investigation of the reversible inhibition of camel (camelus dromedarius) acetylcholinesterase by...
TRANSCRIPT

Camp. &o&em. Physiof. Vol. 10X, No. 2, pp. 323-327, 1993 0306492/93 $6.00 + 0.00 Printed in Great Britain 0 1993 Pergamon Press Ltcl
INVESTIGATION OF THE REVERSIBLE INHIBITION OF CAMEL (CASSIUS ~~~~~~A~U~)
ACETYLCHOLINESTERASE BY TETRACAINE
ABDULAZIZ A. AL-JAFARI
Department of Biochemistry, College of Science, King Saud University, P.O. Box 245.5, Riyadh 11451, Saudi Arabia (Tel. 01 467-5953; Fax 01 467-6462)
(Received 23 November 1992; accepted for ~blica~io~ 17 February 1993)
Abstract-l. The camel erythrocyte membrane bound acetylcholinesterase (AChE) was extracted with the non-ionic detergent Triton X-100 and some of its kinetics parameters were studied. In addition the effect of tetracaine hydrochloride on AChE was also investigated.
2. The Michael&Menten constant (&,) for the hydrolysis of acetylthiocholine iodide was found to be 7 x low5 M and the V,, was 21.2 ~mol~hr/mg protein.
3. Tetracaine (0.025-~.80 mM) reversibly inhibited the AChE activity (2582%) in a concentration- dependent manner, the tcB being about 0.12 mM.
4. The Lineweaver-Burk plot and its secondary plots indicated that the nature of this inhibition is of the linear mixed type. This mixed type inhibition system is considered to be composed of partial competitive and pure non-competitive in nature.
5. The values of $(s~Opj and Y,(inarceptj were estimated as 0.127 mM and 0.263 mM, respectively, by a secondary replot of primary double reciprocal plot of Lineweaver-Burk plot and Dixon plot.
6. &/K, ratio shows that tetracaine has a greater aflinity of binding to the active site than to a peripheral site.
INTRODUCTION
A~tylchoIin~sterase (AChE; 3.1.1.7) is present in large amounts in human erythrocytes and is mainly located in particular areas of the central and parasympathetic nervous systems. In general terms the principal role of AChE is concerned with the final stages of impulse transmission at cholinergic synapses involving the rapid hydrolysis of the neurotrans- mittor acetylcholine (Ott, 1985). The role of AChE in red blood cells is unclear but its presence has activated interest in studying various parameters such as its purification, characterization, inhibition kinetic studies by various inhibitors, molecular struc- ture, its relationship to cell maturation and its varying activity in certain diseases etc. in various animals (Lawson and Barr, 1987). Human erythro- cyte AChE is membrane-bound with its catalytic activity directed totally towards the outside of the membrane (Frenkel et al., 1980). AChE can be solubilized with detergents such as Triton X-100 su~esting that it is an integral membrane protein (Ott et al., 1975).
Erythrocyte AChE has been investigated in man, bovine, pig, ox, rat, mouse, horse, and sheep (Silman and Futerman, 1987) but so far there are no reports on the enzyme from camels. The camel is common to many areas of the world having adapted to the ~vironment both anatomically and bi~hemically (Barakat and Fattah, 1970). It is possible, therefore, that camel erythrocyte AChE shows variations from the enzyme obtained from other sources. For
example, the molecular forms of AChE and its kinetic parameters vary from tissue to tissue within the same animal. These variations reflect the specific oper- ational requirements at the different types of synapses at which these molecular forms of AChE function (Fedorov, 1981).
The aim of the present investigation is to study the interaction of tetracaine with a new source of acetylcholinesterase i.e. camel erythrocyte membrane bound AChE. Moreover, the investigation may provide some insight in to the nature of camel erythrocyte AChE.
MATERIALS AND METHODS
The origin of some of the chemicals used in this study was as follows: acetylthiocholine iodide (ASCh, used as substrate) came from BDH Chemicals Ltd., Poole, (England); 5,5’-dithiobis-(2-nitro) benzoic acid (DTNB) was purchased from Sigma, tetracaine hydro~hlo~de was obtained from Pfaltz & Bauer; Inc., (New York) and bovine serum albumin was obtained from Fluka. All other chemicals used were of analytical reagent grade.
