is there a relationship between polycyclic aromatic hydrocarbon
TRANSCRIPT

UC IrvineUC Irvine Electronic Theses and Dissertations
TitleIs there a Relationship Between Polycyclic Aromatic Hydrocarbon Exposure and Ovarian Reserve Markers in Humans?
Permalinkhttps://escholarship.org/uc/item/54z6194j
AuthorNelson, Gillian
Publication Date2016-01-01
LicenseCC BY-NC-ND 4.0 Peer reviewed|Thesis/dissertation
eScholarship.org Powered by the California Digital LibraryUniversity of California

UNIVERSITY OF CALIFORNIA,
IRVINE
Is there a Relationship Between Polycyclic Aromatic Hydrocarbon Exposure and Ovarian
Reserve Markers in Humans?
THESIS
submitted in partial satisfaction of the requirements
for the degree of
MASTERS OF SCIENCE
in Environmental Health Sciences
by
Gillian Loraine Nelson, MD
Thesis Committee:
Professor Ulrike Luderer, MD, PhD, MPH, Irvine, Chair
Professor Dean Baker, MD, MPH
Associate Professor Jun Wu, PhD
2016

Copyright 2016 Gillian Loraine Nelson, MD

ii
TABLE OF CONTENTS
Page
LIST OF FIGURES iii
LIST OF TABLES iv-v
ACKNOWLEDGMENTS vi
ABSTRACT OF THE THESIS vii
1.0 INTRODUCTION 1
2.0 FEMALE REPRODUCTIVE PHYSIOLOGY 31
3.0 CONFOUNDERS OF OVARIAN RESERVE 39
4.0 STUDY OBJECTIVES 41
5.0 METHODS 41
6.0 STATISTICAL ANALYSIS METHODOLOGY 47
7.0 RESULTS 49
8.0 DISCUSSION 76
9.0 CONCLUSION 82
REFERENCES 84

iii
LIST OF FIGURES
Page
Figure 1 Benzo[ a ]pyrene Structure 3
Figure 2 Metabolism of Benzo[a]pyrene and Phenanthrene 14
Figure 3 Folliculogenesis 35
Figure 4 AMH Decreases With Age 38
Figure 5 AMH Parallels Antral Follicle Count 39
figure 6 OH-PAH Principal Components Factor Analysis 55

iv
LIST OF TABLES
Page
Table 1: PAH Urinary Metabolite names and abbreviations 17
Table 2: PAH Parent Compound and OH-PAH metabolites evaluated in this thesis 24
Table 3: Participant Characteristics (Demographics) 50
Table 4: Participants' and NHANES PAH Geometric Means 51
Table 5: Correlations among mean OH-PAH metabolites in cycle 1 52
Table 6: Correlations among mean OH-PAH metabolites in cycle 2 52
Table 7: Correlations among mean OH-PAH metabolites in cycle 3 53
Table 8: Principal Component Factor Analysis of OH-PAHs 54
Table 9: Correlations of natural log transformed Endocrine Endpoints 56
Table 10: Correlations between age and BMI with OH-PAH metabolites 56
Table 11: Correlations of Potential Confounders with Ovarian Reserve Markers 57
Table 12: Bivariate Associations between OH-PAH Metabolites and Race/Ethnicity 59
Table 13: Bivariate Associations between OH-PAH Metabolites and Smoking Status 60
Table 14: Bivariate Associations between OH-PAH Metabolites and EtOH 61
Table 15: Bivariate Associations of OH-PAH Metabolites and Caffeinated Coffee 62
Table 16: Bivariate Associations between OH-PAH metabolites and Caffeinated Tea 62
Table 17: Bivariate Associations between OH-PAH Metabolites and Soda/Energy Drink 63

v
Page
Table 18: Bivariate Associations of OH-PAH Metabolites and Education 64
Table 19: Bivariate Associations of OH-PAH Metabolites and Family Income past year 65
Table 20: Bivariate associations of Race/Ethnicity and Ovarian Reserve Markers 67
Table 21: Bivariate Associations of Cigarette use and Ovarian Reserve Markers 67
Table 22: Bivariate Associations between ETOH and Ovarian Reserve Markers 68
Table 23: Bivariate Associations of Caffeinated Coffee and Ovarian Reserve Markers 68
Table 24: Bivariate Associations between Caffeinated Tea and Ovarian Reserve Markers 69
Table 25: Bivariate Associations of Soda/Energy Drink and Ovarian Reserve Markers 69
Table 26: Bivariate Associations between Education level and Ovarian Reserve Markers 70
Table 27: Bivariate Associations of Family Income past year and Ovarian Reserve Markers 70
Table 28: AMH Multiple Linear Regression Model 71
Table 29: Inhibin B Multiple Linear Regression Model 71
Table 30: FSH Multiple Linear Regression Model 72
Table 31: AMH Effect Modification by GSTT1 74
Table 32: AMH Effect Modification by GSTM1 74
Table 33: Inhibin B Effect Modification by GSTT1 75
Table 34: Inhibin B Effect Modification by GSTM1 75
Table 35: FSH Effect Modification by GSTT1 75
Table 36: FSH Effect Modification by GSTM1 76

vi
ACKNOWLEDGMENTS
I will be forever grateful and proud to have learned from my committee chair, Dr. Ulrike
Luderer during my journey through this thesis project. I sincerely thank her for her valuable
time, unmatched knowledge, and endless patience with me during this endeavor.
I am equally proud and sincerely thankful to have been given the opportunity to have learned
from Drs. Dean Baker and Jun Wu. My thesis project would not have been possible without the
input and patience of every one of the committee members.

vii
ABSTRACT OF THESIS
Is there a Relationship Between Polycyclic Aromatic Hydrocarbon Exposure and
Ovarian Reserve Markers in Humans?
by
Gillian Nelson, MD
Master of Science in Environmental Health Sciences
University of California, Irvine, 2016
Ulrike Luderer, MD, PhD, MPH, Chair
About 12% of American women suffer from infertility, and premature ovarian failure
(POF) is a major cause of infertility. Most cases of POF are of unknown etiology. Environmental
toxicants such as polycyclic aromatic hydrocarbons (PAHs) have been shown in animal models
and human tissue cultures to destroy ovarian primordial follicles. The aim of this study is to
assess whether PAH exposure is associated with diminished ovarian reserve in humans.
Hypotheses: Women with higher levels of PAH exposure will have decreased ovarian
reserve compared to women with lower levels of PAH exposure. Women who have enzyme
polymorphisms that alter Phase II metabolism of PAHs will have increased susceptibility to the
ovarian toxicity of PAHs compared to women who do not have these polymorphisms.
Methods: This cross-sectional data analysis of a longitudinal study involved 51 women of
reproductive age who resided in Orange County, California. Urinary monohydroxylated PAH
metabolites (OH-PAHs) were used as biomarkers for exposure to PAHs. Anti-Mullerian hormone

viii
(AMH), follicle-stimulating hormone (FSH) and inhibin B served as markers of ovarian reserve.
Null polymorphisms in the Phase II enzymes GSTT1 and GSTM1 were determined. Multiple
linear regression models were generated and Principal Component Factor analysis with varimax
rotation was used to assess co-linearity of the OH-PAHs. The OH-PAHs with the strongest
associations with each of the ovarian reserve markers were forced into the regression models.
Potential confounders identified in bivariate analyses were entered into the models and
retained if they changed the beta coefficients for the OH-PAHs by 10% or more.
Results: Concentrations of urinary OH-PAHs in the study population were similar to those
reported for women in the National Health and Nutrition Examination Survey (NHANES'). The
OH-PAHs with the strongest associations with the ovarian reserve markers were 2-hydroxy
naphthalene (NAP2) and 9-hydroxy fluorene (FLUO9) for the AMH model, 3- hydroxy fluorene
(FLUO3) for the inhibin B model, and 3-hydroxy phenanthrene (PHEN3) for the FSH model.
Factor analysis identified two factors that explained 37% and 63 % of the variation in the OH-
PAHs, respectively. Factor 1 primarily loaded on 1-hydroxy phenanthrene (PHEN1), 2-hydroxy
phenanthrene (PHEN2), PHEN3, and FLUO9. Factor 2 primarily loaded on 2-hydroxy fluorene
(FLUO2), FLUO3, 1-hydroxy naphthalene (NAP1), and NAP2. In the final models, the beta
coefficients for the OH-PAHs were in the expected direction (negative for AMH and inhibin B
models and positive for the FSH model), but none of them was statistically significant at the
P=0.05 level.
We examined effect modification by GSTM1 and GSTT1 null polymorphisms in stratified
analyses. Our models revealed non-significant associations between OH-PAHs and ovarian

ix
reserve markers in the expected directions for the strata with ≥ 1 allele and no or opposite
than expected associations for the strata with null alleles. However, the numbers in each
stratum were very small.
Conclusions: Our findings are suggestive that environmental exposure to PAHs may be
associated with decreased ovarian reserve. Larger studies are needed and could be conducted
with techniques used in this pilot study.

1
1.0 INTRODUCTION
1.1 Polycyclic Aromatic Hydrocarbons (PAHs)
The International Union of Pure and Applied Chemistry (IUPAC) defines PAHs as
compounds consisting of only carbon and hydrogen atoms with multiple aromatic rings (two to
six )(Levy, B et. al., 2011). PAHs' rings are fused, such that two adjacent benzene rings share
two carbons (CDC, 2013). PAHs are a group of over a hundred organic chemicals found in
nature in mixtures. Most PAHs are created during combustion and pyrolysis processes of
anthropogenic and natural origin. Many PAHs are emitted from incomplete combustion of
organic compounds (Domingo, JL et al., 2015). Naphthalene is the only PAH that is commercially
produced in the US (CDC, 2013). Most PAHs persist in the environment and are insoluble in
water. All the PAHs are solids although some have low vapor pressures and are volatile, such as
naphthalene (ATSDR, 1995). The manufactured or pure PAHs, range from colorless to white or
pale yellow-green solids (USEPA, 2006). The large PAHs (composed of 5 or more rings) such as
7,12-dimethylbenzanthracene (DMBA), 3-methylcholanthrene (3-MC) and benzo[a]pyrene
(BaP) have been the most studied in general and with regard to the ovary.
Naphthalene (two rings) is a PAH used as an intermediate (NAP2) in the manufacturing of
pesticides, as a precursor for various dyestuffs, pigments, in rubber processing chemicals and
other miscellaneous chemicals such as toilet bowel deodorants, tanning agents, and
pharmaceuticals. Fluorene (three rings) is used as an intermediate in several chemical
processes such as forming pesticides, to form polyradicals for resins and in manufacturing
dyestuffs; anthracene (three rings) is used in the manufacturing of dyes, lubricating materials
and thermosetting plastics (Case Studies, 2009; ATSDR, 2013; ATSDR, 1995; USEPA, 2006; CDC,

2
2013; Ladou, M et al., 2014; Collin, G et al., 2003). PAH compounds with up to three or four
aromatic rings (phenanthrene, naphthalene, and pyrene) predominate in mixtures (Ladou, M et
al., 2014).
PAHs with five or six rings are classified as carcinogenic, mutagenic, and
immunosuppressive (Ladou, M et al., 2014). The International Agency for Research on Cancer
(IARC) and the US National Toxicology Program (NTP) have classified specific PAH-containing
chemical mixtures (e.g., soot, coke oven emissions, coal tars and coal tar pitches) as human
carcinogens. The NTP determined in 2000 that naphthalene is reasonably anticipated to be a
human carcinogen (NTP, 2005). IARC classifies naphthalene as a possible human carcinogen
(CDC, 2013, Preuss, R et al., 2003). Because of these health effect concerns, the US
Occupational Safety and Health Administration (OSHA) has developed criteria on the allowable
levels of these chemicals in the workplace (CDC, 2013).
Each ring structure in a PAH has a unique ultraviolet spectrum, dictating the unique
absorbance of each isomer, which is used in the identification of that PAH. BaP is internationally
used as a marker compound for PAH measurements in air and is classified as the primary ( IARC
Group 1) carcinogen in tobacco smoke (Angerer, J et al., 1997; Levy, BS et al., 2011). Figure 1 is
a structural representation of BaP:

3
Figure 1. Benzo[a]pyrene (BaP)
The Agency for Toxic Substances and Disease Registry (ATSDR) identified 17 PAHs as being
of greatest health concerns (ATSDR, 1995). These PAHs were selected using the following four
criteria: (1) toxicity (2) potential for human exposure (3) frequency of occurrence at the United
States Environmental Protection Agency's (USEPA) National Priorities List (NPL) of hazardous
waste sites, and (4) the extent of information available. The 17 PAHs are listed below.
• acenaphthene • acenaphthylene • anthracene • benz[a]anthracene • benzo[a]pyrene •
benzo[e]pyrene • benzo[b]fluoranthene • benzo[g,h,i]perylene • benzo[j]fluoranthene •
benzo[k]fluoranthene • chrysene • dibenz[a,h]anthracene • fluoranthene • fluorene • indeno[
1,2,3-c,d]pyrene • phenanthrene • pyrene
The USEPA priority air pollutant list of 16 PAHs includes those that have less than two dozen
ring carbons or 4 to 6 rings and overlaps with the ATSDR's list except that naphthalene is
included and benzo[e]pyrene and benzo[j]fluoranthene are excluded (NCBI).

4
1.2 PAH Sources and Exposure Pathways
1.2.1 Air
There are multiple sources of airborne PAH mixtures. Important non-natural PAH sources in
air include tobacco smoke, motor vehicle exhaust (major source in urban and suburban areas),
residential and industrial heating sources, coal, crude oil and natural gas processing, and waste
incineration (CDC, 2013; ATSDR, 1995). As previously mentioned, it is generally agreed upon in
the literature that naphthalene, phenanthrene, and pyrene are dominant constituents in PAH
mixtures. Diesel exhaust has PAH adsorbed to particulates (primarily fluoranthrene,
phenanthrene, and anthracene) and PAH vapor emissions dominated by phenanthrene and
anthracene (Westerholm, R et al., 1994; Rogge, WF et al., 1994; Kelly, G et al., 1993). Two or
three ring PAHs are usually emitted in gaseous form and condense quickly, adsorbing onto
particles in soot or particulate matter air pollution (WHO, 1987). Fluorene is mostly detected in
the vapor phase (but present in air particulates) of various PAH emission sources, including coal
tar pitch, petroleum refineries, diesel exhaust fumes, and tobacco smoke, where it is the
second (BaP is the first as mentioned above) most abundant PAH (Fang, GC et al., 2006; Ding, YS
et al., 2005). Larger, five or more ring PAHs are emitted in the particulate phase (Byeong-Kyu, L
et al., 2010). The smaller or fine particulates (e.g., particulate matter equal to or less than 2.5
millimeters (PM 2.5)) have higher concentrations of PAHs than larger or coarse particulates
(Boström, CE et al., 2002; Rehwagen, M et al., 2005). Naphthalene is a constituent of diesel and
jet fuel (Clark, CR et al., 1982; McDougal, JN et al., 2000). Cigarette smoke contributes
significantly to the naphthalene exposure of humans (Schmeltz , I et al., 1976; Hoffmann, D et
al. 2001). Together these exposures lead to a background burden of PAH exposure for the

5
general population. Natural airborne sources of PAHs include volcanoes, forest fires, crude oil,
and shale oil (Case Series, 2009; ATSDR, 1995). PAH concentrations in the ambient air have
shown seasonal variation (IPCS, 1998; Rehwagen, M et. al., 2005).
Indoors, PAHs exist in the vapor phase, with the remainder partitioning to suspended or re-
suspended particles as well as dust deposited on room surfaces (settled dust). Inhaled indoor
air (both gas and particle phases) and ingestion of settled house dust are two prominent
pathways of human exposure to PAHs (Harrad, S et al., 2010). Indoor concentrations of PAHs
are influenced by outdoor concentrations (Lioy, PL et al., 1988), and indoor sources include
tobacco smoke, unvented radiant and convective kerosene space heaters, and gas cooking and
heating appliances (ATSDR, 1995). The levels of PAHs vary with different types of tobacco
varieties (Ding, YS et al., 2008). Households with smokers and gas stoves have the highest
concentration of PAHs (ATSDR, 1995). Typical ambient and residential air concentrations of
PAHs are in the nanogram per meter cubed (ng/m3) range in the US (Jung, KH et al., 2014;
Naumova, YY et al., 2002).
1.2.2 Water
PAHs have been found in some drinking water supplies in the US. Background levels of PAHs
in drinking water range from 4 to 24 nanograms per liter (ng/L) (ATSDR, 1995). Concentrations
of BaP in treated drinking water have been shown to range from 0.0002 to 0.024 micrograms
per liter (μg/L) (WHO, 1987; IARC, 1983), which are generally lower than those in untreated
water and about 100-fold lower than the USEPA's drinking water standard (0.2 parts per billion
(ppb), since chlorination eliminates aqueous PAHs (Kennish, MJ 1996; Case Studies, 2009).
These treatments decrease personal exposure to PAHs surface water deposition of airborne

6
PAHs. Atmospheric fallout includes wet and dry depositions of particulate and vapor phases of
PAHs during precipitation, as some are semi-volatile compounds such as pyrene (Guo, W et al.,
2007; Ma, WL et al., 2010; Zhang, Y et al., 2004). Surface water PAH contamination also comes
from municipal waste water discharge, urban storm water runoff, runoff from coal storage
areas , effluents from wood treatment plants and other industries, oil spills such as during oil
shipment at sea, and petroleum processing (ATSDR, 1995) as PAHs can leach from soil into
water (Case Studies, 2009).
1.2.3 Soil
Srogi reported in 2007 that the US was second only to Austria in soil concentrations of PAHs
at 59 microgram per gram (μg/g) compared to 79 μg/g (Srogi, K 2007). PAHs are found in soil
near areas where wood, coal, gasoline, and other products have been burned, such as former
manufactured-gas-factory sites and wood-preserving facilities (ATSDR, 1995). Documented
levels near oil refineries in the past have been as high as 200,000 micrograms per kilogram (μg
/kg) of dried soil (Case Studies, 2009). Soil samples obtained near cities and areas with heavy
traffic have generally averaged less than 2,000 μg/kg (IARC, 1973; Case Studies, 2009). There is
a national waste site clean-up effort administered by the EPA based on a mandate from the
Comprehensive Environmental Response, Compensation, and Liability Act. The US EPA's
Superfund Program locates, investigates, and cleans up the worst hazardous waste sites in the
US. Fifty-four PAHs have been identified at a number of NPL hazardous waste sites (NCBI).
There are two NPL sites in Orange County California near which participants in the present
study may have lived. The first site is the El Toro Marine Corps Air Station in Santa Ana, which is
a former location of the Fleet Marine Forces in the Pacific Ocean. The area around the site

7
changed from an agricultural area to an urban area (NETR, 1990). The second area is the McColl
site in Fullerton which is an inactive waste disposal facility covering 8 acres. The residents are
mostly exposed to waste from oil refinery acid sludge, which is predominantly sulfur and
hydrocarbons - including PAHs. They also have inhalation exposure to organic gas mixtures,
which would include PAHs (NETR, 1983).
1.2.4 Food
Up to 70% of PAH exposure for nonsmokers in a non-occupational setting can be associated
with diet (Phillips, DH 1999). PAHs are found in caffeinated tea, caffeinated roasted coffee,
cereals, grains, soy, flour, bread, vegetables, refined vegetable oil, fruits, meat, and processed
or pickled foods (Levy, B et al., 2011). Food grown in contaminated soil or air may also contain
PAHs. A few crops such as wheat, rye, and lentils, may synthesize PAHs or absorb them through
water, air, or soil (Menzie, CA et al., 1992; Shabad, LM et al., 1972; Grimmer, G, 1968; IARC,
1973). Smoked meat and fish contain more PAHs than do uncooked products with up to 2.0
μg/kg of BaP detected in smoked fish (Case Studies, 2009). Cooking the meat (muscle meat
such as beef, pork, poultry, and fish) or other food at high temperature increases the amount of
PAHs, which are formed from the fat and the juices of the meats in the food (Jägerstad, M et
al., 2005). For example, the PAH level for charred meat can be as high as 10–20 µg/kg (Phillips,
DH 1999) which has been shown to be 3 to 5 times more PAHs compared to grilling (Mc Donald,
JD et al., 2003). Rose et al., reported that barbecuing with charcoal plus wood chips resulted in
the formation of BaP in most foods; barbecuing beef burgers over charcoal without using wood
chips gave the highest levels in the study as they were closer to the heat source (Rose, M et al.,
2015). Epidemiology studies support these findings, such as the Li et al., study that reported

8
urinary concentrations of PAHs after consumption of barbecued chicken were in similar ranges
or as high as occupational exposure to PAHs (Li, Z et al., 2012). Secondly, self-reported
grilled/broiled meat consumption was reported to be associated with increased concentrations
of most PAH metabolites in a cross-sectional analysis of 5,060 participants over 6 years of age
using data from the 1999-2002 National Health and Nutrition Examination Survey (NHANES)
(Suwan-ampai, P et al., 2009). Third, Vyskocil et al. reported that food consumption
represented the main source of PAH exposure for children in big cities that were not heavily
polluted by PAH from industrial sources (Vyskocil, A et al., 2000). Finally, Kim, SR et al., found in
their study of nonsmoking women that for fluorene and phenanthrene, ingested doses were
the dominant pathway of exposure (Kim, SR et al., 2008). The level of PAHs in the typical US
diet is less than 2 µg/kg of food (ATSDR, 1995). However, the recent reports on PAHs and diet
by Ma, Y et al., 2015, for example indicate that US dietary exposure to PAHs ranges from 0.1 to
55 μg/day, depending on the amount of kgs of food intake (Ma et al., 2015). Zelinkova, Z et al.,
2015 reviewed the occurrence of the 16 USEPA air pollutant priority list of PAHs in food in the
US and in Europe. Their findings were consistent with other authors' prior reports on federal
guidelines that address the consumption of foods containing PAHs. Transplacental and
lactational routes are important exposure routes to PAHs for fetuses and infants. PAH
metabolites are excreted in breast milk and they readily cross the placenta (Case Studies, 2009).
The World Cancer Research Fund/American Institute for Cancer Research issued a report in
2007 recommending guidelines for the consumption of red and processed (including smoked)
meats; however, no recommendations were provided for PAH levels in smoked meat products

9
(WCRF/AICR, 2010). To date, no maximum limits for PAH contents of pre-cooked foodstuffs
have been implemented into the US legislation (Zelinkova, Z et al., 2015).
1.2.5 Topical Medications
Coal tar shampoos, lotions, and creams found in prescription and nonprescription formulas
for treatment of psoriasis, eczema, and dandruff contain PAHs (ATSDR, 1995; Case Studies,
2009). Disdier et. al., found fluorene, phenanthrene, pyrene, and naphthalene, among other
PAHs in an analysis of coal tar creams (Disdier, B et al., 2000).
1.2.6 Occupational Exposure
The highest risks of PAH exposure to workers are in industries or trades involving coal and
coal products (Case Studies, 2009). Specific jobs in these industries include but are not limited
to chimney sweeps , fishermen using coal tar on nets, machinists, printers, steel foundry
workers (Case Studies, 2009). Workers such as engine mechanics and drivers are exposed to
PAHs by inhaling auto and diesel engine exhaust. Other workers are exposed by using products
that contain PAHs in the following industries: mining, oil refining, chemical production, and the
electrical industry's graphite electrode workers (ATSDR, 1995; Case Studies, 2009). Specifically,
truck drivers, underground miners, and railroad workers have documented exposures to
methylated naphthalenes and phenanthrenes from diesel exhaust exposure and utility workers
to fluorene (Ladou, J et al., 2014).
Creosote is used as a wood preservative by carpenters, farmers, railroad workers, and
telegraph pole workers and in tunnel construction. It's major constituent is naphthalene and its
alkyl homologues. It is impregnated into lumber under high pressure. It is a constituent of fuel

10
oil, a lubricant for die molds, as well as pitch for roofing (Ladou, J et al., 2014; Siegel, E et al.,
1986). According to the US Department of Labor's April 2013 report, the number of women
employed in the US construction industry grew by 81.3% from 1985 to 2007 (US Bureau Labor
Statistics, 2010).
1.3 PAH Biodegradation
The degradation of PAHs in the environment occurs through photo and chemical oxidation
as well as biotic metabolism in estuarine and marine systems. In seawater, PAHs are degraded
directly by photolysis reactions or photo-induced reactions involving a molecule of ozone
oxygen, hydroxyl radicals, and other oxidizing agents in the upper part of the water column
(Kennish, MJ 1996). Bacteria under aerobic conditions, compared to fungi and aquatic fauna,
more quickly degrade the majority of PAHs, especially in highly contaminated sites such as oil
spills (Kennish, MJ 1996).
1.4 PAH Toxicokinetics
1.4.1 Absorption
PAH's are highly lipid soluble and are well-absorbed from the lung, gut, and skin of
mammals (WHO, 1987). In a 12 week exposure of rats to either inhaled (radio-labeled on
carbon number14 (14C)-BaP adsorbed on carbon black particles or inhaled pure BaP , the
authors reported a 100-fold higher level of 14C in the lungs for the adsorbed BaP (Wolff, RK et
al., 1989). Biphasic respiratory clearance of PAHs have been shown in female rats where the
bronchial airways partially clear some of the particles during mucociliary transport mechanisms,
leaving the rest to penetrate into the bronchial epithelial cells where metabolism takes place
(WHO, 1987). In addition to inhalation, BaP, phenanthrene and pyrene have been shown to

11
rapidly penetrate the skin of mice and rats. In mice after dermal exposure, 80% of BaP was
recovered from feces after 7 days, while a total of 42% was recovered from feces and urine in
rats (Sanders, CL et al., 1984; Yang, JJ et al., 1986).
All routes of absorption have been illustrated in human studies. In humans, they are largely
absorbed from carrier particles through the respiratory route (Ladou, J et al., 2014) as well as
through the gut. Human skin absorption has been demonstrated after using topical BaP (Kao, J
et al., 1985). Evidence of absorption measured with urinary levels of 1-hydroxypyrene (PYR1),
the urinary metabolite of pyrene, has been found in patients treated with topical coal tar
applications (Larsen, JC 1995). Absorption after inhalation and ingestion of PAHs has also been
demonstrated in workers exposed to creosote oil, in coal tar distillery workers, and in road-
surfacing. Evidence of absorption through inhalation and ingestion in coke-oven workers, other
workers exposed to bitumen fumes while working with binders to waterproof roofs and
workers in crude oil refineries has been reported (Larsen, JC 1995). Respiratory absorption was
also illustrated in coke-oven workers and city residents where significant correlations were
obtained between the metabolite PYR1 and levels of pyrene and BaP in the ambient air (Larsen,
JC 1995).
1.4.2 Distribution
PAHs are rapidly and widely distributed in organisms independent of the route of exposure.
Similar patterns of distribution were found with intrathecal, intravenous, and subcutaneous
exposures to PAHs in rats and mice. Detectable BaP levels are observed in most internal organs
from minutes to hours after administration with the highest levels in the liver (IARC, 1983; Foth,
H et al., 1988). Fatty tissues are significant storage depots for PAHs without accumulation.

12
Hepatobiliary excretion results in an abundance of PAH metabolites distributed in the
gastrointestinal tract (Schlede, E et al., 1970; Wiersma, DA 1983). BaP readily crosses the
placental barrier of rats and mice (Neubert, D et al., 1988; Withey, JI et al., 1993), consistent
with the fetal and developmental toxicity of the substance (WHO, 1987).
1.4.3 Metabolism
Metabolizing capacity of PAHs varies among animal tissues. The highest metabolizing
capacity in descending order takes place in liver, lung, intestinal mucosa, skin and kidneys. PAH
metabolism may also take place in nasal tissues, mammary glands, spleen, brain, hair follicles,
erythrocytes, platelets, leukocytes, gonads, placenta and uterus. Animal and human fetal
tissues metabolize PAHs at a lower rate compared to their respective adult tissues (Anderson,
LM et al., 1989). The ovary has been shown to metabolize PAHs (Rekhadevi, PV et al., 2014).
Phase I metabolism in the ovary involves the cytochrome P450 (CYP) enzymes (Bhattacharya, P
et al., 2012; Klaassen, C et al., 2008) and microsomal epoxide hydrolases (EHs), similar to other
tissues.
The major pathways involved in the metabolic transformation of PAHs are the CYP pathway
mentioned above and the aldo-keto reductase pathway (AKR pathway) (Zhang, L et al., 2012).
In most cases, oxidation of PAHs by CYP enzymes is an initial step in the activation process to
produce the polar biochemically reactive electrophilic species capable of interacting with
cellular macromolecules, particularly nucleic acids and proteins (Xue, W et al., 2005). See BaP
and phenanthrene in Figure 2 for example. Specifically, PAHs are activated by the microsomal
mixed function oxidases, mainly CYP1A1 and CYP1B1, which oxidize the non-polar PAHs into
polar hydroxy and reactive epoxide-derivative intermediates. The reactive epoxides are

13
enzymatically metabolized by microsomal EHs to dihydrodiols or spontaneously convert to
phenols (Ladou, J et al., 2014; Shimata, T et al., 2004; Hall, M et al., 1989 Glatt, HR et al., 1987).
The aryl hydrocarbon receptor (AHR) is in the cytosol and is responsible for the inducible
expression of several xenobiotic-metabolizing enzymes including CYP1A1 and CYP1B1 by PAHs
(Shimada, T et al., 2004). AHR is a receptor for low molecular weight sterically planar ligands
that are approximately three benzene rings in size (such as the PAHs examined in the current
study), the most widely recognized being, 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) (Denison,
MS et al., 2003). AHR is expressed in many tissues, including the ovaries (Furue, M et al., 2014;
Schnekenburger, M et al., 2007).
The AKRs are a superfamily of monomeric nicotinamide adenine dinucleoside phosphate-
oxidase (NADPH)-dependent oxidoreductases in the cytosol (Disdier, B et al., 2000; Zhang, L et
al., 2012). The PAH phenanthrene is metabolized through this later pathway along with BaP
(Xue, W et al., 2005). Xue, et al., reported on dihydrodiol metabolism. Their metabolites are
oxidized through the AKR pathway as an alternative. The dihydrodiol dehydogenases are
converted to ketols, which then tautomerize to catechols. The later, oxidizes first to an o-semi-
quinone anion radical and hydrogen peroxide and then to o-quinone and superoxide anion
radical (Xue, W et al., 2005).

14
Figure 2. Cancer Epidemiology Biomarkers Prev., October 1, 2006 vol. 15 no. 10 1805-1811
Metabolism of BaP to trans, anti-BaP-tetraol via BPDE (metabolic activation pathway) and to phenols, such as 3-
HO-BaP (metabolic detoxification pathway). (top) Metabolism of phenanthrene (Phe) to PheT and phenanthrols,
such as 3-HOPhe. (bottom).
PAHs are not considered toxic as parent compounds (Chaloupka, K et al., 1994). They
require activation to become toxic. The structures and reactions in Figure 2 illustrate that
phenanthrene metabolism is similar to BaP's. The lower half of Figure 2 illustrates
phenanthrene with the generic symbols for CYP1A1 and 1B1. Phenanthrene is oxidized by
CYP1A1 and 1B1 to an epoxide (represented by Phen 1,2 -epoxide –the cyclic ether with the 3-
ring atoms, commonly referred to as an aromatic epoxide or arene oxide or
oxirane)(Anzenbacher, P et al., 2012). EHs are hydrolytic enzymes that use water as a cofactor
but instead of breaking the substrate apart like other hydrolytic enzymes, they add water to the
substrate (Phen 1,2- epoxide) producing a dihydrodiol (represented by Phen1,2 diol in Figure 2).

