interactions between bacillus subtilis early spore coat morphogenetic proteins
TRANSCRIPT

R E S E A R C H L E T T E R
InteractionsbetweenBacillus subtilis early spore coatmorphogenetic proteinsDenisa Mullerova, Daniela Krajcıkova & Imrich Barak
Institute of Molecular Biology, Slovak Academy of Sciences, Bratislava, Slovakia
Correspondence: Imrich Barak, Institute of
Molecular Biology, Slovak Academy of
Sciences, Dubravska cesta 21, Bratislava,
Slovakia. Tel.: 1421 2 593 074 18; fax: 1421
2 593 074 16; e-mail: [email protected]
Received 1 June 2009; accepted 13 July 2009.
Final version published online 24 August 2009.
DOI:10.1111/j.1574-6968.2009.01737.x
Editor: Ezio Ricca
Keywords
Bacillus subtilis ; spore coat; protein–protein
interactions; yeast two-hybrid assay; pull-down
assay.
Abstract
When challenged by stresses such as starvation, the soil bacterium Bacillus subtilis
produces an endospore surrounded by a proteinaceous coat composed of 4 70
proteins that are organized into three main layers: an amorphous undercoat,
lightly staining lamellar inner coat and electron-dense outer coat. This coat
protects the spore against a variety of chemicals or lysozyme. Mutual interactions
of the coat’s building blocks are responsible for the formation of this structurally
complex and extraordinarily resistant shell. However, the assembly process of
spore coat proteins is still poorly understood. In the present work, the main focus
is on the three spore coat morphogenetic proteins: SpoIVA, SpoVID and SafA.
Direct interaction between SpoIVA and SpoVID proteins was observed using a
yeast two-hybrid assay and verified by coexpression experiment followed by
Western blot analysis. Coexpression experiments also confirmed previous findings
that SpoVID and SafA directly interact, and revealed a novel interaction between
SpoIVA and SafA. Moreover, gel filtration analysis revealed that both SpoIVA and
SpoVID proteins form large oligomers.
Introduction
The sporulation process in Bacillus subtilis is accomplished
by formation of a proteinaceous spore coat that grants the
cell protection against oxidizing agents, lytic enzymes and
toxic molecules (Setlow, 2000, 2003) and enables the dor-
mant cell to endure for long periods of time (Nicholson
et al., 2000). This protein complex is composed of 4 70
species and creates a double-layered structure visible under
electron microscope: the inner coat, which is formed by
three to six fine, lightly staining lamellae in juxtaposition to
each other and the outer coat, thicker and darker than the
inner layer, coarsely striated (Driks, 1999; Henriques &
Moran, 2000, 2007; Kim et al., 2006).
Although the spore coat consists of large number of
proteins, only a small group of them plays a crucial role in
coat morphology. Hence, these proteins are referred to as
morphogenetic (Driks & Setlow, 2000; Driks, 2002; Henriques
et al., 2004; Kim et al., 2006). The first essential morphoge-
netic protein engaged in spore coat assembly is SpoIVA.
Transcription of the spoIVA gene commences early, 2 h after
the onset of sporulation, under control of sE transcription
factor from two closely spaced promoters. spoIVA mutant
produces some coat material but this material is misassembled
as swirls and accumulates in the mother cell cytosol (Roels
et al., 1992). The peptidoglycan cortex layer, which lies just
underneath the coat, is also misformed in mutant cells,
indicating that SpoIVA is necessary for proper formation of
both the cortex and the coat (Coote, 1972; Piggot & Coote,
1976). Catalano et al. (2001) predicted that these two func-
tions of SpoIVA are not genetically separable and are not
encoded by distinct domains of the protein. Deposition of
SpoIVA protein is connected to, and serves as a marker for, the
septation and engulfment process, because SpoIVA protein
tracks along the enveloping membrane surrounding the fore-
spore and creates a shell adjacent to this membrane (Pogliano
et al., 1995). Price & Losick (1999) revealed that the extreme
C-terminal portion of SpoIVA is critical for its targeting to the
forespore and depends on small peptide SpoVM (Levin et al.,
1993). These authors (Price & Losick, 1999) also predicted an
oligomerization domain outside the C terminus of SpoIVA
protein, which might be responsible for interaction between
SpoIVA molecules. Further study showed that such a domain
does indeed exist (Ramamurthi & Losick, 2008). SpoIVA
possesses a canonical Walker A box that binds and hydrolyzes
ATP, which is needed for multimerization and assembly of
FEMS Microbiol Lett 299 (2009) 74–85c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
MIC
ROBI
OLO
GY
LET
TER
S

SpoIVA into filamentous structures forming a shell encasing
the forespore (Ramamurthi & Losick, 2008).
SpoIVA forms a basement layer on top of which other spore
coat proteins are deposited. Proper assembly of SpoIVA is a
prerequisite for the recruitment of another morphogenetic
and structural component of the coat – CotE. Expression of
the cotE gene is switched on early in the mother cell and is
under control of sE transcription factor. CotE protein assem-
bles in very much the same fashion as SpoIVA, as a coating
around the forespore, but the formation of CotE shell is
delayed compared with SpoIVA and is separated from the
forespore surface by a 75-nm-wide gap. This space is called a
matrix and it is where infiltration of the inner coat compo-
nents takes place (Driks et al., 1994; Pogliano et al., 1995).
Thus, CotE sits at the edge of the matrix, between the inner
and outer coat layer, and from this position guides the correct
formation of the latter. In cotE mutant cells, the outer coat is
missing (Zheng et al., 1988).
Apart from SpoIVA, another protein is involved in the
attachment of CotE to the forespore. This protein is a product
of the spoVID gene, which is transcribed by sE-containing
RNA polymerase and codes for acidic, 575-residue-long
protein (Beall et al., 1993). SpoVID is not responsible for the
initial targeting of CotE to the forespore but for the continued
attachment of CotE and the coat in the later stages of
sporulation. In spoVID null mutant, a defect similar to that
of spoIVA mutant was observed; the coat material accumu-
lated in the mother cell cytoplasm as swirls instead of shell
around the forespore. However, spoVID mutant cells possess
normal-looking cortex (Beall et al., 1993; Driks et al., 1994).
The SpoVID protein sequence contains a cell wall-binding
LysM motif, facilitating its deployment to the cortex–coat
boundary (Ozin et al., 2000; Costa et al., 2006). The same
motif was observed in the N-terminal part of another protein
– SafA (SpoVID-associated factor A) (Kodama et al., 1999).
SpoVID and SafA accumulate early in the sporulation
process and localize to the matrix region (Ozin et al., 2000).
Localization of SafA to the pre-engulfed forespore is an
early, SpoVID- and SpoIVA-dependent event. Yeast two-
hybrid experiments as well as in vitro pull-down assays
revealed that SpoVID and SafA proteins interact directly
(Ozin et al., 2001). The interaction is mediated by two
motifs present in SafA protein sequence: the PYYH motif in
the C-terminal half of SafA, and a region just downstream of
the LysM domain consisting of 13 residues. In SpoVID
protein sequence, amino acids 1–202 are involved in contact
with SafA. Besides SafA, other interacting partners were
suggested for SpoVID protein, among them SpoIVA (Ozin
et al., 2001; Costa et al., 2006). SafA mutant spores are
susceptible to lysozyme and form spores with abnormal
coats that lack several coat protein components, an impact
not as severe as that of spoVID null mutant (Takamatsu
et al., 1999; Ozin et al., 2000).
