isotope analyses of molecular and total organic carbon from miocene sediments
TRANSCRIPT

PII S0016-7037(99)00151-9
Isotope analyses of molecular and total organic carbon from Miocene sediments
MARK PAGANI,* K ATHERINE H. FREEMAN, and MICHAEL A. ARTHUR
Department of Geosciences, The Pennsylvania State University, University Park, Pennsylvania 16872 USA
(Received August11, 1998;accepted in revised form March31, 1999)
Abstract—Carbon-isotope compositions ofn-alkanes, pristane and phytane, and total organic carbon weremeasured and compared against isotopic trends of coeval alkadienones from Miocene sediments containingvery low organic-carbon contents. Compound-specific isotope analysis ofn-alkanes and isoprenoid lipids, inconjunction with abundance distributions ofn-alkanes reveal the influence of terrestrially derived organiccarbon at all sites analyzed. In general,n-alkanes are derived from allochthonous sources with the exceptionof n-C37 from site 516, which appears genetically related to coeval alkadienones. Further, pristane and phytanefrom pelagic sites 608 and 516 apparently derive from terrestrial sources as well, although a marine origincannot be excluded.
dTOC values lack a coherent relationship to %TOC andd13C37:2. Differential alteration and mixing ofdiverse isotopic signals most likely contribute to temporal variation and spatial differences indTOC. Therefore,when working with sediments from oligotrophic settings, we do not recommenddTOC as an indicator ofphytoplanktond13C values. Copyright © 1999 Elsevier Science Ltd
1. INTRODUCTION
The total isotope effect associated with marine photoautotro-phic carbon fixation («p) is, in part, proportional to the con-centration of available CO2 (Degens et al., 1968; Wong andSackett, 1978; Rau et al., 1989, 1992; Hinga et al., 1994,Bidigare et al., 1997). This relationship provides the incentiveto interpret isotopic trends of marine sedimentary organic car-bon (dTOC) as a record of thed13C of planktonic biomass inorder to facilitate estimates of ancient surface-water [CO2(aq)]variability (Arthur et al., 1985; Rau et al., 1991; Pedersen et al.,1991; Fontugue and Calvert, 1992; Bentaleb et al., 1996;Raymo et al., 1996). However, the use ofdTOC to reconstructpaleo-CO2 is complicated by several factors. Invariably,dTOC
results from an integration of primary, secondary, and alloch-thonous (such as aerially transported terrestrial organic carbon)isotopic signals (Gagosian and Peltzer, 1986; Simo` et al., 1991;Prahl et al., 1989; Ohkouchi et al., 1997; Villanueva et al.,1997). Therefore, the potential exists that thedTOC values ofsediments derived from low productivity waters (Prahl et al.,1989) or shelf settings (Prahl et al., 1994) do not reflect theisotopic character of in situ primary production. Further, car-bon-isotope compositions of individual photosynthesizers aregreatly influenced by a variety of other factors, such as thecarbon substrate fixed, mechanisms of carbon transport, thespecific carboxylase enzyme mediating fixation (e.g., Goerickeet al., 1994), growth rate (Laws et al., 1995; Bidigare et al.,1997) and cell geometry (Rau, 1996; Popp et al., 1998). Thesecontrols can provide a strong ecological influence on the iso-topic character of bulk sedimentary organic carbon. Therefore,given our current understanding of the processes affectingisotopic compositions among marine algae and bulk sedimen-tary organic carbon (Corg), appraisal of ancient surface-waterCO2 is most refined through application of compound-specific
isotope analysis of molecular markers derived from phyto-plankton that utilize diffusively transported CO2(aq) as thesubstrate for photosynthesis (Bidigare et al., 1997).
Alkadienones (heptatriaconta-15E, 22E-dien-2-one), bio-markers exclusive to some haptophyte algae (Marlowe et al.,1984; Conte et al., 1994), are demonstrably effective recordersof surface-water chemistry in modern waters (Bidigare et al.,1997). Moreover,d13C trends of alkadienones in Pleistoceneand Holocene sediments (Jasper and Hayes, 1990; Jasper et al.,1994) demonstrate their effectiveness in recording ancient sur-face-water chemistry and water-column stability. For example,Jasper et al. (1994) evaluated variations in surface-water CO2
in the eastern equatorial Pacific across Pleistocene glacial/interglacial intervals using«p trends derived from isotopicrecords of coeval alkadienones and surface-water foraminifera.They concluded that higher rates of upwelling during glacialepisodes effectively increased the atmosphere-water CO2 dis-equilibrium, promoting a higher CO2 flux to the atmosphere.We applied a similar approach to Miocene-aged sediments inan effort to characterize global trends in atmospheric CO2 inrelation to climate change (Pagani et al., 1999). This studyutilized sediments from oligotrophic regions in order to mini-mize the importance of growth rate variability on the isotopiccharacter of haptophyte biomass, thereby allowing [CO2(aq)] todominate records of«p. Though the occurrence of alkenones iswell-documented throughout sedimentary records of the Ceno-zoic (e.g., Brassell, 1993), low-productivity regions inevitablyyield low total organic and biomarker concentrations which canlimit isotope measurements of some molecular markers. There-fore, if low-productivity sites prove most effective in the anal-ysis of paleo-[CO2], it would be advantageous for future studiesif isotopic compositions of bulk Corg and other generalist bi-omarkers are shown to track effectively the isotopic characterof coeval alkadienones.
This study evaluates some of the organic characteristics ofMiocene-aged sediments from oligotrophic regions. Specifi-cally, the relative abundance ofn-alkanes and alkadienones,
* Author to whom correspondence should be addressed.† Present address:Earth Science Department, University of California,Santa Cruz, California 95064 USA.
Pergamon
Geochimica et Cosmochimica Acta, Vol. 64, No. 1, pp. 37–49, 2000Copyright © 1999 Elsevier Science LtdPrinted in the USA. All rights reserved
0016-7037/00 $20.001 .00
37

and isotopic compositions of bulk Corg, n-alkanes, pristane,phytane, and alkadienones are compared and assessed in termsof source and environment.
2. SITE DESCRIPTIONS
Samples for this study were used in previous research toreconstruct MiocenepCO2 trends using the isotopic composi-tions of alkadienones and shallow-dwelling planktonic forami-nifera (Pagani, 1998; Pagani et al., 1999). Site selection (Fig. 1)was determined from evidence for continuous sedimentationand inferred low-productivity surface waters characterized bylow and steady sedimentation rates, low total organic carbon,and calcareous lithologies.
Deep Sea Drilling Project (DSDP) site 608 (42°50.2059N;23°05.2529W) is located on the eastern side of the NorthAtlantic Ridge on the southern flank of the King’s Troughtectonic complex. Positioned in the subtropical North AtlanticGyre, pelagic sedimentation prevailed, accounting for primarilynannofossil-foraminifera chalk deposition during the Miocene.
DSDP site 588 (26°06.79S; 161°13.69E) was drilled on theLord Howe Rise in the southwest Pacific Ocean. Its warmsubtropical locality provides a record of low, steady sedimen-tation and upper water-column stability through the studiedtime interval (Flower and Kennett, 1993).
DSDP site 516 (30°16.599S, 35°17.109W) is located in thewestern South Atlantic Ocean on the Rio Grande Rise, effec-tively isolating this site from terrigenous sediment transport.Sediments are primarily composed of nannofossil-foraminiferaland nannofossil oozes.
Ocean Drilling Program (ODP) site 730 (17°43.885°N;57°41.519°E) is positioned in the northwest Arabian Sea on theOman margin. Currently this region experiences intense sea-sonal upwelling in response to the Indian Monsoonal system.However, middle Miocene-aged sediments used for this studypredate the onset of upwelling (Burckle, 1989; Nigrini, 1991;Kroon et al., 1991). Sediments are primarily foraminifer-bear-ing nannofossil chalk containing low Corg (Pagani, 1998) andlow to no siliceous microfossil content (Shipboard Scientific
Party, 1991). Apparent sedimentation rates ranged between 2.5to 8.5 cm/1000 yrs. These rates are 2 to 4 times higher thanthose found at sites 588, 608, and 516, suggesting higher ratesof productivity prevailed in the northern Arabian Sea during themiddle Miocene. Though growth rates were probably amplifiedon the Oman margin relative to sites 588 and 608, highersedimentation rates are due, in part, to foraminifer-bearingturbidites and higher siliciclastic input (Shipboard ScientificParty, 1991).
3. ANALYTICAL METHODS
3.1. Sediment Extraction
Samples from DSDP/ODP were dried and weighed. Sediments werehydrated with distilled, deionized, and dichloromethane/hexane-ex-tracted H2O (5–10 mL), and then subjected to Soxhlet extraction in a2:1 azeotrope of dichloromethane and distilled methanol for 24 hr. A5% NaCl solution was added to the total solvent extract to form twophases, and the neutral lipid fraction in the dichloromethane phase wasisolated by separatory funnel. The remaining aqueous phase waswashed three more times with dichloromethane and resulting neutrallipid fractions were collected and combined. The resulting total lipidextract was separated into compound classes by silica-gel columnchromatography (Pagani, 1998).
Cyclic and branched alkanes were separated from normal and isoal-kanes by adduction with urea (Michalczyk, 1985). The hydrocarbonfraction was dried under a stream of N2 and subsequently dissolved in200 mL each of methanol saturated with urea, pentane, and acetone.Samples were refrigerated for 30 min and then evaporated under N2.The resulting urea crystals were extracted with hexane, yielding cyclic/branched alkanes. Remaining urea crystals were dissolved in 500mL ofdichloromethane/hexane-extracted H2O and extracted with hexane toyield then-alkane fraction.