Enzyme preparation
Camel blood was collected from healthy young male camels (Came&s dromedarius) during slaughter- ing at a local slaughter house in the presence of Na-EDTA, anticoagulant (1.5 mg/ml). The blood
323
324 A. A. AL-JAFARI
was kept in an ice-box for 1 hr until further process- ing. Erythrocytes were separated from plasma by centrifugation at 1000 g for 30 min at 4°C and the plasma and the buffy coat removed by aspiration; the cells were then washed twice with 10 volumes of 310 imOsM, Tris/HCl buffer, pH 7.6, by re-centrifu- gation as described above. The washed cells were resuspended in the same 310 imOsM, isotonic Tris buffer to a haematocrit of 50% and mixed well by inversion for 1 min. The 1 volume of 50% haemat- ocrit was added drop by drop with constant stirring in 7.5 volumes of lysis buffer (20 imOsM Tris/HCl buffer, pH 7.8, 10 mM imidazole, 1 mM NaCl and 1 mM MgCl,). After lysis, the “ghosts” were har- vested by centrifugation at 20,000 g for 40 min at 4°C and the supernatant was taken out by suction. The “ghosts” were washed 4 times with lysis buffer while the supernatant fractions of each washing were kept for analysis of AChE activity (not shown). The finally washed colourless ghosts were resuspended in 25 mM Tris/HCl buffer/l20 mM NaCl, pH 7.4 for estimation of AChE activity as well as protein concentration (Fleischer and Packer, 1974; Beauregard and Roufo- galis, 1979).
Camel erythrocyte membranes (1.8 mg/ml) were
solubilized with 1% Triton X-100. After centrifu- gation at 100,OOOg for 1 hr at 4°C (45 Ti rotor, Beckman ultracentrifuge L8-80) the AChE activity was quantitatively (70% yield with 4.7 purification factor) recovered in the supernatant.
Assay of AChE activity
AChE activity was determined at 37°C as described
by Ellman et al. (1961). The assay mixture (3 ml) contained 0.5 mM ASCh, 0.150 mM DTNB and 8.9 pg/ml as a native membrane protein while 18.5 pg/ml Triton solubilized protein in 25 mM Tris/HCl buffer/l20mM NaCl, pH 7.4. The 5 min incubation time was selected for the enzyme assay, after preliminary experiments performed to ensure that enzyme activity is linear with respect to reaction time and the enzyme concentration employed.