In this study, the potential direct protein–protein inter-
actions between three major morphogenetic proteins
SpoIVA, SpoVID and SafA were examined using yeast two-
hybrid assay and by coexpression followed by an in vitro pull-
down experiment. Also investigated were the portions of the
proteins responsible for this interaction. Further on, isolated
SpoIVA and SpoVID were subjected to gel filtration analysis
to gain a better insight into their oligomerization properties.
Materials and methods
Strains and cultivation media
The Escherichia coli strain MM294 (endA1 hsdR17 supE44
thi-1 recA1) (Backman et al., 1976) was used for cloning and
amplification of all recombinant plasmids. Escherichia coli
strain BL21(DE3) [F� ompT hsdSB (rB�mB
�) gal dcm (DE3)]
(Novagen) was used for expression and overproduction of
histidine-tagged fusion proteins. Escherichia coli strain
AD202 was used for expression and overproduction of the
glutathione-S-transferase (GST) fusion SpoIVA protein.
Luria–Bertani (LB) medium with appropriate antibiotics
was used for maintenance and growth of E. coli strains.
Bacillus subtilis strain PY79 (Youngman et al., 1984) was
cultivated in LB media and its genomic DNA served for
amplification of genes encoding spore coat proteins.
Saccharomyces cerevisiae strain MaV203 (Vidal, 1997)
(MATa , leu2-3, 112, trp1-901, his3D200, ade2-101, gal4D,
gal80D, SPAL10<URA3, GAL1<lacZ, HIS3UAS GAL1<
HIS3@LYS2, can1R, cyh2R) was used for the yeast two-hybrid
assay. Yeasts were cultivated in media recommended by the
manufacturer (Invitrogen). Rich YPAD medium was used
for routine growth of yeasts, and synthetic complete (SC)
medium, omitting specific amino acids according to selec-
tion requirements, was used for examining protein–protein
interactions.
Construction of yeast plasmids
The ProQuestTM Two-Hybrid system with Gateways tech-
nology (Invitrogen) was used. Coding regions of spoIVA,
spoVID and safA genes were amplified by PCR using
Y2HspoIVA50, Y2HspoVID50 and Y2HsafA50 as sense primers
and Y2HspoIVA30, Y2HspoVID30 and Y2HsafA30 as antisense
primers. Primers were flanked by attB cloning sites necessary
for fragment insertion into pDONR221 vector by BP reaction,
generating an entry clone. In the next step, genes from the
entry clones were transferred into yeast destination vectors
pDEST22 and pDEST32 via LR reaction. Site-specific recom-
bination generates in-frame fusions with the GAL4 DNA-
binding domain (in pDEST32) or the GAL4 activation
domain (in pDEST22). In pDONR221 vector, the kanamycin
resistance gene serves as a selection marker. Vector pDEST32
includes the LEU2 gene for selection in yeast on medium
FEMS Microbiol Lett 299 (2009) 74–85 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
75Interactions of early spore coat proteins

lacking the leucine and gentamicin resistance gene for main-
tenance in E. coli. Vector pDEST22 contains a TRP1 gene to
compensate yeast’s tryptophan auxotrophy, and an ampicillin
resistance gene enabling E. coli carrying this plasmid to grow
in media containing ampicillin.
Truncated mutants of the spoIVA gene were prepared via
PCR amplification using two sets of primers: Y2HspoIVA50 and
TDspoIVA30, respectively, thus creating spA2 (1200 bp) trun-
cated mutant; a second set of primers, lTDspoIVA50 and
Y2HspoIVA30, respectively, generated spA4 (1173 bp) truncated
mutant of spoIVA. All primers were flanked by attB cloning sites.
PCR-generated truncated mutants were cloned into pDONR221
and pDEST32/pDEST22 vectors as described above.
Truncated mutants of the spoVID gene were also amplified
by PCR using two sets of primers: spD2 truncated version
(1194-bp) of spoVID was amplified by PCR using primers
Y2HspoVID50 and spoVID39930; 1128-bp truncated mutant of
the spoVID gene, named spD3, was prepared via PCR with
primers spoVID201–57550 and Y2HspoVID30, respectively. All
primers were flanked by attB cloning sites. PCR-generated
truncated mutants were cloned into pDONR221 and
pDEST32/pDEST22 vectors as described above.
In all PCR amplifications, chromosomal DNA of B. sub-
tilis PY79 was used as a template. PCR products were
purified using QIA quick Gel Extraction Kit (Qiagen). New
plasmid constructs were verified by restriction analysis and
sequencing.
Construction of expression plasmids
DNA manipulations were carried out according to Sambrook
et al. (1989). The entire spoVID coding region (1728 bp) and
a 1200-bp-long fragment of spoIVA (1479 bp) were ampli-
fied by PCR using sense primers SpoVID50 and SpoIVA50,
respectively, and antisense primers SpoVID30 and SpoIVA30,
respectively (Table 1). PCR products were digested with
NdeI and XhoI restriction enzymes and cloned between the
same sites of pET28a expression vector (Novagen), yielding
expression plasmids pETspoIVA and pETspoVID, respec-
tively. Inserted genes carry a six-histidine-tag fused to their
N terminus.
For coexpression and pull-down experiments, full-length
spoIVA and spoVID genes were amplified by PCR. For
amplification of spoIVA, sense primer duetSpA50, carrying
BglII restriction site, and antisense primer duetSpA30,
carrying XhoI restriction site, were used. Primers duetSpD-
his50 and duetSpD-his30 containing BamHI and PstI restric-
tion sites were used to amplify the spoVID gene (Table 1).