Compound abundances were determined using a Hewlett Packard5890 Series II gas chromatograph fitted with an on-column injector anda fused silica, DB-1 phase column (60 m3 0.32 mm I.D., 0.25mm filmthickness). Heptatriaconta-15E, 22E-dien-2-ones were identified byGC/MS, and subsequently identified by comparison of elution times.Temperature program was 60–200°C @ 20°C/min, then 200–320°C @5°C/min, (isothermal for 35 min). Pristane, phytane, andn-alkaneswere identified through comparison of elution times with known stan-dards. Temperature program utilized was 60–200°C @ 10°C/min, then200–320°C @ 4°C/min, (isothermal for 30 min).
Fig. 1. Site location map.
38 M. Pagani, K. H. Freeman, and M. A. Arthur

3.2. Total Organic Carbon
Sediments were ground by mortar and pestle, weighed, acidified in10% HCl, and sonicated for 1 h. Carbonate-free residue was filteredthrough a Whatman quartz microfibre filter with a 0.6mm pore size.Filters were dried and combusted under vacuum with CuO at 850°C for12 h. Following combustion, samples were purified and expanded intoa known volume and pressure was recorded, providing calculation ofthe percent of total organic carbon (%TOC). %TOC are reported basedon total sediment weight. Sample weights were not corrected forpotential salt residues. Prior to acidification ground samples were splitand entire analysis was performed in duplicate, though limited samplesize for many allowed for only a single analysis. Duplicate analyses of%TOC provided an average precision of 0.004%. It should be notedthat acidification and filtering will result in the loss of some dissolvedorganic matter. This loss was not quantified for this study.
3.3. Isotope Analyses
Compound-specific isotope analyses were performed using a GC-combustion system connected to a Finnigan MAT 252 mass spectrom-eter (Merritt et al., 1995). Compounds eluting from a gas chromato-graph are combusted over nickel and platinum at 1000°C. Water isremoved through a refrigerated Nafion permeable membrane (Leckroneand Hayes, 1997), externally flushed with helium, and the resultingpurified CO2 is introduced into the mass spectrometer. Isotopic com-positions are calculated relative to the Peedee belemnite (PDB) bycomparison with NBS-19 (Ricci et al., 1994):d13C 5 [(RSA/RST) 2 1]3 103; where R represents the13C/12C abundance ratio, and subscriptsSA and ST represent the sample and PDB standard, respectively. Inmost cases, isotopic values reported represent the average of two ormore measurements. Based on co-injected standards, accuracy in iso-tope determinations forn-alkanes and isoprenoids was60.54‰ and60.61‰, respectively. Uncertainties for TOC isotope analyses, deter-mined by measurements of standards with similarly low TOC contents,was60.05‰.
4. STRATIGRAPHY AND CORRELATIONS
Age models used for comparisons were developed by lin-early interpolating between magnetostratigraphic, stable-isoto-pic and biostratigraphic datums, calibrated to the most recentGeomagnetic Polarity Time Scale (Cande and Kent, 1992;Berggren et al., 1995). Age model parameters for sites 588,
608, and 730 were previously published (Pagani et al., 1999).Datums for site 516 are shown in Table 1.
Age control for site 730 is less constrained compared to sites588, 608, and 516. In an effort to improve age assignments,87Sr/86Sr compositions of well-preserved planktonic foraminif-era were measured and age estimates (Pagani et al., 1999) werederived through comparison with strontium-isotope-age curvesconstructed from sites 289 and 588 (Hodell and Woodruff,1994).
5. RESULTS
5.1. n-Alkane Distributions
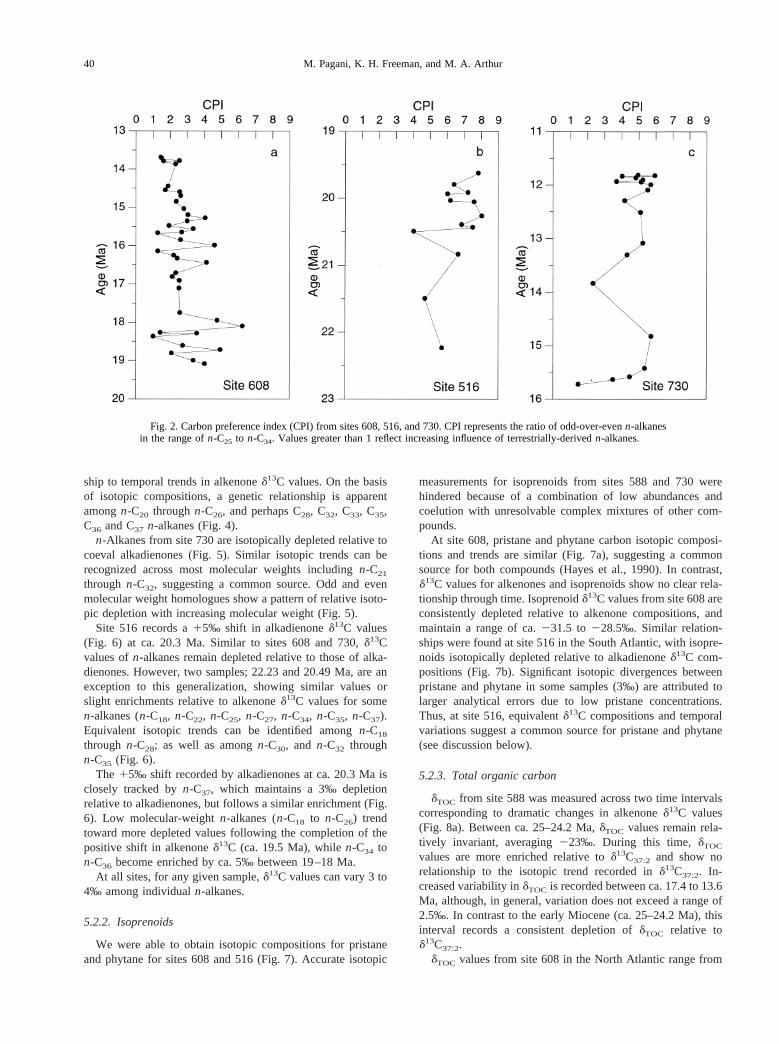
n-Alkanes were recovered from sites 608, 516, and 730, andwere absent at site 588. In general, high-molecular weightn-alkane homologues (n-C25 to n-C33) from all sites are char-acterized by odd-over-even carbon-number predominance, withn-C31 consistently representing the highest relative concentra-tion (Fig. 2).
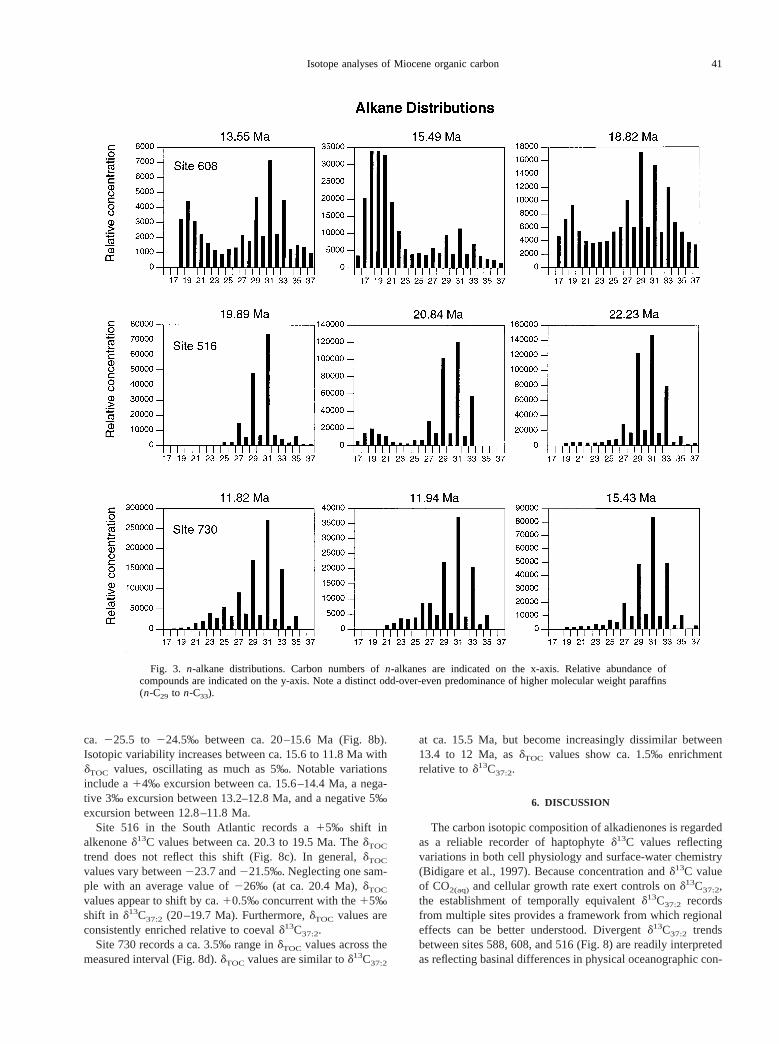
Bimodal paraffin distributions characterize most samplesfrom site 608, and some from site 516, with maxima in bothshort-chain (n-C18, n-C19, n-C20) and long-chain alkanes (n-C29, n-C31, n-C33) (Fig. 3). In general,n-C19 appears in thehighest relative concentration within the short-chain homo-logues; however, in some samplesn-C18 or n-C20 dominate.