15
These diols are much less reactive, thus the actions of EHs are generally accepted as
detoxification. The diols usually spontaneously rearrange to phenols from the arene oxides
(Anzenbacher, P et al., 2012; ATSDR, 2005). However, the diols can be re-oxidized by CYP1A1
and 1B1 to a more reactive dihydrol expoxide (anti-PheDE). The dihydrodiol epoxides react
with DNA forming adducts during Phase I metabolism. Detoxification of PAH dihydrodiol
epoxides occur in Phase II metabolism predominantly through glutathione (GSH) conjugation
(Jernstrom, B et al., 1996; Romert, L et al., 1989). GSH is a ubiquitous antioxidant compound,
present in cells, and composed of glycine, cysteine and glutamic acid. GSH xenobiotic
conjugation is catalyzed by the glutathione S-transferase (GST) family of enzymes, which
include the isoforms alpha, pi, mu, omega and theta, which comprise about 10% of the total
cellular protein (Klaassen, C et al., 2008). Pi, Alpha, and Mu class human GSTs enzymes have
been reported to be active in catalyzing GSH conjugation of PAH dihydrodiol epoxides
(Jernstrom, B et al., 1996; Seidel, A et al. 1998).
Fluorene is a tricyclic PAH examined in this thesis. Fluorene along with other tricyclic PAH
parent compounds has been shown to induce the CYP system. A study showed that tricyclic
PAHs induced hepatic CYP1A2 gene expression in B6C3F1 mice by an AHR-independent
pathway (Chaloupka, K et al., 1994). The CYP1A2 protein is the major CYP1A inducible
isoenzyme in humans and preferentially catalyzes the oxidative metabolism of diverse
substrates, including methoxyresorufin (O-dealkylation) and 17 beta-estradiol (E2) (2-
hydroxylation)(Chaloupka, K et al., 1994; Okey, AB 1990). The study by Chaloupka, et al.,
showed that all six tricyclic PAHs tested significantly induced hepatic microsomal
methoxyresorufin 0-deethylase (MROD) activity in B6C3F1 mice. CYP1A2 was induced by the

16
reconstituted 3-ring PAH mixture as well, which contained acenaphthene, acenaphthylene,
anthracene, phenanthrene, fluorene, and dibenzofuran (38%)(Chaloupka, K et al., 1994). These
findings implicate PAHs as ovotoxicants through induction of CYP1A enzymes independent of
the AHR pathway discussed above in reference to larger PAHs (> 4 rings) such as 3-MC (that
bind the AHR to induce CYP1A1 and CYP1A2 gene expression and ovotoxicity in rodents)(0key,
AB 1990). Estradiol (E2) oxidation by enzyme induction by PAHs in this manner can also result in
lower levels of E2 in women leading to irregular ovulation and/or anovulatory cycles (see
discussion on the menstrual cycle).
Half-lives for OH-PAHs have been reported in multiple human studies. PAHs in general
have short half-lives (hours to a couple of days)(ATSDR, 1995; Li, Z et al. 2008). Motorykin et al.,
2015 reported a half-life for the parent compound pyrene as 3.3 hours(h) with overall half-lives
of the OH-PAHs being higher with a range of 1.7 h for FLUO9 to 7 h for FLUO3. Li reported the
half-lives of NAP2 and FLUO3 to be 2.5 h and 6.1 h, respectively (Li, Z et al., 2012). Buchet
reported the half-life of PYR1 as 18 h (95% CI : 16.1 h, 19.8 h)(Buchet, JP et al., 1992), whereas
Buckley and Lioy estimated it to be 4.4 h (3.1 h to 5.9 h)(Buckley, J et al., 1992). Smith's group
estimated the half-lives of NAP1 and NAP2 as 13.6h (95% CI: 9.5h, 24.0 h) and 15.9 h (95% CI:
12.0 h, 23.8 h) respectively (Smith, KW et al., 2012). Li, Z et al., in 2012 reported median half-
lives with confidence intervals for all of the OH-PAHs that will be examined in this thesis. See
Table 1.
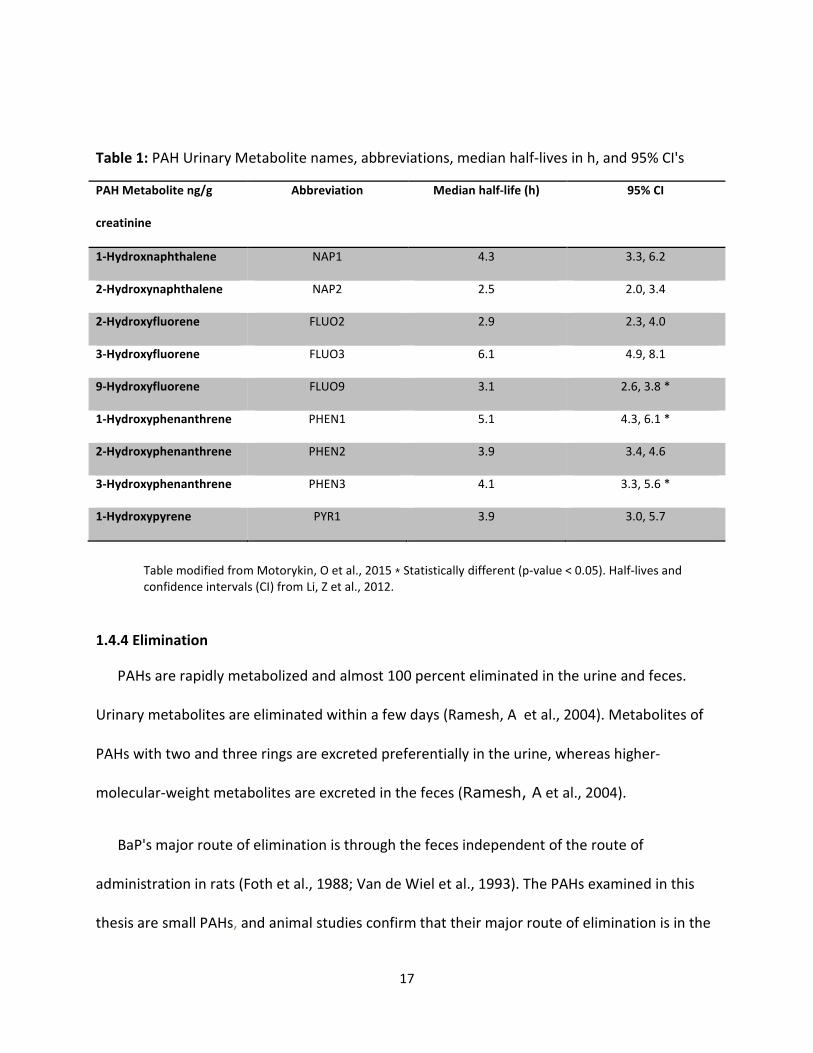
17
Table 1: PAH Urinary Metabolite names, abbreviations, median half-lives in h, and 95% CI's
PAH Metabolite ng/g
creatinine
Abbreviation Median half-life (h) 95% CI
1-Hydroxnaphthalene NAP1 4.3 3.3, 6.2
2-Hydroxynaphthalene NAP2 2.5 2.0, 3.4
2-Hydroxyfluorene FLUO2 2.9 2.3, 4.0
3-Hydroxyfluorene FLUO3 6.1 4.9, 8.1
9-Hydroxyfluorene FLUO9 3.1 2.6, 3.8 *
1-Hydroxyphenanthrene PHEN1 5.1 4.3, 6.1 *
2-Hydroxyphenanthrene PHEN2 3.9 3.4, 4.6
3-Hydroxyphenanthrene PHEN3 4.1 3.3, 5.6 *
1-Hydroxypyrene PYR1 3.9 3.0, 5.7
Table modified from Motorykin, O et al., 2015 ⁎ Statistically different (p-value < 0.05). Half-lives and
confidence intervals (CI) from Li, Z et al., 2012.
1.4.4 Elimination
PAHs are rapidly metabolized and almost 100 percent eliminated in the urine and feces.
Urinary metabolites are eliminated within a few days (Ramesh, A et al., 2004). Metabolites of
PAHs with two and three rings are excreted preferentially in the urine, whereas higher-
molecular-weight metabolites are excreted in the feces (Ramesh, A et al., 2004).
BaP's major route of elimination is through the feces independent of the route of
administration in rats (Foth et al., 1988; Van de Wiel et al., 1993). The PAHs examined in this
thesis are small PAHs, and animal studies confirm that their major route of elimination is in the

18
urine. Animal studies of administered naphthalene for example, showed it is eliminated as
metabolites in the urine: 77–93% in urine, 6–7% in feces following oral administration (Bakke, J
et al. 1985), and 70–87% in urine, 6–14% in expired air, 2–4% in feces following dermal
administration (Turkall, RM et al. 1994). Bakke et al. (1985) recovered 7–20% of an
administered dose as NAP1 and NAP2 in rats’ urine. Kilanowicz et al., (1999) also reported that
over 88% of intraperitoneally administered naphthalene was eliminated in urine during the first
72 h in rats. Maximum levels in plasma were observed at the second hour post-administration,
following a biphasic decline with half-lives of 0.8 h in phase I and 99 h in phase II. These data
are largely confirmed in human studies involving urinary NAP1 excretion (1 to 2 h in phase I and
14 to 46 h in phase II) (Heikkila , P et al., 1995). Additionally, Bieniek (1994) determined a mean
urinary excretion rate of 57 mg/h, with a half-life of approximately 4 h in occupationally
exposed workers employed in distillation of naphthalene oil.
Excretion kinetics involving carbon labeled PAHs (Grova, N et al., 2002) showed that
phenanthrene and pyrene behaved identically, with low milk transfer and high excretion in the
urine 103 h after a single oral ingestion in lactating goats. The feces excretion kinetics
supported Foth and Van de Wiel's finding for BaP, showing that 88.2% of the ingested BaP
radioactivity was recovered in the feces. The radioactivity detected in feces due to
phenanthrene and pyrene were 21.7% and 25.5%, respectively, further supporting their major
urinary excretory route.
Animal and human studies demonstrate that fluorene is also predominantly, if not totally,
eliminated in the urine. Dewhurst, F et al., (1962) reported that fluorene resembled

19
naphthalene in its metabolism as it is almost completely absorbed in the gut and its metabolites
excreted mainly, if not entirely, in the urine of rats and rabbits. Further, as far back as 1948,
Neish reported finding metabolites obtained from the urine of rabbits after ingesting fluorene
were FLUO2 and the glucuronide of this phenol. He also concluded from his study that little if
any unchanged fluorene was excreted in the feces, which was in agreement with prior studies
dating 5 years back (Neish, WJ, 1948). The data on excretion of PAHs has remained consistent
over the years.
Motorykin et al., (2015), reported post-inhalation excretion rates of 4 parent PAHs and 10
OH-PAHs during the smoking of salmon, and Santiago-Delago, L et al.(2015) reported on
excretion of the same compounds after the consumption of smoked salmon. The maximum
concentration excretions were higher for ingestion exposure compare to inhalation for most of
the metabolites (Motorykin, O et al., 2015).
1.4.5 PAH-induced Ovarian Toxicity
PAH-induced ovotoxicity has been assessed in epidemiology studies of women smokers over
the past 5 decades. It has long been documented that women smokers experience a 1 to 4 year
earlier age onset of menopause (Jick H et al., 1977). Multiple epidemiology studies reveal a
strong association between cigarette smoking and impaired fertility (Mark-Kappeler CJ et al.,
2011). The risk factors for premature ovarian failure( POF) established to date include rare
autosomal mutations, disorders of the X-chromosome, treatment with anti-neoplastic drugs,
pelvic ionizing radiation, and cigarette smoking. Animal studies have been relied upon to
confirm PAH-induced primordial follicle depletion which is the true marker of ovarian reserve,

20
because it is not possible to retrieve primordial follicles from humans without destroying their
ovaries. Mattison, DR et al., found that the diol epoxide metabolites of BaP were the ovotoxic
metabolites in B6 and F1 mice, decreasing ovarian primordial and primary follicle counts
(Mattison, et al., 1989). Higher animal species such as rhesus monkey have demonstrated
gonadotropin-dependent metabolism of DMBA in the ovary as well (Bengtsson, M et al., 1989).
DMBA-induced primordial and primary follicle destruction were later illustrated in a dose and
time-dependent manner in grafted human ovarian tissues (Matikainen, T et al. 2001). The
evidence that the mechanism of PAH-induced ovotoxicity involves CYP metabolism was further
confirmed when GSH was shown to prevent DMBA-induced apoptosis and the rise in reactive
oxygen species (ROS) that occurs prior to apoptosis in cultured pre-ovulatory rat follicles (Tsai-
Turton, M et al., 2007). Finally, Archibong illustrated that inhaled BaP dose-dependently-
induced ovarian endocrine disruption in F-344 adult rats prior to mating. The results revealed
decreased ovulation rates and number of pups per litter in BaP exposed rats at low dose
compared to controls. The E2 and luteinizing hormone (LH) plasma levels at proestrus were also
significantly reduced. Finally, the activity of the AHR activity in ovarian and liver tissues as well
as the concentrations of BaP 7,8-diol and 3,6-dione metabolites increased in an exposure
concentration-dependent manner (Archibong, E et al., 2012).
Epidemiology studies have examined PAHs with regard to female fertility and in vitro
fertilization assessments and have supported prior animal and other epidemiology studies
implicating PAH-induced female suboptimal fertility. Neal found BaP in the ovarian follicular
fluid of women who did not conceive to be significantly higher than that of women who did
conceive during IVF (Neal, M et al., 2008). Cigarette smoke has been shown in animal studies to

21
destroy primordial follicles. Tuttle documented mainstream smoke exposure in mice and
reported cigarette smoke induced a significant reduction in the number of primordial follicles
(Tuttle, AM et al., 2009). Finally, PAH-induced ovotoxicty has been established for decades, yet
research to advance this knowledge for clinical application is still lacking. There is an urgent
need to support the use of this knowledge to help mitigate fertility problems in women.
Bhattachary states " it is well-known that ovaries express xenobiotic bioactivating and
detoxifying enzymes, yet the knowledge gaps remain for established ovotoxicants" , which
acknowledges the need for studies like the current study (Bhattachary, P et al., 2012).
1.4.6 Effects of Genetic Polymorphism on PAH Metabolism
Genetic polymorphisms in metabolic enzymes, such as the CYP family, are known to
influence individual metabolism of xenobiotic compounds (Strickland, P et al., 1999). Genetic
polymorphisms in potential PAH metabolizing phase I and II enzymes have been shown to
contribute to inter-individual differences in urine concentration of PAH metabolites in
occupationally and non-occupationally exposed populations.
Multiple human studies have identified many genetic polymorphisms in CYP 1A1 and 1B1,
some of which have been linked with susceptibilities to lung and breast cancer, however race-
related differences in occurrence of genetic polymorphisms in these CYP species make it
difficult to interpret these studies as there are also reports of no relationship of polymorphisms
of CYP1A1 and 1B1 to cancer susceptibility (Nakachi, K et al., 1993; Watanabe, J et al., 2000;
Kelsey, KT et al., 1994; Nebert, DW et al., 1999). Abnet et al., investigated multiple
polymorphisms, including CYP1A1 and CYP1B1 polymorphisms, on the urinary PYR1 glucuronide
(PYR1G) concentrations in 199 healthy subjects and found CYP1B1 to have a strong association

22
with PYR1G concentrations in that population and a modest but significant associations with
one variant in each of the AHR, CYP1A1 and CYP1A2 genes (Abnet, CC et al., 2007). Another
study evaluated the correlation between external PAH exposure and biomarkers of exposure
and investigated to what extent genetic polymorphisms in metabolic enzymes could explain
inter-individual variation in PYR1 levels. They found that the part of the variance that could be
explained by differences in biotransformation genotypes seemed to be of the same order of
magnitude as the variance explained by differences in exposure. They concluded that PYR1, in
addition to being a sensitive indicator of recent human exposure to PAHs, may also to some
extent reflect the inter-individual variation in susceptibility to PAHs (Alexandrie, AK et al.,
2000). Yang et al., reported polymorphisms in metabolic enzymes CYP2E1, GSTM1, and N-
acetyltransferase (NAT2) influenced urinary naphthol levels and also suggested that the GSTT1
null genetic polymorphism has the potential to affect the biological monitoring of PAHs with
PYR1, and might act as a genetic factor in PAH-related toxicity in subjects who did not have
occupational PAH exposure (Yang, M et al., 1999).
Inter-individual variability in the excretion of urinary OH-PAHs has been shown with
different GSTM1 and GSTT1 genotypes. Hong's group found in their study with bus drivers, that
the excretion of PYR1G in GSTM1-deficient smokers was higher than that in GSTM1-positive
smokers. In addition, PYR1G excretion was higher in GSTT1-positive smokers than in GSTT1-
deficient smokers (Hong, YC et al., 1999). Another group found significantly increased PYR1
levels (p <0.001) in PAH occupationally exposed coke oven workers but no occupational
induction of the effect biomarkers (DNA strand breaks for example) after adjustment for
cigarette smoking. Their multi-variate analysis did not show a significant influence of GSTM1

23
and GSTT1 polymorphisms on any of their biomarkers (van Delft, JH et al., 2001). The later
finding was consistent with Kawamoto's group's report of no effect of polymorphisms in
CYP1A1 or GSTM1 on urinary PYR1 (Kawamoto, T et al., 1998). More recently, Petchpoung's
group found that urinary PYR1 concentrations were associated with exposure to air pollution,
cigarette smoking and polymorphisms of CYP1A1 and GSTM1 but not with polymorphisms in
GSTT1 (Petchpoung, K et al., 2011). Finally, the literature is mixed with regards to different
ethnic/racial groups, regarding the importance of polymorphisms in PAH metabolism. For
example, as opposed to the findings of Hong's group cited earlier, Lee's group found that
smoking had a more profound effect on urinary OH-PAH metabolites than did genetic
polymorphism in Koreans. They investigated Korean aircraft maintenance workers' genetic
polymorphisms in CYP1A1 , CYP2E1, and GSTM1 and GSTT1 and their effects on PYR1 and NAP2
levels as well as estimated their levels of exposures to PAHs (Lee, CY et al., 2001). Nan's group
examined the same polymorphisms in Korean coke oven workers and Korean university
students, and found that In the occupationally exposed population, CYP2E1 and GSTM1
appeared to play an important role in the metabolism of pyrene and naphthalene. However, in
individuals that were not occupationally exposed to PAHs, GSTM1 and smoking seemed to
influence the urinary concentration of NAP2 (Nan, HM et al., 2001). Lastly, there are also
differences in gene allele frequencies within races such as those found among Asians. Garte's
group found a significant difference between Japanese and other Asians for GSTM1 and GSTT1,
with the Japanese showing lower frequencies of both deletions as well as major and significant
differences in frequencies between Caucasians for these deletions (Garte, S et al., 2001). All of
these findings indicate that more work needs to be done in order to understand the role of

24
polymorphisms in individuals' PAH metabolism and ultimate susceptibility to PAH-induced
ovotoxicity.
1.5 PAH Biomonitoring
Table 2 illustrates PAH parent compounds, their structures, and urinary hydroxylated
metabolite names.
Table 2: PAH Parent Compounds and OH-PAH metabolites evaluated in this thesis
PAH Parent PAH Parent Structure Metabolite (s)
Naphthalene
1-Hydroxynapthalene
2-Hydroxynapthalene
Fluorene
2-Hydroxyfluorene
3-Hydroxyfluorene
9-Hydroxyfluorene
Phenanthrene
1-Hydroxyphenanthrene
2-Hydroxyphenanthrene
3-Hydroxyphenanthrene
4-Hydroxyphenanthrene*
Pyrene
1-Hydroxypyrene
PAHs in the National Bio-monitoring Program. * Not measured in this thesis.

25
Specific PAHs evaluated in this thesis are hydroxylated metabolites of small PAHs. The best
studied PAHs (DMBA, 3-MC, and BaP) in terms of ovarian toxicity have been established in
multiple investigations to destroy primordial follicles in laboratory animals. There is sufficient
evidence for using small urinary OH-PAHs as biomarkers of exposure to larger PAHs because
larger PAH metabolites such as BaP metabolites have been found in urine samples along with
the urinary OH-PAH metabolites involved in this thesis in several studies. Buratti, M et al.,
detected 3-hydroxybenzo[a]pyrene in spot urine samples containing 15 inhaled priority PAHs
that included FLUO2, NAP2, PHEN3, and PYR1 collected at three different times during a work
week from asphalt workers (Buratti, M et al., 2007). Moreover, small urinary metabolites OH-
PAHs used in this thesis have been found to represent inhalation exposure to BaP in multiple
studies. Merlo's group investigated the relationships between exposure to ambient air PAHs
and PYR1 and indoor air pollution using BaP air measurements as an index compound of PAH
levels in the air in traffic police officers and indoor subjects respectively. Stratified analysis
showed higher levels of PYR1 in the urine of those who smoked 15 or less cigarettes per day
and who carried the CYP1A1 polymorphism relative to those who didn't (Merlo, F et al., 1998).
Finally, since large PAHs are mostly eliminated in the feces, they are impractical in terms of
specimen collection in human studies.
A number of techniques have been developed for the biological monitoring of human
exposures to PAHs. Among them are PAHs and their metabolites in blood (serum) and urine,
measurement of mutagenicity in urine and feces, chromosome aberrations and sister
chromatid exchanges in peripheral blood lymphocytes, and DNA and protein adduct
measurement in the serum and in other tissues (Larsen, JC et al., 1995). The exchange of sister

26
chromatids, chromosomal aberrations, and mutagenicity of urine and feces have poor
specificity for PAHs (Hatijan, BA et al., 1995; Popp, W et al., 1995; Perera, FP et al., 1994;
Santella, RM et al., 1994). High performance liquid chromatography (HPLC) was shown in the
past to be more specific in the detection of PAH-DNA adducts than enzyme-linked
immunosorbent assay (ELISA), however, it was concluded that most of the analytical methods
used to determine PAH-DNA adducts were too nonspecific to produce accurate and comparable
results (Angerer, J et al., 1997). Ferreira, et al., examined employees at a graphite electrode
production company and in a foundry for internal doses of PAHs by measuring protein adducts
from hemoglobin in an attempt to avoid measurement errors introduced by DNA repair
mechanism found in prior studies using DNA-adducts. His group found increasing adduct
concentrations and increased diagnostic sensitivity, but found a weak correlation coefficient
(0.26) (although a statistically significant relationship) between the external exposure (sum of
13 PAHs) and the internal dose (level of hemoglobin adducts) of PAHs (Ferreira Júnior, MF et al.,
1994). Tas, S et al., assessed the risk of exposure to PAHs in steel foundries and a graphite
electrode producing plant using albumin and HPLC. They found that the relationship between
the BaP-diol epoxide adducts to albumin (BPDE-alb) and the intensity of exposure from
airborne personal sampling to PAHs and PYR1 excretion in post-shift urine to be significantly
higher in exposed workers compared to controls. However, there was a wide inter-individual
variation between subjects' adduct levels with the same level of occupational exposure
measured with airborne personal sampling. Therefore they concluded that as long as BPDE-
protein adduct levels had not been demonstrated to be better related to the risk of adverse
genotoxic effects caused by PAHs in target organs than the PYR1 level in urine, the latter

27
remained the most practical biological parameter for assessing the risk of exposure to PAHs
(Tas, S et al., 1994). The sensitivity and specificity issues of the serum and urine biomarkers
persisted throughout the literature over the years and helped shape the current preferred
methods of biomonitoring of PAH exposure in humans.
Metabolites of parent PAHs smaller than pyrene are usually excreted in the urine in the
form of OH-PAHs, facilitating their effectiveness as biomarkers of PAH exposure (Dor, F et al.
1999; Li, Z et al. 2008). PYR1 has been found to be a good biomarker of exposure in both
environmental and occupational studies (ATSDR, 1995; Li, Z et al. 2008). Urinary PAH
metabolite biomonitoring of PYR1 has proven to be sensitive and reliable in many studies
(Gerde P et al., 2001; Campo, L et al., 2006; McClean, MD et al., 2004;IPCS, 1998).
Biomonitoring of OH-PAHs in the US is part of an ongoing effort to reduce and monitor
particulate matter in air pollution. Asphalt is a source of occupational and environmental
particulate matter through bitumen fumes. PYR1 has been useful in the study of high
occupational exposure to bitumen fumes produced when it is stored and handled at
temperatures up to 200-230 Centigrade (C) (Mc Clean, MD et al., 2004). Historically, the relative
ease of measurement of PYR1 and its' prevalence in human urine have been instrumental in
epidemiological studies aimed at illustrating the need for biomarkers of fine particle exposure.
Some of those studies showed apparent associations between ambient particulate matter
concentrations and health effects among adults. They also illustrated the insufficiency of the
current regulation for particulate matter levels in the US and other countries that address the
soot and sulphur dioxide from the 1950's, at preventing respiratory adverse health effects and

28
mortality (Pope, CA 1989; Schwartz, J et al., 1992; Dockery, DW et al., 1993; Pope, CA et al.,
1995). Since the early 2000's, the Centers for Disease Control and Prevention's NHANES has
used urinary OH-PAH concentrations to establish reference range concentrations for the US
population, and to set benchmarks for future epidemiologic and biomonitoring studies (Li, Z et
al., 2008). The survey focuses on the PAHs of concern to IARC, the NTP, and on the ASTDR's and
USEPA's NPL.
Pyrene is 90% metabolized into PYR1 and it has a high concentration in all mixtures of PAHs
(Cattaneo, A et al., 2010). There are positive correlations between external exposure to pyrene
and its metabolite PYR1 (Cattaneo, A et al., 2010). The NHANES reports on urinary OH-PAH's
from samples of thousands of people including children, adolescents and adult in the US
population in two year intervals, which support these statements. The NHANES reports are a
reliable assessment of human exposure and excretory route. The report from 2001 and 2002, of
22 OH-PAH urinary metabolites showed detectable levels ranging from nearly 100% for the
metabolites of naphthalene, fluorene, phenanthrene, and pyrene, to less than 5% for
metabolites from parent compounds with higher molecular weight such as chrysene,
benzo[c]phenanthrene, and benz[a]anthracene (Li, Z, et al., 2008), which supports the prior
sized-based statements regarding major routes of excretions for PAHs. Further, the geometric
mean for PYR1—the most commonly used biomarker for PAH exposure—was 49.6 ng/L urine,
or 46.4 ng/g creatinine in that study. These authors found that children (ages 6–11) generally
have higher levels than adolescents (ages 12–19) or adults (ages 20 and older). Their model-
adjusted, least-square geometric means for PYR1 were 87, 53 and 43 ng/L for children,
adolescents (ages 12–19) and adults (ages 20 years and older), respectively. Log-transformed

29
concentrations for major detectable OH-PAHs were significantly correlated with each other.
The correlation coefficients between PYR1 and other metabolites ranged from 0.17 to 0.63 (Li, Z
et al., 2008), which illustrates that PYR1 is correlated with some PAHs more than others.
Motorykin, O et al., illustrated the usefulness of the NHANES reference values in their study of
the metabolism and excretion rates of different OH- PAHs after oral and inhalation of PAHs,
when they used the NHANES adult geometric means from the 2008 tables for urinary creatinine
adjusted OH-PAHs to compare to their values from study participants. Their baseline values
were in the range of the 25th to 75th percentile of NHANES concentrations (Motorykin, O et al.,
2015).
Examples of other studies using PAH urinary metabolites as the main biomonitoring
measurement tool include a controlled human exposure study for the significance of ingested
PAHs in addition to occupational exposures to PAHs. The investigators used urinary metabolites
to demonstrate that diet in addition to occupational exposure need to be considered in
exposure assessments of PAHs (Buckely, TJ et al., 1992). Urinary excretion of multiple PAH
metabolite biomarkers were used to assess the exposure of soil remediation workers on a
former creosote wood impregnation site polluted with creosote oil. The PAH biomarker data
(concentrations and diurnal excretion profiles) showed significant work-related exposure in
both non-smoking and smoking participants (Elovaara, E et al., 2006).
There are inter and intra-individual PAH concentration variability throughout the day.
Studies using urinary PAH metabolites as biomarkers of exposure have used 24-hour composite
urine samples as well as first morning void measurements in occupational and environmental

30
studies to represent daily PAH exposure. The internal validity of these studies have been
adversely affected by 24 hour urine collection attempts due to compliance issues involving
forgetfulness, inconvenience, and misplaced samples or poor sample container capacities,
contamination during storage, or loss of urine (Garde, AH et al., 2004). Han, IK et al. (2008),
investigated whether the convenience of using the first morning voids for the PYR1
concentrations can reflect the daily average PYR1 concentrations. They also investigated
whether creatinine adjustment could reduce the inter and/or intra-individual differences
between the first morning urine and 24-h composite urine measurements, since historically
researchers have used it to adjust for the large inter and intra-individual urine output volume
rates. This method is still controversial because diurnal variation in creatinine excretion also
varies with intake of meat, age, gender, and other factors (Boeniger, MF et al., 1993; Barr, DB et
al., 2005). Han reported adult coke oven workers versus non-coke oven workers' significant
(40% to 62%) intra-individual differences between the first morning voids and 24-h PYR1
concentrations. The first morning PYR1 was substantially correlated with 24-h PYR1 and
appeared to discriminate occupationally exposed workers from those not so exposed.
Creatinine adjustments of PYR1 concentrations reduced the intra-individual difference between
first morning and 24-h composite urine measurements by approximately 10% but did not
significantly improve overall correlations between the two measurements, although winter 24-
h urine and spot urine data correlated higher than fall data (Han, IK et al., 2008). Urinary OH-
PAH biomonitoring is the measurement tool of exposure in this thesis. Inter and intra-individual
differences in PAH urinary metabolite concentrations will not pose a methods concern because
the endpoint of analysis is the average of measurements taken over a six month period in this

31
convenience sample where the interest is in assessment of the long-term effects of PAHs on
ovarian reserve markers.
2.0 Female Reproductive Physiology
2.1 Hypothalamic-Pituitary Ovarian Axis
Environmental and internal stimuli trigger gonadotropin releasing hormone (GnRH), a
decapeptide, synthesized and released by specific neuronal endings in the anterior and
mediobasal hypothalamus in a constant pulsatile pattern (Homburg, R, 2014). GnRH reaches
the anterior pituitary gland through portal vessels that connect the hypothalamus to the
anterior pituitary gland (Devoto, L et al., 2000). In response, the anterior pituitary gland
releases FSH and LH (gonadotropins), which act on the ovarian follicle cells (Homburg, R, 2014).
FSH and LH stimulate the ovary to produce the sex steroids (E2 and progesterone) (Dekker, M
1999; Homburg, R, 2014). Inhibins are hormones secreted by the granulosa cells of small
ovarian follicles that selectively inhibit the anterior pituitary FSH secretion (Welt, CK et al.,
2001; Homburg, R, 2014). Inhibin A and inhibin B dimers differ in their pattern of secretion.
Inhibin A concentrations are low during most of the follicular phase of the menstrual cycle, but
start to rise during its latest stages and peak in the mid-luteal phase. In contrast, inhibin B
concentrations start rising early in the follicular phase, paralleling but lagging behind the FSH
rise (Homburg, R, 2014). Inhibin B reflects the size of the follicle cohort (Homburg, R, 2014).
During the menstrual cycle, the effect of inhibins, E2 and progesterone (on the hypothalamus
and anterior pituitary) are mainly inhibitory (Homburg, R, 2014). However, when E2 surpasses a
threshold level, its feedback switches from negative to positive on the hypothalamus and
anterior pituitary gland (Homburg, R, 2014). FSH stimulates the growth of antral follicles,

32
inhibin synthesis, E2 production, as well as granulosa cell proliferation and differentiation in the
ovary (Homburg, R, 2014; Boundless, 2013).
2.2 Menstrual Cycle
The menstrual cycle lasts from the onset of menses (the first day) until the next onset of
menses (Boundless, 2013). During the follicular phase (days 5-15) basal secretion of LH and FSH
occur, and E2 secretion predominates (Homburg, R, 2014). E2 secretion, by the large ovarian
follicles, increases gradually throughout the first 11 days of the cycle. As days 13 to 15
approach, there is a dramatic rise in estrogen followed by an LH surge (upon which meiosis is
resumed in the oocyte), and to a lesser extent an FSH surge, which trigger the dominant follicle
to ovulate (Adashi, EY et al., 1994; Boundless, 2013; Homburg, R, 2014). After ovulation, the
dominant follicle reorganizes to produce the corpus luteum (Adashi, EY et al., 1994). The
possibility of fertilization of the ovum and implantation takes place about day 21 , after which
the luteal phase ends with return of basal secretion of FSH and LH. The corpus luteum grows
and secretes progesterones after ovulation to prepare the uterus for implantation of a fertilized
ovum and to maintain pregnancy (Boundless 2013). If fertilization does not occur, the corpus
luteum spontaneously regresses (Adashi, EY et al., 1994) which results in menstruation, and the
negative feedback effects of E2, progesterone and inhibin A on FSH secretion are suddenly lost
so that FSH is secreted in relatively large quantities during menstruation itself. The granulosa
cells in the ovary have FSH receptors and the theca cells in the ovary have LH receptors
(Homburg, R, 2014).
At the beginning or menses phase of each cycle, a group of the most mature follicles (2–5
mm in diameter) is recruited for further growth, granulosa cell differentiation and

33
multiplication (Homburg, R, 2014). The follicles more sensitive to FSH rather than those less
mature are selected for further development at the time of the FSH inter-cycle rise (Homburg,
R, 2014). The follicle most sensitive to FSH will ovulate and use FSH to increase aromatase
activity and produce E2s and inhibin B (Homburg, R, 2014). Inhibin B is secreted from the small
and large antral follicles. As FSH concentrations fall in response to rising E2 and inhibin B levels
it becomes less available and only the most sensitive follicle with the lowest threshold for a
response to FSH, survives and continues to thrive and produce the most E2 and LH receptors.
The induction of LH receptors on the largest developing follicle(s) (>10mm diameter) allows
response to LH, which leads to E2 production and follicular growth in the mid-late follicular
phase. It also enables LH to have a role in the development of the dominant follicle in the late
follicular phase and prepare it for the oncoming LH surge (Homburg, R, 2014).
2.3 Folliculogenesis
At the follicular level, the primordial follicles (small preantral follicle) are quiescent and
arrested in the diplotene stage of the first meiotic prophase (in second trimester of pregnancy
and onward)( Zheng, W et al., 2012; Juengal, JL et al., 2005). They have a single incomplete flat
cell layer of granulosa cells. Throughout life, primordial follicles continuously enter the growing
pool, and a small fraction of these recruited follicles progress successfully through all stages of
maturation, (primordial, primary, secondary (large preantral), antral, preovulatory/large antral)
to the ovulatory stage (Parrot, JA et al., 1999; Otsuka, F et al., 2011). As the follicles mature
their granulosa cells differentiate and become cuboidal and proliferate. Once there is a single
layer of granulosa cells they are called primary follicles. When there is more than one layer,
they are called secondary follicles. At this point a basement membrane is formed to separate

34
the theca cells from the granulosa cells (Zoller, LC 1991). The first meiotic division of the oocyte
is completed as a result of the preovulatory LH surge ( Boundless, 2013). AMH, a dimeric
glycoprotein and member of the transforming growth factor-beta family (Homburg, R, 2014), is
secreted by the granulosa cells of the primary, secondary, and small antral follicles (van Houten,
EL et al., 2010). See Figure 3.
Folliculogenesis is under paracrine regulation. AMH is gonadotropin-independent (Iorio, R et
al., 2014) and regulates folliculogenesis at the two extremes (Homburg, R, 2014; Durlinger, AL
et al., 2002). First, it restricts the progress of developing primordial follicles (Durlinger, AL et al.,
1999) and second it inhibits the sensitivity of antral follicles to FSH (in selection of the dominant
follicle). It also inhibits aromatase activity that catalyzes E2 production during an ovulatory
cycle (Durlinger, AL et al., 2002; Homburg, R, 2014). As previously mentioned, the first meiosis
is completed just before ovulation. The oocyte only completes the second meiosis if it is
fertilized (Zoller, LC 1991). Because of the established roles of AMH and the fact that it is
gonadotropin-independent, it serves as a reliable indirect marker of the number of small antral
follicles in a cohort available for ovarian stimulation- a very sensitive marker of ovarian reserve
irrespective of age (Homburg, R, 2014). See Figure 3 for a pictorial representation of the ovarian
follicle stages that secrete the markers of ovarian reserve evaluated in this thesis (AMH, inhibin
B, and FSH). It illustrates the selective activity of AMH, FSH, and inhibin B on folliculogenesis
and the author's proposal that Inhibin B may have a paracrine positive influence on
folliculogenesis (Hillier, SG 1991, Findlay, JK et al., 2000, Visser et al. 2006, Broekmans, FJ et al.
2008).
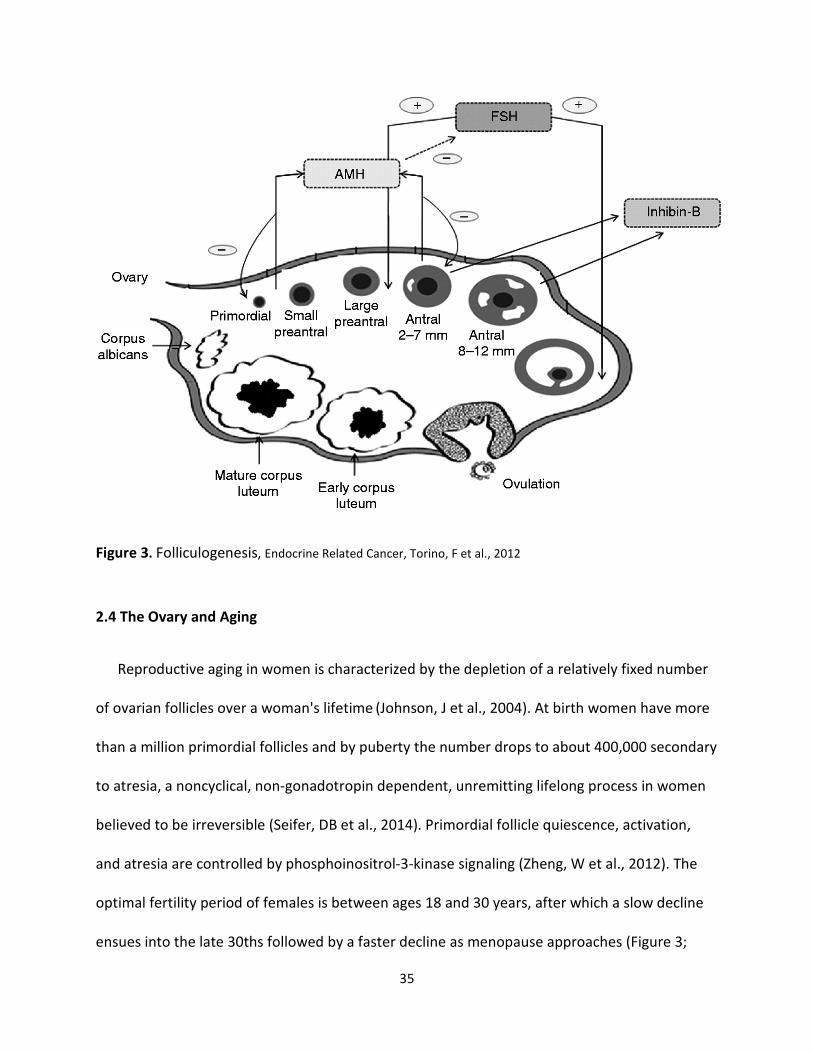
35
Figure 3. Folliculogenesis, Endocrine Related Cancer, Torino, F et al., 2012
2.4 The Ovary and Aging
Reproductive aging in women is characterized by the depletion of a relatively fixed number
of ovarian follicles over a woman's lifetime (Johnson, J et al., 2004). At birth women have more
than a million primordial follicles and by puberty the number drops to about 400,000 secondary
to atresia, a noncyclical, non-gonadotropin dependent, unremitting lifelong process in women
believed to be irreversible (Seifer, DB et al., 2014). Primordial follicle quiescence, activation,
and atresia are controlled by phosphoinositrol-3-kinase signaling (Zheng, W et al., 2012). The
optimal fertility period of females is between ages 18 and 30 years, after which a slow decline
ensues into the late 30ths followed by a faster decline as menopause approaches (Figure 3;