Table 1. Oligonucleotides used for cloning
Primers Sequence 50–30
SpoIVA50 AGGAGGAGGCATATGGAAAAGGTCGATATTTTCAAGG
SpoIVA30 AGGAGGAGGGCTCGAGTCAGATCGACGGAGCGAC
SpoVID50 GGAGGAGGACATATGCCGCAAAATCATCGATTG
SpoVID30 TGGTGGTGGCTCGAGTTACGCATGGCTATTTTTATATTG
DuetSpA50 AGGAGGAGGAGATCTCGAAAAGGTCGATATTTTCAAGG
DuetSpA30 AGGAGGAGGCTCGAGTTACAGGATGATGGCGATTAAG
DuetSpD50 GGAGGAGGACATATGCCGCAAAATCATCGATTG
DuetSpD30 TGGTGGTGGCTCGAGTTACGCATGGCTATTTTTATATTG
DuetSpD-his50 GGAGGAGGAGGATCCGCCGCAAAATCATCGATTGC
DuetSpD-his30 TGGTGGTGGCTGCAGTTACGCATGGCTATTTTTATATTG
SafA50 Duet CATCATCATGGATCCGAAAATCCATATCGTTCAAAAAGG
SafA30 Duet CATCATCATCTGCAGTTACTCATTTTCTTCTTCCGGAC
spA(GEX)50 AGGAGGAGGGGATCCGAAAAGGTCGATATTTTCAAGG
spA(GEX)30 TGGTGGTGGCTCGAGTTACAGGATGATGGCGATTAA
Y2HspoIVA50 GGGGACAAGTTTGTACAAAAAAGCAGGCTTCGAAAAGGTCGATATTTTCAAGGATATC
Y2HspoIVA30 GGGGACCACTTTGTACAAGAAAGCTGGGTCTTACAGGATGATGGCGATTAAGC
Y2HspoVID50 GGGGACAAGTTTGTACAAAAAAGCAGGCTTCCCGCAAAATCATCGATTGCAATTTTC
Y2HspoVID30 GGGGACCACTTTGTACAAGAAAGCTGGGTCTTACGCATGGCTATTTTTATATTGAGG
Y2HsafA50 GGGGACAAGTTTGTACAAAAAAGCAGGCTTCAAAATCCATATCGTTCAAAAAGG
Y2HsafA30 GGGGACCACTTTGTACAAGAAAGCTGGGTCTTACTCATTTTCTTCTTCCGGAC
ITDspoIVA50 GGGGACAAGTTTGTACAAAAAAGCAGGCTTCGTGCCCGGCGCTAAAGG
TDspoIVA30 GGGGACCACTTTGTACAAGAAAGCTGGGTCTCAATGGATCGACGGAGCGAC
SpoVID201–57550 GGGGACAAGTTTGTACAAAAAAGCAGGCTTCCTCCGTGAAGAACTGGAGAC
SpoVID39930 GGGGACCACTTTGTACAAGAAAGCTGGGTCTCAATTGAAATGGAAATGTAAATCGGCATTG
Restriction enzymes sides are underlined. Sites required by Gateway cloning are double underlined.
FEMS Microbiol Lett 299 (2009) 74–85c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
76 D. Mullerova et al.

PCR products were digested with corresponding enzymes
and ligated into pETDuet-1 vectors (Novagen), digested
with the same enzymes, thus creating pETDuet(spoIVA) and
pETDuet(his-spoVID) expression plasmids where the
spoVID gene was N-terminally tagged with His tag. Further
on, pETDuet(his-spoVID) plasmid was digested with BglII
and XhoI restriction enzymes and the PCR-amplified
spoIVA gene, digested with the same enzymes, and cloned into
it to form pETDuet(spoIVA1his-spoVID) plasmid, coexpres-
sing both proteins (SpoIVA and SpoVID, respectively).
For coexpression and triple pull-down assay, the gene-
encoding spoIVA was cut out from pETDuet(spoIVA) using
restriction enzymes BglII and XhoI and recloned into
pACYCDuet-1 vector (Novagen) digested with the same
enzymes, thus generating pACYC(spoIVA). Genes for safA
and spoVID were both cloned into the same pETDuet-1
vector with safA carrying the N-terminal His tag. Primer
pair SafA50 Duet and SafA30 Duet (Table 1) was used to
PCR-amplify the coding region of safA (1164 bp). The
resulting PCR product was digested with restriction en-
zymes BamHI and PstI, respectively, and ligated with
pETDuet-1 vector cleaved with corresponding enzymes,
thus creating pETDuet(his-safA). The coding sequence of
spoVID was generated by PCR using primers duetSpD 50 and
duetSpD 30 (Table 1), respectively. The product was subse-
quently digested with NdeI and XhoI restriction enzymes
and cloned into pETDuet(his-safA), which was digested
accordingly, creating pETDuet(spD1his-safA).
For the in vitro pull-down assay, primer pairs
spA(GEX)50 and spA(GEX)30 (Table 1) were used for PCR
amplification of the coding region of spoIVA gene. The
resulting product was digested with BamHI and XhoI
restriction enzymes and cloned between the same sites of
pGEX-4T-1 vector (Amersham Pharmacia). In this recom-
binant plasmid, named pGEXspA, the spoIVA gene carries
the N-terminal GST tag.
In all PCR amplifications, chromosomal DNA of B.
subtilis PY79 was used as a template. PCR products were
purified using QIA quick Gel Extraction Kit (Qiagen). New
plasmid constructs were verified by restriction analysis.
Expression and purification of proteins andantibody production
Expression plasmids were transformed into E. coli strain
BL21(DE3) and bacteria were cultivated in LB medium
supplemented with appropriate antibiotic at 37 1C until the
cells reached an OD600 nm between 0.5 and 0.7. Then the
expression of recombinant protein was induced by addition
of isopropyl-b-D-1-thiogalactopyranoside (IPTG) at 1 mM
concentration and cells were further incubated with shaking
at 28 1C for 3 h. Bacteria were collected by centrifugation,
resuspended in solubilization buffer (20 mM Tris-Cl pH 8.0,
150 mM NaCl) and lysed by sonication (total time of 2 min,
with 10-s bursts and 35-s cooling periods). The cells with
expressed SpoIVA protein were resuspended in solubiliza-
tion buffer with 8 M urea, as SpoIVA is produced, to a large
extent, as an insoluble protein. Cell debris was removed
by centrifugation for 25 min at c. 73 000 g in a Beckman
centrifuge. Supernatant was applied on 1-mL Ni21-Sephar-
ose HP chelating column (Amersham Biosciences). Proteins
were eluted by a 4-mL concentration step gradient of
100 mM up to 1 M imidazole. Proteins were subsequently
used for antibody production in mice.
Escherichia coli strain AD202 was transformed by GST
fusion plasmid pGEX-4T-1 and cultivated in LB medium
with appropriate antibiotic at 37 1C until OD600 nm reached
0.5–0.7. Expression was then induced with 1 mM IPTG and
cells were further grown at 30 1C with shaking for 3 h before
harvesting by centrifugation. Cells were then processed
according to a GST resin manufacturer’s protocol (bio-
WORLD). Fusion protein was eluted by boiling the GST
Sepharose resin with an equal amount of 2� sodium
dodecyl sulfate (SDS) sample buffer for 1 min.
Pull-down assay of purified GST-SpoIVA andHis-tagged SpoVID
Recombinant His-SpoVID protein expressed from
pET(spoVID) plasmid was produced and purified as de-
scribed in the previous section. Purified His-SpoVID was
subsequently incubated with GST-SpoIVA bound to glu-
tathione Sepharose beads and with beads alone. The mixture
was then washed and proteins were eluted as recommended
by the manufacturer (bioWORLD). Interacting proteins
were resolved by SDS-10% polyacrylamide gel electrophor-
esis (PAGE) and identified by Western blot analysis.
Pull-down assay of coexpressed proteins
Recombinant proteins expressed from pETDuet-1 and
pACYCDuet-1 vectors were produced and purified as described
in the previous section. Interacting proteins were resolved by
SDS-10% PAGE and identified by Western blot analysis.