5.2. Isotope Compositions
5.2.1. n-Alkanes
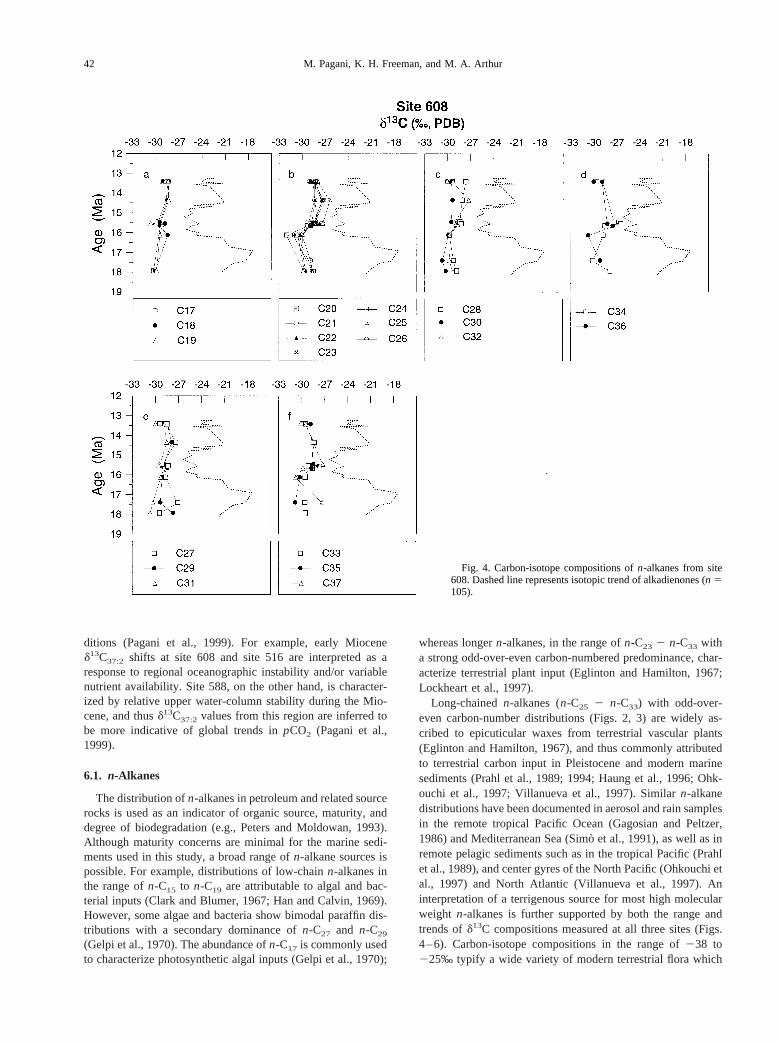
Carbon isotopic compositions ofn-alkanes and alkadienonesare displayed in Figures 4–6. At site 608, large variations inalkenoned13C values (ca. 10‰) occur throughout the studiedinterval (Fig. 4). However, individualn-alkanes lack similarisotopic variations and trends. Instead,d13C values ofn-al-kanes are consistently depleted relative to alkenones, maintain-ing values in the range of227 to 232‰. Temporal variationfor individual alkanes do not exceed 4‰ and show no relation-
Table 1. Age model parameters.
Site/Hole Datum Depth (m) Age (Ma) Ref*
516 LAD Globoquadrina dehiscens 53.5 5.8 1,2516 LAD Sphenolithus heteromorphus 71.1 13.6 1,2516 LAD Praeorbulina sicana 72.2 14.8 1,2516 FAD Praeorbulina glomerosa 88.3 16.1 1,2516 FAD P. sicana 93.1 16.4 1,2516 FAD Globorotalia birnageae 103 16.7 1,2516 LAD Globorotalia semivera 109.6 17.3 1,2516 LAD G. dehiscens(forma spinosa) 129 17.9 1,2516 FAD Globogerinoides altiapertura 170 20.5 1,2516 LAD Globorotalia kulgeri 178 21.5 1,2516F FADG. dehiscens(forma spinosa) 180 22.2 1,2516F FADDiscoaster druggi 198 23.2 1,2516F FADGloborotalia kugleri 208.2 23.8 1,2516F FADGlobogerinoides primordius(common) 215 24.3 1,2516F LAD Sphenolithus ciperoensis 246 24.75 1,2
FAD 5 First appearance datum.LAD 5 Last appearance datum.* The first number of the reference column refers to the identification of the datum level, the second number refers to the age assignment: (1)
Berggren et al., 1983; (2) Berggren et al., 1985.
39Isotope analyses of Miocene organic carbon
ship to temporal trends in alkenoned13C values. On the basisof isotopic compositions, a genetic relationship is apparentamongn-C20 throughn-C26, and perhaps C28, C32, C33, C35,C36 and C37 n-alkanes (Fig. 4).
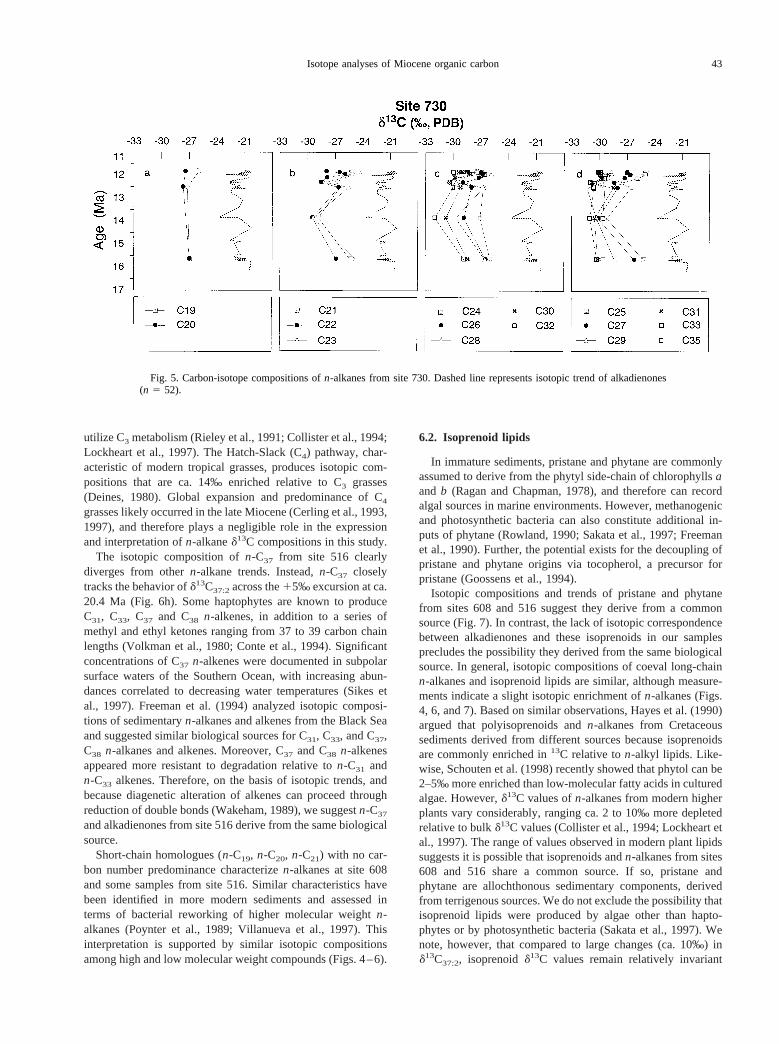
n-Alkanes from site 730 are isotopically depleted relative tocoeval alkadienones (Fig. 5). Similar isotopic trends can berecognized across most molecular weights includingn-C21
through n-C32, suggesting a common source. Odd and evenmolecular weight homologues show a pattern of relative isoto-pic depletion with increasing molecular weight (Fig. 5).
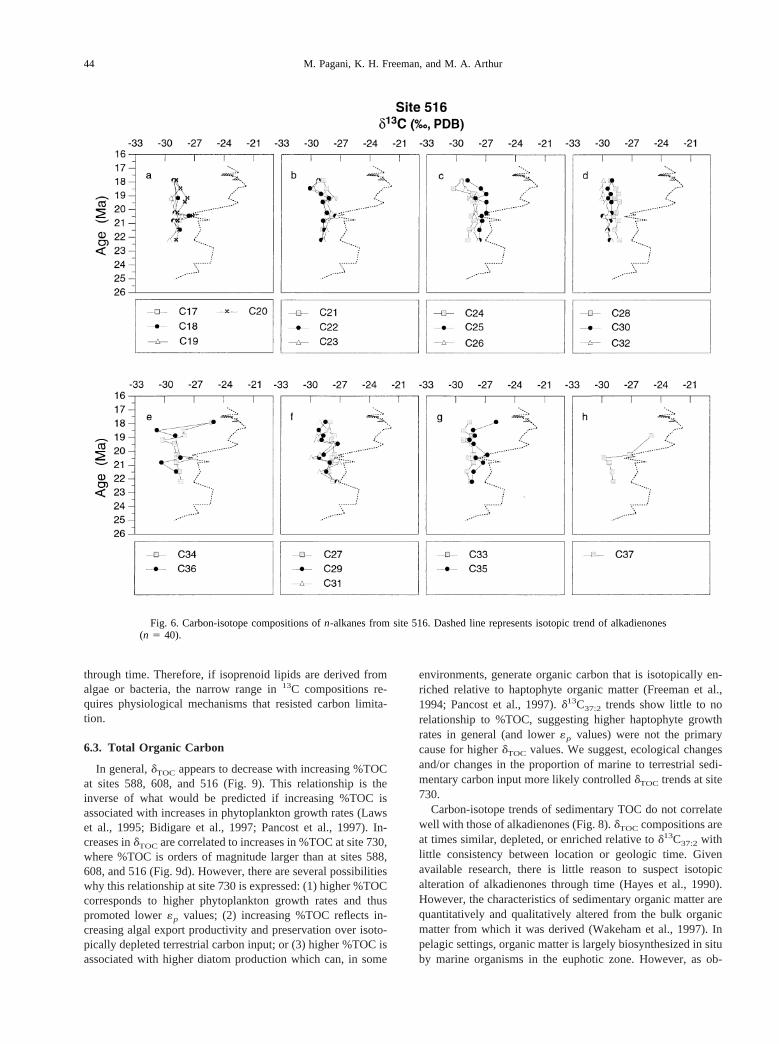
Site 516 records a15‰ shift in alkadienoned13C values(Fig. 6) at ca. 20.3 Ma. Similar to sites 608 and 730,d13Cvalues ofn-alkanes remain depleted relative to those of alka-dienones. However, two samples; 22.23 and 20.49 Ma, are anexception to this generalization, showing similar values orslight enrichments relative to alkenoned13C values for somen-alkanes (n-C18, n-C22, n-C25, n-C27, n-C34, n-C35, n-C37).Equivalent isotopic trends can be identified amongn-C18
through n-C28; as well as amongn-C30, and n-C32 throughn-C35 (Fig. 6).
