M., Hawkins, T., Plengvidhya, V., Welker, D., Hughes, J., Goh, Y., Benson, A., Baldwin, K., Lee, J.H ., Diaz-Muniz, I., Dosti, B., Smeianov, V., Wechter, W., Barabote, R., Lorca, G., Altermann, E., Barrangou, R., Ganesan, B., Xie, Y., Rawsthorne, H., Tamir, D., Parker, C., Breid t, F., Broadbent, J., Hutkins, R., O'Sullivan, D., Steele, J., Unlu, G., Saier, M., Klaenhammer, T., Richardson, P., Kozyavkin, S., Weimer, B. and Mills, D. (2006) Comparative genomics of the lactic acid bacteria. Proc Natl Acad Sci U S A 103, 15611-15616.
Martel, A., Meulenaere, V., Devriese, L.A., Decostere, A. and Haesebrouck, F. (2003) Macrolide and lincosamide resistance in the gram -positive nasal and tonsillar flora of pigs. Microb Drug Resist 9, 293-297.
Mättö, J., van Hoek, A.H., Domig, K.J., Saarela, M., Florez, A.B., Brockmann, E., Amtmann, E., Mayo, B., Aarts, H .J. and Danielsen, M. (2007) Susceptibility of human and probiotic Bifidobacterium spp. to selected antibiotics as determined by the Etest method. Int Dairy J 17, 1123-1131.
Mitsuoka, T. (1992) The human gastrointestinal tract. In The Lactic Acid Bacteria. The Lactic Acid Bacteria in Health and Disease. ed . Wood, B.J.B. pp. 49-114. London, U.K.: Elsevier Applied Science.
Morelli, L., Sarra, P.G. and Bottazzi, V. (1988) In vivo transfer of pAM beta 1 from Lactobacillus reuteri to Enterococcus faecalis. J Appl Bacteriol 65, 371-375.
Myllyniemi, A., Pitkälä, A., Heiska, H., Kivilahti-Mäntylä, K., Kaartinen, L. and Koppinen, J. (2007) FIN-RES-Vet 2005-2006. Finnish veterinary antimicrobial resistance monitoring and consumption of antimicrobial agents. Evira publications 22, 1-56.
Naito, S., Hayashidani, H ., Kaneko, K., Ogawa, M. and Benno, Y. (1995) Development of intestinal lactobacilli in normal piglets. J Appl Bacteriol 79, 230-236.
Nam, H., Whang, K. and Lee, Y. (2007) Analysis of vaginal lactic acid producing bacteria in healthy women. J Microbiol 45, 515-520.

69
Neu, H.C. (1992) The crisis in antibiotic resistance. Science 257, 1064-1073. Nousiainen, J., Javanainen, P., Setälä, J. and von Wright, A. (2004) Lactic acid
bacteria as animal probiotics. In Lactic Acid Bacteria. Microbiological and Functional Aspects. ed . Salminen, S., von Wright, A. and Ouwehand, A. pp. 547-580. New York, U.S.A.: Marcel Dekker Inc.
Ouoba, L.I., Lei, V. and Jensen, L.B. (2008) Resistance of potential probiotic lactic acid bacteria and bifidobacteria of African and European origin to antimicrobials: determination and transferability of the resistance genes to other bacteria. Int J Food Microbiol 121, 217-224.
Patterson, A.J., Rincon, M.T., Flint, H .J. and Scott, K.P. (2007) Mosaic tetracycline resistance genes are widespread in human and animal fecal samples. Antim Agents Chemother 51, 1115-1118.
Plengvidhya, V., Breidt, F.,Jr, Lu, Z. and Fleming, H.P. (2007) DNA fingerprinting of lactic acid bacteria in sauerkraut fermentations. Appl Environ Microbiol 73, 7697-7702.
Pu, Z.Y., Dobos, M., Limsowtin, G.K. and Powell, I.B. (2002) Integrated polymerase chain reaction-based procedures for the detection and identification of species and subspecies of the Gram -positive bacterial genus Lactococcus. J Appl Microbiol 93, 353-361.