Western blot analysis
Purified protein samples were boiled for 10 min with
identical volumes of 2� SDS sample buffer and resolved by
SDS-10% PAGE. Fractionated proteins were subsequently
transferred onto nitrocellulose membrane (Hybond ECL,
Amersham Biosciences), which was then blocked in 5%
nonfat milk in TBST buffer to avoid nonspecific protein
binding. Proteins bound to the membrane were detected
by polyclonal antibodies against SpoIVA and SpoVID pro-
teins at 1 : 2000 dilution and by monoclonal anti-His-tag
FEMS Microbiol Lett 299 (2009) 74–85 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
77Interactions of early spore coat proteins

antibody (Novagen) at 1 : 1000 dilution. As a secondary
antibody, anti-mouse HRP conjugated antibody at 1 : 5000
dilution was used (Promega).
Yeast two-hybrid assay of protein--protein interactions
Saccharomyces cerevisiae strain MaV203 was cotransformed
with both pDEST32 (bait vector) and pDEST22 (prey
vector) yeast plasmids carrying the genes of interest, by the
lithium-acetate method (Gietz & Woods, 2002). Transfor-
mants were grown for 72 h on SC dropout medium lacking
amino acids leucine (Leu) and tryptophan (Trp). Large
colonies were picked up and resuspended in 70 mL of sterile
water and 2 mL droplets were applied on selection plates.
Yeast cells, in which the protein–protein interaction induced
the expression of three reporter genes (HIS3, URA3 and
lacZ), were able to grow on SC-Leu-Trp-His1100 mM 3AT
and SC-Leu-Trp-Ura plates, and their colony color changed
to blue in the presence of X-gal (5-bromo-4-chloro-3-
indolyl-b-D-galactoside). Autoactivation of reporter genes
was excluded by testing the yeast strains after cotransforma-
tion of all bait vectors with pEAXP-AD502 vector and all
prey vectors with pDBLeu vector.
Quantitative b -galactosidase assay inliquid culture
The strength of the interactions was measured by b-galacto-
sidase activity using o-nitrophenol-b-D-galactopyranoside
as a substrate according to the manufacturer’s protocol.
Activity of b-galactosidase was expressed in Miller units
[1 MU = 1000�OD420 nm/(t�V�OD600 nm); where t is the
time in incubation in minutes; V the volume of culture used
in the assay; OD420 nm the absorbance of o-nitrophenol;
OD600 nm the cell density of the culture]. The number of
Miller units (Table 2) represents the average values of at least
five independent isolated colonies and each sample was
assayed in triplicate to reduce variability.
Verification of candidate clones --retransformation assay
Positive interactions between two proteins were reassessed
for false positives by retransformation assay. The cells from
single colony were grown in SC-Leu-Trp (10 mL) at 30 1C
for 24 h. Centrifuged cells were disrupted mechanically
by vortexing with autoclaved acid-washed glass beads
for 2 min and plasmids were isolated using QIAprep Spin
Miniprep Kit (Qiagen) according to a protocol supplied.
Plasmids were transformed into E. coli XL1 Blue and trans-
formants were cultivated on LB plates with 100 mg mL�1
ampicillin to selectively isolate pDEST22 plasmids and
10 mg mL�1 gentamicin to selectively isolate pDEST32 plas-
mids. Isolated plasmids were verified by restriction analysis
and reintroduced into MaV203 yeast strain. Transformants
were grown on selection plates to reproduce the original
phenotype.
Analysis of proteins by size-exclusionchromatography
The Superose 12 column 10/300 GL (Amersham Pharmacia)
was calibrated with low-molecular-weight calibration pro-
teins (Amersham Pharmacia) in 50 mM Tris-HCl, 150 mM
NaCl, pH 8.0. Exclusion limit of this column (Mr) is
approximately 2� 106 proteins and its optimal separation
range (globular proteins) is 1� 103–3� 105. Purified re-
combinant proteins were loaded in total volume of 0.5 mL.
Chromatography was performed at a flow rate of
0.5 mL min�1. The eluted proteins were detected with UV
detector at 280 nm and confirmed by Western blot analysis.
Results
SpoIVA and SpoVID directly interact in the yeasttwo-hybrid system
In our previous work (Krajcikova et al., 2009), the main
focus involved uncovering novel direct protein–protein
interactions between spore coat proteins. Among those
examined by the yeast two-hybrid system and giving a
positive signal were two morphogenetic proteins, SpoIVA
and SpoVID. Additionally, an earlier study carried out by
Costa et al. (2006) showed the localization dependency of
SpoVID on SpoIVA protein and proposed that a direct
interaction may exist between them.
The coding regions of both proteins were fused either to
the GAL4 activation domain (AD) or the DNA-binding
domain (DB) and plasmids were introduced into S. cerevi-
siae strain MaV203. Interaction between proteins of interest
in this study was revealed by reconstitution of active GAL4
transcription factor when screening the combination
SpoIVA-AD and SpoIVD-DB. The expression of three
Table 2. b-Galactosidase activity of interacting proteins expressed in
Miller units, using ONPG as a substrate
Fusion protein DB/AD
b-Galactosidase
(Miller units)
SpoVID/SpoIVA 7.7�2.3
SpoVID201–575/SpoIVA 6.7�2.7
SpoVID/pEAXP-AD502 0.2�0.1
SpoVID201–575/pEAXP-AD502 0.1�0.0
Measured Miller units were averaged and the standard deviations were
calculated. The table lists the levels of lacZ expression in yeasts carrying
vectors with our genes of interest.
ONPG, o-nitrophenol-b-D-galactopyranoside.
FEMS Microbiol Lett 299 (2009) 74–85c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
78 D. Mullerova et al.

reporter genes was activated, allowing the cells to grow on
various selection plates as well as changing the colony color
to blue on X-gal assay (Fig. 1a). However, when SpoIVA was
fused to the DNA-binding domain and SpoVID to the
activation domain the interaction did not occur (Fig. 1a)
or was below the detection limit of this method. One
possible explanation for this bias might be misfolding of
the protein or steric hindrance when combined with a
certain domain of GAL4. Nonetheless, these results suggest
that there may be a direct interaction between these two
morphogenetic proteins. The interaction of SpoIVA with
itself as well as SpoVID with itself was not detected by the
yeast two-hybrid experiment. This observation, however,
does not rule out the possibility of oligomerization of these
proteins. Polymerization of SpoIVA molecules was ob-
served in the work of Price & Losick (1999). In agreement
with previous results mentioned above, Ozin et al. (2001)
also found no evidence of SpoVID self-interaction using
yeast two-hybrid assay.
Positive interaction between SpoIVA and SpoVID was
confirmed by retransformation assay (not shown) and by
measurement of b-galactosidase activity of interacting pro-
teins. The strength of interaction is expressed in Miller units
and in the system used may vary from weak to very strong
(Invitrogen, ProQuestTM Two-Hybrid System with Gate-
ways Technology instruction manual) and was also de-
scribed in the study of Krajcikova et al. (2009). When
compared with control strains carrying interacting pro-
teins, the protein–protein interaction between SpoIVA and
SpoVID is moderately strong (Table 2).