The 15‰ shift recorded by alkadienones at ca. 20.3 Ma isclosely tracked byn-C37, which maintains a 3‰ depletionrelative to alkadienones, but follows a similar enrichment (Fig.6). Low molecular-weightn-alkanes (n-C18 to n-C26) trendtoward more depleted values following the completion of thepositive shift in alkenoned13C (ca. 19.5 Ma), whilen-C34 ton-C36 become enriched by ca. 5‰ between 19–18 Ma.
At all sites, for any given sample,d13C values can vary 3 to4‰ among individualn-alkanes.
5.2.2. Isoprenoids
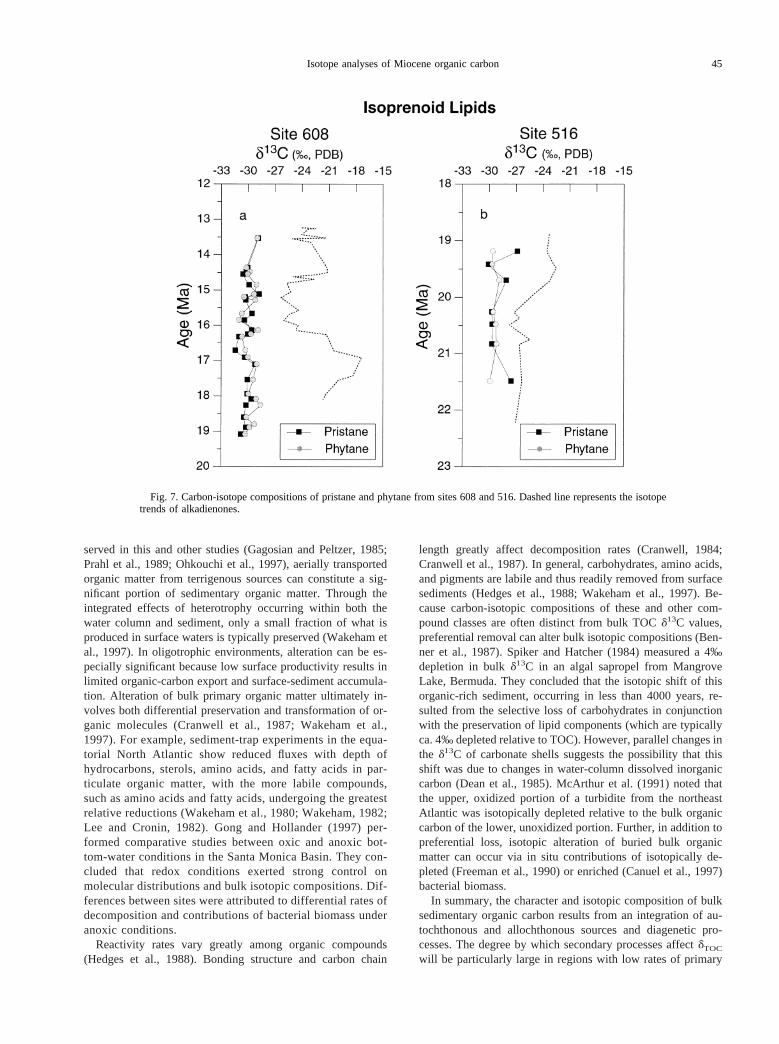
We were able to obtain isotopic compositions for pristaneand phytane for sites 608 and 516 (Fig. 7). Accurate isotopic
measurements for isoprenoids from sites 588 and 730 werehindered because of a combination of low abundances andcoelution with unresolvable complex mixtures of other com-pounds.
At site 608, pristane and phytane carbon isotopic composi-tions and trends are similar (Fig. 7a), suggesting a commonsource for both compounds (Hayes et al., 1990). In contrast,d13C values for alkenones and isoprenoids show no clear rela-tionship through time. Isoprenoidd13C values from site 608 areconsistently depleted relative to alkenone compositions, andmaintain a range of ca.231.5 to 228.5‰. Similar relation-ships were found at site 516 in the South Atlantic, with isopre-noids isotopically depleted relative to alkadienoned13C com-positions (Fig. 7b). Significant isotopic divergences betweenpristane and phytane in some samples (3‰) are attributed tolarger analytical errors due to low pristane concentrations.Thus, at site 516, equivalentd13C compositions and temporalvariations suggest a common source for pristane and phytane(see discussion below).
5.2.3. Total organic carbon
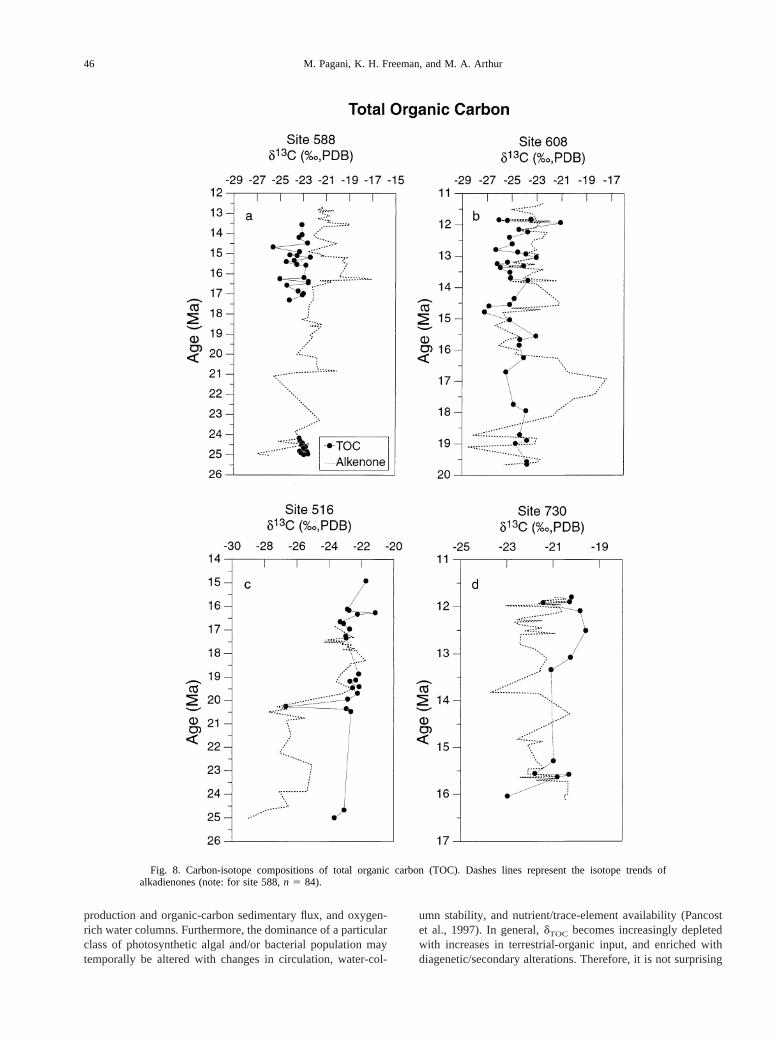
dTOC from site 588 was measured across two time intervalscorresponding to dramatic changes in alkenoned13C values(Fig. 8a). Between ca. 25–24.2 Ma,dTOC values remain rela-tively invariant, averaging223‰. During this time,dTOC
values are more enriched relative tod13C37:2 and show norelationship to the isotopic trend recorded ind13C37:2. In-creased variability indTOC is recorded between ca. 17.4 to 13.6Ma, although, in general, variation does not exceed a range of2.5‰. In contrast to the early Miocene (ca. 25–24.2 Ma), thisinterval records a consistent depletion ofdTOC relative tod13C37:2.
dTOC values from site 608 in the North Atlantic range from
Fig. 2. Carbon preference index (CPI) from sites 608, 516, and 730. CPI represents the ratio of odd-over-evenn-alkanesin the range ofn-C25 to n-C34. Values greater than 1 reflect increasing influence of terrestrially-derivedn-alkanes.
40 M. Pagani, K. H. Freeman, and M. A. Arthur
ca. 225.5 to 224.5‰ between ca. 20–15.6 Ma (Fig. 8b).Isotopic variability increases between ca. 15.6 to 11.8 Ma withdTOC values, oscillating as much as 5‰. Notable variationsinclude a14‰ excursion between ca. 15.6–14.4 Ma, a nega-tive 3‰ excursion between 13.2–12.8 Ma, and a negative 5‰excursion between 12.8–11.8 Ma.