Rachman, C.N., Kabadjova, P., Prevost, H . and Dousset, X. (2003) Identification of Lactobacillus alimentarius and Lactobacillus farciminis with 16S-23S rDNA intergenic spacer region polymorphism and PCR amplification using species-specific oligonucleotide. J Appl Microbiol 95, 1207-1216.
Ravelo, C., Magarinos, B., Lopez-Romalde, S., Toranzo, A.E. and Romalde, J.L. (2003) Molecular fingerprinting of fish-pathogenic Lactococcus garvieae strains by random amplified polymorphic DNA analysis. J Clin Microbiol 41, 751-756.
Reuter, G. (2001) The Lactobacillus and Bifidobacterium microflora of the human intestine: composition and succession. Curr Issues Intest Microbiol 2, 43-53.
Ringø, E. and Gatesoupe, F. (1998) Lactic acid bacteria in fish: a review . Aquaculture 160, 177-203.
Roberts, M.C. (1996) Tetracycline resistance determinants: mechanisms of action, regulation of expression, genetic mobility, and d istribution. FEMS Microbiol Rev 19, 1-24.
Roberts, M.C. (2005) Update on acquired tetracycline resistance genes. FEMS Microbiol Lett 245, 195-203.
Rossetti, L., Carminati, D., Zago, M. and Giraffa, G. (2009) A qualified presumption of safety approach for the safety assessment of Grana Padano whey starters. Int J Food Microbiol 130, 70-73.
Salyers, A.A., Gupta, A. and Wang, Y. (2004) Human intestinal bacteria as reservoirs for antibiotic resistance genes. Trends Microbiol 12, 412-416.
Salyers, A.A. and Shoemaker, N.B. (1996) Resistance gene transfer in anaerobes: new insights, new problems. Clin Infect Dis 23 Suppl 1, S36-43.

70
Santos, E.M., Jaime, I., Rovira, J., Lyhs, U., Korkeala, H . and Bjorkroth, J. (2005) Characterization and identification of lactic acid bacteria in "morcilla de Burgos". Int J Food Microbiol 97, 285-296.
Schillinger, U., Boehringer, B., Wallbaum, S., Caroline, L., Gonfa, A., Hu ch Nee Kostinek, M., Holzapfel, W.H. and Franz, C.M. (2008) A genus-specific PCR method for d ifferentiation between Leuconostoc and Weissella and its application in identification of heterofermentative lactic acid bacteria from coffee fermentation. FEMS Microbiol Lett 286, 222-226.
Schleifer and Ludwig (1995) Phylogeny of the genus Lactobacillus and related genera. Syst Appl Microbiol 18, 461-467.
Singer, R.S., Finch, R., Wegener, H .C., Bywater, R., Walters, J. and Lipsitch, M. (2003) Antibiotic resistance--the interplay between antibiotic use in animals and human beings. Lancet Infect Dis 3, 47-51.
Stiles, M.E. and Holzapfel, W.H. (1997) Lactic acid bacteria of foods and their current taxonomy. Int J Food Microbiol 36, 1-29.
Tannock, G.W. (1987) Conjugal transfer of plasmid pAM beta 1 in Lactobacillus reuteri and between lactobacilli and Enterococcus faecalis. Appl Environ Microbiol 53, 2693-2695.
Tannock, G.W. (2004) A special fondness for lactobacilli. Appl Environ Microbiol 70, 3189-3194.
Tannock, G.W., Fuller, R. and Pedersen, K. (1990) Lactobacillus succession in the piglet d igestive tract demonstrated by plasmid profiling. Appl Environ Microbiol 56, 1310-1316.
Teale, C.J. (2002) Antimicrobial resistance and the food chain. J Appl Microbiol 92 Suppl, 85S-9S.
Temmerman, R., Huys, G. and Swings, J. (2004) Identification of lactic acid bacteria: culture-dependent and culture-independent methods. Trends Food Sci Tech 15, 348-359.
Torriani, S., Felis, G.E. and Dellaglio, F. (2001) Differentiation of Lactobacillus plantarum, L. pentosus, and L. paraplantarum by recA gene sequence analysis and multiplex PCR assay with recA gene-derived primers. Appl Environ Microbiol 67, 3450-3454.