Whole sequence of SpoIVA is necessary forinteraction with SpoVID
To identify regions of SpoIVA protein that might be respon-
sible for its contact with SpoVID, two truncated mutant
versions were prepared. SpoIVA protein consists of 492
amino acid residues and the first SpoIVA mutant, herein
named SpA2, lacking 91 residues on its C terminus
(SpoIVA1–401), was fused with both DB and AD of GAL4
transcription factor and subjected to yeast two-hybrid
analysis with full-length SpoVID protein. The second trun-
cated mutant, SpA4, missing 102 N-terminal amino acids
(SpoIVA103–492), was also analyzed for direct interaction
with full-length SpoVID in S. cerevisiae MaV203 cells. None
of the truncated mutants of SpoIVA was able to interact
Fig. 1. Yeast two-hybrid analysis of protein–
protein interactions. (a) Saccharomyces cerevisiae
MaV203 cells bearing two fusion plasmids, one
with the first gene of interest fused to activation
domain (AD) of GAL4 transcription factor and
another with the second gene of interest fused to
the DNA-binding domain (DB) of GAL4, are able
to grow on the SC-Leu-Trp selection plate. Only
direct interaction between two proteins of
interest enables the yeast cells to grow on media
lacking uracil (SC-Leu-Trp-Ura), lacking histidine
and containing 100 mM 3AT (SC-Leu-Trp-His
1100 mM 3AT) or to give a blue color with
X-gal (see Materials and methods). (b) Mutual
interactions between full-length SpoIVA and
SpoVID, respectively, and their truncated forms:
SpA2–SpoIVA1–401; SpA4–SpoIVA103–492;
SpD2– SpoVID1–399; SpD3–SpoVID201–575.
Fusions with the activation domain (AD) are
placed horizontally, with the binding domain
(DB) vertically (a). (c) Positive control (PC) and
negative control (NC) on selection plates.
The previously identified interaction between
DB(CotE)/AD(CotE) (Krajcikova et al., 2009)
served as a positive control and pDBLeu/
pEXP-AD502 (GAL4 domains in expression
vectors) served as a negative control.
FEMS Microbiol Lett 299 (2009) 74–85 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
79Interactions of early spore coat proteins
directly with its partner or reconstitute an active GAL4 using
the yeast two-hybrid system (Fig. 1b).
An earlier study examined a set of random point
mutants of spoIVA and concluded that for localization and
coat assembly purposes, regions spread throughout the
whole sequence of SpoIVA are necessary (Catalano et al.,
2001). Therefore, it could also be assumed, as for the
interaction with SpoVID protein, that there is probably not
a specific domain created by adjacent residues of SpoIVA
that is responsible for this specific protein–protein contact.
Rather, a folding process brings together distant amino acids
recognizing interaction patterns on their partner. Another
explanation is that the truncated form of SpoIVA could be
unstable.
C-terminal region of SpoVID is crucial forinteraction with SpoIVA
Previous works (Ozin et al., 2000, 2001; Costa et al., 2006)
revealed that the last 50 residues of the C terminus of
SpoVID represent a cell wall-binding motif responsible for
targeting this protein to the cortex/coat interface. Costa
et al. (2006) also identified a region of 202 residues in the
N-terminal part of SpoVID necessary for interaction with
SafA. Here, the regions of SpoVID crucial for interaction
with SpoIVA were analyzed by creating two truncated
versions of SpoVID protein. First, the deletion mutant,
hereafter named SpD2 (SpoVID1–399), lacks 176 amino acids
from the C-terminal region. Secondly, the mutant, desig-
nated SpD3 (SpoVID201–575), lacks 200 N-terminal amino
acids. Both truncated forms of SpoVID were fused to the DB
and AD domains of GAL4, respectively, and were analyzed
for direct interaction with full-length SpoIVA in MaV203
yeast cells (Fig. 1b). The results from the yeast two-hybrid
experiments with truncated mutants showed that the first
200 amino acids of the N terminus of SpoVID are dispen-
sable for contact with full-length SpoIVA. Thus, some
uncharacterized amino acids in the C-terminal part of
SpoVID protein confer the ability to interact with SpoIVA.
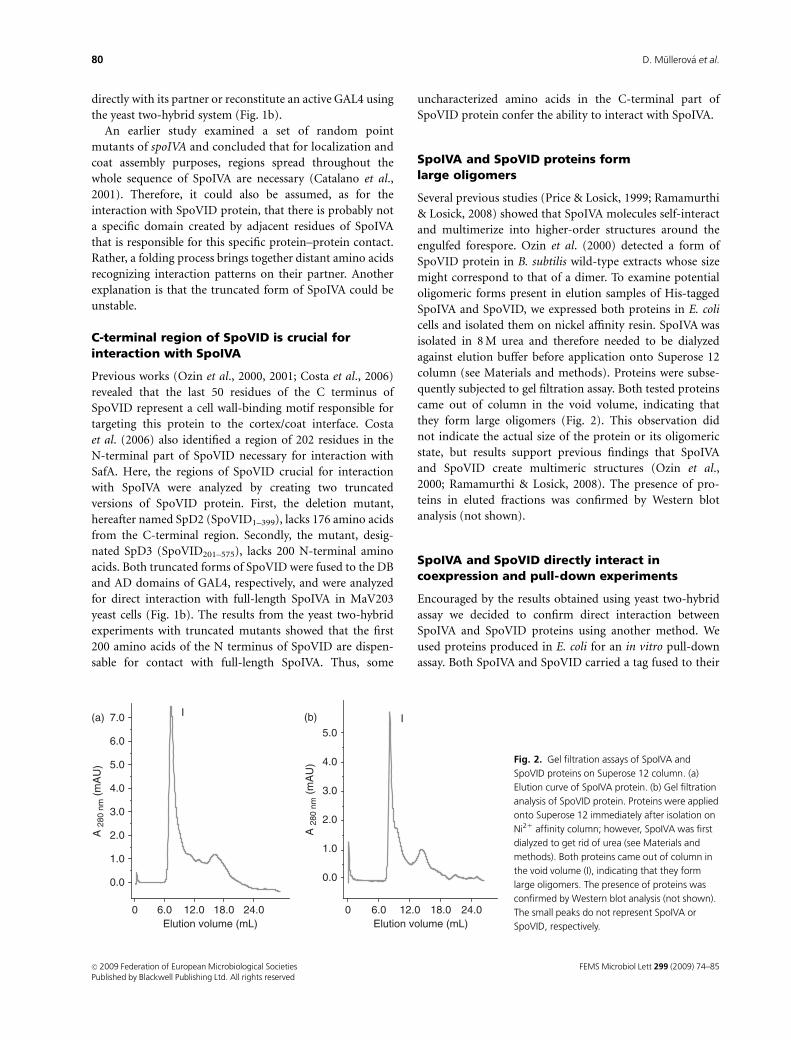
SpoIVA and SpoVID proteins formlarge oligomers
Several previous studies (Price & Losick, 1999; Ramamurthi
& Losick, 2008) showed that SpoIVA molecules self-interact
and multimerize into higher-order structures around the
engulfed forespore. Ozin et al. (2000) detected a form of
SpoVID protein in B. subtilis wild-type extracts whose size
might correspond to that of a dimer. To examine potential
oligomeric forms present in elution samples of His-tagged
SpoIVA and SpoVID, we expressed both proteins in E. coli
cells and isolated them on nickel affinity resin. SpoIVA was
isolated in 8 M urea and therefore needed to be dialyzed
against elution buffer before application onto Superose 12
column (see Materials and methods). Proteins were subse-
quently subjected to gel filtration assay. Both tested proteins
came out of column in the void volume, indicating that
they form large oligomers (Fig. 2). This observation did
not indicate the actual size of the protein or its oligomeric
state, but results support previous findings that SpoIVA
and SpoVID create multimeric structures (Ozin et al.,
2000; Ramamurthi & Losick, 2008). The presence of pro-
teins in eluted fractions was confirmed by Western blot
analysis (not shown).