Site 516 in the South Atlantic records a15‰ shift inalkenoned13C values between ca. 20.3 to 19.5 Ma. ThedTOC
trend does not reflect this shift (Fig. 8c). In general,dTOC
values vary between223.7 and221.5‰. Neglecting one sam-ple with an average value of226‰ (at ca. 20.4 Ma),dTOC
values appear to shift by ca.10.5‰ concurrent with the15‰shift in d13C37:2 (20–19.7 Ma). Furthermore,dTOC values areconsistently enriched relative to coevald13C37:2.
Site 730 records a ca. 3.5‰ range indTOC values across themeasured interval (Fig. 8d).dTOC values are similar tod13C37:2
at ca. 15.5 Ma, but become increasingly dissimilar between13.4 to 12 Ma, asdTOC values show ca. 1.5‰ enrichmentrelative tod13C37:2.
6. DISCUSSION
The carbon isotopic composition of alkadienones is regardedas a reliable recorder of haptophyted13C values reflectingvariations in both cell physiology and surface-water chemistry(Bidigare et al., 1997). Because concentration andd13C valueof CO2(aq) and cellular growth rate exert controls ond13C37:2,the establishment of temporally equivalentd13C37:2 recordsfrom multiple sites provides a framework from which regionaleffects can be better understood. Divergentd13C37:2 trendsbetween sites 588, 608, and 516 (Fig. 8) are readily interpretedas reflecting basinal differences in physical oceanographic con-
Fig. 3. n-alkane distributions. Carbon numbers ofn-alkanes are indicated on the x-axis. Relative abundance ofcompounds are indicated on the y-axis. Note a distinct odd-over-even predominance of higher molecular weight paraffins(n-C29 to n-C33).
41Isotope analyses of Miocene organic carbon
ditions (Pagani et al., 1999). For example, early Miocened13C37:2 shifts at site 608 and site 516 are interpreted as aresponse to regional oceanographic instability and/or variablenutrient availability. Site 588, on the other hand, is character-ized by relative upper water-column stability during the Mio-cene, and thusd13C37:2 values from this region are inferred tobe more indicative of global trends inpCO2 (Pagani et al.,1999).
6.1. n-Alkanes
The distribution ofn-alkanes in petroleum and related sourcerocks is used as an indicator of organic source, maturity, anddegree of biodegradation (e.g., Peters and Moldowan, 1993).Although maturity concerns are minimal for the marine sedi-ments used in this study, a broad range ofn-alkane sources ispossible. For example, distributions of low-chainn-alkanes inthe range ofn-C15 to n-C19 are attributable to algal and bac-terial inputs (Clark and Blumer, 1967; Han and Calvin, 1969).However, some algae and bacteria show bimodal paraffin dis-tributions with a secondary dominance ofn-C27 and n-C29
(Gelpi et al., 1970). The abundance ofn-C17 is commonly usedto characterize photosynthetic algal inputs (Gelpi et al., 1970);
whereas longern-alkanes, in the range ofn-C23 2 n-C33 witha strong odd-over-even carbon-numbered predominance, char-acterize terrestrial plant input (Eglinton and Hamilton, 1967;Lockheart et al., 1997).
Long-chainedn-alkanes (n-C25 2 n-C33) with odd-over-even carbon-number distributions (Figs. 2, 3) are widely as-cribed to epicuticular waxes from terrestrial vascular plants(Eglinton and Hamilton, 1967), and thus commonly attributedto terrestrial carbon input in Pleistocene and modern marinesediments (Prahl et al., 1989; 1994; Haung et al., 1996; Ohk-ouchi et al., 1997; Villanueva et al., 1997). Similarn-alkanedistributions have been documented in aerosol and rain samplesin the remote tropical Pacific Ocean (Gagosian and Peltzer,1986) and Mediterranean Sea (Simo` et al., 1991), as well as inremote pelagic sediments such as in the tropical Pacific (Prahlet al., 1989), and center gyres of the North Pacific (Ohkouchi etal., 1997) and North Atlantic (Villanueva et al., 1997). Aninterpretation of a terrigenous source for most high molecularweight n-alkanes is further supported by both the range andtrends ofd13C compositions measured at all three sites (Figs.4–6). Carbon-isotope compositions in the range of238 to225‰ typify a wide variety of modern terrestrial flora which
Fig. 4. Carbon-isotope compositions ofn-alkanes from site608. Dashed line represents isotopic trend of alkadienones (n 5105).
42 M. Pagani, K. H. Freeman, and M. A. Arthur
utilize C3 metabolism (Rieley et al., 1991; Collister et al., 1994;Lockheart et al., 1997). The Hatch-Slack (C4) pathway, char-acteristic of modern tropical grasses, produces isotopic com-positions that are ca. 14‰ enriched relative to C3 grasses(Deines, 1980). Global expansion and predominance of C4
grasses likely occurred in the late Miocene (Cerling et al., 1993,1997), and therefore plays a negligible role in the expressionand interpretation ofn-alkaned13C compositions in this study.
The isotopic composition ofn-C37 from site 516 clearlydiverges from othern-alkane trends. Instead,n-C37 closelytracks the behavior ofd13C37:2across the15‰ excursion at ca.20.4 Ma (Fig. 6h). Some haptophytes are known to produceC31, C33, C37 and C38 n-alkenes, in addition to a series ofmethyl and ethyl ketones ranging from 37 to 39 carbon chainlengths (Volkman et al., 1980; Conte et al., 1994). Significantconcentrations of C37 n-alkenes were documented in subpolarsurface waters of the Southern Ocean, with increasing abun-dances correlated to decreasing water temperatures (Sikes etal., 1997). Freeman et al. (1994) analyzed isotopic composi-tions of sedimentaryn-alkanes and alkenes from the Black Seaand suggested similar biological sources for C31, C33, and C37,C38 n-alkanes and alkenes. Moreover, C37 and C38 n-alkenesappeared more resistant to degradation relative ton-C31 andn-C33 alkenes. Therefore, on the basis of isotopic trends, andbecause diagenetic alteration of alkenes can proceed throughreduction of double bonds (Wakeham, 1989), we suggestn-C37
and alkadienones from site 516 derive from the same biologicalsource.
Short-chain homologues (n-C19, n-C20, n-C21) with no car-bon number predominance characterizen-alkanes at site 608and some samples from site 516. Similar characteristics havebeen identified in more modern sediments and assessed interms of bacterial reworking of higher molecular weightn-alkanes (Poynter et al., 1989; Villanueva et al., 1997). Thisinterpretation is supported by similar isotopic compositionsamong high and low molecular weight compounds (Figs. 4–6).
6.2. Isoprenoid lipids
In immature sediments, pristane and phytane are commonlyassumed to derive from the phytyl side-chain of chlorophyllsaand b (Ragan and Chapman, 1978), and therefore can recordalgal sources in marine environments. However, methanogenicand photosynthetic bacteria can also constitute additional in-puts of phytane (Rowland, 1990; Sakata et al., 1997; Freemanet al., 1990). Further, the potential exists for the decoupling ofpristane and phytane origins via tocopherol, a precursor forpristane (Goossens et al., 1994).
Isotopic compositions and trends of pristane and phytanefrom sites 608 and 516 suggest they derive from a commonsource (Fig. 7). In contrast, the lack of isotopic correspondencebetween alkadienones and these isoprenoids in our samplesprecludes the possibility they derived from the same biologicalsource. In general, isotopic compositions of coeval long-chainn-alkanes and isoprenoid lipids are similar, although measure-ments indicate a slight isotopic enrichment ofn-alkanes (Figs.4, 6, and 7). Based on similar observations, Hayes et al. (1990)argued that polyisoprenoids andn-alkanes from Cretaceoussediments derived from different sources because isoprenoidsare commonly enriched in13C relative ton-alkyl lipids. Like-wise, Schouten et al. (1998) recently showed that phytol can be2–5‰ more enriched than low-molecular fatty acids in culturedalgae. However,d13C values ofn-alkanes from modern higherplants vary considerably, ranging ca. 2 to 10‰ more depletedrelative to bulkd13C values (Collister et al., 1994; Lockheart etal., 1997). The range of values observed in modern plant lipidssuggests it is possible that isoprenoids andn-alkanes from sites608 and 516 share a common source. If so, pristane andphytane are allochthonous sedimentary components, derivedfrom terrigenous sources. We do not exclude the possibility thatisoprenoid lipids were produced by algae other than hapto-phytes or by photosynthetic bacteria (Sakata et al., 1997). Wenote, however, that compared to large changes (ca. 10‰) ind13C37:2, isoprenoidd13C values remain relatively invariant
Fig. 5. Carbon-isotope compositions ofn-alkanes from site 730. Dashed line represents isotopic trend of alkadienones(n 5 52).
43Isotope analyses of Miocene organic carbon
through time. Therefore, if isoprenoid lipids are derived fromalgae or bacteria, the narrow range in13C compositions re-quires physiological mechanisms that resisted carbon limita-tion.