To study the in vitro effect of tetracaine, the enzyme was preincubated with tetracaine at 37°C for 5 min prior to the addition of substrate.
Time dependence
Tetracaine (0.05 mM) was incubated with AChE
for up to 60 min before addition of substrate. Tetra- Caine was added after the substrate to another sample of each set was assayed in the absence of tetracaine and served as a control. All 12 assays were run simultaneously. The experiment was repeated twice more and was also conducted 3 times with tetracaine present at 0.12 mM and 0.30 mM.
Substrate effects
The activity of AChE in the presence and absence of 0.30 mM tetracaine respectively was recorded in 3.0 ml buffer with a 0.025 mM concentration of
OL I #
0 20 40 60
Preincubation time
( min )
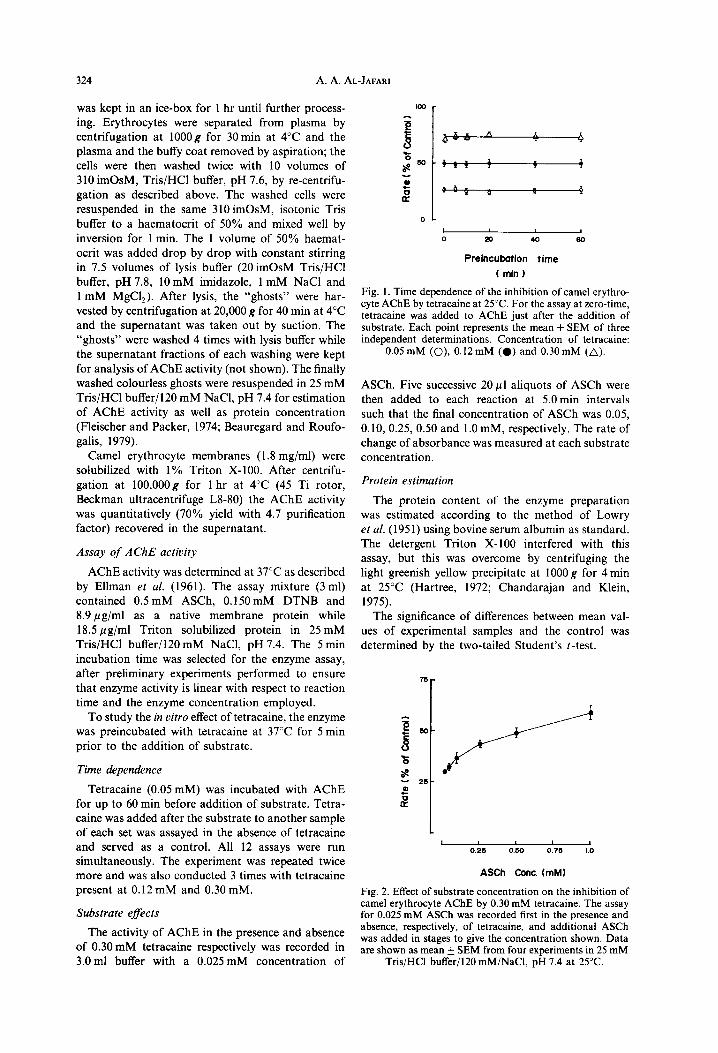
Fig. 1. Time dependence of the inhibition of camel erythro- cyte AChE by tetracaine at 25°C. For the assay at zero-time, tetracaine was added to AChE just after the addition of substrate. Each point represents the mean k SEM of three independent determinations. Concentration of tetracaine:
0.05 mM (O), 0.12mM (0) and 0.30mM (A).
ASCh. Five successive 20~1 aliquots of ASCh were then added to each reaction at 5.0 min intervals such that the final concentration of ASCh was 0.05, 0.10, 0.25, 0.50 and 1.0 mM, respectively. The rate of change of absorbance was measured at each substrate concentration.
Protein estimation
The protein content of the enzyme preparation was estimated according to the method of Lowry et al. (1951) using bovine serum albumin as standard. The detergent Triton X-100 interfered with this assay, but this was overcome by centrifuging the light greenish yellow precipitate at 1000 g for 4 min at 25°C (Hartree, 1972; Chandarajan and Klein, 1975).
The significance of differences between mean val- ues of experimental samples and the control was determined by the two-tailed Student’s t-test.