SpoIVA and SpoVID directly interact incoexpression and pull-down experiments
Encouraged by the results obtained using yeast two-hybrid
assay we decided to confirm direct interaction between
SpoIVA and SpoVID proteins using another method. We
used proteins produced in E. coli for an in vitro pull-down
assay. Both SpoIVA and SpoVID carried a tag fused to their
0 6.0 12.0 18.0 24.0
0.0
1.0
2.0
3.0
4.0
5.0
(b)
Elution volume (mL)
I
0 6.0 12.0 18.0 24.0
0.0
1.0
2.0
3.0
4.0
5.0
6.0
7.0
A 2
80 n
m (
mA
U)
A 2
80 n
m (
mA
U)
(a)
Elution volume (mL)
I
Fig. 2. Gel filtration assays of SpoIVA and
SpoVID proteins on Superose 12 column. (a)
Elution curve of SpoIVA protein. (b) Gel filtration
analysis of SpoVID protein. Proteins were applied
onto Superose 12 immediately after isolation on
Ni21 affinity column; however, SpoIVA was first
dialyzed to get rid of urea (see Materials and
methods). Both proteins came out of column in
the void volume (I), indicating that they form
large oligomers. The presence of proteins was
confirmed by Western blot analysis (not shown).
The small peaks do not represent SpoIVA or
SpoVID, respectively.
FEMS Microbiol Lett 299 (2009) 74–85c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
80 D. Mullerova et al.

N termini: SpoIVA had a GST tag and SpoVID carried a His
tag critical for its purification (see Materials and methods).
The GST-SpoIVA, immobilized on glutathione affinity ma-
trix, was intended to capture His-tagged SpoVID from the
solution incubated on glutathione Sepharose beads. After
several washes, the presence of retained proteins was
detected by Western blot using antibodies against SpoIVA
and SpoVID, respectively, as well as anti-His-tag antibody
(not shown). His-SpoVID was not pulled down by GST-
SpoIVA on glutathione beads. Only GST-SpoIVA fusion
protein was detected, which migrated at approximately
80 kDa (not shown). There are several possible explanations
for why we failed to detect a direct interaction between
SpoIVA and SpoVID using GST pull-down. One explanation
consists in the folding process of fusion proteins GST and
SpoIVA. GST is a relatively large tag (26 kDa) and it might
have disrupted folding of SpoIVA and hampered constitution
of the correct conformation of SpoIVA. Another reason could
be that GST tag shields the site of interaction on SpoIVA
protein sequence, causing a steric barrier. Lastly, either one or
both interacting proteins may undergo a multimerization
process when isolated separately which prevents the sites of
interaction from coming into contact.
To overcome these obstacles, we employed coexpression
of the proteins of interest. Coexpression plasmids are
designed for simultaneous expression of two or more genes,
allowing proteins to interact immediately after the transla-
tion. The gene coding for SpoVID protein was cloned in-
frame with N-terminal His tag, while the second protein,
SpoIVA, carried no tag. After the expression in E. coli, a
pull-down experiment on the Ni21 affinity column and
Western blot, His-tagged SpoVID clearly pulled down
untagged SpoIVA protein (Fig. 3a, lanes 5–8). SpoIVA
protein without His tag bound only slightly to the nickel
affinity matrix (Fig. 3a, lane 2) and was eluted by up to
0.2 M imidazole.
It may be concluded that the direct interaction between
SpoIVA and SpoVID proteins identified by the yeast two-
hybrid system was confirmed by coexpression and subsequent
pull-down experiment.
SpoIVA0.1 0.2 0.3 1.0
SpoIVA+SafA-His0.1 0.2 0.3 1.0 0.1 0.2 0.3 1.0
SpoIVA+SpoVID+SafA-His
5572
(a)
1 2 3 4 5 6 7 8 9 10 11 12
SpoVID SpoVID+SafA-His SpoIVA+SpoVID+SafA-His
95130
1 2 3 4 5 6 7 8 9 10 11 12
(b)
1 2 3 4 5 6 7 8 9 10 11 12
SpoVID+SafA-His SpoIVA+SafA-His SpoIVA+SpoVID+SafA-His0.1 0.2 0.3 1.0
0.1 0.2 0.3 1.0 0.1 0.2 0.3 1.0 0.1 0.2 0.3 1.0
0.1 0.2 0.3 1.0 0.1 0.2 0.3 1.0
55
(c)
130
95
SpoVID-His SpoIVA+SpoVID-His
1 2 3 4 5 6 7 8
(d)
72
55
SpoIVA0.1 0.2 0.3 1.0 0.1 0.2 0.3 1.0
0.1 0.2 0.3 1.0 0.1 0.2 0.3 1.0
SpoIVA+SpoVID-His
1 2 3 4 5 6 7 8
(e)
SpoIVA
SpoVID
SpoVID
SpoIVA
SafA
Fig. 3. Western blots of pull-down analyses
of coexpressed SpoVID, SpoIVA and SafA in
Escherichia coli. The pull-down assays were
performed on Ni21 column: SpoVID was
His-tagged in (a, b) and SafA in (c–e). Proteins
were probed with polyclonal anti SpoIVA
antibody (a, c), polyclonal anti SpoVID antibody
(d) and monoclonal anti-His-tag antibody (b, e).
Above each lane is the concentration of imidazole
(mol L�1) used to elute proteins from the nickel
column. Small amounts of untagged proteins
(a, c, d – lanes 1–4) are retained slightly on Ni21
column, eluted already by 0.1 M (d – lane 1) or
0.2 M (a, c – lane 2) concentrations of imidazole.
On the other hand, untagged SpoIVA and SpoVID
are present in the sample eluted with 0.3 M
imidazole in the presence of His-SafA (c, d – lanes
9–12), indicating that they are pulled-down by
His-SafA protein. In the same way, SpoIVA is
pulled-down by His-SpoVID (a – lanes 5–8).
(b, e) Elution profiles of His-tagged proteins:
SpoVID (b) and SafA (e). Molecular masses (in
kDa) are indicated on the right side of each panel.