6.3. Total Organic Carbon
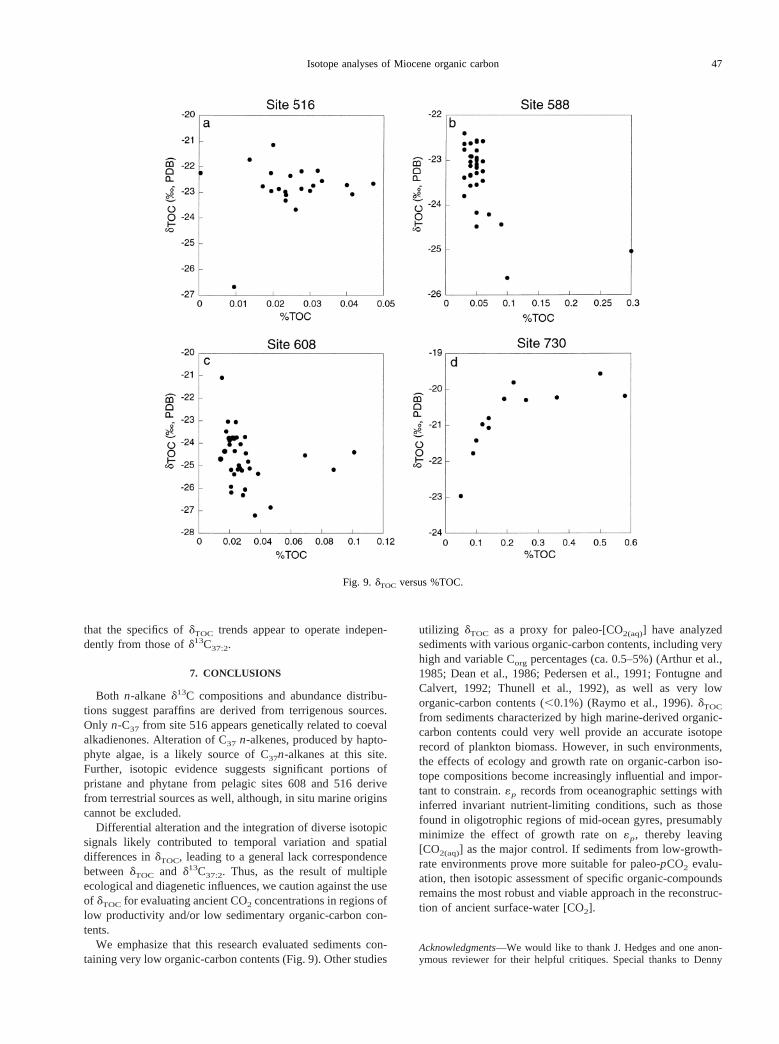
In general,dTOC appears to decrease with increasing %TOCat sites 588, 608, and 516 (Fig. 9). This relationship is theinverse of what would be predicted if increasing %TOC isassociated with increases in phytoplankton growth rates (Lawset al., 1995; Bidigare et al., 1997; Pancost et al., 1997). In-creases indTOC are correlated to increases in %TOC at site 730,where %TOC is orders of magnitude larger than at sites 588,608, and 516 (Fig. 9d). However, there are several possibilitieswhy this relationship at site 730 is expressed: (1) higher %TOCcorresponds to higher phytoplankton growth rates and thuspromoted lower«p values; (2) increasing %TOC reflects in-creasing algal export productivity and preservation over isoto-pically depleted terrestrial carbon input; or (3) higher %TOC isassociated with higher diatom production which can, in some
environments, generate organic carbon that is isotopically en-riched relative to haptophyte organic matter (Freeman et al.,1994; Pancost et al., 1997).d13C37:2 trends show little to norelationship to %TOC, suggesting higher haptophyte growthrates in general (and lower«p values) were not the primarycause for higherdTOC values. We suggest, ecological changesand/or changes in the proportion of marine to terrestrial sedi-mentary carbon input more likely controlleddTOC trends at site730.
Carbon-isotope trends of sedimentary TOC do not correlatewell with those of alkadienones (Fig. 8).dTOC compositions areat times similar, depleted, or enriched relative tod13C37:2 withlittle consistency between location or geologic time. Givenavailable research, there is little reason to suspect isotopicalteration of alkadienones through time (Hayes et al., 1990).However, the characteristics of sedimentary organic matter arequantitatively and qualitatively altered from the bulk organicmatter from which it was derived (Wakeham et al., 1997). Inpelagic settings, organic matter is largely biosynthesized in situby marine organisms in the euphotic zone. However, as ob-
Fig. 6. Carbon-isotope compositions ofn-alkanes from site 516. Dashed line represents isotopic trend of alkadienones(n 5 40).
44 M. Pagani, K. H. Freeman, and M. A. Arthur
served in this and other studies (Gagosian and Peltzer, 1985;Prahl et al., 1989; Ohkouchi et al., 1997), aerially transportedorganic matter from terrigenous sources can constitute a sig-nificant portion of sedimentary organic matter. Through theintegrated effects of heterotrophy occurring within both thewater column and sediment, only a small fraction of what isproduced in surface waters is typically preserved (Wakeham etal., 1997). In oligotrophic environments, alteration can be es-pecially significant because low surface productivity results inlimited organic-carbon export and surface-sediment accumula-tion. Alteration of bulk primary organic matter ultimately in-volves both differential preservation and transformation of or-ganic molecules (Cranwell et al., 1987; Wakeham et al.,1997). For example, sediment-trap experiments in the equa-torial North Atlantic show reduced fluxes with depth ofhydrocarbons, sterols, amino acids, and fatty acids in par-ticulate organic matter, with the more labile compounds,such as amino acids and fatty acids, undergoing the greatestrelative reductions (Wakeham et al., 1980; Wakeham, 1982;Lee and Cronin, 1982). Gong and Hollander (1997) per-formed comparative studies between oxic and anoxic bot-tom-water conditions in the Santa Monica Basin. They con-cluded that redox conditions exerted strong control onmolecular distributions and bulk isotopic compositions. Dif-ferences between sites were attributed to differential rates ofdecomposition and contributions of bacterial biomass underanoxic conditions.
Reactivity rates vary greatly among organic compounds(Hedges et al., 1988). Bonding structure and carbon chain
length greatly affect decomposition rates (Cranwell, 1984;Cranwell et al., 1987). In general, carbohydrates, amino acids,and pigments are labile and thus readily removed from surfacesediments (Hedges et al., 1988; Wakeham et al., 1997). Be-cause carbon-isotopic compositions of these and other com-pound classes are often distinct from bulk TOCd13C values,preferential removal can alter bulk isotopic compositions (Ben-ner et al., 1987). Spiker and Hatcher (1984) measured a 4‰depletion in bulkd13C in an algal sapropel from MangroveLake, Bermuda. They concluded that the isotopic shift of thisorganic-rich sediment, occurring in less than 4000 years, re-sulted from the selective loss of carbohydrates in conjunctionwith the preservation of lipid components (which are typicallyca. 4‰ depleted relative to TOC). However, parallel changes inthe d13C of carbonate shells suggests the possibility that thisshift was due to changes in water-column dissolved inorganiccarbon (Dean et al., 1985). McArthur et al. (1991) noted thatthe upper, oxidized portion of a turbidite from the northeastAtlantic was isotopically depleted relative to the bulk organiccarbon of the lower, unoxidized portion. Further, in addition topreferential loss, isotopic alteration of buried bulk organicmatter can occur via in situ contributions of isotopically de-pleted (Freeman et al., 1990) or enriched (Canuel et al., 1997)bacterial biomass.
In summary, the character and isotopic composition of bulksedimentary organic carbon results from an integration of au-tochthonous and allochthonous sources and diagenetic pro-cesses. The degree by which secondary processes affectdTOC
will be particularly large in regions with low rates of primary
Fig. 7. Carbon-isotope compositions of pristane and phytane from sites 608 and 516. Dashed line represents the isotopetrends of alkadienones.
45Isotope analyses of Miocene organic carbon
production and organic-carbon sedimentary flux, and oxygen-rich water columns. Furthermore, the dominance of a particularclass of photosynthetic algal and/or bacterial population maytemporally be altered with changes in circulation, water-col-
umn stability, and nutrient/trace-element availability (Pancostet al., 1997). In general,dTOC becomes increasingly depletedwith increases in terrestrial-organic input, and enriched withdiagenetic/secondary alterations. Therefore, it is not surprising
Fig. 8. Carbon-isotope compositions of total organic carbon (TOC). Dashes lines represent the isotope trends ofalkadienones (note: for site 588,n 5 84).
46 M. Pagani, K. H. Freeman, and M. A. Arthur
that the specifics ofdTOC trends appear to operate indepen-dently from those ofd13C37:2.
7. CONCLUSIONS
Both n-alkaned13C compositions and abundance distribu-tions suggest paraffins are derived from terrigenous sources.Only n-C37 from site 516 appears genetically related to coevalalkadienones. Alteration of C37 n-alkenes, produced by hapto-phyte algae, is a likely source of C37n-alkanes at this site.Further, isotopic evidence suggests significant portions ofpristane and phytane from pelagic sites 608 and 516 derivefrom terrestrial sources as well, although, in situ marine originscannot be excluded.
Differential alteration and the integration of diverse isotopicsignals likely contributed to temporal variation and spatialdifferences indTOC, leading to a general lack correspondencebetweendTOC and d13C37:2. Thus, as the result of multipleecological and diagenetic influences, we caution against the useof dTOC for evaluating ancient CO2 concentrations in regions oflow productivity and/or low sedimentary organic-carbon con-tents.
We emphasize that this research evaluated sediments con-taining very low organic-carbon contents (Fig. 9). Other studies
utilizing dTOC as a proxy for paleo-[CO2(aq)] have analyzedsediments with various organic-carbon contents, including veryhigh and variable Corg percentages (ca. 0.5–5%) (Arthur et al.,1985; Dean et al., 1986; Pedersen et al., 1991; Fontugne andCalvert, 1992; Thunell et al., 1992), as well as very loworganic-carbon contents (,0.1%) (Raymo et al., 1996).dTOC
from sediments characterized by high marine-derived organic-carbon contents could very well provide an accurate isotoperecord of plankton biomass. However, in such environments,the effects of ecology and growth rate on organic-carbon iso-tope compositions become increasingly influential and impor-tant to constrain.«p records from oceanographic settings withinferred invariant nutrient-limiting conditions, such as thosefound in oligotrophic regions of mid-ocean gyres, presumablyminimize the effect of growth rate on«p, thereby leaving[CO2(aq)] as the major control. If sediments from low-growth-rate environments prove more suitable for paleo-pCO2 evalu-ation, then isotopic assessment of specific organic-compoundsremains the most robust and viable approach in the reconstruc-tion of ancient surface-water [CO2].