36
Hansen, KR et al., 2008). The average age of menopause is 50 - 51 years (range 45 to 59) (Gold,
EB et al., 2001). At menopause, the follicle count becomes essentially zero. Assuming fixed time
differences between the reproductive milestones perimenopause (the time period when
ovulation and menses becomes irregular) and menopause, fertility will not be lost completely
during that time period which lasts about for 4 years on average (Nikolaou, D et al., 2003).
2.5 Ovarian Reserve
Ovarian reserve refers to the number and quality of remaining primordial follicles, which is
the measure of a women's reproductive potential or fecundity. Clinically, diminished ovarian
reserve (DOR) has been defined as reduced response to ovarian stimulation in women of
reproductive age with regular menses and without any other anatomical or other non-
hormonal reason preventing the ability to reproduce (Scott, RT et al., 1995). It indicates that
there is a low number and/or impaired development of primordial follicles. Women with
markedly DOR have low chances of conception with their own gametes, even with assisted
reproductive technologies (Sharara, FI et al., 1998). Diminishing ovarian reserve is normal
perimenopausally, and happens in women during their mid to late thirties and at times earlier
(Scott, RT et al., 1995). In 2002, 7.4 % of married women in the US experienced some type of
infertility (Stephen, EH et al., 2006). Specifically 10.9% of US women aged 15 to 44 years, which
is within the age range of the thesis study participants, experience impaired fecundity based on
data from 2006 to 2010 (CDC, 2015). Premature ovarian failure (POF) affects 1% of the general
population (Coulam, CB et al., 1986) and refers to women who reach menopause before the
age of 40 by means other than medical intervention (Coulam, CB et al., 1982). The women (with

37
intact, functioning ovaries and no other known reason for delayed conception) who don't meet
the criteria for POF are classified as having DOR (Scott, RT et al., 1995).
2.6 Markers of Ovarian Reserve
As indicated earlier, true ovarian reserve can only be measured with primordial follicle
count, which is impossible in humans without destroying the ovary. Therefore, indirect
measures of ovarian reserve are relied upon and involve measurements of the hormones
involved in folliculogenesis discussed above for assessment of ovarian reserve both in research
and in clinical practice in humans. Serum levels of FSH, AMH, inhibin B, and E2 are all used in
assessment of ovarian reserve. Measurements of antral follicle count (AFC) with ultrasound is
also used in clinical assessments of ovarian reserve. The ovarian reserve markers that are
examined in this thesis (AMH, inhibin B, and FSH) will be the focus of the remainder of this
discussion. Low levels of inhibin B indicates a reduction in the small and large antral follicle
count and have been used as a test of functional reserve of ovaries to facilitate prediction of
low response to in vitro fertilization standard protocols (Sharara, FI et al., 1998). Clinically, an
increase in a women's basal FSH levels measured on day 3 of the menstrual cycle occurs due to
follicle depletion and is used to classify women as having DOR (Scott, RT et al., 1995; Karande,
V et al., 1999) and poor oocyte quality (Karande, V et al.,1999). The latter two ovarian reserve
markers fluctuate with the menstrual cycle as discussed above. AMH secretion is gonadotropin-
independent which means it stays consistent throughout the menstrual cycle, making it the
superior indirect marker of ovarian reserve over FSH and inhibin B. Both median and mean
AMH serum levels decrease with age and the wide year to year variation in values reflects
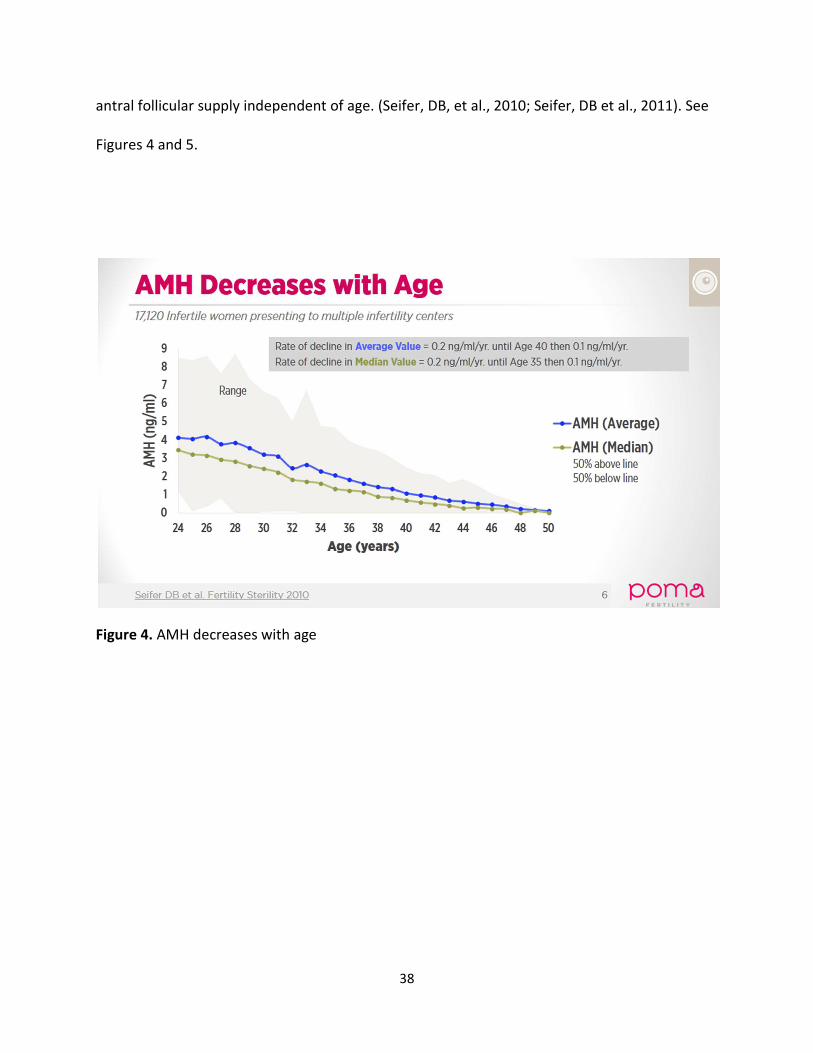
38
antral follicular supply independent of age. (Seifer, DB, et al., 2010; Seifer, DB et al., 2011). See
Figures 4 and 5.
Figure 4. AMH decreases with age

39
AMH Parallels Antral Follicle Count
Figure 5. AMH parallels antral follicle count, Modified from Seifer, DB, et al., Fertility and Sterility, 2011
3.0 Confounders of Ovarian Reserve
A few studies have explored factors that may affect ovarian reserve. One study examined
body mass index (BMI) and ovarian reserve markers, including markers evaluated in this thesis,
to assess the association between BMI and ovarian reserve in late reproductive age (40-52)
women. Su et al., reported that obese (BMI > 30) women had significantly lower levels of AMH
(p =0.02) and inhibin B (p=0.08) than normal (BMI < 25) weight, age-matched women
respectively. FSH, AFC, and E2 were not found to be associated with BMI and the AMH
differences appeared to result from physiological processes other than decreased ovarian
reserve (Su, HI et al., 2008).

40
Pal, et al.,(2010) reported in a cross sectional study of infertile women that FSH, Mullerian
Inhibitor Substance (same as AMH), and inhibin B are associated with chronic stress measured
by morning cortisol levels and psychosocial stressor indicators. Age, family history of early
menopause and chronic stress were all predictive of DOR (Pal, L et al., 2010). Finally, Kinney, A
et al., investigated whether alcohol, caffeine, and smoking were associated with FSH, inhibin B,
E2, and AFC as indicators of ovarian age. They found that current smoking increased FSH (95%
CI 0.04-0.39), but had no influence on the other measures and neither alcohol or caffeine was
related to any ovarian age indicator (Kinney, A et al., 2007). The etiology of DOR is likely
multifaceted and will take research efforts such as those involved in this thesis to positively
impact the future of women in the US and likely the world, who suffer from its'consequences.
To the best of the authors' knowledge, the research involved in this thesis is the first to
investigate polycyclic aromatic carbon-induced ovotoxicity using serum markers of ovarian
reserve in humans.

41
4.0 Study Objective
The study objective was to evaluate the association between PAH exposure determined by
urinary biomonitoring of 9 OH-PAH metabolites and three serum markers of ovarian reserve,
AMH, inhibin B, and FSH in reproductive aged women. We hypothesized that women with
higher PAH exposure levels would have decreased ovarian reserve (decreased AMH and inhibin
B, and increased FSH) compared to women with lower levels of PAH exposure. A second
hypothesis was that a woman's susceptibility to PAH effects on ovarian reserve would be
affected by whether she had polymorphisms in enzymes that detoxify PAHs (homozygous null,
GSTM1 and GSTT1). We hypothesized that women who were homozygous null GSTM1 and or
GSTT1 would have lower ovarian reserve markers compared to women who were not.
5.0 Methods
5.1 Study Design
This is a cross sectional study analysis of data collected from a prospective pilot study
investigating whether the average exposure to PAHs over time, measured as the average
concentration of urinary OH-PAHs collected over a 6 month period, had a dose-dependent
adverse effect on ovarian reserve markers in reproductive aged women and whether the effect
was modified by polymorphisms of PAH metabolizing enzymes. The general aim was to
understand how toxicants cause ovarian dysfunction in order to develop future preventative
measures. This study aimed to measure OH-PAH metabolite excretion profiles in the urine and
baseline serum ovarian reserve markers and genetic polymorphisms in women who did not
intend to become pregnant during the study period. Fifty-one women who were not on
hormonal therapy were selected for the urinary PAH biomonitoring group from a convenience

42
sample of women who did not live in segments selected for the National Children's Study (NCS).
This pilot study was designed to test whether the methods used would be feasible to use in a
study with a larger sample size, as an adjunct to the planned NCS. Initially, eligible women were
identified and recruited by door to door household contact with fliers, follow-up telephone and
email contact by study staff. Subsequently, when the NCS shifted to other recruitment
strategies, eligible women were recruited at public events such as health fairs at universities
and colleges, work places and those sponsored by community groups. Visits to the study center,
questionnaires, diaries, email, telephone, mail, and web-based communications, facilitated by
the study staff were used for data collection and education.
5.2 Description of Study Population (Participant Sampling Frame)
5.2.1 Inclusion and Exclusion Criteria
Eligible women were between the ages of 18 and 44, residing in Orange County California,
who were not pregnant currently, not planning to conceive, and not using hormonal
contraception. Exclusion criteria included ovarian failure (surgical, infertility treatments),
history of chemotherapy, pelvic radiation, pelvic inflammatory disease, polycystic ovarian
syndrome, endometriosis, women without menses for 5 or more months, inability to speak
English or Spanish, inability to understand the consent form, and residence in one of the
Orange County, CA NCS segments.

43
5.3 Study Encounters and Specimen Processing
5.3.1 Baseline Study Visit
After completing informed consent, the recruited women scheduled baseline evaluations at
one of two Orange County locations of the UC Irvine Institute for Clinical and Translational
Science (ICTS) on dates projected to be 5 to 9 days prior to their next expected menstrual cycle.
ICTS nursing staff measured height, weight, and blood pressures and collected blood samples.
Study staff administered baseline questionnaires, which were adapted from the standardized
preconception questionnaire developed for the NCS (NCS, 2007) and from the NCS First
Trimester Maternal In-Person Questionnaire (NCS, 2007). They obtained the demographic
information (age, birthplace, ethnicity, household income, education), medical history (cancer,
chronic illnesses, medication use), reproductive history (menstrual history, pregnancy and
contraceptive history, fertility and reproductive problems), tobacco, alcohol use (EtOH),
exercise, and illicit drug use history, environmental tobacco smoke exposure, exercise history,
occupational history (job title, industry, exposure to specific chemicals and hazards), use of
cleaning agents and pesticides in the home and yard, as well as pets. Participants were
provided with urine collection kits and instructions for urine collection and storage.
Daily Diary: Participants were given and asked to record daily information about occurrence of
menstrual bleeding, heaviness of menstrual flow (assessed with the help of a menstrual
pictogram, as described by Wyatt (Wyatt, KM et al. 2001), use of contraception, number of
cigarettes smoked, illness, use of medications, use of alcohol, and whether they had sexual
intercourse (Tingen, C et al., 2004).

44
5.3.2 Measurement of Urinary Hydroxylated PAH Metabolites
Participants were instructed to collect one urine sample per menstrual cycle (on cycle day
10) into a plastic beaker, pour 10 ml from the beaker into each of 3 tubes, and to store them in
the box provided with the kit in their home freezers for pickup by study staff every month.
Urine samples were used for the measurement of hydroxylated PAH metabolites as biomarkers
of PAH exposure (Li, Z et al., 2008), and creatinine. Urine samples for measurements of 9
urinary OH-PAHs were collected over a 6 menstrual cycle period (2010-2012). Urinary OH-PAH
metabolites were measured for each subject for two (2 participants) or three (49 participants)
cycles during the testing period by the California Department of Public Health Environmental
Health Laboratory using procedures developed by the Center for Disease Control and
Prevention NHANES study (CDC, 2013; Li, Z et al., 2008). If a participant only had collected two
or three cycles of urinary hormone data, (collected for a different aim of this data set) then OH-
PAHs were measured in all cycles, as the original plan for the other aim was to get hormone
data on 6 cycles over the 6 month study period. If the participant collected urinary hormone
data for more than three cycles, then the first three cycles with the fewest missing days of
urinary hormone data were selected for OH-PAH measurements. Urine samples were
enzymatically deconjugated and subjected to solid phase extraction, followed by gas
chromatography/isotope dilution high resolution mass spectrometry for nine PAH metabolites,
NAP1 and NAP2, PYR1, PHEN1, PHEN2 and PHEN3, and FLUO2, FLUO3, and FLUO9, (Romanoff,
LC et al., 2006; Li, Z et al., 2008). Urinary OH-PAH concentrations were expressed per gram
urinary creatinine or unadjusted OH-PAH concentrations were used in separate analysis. A

45
small percentage of analytes were below the limits of detection (LOD) of the assay. These were
set to the LOD divided by the square root of two (Ogden, TL et al., 2010).
Participants also performed human chorionic gonadotropin Clearblue® Easy pregnancy tests
(Swiss Precision Diagnostics) by urinary dipstick if their menses onset had not occurred within
10 days after the expected date.
5.3.3 Serum Ovarian Reserve Markers
One 10 ml tube of blood was collected and allowed to clot at room temperature, then
centrifuged at room temperature for 20 min at 1500xg. The serum was then removed for
measurements of AMH, FSH, and inhibin B and stored at –70°C until assays were performed.
AMH was measured in ng/ml, inhibin B in ng/L, and FSH in units per liter (U/L). De-identified
frozen serum samples were shipped by express mail on dry ice to Dr. Jenny Visser’s laboratory
(Erasmus Medical Center, Rotterdam, The Netherlands) for hormone analyses (AMH, FSH, and
inhibin B immunoassays). AMH was measured using two monoclonal mouse anti-human
antibodies directed against two different antigenetic determinants of AMH (Kevenaar, ME et
al., 2006). FSH was measured by chemiluminescent immunoassay (Diagnostic Products
Corporation, Los Angeles), in the same manner as AMH. Serum inhibin B was measured using
enzyme-linked immunoassay (ELISA) (Serotec Limited, Kidlington, UK), which used two
antibodies against the alpha and beta-subunits of the inhibin B molecule (de Vet, A et al., 2002).
For all assays, each subject sample was run in duplicate and the average of the two samples
were used for statistical analyses and data presentation.

46
5.3.4 GSTM1 and GSTT1 null polymorphism Genotyping
Genotyping for the GSTM1 and GSTT1 null polymorphisms was performed using DNA
extracted from clotted whole blood samples, according to the QIAGEN Supplementary Protocol:
"Purification of archive-quality DNA from clotted whole blood using Clotspin Baskets (Qiagen
product no. 158932) and the Gentra Puregene Blood Kit" (Qiagen Cat#158389) by the UC Irvine
ICTS laboratory. Tagman Copy Number Assays were used (Assay # Hs00010004_cn for GSTT1
and Hs02575461_cn for GSTM1). In these assays, RNase P (RNAP) was used as an internal
control representing a single-copy reference gene (1/1 genotype). Reaction volumes were 20 μl
and they contained: 4 μl of water, 1 μL of 20x TagMan@ primer/probe mix for target gene
(FAM labeled), 1μl of 20x TagMan@ primer/probe mix for RNAP(VIC labeled), 10 ul of 2x
TagMan @ Genotyping Mast Mix (Applied Biosystems), and 4μl of genomic DNA at a
concentration of 5 ng/μl. Reactions were run on a 7900HT Thermocycler under these
conditions: 95 degree Celsius (C) hot-start for 10 min, followed by 40 cycles of 95 degree
centigray for 15 seconds (s), at 60 degrees C for 1 min. Each sample was run in quadruplicate,
with the deltaCt averaged over the 4 replicates. Data was analyzed using Copy Caller Software,
supplied by Applied Biosystems, which determined gene copy number.
56 to 77 % of the Caucasian population have the GSTM1 null genotype (Farin, FM et al.,
2001; Wang H et al., 2002; Wenslaff, AS et al., 2005; McCarroll, SA et al., 2006) or deficiency,
compared to 44 to 23% who have a copy of the gene. 21.3 % of the Southern California Hispanic
population have the GSTT1 null genotype compared to 78.7 % who have a copy of the gene
(Gilliland, FD et al., 2002). 40.9% of the Southern California Hispanic population have the

47
GSTM1 null genotype, compared to 59.1 % who have a copy of the gene (Gilliland, FD et al.,
2002).
6.0 Statistical Analysis Methodology
6.1 Exposure Variables
Creatinine adjusted OH-PAH urinary metabolites were natural-log (In) transformed for right
skewed distributions and the transformed variables were found to be normally distributed. The
natural-log transformations were used for the remainder of the analysis. Correlations among
the In-OH-PAHs were calculated using Pearson correlation coefficients. We used the arithmetic
mean concentrations of all 3 cycles for each In-OH- PAH metabolite because we were interested
in long-term exposure effects on ovarian reserve markers. One-way analysis of variance
(ANOVA) was used to test for bivariate associations between alcohol consumption (EtOH),
income, education and race/ethnicity and OH-PAH urinary metabolites. Independent sample t-
test (equality of variance was not assumed) were used to test for bivariate associations
between cigarette use, caffeinated coffee, caffeinated tea, soda/energy drink and the averages
of each OH-PAH urinary metabolite collected over the study period.
6.2 Outcome Variables
The ovarian reserve markers were found to be right skewed and natural-log transformed for
normal distribution. The natural-log transformed data was used for all subsequent analyses.
Pearson correlations were performed on the ovarian reserve markers as well as between the
ovarian reserve markers and the age and BMI of the participants for bivariate analysis. ANOVA
was used to test for bivariate associations between EtOH, income, education and race/ethnicity
and ovarian reserve markers. Independent sample T test were used to test for bivariate

48
associations between cigarette use, caffeinated coffee, caffeinated tea, and soda/energy drink
and ovarian reserve markers AMH, inhibin B, and FSH.
6.3 Adjusted Measures of Association
Multiple linear regression models were used to investigate associations between the OH-
PAH urinary markers and serum concentrations of the ovarian reserve markers with
adjustments for important covariates. Each outcome marker was regressed against the OH-
PAHs, and OH-PAHs were retained in the models at a significance level of 0.20. Age, BMI,
education, caffeinated tea, soda/energy drink and EtOH were selected as covariates, using a
cut-off of p < 0.20 for their associations as potential confounders with the exposure and
outcome variables in the bivariate analysis. Caffeinated coffee and cigarette use were not
included because they are considered sources of PAH exposure. Family income was not chosen
to represent socio-economic status because there were too many (N=7) missing values. All pre-
selected covariates were entered into the Statistical Program for Social Sciences (SPSS) program
while building the models for the 3 outcome variables (The Enter Method). The covariates were
retained in the models if they changed the standardized beta coefficient of the PAH(s) more
than 10%.
Principal Component Factor analysis with variance maximizing (varimax) rotation was used
to reduce the dimensions in the PAH predictors to better understand the multicollinearity of
the PAHs.

49
6.3.1 Effect Modification of Polymorphisms
Bivariate associations between the genotypes GSTM1 and GSTT1 were investigated with
independent t-test with all 9 of the OH-PAH metabolites and none were found to be significant
(p > 0.05). Bivariate associations between GSTM1 and GSTT1 and the ovarian reserve marker
endpoints (AMH, inhibin B, and FSH) were investigated using independent t-test. To evaluate
the effect modification of the homozygous null GSTM1 and homozygous null GSTT1, split file,
compare group analysis were performed. The participants were stratified on the null genotype
for both polymorphisms and compared to all others in separate multivariable analyses. The
adjusted multivariable models for the ovarian reserve markers were run comparing the
homozygous null genotypes to the heterozygous/wild-type genotype group. All statistical
analyses were done with SPSS version 23.
7.0 RESULTS
The mean age of the participants was 29.86 (SD 6.46) and the mean BMI was 24.9 (SD 5.29),
which is considered normal weight. Fifty three percent of the participants had a bachelors
degree or higher and just over half consumed a moderate amount of alcohol (55%) on a regular
basis. Table 3 summarizes selected participant characteristics.

50
Table 3: Participant Characteristics (Demographics)
Characteristic Mean SD
Age (years)at baseline 29.90 6.50
BMI (Kg/m2)* 24.90 5.30
Number Percent
Race/Ethnicity
Hispanic 11 21.60
Non-Hispanic White 22 43.10
Non-Hispanic Asian 14 27.50
Non-Hispanic Black 4 7.80
Education - **
Some High School/High School)/Some
College/Vocational Program
23 45.10
Bachelor's Degree 16 31.40
Graduate/Doctoral Degree 11 21.60
Ever Smoked or Current Smoker
Yes 5 9.80
< 1per month - no 46 90.20
Alcohol Use
> 1 a week 12 24
>1 per month to 1/week 16 31
Never to <1/month 21 41
Family Income in the past year****
< $ 50,000 12 23.50
> $ 50,000 - > $100,000 14 27.40
> $100.000 18 35.20
Caffeinated Beverages (Yes Routinely)
Coffee 28 55
Tea 21 41
Soda/Energy Drinks 12 24
* 3 missing BMIs
* *1 missing Education level
*** 2 missing Alcohol
*** *7 missing Family Income/Median household
income for Orange County 2009-2013 = 75K
7.1 PAH Metabolite Measurements
The participants' geometric mean (GM) concentrations of PAHs for the unadjusted (ng/L) and
the creatinine adjusted (ng/g creatinine) data were compared to the geometric means and 95%
CIs for women from the NHANES data for the US general population during the study period
(Table 4). Creatinine unadjusted NAP2, FLUO3, FLUO9, and PHEN3 did not fall within the
NHANES 95% CI for the geometric means, but did fall within the 95% CI for the 75th percentile.
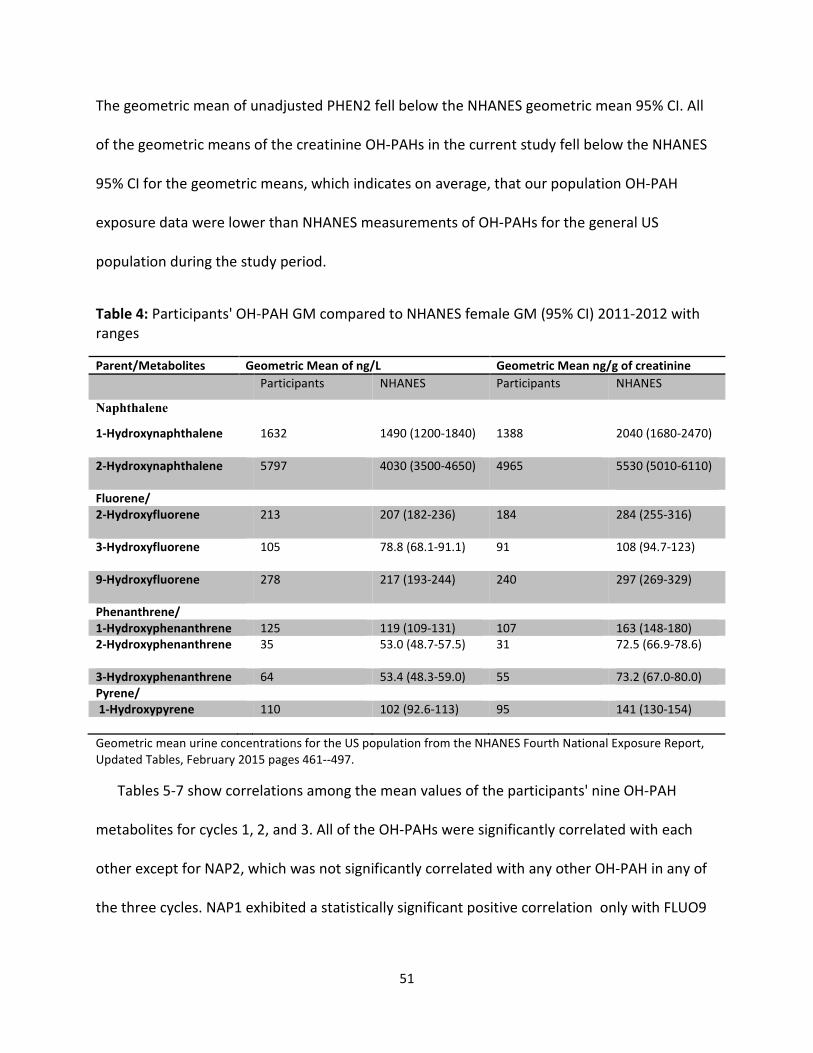
51
The geometric mean of unadjusted PHEN2 fell below the NHANES geometric mean 95% CI. All
of the geometric means of the creatinine OH-PAHs in the current study fell below the NHANES
95% CI for the geometric means, which indicates on average, that our population OH-PAH
exposure data were lower than NHANES measurements of OH-PAHs for the general US
population during the study period.
Table 4: Participants' OH-PAH GM compared to NHANES female GM (95% CI) 2011-2012 with
ranges
Parent/Metabolites Geometric Mean of ng/L Geometric Mean ng/g of creatinine
Participants NHANES Participants NHANES
Naphthalene
1-Hydroxynaphthalene 1632 1490 (1200-1840) 1388 2040 (1680-2470)
2-Hydroxynaphthalene 5797 4030 (3500-4650) 4965 5530 (5010-6110)
Fluorene/
2-Hydroxyfluorene
213 207 (182-236) 184 284 (255-316)
3-Hydroxyfluorene
105 78.8 (68.1-91.1) 91 108 (94.7-123)
9-Hydroxyfluorene
278 217 (193-244) 240 297 (269-329)
Phenanthrene/
1-Hydroxyphenanthrene 125 119 (109-131) 107 163 (148-180)
2-Hydroxyphenanthrene
35 53.0 (48.7-57.5) 31 72.5 (66.9-78.6)
3-Hydroxyphenanthrene 64 53.4 (48.3-59.0) 55 73.2 (67.0-80.0)
Pyrene/
1-Hydroxypyrene 110 102 (92.6-113) 95 141 (130-154)
Geometric mean urine concentrations for the US population from the NHANES Fourth National Exposure Report,
Updated Tables, February 2015 pages 461--497.
Tables 5-7 show correlations among the mean values of the participants' nine OH-PAH
metabolites for cycles 1, 2, and 3. All of the OH-PAHs were significantly correlated with each
other except for NAP2, which was not significantly correlated with any other OH-PAH in any of
the three cycles. NAP1 exhibited a statistically significant positive correlation only with FLUO9
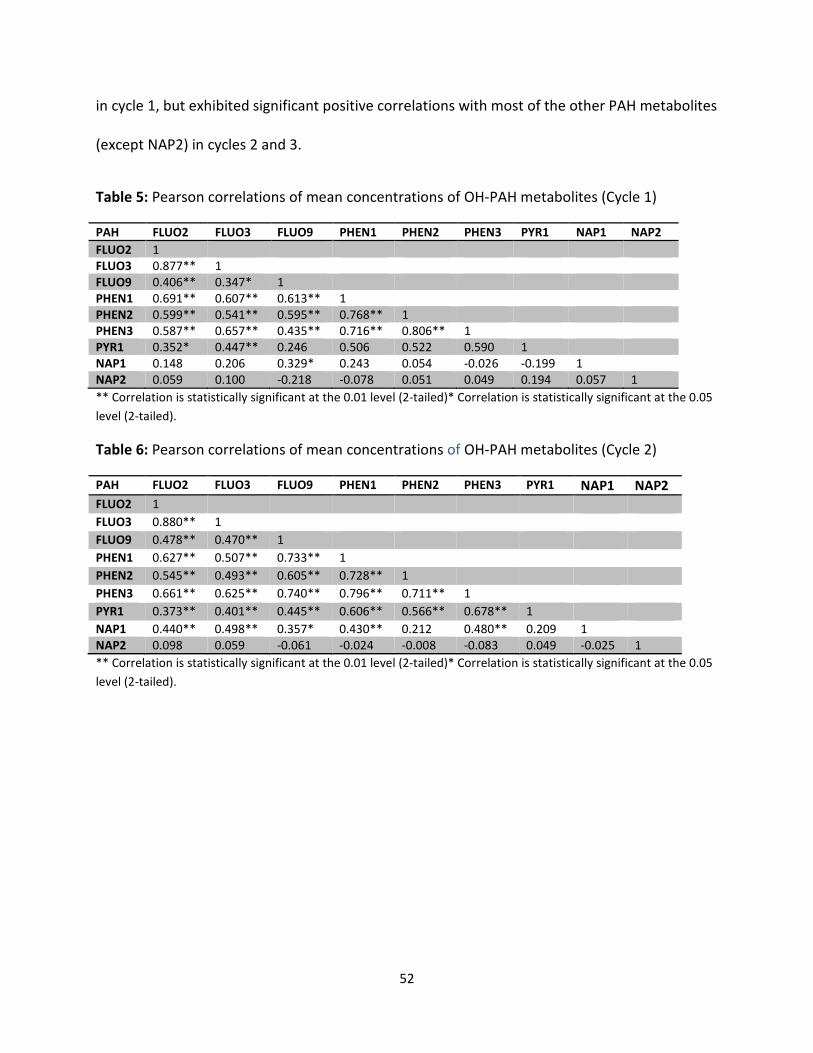
52
in cycle 1, but exhibited significant positive correlations with most of the other PAH metabolites
(except NAP2) in cycles 2 and 3.
Table 5: Pearson correlations of mean concentrations of OH-PAH metabolites (Cycle 1)
PAH FLUO2 FLUO3 FLUO9 PHEN1 PHEN2 PHEN3 PYR1 NAP1 NAP2
FLUO2 1
FLUO3 0.877** 1
FLUO9 0.406** 0.347* 1
PHEN1 0.691** 0.607** 0.613** 1
PHEN2 0.599** 0.541** 0.595** 0.768** 1
PHEN3 0.587** 0.657** 0.435** 0.716** 0.806** 1
PYR1 0.352* 0.447** 0.246 0.506 0.522 0.590 1
NAP1 0.148 0.206 0.329* 0.243 0.054 -0.026 -0.199 1
NAP2 0.059 0.100 -0.218 -0.078 0.051 0.049 0.194 0.057 1
** Correlation is statistically significant at the 0.01 level (2-tailed)* Correlation is statistically significant at the 0.05
level (2-tailed).
Table 6: Pearson correlations of mean concentrations of OH-PAH metabolites (Cycle 2)
PAH FLUO2 FLUO3 FLUO9 PHEN1 PHEN2 PHEN3 PYR1 NAP1 NAP2
FLUO2 1
FLUO3 0.880** 1
FLUO9 0.478** 0.470** 1
PHEN1 0.627** 0.507** 0.733** 1
PHEN2 0.545** 0.493** 0.605** 0.728** 1
PHEN3 0.661** 0.625** 0.740** 0.796** 0.711** 1
PYR1 0.373** 0.401** 0.445** 0.606** 0.566** 0.678** 1
NAP1 0.440** 0.498** 0.357* 0.430** 0.212 0.480** 0.209 1
NAP2 0.098 0.059 -0.061 -0.024 -0.008 -0.083 0.049 -0.025 1
** Correlation is statistically significant at the 0.01 level (2-tailed)* Correlation is statistically significant at the 0.05
level (2-tailed).
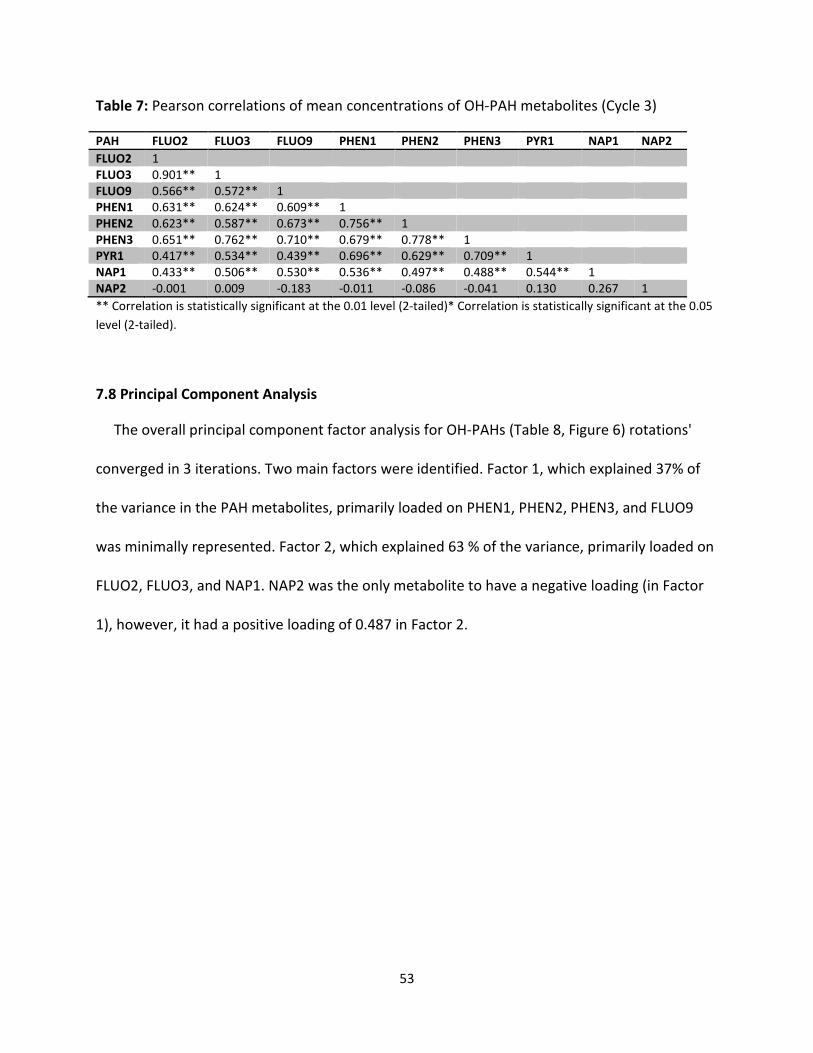
53
Table 7: Pearson correlations of mean concentrations of OH-PAH metabolites (Cycle 3)
PAH FLUO2 FLUO3 FLUO9 PHEN1 PHEN2 PHEN3 PYR1 NAP1 NAP2
FLUO2 1
FLUO3 0.901** 1
FLUO9 0.566** 0.572** 1
PHEN1 0.631** 0.624** 0.609** 1
PHEN2 0.623** 0.587** 0.673** 0.756** 1
PHEN3 0.651** 0.762** 0.710** 0.679** 0.778** 1
PYR1 0.417** 0.534** 0.439** 0.696** 0.629** 0.709** 1
NAP1 0.433** 0.506** 0.530** 0.536** 0.497** 0.488** 0.544** 1
NAP2 -0.001 0.009 -0.183 -0.011 -0.086 -0.041 0.130 0.267 1
** Correlation is statistically significant at the 0.01 level (2-tailed)* Correlation is statistically significant at the 0.05
level (2-tailed).
7.8 Principal Component Analysis
The overall principal component factor analysis for OH-PAHs (Table 8, Figure 6) rotations'
converged in 3 iterations. Two main factors were identified. Factor 1, which explained 37% of
the variance in the PAH metabolites, primarily loaded on PHEN1, PHEN2, PHEN3, and FLUO9
was minimally represented. Factor 2, which explained 63 % of the variance, primarily loaded on
FLUO2, FLUO3, and NAP1. NAP2 was the only metabolite to have a negative loading (in Factor
1), however, it had a positive loading of 0.487 in Factor 2.
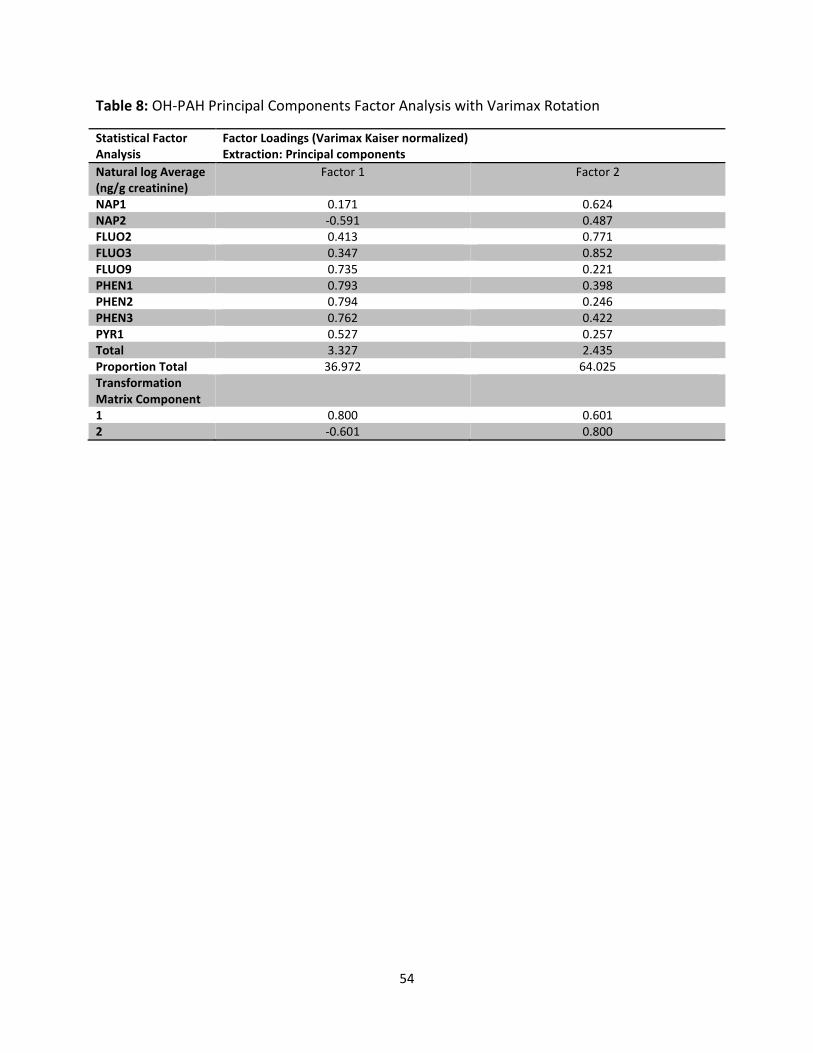
54
Table 8: OH-PAH Principal Components Factor Analysis with Varimax Rotation
Statistical Factor
Analysis
Factor Loadings (Varimax Kaiser normalized)
Extraction: Principal components
Natural log Average
(ng/g creatinine)
Factor 1 Factor 2
NAP1 0.171 0.624
NAP2 -0.591 0.487
FLUO2 0.413 0.771
FLUO3 0.347 0.852
FLUO9 0.735 0.221
PHEN1 0.793 0.398
PHEN2 0.794 0.246
PHEN3 0.762 0.422
PYR1 0.527 0.257
Total 3.327 2.435
Proportion Total 36.972 64.025
Transformation
Matrix Component
1 0.800 0.601
2 -0.601 0.800
55
Figure 6: OH- PAH Principal Components Factor Analysis with Varimax Rotation.
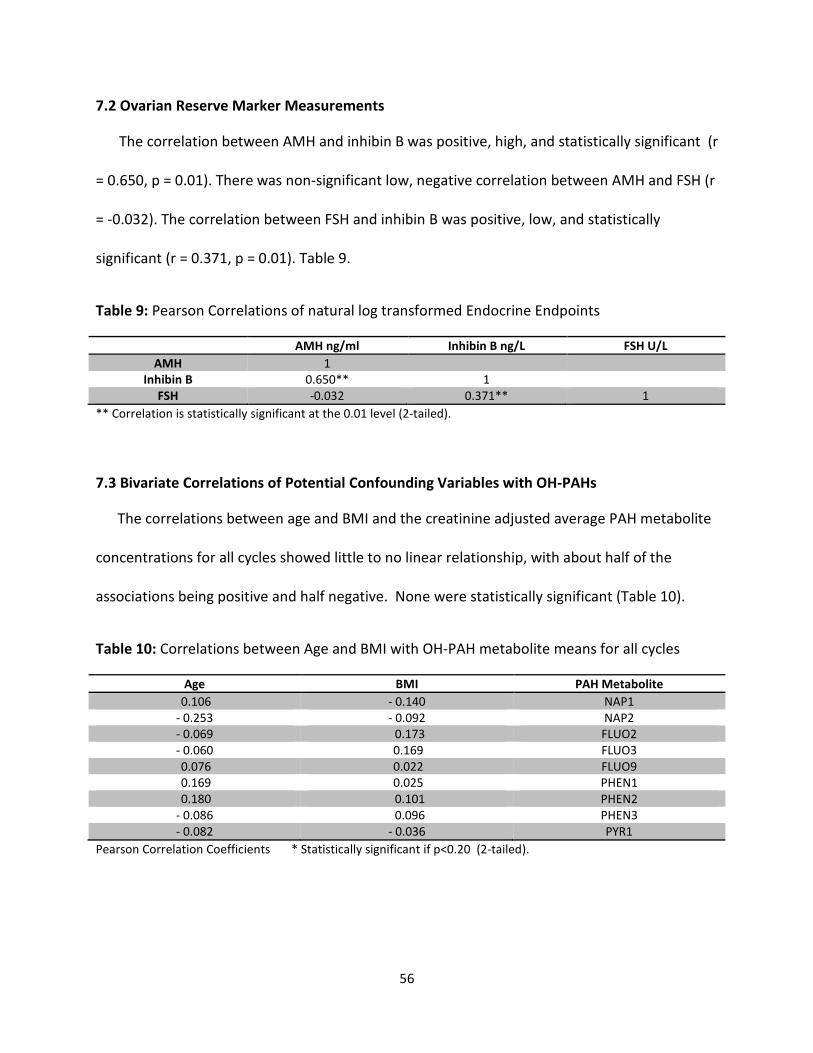
56
7.2 Ovarian Reserve Marker Measurements
The correlation between AMH and inhibin B was positive, high, and statistically significant (r
= 0.650, p = 0.01). There was non-significant low, negative correlation between AMH and FSH (r
= -0.032). The correlation between FSH and inhibin B was positive, low, and statistically
significant (r = 0.371, p = 0.01). Table 9.
Table 9: Pearson Correlations of natural log transformed Endocrine Endpoints
AMH ng/ml Inhibin B ng/L FSH U/L
AMH 1
Inhibin B 0.650** 1
FSH -0.032 0.371** 1
** Correlation is statistically significant at the 0.01 level (2-tailed).
7.3 Bivariate Correlations of Potential Confounding Variables with OH-PAHs
The correlations between age and BMI and the creatinine adjusted average PAH metabolite
concentrations for all cycles showed little to no linear relationship, with about half of the
associations being positive and half negative. None were statistically significant (Table 10).
Table 10: Correlations between Age and BMI with OH-PAH metabolite means for all cycles
Age BMI PAH Metabolite
0.106 - 0.140 NAP1
- 0.253 - 0.092 NAP2
- 0.069 0.173 FLUO2
- 0.060 0.169 FLUO3
0.076 0.022 FLUO9
0.169 0.025 PHEN1
0.180 0.101 PHEN2
- 0.086 0.096 PHEN3
- 0.082 - 0.036 PYR1
Pearson Correlation Coefficients * Statistically significant if p<0.20 (2-tailed).