FEMS Microbiol Lett 299 (2009) 74–85 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
81Interactions of early spore coat proteins

SpoIVA as well as SpoVID interact directly withSafA in coexpression and triple pull-down assay
Several previous studies (Ozin et al., 2000, 2001; Costa et al.,
2006) showed that there is a direct contact between two
morphogenetic proteins, SpoVID and SafA. Their mutual
association was observed by coimmunoprecipitation (Ozin
et al., 2000) and the direct contact was confirmed by the
yeast two-hybrid system and in vitro pull-down assay (Ozin
et al., 2001). The same work pointed out the dependency of
SafA on SpoIVA in targeting the forespore. Thus we decided
to include the third morphogenetic protein, SafA, in our
coexpression experiments.
Escherichia coli cells were cotransformed with compatible
vectors pETDuet-1, carrying spoVID and 50 end His-tagged
safA, and with pACYCDuet-1, carrying untagged spoIVA
gene. We also transformed single pETDuet-1 carrying His-
SafA and SpoVID, to confirm their mutual interaction, as
well as a combination of pETDuet(his-safA) and pACYC
(spoIVA). Expression of proteins was followed by pull-down
assay on nickel-conjugated resin and the presence of pro-
teins of interest was probed by Western blot using anti-
His-tag, anti-SpoIVA and anti-SpoVID antibodies (see
Materials and methods). Immunodetection confirmed the
direct interaction between SpoVID and SafA proteins and
revealed a novel, albeit anticipated (Ozin et al., 2001), direct
interaction between SafA and SpoIVA. His-SafA pulled
down both SpoVID and SpoIVA independently (Fig. 3c, d,
lanes 5–8) but also when both were coexpressed in E. coli
(Fig. 3c, d, lanes 9–12). Proteins with no His tag were eluted
early from the column, at 0.1 M concentration of imidazole
(Fig. 3c, d, lane 1). Elution profiles of His-tagged SafA
expressed with either one or both untagged proteins are
shown in Fig. 3e. As shown in Fig. 3c (lanes 1–3) and Fig. 3d
(lanes 1–3), untagged SpoVID and SpoIVA apparently
possess some low innate affinity to nickel resin, as they
remained attached to the column even after several washes.
Although the same amounts of cell extract were used in all
cases, the concentrations of SpoVID and SpoIVA in these
samples were higher than when two or three proteins were
coexpressed. Key to this observation is that the level of
expression of single protein from pETDuet-1 vector is
higher than the level of expression of two or more proteins
(Novagen, Duet vectors manual). Therefore, we would
expect to detect no signals in lanes 1–3 (Fig. 3c, d) if equal
amounts of proteins are loaded onto the column, as for pull-
down samples of a mixture of interacting proteins. All the
results presented here strongly indicate the existence of
mutual interactions between SpoVID, SpoIVA and SafA.
Discussion
Bacillus subtilis spore coat is built up of 4 70 protein
components (Henriques et al., 2004; Kim et al., 2006). Their
mutual direct or indirect interactions lead to formation of
an intricate composite structure whose properties enable
the spore to endure various assaults. From this protein
network, only a small subset of components play a morpho-
genetic role (Driks, 1999; Henriques & Moran, 2000;
McPherson et al., 2005). In this study, we demonstrated that
two morphogenetic spore coat proteins, SpoIVA and
SpoVID, interact directly. For this interaction to occur, the
C-terminal portion of SpoVID and full-length SpoIVA
protein are crucial. We also showed that both SpoIVA and
SpoVID proteins interact with SafA either independently or
simultaneously.
Yeast two-hybrid experiments revealed a positive interac-
tion when full-length SpoIVA was fused to the GAL4
activation domain and full-length SpoVID to the DNA-
binding domain but not vice versa, which is not a rare
observation (Seyler et al., 1997). In the search for domains
of each protein, responsible for their direct interaction,
N-terminal and C-terminal truncated mutants of SpoVID
and SpoIVA were prepared. No truncated mutant of SpoIVA
protein interacted with full-length SpoVID (Fig. 1b), in-
dicating that full-length protein is necessary for direct
contact with SpoVID. On the other hand, 200 residues
of the N-terminal part of SpoVID are dispensable for
interaction with full-length SpoIVA, implying that amino
acids mediating contact with SpoIVA are present in the
C-terminal part of SpoVID.
Previous research (Ozin et al., 2001; Costa et al., 2006)
focused on searching for direct interactions between pro-
teins used GST as a tag in pull-down experiments. Using this
method, these studies confirmed direct interaction between
two morphogenetic proteins, SpoVID and SafA, initially
observed by the yeast two-hybrid system. Similarly, here it
was necessary to confirm the direct interaction between the
proteins of interest, SpoIVA and SpoVID, by the in vitro
pull-down assay. Unfortunately, experiments using GST as a
tag were not successful, GST-SpoIVA failed to pull-down
His-SpoVID. As SpoIVA is an insoluble protein, we believed
that GST tag might enhance its solubility; instead, it
probably prevented SpoIVA from contacting SpoVID. Like-
wise, in the yeast two-hybrid system, SpoVID and SpoIVA
interacted only when fused to a particular domain of GAL4;
it might be possible that GST is not a suitable fusion partner
for SpoIVA and that SpoVID, as in the studies of Ozin et al.
(2001) and Costa et al. (2006), would in fusion with GST,
occupy the correct conformation.
Ozin et al. (2000) detected two forms of SpoVID pro-
tein in B. subtilis wild-type extracts: the 66 and 120 kDa
forms. The predicted weight of the protein monomer is
65 kDa. Our heterologous proteins expressed in E. coli
migrated on Western blots at approximately a mass which
may correspond to a dimer. Results obtained from gel
filtration assay indicate that SpoVID protein may undergo
FEMS Microbiol Lett 299 (2009) 74–85c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
82 D. Mullerova et al.

a multimerization process. The same result was observed
with SpoIVA protein. The denaturing conditions of SDS-
PAGE gel break down the oligomeric structures of SpoVID
and SpoIVA in such a way that they migrate as smaller
species. The predicted weight of SpoIVA is 55 kDa. How-
ever, previous and present results suggest that SpoVID and
SpoIVA proteins may be present in several forms within
the cell. Is the oligomerization process of these proteins
simply the consequence of spore maturation or do their
different forms serve distinct functions? The model of
SpoIVA assembly proposed by Ramamurthi & Losick
(2008) shows that individual SpoIVA molecules are teth-
ered to forespore surface by SpoVM (Price & Losick, 1999)
and oligomerization occurs subsequently upon hydrolysis
of ATP and is an irreversible process. This finding indicates
that only monomeric molecules are capable of recognition
of the assembly site and hence are essential for assembly
initiation. In view of this polymerization evidence, it is
surprising that our yeast two-hybrid screen did not reveal
self-interaction of either SpoIVA or SpoVID. On the other
hand and in agreement with results presented here, Ozin
et al. (2001) also found no evidence of interaction between
SpoVID molecules using the yeast two-hybrid system. It
may be that one limitation of this system is that it is not
able to reveal all existing interactions between tested
proteins. One of them is a steric barrier when protein
is fused to a particular domain of GAL4 transcription
factor. Another limitation may consist in incorrect fold-
ing and instability of such a protein. The last possible
explanation is impaired targeting of heterologous fusion
proteins into the yeast nucleus. Similar problems were
encountered when testing SpoVID–SafA and SpoIVA–SafA
interactions by the yeast two-hybrid system. A direct
interaction of SpoVID and SafA has already been observed
using the yeast two-hybrid system (Ozin et al., 2001), but
in that work they used a different two-hybrid system
(Clontech, Matchmaker) which utilizes high copy vectors
and is therefore more sensitive. Further experiments and/
or other methods need to be employed to elucidate this
obvious discrepancy.