Tynkkynen, S., Singh, K.V. and Varmanen, P. (1998) Vancomycin resistance factor of Lactobacillus rhamnosus GG in relation to enterococcal vancomycin resistance (van) genes. Int J Food Microbiol 41, 195-204.
Urso, R., Comi, G. and Cocolin, L. (2006) Ecology of lactic acid bacteria in Italian fermented sausages: isolation, iden tification and molecular characterization. Syst Appl Microbiol 29, 671-680.
van den Bogaard , A.E. and Stobberingh, E.E. (1996) Time to ban all antibiotics as animal growth-promoting agents? Lancet 348, 619.
van Elsas, J.D. (1992) Antibiotic resistance gene transfer in the environment: an overview. In Genetic Interactions Among Microorganisms in the Natural Environment. ed . Wellington, E.M. and van Elsas, J.D. pp. 17-39. Oxford , United Kingdom: Pergamon Press.
van Hoek, A.H., Mayrhofer, S., Domig, K.J., Florez, A.B., Ammor, M.S., Mayo, B. and Aarts, H .J. (2008a) Mosaic tetracycline resistance genes and their

71
flanking regions in Bifidobacterium thermophilum and Lactobacillus johnsonii. Antimicrob Agents Chemother 52, 248-252.
van Hoek, A.H.A.M. and Aarts, H .J.M. (2008b) Microarray based detection of antibiotic resistance genes in Salmonella. Food Anal Methods 1, 95-108.
van Hoek, A.H., Margolles, A., Domig, K.J., Korhonen, J., Żyska -Krzesińska, J., Bardowski, J., Danielsen. M., Huys, G., Morelli, L. and Aarts, H . (2008c) Molecular assessment of erythromycin and tetrcycline resistance genes in lactic acid bacteria and bifidobacteria and their relation to the phenotypic resistance. Int J Prob Preb 3, 271-280.
Walter, J. In Probiotics and Prebiotics. Scientific Aspects ed . Tannock, G.W. pp. 51-82: Caister Academic Press.
Walter, J., Hertel, C., Tannock, G.W., Lis, C.M., Munro, K. and Hammes, W. (2001) Detection of Lactobacillus, Pediococcus, Leuconostoc and Weissella species in human feces by using group -specific PCR primers and denaturing gradient gel electrophoresis. Appl Env Microb 67, 2578-2585.
Yin, Q. and Zheng, Q. (2005) Isolation and identification of the dominant Lactobacillus in gut and faeces of pigs using carbohydrate fermentation and 16S rDNA analysis. J Biosci Bioeng 99, 68-71.
Yun, J.H., Lee, K.B., Sung, Y.K., Kim, E.B., Lee, H.G. and Choi, Y.J. (2009) Isolation and characterization of potential probiotic lactobacilli from pig feces. J Basic Microbiol 49, 220-226.
Zhou, J.S., Pillidge, C.J., Gopal, P.K. and Gill, H .S. (2005) Antibiotic susceptibility profiles of new probiotic Lactobacillus and Bifidobacterium strains. Int J Food Microbiol 98, 211-217.

Publications of the University of Eastern FinlandDissertations in Forestry and Natural Sciences
Publications of the University of Eastern Finland
Dissertations in Forestry and Natural Sciences
isbn 978-952-61-0096-8
Jenni Korhonen
Antibiotic Resistance ofLactic Acid Bacteria
Resistance to antibiotics is a common
characteristic in the world of bacteria.
Hitherto, only little attention has
been paid to the antimicrobial
susceptibilities of beneficial lactic acid
bacteria, on the opposite to several
pathogenic bacterial species. However,
there is some concern that antibiotic
resistance in lactic acid bacteria could
be transferred to pathogenic bacterial
species, complicating the treatment of
a diesease or infection and lead to the
spread of antibiotic-resistant bacteria.
Studies were conducted to isolate and
identify lactic acid bacteria of animal
and human origins, to evaluate their
antimicrobial resistance patterns
and to propose minimum inhibitory
concentrations for Lactobacillus species.
dissertatio
ns | 0
07 | Jenn
i Ko
rh
on
en | A
ntib
iotic R
esistan
ce of La
ctic Acid
Ba
cteria
Jenni Korhonen
Antibiotic Resistance ofLactic Acid Bacteria