57
7.5 Bivariate Correlations of Ovarian Reserve Markers with potential Confounding Variables
Table 11: Correlations of Potential Confounders with Ovarian Reserve Markers
Age BMI
AMH (ng/ml) - 0.280* - 0.293*
Inhibin B (ng/L) - 0.465** - 0.287*
FSH (U/L) 0.025 0.225
Pearson Correlations *Correlation is significant at the 0.05 level (2-tailed) **Correlation is significant at the 0.01
level (2-tailed).
There were negative associations between age and BMI with AMH and inhibin B that were
significant. There were 11 participants 35 or older. FSH was not significantly correlated with
BMI or age (Table 11).
7.6 Bivariate Analysis of PAHs and Ovarian Reserve Markers with Participant Characteristics
Cigarette use, EtOH, and education were associated with more than two hydroxylated PAH
metabolites in the analyses of associations between potential confounders and OH- PAH
average concentrations across cycles per woman (Tables 13, 14 and 18). Only 5 participants
used cigarettes. Despite this low number of smokers, cigarette use was associated with 5 of the
9 (> 50%) OH-PAH metabolites, with higher concentrations of the metabolites in smokers. There
were less than two OH-PAH associations that are different by strata, observed between
race/ethnicity, caffeinated coffee, caffeinated tea, soda/energy drink, and family income in the
past year, using OH- PAH average concentrations across cycles per woman (Tables 12, 15, 16,
17, and 19).
Bivariate associations were conducted with independent t-test between the genotypes
GSTM1 and GSTT1 with all 9 of the OH-PAH metabolites and none were found to be significant
(p > 0.05) (data not shown). Bivariate associations between GSTM1 and GSTT1 and the ovarian
reserve marker endpoints (AMH, inhibin B, and FSH) were investigated using independent t-test

58
as well and there was a statistically significant association between GSTM1 and AMH (p = 0.04).
There were no other statistically significant associations between the genotypes and the
ovarian reserve markers observed (data not shown).
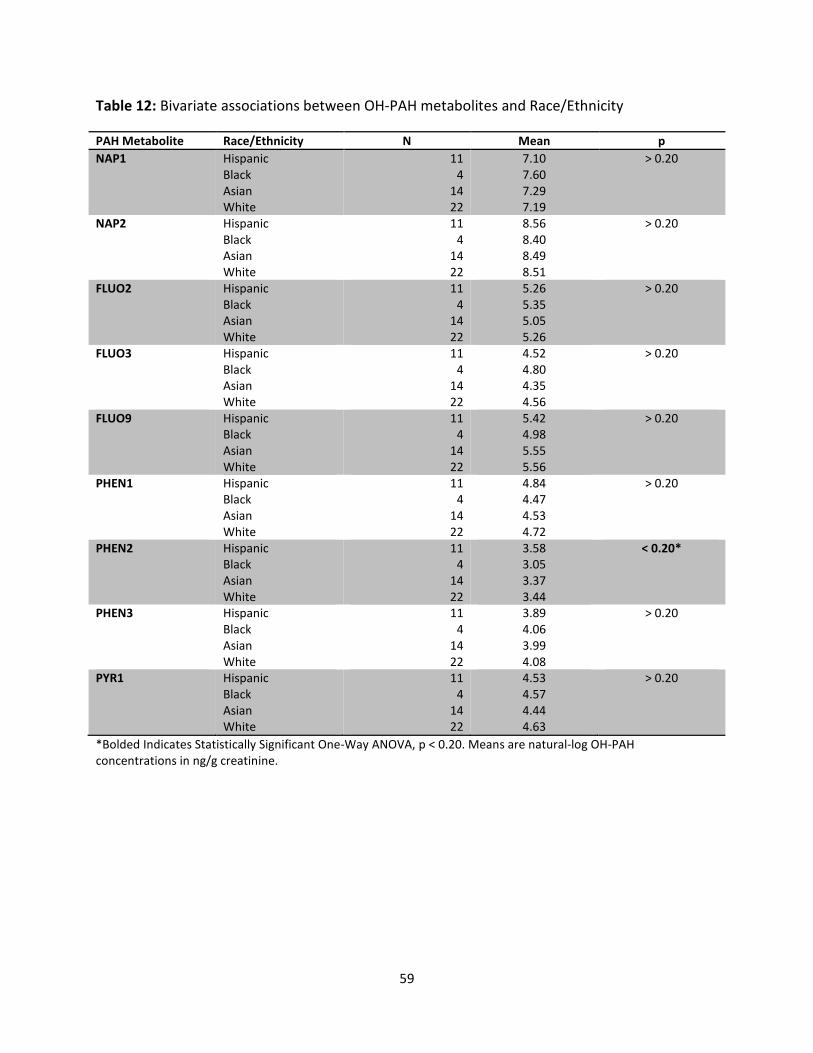
59
Table 12: Bivariate associations between OH-PAH metabolites and Race/Ethnicity
PAH Metabolite Race/Ethnicity N Mean p
NAP1 Hispanic
Black
Asian
White
11
4
14
22
7.10
7.60
7.29
7.19
> 0.20
NAP2 Hispanic
Black
Asian
White
11
4
14
22
8.56
8.40
8.49
8.51
> 0.20
FLUO2 Hispanic
Black
Asian
White
11
4
14
22
5.26
5.35
5.05
5.26
> 0.20
FLUO3 Hispanic
Black
Asian
White
11
4
14
22
4.52
4.80
4.35
4.56
> 0.20
FLUO9 Hispanic
Black
Asian
White
11
4
14
22
5.42
4.98
5.55
5.56
> 0.20
PHEN1 Hispanic
Black
Asian
White
11
4
14
22
4.84
4.47
4.53
4.72
> 0.20
PHEN2 Hispanic
Black
Asian
White
11
4
14
22
3.58
3.05
3.37
3.44
< 0.20*
PHEN3 Hispanic
Black
Asian
White
11
4
14
22
3.89
4.06
3.99
4.08
> 0.20
PYR1 Hispanic
Black
Asian
White
11
4
14
22
4.53
4.57
4.44
4.63
> 0.20
*Bolded Indicates Statistically Significant One-Way ANOVA, p < 0.20. Means are natural-log OH-PAH
concentrations in ng/g creatinine.
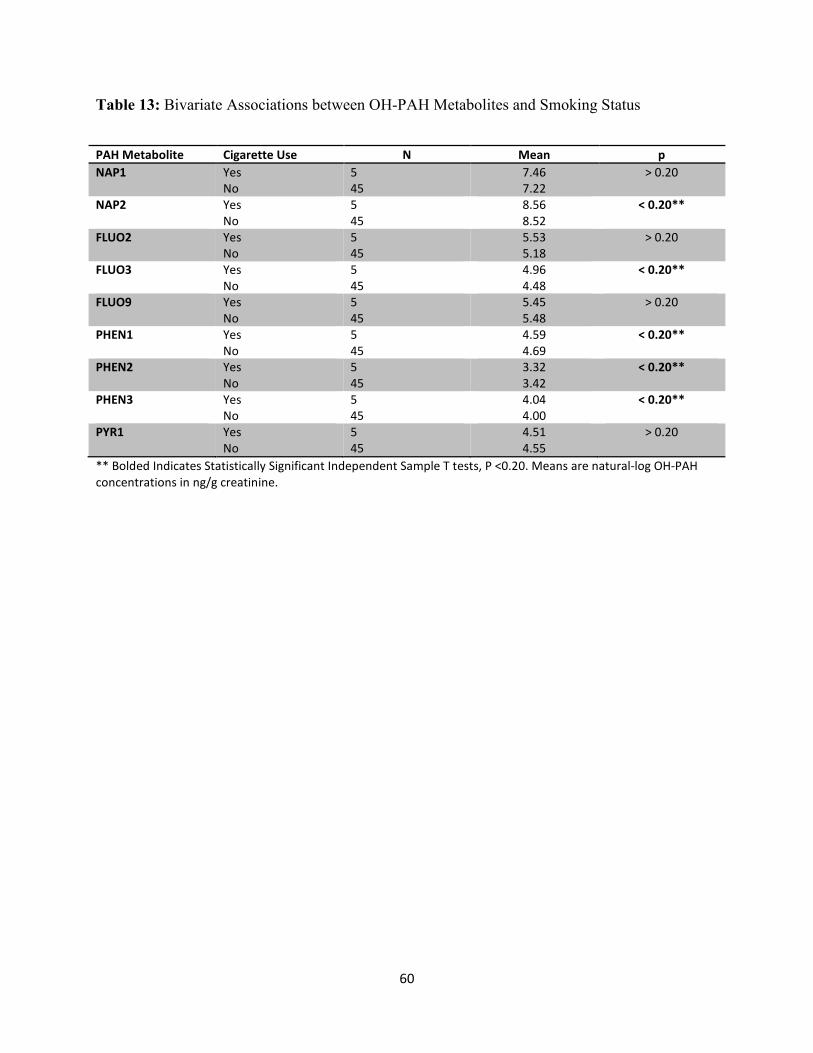
60
Table 13: Bivariate Associations between OH-PAH Metabolites and Smoking Status
PAH Metabolite Cigarette Use N Mean p
NAP1 Yes
No
5
45
7.46
7.22
> 0.20
NAP2 Yes
No
5
45
8.56
8.52
< 0.20**
FLUO2 Yes
No
5
45
5.53
5.18
> 0.20
FLUO3 Yes
No
5
45
4.96
4.48
< 0.20**
FLUO9 Yes
No
5
45
5.45
5.48
> 0.20
PHEN1 Yes
No
5
45
4.59
4.69
< 0.20**
PHEN2 Yes
No
5
45
3.32
3.42
< 0.20**
PHEN3 Yes
No
5
45
4.04
4.00
< 0.20**
PYR1 Yes
No
5
45
4.51
4.55
> 0.20
** Bolded Indicates Statistically Significant Independent Sample T tests, P <0.20. Means are natural-log OH-PAH
concentrations in ng/g creatinine.
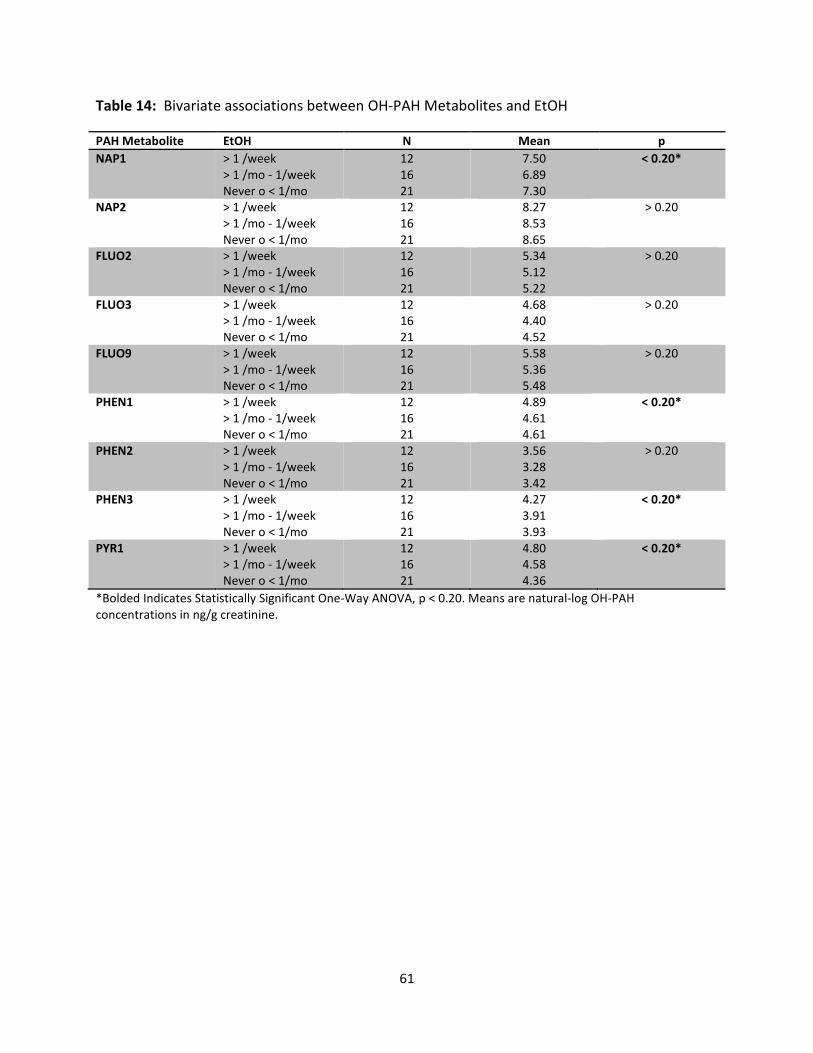
61
Table 14: Bivariate associations between OH-PAH Metabolites and EtOH
PAH Metabolite EtOH N Mean p
NAP1 > 1 /week
> 1 /mo - 1/week
Never o < 1/mo
12
16
21
7.50
6.89
7.30
< 0.20*
NAP2 > 1 /week
> 1 /mo - 1/week
Never o < 1/mo
12
16
21
8.27
8.53
8.65
> 0.20
FLUO2 > 1 /week
> 1 /mo - 1/week
Never o < 1/mo
12
16
21
5.34
5.12
5.22
> 0.20
FLUO3 > 1 /week
> 1 /mo - 1/week
Never o < 1/mo
12
16
21
4.68
4.40
4.52
> 0.20
FLUO9 > 1 /week
> 1 /mo - 1/week
Never o < 1/mo
12
16
21
5.58
5.36
5.48
> 0.20
PHEN1 > 1 /week
> 1 /mo - 1/week
Never o < 1/mo
12
16
21
4.89
4.61
4.61
< 0.20*
PHEN2 > 1 /week
> 1 /mo - 1/week
Never o < 1/mo
12
16
21
3.56
3.28
3.42
> 0.20
PHEN3 > 1 /week
> 1 /mo - 1/week
Never o < 1/mo
12
16
21
4.27
3.91
3.93
< 0.20*
PYR1 > 1 /week
> 1 /mo - 1/week
Never o < 1/mo
12
16
21
4.80
4.58
4.36
< 0.20*
*Bolded Indicates Statistically Significant One-Way ANOVA, p < 0.20. Means are natural-log OH-PAH
concentrations in ng/g creatinine.
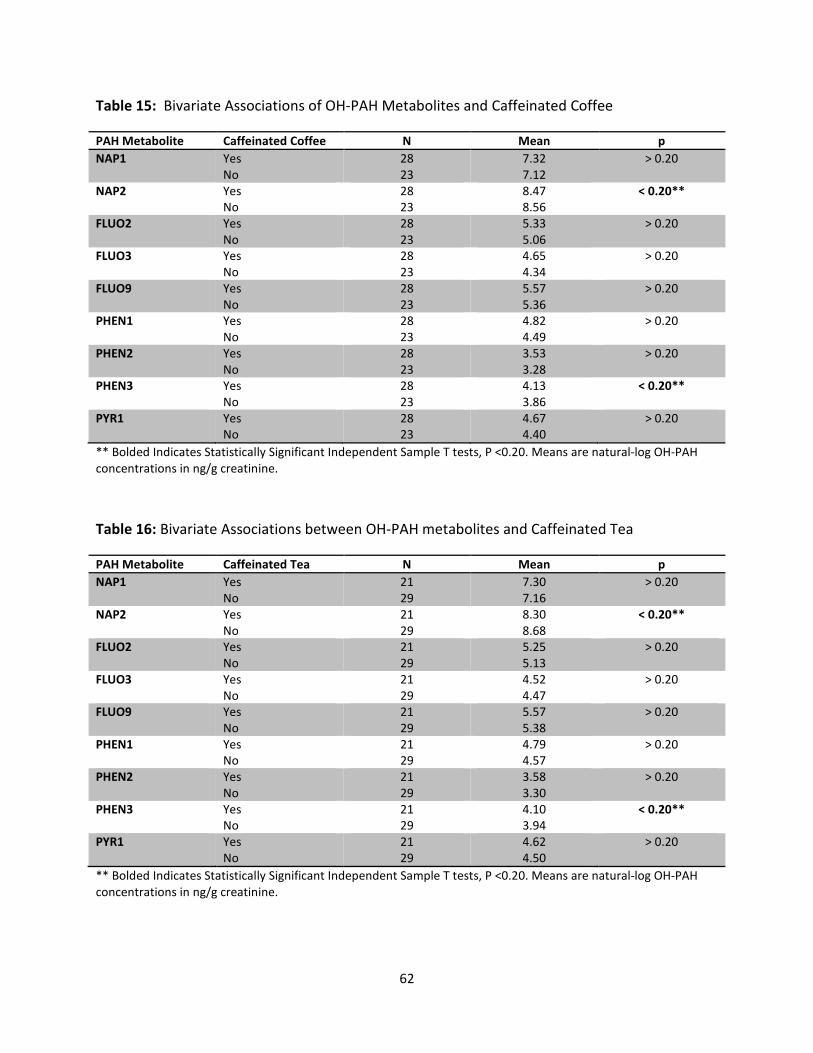
62
Table 15: Bivariate Associations of OH-PAH Metabolites and Caffeinated Coffee
PAH Metabolite Caffeinated Coffee N Mean p
NAP1 Yes
No
28
23
7.32
7.12
> 0.20
NAP2 Yes
No
28
23
8.47
8.56
< 0.20**
FLUO2 Yes
No
28
23
5.33
5.06
> 0.20
FLUO3 Yes
No
28
23
4.65
4.34
> 0.20
FLUO9 Yes
No
28
23
5.57
5.36
> 0.20
PHEN1 Yes
No
28
23
4.82
4.49
> 0.20
PHEN2 Yes
No
28
23
3.53
3.28
> 0.20
PHEN3 Yes
No
28
23
4.13
3.86
< 0.20**
PYR1 Yes
No
28
23
4.67
4.40
> 0.20
** Bolded Indicates Statistically Significant Independent Sample T tests, P <0.20. Means are natural-log OH-PAH
concentrations in ng/g creatinine.
Table 16: Bivariate Associations between OH-PAH metabolites and Caffeinated Tea
PAH Metabolite Caffeinated Tea N Mean p
NAP1 Yes
No
21
29
7.30
7.16
> 0.20
NAP2 Yes
No
21
29
8.30
8.68
< 0.20**
FLUO2 Yes
No
21
29
5.25
5.13
> 0.20
FLUO3 Yes
No
21
29
4.52
4.47
> 0.20
FLUO9 Yes
No
21
29
5.57
5.38
> 0.20
PHEN1 Yes
No
21
29
4.79
4.57
> 0.20
PHEN2 Yes
No
21
29
3.58
3.30
> 0.20
PHEN3 Yes
No
21
29
4.10
3.94
< 0.20**
PYR1 Yes
No
21
29
4.62
4.50
> 0.20
** Bolded Indicates Statistically Significant Independent Sample T tests, P <0.20. Means are natural-log OH-PAH
concentrations in ng/g creatinine.
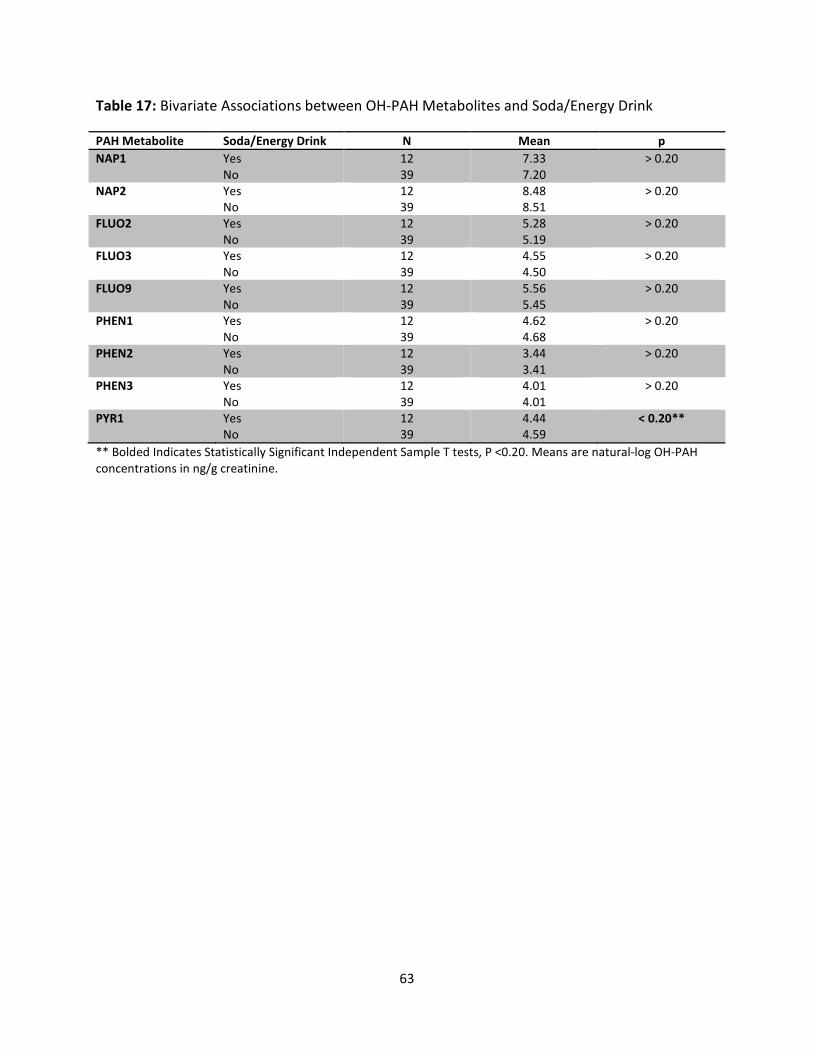
63
Table 17: Bivariate Associations between OH-PAH Metabolites and Soda/Energy Drink
PAH Metabolite Soda/Energy Drink N Mean p
NAP1 Yes
No
12
39
7.33
7.20
> 0.20
NAP2 Yes
No
12
39
8.48
8.51
> 0.20
FLUO2 Yes
No
12
39
5.28
5.19
> 0.20
FLUO3 Yes
No
12
39
4.55
4.50
> 0.20
FLUO9 Yes
No
12
39
5.56
5.45
> 0.20
PHEN1 Yes
No
12
39
4.62
4.68
> 0.20
PHEN2 Yes
No
12
39
3.44
3.41
> 0.20
PHEN3 Yes
No
12
39
4.01
4.01
> 0.20
PYR1 Yes
No
12
39
4.44
4.59
< 0.20**
** Bolded Indicates Statistically Significant Independent Sample T tests, P <0.20. Means are natural-log OH-PAH
concentrations in ng/g creatinine.
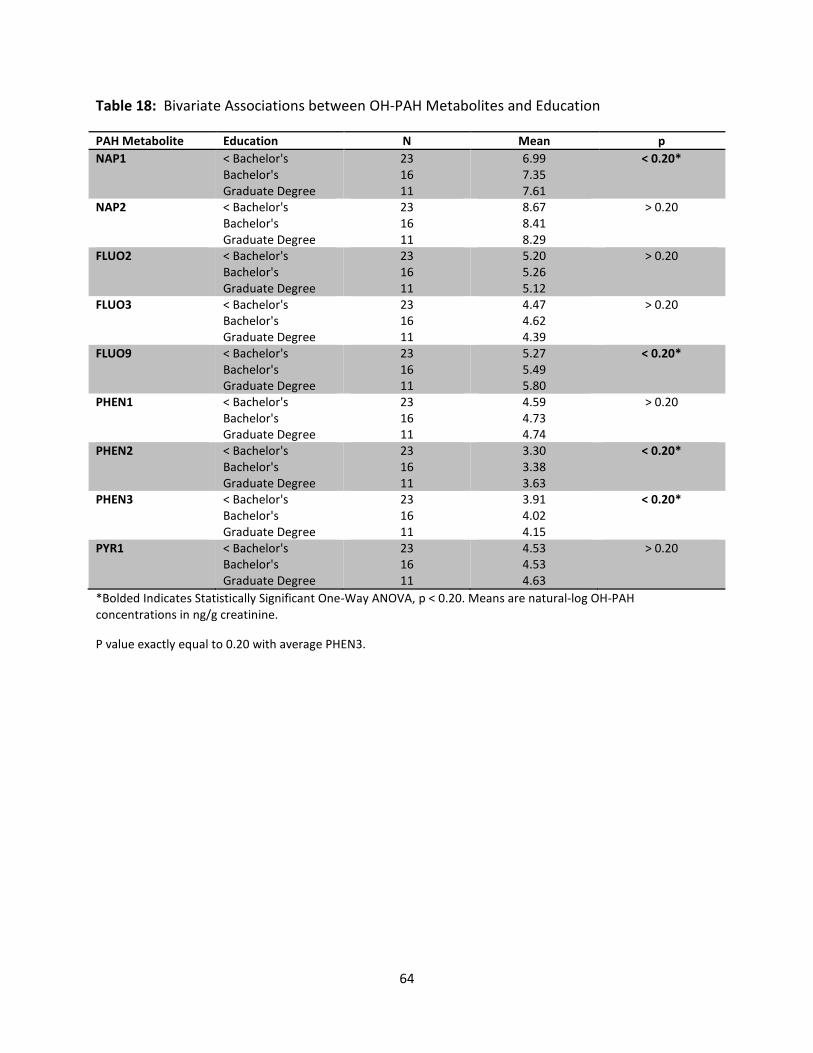
64
Table 18: Bivariate Associations between OH-PAH Metabolites and Education
PAH Metabolite Education N Mean p
NAP1 < Bachelor's
Bachelor's
Graduate Degree
23
16
11
6.99
7.35
7.61
< 0.20*
NAP2 < Bachelor's
Bachelor's
Graduate Degree
23
16
11
8.67
8.41
8.29
> 0.20
FLUO2 < Bachelor's
Bachelor's
Graduate Degree
23
16
11
5.20
5.26
5.12
> 0.20
FLUO3 < Bachelor's
Bachelor's
Graduate Degree
23
16
11
4.47
4.62
4.39
> 0.20
FLUO9 < Bachelor's
Bachelor's
Graduate Degree
23
16
11
5.27
5.49
5.80
< 0.20*
PHEN1 < Bachelor's
Bachelor's
Graduate Degree
23
16
11
4.59
4.73
4.74
> 0.20
PHEN2 < Bachelor's
Bachelor's
Graduate Degree
23
16
11
3.30
3.38
3.63
< 0.20*
PHEN3 < Bachelor's
Bachelor's
Graduate Degree
23
16
11
3.91
4.02
4.15
< 0.20*
PYR1 < Bachelor's
Bachelor's
Graduate Degree
23
16
11
4.53
4.53
4.63
> 0.20
*Bolded Indicates Statistically Significant One-Way ANOVA, p < 0.20. Means are natural-log OH-PAH
concentrations in ng/g creatinine.
P value exactly equal to 0.20 with average PHEN3.
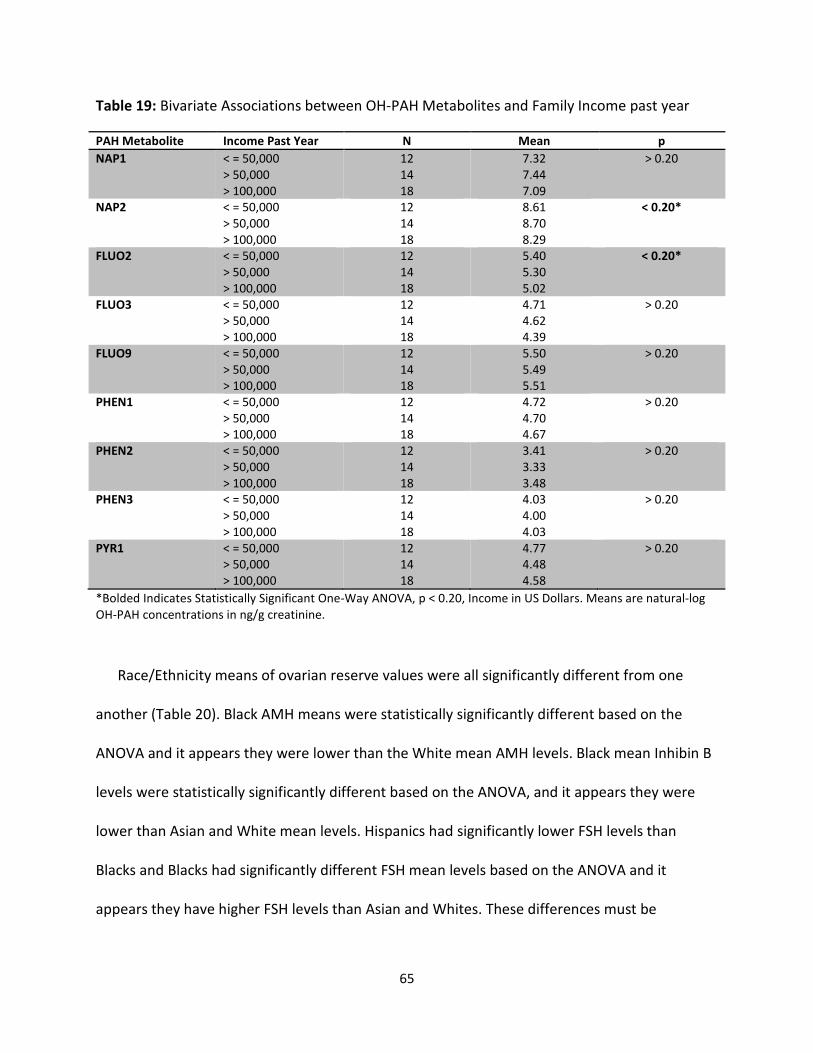
65
Table 19: Bivariate Associations between OH-PAH Metabolites and Family Income past year
PAH Metabolite Income Past Year N Mean p
NAP1 < = 50,000
> 50,000
> 100,000
12
14
18
7.32
7.44
7.09
> 0.20
NAP2 < = 50,000
> 50,000
> 100,000
12
14
18
8.61
8.70
8.29
< 0.20*
FLUO2 < = 50,000
> 50,000
> 100,000
12
14
18
5.40
5.30
5.02
< 0.20*
FLUO3 < = 50,000
> 50,000
> 100,000
12
14
18
4.71
4.62
4.39
> 0.20
FLUO9 < = 50,000
> 50,000
> 100,000
12
14
18
5.50
5.49
5.51
> 0.20
PHEN1 < = 50,000
> 50,000
> 100,000
12
14
18
4.72
4.70
4.67
> 0.20
PHEN2 < = 50,000
> 50,000
> 100,000
12
14
18
3.41
3.33
3.48
> 0.20
PHEN3 < = 50,000
> 50,000
> 100,000
12
14
18
4.03
4.00
4.03
> 0.20
PYR1 < = 50,000
> 50,000
> 100,000
12
14
18
4.77
4.48
4.58
> 0.20
*Bolded Indicates Statistically Significant One-Way ANOVA, p < 0.20, Income in US Dollars. Means are natural-log
OH-PAH concentrations in ng/g creatinine.
Race/Ethnicity means of ovarian reserve values were all significantly different from one
another (Table 20). Black AMH means were statistically significantly different based on the
ANOVA and it appears they were lower than the White mean AMH levels. Black mean Inhibin B
levels were statistically significantly different based on the ANOVA, and it appears they were
lower than Asian and White mean levels. Hispanics had significantly lower FSH levels than
Blacks and Blacks had significantly different FSH mean levels based on the ANOVA and it
appears they have higher FSH levels than Asian and Whites. These differences must be