Acknowledgments—We would like to thank J. Hedges and one anon-ymous reviewer for their helpful critiques. Special thanks to Denny
Fig. 9. dTOC versus %TOC.
47Isotope analyses of Miocene organic carbon

Walizer and Gabriel Montemurro for all their invaluable help in pro-cessing samples. Samples for this project were provided by DSDP/ODP. This research was funded by grants from NSF and JOI/USSAC.
REFERENCES
Arthur M. A., Walter D. E., and Claypool G. E. (1985) Anomalous13Cenrichment in modern marine organic carbon.Nature315,216–218.
Benner R., Fogel M. L., Sprague E. K., and Hodson R. E. (1987)Depletion of 13C in lignin and its implications for stable carbonisotope studies.Nature329,708–710.
Bentaleb I., Fontugne M., Descolas-Gros C., Girardin C., Mariotti A.,Pierre C., Brunet C., and Poisson A. (1996) Organic carbon isotopiccomposition of phytoplankton and sea-surfacepCO2 reconstructionsin the Southern Indian Ocean during the last 50,000 yr.Org. Geo-chem.4, 399–410.
Berggren W. A., Aubury M. P., and Hamilton N. (1983) Neogenemagnetostratigraphy of Deep Sea Drilling Project site 516 (RioGrand Rise, South Atlantic).Init. Rep. Deep Sea Drill. Proj.72,675–713.
Berggren W. A., Kent D. V., Swisher III C. C., and Aubry M. P. (1995)A revised Cenozoic geochronology and chronostratigraphy. InGeo-chronology, Time Scales and Global Stratigraphic Correlation(ed.W. A. Berggren et al.). Special Publication No. 54. pp. 129–212.Society for Sedimentary Geology.
Bidigare R. R., Fluegge A., Freeman K. H., Hanson D. L., Hayes J. M.,Hollander D., Jasper J. P., King L. L., Laws E. A., Milder J., MilleroF. J., Pancost R., Popp B. N., Steinberg P. A., and Wakeham S. G.(1997) Consistent fractionation of13C in nature and in the labora-tory: Growth-rate effects in some haptophyte algae.Global Biogeo-chem. Cycles11, 279–292.
Brassell S. C. (1993) Applications of biomarkers for delineating marinepaleoclimatic fluctuations during the Pleistocene. InOrganic Geo-chemistry: Principles and Application(ed. M. H. Engel and S. A.Macko). pp. 699–738. Plenum Press.
Burckle L. H. (1989) Distribution of diatoms in sediments of thenorthern Indian Ocean: Relationship to physical oceanography.Mar.Micropaleont.15, 53–65.
Cande S. C. and Kent D. V. (1992) A new geomagnetic polarity timescale for the Late Cretaceous and Cenozoic.J. Geophys. Res.97,13917–13951.
Canuel E. A., Freeman K. H., and Wakeham S. G. (1997) Isotopiccompositions of lipid biomarker compounds in estuarine plants andsurface sediments.Limnol. Oceanogr.42, 1570–1583.
Cerling T. E., Wang Y., and Quade J. (1993) Expansion of C4 eco-systems as an indicator of global ecological change in the lateMiocene.Nature361,344–345.
Cerling T. E., Harris J. M., MacFadden B. J., Leakey M. G., Quade J.,Eisenmann V., and Ehleringer J. R. (1997) Global vegetation changethrough the Miocene/Pliocene boundary.Nature389,153–158.
Clark R. C. J. and Blumer M. (1967) Distribution ofn-paraffins inmarine organisms and sediments.Limnol. Oceanogr.12, 79–87.
Collister J. W., Rieley G., Stern B., Eglinton G., and Fry B. (1994)Compound-specificd13C of leaf lipids from plants with differingcarbon dioxide metabolisms.Org. Geochem.21, 619–627.
Conte M. H., Volkman J. K., and Eglinton G. (1994) Lipid biomarkersof the Haptophyta. InThe Haptophyte Algae(ed. J. C. Green et al.).pp. 351–377. Clarendon Press.
Cranwell P. A. (1984) Lipid geochemistry of sediments from UptonBroad, a small productive lake.Org. Geochem.7, 25–27.
Cranwell P. A., Eglinton G., and Robinson N. (1987) Lipids of aquaticorganisms as potential contributors to lacustrine sediments-II.Org.Geochem.6, 513–527.
Dean W. E., Arthur M. A., and Claypool G. E. (1986) Depletion of13Cin Cretaceous marine organic matter: Source, diagenetic, or environ-mental signal?Mar. Geol.70, 119–157.
Degens E. T., Guillard R. R. L., Sackett W. M., and Hellebust J. A.(1968) Metabolic fractionation of carbon isotopes in marine plank-ton-I. Temperature and respiration experiments.Deep Sea Res.15,1–9.
Deines P. (1980) The isotopic composition of reduced organic carbon.In Handbook of Environmental Isotope Geochemistry(ed. P. Fritz etal.). 1, The Terrestial Environment. pp. 329–345. Elsevier.
Eglinton G. and Hamilton R. J. (1967) Leaf epicuticular waxes.Science156,1322–1334.
Flower B. P. and Kennett J. P. (1993) Middle Miocene ocean-climatetransition: High-resolution oxygen and carbon isotopic records fromDeep Sea Drilling Project site 588A, southwest Pacific.Paleocean-ography8, 811–843.
Fontugne M. R. and Calvert S. E. (1992) Late Pleistocene variability ofthe carbon isotopic composition of organic matter in the easternMediterranean: Monitor of changes in carbon sources and atmo-spheric CO2 concentrations.Paleoceanography7, 1–20.
Freeman K. H., Hayes J. M., Trendel J., and Albrecht P. (1990)Evidence from carbon isotope measurements for diverse origins ofsedimentary hydrocarbons.Nature343,254–256.
Freeman K. H., Boreham C. J., Summons R. E., and Hayes J. M. (1994)The effect of aromatization on the isotopic compositions of hydro-carbons during early diagenesis.Org. Geochem.21, 1037–1049.
Freeman K. H., Wakeham S. G., and Hayes J. M. (1994) Predictiveisotopic biogeochemistry: Hydrocarbons from anoxic marine basins.Org. Geochem.21, 629–644.
Gagosian R. B. and Peltzer E. T. (1986) The importance of atmosphericinput of terrestrial organic material to deep sea sediments.Org.Geochem.10, 661–669.
Gelpi E., Schneider J., Mann J., and Oro` J. (1970) Hydrocarbons ofgeochemical significance in microscopic algae.Phytochem.9,603–612.
Goericke R., Montoya J. P., and Fry B. (1994) Physiology of isotopefractionation in algae and cyanobacteria. InStable Isotopes in Ecology(ed. K. Lajtha et al.). pp. 187–221. Blackwell Scientific Publications.
Gong C. and Hollander D. J. (1997) Differential contribution of bac-teria to sedimentary organic matter in oxic and anoxic environments,Santa Monica Basin, California.Org. Geochem.26, 545–563.
Goossens H., de Leeuw J. W., Schenck P. A., and Brassell S. C. (1984)Tocopherols as likely precursors of pristane in ancient sediments andcrude oils.Nature312,440–442.
Han J. and Calvin M. (1969) Hydrocarbon distribution of algae andbacteria, and microbiological activity in sediments.Proc. NationalAcad. Sci.64, 436–443.
Hayes J. M., Freeman K. H., Popp B. N., and Homan C. H. (1990)Compound-specific isotopic analyses: A novel tool for reconstruc-tion of ancient bigeochemical processes.Org. Geochem.16, 1115–1128.
Hedges J. I., Clark W. A., and Cowie G. L. (1988) Fluxes andreactivities of organic matter in a coastal marine bay.Limnol. Ocean-ogr. 33, 1137–1152.
Hinga K. R., Arthur M. A., Pilson M. E. Q., and Whitaker D. (1994)Carbon isotope fractionation by marine phytoplankton in culture:The effects of CO2 concentration,pH, temperature, and species.Global Biogeochem. Cycles8, 91–102.
Hodell D. A. and Woodruff F. (1994) Variations in the strontiumisotopic ratio of seawater during the Miocene: Stratigraphic andgeochemical implications.Paleoceanography9, 405–426.
Huang Y., Eglinton G., Ineson P., Latter P. M., Bol R., and HarknessD. D. (1997) Absence of carbon isotope fractionation of individualn-alkanes in a 23-year field decomposition experiment withCallunavulgaris. Org. Geochem.26, 497–501.
Jasper J. P. and Hayes J. M. (1994) Reconstruction of paleoceanicpCO2 levels from the carbon isotopic compositions of sedimentarybiogenic components. InCarbon Cycling in the Glacial Ocean:Constraints on the Ocean’s Role in Global Change(ed. R. Aahn etal.). NATO ASI Series, I 17. pp. 323–341. Springer.
Jasper J. P., Mix A. C., Prahl F. G., and Hayes J. M. (1994) Photo-synthetic fractionation of13C and concentrations of dissolved CO2 inthe central equatorial Pacific during the last 255,000 years.Pale-oceanography6, 781–798.
Kroon D., Steens T. N. F., and Troelstra S. R. (1991) Onset ofmonsoonal related upwelling in the western Arabian Sea as revealedby planktonic foraminifera.Proc. Ocean Drill. Program, Sci. Res.117,257–263.
Laws E. A., Popp B. N., Bidigare R. R., Kennicut M. C., and MackoS. A. (1995) Dependence of phytoplankton carbon isotopic compo-sition on growth rate and [CO2]aq: Theoretical considerations andexperimental results.Geochim. Cosmochim. Acta59, 1131–1138.