L
1 I
0.21 0.50 0.71 1.0
ASCh Cont. (mM)
Fig. 2. Effect of substrate concentration on the inhibition of camel erythrocyte AChE by 0.30 mM tetracaine. The assay for 0.025 mM ASCh was recorded first in the oresence and absence, respectively, of tetracaine, and additional ASCh was added in stages to give the concentration shown. Data are shown as mean + SEM from four experiments in 25 mM
Tris/HCl buffer/l20 mM/NaCl, pH 7.4 at 25°C.
Reversible inhibition of acetylcholinesterase by tetracaine 325
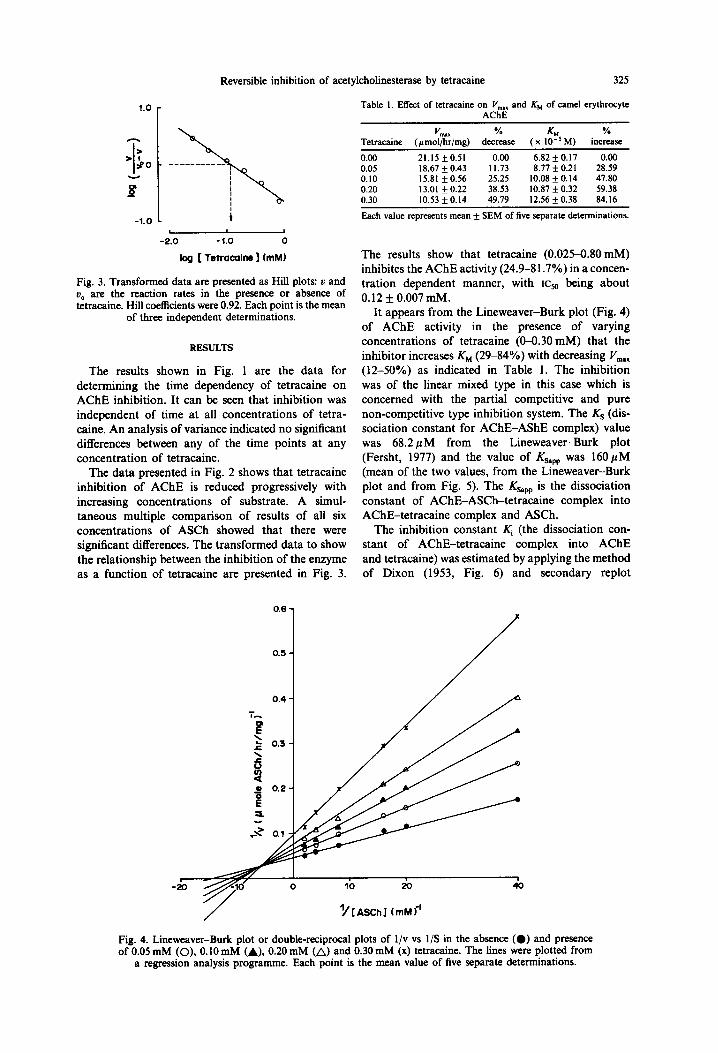
Table 1. Et&t of tetracaine on I’,,,,, and Kkl of camel erythrocyte AChE
V mm % K” % Tetracaine (pmol/hr/mg) decrease ( x 10m5 M) increase
0.00 21.15 +0.51 0.00 6.82 f 0.17 0.00 0.05 18.67 + 0.43 11.73 8.77 f 0.21 28.59 0.10 15.81 + 0.56 25.25 10.08 f 0.14 47.80 0.20 13.01 f 0.22 38.53 10.87 f 0.32 59.38 0.30 10.53 f 0.14 49.79 12.56 f 0.38 84.16
-1.0 0 I
-2.0 - 1.0 0
log [ Tetrocoim I (mM)
Fig. 3. Transformed data are presented as Hill plots: u and U, are the reaction rates in the presence or absence of tetracaine. Hill coefficients were 0.92. Each point is the mean
of three independent determinations.
RESULTS
The results shown in Fig. 1 are the data for determining the time dependency of tetracaine on AChE inhibition. It can be seen that inhibition was independent of time at all concentrations of tetra- caine. An analysis of variance indicated no significant differences between any of the time points at any concentration of tetracaine.
The data presented in Fig. 2 shows that tetracaine inhibition of AChE is reduced progressively with increasing concentrations of substrate. A simul- taneous multiple comparison of results of all six concentrations of ASCh showed that there were significant differences. The transformed data to show the relationship between the inhibition of the enzyme as a function of tetracaine are presented in Fig. 3.
Each value represents mean + SEM of five separate determinations.