66
interpreted with caution because they are based on very small numbers, as there were only
four black participants.
The EtOH with ovarian reserve markers were only significant for AMH (Table 22). The mean
AMH was significantly different based on the t-tests, and it appears higher in the group who
drank the most alcohol compared to the group who drank the least amount. There was a
statistically different level of FSH in coffee drinkers based on the t-tests, and it appears that
there is a lower level of FSH in coffee drinkers (Table 23) compared to non-coffee drinkers.
Women who did not drink tea had higher mean levels of all the ovarian reserve markers
compared to those who did (Table 24). The mean AMH levels were significantly different based
on the t-tests, and it appears higher in those who drank soda/energy drink compared to those
who did not (Table 25). There were statistically significant differences in inhibin B levels based
on the t-tests, and it appears that there was a higher mean Inhibin B levels among those with a
graduate degree or higher. The lowest statistically significant Inhibin B level occurred in the
group with at least a Bachelor’s degree (Table 26). There were no statistically significant
differences in the means of income observed for any ovarian reserve marker (Table 27).
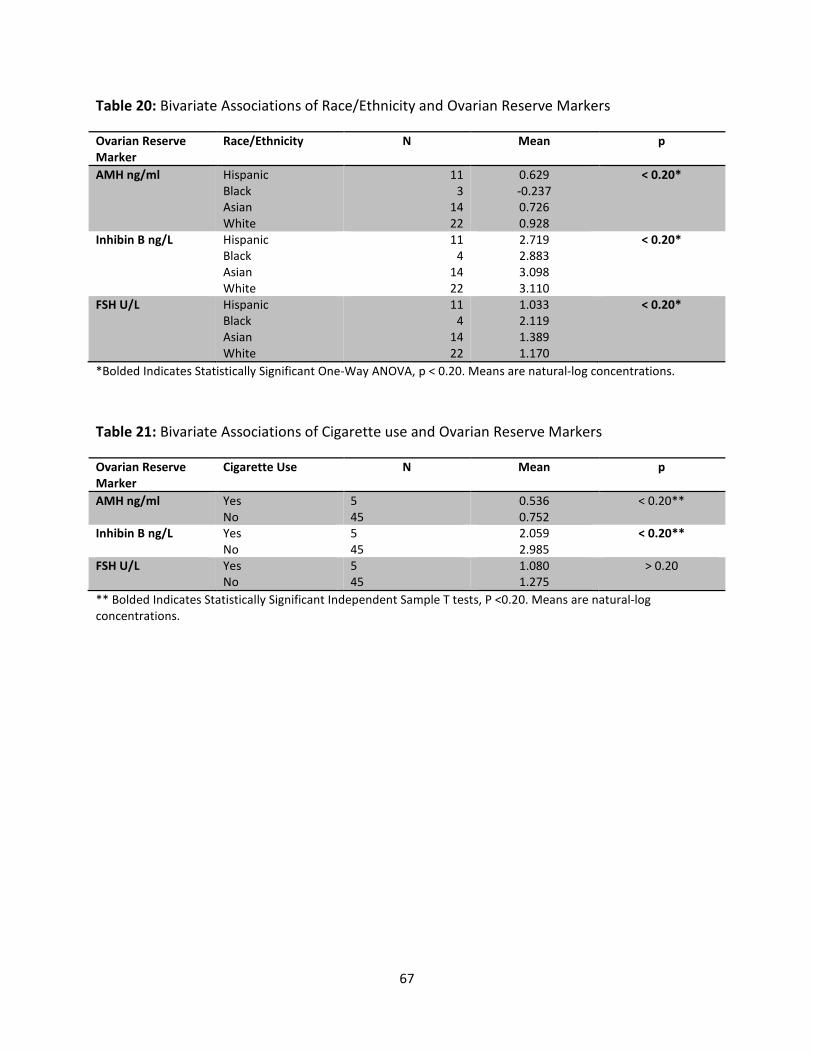
67
Table 20: Bivariate Associations of Race/Ethnicity and Ovarian Reserve Markers
Ovarian Reserve
Marker
Race/Ethnicity N Mean p
AMH ng/ml Hispanic
Black
Asian
White
11
3
14
22
0.629
-0.237
0.726
0.928
< 0.20*
Inhibin B ng/L Hispanic
Black
Asian
White
11
4
14
22
2.719
2.883
3.098
3.110
< 0.20*
FSH U/L Hispanic
Black
Asian
White
11
4
14
22
1.033
2.119
1.389
1.170
< 0.20*
*Bolded Indicates Statistically Significant One-Way ANOVA, p < 0.20. Means are natural-log concentrations.
Table 21: Bivariate Associations of Cigarette use and Ovarian Reserve Markers
Ovarian Reserve
Marker
Cigarette Use N Mean p
AMH ng/ml Yes
No
5
45
0.536
0.752
< 0.20**
Inhibin B ng/L Yes
No
5
45
2.059
2.985
< 0.20**
FSH U/L Yes
No
5
45
1.080
1.275
> 0.20
** Bolded Indicates Statistically Significant Independent Sample T tests, P <0.20. Means are natural-log
concentrations.
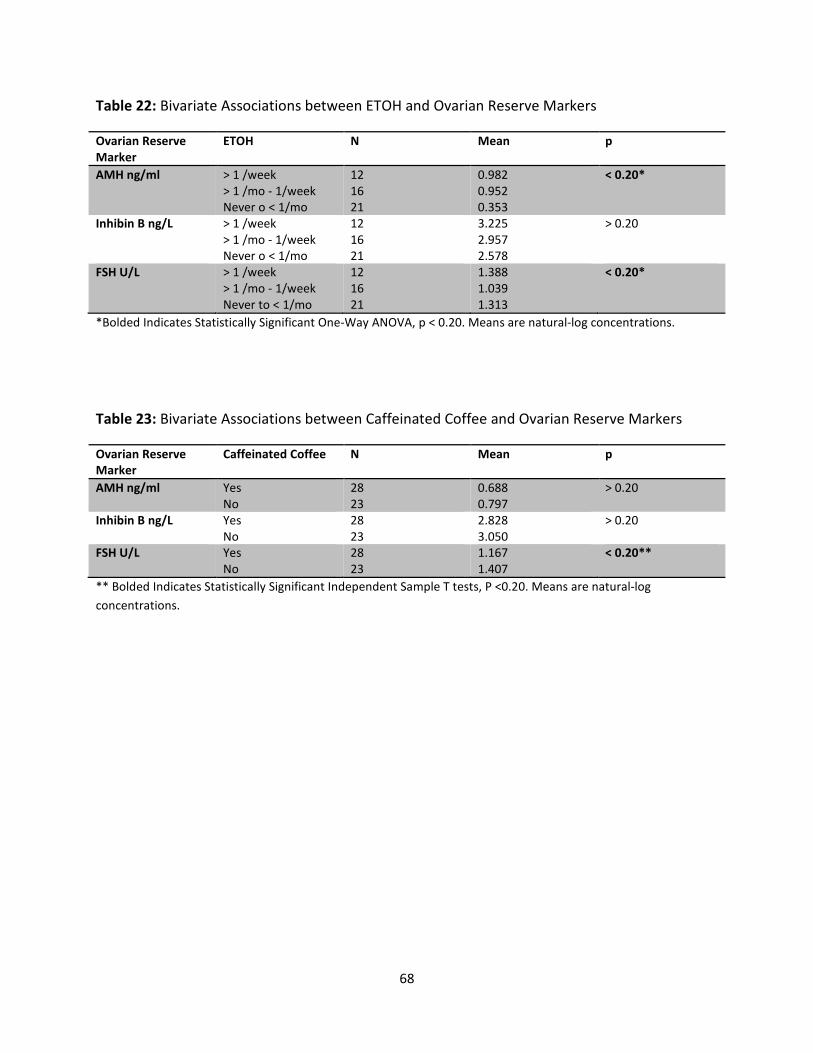
68
Table 22: Bivariate Associations between ETOH and Ovarian Reserve Markers
Ovarian Reserve
Marker
ETOH N Mean p
AMH ng/ml > 1 /week
> 1 /mo - 1/week
Never o < 1/mo
12
16
21
0.982
0.952
0.353
< 0.20*
Inhibin B ng/L > 1 /week
> 1 /mo - 1/week
Never o < 1/mo
12
16
21
3.225
2.957
2.578
> 0.20
FSH U/L > 1 /week
> 1 /mo - 1/week
Never to < 1/mo
12
16
21
1.388
1.039
1.313
< 0.20*
*Bolded Indicates Statistically Significant One-Way ANOVA, p < 0.20. Means are natural-log concentrations.
Table 23: Bivariate Associations between Caffeinated Coffee and Ovarian Reserve Markers
Ovarian Reserve
Marker
Caffeinated Coffee N Mean p
AMH ng/ml Yes
No
28
23
0.688
0.797
> 0.20
Inhibin B ng/L Yes
No
28
23
2.828
3.050
> 0.20
FSH U/L Yes
No
28
23
1.167
1.407
< 0.20**
** Bolded Indicates Statistically Significant Independent Sample T tests, P <0.20. Means are natural-log
concentrations.
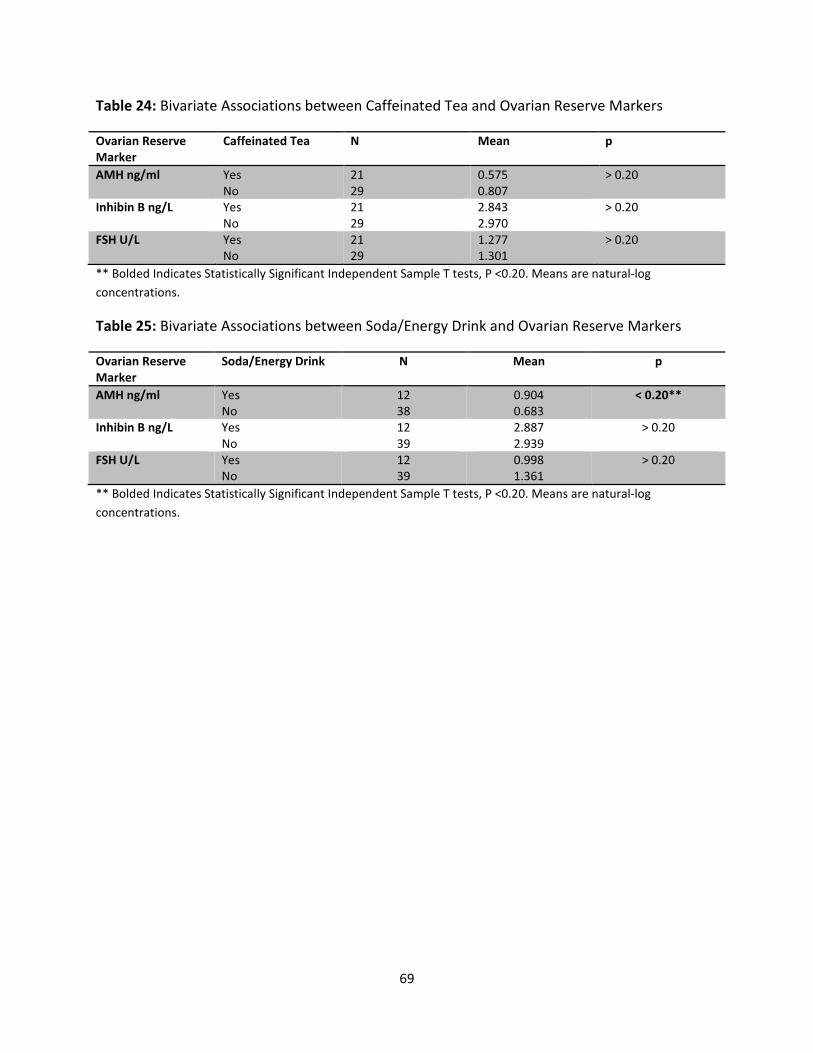
69
Table 24: Bivariate Associations between Caffeinated Tea and Ovarian Reserve Markers
Ovarian Reserve
Marker
Caffeinated Tea N Mean p
AMH ng/ml Yes
No
21
29
0.575
0.807
> 0.20
Inhibin B ng/L Yes
No
21
29
2.843
2.970
> 0.20
FSH U/L Yes
No
21
29
1.277
1.301
> 0.20
** Bolded Indicates Statistically Significant Independent Sample T tests, P <0.20. Means are natural-log
concentrations.
Table 25: Bivariate Associations between Soda/Energy Drink and Ovarian Reserve Markers
Ovarian Reserve
Marker
Soda/Energy Drink N Mean p
AMH ng/ml Yes
No
12
38
0.904
0.683
< 0.20**
Inhibin B ng/L Yes
No
12
39
2.887
2.939
> 0.20
FSH U/L Yes
No
12
39
0.998
1.361
> 0.20
** Bolded Indicates Statistically Significant Independent Sample T tests, P <0.20. Means are natural-log
concentrations.
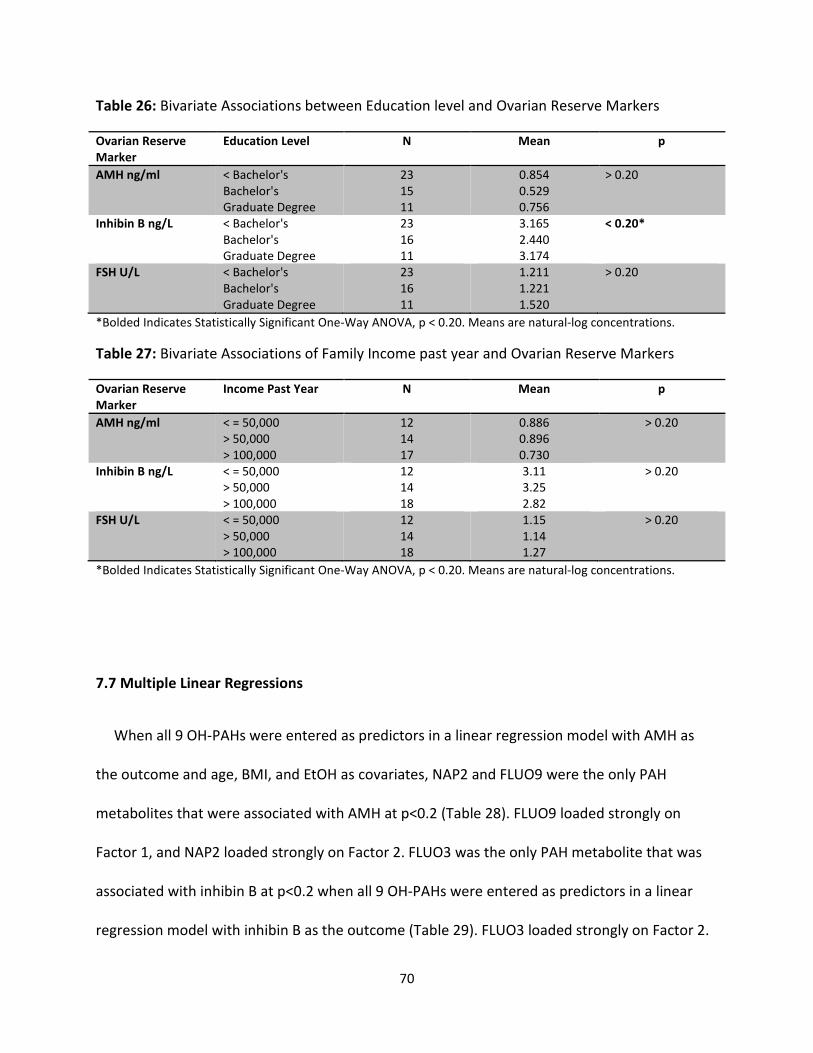
70
Table 26: Bivariate Associations between Education level and Ovarian Reserve Markers
Ovarian Reserve
Marker
Education Level N Mean p
AMH ng/ml < Bachelor's
Bachelor's
Graduate Degree
23
15
11
0.854
0.529
0.756
> 0.20
Inhibin B ng/L < Bachelor's
Bachelor's
Graduate Degree
23
16
11
3.165
2.440
3.174
< 0.20*
FSH U/L < Bachelor's
Bachelor's
Graduate Degree
23
16
11
1.211
1.221
1.520
> 0.20
*Bolded Indicates Statistically Significant One-Way ANOVA, p < 0.20. Means are natural-log concentrations.
Table 27: Bivariate Associations of Family Income past year and Ovarian Reserve Markers
Ovarian Reserve
Marker
Income Past Year N Mean p
AMH ng/ml < = 50,000
> 50,000
> 100,000
12
14
17
0.886
0.896
0.730
> 0.20
Inhibin B ng/L < = 50,000
> 50,000
> 100,000
12
14
18
3.11
3.25
2.82
> 0.20
FSH U/L < = 50,000
> 50,000
> 100,000
12
14
18
1.15
1.14
1.27
> 0.20
*Bolded Indicates Statistically Significant One-Way ANOVA, p < 0.20. Means are natural-log concentrations.
7.7 Multiple Linear Regressions
When all 9 OH-PAHs were entered as predictors in a linear regression model with AMH as
the outcome and age, BMI, and EtOH as covariates, NAP2 and FLUO9 were the only PAH
metabolites that were associated with AMH at p<0.2 (Table 28). FLUO9 loaded strongly on
Factor 1, and NAP2 loaded strongly on Factor 2. FLUO3 was the only PAH metabolite that was
associated with inhibin B at p<0.2 when all 9 OH-PAHs were entered as predictors in a linear
regression model with inhibin B as the outcome (Table 29). FLUO3 loaded strongly on Factor 2.

71
PHEN3 was the only PAH metabolite that was associated with FSH at p<0.2 when all 9 OH-PAHs
were entered as predictors in a linear regression model with FSH as the outcome (Table 30).
PHEN3 loaded strongly on Factor 1.
The next step in the modeling was to enter the potential confounders identified for each
outcome variable in the bivariate analyses into the regression models. These potential
confounders were retained in the final models only if they altered the adjusted beta
coefficients for the PAHs by 10% or more. Thus age, BMI, and EtOH were retained in the final
AMH model (Table 28), age and EtOH were retained in the final inhibin B model (Table 29), and
BMI and education were retained in the final FSH model (Table 30). The final AMH, inhibin B,
and FSH models explain 15.1 % , 27 % and 1.9% of the variations in the outcome variables
respectively (Tables 28-30).
Table 28: AMH Multiple Linear Regression Model
AMH = B 80% CI p
6.698 + 1.938, 11.458 0.007*
NAP2 - 0.222 - 0.675, 0.115 0.159
FLUO9 - 0.220 - 0.643, 0.096 0.142
Age - 0.233 - 0.068, 0.009 0.126
BMI - 0.183 - 0.072, 0.018 0.233
EtOH - 0.243 - 0.541, 0.062 0.116
Adjusted R squared 0.151 Model p = 0.042 B = Standardized Beta * Significant if p < 0.05. + = intercept
Table 29: Inhibin B Multiple Linear Regression Model
Inhibin B = B 80% CI p
6.963 + 5.192, 8.734 0.000*
FLUO3 - 0.117 - 0.549, 0.088 0.352
Age - 0.493 - 0.102, -0.052 0.000*
EtOH - 0.277 - 0.556, -0.147 0.030*
Adjusted R squared 0.270 Model p = 0.001 B = Standardized Beta * Significant if p < 0.05. + = intercept
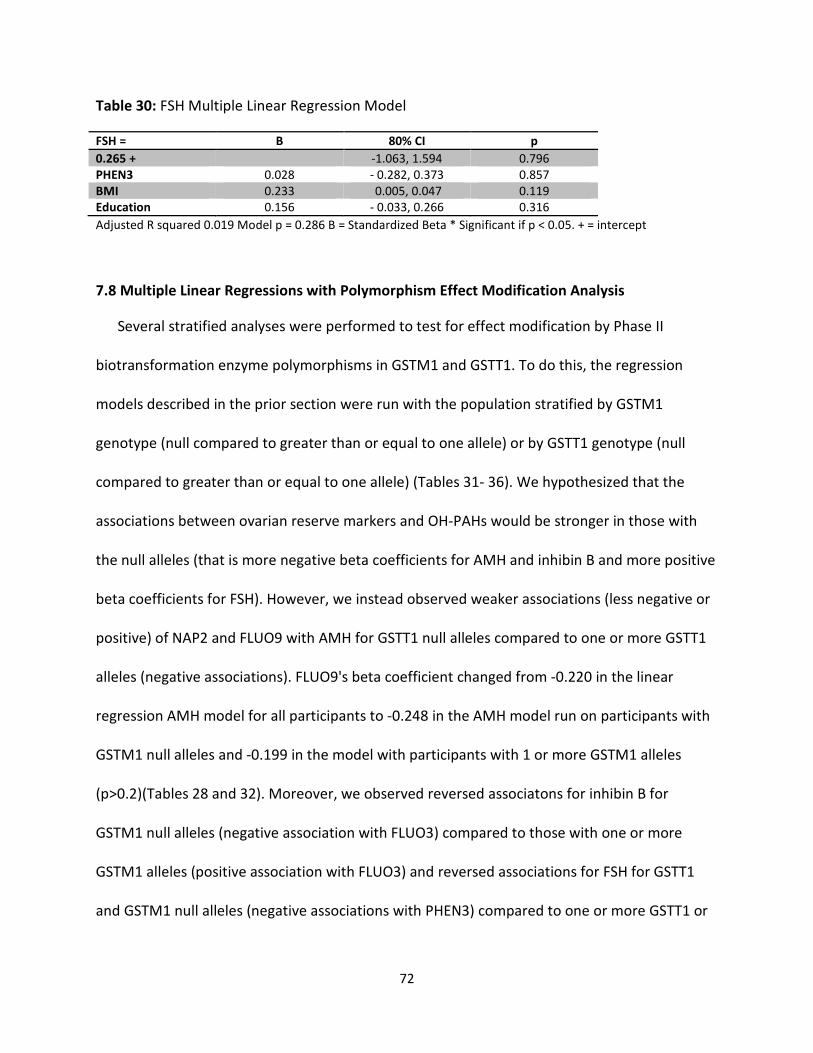
72
Table 30: FSH Multiple Linear Regression Model
FSH = B 80% CI p
0.265 + -1.063, 1.594 0.796
PHEN3 0.028 - 0.282, 0.373 0.857
BMI 0.233 0.005, 0.047 0.119
Education 0.156 - 0.033, 0.266 0.316
Adjusted R squared 0.019 Model p = 0.286 B = Standardized Beta * Significant if p < 0.05. + = intercept
7.8 Multiple Linear Regressions with Polymorphism Effect Modification Analysis
Several stratified analyses were performed to test for effect modification by Phase II
biotransformation enzyme polymorphisms in GSTM1 and GSTT1. To do this, the regression
models described in the prior section were run with the population stratified by GSTM1
genotype (null compared to greater than or equal to one allele) or by GSTT1 genotype (null
compared to greater than or equal to one allele) (Tables 31- 36). We hypothesized that the
associations between ovarian reserve markers and OH-PAHs would be stronger in those with
the null alleles (that is more negative beta coefficients for AMH and inhibin B and more positive
beta coefficients for FSH). However, we instead observed weaker associations (less negative or
positive) of NAP2 and FLUO9 with AMH for GSTT1 null alleles compared to one or more GSTT1
alleles (negative associations). FLUO9's beta coefficient changed from -0.220 in the linear
regression AMH model for all participants to -0.248 in the AMH model run on participants with
GSTM1 null alleles and -0.199 in the model with participants with 1 or more GSTM1 alleles
(p>0.2)(Tables 28 and 32). Moreover, we observed reversed associatons for inhibin B for
GSTM1 null alleles (negative association with FLUO3) compared to those with one or more
GSTM1 alleles (positive association with FLUO3) and reversed associations for FSH for GSTT1
and GSTM1 null alleles (negative associations with PHEN3) compared to one or more GSTT1 or