Leckrone K. J. and Hayes J. M. (1997) Efficiency and temperature-
48 M. Pagani, K. H. Freeman, and M. A. Arthur

dependence of water removal by membrane dryers.Anal. Chem.69,911–918.
Lee C. and Cronin C. (1982) The vertical flux of particulate organicnitrogen in the sea: Decomposition of amino acids in the Peruupwelling area and the equatorial Atlantic.J. Mar. Res.40,227–251.
Lockheart M. J., Van Bergen P. F., and Evershed R. P. (1997) Varia-tions in the stable carbon isotope compositions of individual lipidsfrom the leaves of modern angiosperms: Implications for the study ofhigher land plant-derived sedimentary organic matter.Org. Geo-chem.26, 137–153.
Marlowe I. T., Green J. C., Neal A. C., Brassell S. C., Eglinton G., andCourse P. A. (1984) Long chain (n-C37–C39) alkenones in thePrymnesiophycea. Distribution of alkenones and other lipids andtheir taxonomic significance.Br. Phycol. J.19, 203–216.
McArthur J. M., Tyson R. V., Thomson J., and Mattey D. (1992) Earlydiagenesis of marine organic matter: Alteration of the carbon isoto-pic composition.Mar. Geol.105,51–61.
Michalczyk G. (1985) Determination ofn- and iso-paraffins in hydro-carbon waxes—comparative results of analyses by gas chromatog-raphy, urea adduction, and molecular sieve adsorption.Fette-Seifen-Anstrichmittel87, 481–486.
Nigrini C. (1991) Composition and biostratigraphy of radiolarian as-semblages from an area of upwelling (northwestern Arabian Sea,Leg 117).Proc. Ocean Drill. Program, Sci. Res.117,89–126.
Ohkouchi N., Kawamura K., and Taira A. (1997) Molecular paleocli-matology: Reconstruction of climate variabilities in the late Quater-nary.Org. Geochem.27, 173–183.
Pagani M. (1998) Miocene atmospheric carbon dioxide concentrationsand paleoceanography: Constraints from compound-specific isotopecompositions. Ph.D. dissertation, Pennsylvania State Univ.
Pagani M., Arthur M. A., and Freeman K. H., (1999) Miocene evolu-tion of atmospheric carbon dioxide.Paleoceanography14,273–292.
Pancost R. D., Freeman K. H., Wakeham S. G., and Robertson C. Y.(1997) Controls on carbon-isotope fractionation by diatoms in thePeru upwelling region.Geochim. Cosmochim. Acta61, 4983–4991.
Pedersen T. F., Nielson B., and Pickering M. (1991) Timing of lateQuaternary productivity pulses in the Panama Basin and implicationsfor atmospheric CO2. Paleoceanography6, 657–677.
Peters K. E. and Moldowan J. M. (1993)The Biomarker Guide,Interpreting Molecular Fossils in Petroleum and Ancient Sediments.Prentice Hall.
Popp B. N., Laws E. A., Bidigare R. R., Dore J. E., Hanson K. L., andWakeham S. G. (1998) Effect of phytoplankton cell geometry oncarbon isotopic fractionation.Geochim. Cosmochim. Acta62,69 –77.
Poynter J. G., Farrimond P., Robinson N., and Eglinton G. (1989)Aeolian-derived higher plant lipids in the marine sedimentaryrecord: Links with palaeoclimate. InPaleoclimatology and Paleo-meteorology: Modern and Past Patterns of Global AtmosphericTransport (ed. M. Leinen et al.). pp. 435–462. Kluwer AcademicPublishers.
Prahl F. G., Muehlhausen L. A., and Lyle M. (1989) An organicgeochemical assessment of oceanographic conditions at MANOPsite over the past 26,000 years.Paleoceanography4, 495–510.
Prahl F. G., Ertel J. R., Goni M. A., Sparrow M. A., and EversmeyerB. (1994) Terrestrial organic carbon contributions to sediments onthe Washington margin.Geochim. Cosmochim. Acta58,3035–3048.
Ragan M. A. and Chapman D. J. (1978)A Biochemical Phylogeny ofthe Protists. Academic Press.
Rau G. H., Takahashi T., and Des Marais D. J. (1989) Latitudinalvariations in planktond13C: Implications for CO2 and productivity inpast oceans.Nature341,516–518.
Rau G. H., Froelich P. N., Takahashi T., and Des Marais D. J. (1991)Does sedimentary organicd13C record variations in Quaternaryocean [CO2(aq)]? Paleoceanography6, 335–347.
Rau G. H., Takahashi T., Des Marais D. J., Repeta D. J., and MartinJ. H. (1992) The relationship betweend13C of organic matter and[CO2(aq)] in ocean surface water: Data from a JGOFS site in thenortheast Atlantic Ocean and a model.Geochim. Cosmochim. Acta56, 1413–1419.
Rau G. H., Riebesell U., and Wolf-Gladrow D. (1996) A model of
photosynthetic13C fractionation by marine phytoplankton based ondiffusive molecular CO2 uptake.Mar. Ecol. Prog. Ser.133,275–285.
Raymo M. E. and Horowitz M. (1996) Organic carbon paleo-pCO2 andmarine-ice core correlations and chronology.Geophy. Res. Lett.23,367–370.
Ricci M. P., Merritt D. A., Freeman K. H., and Hayes J. M. (1994)Acquisition and processing of data for isotope-ratio-monitoring massspectrometry.Org. Geochem.21, 561–571.
Rieley G., Collier R. J., Jones D. M., Eglinton G., Eakin P. A., andFallick A. E. (1991) Sources of sedimentary lipids deduced fromstable carbon-isotope analyses of individual compounds.Nature352,425–427.
Rontani J.-F., Cuny P., and Beker B. (1997) Stability of long-chainalkenones in senescing cells ofEmiliania huxleyi: effect of photo-chemical and aerobic microbial degradation on the alkenone unsat-uration ratio (U37
K9). Org. Geochem.26, 503–509.Rowland S. J. (1990) Production of acyclic isoprenoid hydrocarbons by
laboratory maturation of methanogenic bacteria.Org. Geochem.15,9–16.
Sakata S., Hayes J. M., McTaggart A. R., Evans R. A., Leckrone K. J., andTogasaki R. K. (1997) Carbon isotopic fractionation associated withlipid biosynthesis by a cyanobacterium: relevance for interpretation ofbiomarker records.Geochim. Cosmochim. Acta61, 5379–5389.
Schouten S., Klein Breteler W. C. M., Blokker P., Schogt N., RijpstraI. C., Grice K., Baas M., and Sinninghe Damste` J. S. (1998) Bio-synthetic effects on the stable carbon isotopic compositions of algallipids: implications for deciphering the carbon isotopic biomarkerrecord.Geochem. Cosmochim. Acta62, 1397–1406.
Shipboard S. P. (1991) Site 730.Proc. Ocean Drill. Program117,555–584.
Sikes E. L., Volkman J. K., Robertson L. G., and Pichon J. (1997)Alkenones and alkenes in surface waters and sediments of theSouthern Ocean: Implications for paleotemperature estimation inpolar regions.Geochim. Cosmochim. Acta61, 1495–1505.
Simo R., Colom-Altes M., Grimalt J. O., and Albaige`s J. (1991)Background levels of atmospheric hydrocarbons, sulphate and nitrateover the western Mediterranean.Atmos. Envir.25A, 1463–1471.
Spiker E. C. and Hatcher P. G. (1984) Carbon isotope fractionation ofsapropelic organic matter during early diagenesis.Org. Geochem.5,283–290.
Thunell R. C., Qingmin M., Calvert S. E., and Pedersen T. F. (1992)Glacial-Holocene biogenic sediment patterns in the south China Sea:Productivity variations and surface waterpCO2. Paleoceanography7, 143–162.
Villanueva J., Grimalt J. O., Cortijo E., Vidal L., and Labeyrie L.(1997) A biomarker approach to the organic matter deposited in theNorth Atlantic during the last climatic cycle.Geochim. Cosmochim.Acta 61, 4633–4646.
Volkman J. K., Eglinton G., Corner E. D., and Sargent J. R. (1980)Novel unsaturated straight-chain C37–C39 methyl and ethyl ketonesin marine sediments and a coccolithophoreEmiliania huxleyi. InAdvances in Organic Geochemistry 1979; Proceedings of the NinthInternational Meeting on Organic Geochemistry(ed. A. G. Douglaset al.). pp. 219–227. Pergamon Press.
Wakeham S. G. (1982) Organic matter from a sediment trap experi-ment in the North Atlantic: wax esters, steryl esters, triacylglycerolsand alkyldiacylglycerols.Geochim. Cosmochim. Acta46, 2239–2257.
Wakeham S. G. (1989) Reduction of stenols to stanols in particulatematter at oxic-anoxic boundaries in sea water.Nature342,787–790.
Wakeham S. G., Farrington J. W., Gagosian R. B., Lee C., de Baar H.,Nigrelli G. E., Tripp B. W., Smith S. O., and Frew N. W. (1980)Organic matter fluxes from sedimentary traps in the equatorial NorthAtlantic. Nature286,789–800.
Wakeham S. G., Lee C., Hedges J. I., Hernes P. J., and Peterson M. L.(1997) Molecular indicators of diagenetic status in marine organicmatter.Geochim. Cosmochim. Acta61, 5363–5369.
Wong W. W. and Sackett W. M. (1978) Fractionation of stable carbonisotopes by marine phytoplankton.Geochim. Cosmochim. Acta42,1809–1815.
49Isotope analyses of Miocene organic carbon