The results show that tetracaine (0.025-0.80mM) inhibites the AChE activity (24.9-81.7%) in a concen- tration dependent manner, with I+, being about 0.12 + 0.007 mM.
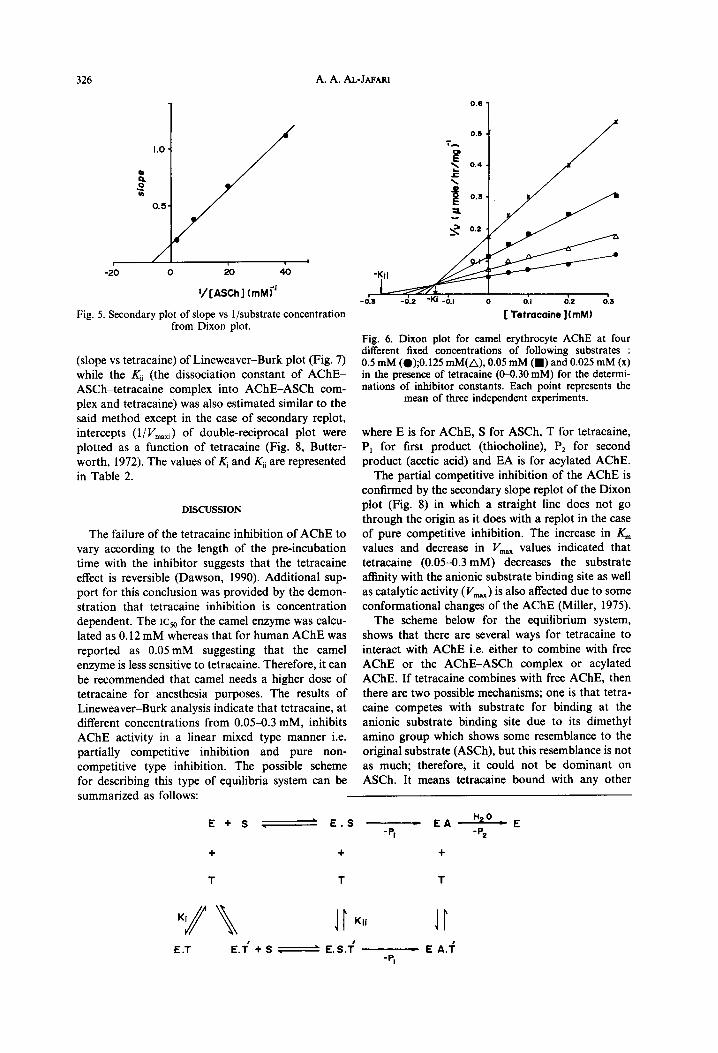
It appears from the Lineweaver-Burk plot (Fig. 4) of AChE activity in the presence of varying concentrations of tetracaine (O-0.30 mM) that the inhibitor increases KM (29-84%) with decreasing I’,,,., (12-50%) as indicated in Table 1. The inhibition was of the linear mixed type in this case which is concerned with the partial competitive and pure non-competitive type inhibition system. The KS (dis- sociation constant for AChE-AShE complex) value was 68.2pM from the Lineweaver-Burk plot (Fersht, 1977) and the value of Ksapp was 160pM (mean of the two values, from the Lineweaver-Burk plot and from Fig. 5). The Ksapp is the dissociation constant of AChE-ASCh-tetracaine complex into AChE-tetracaine complex and ASCh.
The inhibition constant Ki (the dissociation con- stant of AChE-tetracaine complex into AChE and tetracaine) was estimated by applying the method of Dixon (1953, Fig. 6) and secondary replot
0.6 -
0.5 -
‘/[ASChl tmMf’
Fig. 4. Lineweaver-Burk plot or double-reciprocal plots of l/v vs l/S in the absence (0) and presence of 0.05 mM (O), 0.10 mM (A), 0.20 mM (A) and 0.30 mM (x) tetracaine. The lines were plotted from
a regression analysis programme. Each point is the mean value of five separate determinations.
326 A. A. AL-JAFARI
I
0 20 40
‘/[ASCh] bnkd
Fig. 5. Secondary plot of slope vs l/substrate concentration from Dixon plot.
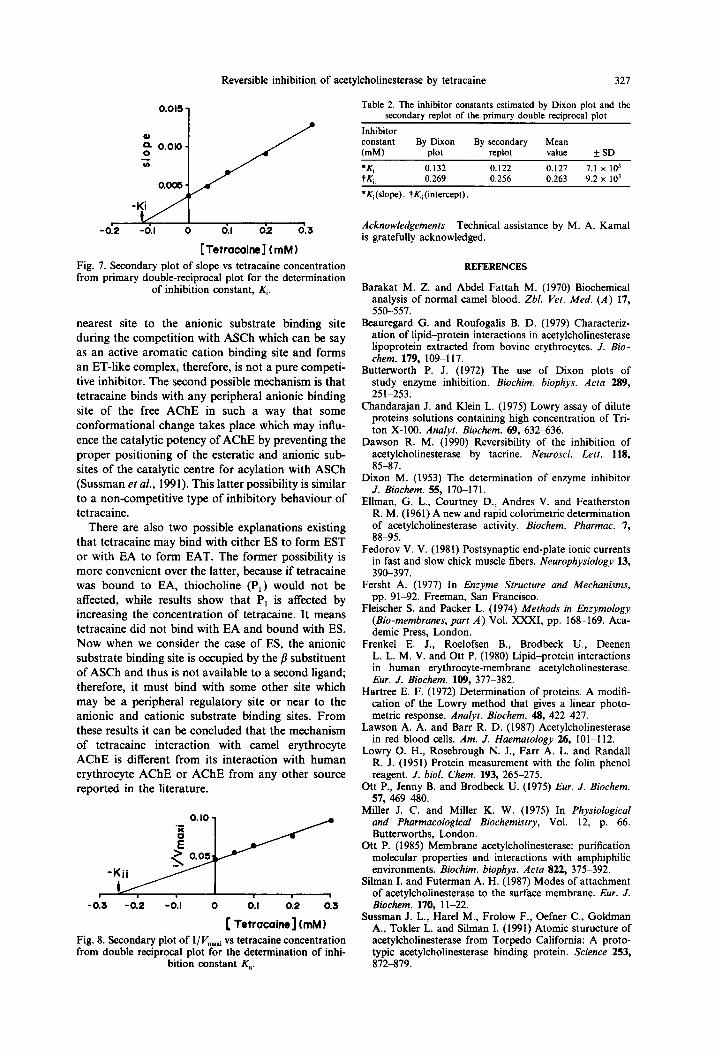
(slope vs tetracaine) of Lineweaver-Burk plot (Fig. 7) while the Ki, (the dissociation constant of AChE- ASCh-tetracaine complex into AChE-ASCh com- plex and tetracaine) was also estimated similar to the said method except in the case of secondary replot, intercepts (l/ Vmati) of double-reciprocal plot were plotted as a function of tetracaine (Fig. 8, Butter- worth, 1972). The values of $ and Kii are represented in Table 2.