73
GSTM1 alleles (positive associations with PHEN3). FLUO3's beta coefficient changed from -0.117
in the linear regression inhibin B model for all participants to -0.342 in the inhibin B model run
for participants with GSTM1 null and to 0.066 for participants with 1 or more GSTM1 allele
(P<0.05)(Tables 29 and 34). The associations persisted when covariates such as BMI, EtOH, and
age were added or removed from the models (not shown). There was a significant association
between GSTM1 and AMH (p=0.04) in the bivariate analysis of the polymorphisms and the
ovarian reserve markers (data not shown). However, the GSTM1 genotype did not show a
significant effect modification in the AMH model (Table 32). Age remained significant in the
expected direction in the models run on participants in all of the models for inhibin B
(p<0.05)(Tables 33 and 34), except in the model run on participants with GSTM1 null genotype
(p>0.2)(Table 34). BMI was significant and in the expected direction for the model run on
participants with GSTM1 null genotype in the regression model for FSH (p<0.05)(Table 36). Age
did not reman a significant predictor of AMH in either model for effect modification by GST
polymorphisms constructed in this study (Tables 31 and 32).
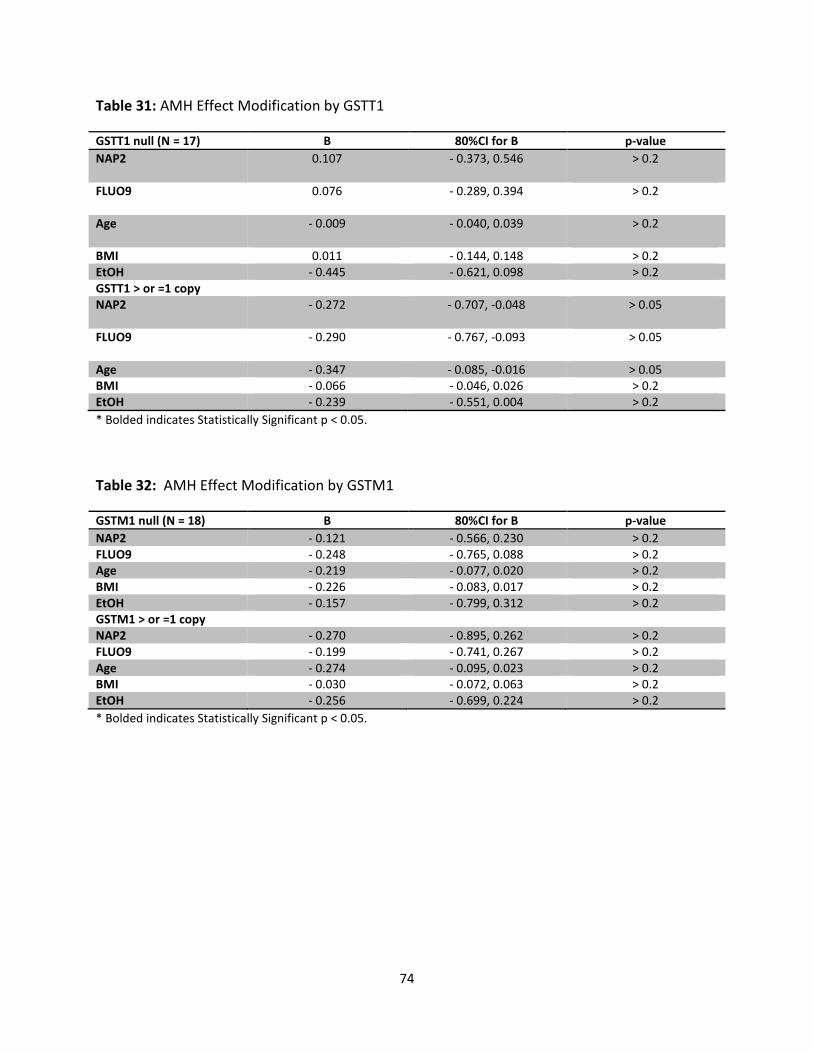
74
Table 31: AMH Effect Modification by GSTT1
GSTT1 null (N = 17) B 80%CI for B p-value
NAP2 0.107 - 0.373, 0.546 > 0.2
FLUO9 0.076 - 0.289, 0.394 > 0.2
Age - 0.009 - 0.040, 0.039 > 0.2
BMI 0.011 - 0.144, 0.148 > 0.2
EtOH - 0.445 - 0.621, 0.098 > 0.2
GSTT1 > or =1 copy
NAP2 - 0.272 - 0.707, -0.048 > 0.05
FLUO9 - 0.290 - 0.767, -0.093 > 0.05
Age - 0.347 - 0.085, -0.016 > 0.05
BMI - 0.066 - 0.046, 0.026 > 0.2
EtOH - 0.239 - 0.551, 0.004 > 0.2
* Bolded indicates Statistically Significant p < 0.05.
Table 32: AMH Effect Modification by GSTM1
GSTM1 null (N = 18) B 80%CI for B p-value
NAP2 - 0.121 - 0.566, 0.230 > 0.2
FLUO9 - 0.248 - 0.765, 0.088 > 0.2
Age - 0.219 - 0.077, 0.020 > 0.2
BMI - 0.226 - 0.083, 0.017 > 0.2
EtOH - 0.157 - 0.799, 0.312 > 0.2
GSTM1 > or =1 copy
NAP2 - 0.270 - 0.895, 0.262 > 0.2
FLUO9 - 0.199 - 0.741, 0.267 > 0.2
Age - 0.274 - 0.095, 0.023 > 0.2
BMI - 0.030 - 0.072, 0.063 > 0.2
EtOH - 0.256 - 0.699, 0.224 > 0.2
* Bolded indicates Statistically Significant p < 0.05.
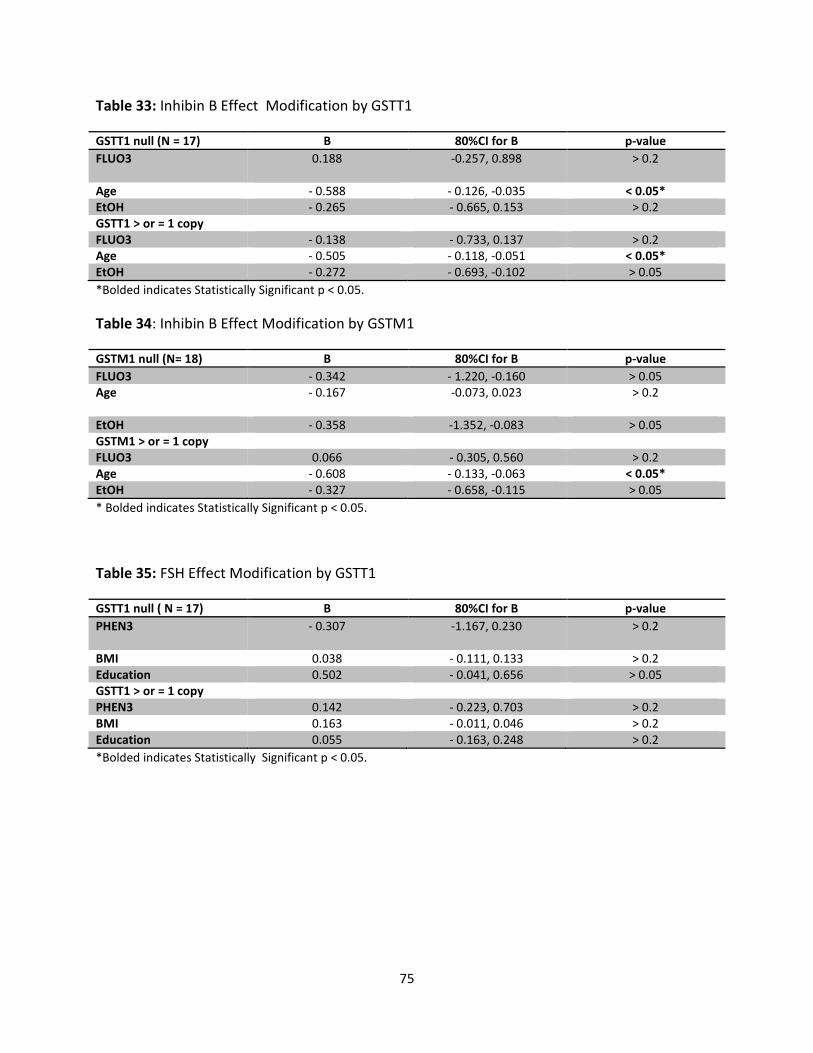
75
Table 33: Inhibin B Effect Modification by GSTT1
GSTT1 null (N = 17) B 80%CI for B p-value
FLUO3 0.188 -0.257, 0.898 > 0.2
Age - 0.588 - 0.126, -0.035 < 0.05*
EtOH - 0.265 - 0.665, 0.153 > 0.2
GSTT1 > or = 1 copy
FLUO3 - 0.138 - 0.733, 0.137 > 0.2
Age - 0.505 - 0.118, -0.051 < 0.05*
EtOH - 0.272 - 0.693, -0.102 > 0.05
*Bolded indicates Statistically Significant p < 0.05.
Table 34: Inhibin B Effect Modification by GSTM1
GSTM1 null (N= 18) B 80%CI for B p-value
FLUO3 - 0.342 - 1.220, -0.160 > 0.05
Age - 0.167 -0.073, 0.023 > 0.2
EtOH - 0.358 -1.352, -0.083 > 0.05
GSTM1 > or = 1 copy
FLUO3 0.066 - 0.305, 0.560 > 0.2
Age - 0.608 - 0.133, -0.063 < 0.05*
EtOH - 0.327 - 0.658, -0.115 > 0.05
* Bolded indicates Statistically Significant p < 0.05.
Table 35: FSH Effect Modification by GSTT1
GSTT1 null ( N = 17) B 80%CI for B p-value
PHEN3 - 0.307 -1.167, 0.230 > 0.2
BMI 0.038 - 0.111, 0.133 > 0.2
Education 0.502 - 0.041, 0.656 > 0.05
GSTT1 > or = 1 copy
PHEN3 0.142 - 0.223, 0.703 > 0.2
BMI 0.163 - 0.011, 0.046 > 0.2
Education 0.055 - 0.163, 0.248 > 0.2
*Bolded indicates Statistically Significant p < 0.05.
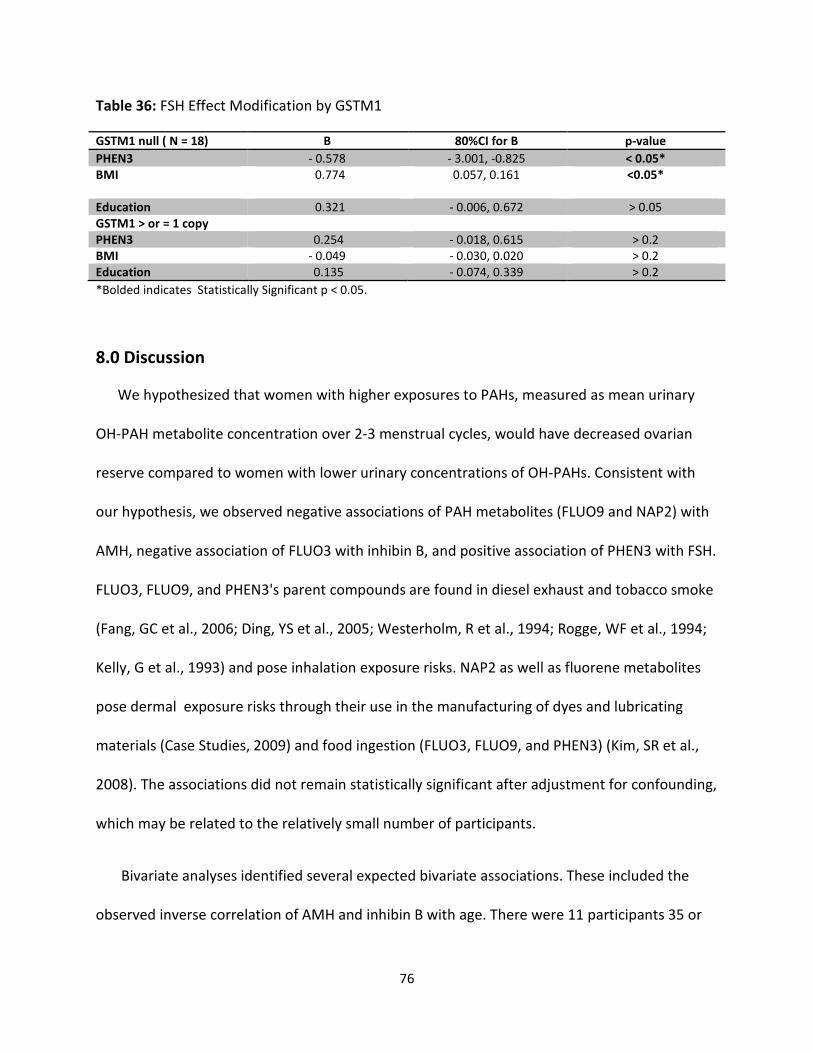
76
Table 36: FSH Effect Modification by GSTM1
GSTM1 null ( N = 18) B 80%CI for B p-value
PHEN3 - 0.578 - 3.001, -0.825 < 0.05*
BMI 0.774 0.057, 0.161 <0.05*
Education 0.321 - 0.006, 0.672 > 0.05
GSTM1 > or = 1 copy
PHEN3 0.254 - 0.018, 0.615 > 0.2
BMI - 0.049 - 0.030, 0.020 > 0.2
Education 0.135 - 0.074, 0.339 > 0.2
*Bolded indicates Statistically Significant p < 0.05.
8.0 Discussion
We hypothesized that women with higher exposures to PAHs, measured as mean urinary
OH-PAH metabolite concentration over 2-3 menstrual cycles, would have decreased ovarian
reserve compared to women with lower urinary concentrations of OH-PAHs. Consistent with
our hypothesis, we observed negative associations of PAH metabolites (FLUO9 and NAP2) with
AMH, negative association of FLUO3 with inhibin B, and positive association of PHEN3 with FSH.
FLUO3, FLUO9, and PHEN3's parent compounds are found in diesel exhaust and tobacco smoke
(Fang, GC et al., 2006; Ding, YS et al., 2005; Westerholm, R et al., 1994; Rogge, WF et al., 1994;
Kelly, G et al., 1993) and pose inhalation exposure risks. NAP2 as well as fluorene metabolites
pose dermal exposure risks through their use in the manufacturing of dyes and lubricating
materials (Case Studies, 2009) and food ingestion (FLUO3, FLUO9, and PHEN3) (Kim, SR et al.,
2008). The associations did not remain statistically significant after adjustment for confounding,
which may be related to the relatively small number of participants.
Bivariate analyses identified several expected bivariate associations. These included the
observed inverse correlation of AMH and inhibin B with age. There were 11 participants 35 or

77
older in this study. The inverse correlations of AMH and inhibin B with BMI was also expected.
FSH had positive correlations with age and BMI, but the correlations were low and not
statistically significant. We expected these results since AMH and inhibin B are made by the
granulosa cells, and AMH doesn’t fluctuate with the menstrual cycle phase. AMH is thus poorly
correlated with both FSH and inhibin B. FSH is not made by the granulosa cells but gets
feedback inhibition of E2 from growing follicles. Early reproductive age women predominate in
this study, with only 6 participants over 37 years old. Six participants had BMIs over 30 in this
study. Su et al., found obesity (BMI > 30) in late reproductive age (40-52) women lowered
serum AMH (77%, p 0.02) and inhibin B (24%, p = 0.08) compared to normal weight (BMI < 25)
women, but had no effect on serum FSH and E2 levels. The group concluded that the lower
AMH levels resulted from physiologic processes other than decreased ovarian reserve (Su, HI et
al., 2008). The finding of lower AMH levels among blacks compared to all other participants,
and the higher FSH levels in blacks likely reflected random variation.
During the construction of the final multiple regression models, we regressed all of the PAHs
against each ovarian reserve marker separately and observed the expected directions of
associations between the PAHs and the ovarian reserve models with most of the PAHs. At least
one of the PAHs was observed to have an opposite association with a marker of ovarian reserve
(FLUO3 and FSH), but this association did not approach statistical significance.
The multiple linear regressions were limited by the small sample size, the collinearity of the
PAHs (Tables 5-7), the fact that ovarian reserve markers inhibin B and FSH fluctuate with the
menstrual cycle , and baseline serum collections of ovarian reserve markers were outside of the

78
study design intent to collect on 5 to 9 days prior to the next expected menstruation. These
factors explain why two of the three final models were not significant (Tables 29 and 30, inhibin
B , p = 0.352 and FSH, p = 0.857). All of the aforementioned may also explain the strength of the
OH-PAHs' associations in the multiple linear regressions with the ovarian reserve markers when
all 9 OH-PAHs were included in the regressions. Unexplained confounding may have also played
a role in the outcomes of the regression models, as we had 7 missing values in income (Table 1),
which precluded it from our regression models as a surrogate for SES, over half (28 or 55%) of
the participants drank coffee (Table 1) which is a source of PAHs and was therefore excluded
from the regression analysis as well as cigarette use, which was under-represented in this small
sample size (5 participants smoked cigarettes, Table 1).
The collinearity of the PAHs was addressed in the principal component factor analysis with
varimax rotation (Table 8 and Figure 6), which revealed that 34% of the cumulative variance
was explained by all of the PAH metabolites in the study except for NAP2, and was
predominated by all of the phenanthrene metabolites evaluated in the study, including PHEN3
(0.762) which was retained in the model with FSH (p= 0.857) and FLUO9 (p=0.735), which was
retained in the model with AMH (p = 0.142). About 64.% of the cumulative variance was
explained predominantly by FLUO3 (which had a principal component factor analysis
coefficient of 0.852) which was retained in the model with inhibin B (p= 0.352) and FLUO2
(0.771), and the parent compound naphthalene, where NAP2 (0.487) was retained in the model
with AMH (p = 0.159). The Principal Component analysis identified weights of individual OH-
PAHs in the overall mixture of PAHs which does not address whether they were associated
with the outcome variables. For example, even though all of the OH-metabolites of

79
phenanthrene loaded stronger on Factor 1 compared to FLUO9 and NAP2 loaded less strongly
on Factor 2 compared to NAP1, FLUO2, and FLUO3, the multivariable analysis selected OH-PAHs
(different) that were most strongly associated with the 3 outcome variables in the models
(Tables 28 - 30). The clinical importance of associating specific PAHs with the various ovarian
reserve monitors would help construct future exposure risk models for various exposures to
PAHs as they exist in mixtures. However, there was not enough evidence in this small pilot
study to reveal which specific PAHs are associated with the different markers of ovarian
reserve. Despite the use of principal component analysis, the sample sizes were too small (the
resulting factor analysis supported the associations found in the adjusted regression analysis).
Overall, the PAHs were not significant (p > 0.05) in the models compared to the confounders
(Tables 29 - 30).
Our cohort had similar mean PAH levels as the NHANES data (Table 4), and therefore,
although we had a small convenience sample, we had good representation of excretion profiles
for the part of the general US population of interest in this study. We found significant
associations between all of the parent PAHs and cigarette use as expected (Table 13) even
though we had a small number (5) of participants who reported cigarette use (9.8%) (Van Rooij,
JG et al., 1994). More than half (55%) of the participants reported drinking coffee, however we
did not obtain information on whether they drank boiled and/or brewed coffee (brewed
contains more PAHs) (Tfouni, SAV et al., 2013). This may explain why we only found significant
associations between two PAH parent compounds and coffee drinkers (Table 15). We observed
associations between EtOH and 4 OH-PAHs (NAP1, PHEN1, PHEN3, and PYR1), (p < 0.20) when
all 9 OH-PAHs were evaluated, however, no consistent patterns were observed (Table 14).

80
There is a dose-dependent use of CYP2E1 to metabolize alcohol with acute alcohol intake (Seitz,
HK et al., 2006), however, the use of this enzyme to explain the individual participants' ovarian
reserve marker levels or PAH levels as they relate to EtOH were not examined in this thesis.
Moreover, Van Rooij et al., did not find a significant association between alcohol consumption
and baseline urinary excretion of PYR1 in non-occupationally exposed participants compared to
the major significant findings associated with their dietary PAH intake, age, and tobacco
consumption (Van Rooij, JG et al., 1994). This study did not find significant associations
between PAHs and age or BMI (Table 10) which was expected due to the narrow age-range (18-
44 years) and BMI range (SD 5.29) of the study participants (Table 1).
Effect modification analysis was use to assess whether inter-individual variations in PAH
ovotoxicity could be explained by polymorphisms in the enzymes GSTM1 and GSTT1 that
detoxify PAHs. The results did not prove or disprove our hypothesis. The negative association of
FLUO9 with GSTM1 null in the AMH model was the strongest but not significant (Table 32).
Evaluations of the GSTM1 null and GSTT1 null did not reveal any associations between the PAHs
in the inhibin B and FSH models (Tables 33 -36), partly because of the small sample size, which
leads to the wide confidence intervals. GST polymorphisms have been associated with some
PAHs in some studies and not in others, and therefore our findings were not entirely surprising
(Liu, M et al., 2013; Gao, Q et al., 2011; Mc Carty, KM et al., 2009).
Our stratified regression models revealed associations between OH-PAHs and ovarian
reserve markers in the unexpected (though insignificant) directions for the strata with null
alleles for inhibin B and AMH in the GSTT1 null regression and for FSH in both the GSTT1 and

81
GSTM1 null regressions. The nonsignificant associations persisted when covariates such as BMI,
ETOH, and age were added or removed from the models (data not shown). Both non-deleted
GST genotypes in this study changed PAH coefficients in directions expected for the deleted
genotypes and there were stronger associations with PAHs in the non-deleted genotypes
compared to the deleted group, however, none were significant. Interestingly, despite the
significant association between GSTM1 and AMH (p = 0.04) in our bivariate analysis of the
polymorphisms and the ovarian reserve markers (data not shown), we did not observe a GSTM1
null genotype-effect modification in the AMH model. This may indicate that GST is involved in
the metabolism of AMH. To the best of the author's knowledge, there are no reports of GST's
associated with ovarian reserve markers as we observed in our bivariate analysis between
GSTM1 null and AMH (data not shown).
Limitations of this study included a small sample size and our inability to collect serum
samples for the ovarian reserve markers in the study time-frame. If the samples were collected
in the study time-frame (5 to 9 days prior to the next expected menstrual cycle), then we may
have been able to capture more meaningful differences in the FSH levels and possibly have
more significant associations with OH-PAHs. Significant differences between covariate groups
with respect to the outcome variables could have been assessed by using pairwise t-testing. The
results could have been compared with prior research on these topics (Kinney, A et al., 2007),
however, that was not the focus of this thesis.
This pilot study demonstrated that the recruitment strategies, sample collection procedures,
measurements used, and biomarker used are efficacious and would serve as robust analysis of

82
the hypotheses addressed in this thesis in a larger sample size. It also validated the techniques
used for genotyping polymorphisms in PAH metabolizing enzyme genes. To the best of authors'
knowledge, this is the first study evaluating the effects of PAHs on ovarian reserve in human
subjects along with whether their effects are modified by polymorphisms in PAH-detoxifying
metabolic enzymes.
9.0 Conclusion
In conclusion, we analyzed the long-term effects of exposure to PAHs on ovarian reserve
markers, AMH, inhibin B, and FSH and whether there were effect modifications by GSTM1 and
GSTT1 genetic polymorphisms in a cohort of reproductive age women. We found a trend
suggesting that women exposed to higher concentrations of PAHs over time would have lower
ovarian reserve as measured by serum markers compared to women exposed to lower
concentrations. The results of this pilot study, although limited by statistical power, adds to the
body of knowledge in the field of reproductive toxicology with regards to PAH-induced
ovotoxicity. We did not prove or disprove PAH-induced ovotoxicity in this particular pilot study.
Nor did we illustrate that GSTM1 null and GSTT1 null polymorphisms add to the effects of the
PAH-induced decreased ovarian reserve.
To the best of the authors' knowledge, this is the first study to investigate the association
between PAHs and diminished ovarian reserve in humans and possible mechanisms of inter-
individual variations in those associations with effect modification analysis involving enzyme
polymorphisms (GST's) in the detoxification of PAHs. This study is a step in the right direction
towards a preventative approach to diminished ovarian reserve and contributes to the body of

83
knowledge aimed at addressing air pollution policy and smoking cessation efforts. This study
could be an inspiration to nutrition investigators to design studies involving PAHs that would
help create guideline recommendations for consumption and processing of red meat (Kaiser, J
1997; Browner, CM 1997; Lijinsky, W et al., 1964; Lioy, PJ 1988). This pilot study also illustrated
the feasibility of the study design and methods. Finally, data from this study (small sample size)
could be used in a power analysis for a larger study examining the associations between OH-
PAHs and ovarian reserve markers.

84
REFERENCES
Abnet, C. C., Fagundes, R. B., Strickland, P. T., Kamangar, F., Roth, M. J., Taylor, P. R., & Dawsey,
S. M. (2007). The influence of genetic polymorphisms in Ahr, CYP1A1, CYP1A2, CYP1B1,
GST M1, GST T1 and UGT1A1 on urine 1-hydroxypyrene glucuronide concentrations in
healthy subjects from Rio Grande do Sul, Brazil. Carcinogenesis, 28(1), 112-117.
doi:10.1093/carcin/bgl131
Adashi, E. Y. (1994). Endocrinology of the ovary. Hum Reprod, 9(5), 815-827.
Agency for Toxic Substances and Disease Registry. (June 24, 2013).
Agency for Toxic Substances and Disease Registry (ATSDR). (1995). Toxicological Profile for
Polycyclic Aromatic Hydrocarbons 1995 [online].
Al-Qahtani, A., Al-Ahdal, M., Abdo, A., Sanai, F., Al-Anazi, M., Khalaf, N., . . . Almajhdi, F. N.
(2012). Toll-like receptor 3 polymorphism and its association with hepatitis B virus
infection in Saudi Arabian patients. J Med Virol, 84(9), 1353-1359.
doi:10.1002/jmv.23271
Al-Qahtani, A., Al-Anazi, M., Viswan, N. A., Khalaf, N., Abdo, A. A., Sanai, F. M., . . . Al-Ahdal, M.
(2012). Role of single nucleotide polymorphisms of KIF1B gene in HBV-associated viral
hepatitis. PLoS One, 7(9), e45128. doi:10.1371/journal.pone.0045128
Al-Qahtani, A. A., Kessie, G., Dela Cruz, D., Al-Faleh, F. Z., & Al-Ahdal, M. N. (2010).
Quasispecies of genotype 4 of hepatitis C virus genomes in Saudi patients managed
with interferon alfa and ribavirin therapy. Ann Saudi Med, 30(2), 109-114.
doi:10.4103/0256-4947.60515
Alexandrie, A. K., Warholm, M., Carstensen, U., Axmon, A., Hagmar, L., Levin, J. O., . . . Rannug,
A. (2000). CYP1A1 and GSTM1 polymorphisms affect urinary 1-hydroxypyrene levels
after PAH exposure. Carcinogenesis, 21(4), 669-676.
Altshuler, D., Brooks, L., Chakravart, A., Collins, F., Daly, M., & Donnelly, P. (2005). A Haplotype
Map of the Human Genome. Nature, 437(Consortium IH), 1299-1320.
Anderson, L. M., Jones, A. B., Miller, M. S., & Chauhan, D. P. (1989). Metabolism of
transplacental carcinogens. IARC Sci Publ(96), 155-188.
Angerer, J., Mannschreck, C., & Gündel, J. (1997a). Biological monitoring and biochemical
effect monitoring of exposure to polycyclic aromatic hydrocarbons. Int Arch Occup
Environ Health, 70(6), 365-377.

85
Angerer, J., Mannschreck, C., & Gündel, J. (1997b). Occupational exposure to polycyclic
aromatic hydrocarbons in a graphite-electrode producing plant: biological monitoring
of 1-hydroxypyrene and monohydroxylated metabolites of phenanthrene. Int Arch
Occup Environ Health, 69(5), 323-331.
Anzenbacher, P., & Zanger, U. (2012). Metabolism of Drugs and other Xenobiotics. Wiley
Online Library: Wiley.
Archibong, A. E., Ramesh, A., Inyang, F., Niaz, M. S., Hood, D. B., & Kopsombut, P. (2012).
Endocrine disruptive actions of inhaled benzo(a)pyrene on ovarian function and fetal
survival in fisher F-344 adult rats. Reprod Toxicol, 34(4), 635-643.
doi:10.1016/j.reprotox.2012.09.003
ATSDR, Agency for Toxic Substances Disease Registry. (2005).
Bakke, J., Struble, C., Gustafsson, J. A., & Gustafsson, B. (1985). Catabolism of premercapturic
acid pathway metabolites of naphthalene to naphthols and methylthio-containing
metabolites in rats. Proc Natl Acad Sci U S A, 82(3), 668-671.
Barbakadze, L., Kristesashvili, J., Khonelidze, N., & Tsagareishvili, G. (2015). The correlations of
anti-mullerian hormone, follicle-stimulating hormone and antral follicle count in
different age groups of infertile women. Int J Fertil Steril, 8(4), 393-398.
Barbeau, D., Persoons, R., Marques, M., Hervé, C., Laffitte-Rigaud, G., & Maitre, A. (2014).
Relevance of urinary 3-hydroxybenzo(a)pyrene and 1-hydroxypyrene to assess
exposure to carcinogenic polycyclic aromatic hydrocarbon mixtures in metallurgy
workers. Ann Occup Hyg, 58(5), 579-590. doi:10.1093/annhyg/meu004
Barr, D. B., Wilder, L. C., Caudill, S. P., Gonzalez, A. J., Needham, L. L., & Pirkle, J. L. (2005).
Urinary creatinine concentrations in the U.S. population: implications for urinary
biologic monitoring measurements. Environ Health Perspect, 113(2), 192-200.
Bengtsson, M., & Mattison, D. R. (1989). Gonadotropin-dependent metabolism of 7,12-
dimethylbenz(a)anthracene in the ovary of rhesus monkey. Biochem Pharmacol, 38(11),
1869-1872.
Bhattacharya, P., & Keating, A. F. (2012). Impact of environmental exposures on ovarian
function and role of xenobiotic metabolism during ovotoxicity. Toxicol Appl Pharmacol,
261(3), 227-235. doi:10.1016/j.taap.2012.04.009
Bieniek, G. (1994). The presence of 1-naphthol in the urine of industrial workers exposed to
naphthalene. Occup Environ Med, 51(5), 357-359.

86
Boeniger, M. F., Lowry, L. K., & Rosenberg, J. (1993). Interpretation of urine results used to
assess chemical exposure with emphasis on creatinine adjustments: a review. Am Ind
Hyg Assoc J, 54(10), 615-627. doi:10.1080/15298669391355134
Bogovski, P. (1983). IARC Monographs On The Evaluations on the Carcinogenic Risk Of
Chemicals To Humans. Retrieved from Lyon, France:
Boström, C. E., Gerde, P., Hanberg, A., Jernström, B., Johansson, C., Kyrklund, T., . . .
Westerholm, R. (2002). Cancer risk assessment, indicators, and guidelines for polycyclic
aromatic hydrocarbons in the ambient air. Environ Health Perspect, 110 Suppl 3, 451-
488.
Boundless. (2013). Hormone regulation of the female reproductive cycle Boundless Anatomy
and Physiology (Vol. 4).
Bourne, M. C., & Young, L. (1934). The metabolism of naphthalene in rabbits. Biochem J, 28(3),
803-808.
Broekmans, F. J., Visser, J. A., Laven, J. S., Broer, S. L., Themmen, A. P., & Fauser, B. C. (2008a).
Anti-Müllerian hormone and ovarian dysfunction. Trends Endocrinol Metab, 19(9), 340-
347. doi:10.1016/j.tem.2008.08.002
Broekmans, F. J., Visser, J. A., Laven, J. S., Broer, S. L., Themmen, A. P., & Fauser, B. C. (2008b).
Anti-Müllerian hormone and ovarian dysfunction. Trends Endocrinol Metab, 19(9), 340-
347. doi:10.1016/j.tem.2008.08.002
Buchet, J. P., Gennart, J. P., Mercado-Calderon, F., Delavignette, J. P., Cupers, L., & Lauwerys, R.
(1992). Evaluation of exposure to polycyclic aromatic hydrocarbons in a coke
production and a graphite electrode manufacturing plant: assessment of urinary
excretion of 1-hydroxypyrene as a biological indicator of exposure. Br J Ind Med,
49(11), 761-768.
Buckley, T. J., & Lioy, P. J. (1992a). An examination of the time course from human dietary
exposure to polycyclic aromatic hydrocarbons to urinary elimination of 1-
hydroxypyrene. Br J Ind Med, 49(2), 113-124.
Buckley, T. J., & Lioy, P. J. (1992b). An examination of the time course from human dietary
exposure to polycyclic aromatic hydrocarbons to urinary elimination of 1-
hydroxypyrene. Br J Ind Med, 49(2), 113-124.
Buratti, M., Campo, L., Fustinoni, S., Cirla, P. E., Martinotti, I., Cavallo, D., & Foa, V. (2007).
Urinary hydroxylated metabolites of polycyclic aromatic hydrocarbons as biomarkers of
exposure in asphalt workers. Biomarkers, 12(3), 221-239.
doi:10.1080/13547500601100110

87
Byeong-Kyu, L., & Vu, V. T. (2010). Sources, Distribution and Toxicity of Polycyclic Aromatic
Hydrocarbons (PAHS) in Particulate Matter: CC BY-NC-SA3.0.
Béranger, R., Hoffmann, P., Christin-Maitre, S., & Bonneterre, V. (2012). Occupational
exposures to chemicals as a possible etiology in premature ovarian failure: a critical
analysis of the literature. Reprod Toxicol, 33(3), 269-279.
doi:10.1016/j.reprotox.2012.01.002
Campo, L., Addario, L., Buratti, M., Scibetta, L., Longhi, O., Valla, C., . . . Fustinoni, S. (2006).
Biological monitoring of exposure to polycyclic aromatic hydrocarbons by
determination of unmetabolized compounds in urine. Toxicol Lett, 162(2-3), 132-138.
doi:10.1016/j.toxlet.2005.09.024
Cancer Epidemiological Biomarkers Preview. (October 1, 2006). (vol 15 no. 10 pages 1805-1811
ed.).
Cannady, E. A., Dyer, C. A., Christian, P. J., Sipes, I. G., & Hoyer, P. B. (2002). Expression and
activity of microsomal epoxide hydrolase in follicles isolated from mouse ovaries.
Toxicol Sci, 68(1), 24-31.
Carlsson, I. B., Scott, J. E., Visser, J. A., Ritvos, O., Themmen, A. P., & Hovatta, O. (2006). Anti-
Müllerian hormone inhibits initiation of growth of human primordial ovarian follicles in
vitro. Hum Reprod, 21(9), 2223-2227. doi:10.1093/humrep/del165
Case Studies in Environmental Medicine Toxicity of Polycyclic Aromatic Hydrocarbons (PAHs).
(2009). Agency for Toxic Substances and Disease Registry.
Cattaneo, A., Taronna, M., Consonni, D., Angius, S., Costamagna, P., & Cavallo, D. M. (2010).
Personal exposure of traffic police officers to particulate matter, carbon monoxide, and
benzene in the city of Milan, Italy. J Occup Environ Hyg, 7(6), 342-351.
doi:10.1080/15459621003729966
Center for Disease Control and Prevention, Infertility. (2015).
Center for Disease Control and Prevention CDC 24/7. (2013).
Cerniglia, C. E., Van Baalen, C., & Gibson, D. T. (1980). Metabolism of Naphthalene by the
Cyanobacterium. Oscillatoria sp, Strain JCM, 116, 485-494.
Chaloupka, K., Santostefano, M., Goldfarb, I. S., Liu, G., Myers, M. J., Tsyrolv, I. B., . . . Safe, S.
(1994). Aryl hydrocarbon (Ah) receptor-independent induction of Cyp1a2 gene
expression by acenaphthylene and related compounds in B6C3F1 mice. Carcinogenesis,
15(12), 2835-2840.

88
Chandra, A., Martinez, G. M., Mosher, W. D., Abma, J. C., & Jones, J. (2005). Fertility, family
planning, and reproductive health of U.S. women: data from the 2002 National Survey
of Family Growth. Vital Health Stat 23(25), 1-160.
Chen, S. L., Xia, R., Chen, X., Luo, Y. Q., Wang, L. L., Wu, Y. Q., . . . Zheng, H. Y. (2011).
[Prediction of ovarian reserve, poor response and pregnancy outcome based on basal
antral follicle count and age in patients undergoing in vitro fertilization-embryo
transfer]. Nan Fang Yi Ke Da Xue Xue Bao, 31(4), 572-577.
Clark, C. R., Dutcher, J. S., Brooks, A. L., McClellan, R. O., Marshall, W. F., & Naman, T. M.
(1982). Mutagenicity of diesel exhaust particle extracts: influence of driving cycle and
environmental temperature. Fundam Appl Toxicol, 2(4), 153-157.
Clark, C. R., McClellan, R. O., Marshall, W. F., Naman, T. M., & Seizinger, D. E. (1982).
Mutagenicity of diesel exhaust particle extracts: influence of non-petroleum fuel
extenders. Arch Environ Contam Toxicol, 11(6), 749-752.
Collin, Gerd Höke, Hartmut Greim, Helmut. (2003). (Weinheim Ed.): Wiley-VCH.
Coulam, C. B. (1982). Premature gonadal failure. Fertil Steril, 38(6), 645-655.
Coulam, C. B., Adamson, S. C., & Annegers, J. F. (1986). Incidence of premature ovarian failure.
Obstet Gynecol, 67(4), 604-606.
Crofts, F., Taioli, E., Trachman, J., Cosma, G. N., Currie, D., Toniolo, P., & Garte, S. J. (1994).
Functional significance of different human CYP1A1 genotypes. Carcinogenesis, 15(12),
2961-2963.
Crowell, S. R., Hanson-Drury, S., Williams, D. E., & Corley, R. A. (2014). In vitro metabolism of
benzo[a]pyrene and dibenzo[def,p]chrysene in rodent and human hepatic microsomes.
Toxicol Lett, 228(1), 48-55. doi:10.1016/j.toxlet.2014.04.004
de Vet, A., Laven, J. S., de Jong, F. H., Themmen, A. P., & Fauser, B. C. (2002). Antimüllerian
hormone serum levels: a putative marker for ovarian aging. Fertil Steril, 77(2), 357-362.
Dekker, M., Harvey, P., & Heindel, D. a. (1999). Two-cell Two-gonadotropin Theory of Ovarian
Steroid Synthesis. In D. a. Heindel (Ed.), Reproductive and Developmental Toxicology
(Vol. 10, pp. 772). New York: John Wiley & Sons, Ltd.
Denison, M. S., & Nagy, S. R. (2003). Activation of the aryl hydrocarbon receptor by structurally
diverse exogenous and endogenous chemicals. Annu Rev Pharmacol Toxicol, 43, 309-
334. doi:10.1146/annurev.pharmtox.43.100901.135828

89
Devoto, L., Vega, M., Kohen, P., Castro, A., Castro, O., Christenson, L. K., . . . Strauss, J. F.
(2000). Endocrine and paracrine-autocrine regulation of the human corpus luteum
during the mid-luteal phase. J Reprod Fertil Suppl, 55, 13-20.
Dewailly, D., Andersen, C. Y., Balen, A., Broekmans, F., Dilaver, N., Fanchin, R., . . . Anderson, R.
A. (2014). The physiology and clinical utility of anti-Mullerian hormone in women. Hum
Reprod Update, 20(3), 370-385. doi:10.1093/humupd/dmt062
Dewhurst, F. (1962). The hydroxylation of fluorene in the rat and the rabbit. Br J Cancer, 16,
371-377.
Ding, Y. S., Trommel, J. S., Yan, X. J., Ashley, D., & Watson, C. H. (2005). Determination of 14
polycyclic aromatic hydrocarbons in mainstream smoke from domestic cigarettes.
Environ Sci Technol, 39(2), 471-478.
Ding, Y. S., Zhang, L., Jain, R. B., Jain, N., Wang, R. Y., Ashley, D. L., & Watson, C. H. (2008).
Levels of tobacco-specific nitrosamines and polycyclic aromatic hydrocarbons in
mainstream smoke from different tobacco varieties. Cancer Epidemiol Biomarkers Prev,
17(12), 3366-3371. doi:10.1158/1055-9965.EPI-08-0320
Disdier, B., Arfi, C., Pastor, J., & Pauline, A. (2000). Analysis by GC-MS of Polycyclic Aromatic
Hydrocarbons in a Cream Containing Coal Tar. Polycyclic Aromatic Compounds, 10(1-4),
169-177.
Dockery, D. W., Pope, C. A., Xu, X., Spengler, J. D., Ware, J. H., Fay, M. E., . . . Speizer, F. E.
(1993). An association between air pollution and mortality in six U.S. cities. N Engl J
Med, 329(24), 1753-1759. doi:10.1056/NEJM199312093292401
Domingo, J. L., & Nadal, M. (2015). Human dietary exposure to polycyclic aromatic
hydrocarbons: A review of the scientific literature. Food Chem Toxicol, 86, 144-153.
doi:10.1016/j.fct.2015.10.002
Dor, F., Dab, W., Empereur-Bissonnet, P., & Zmirou, D. (1999). Validity of biomarkers in
environmental health studies: the case of PAHs and benzene. Crit Rev Toxicol, 29(2),
129-168. doi:10.1080/10408449991349195
Durlinger, A. L., Kramer, P., Karels, B., de Jong, F. H., Uilenbroek, J. T., Grootegoed, J. A., &
Themmen, A. P. (1999). Control of primordial follicle recruitment by anti-Müllerian
hormone in the mouse ovary. Endocrinology, 140(12), 5789-5796.
doi:10.1210/endo.140.12.7204
Durlinger, A. L., Visser, J. A., & Themmen, A. P. (2002). Regulation of ovarian function: the role
of anti-Müllerian hormone. Reproduction, 124(5), 601-609.

90
El-Nemr, A., Al-Shawaf, T., Sabatini, L., Wilson, C., Lower, A. M., & Grudzinskas, J. G. (1998).
Effect of smoking on ovarian reserve and ovarian stimulation in in-vitro fertilization and
embryo transfer. Hum Reprod, 13(8), 2192-2198.
Elovaara, E., Mikkola, J., Mäkelä, M., Paldanius, B., & Priha, E. (2006). Assessment of soil
remediation workers' exposure to polycyclic aromatic hydrocarbons (PAH):
biomonitoring of naphthols, phenanthrols, and 1-hydroxypyrene in urine. Toxicol Lett,
162(2-3), 158-163. doi:10.1016/j.toxlet.2005.09.028
Everson, R. B., Sandler, D. P., Wilcox, A. J., Schreinemachers, D., Shore, D. L., & Weinberg, C.
(1986). Effect of passive exposure to smoking on age at natural menopause. Br Med J
(Clin Res Ed), 293(6550), 792.
Fang, G. C., Wu, Y. S., Chen, J. C., Chang, C. N., & Ho, T. T. (2006). Characteristic of polycyclic
aromatic hydrocarbon concentrations and source identification for fine and coarse
particulates at Taichung Harbor near Taiwan Strait during 2004-2005. Sci Total Environ,
366(2-3), 729-738. doi:10.1016/j.scitotenv.2005.09.075
Farin, F. M., Janssen, P., Quigley, S., Abbott, D., Hassett, C., Smith-Weller, T., . . . Checkoway, H.
(2001). Genetic polymorphisms of microsomal and soluble epoxide hydrolase and the
risk of Parkinson's disease. Pharmacogenetics, 11(8), 703-708.
Ferreira Júnior, M. F., Tas, S., dell'Omo, M., Goormans, G., Buchet, J. P., & Lauwerys, R. (1994).
Determinants of benzo(a)pyrenediol epoxide adducts to haemoglobin in workers
exposed to polycyclic aromatic hydrocarbons. Occup Environ Med, 51(7), 451-455.
Findlay, J. K., Drummond, A. E., Britt, K. L., Dyson, M., Wreford, N. G., Robertson, D. M., . . .
Simpson, E. R. (2000). The roles of activins, inhibins and estrogen in early committed
follicles. Mol Cell Endocrinol, 163(1-2), 81-87.
Foth, H., Kahl, R., & Kahl, G. F. (1988a). Pharmacokinetics of low doses of benzo[a]pyrene in
the rat. Food Chem Toxicol, 26(1), 45-51.
Foth, H., Kahl, R., & Kahl, G. F. (1988b). Pharmacokinetics of low doses of benzo[a]pyrene in
the rat. Food Chem Toxicol, 26(1), 45-51.
Fourth National Report of Human Exposure to Environmental Chemicals. (2015). Polycyclic
Aromatic Hydrocarbons (pp. 461-497). Center for Disease Control and Prevention
National Center for Environmental Health
Division of Laboratory Sciences: CDC.
Freour, T., Masson, D., Mirallie, S., Jean, M., Bach, K., Dejoie, T., & Barriere, P. (2008). Active
smoking compromises IVF outcome and affects ovarian reserve. Reprod Biomed Online,
16(1), 96-102.

91
Fuentes, A., Muñoz, A., Pommer, R., Argüello, B., Galleguillos, A., Torres, A., & Jesam, C. (2012).
Decreased anti-Müllerian hormone concentration in follicular fluid of female smokers
undergoing artificial reproductive techniques. Chemosphere, 88(4), 403-406.
doi:10.1016/j.chemosphere.2012.02.054
Funk, M., Gath, I., Seidel, A., Oesch, F., & Platt, K. L. (1995). Conjugation of anti-dihydrodiol
epoxides of benzo[a]pyrene, chrysene, benzo[c]phenanthrene and
dibenz[a,h]anthracene with glutathione catalyzed by cytosol and by the Mu-class
glutathione transferase HTP II from rat liver. Chem Biol Interact, 95(1-2), 189-201.
Furue, M., Takahara, M., Nakahara, T., & Uchi, H. (2014). Role of AhR/ARNT system in skin
homeostasis. Arch Dermatol Res, 306(9), 769-779. doi:10.1007/s00403-014-1481-7
Förster, K., Preuss, R., Rossbach, B., Brüning, T., Angerer, J., & Simon, P. (2008). 3-
Hydroxybenzo[a]pyrene in the urine of workers with occupational exposure to
polycyclic aromatic hydrocarbons in different industries. Occup Environ Med, 65(4),
224-229. doi:10.1136/oem.2006.030809
Gao, Q., Xu, Z., Li, S., Jin, T., Chen, B., Lao, Z., . . . Zhi, Z. (2011). Modification of GSTM1, GSTT1
and GSTP1 gene polymorphisms on urinary 1-hydroxypyrene excretions. Chines Journal
of Industrial Hygiene and Occupational Diseases
Research Support, US Gov't, Non-P.H.S., English Abstract (lang;chi), 29(1), 11-15.
Garde, A. H., Hansen, A. M., Kristiansen, J., & Knudsen, L. E. (2004). Comparison of
uncertainties related to standardization of urine samples with volume and creatinine
concentration. Ann Occup Hyg, 48(2), 171-179.
Garte, S., Gaspari, L., Alexandrie, A. K., Ambrosone, C., Autrup, H., Autrup, J. L., . . . Taioli, E.
(2001). Metabolic gene polymorphism frequencies in control populations. Cancer
Epidemiol Biomarkers Prev, 10(12), 1239-1248.
Gattás, G. J., & Soares-Vieira, J. A. (2000). Cytochrome P450-2E1 and glutathione S-transferase
mu polymorphisms among Caucasians and mulattoes from Brazil. Occup Med (Lond),
50(7), 508-511.
Gelboin, H. V. (1980). Benzo[alpha]pyrene metabolism, activation and carcinogenesis: role and
regulation of mixed-function oxidases and related enzymes. Physiol Rev, 60(4), 1107-
1166.
Gerde, P., Muggenburg, B. A., Lundborg, M., & Dahl, A. R. (2001a). The rapid alveolar
absorption of diesel soot-adsorbed benzo[a]pyrene: bioavailability, metabolism and
dosimetry of an inhaled particle-borne carcinogen. Carcinogenesis, 22(5), 741-749.

92
Gerde, P., Muggenburg, B. A., Lundborg, M., & Dahl, A. R. (2001b). The rapid alveolar
absorption of diesel soot-adsorbed benzo[a]pyrene: bioavailability, metabolism and
dosimetry of an inhaled particle-borne carcinogen. Carcinogenesis, 22(5), 741-749.
Gilliland, F. D., Harms, H. J., Crowell, R. E., Li, Y. F., Willink, R., & Belinsky, S. A. (2002).
Glutathione S-transferase P1 and NADPH quinone oxidoreductase polymorphisms are
associated with aberrant promoter methylation of P16(INK4a) and O(6)-methylguanine-
DNA methyltransferase in sputum. Cancer Res, 62(8), 2248-2252.
Glatt, H. R., & Oesch, F. (1987). Species differences in enzymes controlling reactive epoxides.
Arch Toxicol Suppl, 10, 111-124.
Gold, E. B., Bromberger, J., Crawford, S., Samuels, S., Greendale, G. A., Harlow, S. D., &
Skurnick, J. (2001). Factors associated with age at natural menopause in a multiethnic
sample of midlife women. Am J Epidemiol, 153(9), 865-874.
Gracia, C. R., Sammel, M. D., Freeman, E., Prewitt, M., Carlson, C., Ray, A., . . . Ginsberg, J. P.
(2012). Impact of cancer therapies on ovarian reserve. Fertil Steril, 97(1), 134-140.e131.
doi:10.1016/j.fertnstert.2011.10.040
Greenseid, K., Jindal, S., Zapantis, A., Nihsen, M., Hurwitz, J., & Pal, L. (2009). Declining ovarian
reserve adversely influences granulosa cell viability. Fertil Steril, 91(6), 2611-2615.
doi:10.1016/j.fertnstert.2008.03.065
Grimmer, G. (1968). Carcinogenic hydrocarbons in the human environment (Vol. 108, pp. 529).
Germany: Biochemical Institute for Environmental Carcinogens.
Grimmer, G., & Hildebrandt, A. (1968). [Hydrocarbons in the environment of man. VI. The
contents of polycyclic hydrocarbons in raw vegetable oils]. Arch Hyg Bakteriol, 152(3),
255-259.
Grimmer, G., Jacob, J., Dettbarn, G., & Naujack, K. W. (1997). Determination of urinary
metabolites of polycyclic aromatic hydrocarbons (PAH) for the risk assessment of PAH-
exposed workers. Int Arch Occup Environ Health, 69(4), 231-239.
Grova, N., & Feidt, C. (2002). [14C] Milk, urine, and faeces excretion kinetics in lactating goats
after oral administration of [14C] polycyclic aromatic hydrocarbons. International Dairy
Journal, 12(Laboratoire de Sciences Animales), 1025-1031.
Gruijters, M. J., Visser, J. A., Durlinger, A. L., & Themmen, A. P. (2003). Anti-Müllerian hormone
and its role in ovarian function. Mol Cell Endocrinol, 211(1-2), 85-90.
Guo, W., He, M., Yang, Z., Lin, C., Quan, X., & Wang, H. (2007). Distribution of polycyclic
aromatic hydrocarbons in water, suspended particulate matter and sediment from

93
Daliao River watershed, China. Chemosphere, 68(1), 93-104.
doi:10.1016/j.chemosphere.2006.12.072
Hall, M., Forrester, L. M., Parker, D. K., Grover, P. L., & Wolf, C. R. (1989). Relative contribution
of various forms of cytochrome P450 to the metabolism of benzo[a]pyrene by human
liver microsomes. Carcinogenesis, 10(10), 1815-1821.
Han, I. K., Duan, X., Zhang, L., Yang, H., Rhoads, G. G., Wei, F., & Zhang, J. (2008). 1-
Hydroxypyrene concentrations in first morning voids and 24-h composite urine: intra-
and inter-individual comparisons. J Expo Sci Environ Epidemiol, 18(5), 477-485.
doi:10.1038/sj.jes.7500639
Hansen, K. R., Knowlton, N. S., Thyer, A. C., Charleston, J. S., Soules, M. R., & Klein, N. A. (2008).
A new model of reproductive aging: the decline in ovarian non-growing follicle number
from birth to menopause. Hum Reprod, 23(3), 699-708. doi:10.1093/humrep/dem408
Harrad, S., de Wit, C. A., Abdallah, M. A., Bergh, C., Björklund, J. A., Covaci, A., . . . Webster, T.
F. (2010). Indoor contamination with hexabromocyclododecanes, polybrominated
diphenyl ethers, and perfluoroalkyl compounds: an important exposure pathway for
people? Environ Sci Technol, 44(9), 3221-3231. doi:10.1021/es903476t
Hatjian, B. A., Edwards, J. W., Harrison, J., Williams, F. M., & Blain, P. G. (1995). Ambient,
biological, and biological effect monitoring of exposure to polycyclic aromatic
hydrocarbons (PAHs). Toxicol Lett, 77(1-3), 271-279.
Heikkilä, P., Luotamo, M., Pyy, L., & Riihimäki, V. (1995). Urinary 1-naphthol and 1-pyrenol as
indicators of exposure to coal tar products. Int Arch Occup Environ Health, 67(3), 211-
217.
Hillier, S. G. (1991). Regulatory functions for inhibin and activin in human ovaries. J Endocrinol,
131(2), 171-175.
Hoffmann, D., Hoffmann, I., & El-Bayoumy, K. (2001). The less harmful cigarette: a
controversial issue. a tribute to Ernst L. Wynder. Chem Res Toxicol, 14(7), 767-790.
Homburg, R. (2014). The Mechanism of Ovulation. Global Liberal Women's Medicine, 10(3843),
1756-2226.
Homburg, R., & Crawford, G. (2014). The role of AMH in anovulation associated with PCOS: a
hypothesis. Hum Reprod, 29(6), 1117-1121. doi:10.1093/humrep/deu076
Hong, Y. C., Leem, J. H., Park, H. S., Lee, K. H., Lee, S. J., Lee, C. K., & Kang, D. (1999). Variations
in urinary 1-hydroxypyrene glucuronide in relation to smoking and the modification
effects of GSTM1 and GSTT1. Toxicol Lett, 108(2-3), 217-223.

94
Hu, Y., Li, G., Xue, X., Zhou, Z., Li, X., Fu, J., . . . Qu, Q. (2008). PAH-DNA adducts in a Chinese
population: relationship to PAH exposure, smoking and polymorphisms of metabolic
and DNA repair genes. Biomarkers, 13(1), 27-40. doi:10.1080/13547500701671895
IARC, The International Agency for Research on Cancer. (1973).
Igawa, Y., Keating, A. F., Rajapaksa, K. S., Sipes, I. G., & Hoyer, P. B. (2009). Evaluation of
ovotoxicity induced by 7, 12-dimethylbenz[a]anthracene and its 3,4-diol metabolite
utilizing a rat in vitro ovarian culture system. Toxicol Appl Pharmacol, 234(3), 361-369.
doi:10.1016/j.taap.2008.10.009
International Programme on Chemical Safety (IPCS). (1998). Selected non-heterocyclic
polycyclic aromatic hydrocarbons. 202.
Iorio, R., Castellucci, A., Ventriglia, G., Teoli, F., Cellini, V., Macchiarelli, G., & Cecconi, S. (2014).
Ovarian toxicity: from environmental exposure to chemotherapy. Curr Pharm Des,
20(34), 5388-5397.
Jacob, J., & Seidel, A. (2002). Biomonitoring of polycyclic aromatic hydrocarbons in human
urine. J Chromatogr B Analyt Technol Biomed Life Sci, 778(1-2), 31-47.
Jaffe, R. B. (2009). [Commentary on] Anti-Müllerian hormone and inhibin B in the definition of
ovarian aging and the menopause transition. Obstetrical & Gynecological Survey, 64(2),
111-112.
Jernstrom, B., Funk, M., Frank, H., Mannervik, B., & Seidel, A. (1996). Glutathione S-transferase
A1-1-catalysed conjugation of bay and fjord region diol epoxides or polycyclic aromatic
hydrocarbons with glutathione. Carcinogenesis, 17(7), 1491-1498.
Jernström, B., Seidel, A., Funk, M., Oesch, F., & Mannervik, B. (1992). Glutathione conjugation
of trans-3,4-dihydroxy 1,2-epoxy 1,2,3,4-tetrahydrobenzo[c]phenanthrene isomers by
human glutathione transferases. Carcinogenesis, 13(9), 1549-1555.
Jick, H., & Porter, J. (1977). Relation between smoking and age of natural menopause. Report
from the Boston Collaborative Drug Surveillance Program, Boston University Medical
Center. Lancet, 1(8026), 1354-1355.
Jindal, S., Greenseid, K., Berger, D., Santoro, N., & Pal, L. (2012). Impaired gremlin 1 (GREM1)
expression in cumulus cells in young women with diminished ovarian reserve (DOR). J
Assist Reprod Genet, 29(2), 159-162. doi:10.1007/s10815-011-9684-8
Jinot, J., & Bayard, S. (1996). Respiratory health effects of exposure to environmental tobacco
smoke. Rev Environ Health, 11(3), 89-100.

95
Johnson, J., Canning, J., Kaneko, T., Pru, J. K., & Tilly, J. L. (2004). Germline stem cells and
follicular renewal in the postnatal mammalian ovary. Nature, 428(6979), 145-150.
doi:10.1038/nature02316
Jongeneelen, F. J. (1992). Biological exposure limit for occupational exposure to coal tar pitch
volatiles at cokeovens. Int Arch Occup Environ Health, 63(8), 511-516.
Jongeneelen, F. J. (1994). Biological monitoring of environmental exposure to polycyclic
aromatic hydrocarbons; 1-hydroxypyrene in urine of people. Toxicol Lett, 72(1-3), 205-
211.
Jongeneelen, F. J. (2014). A guidance value of 1-hydroxypyrene in urine in view of acceptable
occupational exposure to polycyclic aromatic hydrocarbons. Toxicol Lett, 231(2), 239-
248. doi:10.1016/j.toxlet.2014.05.001
Jongeneelen, F. J., Bos, R. P., Anzion, R. B., Theuws, J. L., & Henderson, P. T. (1986). Biological
monitoring of polycyclic aromatic hydrocarbons. Metabolites in urine. Scand J Work
Environ Health, 12(2), 137-143.
Juengel, J. L., & McNatty, K. P. (2005). The role of proteins of the transforming growth factor-
beta superfamily in the intraovarian regulation of follicular development. Hum Reprod
Update, 11(2), 143-160. doi:10.1093/humupd/dmh061
Jung, K. H., Liu, B., Lovinsky-Desir, S., Yan, B., Camann, D., Sjodin, A., . . . Miller, R. L. (2014).
Time trends of polycyclic aromatic hydrocarbon exposure in New York City from 2001
to 2012: assessed by repeat air and urine samples. Environ Res, 131, 95-103.
doi:10.1016/j.envres.2014.02.017
Jägerstad, M., & Skog, K. (2005). Genotoxicity of heat-processed foods. Mutat Res, 574(1-2),
156-172. doi:10.1016/j.mrfmmm.2005.01.030
Kaiser, J. (1997). Showdown over clean air science. Science, 277(5325), 466-469.
Kao, J., Patterson, F. K., & Hall, J. (1985). Skin penetration and metabolism of topically applied
chemicals in six mammalian species, including man: an in vitro study with
benzo[a]pyrene and testosterone. Toxicol Appl Pharmacol, 81(3 Pt 1), 502-516.
Karande, V., & Gleicher, N. (1999). A rational approach to the management of low responders
in in-vitro fertilization. Hum Reprod, 14(7), 1744-1748.
Kawamoto, T., Kunugita, N., Katoh, T., & Yang, M. (1998). Effects of life-style and genetic
polymorphisms of CYP1A1 and GSTM1 on urinary 1-hydroxypyrene concentration.
Paper presented at the Proc. intern. Symp. Biol. Monitoring, Seoul.

96
Kawamoto, T., Yang, M., Kim, Y. D., Kim, H., Oyama, T., Isse, T., . . . Uchiyama, I. (2007). Effects
of lifestyle on urinary 1-hydroxypyrene concentration. J Occup Health, 49(3), 183-189.
Kawanami, S., Nguyen, T. T., Inoue, J., Kawai, K., & Horie, S. (2014). Urinary 1-hydroxypyrene
levels among office workers living in industrial areas. J UOEH, 36(1), 1-10.
Kelly, G., Bartle, K., & Clifford, A. (1993). Identification and Quantification of Polycyclic
Aromatic Hydrocarbons in Air Particulate and Diesel Exhaust Particulate Extracts by LC-
GC. Journal of Chromotographic Science, 31(3), 73-76.
Kelsey, K. T., Wiencke, J. K., & Spitz, M. R. (1994). A race-specific genetic polymorphism in the
CYP1A1 gene is not associated with lung cancer in African Americans. Carcinogenesis,
15(6), 1121-1124.
Kennish, M. J. (1996). Polycyclic Aromatic Hydrocarbons. In P. Petralia (Ed.), Practical
Handbook of Estuarine and Marine Pollution (CRC Press Marine Science Series ed., pp.
142-145). Institute of Marine and Coastal Science, New Brunswick, New Jersey: Library
of Congress Catologing.
Kevenaar, M. E., Laven, J. S., Fong, S. L., Uitterlinden, A. G., de Jong, F. H., Themmen, A. P., &
Visser, J. A. (2008). A functional anti-mullerian hormone gene polymorphism is
associated with follicle number and androgen levels in polycystic ovary syndrome
patients. J Clin Endocrinol Metab, 93(4), 1310-1316. doi:10.1210/jc.2007-2205
Kevenaar, M. E., Meerasahib, M. F., Kramer, P., van de Lang-Born, B. M., de Jong, F. H.,
Groome, N. P., . . . Visser, J. A. (2006a). Serum anti-mullerian hormone levels reflect the
size of the primordial follicle pool in mice. Endocrinology, 147(7), 3228-3234.
doi:10.1210/en.2005-1588
Kevenaar, M. E., Meerasahib, M. F., Kramer, P., van de Lang-Born, B. M., de Jong, F. H.,
Groome, N. P., . . . Visser, J. A. (2006b). Serum anti-mullerian hormone levels reflect the
size of the primordial follicle pool in mice. Endocrinology, 147(7), 3228-3234.
doi:10.1210/en.2005-1588
Kevenaar, M. E., Themmen, A. P., Laven, J. S., Sonntag, B., Fong, S. L., Uitterlinden, A. G., . . .
Visser, J. A. (2007). Anti-Müllerian hormone and anti-Müllerian hormone type II
receptor polymorphisms are associated with follicular phase estradiol levels in normo-
ovulatory women. Hum Reprod, 22(6), 1547-1554. doi:10.1093/humrep/dem036
Kilanowicz, A., Czerski, B., & Sapota, A. (1999). The disposition and metabolism of naphthalene
in rats. Int J Occup Med Environ Health, 12(3), 209-219.
Kim, S. R., Halden, R. U., & Buckley, T. J. (2008). Polycyclic aromatic hydrocarbons in human
milk of nonsmoking U.S. women. Environ Sci Technol, 42(7), 2663-2667.

97
Kinney, A., Kline, J., Kelly, A., Reuss, M. L., & Levin, B. (2007). Smoking, alcohol and caffeine in
relation to ovarian age during the reproductive years. Hum Reprod, 22(4), 1175-1185.
doi:10.1093/humrep/del496
Klaassen, C., & Watkins III, J. (2008). Casarett & Doull's Essentials of Toxicology (3rd ed.):
McGraw Hill Company Inc.
Kleiner, H. E., Vulimiri, S. V., Hatten, W. B., Reed, M. J., Nebert, D. W., Jefcoate, C. R., &
DiGiovanni, J. (2004). Role of cytochrome p4501 family members in the metabolic
activation of polycyclic aromatic hydrocarbons in mouse epidermis. Chem Res Toxicol,
17(12), 1667-1674. doi:10.1021/tx049919c
Kummer, N., Martin, J. R., & Pal, L. (2009). Diminished ovarian reserve in a woman with a
balanced 13;21 translocation. Fertil Steril, 91(3), 931.e933-935.
doi:10.1016/j.fertnstert.2008.07.1726
Laboratory decontamination and destruction of carcinogens in laboratory wastes: some N-
nitrosamides. International Agency for Research on Cancer. (1983). IARC Sci Publ(55), 1-
65.
Ladou, J., & Harrison, R. (2014). Current Occupational and Environmental Medicine (5th ed.).
University of California, San Francisco: McGraw Hill Education Companies, Appleton &
Lange.
Larsen, J. C. (1995). Levels of pollutants and their metabolites: exposures to organic
substances. Toxicology, 101(1-2), 11-27.
Laven, J. S., Mulders, A. G., Visser, J. A., Themmen, A. P., De Jong, F. H., & Fauser, B. C. (2004).
Anti-Müllerian hormone serum concentrations in normoovulatory and anovulatory
women of reproductive age. J Clin Endocrinol Metab, 89(1), 318-323.
doi:10.1210/jc.2003-030932
Lawther, P. J., & Waller, R. E. (1976). Coal fires, industrial emissions and motor vehicles as
sources of environmental carcinogens. IARC Sci Publ(13), 27-40.
Lee, C. Y., Lee, J. Y., Kang, J. W., & Kim, H. (2001). Effects of genetic polymorphisms of CYP1A1,
CYP2E1, GSTM1, and GSTT1 on the urinary levels of 1-hydroxypyrene and 2-naphthol in
aircraft maintenance workers. Toxicol Lett, 123(2-3), 115-124.
Levy, B., Wegman, D., Baron, S., & Sokas, R. (2011). Occupational and Environmental Health
(6th ed.). 198 Madison Avenue, New York, New York 10016: Oxford University Press,
Inc.

98
Li, Z., Romanoff, L., Bartell, S., Pittman, E. N., Trinidad, D. A., McClean, M., . . . Sjödin, A. (2012).
Excretion profiles and half-lives of ten urinary polycyclic aromatic hydrocarbon
metabolites after dietary exposure. Chem Res Toxicol, 25(7), 1452-1461.
doi:10.1021/tx300108e
Li, Z., Romanoff, L. C., Lewin, M. D., Porter, E. N., Trinidad, D. A., Needham, L. L., . . . Sjödin, A.
(2010). Variability of urinary concentrations of polycyclic aromatic hydrocarbon
metabolite in general population and comparison of spot, first-morning, and 24-h void
sampling. J Expo Sci Environ Epidemiol, 20(6), 526-535. doi:10.1038/jes.2009.41
Li, Z., Romanoff, L. C., Trinidad, D. A., Hussain, N., Jones, R. S., Porter, E. N., . . . Sjödin, A.
(2006). Measurement of urinary monohydroxy polycyclic aromatic hydrocarbons using
automated liquid-liquid extraction and gas chromatography/isotope dilution high-
resolution mass spectrometry. Anal Chem, 78(16), 5744-5751. doi:10.1021/ac0606094
Li, Z., Sandau, C. D., Romanoff, L. C., Caudill, S. P., Sjodin, A., Needham, L. L., & Patterson, D. G.
(2008). Concentration and profile of 22 urinary polycyclic aromatic hydrocarbon
metabolites in the US population. Environ Res, 107(3), 320-331.
doi:10.1016/j.envres.2008.01.013
Lie Fong, S., Baart, E. B., Martini, E., Schipper, I., Visser, J. A., Themmen, A. P., . . . Laven, J. S.
(2008). Anti-Müllerian hormone: a marker for oocyte quantity, oocyte quality and
embryo quality? Reprod Biomed Online, 16(5), 664-670.
Lie Fong, S., Schipper, I., de Jong, F. H., Themmen, A. P., Visser, J. A., & Laven, J. S. (2011).
Serum anti-Müllerian hormone and inhibin B concentrations are not useful predictors
of ovarian response during ovulation induction treatment with recombinant follicle-
stimulating hormone in women with polycystic ovary syndrome. Fertil Steril, 96(2), 459-
463. doi:10.1016/j.fertnstert.2011.05.084
Lie Fong, S., Schipper, I., Valkenburg, O., de Jong, F. H., Visser, J. A., & Laven, J. S. (2015). The
role of anti-Müllerian hormone in the classification of anovulatory infertility. Eur J
Obstet Gynecol Reprod Biol, 186, 75-79. doi:10.1016/j.ejogrb.2015.01.007
Lie Fong, S., Visser, J. A., Welt, C. K., de Rijke, Y. B., Eijkemans, M. J., Broekmans, F. J., . . . Laven,
J. S. (2012). Serum anti-müllerian hormone levels in healthy females: a nomogram
ranging from infancy to adulthood. J Clin Endocrinol Metab, 97(12), 4650-4655.
doi:10.1210/jc.2012-1440
LIJINSKY, W., & SHUBIK, P. (1964). BENZO(A)PYRENE AND OTHER POLYNUCLEAR
HYDROCARBONS IN CHARCOAL-BROILED MEAT. Science, 145(3627), 53-55.
Lim, J., Lawson, G. W., Nakamura, B. N., Ortiz, L., Hur, J. A., Kavanagh, T. J., & Luderer, U.
(2013). Glutathione-deficient mice have increased sensitivity to transplacental

99
benzo[a]pyrene-induced premature ovarian failure and ovarian tumorigenesis. Cancer
Res, 73(2), 908-917. doi:10.1158/0008-5472.CAN-12-3636
Lin, P. Y., Huang, F. J., Kung, F. T., Chiang, H. J., Lin, Y. J., Lin, Y. C., & Lan, K. C. (2014).
Evaluation of serum anti-Mullerian hormone as a biomarker of early ovarian aging in
young women undergoing IVF/ICSI cycle. Int J Clin Exp Pathol, 7(9), 6245-6253.
Lioy, P. L., Waldman, J. M., Greenberg, A., Harkov, R., & Pietarinen, C. (1988a). The Total
Human Environmental Exposure Study (THEES) to benzo(a)pyrene: comparison of the
inhalation and food pathways. Arch Environ Health, 43(4), 304-312.
Lioy, P. L., Waldman, J. M., Greenberg, A., Harkov, R., & Pietarinen, C. (1988b). The Total
Human Environmental Exposure Study (THEES) to benzo(a)pyrene: comparison of the
inhalation and food pathways. Arch Environ Health, 43(4), 304-312.
Liu, B., & Jia, C. (2015). Effects of profession on urinary PAH metabolite levels in the US
population. Int Arch Occup Environ Health. doi:10.1007/s00420-015-1057-7
Liu, M., Chen, L., Zhou, R., & Wang, J. (2013). Association between GSTM1 polymorphism and
DNA adduct concentration in the occupational workers exposed to PAHs: a meta-
analysis. Gene, 519(1), 71-76. doi:10.1016/j.gene.2013.01.045
Lukaszuk, K., Liss, J., Kunicki, M., Jakiel, G., Wasniewski, T., Woclawek-Potocka, I., & Pastuszek,
E. (2014). Anti-Mullerian hormone (AMH) is a strong predictor of live birth in women
undergoing assisted reproductive technology. Reprod Biol, 14(3), 176-181.
doi:10.1016/j.repbio.2014.03.004
Lyon, I. A. f. R. o. C. (1983). Polycyclic Aromatic Hydrocarbons, Part 1 . Chemical, environmental
and experimental data.
Lyu, Y., Zheng, W., Zheng, T., & Tian, Y. (2014). Biodegradation of polycyclic aromatic
hydrocarbons by Novosphingobium pentaromativorans US6-1. PLoS One, 9(7),
e101438. doi:10.1371/journal.pone.0101438
Ma, W. L., Li, Y. F., Qi, H., Sun, D. Z., Liu, L. Y., & Wang, D. G. (2010). Seasonal variations of
sources of polycyclic aromatic hydrocarbons (PAHs) to a northeastern urban city, China.
Chemosphere, 79(4), 441-447. doi:10.1016/j.chemosphere.2010.01.048
Ma, Y., & Harrad, S. (2015). Spatiotemporal analysis and human exposure assessment on
polycyclic aromatic hydrocarbons in indoor air, settled house dust, and diet: A review.
Environ Int, 84, 7-16. doi:10.1016/j.envint.2015.07.006
MacKenzie, K. M., & Angevine, D. M. (1981). Infertility in mice exposed in utero to
benzo(a)pyrene. Biol Reprod, 24(1), 183-191.

100
Mark-Kappeler, C. J., Hoyer, P. B., & Devine, P. J. (2011). Xenobiotic effects on ovarian
preantral follicles. Biol Reprod, 85(5), 871-883. doi:10.1095/biolreprod.111.091173
Matikainen, T., Perez, G. I., Jurisicova, A., Pru, J. K., Schlezinger, J. J., Ryu, H. Y., . . . Tilly, J. L.
(2001). Aromatic hydrocarbon receptor-driven Bax gene expression is required for
premature ovarian failure caused by biohazardous environmental chemicals. Nat
Genet, 28(4), 355-360. doi:10.1038/ng575
Matikainen, T. M., Moriyama, T., Morita, Y., Perez, G. I., Korsmeyer, S. J., Sherr, D. H., & Tilly, J.
L. (2002). Ligand activation of the aromatic hydrocarbon receptor transcription factor
drives Bax-dependent apoptosis in developing fetal ovarian germ cells. Endocrinology,
143(2), 615-620. doi:10.1210/endo.143.2.8624
Matthews, J., & Gustafsson, J. A. (2006). Estrogen receptor and aryl hydrocarbon receptor
signaling pathways. Nucl Recept Signal, 4, e016. doi:10.1621/nrs.04016
Mattison, D. R. (1980). Morphology of oocyte and follicle destruction by polycyclic aromatic
hydrocarbons in mice. Toxicol Appl Pharmacol, 53(2), 249-259.
Mattison, D. R., Singh, H., Takizawa, K., & Thomford, P. J. (1989). Ovarian toxicity of
benzo(a)pyrene and metabolites in mice. Reprod Toxicol, 3(2), 115-125.
McCarroll, S. A., Hadnott, T. N., Perry, G. H., Sabeti, P. C., Zody, M. C., Barrett, J. C., . . .
Consortium, I. H. (2006). Common deletion polymorphisms in the human genome. Nat
Genet, 38(1), 86-92.
McCarty, K. M., Santella, R. M., Steck, S. E., Cleveland, R. J., Ahn, J., Ambrosone, C. B., . . .
Gammon, M. D. (2009). PAH-DNA adducts, cigarette smoking, GST polymorphisms, and
breast cancer risk. Environ Health Perspect, 117(4), 552-558. doi:10.1289/ehp.0800119
McClean, M. D., Rinehart, R. D., Ngo, L., Eisen, E. A., Kelsey, K. T., & Herrick, R. F. (2004).
Inhalation and dermal exposure among asphalt paving workers. Ann Occup Hyg, 48(8),
663-671. doi:10.1093/annhyg/meh062
McClean, M. D., Rinehart, R. D., Ngo, L., Eisen, E. A., Kelsey, K. T., Wiencke, J. K., & Herrick, R. F.
(2004a). Urinary 1-hydroxypyrene and polycyclic aromatic hydrocarbon exposure
among asphalt paving workers. Ann Occup Hyg, 48(6), 565-578.
doi:10.1093/annhyg/meh044
McClean, M. D., Rinehart, R. D., Ngo, L., Eisen, E. A., Kelsey, K. T., Wiencke, J. K., & Herrick, R. F.
(2004b). Urinary 1-hydroxypyrene and polycyclic aromatic hydrocarbon exposure
among asphalt paving workers. Ann Occup Hyg, 48(6), 565-578.
doi:10.1093/annhyg/meh044

101
McDougal, J. N., & Robinson, P. J. (2002). Assessment of dermal absorption and penetration of
components of a fuel mixture (JP-8). Sci Total Environ, 288(1-2), 23-30.
McLachlan, R. I., Robertson, D. M., Healy, D. L., Burger, H. G., & de Kretser, D. M. (1987).
Circulating immunoreactive inhibin levels during the normal human menstrual cycle. J
Clin Endocrinol Metab, 65(5), 954-961. doi:10.1210/jcem-65-5-954
McNatty, K. P., Smith, P., Moore, L. G., Reader, K., Lun, S., Hanrahan, J. P., . . . Juengel, J. L.
(2005). Oocyte-expressed genes affecting ovulation rate. Mol Cell Endocrinol, 234(1-2),
57-66. doi:10.1016/j.mce.2004.08.013
Menzie, C., Potocki, B., & Santodonato, J. (1992). Exposure to carcinogenic PAHs in the
environment. Environ. Sci. Technol, 26(7), 1278-1284.
Merlo, F., Andreassen, A., Weston, A., Pan, C. F., Haugen, A., Valerio, F., . . . Abbondandolo, A.
(1998). Urinary excretion of 1-hydroxypyrene as a marker for exposure to urban air
levels of polycyclic aromatic hydrocarbons. Cancer Epidemiol Biomarkers Prev, 7(2),
147-155.
Miyata, M., Kudo, G., Lee, Y. H., Yang, T. J., Gelboin, H. V., Fernandez-Salguero, P., . . .
Gonzalez, F. J. (1999). Targeted disruption of the microsomal epoxide hydrolase gene.
Microsomal epoxide hydrolase is required for the carcinogenic activity of 7,12-
dimethylbenz[a]anthracene. J Biol Chem, 274(34), 23963-23968.
Morin, P., Payette, H., Moos, M. K., St-Cyr-Tribble, D., Niyonsenga, T., & De Wals, P. (2003).
Measuring the intensity of pregnancy planning effort. Paediatr Perinat Epidemiol, 17(1),
97-105.
Motorykin, O., Santiago-Delgado, L., Rohlman, D., Schrlau, J. E., Harper, B., Harris, S., . . .
Massey Simonich, S. L. (2015a). Metabolism and excretion rates of parent and hydroxy-
PAHs in urine collected after consumption of traditionally smoked salmon for Native
American volunteers. Sci Total Environ, 514, 170-177.
doi:10.1016/j.scitotenv.2015.01.083
Motorykin, O., Santiago-Delgado, L., Rohlman, D., Schrlau, J. E., Harper, B., Harris, S., . . .
Massey Simonich, S. L. (2015b). Metabolism and excretion rates of parent and hydroxy-
PAHs in urine collected after consumption of traditionally smoked salmon for Native
American volunteers. Sci Total Environ, 514, 170-177.
doi:10.1016/j.scitotenv.2015.01.083
Mukherjee, S., Rodrigues, E., Weker, R., Palmer, L. J., & Christiani, D. C. (2002). 1-
hydroxypyrene as a biomarker of occupational exposure to polycyclic aromatic
hydrocarbons (PAH) in boilermakers. J Occup Environ Med, 44(12), 1119-1125.