DISCUSSION
The failure of the tetracaine inhibition of AChE to vary according to the length of the pre-incubation time with the inhibitor suggests that the tetracaine effect is reversible (Dawson, 1990). Additional sup- port for this conclusion was provided by the demon- stration that tetracaine inhibition is concentration dependent. The ICY,, for the camel enzyme was calcu- lated as 0.12 mM whereas that for human AChE was reported as 0.05 mM suggesting that the camel enzyme is less sensitive to tetracaine. Therefore, it can be recommended that camel needs a higher dose of tetracaine for anesthesia purposes. The results of Lineweaver-Burk analysis indicate that tetracaine, at different concentrations from 0.0543 mM, inhibits AChE activity in a linear mixed type manner i.e. partially competitive inhibition and pure non- competitive type inhibition. The possible scheme for describing this type of equilibria system can be
,-!i -0.3 -0.2 0
[ ‘Gocaine 10;2mMI 0.3
Fig. 6. Dixon plot for camel erythrocyte AChE at four different fixed concentrations of following substrates : 0.5 mM (@);0.125 mM(A), 0.05 mM (W) and 0.025 mM (x) in the presence of tetracaine (SO.30 mM) for the determi- nations of inhibitor constants. Each point represents the
mean of three independent experiments.
where E is for AChE, S for ASCh, T for tetracaine, P, for first product (thiocholine), P2 for second product (acetic acid) and EA is for acylated AChE.
The partial competitive inhibition of the AChE is confirmed by the secondary slope replot of the Dixon plot (Fig. 8) in which a straight line does not go through the origin as it does with a replot in the case of pure competitive inhibition. The increase in K,,, values and decrease in I’,,,,_ values indicated that tetracaine (0.05-0.3 mM) decreases the substrate affinity with the anionic substrate binding site as well as catalytic activity (V,,) is also affected due to some conformational changes of the AChE (Miller, 1975).
The scheme below for the equilibrium system, shows that there are several ways for tetracaine to interact with AChE i.e. either to combine with free AChE or the AChE-ASCh complex or acylated AChE. If tetracaine combines with free AChE, then there are two possible mechanisms; one is that tetra- Caine competes with substrate for binding at the anionic substrate binding site due to its dimethyl amino group which shows some resemblance to the original substrate (ASCh), but this resemblance is not as much; therefore, it could not be dominant on ASCh. It means tetracaine bound with any other
summarized as follows:
E+S -ES -. -5
c EA r,‘” E 2
+ + +
T T T
I E.T E.T+S- . . -ES-i - E A.+
-PI
Reversible inhibition of acetylcholinesterase by tetracaine 327
Table 2. The inhibitor constants estimated by Dixon plot and the secondary replot of the primary double reciprocal plot
42 4.l 0 6.1 oi d.3
[Tetracaine] (mM) Fig. 7. Secondary plot of slope vs tetracaine concentration from primary double-reciprocal plot for the determination
of inhibition constant, K,.
nearest site to the anionic substrate binding site during the competition with ASCh which can be say as an active aromatic cation binding site and forms an ET-like complex, therefore, is not a pure competi- tive inhibitor. The second possible mechanism is that tetracaine binds with any peripheral anionic binding site of the free AChE in such a way that some conformational change takes place which may influ- ence the catalytic potency of AChE by preventing the proper positioning of the esteratic and anionic sub- sites of the catalytic centre for acylation with ASCh (Sussman et al., 1991). This latter possibility is similar to a non-competitive type of inhibitory behaviour of tetracaine.