102
Nakachi, K., Imai, K., Hayashi, S., & Kawajiri, K. (1993). Polymorphisms of the CYP1A1 and
glutathione S-transferase genes associated with susceptibility to lung cancer in relation
to cigarette dose in a Japanese population. Cancer Res, 53(13), 2994-2999.
Nan, H. M., Kim, H., Lim, H. S., Choi, J. K., Kawamoto, T., Kang, J. W., . . . Kwon, E. H. (2001).
Effects of occupation, lifestyle and genetic polymorphisms of CYP1A1, CYP2E1, GSTM1
and GSTT1 on urinary 1-hydroxypyrene and 2-naphthol concentrations. Carcinogenesis,
22(5), 787-793.
Nationwide Environmental Title Research (NETR), LCC. (1983, 1990).
Naumova, Y. Y., Eisenreich, S. J., Turpin, B. J., Weisel, C. P., Morandi, M. T., Colome, S. D., . . .
Wall, S. J. (2002). Polycyclic aromatic hydrocarbons in the indoor and outdoor air of
three cities in the U.S. Environ Sci Technol, 36(12), 2552-2559.
NCBI, National Center for Biotechnical Information, Database, T. N. C. f. B. I. National Institute
of Health.
NCS, N. C. S. http://NCSArchive.s-3.net.
Neal, M. S., Zhu, J., & Foster, W. G. (2008). Quantification of benzo[a]pyrene and other PAHs in
the serum and follicular fluid of smokers versus non-smokers. Reprod Toxicol, 25(1),
100-106. doi:10.1016/j.reprotox.2007.10.012
Neal, M. S., Zhu, J., Holloway, A. C., & Foster, W. G. (2007). Follicle growth is inhibited by
benzo-[a]-pyrene, at concentrations representative of human exposure, in an isolated
rat follicle culture assay. Hum Reprod, 22(4), 961-967. doi:10.1093/humrep/del487
Nebert, D. W., Ingelman-Sundberg, M., & Daly, A. K. (1999). Genetic epidemiology of
environmental toxicity and cancer susceptibility: human allelic polymorphisms in drug-
metabolizing enzyme genes, their functional importance, and nomenclature issues.
Drug Metab Rev, 31(2), 467-487. doi:10.1081/DMR-100101931
Nebert, D. W., Puga, A., & Vasiliou, V. (1993). Role of the Ah receptor and the dioxin-inducible
[Ah] gene battery in toxicity, cancer, and signal transduction. Ann N Y Acad Sci, 685,
624-640.
Neish, W. J. (1948). Metabolism of fluorene in the rabbit. Biochem J, 43(4), 533-537.
Nejat, E. J., Jindal, S., Berger, D., Buyuk, E., Lalioti, M., & Pal, L. (2011). Implications of blood
type for ovarian reserve. Hum Reprod, 26(9), 2513-2517. doi:10.1093/humrep/der199
NETR. (September 8, 1983). National Security, Soil Preparation Services.

103
Neubert, D., & Tapken, S. (1988). Transfer of benzo(a)pyrene into mouse embryos and fetuses.
Arch Toxicol, 62(2-3), 236-239.
Niedbala, R. S., Haley, N., Kardos, S., & Kardos, K. (2002). Automated homogeneous
immunoassay analysis of cotinine in urine. J Anal Toxicol, 26(3), 166-170.
NIH. (2007). National Institute of Health on Alcohol Abuse and Alcoholism.
Nikolaou, D., & Templeton, A. (2003). Early ovarian ageing: a hypothesis. Detection and clinical
relevance. Hum Reprod, 18(6), 1137-1139.
NTER. (February 21, 1990). National Security, Soil Preparation Services.
NTP Technical Reports 410 and 500. (2005). (March 6, 2005 ed.). NTP: Long-term Abstracts &
Reports.
Ogden, T. L. (2010). Handling results below the level of detection. Ann Occup Hyg, 54(3), 255-
256. doi:10.1093/annhyg/mep099
Okey, A. B. (1990). Enzyme induction in the cytochrome P-450 system. Pharmacol Ther, 45(2),
241-298.
Orlandini, C., Regini, C., Vellucci, F. L., Petraglia, F., & Luisi, S. (2015). Genes involved in the
pathogenesis of Premature Ovarian Insufficiency. Minerva Ginecol.
Otsuka, F., McTavish, K. J., & Shimasaki, S. (2011). Integral role of GDF-9 and BMP-15 in ovarian
function. Mol Reprod Dev, 78(1), 9-21. doi:10.1002/mrd.21265
Pal, L., Bevilacqua, K., & Santoro, N. F. (2010). Chronic psychosocial stressors are detrimental
to ovarian reserve: a study of infertile women. J Psychosom Obstet Gynaecol, 31(3),
130-139. doi:10.3109/0167482X.2010.485258
Pal, L., Bevilacqua, K., Zeitlian, G., Shu, J., & Santoro, N. (2008). Implications of diminished
ovarian reserve (DOR) extend well beyond reproductive concerns. Menopause, 15(6),
1086-1094. doi:10.1097/gme.0b013e3181728467
Parrott, J. A., & Skinner, M. K. (1999). Kit-ligand/stem cell factor induces primordial follicle
development and initiates folliculogenesis. Endocrinology, 140(9), 4262-4271.
doi:10.1210/endo.140.9.6994
Pe'er, I., de Bakker, P. I., Maller, J., Yelensky, R., Altshuler, D., & Daly, M. J. (2006). Evaluating
and improving power in whole-genome association studies using fixed marker sets. Nat
Genet, 38(6), 663-667. doi:10.1038/ng1816

104
Peluso, C., Fonseca, F. L., Rodart, I. F., Cavalcanti, V., Gastaldo, G., Christofolini, D. M., . . .
Bianco, B. (2014). AMH: An ovarian reserve biomarker in assisted reproduction. Clin
Chim Acta, 437, 175-182. doi:10.1016/j.cca.2014.07.029
Perera, F. P., Dickey, C., Santella, R., O'Neill, J. P., Albertini, R. J., Ottman, R., . . . Hemminki, K.
(1994). Carcinogen-DNA adducts and gene mutation in foundry workers with low-level
exposure to polycyclic aromatic hydrocarbons. Carcinogenesis, 15(12), 2905-2910.
Petchpoung, K., Kaojarern, S., Yoovathaworn, K., Sura, T., & Sirivarasai, J. (2011). The influence
of metabolic gene polymorphisms on urinary 1-hydroxypyrene concentration in Thai
bus drivers. Environ Toxicol Pharmacol, 31(1), 160-164. doi:10.1016/j.etap.2010.10.006
Phillips, D. H. (1996). DNA adducts in human tissues: biomarkers of exposure to carcinogens in
tobacco smoke. Environ Health Perspect, 104 Suppl 3, 453-458.
Phillips, D. H. (1999). Polycyclic aromatic hydrocarbons in the diet. Mutat Res, 443(1-2), 139-
147.
Pollenz, R. S. (2002). The mechanism of AH receptor protein down-regulation (degradation)
and its impact on AH receptor-mediated gene regulation. Chem Biol Interact, 141(1-2),
41-61.
Pope, C. A. (1989). Respiratory disease associated with community air pollution and a steel
mill, Utah Valley. Am J Public Health, 79(5), 623-628.
Pope, C. A., Thun, M. J., Namboodiri, M. M., Dockery, D. W., Evans, J. S., Speizer, F. E., & Heath,
C. W. (1995). Particulate air pollution as a predictor of mortality in a prospective study
of U.S. adults. Am J Respir Crit Care Med, 151(3 Pt 1), 669-674.
doi:10.1164/ajrccm/151.3_Pt_1.669
Popp, W., Vahrenholz, C., Schfll, C., Kraus, R., von Bülow, J., Müller, G., & Norpoth, K. (1995).
Risk estimation in coke-oven workers by determining some biomarkers of carcinogen
exposure. Exp Toxicol Pathol, 47(6), 440-442.
Preuss, R., Angerer, J., & Drexler, H. (2003). Naphthalene--an environmental and occupational
toxicant. Int Arch Occup Environ Health, 76(8), 556-576. doi:10.1007/s00420-003-0458-
1
Puga, A., Ma, C., & Marlowe, J. L. (2009). The aryl hydrocarbon receptor cross-talks with
multiple signal transduction pathways. Biochem Pharmacol, 77(4), 713-722.
doi:10.1016/j.bcp.2008.08.031

105
Rajapaksa, K. S., Cannady, E. A., Sipes, I. G., & Hoyer, P. B. (2007). Involvement of CYP 2E1
enzyme in ovotoxicity caused by 4-vinylcyclohexene and its metabolites. Toxicol Appl
Pharmacol, 221(2), 215-221. doi:10.1016/j.taap.2007.03.009
Rajapaksa, K. S., Sipes, I. G., & Hoyer, P. B. (2007). involvement of microsomal epoxide
hydrolase enzyme in ovotoxicity caused by 7,12-dimethylbenz[a]anthracene. Toxicol
Sci, 96(2), 327-334. doi:10.1093/toxsci/kfl202
Ramesh, A., Archibong, A. E., & Niaz, M. S. (2010). Ovarian susceptibility to benzo[a]pyrene:
tissue burden of metabolites and DNA adducts in F-344 rats. J Toxicol Environ Health A,
73(23), 1611-1625. doi:10.1080/15287394.2010.514225
Ramesh, A., Walker, S. A., Hood, D. B., Guillén, M. D., Schneider, K., & Weyand, E. H. (2004).
Bioavailability and risk assessment of orally ingested polycyclic aromatic hydrocarbons.
Int J Toxicol, 23(5), 301-333. doi:10.1080/10915810490517063
Rehwagen, M., Müller, A., Massolo, L., Herbarth, O., & Ronco, A. (2005). Polycyclic aromatic
hydrocarbons associated with particles in ambient air from urban and industrial areas.
Sci Total Environ, 348(1-3), 199-210. doi:10.1016/j.scitotenv.2004.12.050
Rekhadevi, P. V., Diggs, D. L., Huderson, A. C., Harris, K. L., Archibong, A. E., & Ramesh, A.
(2014). Metabolism of the environmental toxicant benzo(a)pyrene by subcellular
fractions of human ovary. Hum Exp Toxicol, 33(2), 196-202.
doi:10.1177/0960327113489050
Rogge, W. F., Hildemann, L. M., Mazurek, M. A., Cass, G. R., & Simoneit, B. R. (1994). Sources of
fine organic aerosol. 6. Cigaret smoke in the urban atmosphere. Environ Sci Technol,
28(7), 1375-1388. doi:10.1021/es00056a030
Romert, L., Dock, L., Jenssen, D., & Jernström, B. (1989). Effects of glutathione transferase
activity on benzo[a]pyrene 7,8-dihydrodiol metabolism and mutagenesis studied in a
mammalian cell co-cultivation assay. Carcinogenesis, 10(9), 1701-1707.
Rose, M., Holland, J., Dowding, A., Petch, S. R., White, S., Fernandes, A., & Mortimer, D. (2015).
Investigation into the formation of PAHs in foods prepared in the home to determine
the effects of frying, grilling, barbecuing, toasting and roasting. Food Chem Toxicol, 78,
1-9. doi:10.1016/j.fct.2014.12.018
Sadeu, J. C., & Foster, W. G. (2011). Effect of in vitro exposure to benzo[a]pyrene, a component
of cigarette smoke, on folliculogenesis, steroidogenesis and oocyte nuclear maturation.
Reprod Toxicol, 31(4), 402-408. doi:10.1016/j.reprotox.2010.12.006
Saengtienchai, A., Ikenaka, Y., Nakayama, S. M., Mizukawa, H., Kakehi, M., Bortey-Sam, N., . . .
Ishizuka, M. (2014). Identification of interspecific differences in phase II reactions:

106
determination of metabolites in the urine of 16 mammalian species exposed to
environmental pyrene. Environ Toxicol Chem, 33(9), 2062-2069. doi:10.1002/etc.2656
Safety, I. P. o. C., (IPCS), & WHO. (1998). Selected Nonheterocyclic Polycyclic Aromatic
Hydrocarbons. Geneva, Switzerland: World Health Organization.
Sahambi, S. K., Visser, J. A., Themmen, A. P., Mayer, L. P., & Devine, P. J. (2008). Correlation of
serum anti-Müllerian hormone with accelerated follicle loss following 4-
vinylcyclohexene diepoxide-induced follicle loss in mice. Reprod Toxicol, 26(2), 116-
122. doi:10.1016/j.reprotox.2008.07.005
Sancini, A., Tomei, F., Montuori, L., Caciari, T., Andrè, J. C., Rinaldi, G., . . . Ciarrocca, M. (2012).
Environmental and biological monitoring of polycyclic aromatic hydrocarbon levels in
outdoor workers exposed to urban stressor. Ann Ig, 24(5), 429-442.
Sanders, C. L., Skinner, C., & Gelman, R. A. (1984). Percutaneous absorption of [7.10-
14C]benzo[a]pyrene and [7,12-14C]dimethylbenz[a]anthracene in mice. Environ Res,
33(2), 353-360.
Santella, R. M., Nunes, M. G., Blaskovic, R., Perera, F. P., Tang, D., Beachman, A., . . . DeLeo, V.
A. (1994). Quantitation of polycyclic aromatic hydrocarbons, 1-hydroxypyrene, and
mutagenicity in urine of coal tar-treated psoriasis patients and untreated volunteers.
Cancer Epidemiol Biomarkers Prev, 3(2), 137-140.
Sassarini, J., & Lumsden, M. A. (2015). Oestrogen replacement in postmenopausal women. Age
Ageing, 44(4), 551-558. doi:10.1093/ageing/afv069
Schipper, I., Visser, J. A., Themmen, A. P., & Laven, J. S. (2012). Limitations and pitfalls of
antimüllerian hormone measurements. Fertil Steril, 98(4), 823-824.
doi:10.1016/j.fertnstert.2012.07.1105
Schlede, E., Kuntzman, R., & Conney, A. H. (1970). Stimulatory effect of benzo(alpha)pyrene
and phenobarbital pretreatment on the biliary excretion of benzo(alpha)pyrene
metabolites in the rat. Cancer Res, 30(12), 2898-2904.
Schmeltz, I., Tosk, J., & Hoffmann, D. (1976). Formation and determination of naphthalenes in
cigarette smoke. Anal Chem, 48(4), 645-650.
Schnekenburger, M., Peng, L., & Puga, A. (2007). HDAC1 bound to the Cyp1a1 promoter blocks
histone acetylation associated with Ah receptor-mediated trans-activation. Biochim
Biophys Acta, 1769(9-10), 569-578. doi:10.1016/j.bbaexp.2007.07.002

107
Schwartz, J., & Dockery, D. W. (1992). Increased mortality in Philadelphia associated with daily
air pollution concentrations. Am Rev Respir Dis, 145(3), 600-604.
doi:10.1164/ajrccm/145.3.600
Scott, R. T., & Hofmann, G. E. (1995). Prognostic assessment of ovarian reserve. Fertil Steril,
63(1), 1-11.
Scott, R. T., Opsahl, M. S., Leonardi, M. R., Neall, G. S., Illions, E. H., & Navot, D. (1995). Life
table analysis of pregnancy rates in a general infertility population relative to ovarian
reserve and patient age. Hum Reprod, 10(7), 1706-1710.
Seidel, A., Friedberg, T., Löllmann, B., Schwierzok, A., Funk, M., Frank, H., . . . Glatt, H. (1998).
Detoxification of optically active bay- and fjord-region polycyclic aromatic hydrocarbon
dihydrodiol epoxides by human glutathione transferase P1-1 expressed in Chinese
hamster V79 cells. Carcinogenesis, 19(11), 1975-1981.
Seifer, D. B., Baker, V. L., & Leader, B. (2011). Age-specific serum anti-Müllerian hormone
values for 17,120 women presenting to fertility centers within the United States. Fertil
Steril, 95(2), 747-750. doi:10.1016/j.fertnstert.2010.10.011
Seifer, D. B., & Merhi, Z. (2014). Is AMH a regulator of follicular atresia? J Assist Reprod Genet,
31(11), 1403-1407. doi:10.1007/s10815-014-0328-7
Seitz, H. K., & Stickel, F. (2006). Risk factors and mechanisms of hepatocarcinogenesis with
special emphasis on alcohol and oxidative stress. Biol Chem, 387(4), 349-360.
doi:10.1515/BC.2006.047
Shabad, L. M., & Cohan, Y. L. (1972). The contents of benzo(a)pyrene in some crops. Arch
Geschwulstforsch, 40(6), 237-243.
Sharara, F. I., Scott, R. T., & Seifer, D. B. (1998). The detection of diminished ovarian reserve in
infertile women. Am J Obstet Gynecol, 179(3 Pt 1), 804-812.
Sheweita, S. A. (2000). Drug-metabolizing enzymes: mechanisms and functions. Curr Drug
Metab, 1(2), 107-132.
Shimada, T., & Fujii-Kuriyama, Y. (2004). Metabolic activation of polycyclic aromatic
hydrocarbons to carcinogens by cytochromes P450 1A1 and 1B1. Cancer Sci, 95(1), 1-6.
Shimada, T., Sugie, A., Shindo, M., Nakajima, T., Azuma, E., Hashimoto, M., & Inoue, K. (2003).
Tissue-specific induction of cytochromes P450 1A1 and 1B1 by polycyclic aromatic
hydrocarbons and polychlorinated biphenyls in engineered C57BL/6J mice of
arylhydrocarbon receptor gene. Toxicol Appl Pharmacol, 187(1), 1-10.

108
Siegel, E., & Wason, S. (1986). Mothball toxicity. Pediatr Clin North Am, 33(2), 369-374.
Smith, K. W., Proctor, S. P., Ozonoff, A. L., & McClean, M. D. (2012). Urinary biomarkers of
occupational jet fuel exposure among Air Force personnel. J Expo Sci Environ Epidemiol,
22(1), 35-45. doi:10.1038/jes.2011.38
Sobinoff, A. P., Mahony, M., Nixon, B., Roman, S. D., & McLaughlin, E. A. (2011). Understanding
the Villain: DMBA-induced preantral ovotoxicity involves selective follicular destruction
and primordial follicle activation through PI3K/Akt and mTOR signaling. Toxicol Sci,
123(2), 563-575. doi:10.1093/toxsci/kfr195
Sowers, M. R., Eyvazzadeh, A. D., McConnell, D., Yosef, M., Jannausch, M. L., Zhang, D., . . .
Randolph, J. F. (2008). Anti-mullerian hormone and inhibin B in the definition of ovarian
aging and the menopause transition. J Clin Endocrinol Metab, 93(9), 3478-3483.
doi:10.1210/jc.2008-0567
Stephen, E. H., & Chandra, A. (2006). Declining estimates of infertility in the United States:
1982-2002. Fertil Steril, 86(3), 516-523. doi:10.1016/j.fertnstert.2006.02.129
Strickland, P., & Kang, D. (1999). Urinary 1-hydroxypyrene and other PAH metabolites as
biomarkers of exposure to environmental PAH in air particulate matter. Toxicol Lett,
108(2-3), 191-199.
Strogi, K. (2007). Monitoring of environmental exposure to polycyclic aromatic hydrocarbons: a
review. Environmental Chemistry Letters, 5, 169-195.
Strohsnitter, W. C., Hatch, E. E., Hyer, M., Troisi, R., Kaufman, R. H., Robboy, S. J., . . . Noller, K.
L. (2008). The association between in utero cigarette smoke exposure and age at
menopause. Am J Epidemiol, 167(6), 727-733. doi:10.1093/aje/kwm351
Su, H. I., Sammel, M. D., Freeman, E. W., Lin, H., DeBlasis, T., & Gracia, C. R. (2008a). Body size
affects measures of ovarian reserve in late reproductive age women. Menopause,
15(5), 857-861. doi:10.1097/gme.0b013e318165981e
Su, H. I., Sammel, M. D., Freeman, E. W., Lin, H., DeBlasis, T., & Gracia, C. R. (2008b). Body size
affects measures of ovarian reserve in late reproductive age women. Menopause,
15(5), 857-861. doi:10.1097/gme.0b013e318165981e
Sullivan, J., & Krieger, G. (2001). Clinical Environmental Health and Toxic Exposures (2nd ed.).
Philadelphia, PA: Lippincott Wilkins & Wilkes.
Suwan-ampai, P., Navas-Acien, A., Strickland, P., & Agnew, J. (2009). Involuntary Tobacco
Smoke Exposure and Urinary Levels of Polycyclic Aromatic Hydrocarbons in the United
States, 1999 to 2002. Cancer Epidemiology Biomarkers Prev, 18(3).

109
Suzuki, N., Yoshioka, N., Takae, S., Sugishita, Y., Tamura, M., Hashimoto, S., . . . Kawamura, K.
(2015). Successful fertility preservation following ovarian tissue vitrification in patients
with primary ovarian insufficiency. Hum Reprod, 30(3), 608-615.
doi:10.1093/humrep/deu353
Sweeney, B. P., & Bromilow, J. (2006). Liver enzyme induction and inhibition: implications for
anaesthesia. Anaesthesia, 61(2), 159-177. doi:10.1111/j.1365-2044.2005.04462.x
Takizawa, K., Yagi, H., Jerina, D. M., & Mattison, D. R. (1984). Murine strain differences in
ovotoxicity following intraovarian injection with benzo(a)pyrene, (+)-(7R,8S)-oxide, (-)-
(7R,8R)-dihydrodiol, or (+)-(7R,8S)-diol-(9S,10R)-epoxide-2. Cancer Res, 44(6), 2571-
2576.
Tas, S., Buchet, J. P., & Lauwerys, R. (1994). Determinants of benzo[a]pyrene diol epoxide
adducts to albumin in workers exposed to polycyclic aromatic hydrocarbons. Int Arch
Occup Environ Health, 66(5), 343-348.
Tfouni, S., Serrate, C., Leme, F., Camargo, M., Teles, C., Cipolle, K., & Furlani, R. (2013).
Polycyclic aromatic hydrocarbons in coffee brew:Influence of roasting and brewing
procedures in two Coffea cultivars. LWT-Food Science and Technology, 50(2), 526-530.
Tingen, C., Stanford, J. B., & Dunson, D. B. (2004). Methodologic and statistical approaches to
studying human fertility and environmental exposure. Environ Health Perspect, 112(1),
87-93.
Tomic, D., Miller, K. P., Kenny, H. A., Woodruff, T. K., Hoyer, P., & Flaws, J. A. (2004). Ovarian
follicle development requires Smad3. Mol Endocrinol, 18(9), 2224-2240.
doi:10.1210/me.2003-0414
Torino, F., Barnabei, A., De Vecchis, L., Appetecchia, M., Strigari, L., & Corsello, S. M. (2012).
Recognizing menopause in women with amenorrhea induced by cytotoxic
chemotherapy for endocrine-responsive early breast cancer. Endocr Relat Cancer,
19(2), R21-33. doi:10.1530/ERC-11-0199
Tsai-Turton, M., & Luderer, U. (2006). Opposing effects of glutathione depletion and follicle-
stimulating hormone on reactive oxygen species and apoptosis in cultured preovulatory
rat follicles. Endocrinology, 147(3), 1224-1236. doi:10.1210/en.2005-1281
Tsai-Turton, M., Luong, B. T., Tan, Y., & Luderer, U. (2007). Cyclophosphamide-induced
apoptosis in COV434 human granulosa cells involves oxidative stress and glutathione
depletion. Toxicol Sci, 98(1), 216-230. doi:10.1093/toxsci/kfm087
Tsai-Turton, M., Nakamura, B. N., & Luderer, U. (2007). Induction of apoptosis by 9,10-
dimethyl-1,2-benzanthracene in cultured preovulatory rat follicles is preceded by a rise

110
in reactive oxygen species and is prevented by glutathione. Biol Reprod, 77(3), 442-451.
doi:10.1095/biolreprod.107.060368
Turkall, R. M., Skowronski, G. A., Kadry, A. M., & Abdel-Rahman, M. S. (1994). A comparative
study of the kinetics and bioavailability of pure and soil-adsorbed naphthalene in
dermally exposed male rats. Arch Environ Contam Toxicol, 26(4), 504-509.
Tuttle, A. M., Stämpfli, M., & Foster, W. G. (2009). Cigarette smoke causes follicle loss in mice
ovaries at concentrations representative of human exposure. Hum Reprod, 24(6), 1452-
1459. doi:10.1093/humrep/dep023
U.S. Bureau of Labor Statistics, 2010 and Previous Years Current Population Survey. (2010).
United States Environmental Protection Agency. (January 2006).
van de Wiel, J. A., Fijneman, P. H., Duijf, C. M., Anzion, R. B., Theuws, J. L., & Bos, R. P. (1993).
Excretion of benzo[a]pyrene and metabolites in urine and feces of rats: influence of
route of administration, sex and long-term ethanol treatment. Toxicology, 80(2-3), 103-
115.
van Delft, J. H., Steenwinkel, M. S., van Asten, J. G., de Vogel, N., Bruijntjes-Rozier, T. C.,
Schouten, T., . . . Hopmans, P. M. (2001). Biological monitoring the exposure to
polycyclic aromatic hydrocarbons of coke oven workers in relation to smoking and
genetic polymorphisms for GSTM1 and GSTT1. Ann Occup Hyg, 45(5), 395-408.
van Dorp, W., van den Heuvel-Eibrink, M. M., de Vries, A. C., Pluijm, S. M., Visser, J. A., Pieters,
R., & Laven, J. S. (2014). Decreased serum anti-Müllerian hormone levels in girls with
newly diagnosed cancer. Hum Reprod, 29(2), 337-342. doi:10.1093/humrep/det442
van Houten, E. L., Themmen, A. P., & Visser, J. A. (2010). Anti-Müllerian hormone (AMH):
regulator and marker of ovarian function. Ann Endocrinol (Paris), 71(3), 191-197.
doi:10.1016/j.ando.2010.02.016
Van Rooij, J. G., Veeger, M. M., Bodelier-Bade, M. M., Scheepers, P. T., & Jongeneelen, F. J.
(1994). Smoking and dietary intake of polycyclic aromatic hydrocarbons as sources of
interindividual variability in the baseline excretion of 1-hydroxypyrene in urine. Int Arch
Occup Environ Health, 66(1), 55-65.
Viau, C., Vyskocil, A., & Martel, L. (1995). Background urinary 1-hydroxypyrene levels in non-
occupationally exposed individuals in the Province of Québec, Canada, and comparison
with its excretion in workers exposed to PAH mixtures. Sci Total Environ, 163(1-3), 191-
194.

111
Visser, J. A., de Jong, F. H., Laven, J. S., & Themmen, A. P. (2006). Anti-Müllerian hormone: a
new marker for ovarian function. Reproduction, 131(1), 1-9. doi:10.1530/rep.1.00529
Visser, J. A., Schipper, I., Laven, J. S., & Themmen, A. P. (2012). Anti-Müllerian hormone: an
ovarian reserve marker in primary ovarian insufficiency. Nat Rev Endocrinol, 8(6), 331-
341. doi:10.1038/nrendo.2011.224
Visser, J. A., & Themmen, A. P. (2005). Anti-Müllerian hormone and folliculogenesis. Mol Cell
Endocrinol, 234(1-2), 81-86. doi:10.1016/j.mce.2004.09.008
Visser, J. A., & Themmen, A. P. (2014). Role of anti-Müllerian hormone and bone
morphogenetic proteins in the regulation of FSH sensitivity. Mol Cell Endocrinol, 382(1),
460-465. doi:10.1016/j.mce.2013.08.012
Vyskocil, A., Fiala, Z., Chénier, V., Krajak, L., Ettlerova, E., Bukac, J., . . . Emminger, S. (2000).
Assessment of multipathway exposure of small children to PAH. Environ Toxicol
Pharmacol, 8(2), 111-118.
Wang, H., Chu, W., Das, S. K., Ren, Q., Hasstedt, S. J., & Elbein, S. C. (2002). Liver pyruvate
kinase polymorphisms are associated with type 2 diabetes in northern European
Caucasians. Diabetes, 51(9), 2861-2865.
Wang, H., Chu, W. S., Hemphill, C., & Elbein, S. C. (2002). Human resistin gene: molecular
scanning and evaluation of association with insulin sensitivity and type 2 diabetes in
Caucasians. J Clin Endocrinol Metab, 87(6), 2520-2524. doi:10.1210/jcem.87.6.8528
Watanabe, J., Shimada, T., Gillam, E. M., Ikuta, T., Suemasu, K., Higashi, Y., . . . Kawajiri, K.
(2000). Association of CYP1B1 genetic polymorphism with incidence to breast and lung
cancer. Pharmacogenetics, 10(1), 25-33.
Waylen, A. L., Jones, G. L., & Ledger, W. L. (2010). Effect of cigarette smoking upon
reproductive hormones in women of reproductive age: a retrospective analysis. Reprod
Biomed Online, 20(6), 861-865. doi:10.1016/j.rbmo.2010.02.021
WCRF/AICR, Food, Nutrition, Physical Activity and the Prevention of Cancer: A Global
Perspective 2007. (September 27, 2010). Retrieved from
Weenen, C., Laven, J. S., Von Bergh, A. R., Cranfield, M., Groome, N. P., Visser, J. A., . . .
Themmen, A. P. (2004). Anti-Müllerian hormone expression pattern in the human
ovary: potential implications for initial and cyclic follicle recruitment. Mol Hum Reprod,
10(2), 77-83.
Weinberg, C. R., Wilcox, A. J., & Baird, D. D. (1989). Reduced fecundability in women with
prenatal exposure to cigarette smoking. Am J Epidemiol, 129(5), 1072-1078.

112
Welt, C. K., & Schneyer, A. L. (2001). Differential regulation of inhibin B and inhibin a by follicle-
stimulating hormone and local growth factors in human granulosa cells from small
antral follicles. J Clin Endocrinol Metab, 86(1), 330-336. doi:10.1210/jcem.86.1.7107
Welt, C. K., Smith, Z. A., Pauler, D. K., & Hall, J. E. (2001). Differential regulation of inhibin A and
inhibin B by luteinizing hormone, follicle-stimulating hormone, and stage of follicle
development. J Clin Endocrinol Metab, 86(6), 2531-2537. doi:10.1210/jcem.86.6.7597
Wenzlaff, A. S., Cote, M. L., Bock, C. H., Land, S. J., Santer, S. K., Schwartz, D. R., & Schwartz, A.
G. (2005). CYP1A1 and CYP1B1 polymorphisms and risk of lung cancer among never
smokers: a population-based study. Carcinogenesis, 26(12), 2207-2212.
doi:10.1093/carcin/bgi191
Westerholm, R., & Li, H. (1994). A multivariate statistical analysis of fuel-related polycyclic
aromatic hydrocarbon emissions from heavy-duty diesel vehicles. Environ Sci Technol,
28(5), 965-972. doi:10.1021/es00054a032
WHO, W. H. O. R. O. f. E. (1987). Polynuclear aromatic hydrocarbons (PAH) In Air quality
guidelines for Europe (pp. 105-117). Copenhagen.
WHO, Regional Office for Europe, Air Quality Guidelines, Chapter, 5.9, Polycyclic Aromatic
Hydrocarbons. (1987). Copenhagen.
Wiersma, D. A., & Roth, R. A. (1983). Total body clearance of circulating benzo(a)pyrene in
conscious rats: effect of pretreatment with 3-methylcholanthrene and the role of liver
and lung. J Pharmacol Exp Ther, 226(3), 661-667.
Withey, J. R., Law, F. C., & Endrenyi, L. (1991). Pharmacokinetics and bioavailability of pyrene in
the rat. J Toxicol Environ Health, 32(4), 429-447. doi:10.1080/15287399109531494
Withey, J. R., Shedden, J., Law, F. C., & Abedini, S. (1993). Distribution of benzo[a]pyrene in
pregnant rats following inhalation exposure and a comparison with similar data
obtained with pyrene. J Appl Toxicol, 13(3), 193-202.
Wolff, R. K., Bond, J. A., Sun, J. D., Henderson, R. F., Harkema, J. R., Griffith, W. C., . . .
McClellan, R. O. (1989). Effects of adsorption of benzo[a]pyrene onto carbon black
particles on levels of DNA adducts in lungs of rats exposed by inhalation. Toxicol Appl
Pharmacol, 97(2), 289-299.
Wyatt, K. M., Dimmock, P. W., Walker, T. J., & O'Brien, P. M. (2001). Determination of total
menstrual blood loss. Fertil Steril, 76(1), 125-131.
Xue, Q., Kato, D., Kamata, T., Guo, Q., You, T., & Niwa, O. (2013). Human cytochrome P450 3A4
and a carbon nanofiber modified film electrode as a platform for the simple evaluation

113
of drug metabolism and inhibition reactions. Analyst, 138(21), 6463-6468.
doi:10.1039/c3an01313h
Xue, W., & Warshawsky, D. (2005). Metabolic activation of polycyclic and heterocyclic aromatic
hydrocarbons and DNA damage: a review. Toxicol Appl Pharmacol, 206(1), 73-93.
doi:10.1016/j.taap.2004.11.006
Yang, J. J., Roy, T. A., & Mackerer, C. R. (1986). Percutaneous absorption of benzo[a]pyrene in
the rat: comparison of in vivo and in vitro results. Toxicol Ind Health, 2(4), 409-416.
Yang, M., Koga, M., Katoh, T., & Kawamoto, T. (1999). A study for the proper application of
urinary naphthols, new biomarkers for airborne polycyclic aromatic hydrocarbons. Arch
Environ Contam Toxicol, 36(1), 99-108.
Yong, A. G., & Pearce, S. (2013). A Beginners Guide to Factor Analysis: Focusing on Exploratory
Factor Analysis. Tutorials in Quantitative Methods for Psychololgy, (2, 9).
Zeilinkova, Z., & Wenzi, T. (2015). The Occurrence of 16 EPA PAHs in Food - A Review.
Polycyclic Aromatic Compounds, 35(2 - 4), 248-284.
Zenzes, M. T., Puy, L. A., & Bielecki, R. (1998). Immunodetection of benzo[a]pyrene adducts in
ovarian cells of women exposed to cigarette smoke. Mol Hum Reprod, 4(2), 159-165.
Zhang, L., Jin, Y., Huang, M., & Penning, T. M. (2012). The Role of Human Aldo-Keto Reductases
in the Metabolic Activation and Detoxication of Polycyclic Aromatic Hydrocarbons:
Interconversion of PAH Catechols and PAH o-Quinones. Front Pharmacol, 3, 193.
doi:10.3389/fphar.2012.00193
Zhang, X. J., Shi, Z., Lyv, J. X., He, X., Englert, N. A., & Zhang, S. Y. (2015). Pyrene is a Novel
Constitutive Androstane Receptor (CAR) Activator and Causes Hepatotoxicity by CAR.
Toxicol Sci, 147(2), 436-445. doi:10.1093/toxsci/kfv142
Zheng, W., Nagaraju, G., Liu, Z., & Liu, K. (2012a). Functional roles of the phosphatidylinositol
3-kinases (PI3Ks) signaling in the mammalian ovary. Mol Cell Endocrinol, 356(1-2), 24-
30. doi:10.1016/j.mce.2011.05.027
Zheng, W., Nagaraju, G., Liu, Z., & Liu, K. (2012b). Functional roles of the phosphatidylinositol
3-kinases (PI3Ks) signaling in the mammalian ovary. Mol Cell Endocrinol, 356(1-2), 24-
30. doi:10.1016/j.mce.2011.05.027
Zoller, L., Lambalk, C., & de Koning, (1991). C. Ultrasound of the Ovary
Quantitative Analysis of the Membrana Granulosa in Developing and Ovulatory Follicles
(Vol. 9).