There are also two possible explanations existing that tetracaine may bind with either ES to form EST or with EA to form EAT. The former possibility is more convenient over the latter, because if tetracaine was bound to EA, thiocholine (P,) would not be affected, while results show that P, is affected by increasing the concentration of tetracaine. It means tetracaine did not bind with EA and bound with ES. Now when we consider the case of ES, the anionic substrate binding site is occupied by the /l substituent of ASCh and thus is not available to a second ligand; therefore, it must bind with some other site which may be a peripheral regulatory site or near to the anionic and cationic substrate binding sites. From these results it can be concluded that the mechanism of tetracaine interaction with came1 erythrocyte AChE is different from its interaction with human erythrocyte AChE or AChE from any other source reported in the literature.
-0.3 -0:2 -d.l 0 0.J 012 d.3
[ Tetracaine] (rnhd) Fig. 8. Secondary plot of l/V,, vs tetracaine concentration from double reciprocal plot for the determination of inhi-
bition constant K,,.
Inhibitor constant By Dixon By secondary Mean WJ) plot replot value +SD
;:, 0.132 0.269 0.122 0.256 0.127 0.263 9.2 7.1 x x IO’ 103
*K,(slope). t&(intercept).
Acknowledgements-Technical assistance by M. A. Kamal is gratefully acknowledged.
REFERENCES
Barakat M. Z. and Abdel Fattah M. (1970) Biochemical analysis of normal camel blood. Zbl. Vet. Med. (A) 17, 550-557.
Beauregard G. and Roufogalis B. D. (1979) Characteriz- ation of lipid-protein interactions in acetylcholinesterase lipoprotein extracted from bovine erythrocytes. J. Bio- them. 179, 109-117.
Butterworth P. J. (1972) The use of Dixon plots of study enzyme inhibition. Biochirn. biophys. Acta 289, 251-253.
Chandarajan J. and Klein L. (1975) Lowry assay of dilute proteins solutions containing high concentration of Tri- ton X-100. Analyt. Biochem. 69, 632636.
Dawson R. M. (1990) Reversibility of the inhibition of acetylcholinesterase by tacrine. Neurosci. Leff. 118, 85-87.
Dixon M. (1953) The determination of enzyme inhibitor J. Biochem. 55, 17&171.
Ellman, G. L., Courtney D., Andres V. and Featherston R. M. (1961) A new and rapid calorimetric determination of acetylcholinesterase activity. Biochem. Pharmac. 7, 88-95.
Fedorov V. V. (1981) Postsynaptic end-plate ionic currents in fast and slow chick muscle fibers. Neurophysiology 13, 390-397.
Fersht A. (1977) In Enzyme Siructure and Mechanisms, pp. 91-92. Freeman, San Francisco.
Fleischer S. and Packer L. (1974) Methods in Enzymology (Bio-membranes, part A) Vol. XxX1, pp. 168-169. Aca- demic Press, London.
Frenkel E. J., Roelofsen B., Brodbeck U., Deenen L. L. M. V. and Ott P. (1980) Lipid-protein interactions in human erythrocyte-membrane acetylcholinesterase. Eur. J. Biochem. 109, 377-382.
Hartree E. F. (1972) Determination of proteins. A modifi- cation of the Lowry method that gives a linear photo- metric response. Analyt. Biochem. 48, 422427.
Lawson A. A. and Barr R. D. (1987) Acetylcholinesterase in red blood cells. Am. J. Haemafology 26, 101-112.
Lowry 0. H., Rosebrough N. J., Farr A. L. and Randall R. J. (1951) Protein measurement with the folin phenol reagent. J. biol. Chem. 193, 265-275.
Ott P., Jenny B. and Brodbeck U. (1975) Eur. J. Biochem. 57, 469480.
Miller J. C. and Miller K. W. (1975) In Physiological and Pharmacological Biochemistry, Vol. 12, p. 66. Butterworths, London.
Ott P. (1985) Membrane acetylcholinesterase: purification molecular properties and interactions with amphiphilic environments. Biochim. biophys. Acta 822, 375-392.
Silman I. and Futerman A. H. (1987) Modes of attachment of acetylcholinesterase to the surface membrane. Eur. J. Biochem. 170, 1 l-22.
Sussman J. L., Hare1 M., Frolow F., Oefner C., Goldman A., Tokler L. and Silman I. (1991) Atomic sturucture of acetylcholinesterase from Torpedo California: A proto- typic acetylcholinesterase binding protein. Science 253, 872-879.