The coexpression experiment followed by pull-down
assay confirmed the direct interaction between SpoVID and
SpoIVA observed by the yeast two-hybrid system and
revealed a potentially novel interaction between SpoIVA
and SafA. Pull-down assay also confirmed a previously
reported direct interaction between SpoVID and SafA
(Costa et al., 2006). Untagged proteins SpoVID and SpoIVA,
in the presence of His-SafA, were retained on nickel affinity
resin at higher concentrations of imidazole than in the
absence of His-SafA. Moreover, it seems that SpoIVA binds
more strongly to SpoVID–SafA complex than to SafA
protein itself, because a higher concentration of imidazole
is needed to elute SpoIVA from the affinity column, and
more protein is eluted.
Fig. 4. Model of the assembly of early spore coat proteins. SpoIVA, SpoVID and SafA are targeted to the OFM. SpoVID and SafA both contain a LysM
domain with peptidoglycan-binding properties. SpoIVA is tethered to OFM via the small amphipathic peptide SpoVM. In the SpoVID protein sequence,
there are two known regions of interaction (depicted as thick-line rectangles): the N-terminal part of SpoVID, comprising the first 202 amino acid
residues, is responsible for contact with regions A and B of SafA (details in text); the C-terminal part between residues 400 and 575 (thick-line rectangle)
contacts unspecified regions of SpoIVA. Walker A box present in the N-terminal part of SpoIVA is responsible for ATP hydrolysis and multimerization of
SpoIVA protein. See text for more details. Solid arrows indicate direct interactions between proteins: full arrowheads mark known regions of interaction;
open arrowheads label putative sites of interaction. Dotted arrows depict mutual dependencies of various proteins.
FEMS Microbiol Lett 299 (2009) 74–85 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
83Interactions of early spore coat proteins

Based on the latest findings (Costa et al., 2006; Kim et al.,
2006; Henriques & Moran, 2007; Ramamurthi & Losick,
2008) and on results presented here, we propose the follow-
ing model of early spore coat protein assembly (Fig. 4). First,
SpoIVA is recruited via its C terminus to the outer forespore
membrane (OFM) by a small amphipathic peptide called
SpoVM (Price & Losick, 1999). SpoIVA then recruits
SpoVID, whose LysM domain facilitates its deployment to
cortex peptidoglycan beneath the OFM. Results obtained
using yeast two-hybrid assay imply that for this interaction
to occur, the C-terminal part of SpoVID and the whole
sequence of SpoIVA are necessary. In the next step, SafA is
firstly targeted to OFM in a SpoVID-independent event
which probably depends on its LysM domain, and, secondly,
SafA encases the spore upon the direction of SpoVID (Ozin
et al., 2001; Costa et al., 2006). Essential for the interaction
between SafA and SpoVID are regions A (residues 51–63)
and B (PYYH motif, residues 203–206) of SafA and the
N-terminal part of SpoVID (Costa et al., 2006). SafA is
supposed to have an extended conformation ranging from
cortex to outer coat and binding outer coat proteins with its
C terminus (Ozin et al., 2000). However, direct interaction
between SafA and CotE has not been observed yet. SafA
deletion results in loss of several uncharacterized proteins
and also CotG, which is an outer coat component (Taka-
matsu et al., 1999; Kim et al., 2006). From the results
mentioned above and from previous work it may be
suggested that SpoIVA and SpoVID form a complex before
SafA localization to the forespore. Based on the pull-down
results it can be concluded that interaction between SafA and
SpoVID is stronger than that between SafA and SpoIVA. Less
SpoIVA is eluted from the column in comparison with the
amount of SpoVID in the cases when these were separately
coexpressed and pulled down by SafA. It is therefore reason-
able to assume that SpoVID tethers SafA to SpoVID–SpoIVA
complex and the low affinity interaction between SafA and
SpoIVA takes place afterwards. The precise regions of each
protein involved in this interaction still remain to be resolved.
Further studies will reveal more details about how proteins
contact each other and how they polymerize into supramole-
cular structures. This work contributes to the elucidation of
the partial mechanism that enables creation of such an
intricate and endurable structure like the B. subtilis spore coat.
Acknowledgements
The authors gratefully acknowledge Dr Simon Cutting for
preparation of antibodies against SpoIVA and SpoVID
proteins. The work in the author’s laboratory is supported
by grant NMP4-CT-2004-013523 from the EU 6th FP, grant
2/7007/27 from the Slovak Academy of Sciences and grants
from the Slovak Research and Development Agency under
contract No. LPP-0218-06, No. ESF-EC-0106.
References
Backman K, Ptashne M & Gilbert AW (1976) Construction of
plasmids carrying the cI gene of bacteriophage lambda. P Natl
Acad Sci USA 73: 4174–4178.
Beall B, Driks A, Losick R & Moran CP Jr (1993) Cloning and
characterization of a gene required for assembly of the Bacillus
subtilis spore coat. J Bacteriol 175: 1705–1716.
Catalano FA, Meador-Parton J, Popham DL & Driks A (2001)
Amino acids in the Bacillus subtilis morphogenetic protein
SpoIVA with roles in spore coat and cortex formation.
J Bacteriol 183: 1645–1654.
Coote JG (1972) Sporulation in Bacillus subtilis. Characterization
of oligosporogenous mutants and comparison of their
phenotypes with those of asporogenous mutants. J Gen
Microbiol 71: 1–15.
Costa T, Isidro AL, Moran CP Jr & Henriques AO (2006)
Interaction between coat morphogenetic proteins SafA and
SpoVID. J Bacteriol 188: 7731–7741.
Driks A (1999) Bacillus subtilis spore coat. Microbiol Mol Biol R
63: 1–20.
Driks A (2002) Proteins of the spore core and coat. Bacillus
subtilis and its Closest Relatives (Sonenshein AL, Hoch JA &
Losick R, eds), pp. 527–536. American Society for
Microbiology, Washington, DC.
Driks A & Setlow P (2000) Morphogenesis and properties of the
bacterial spore. Prokaryotic Development (Brun YV & Shimkets
LJ, eds), pp. 191–218. American Society for Microbiology,
Washington, DC.
Driks A, Roels S, Beall B, Moran CP Jr & Losick R (1994)
Subcellular localization of proteins involved in the assembly of
the spore coat of Bacillus subtilis. Gene Dev 8: 234–244.
Gietz RD & Woods RA (2002) Transformation of yeast by the
LiAc/SS carrier DNA/PEG method. Method Enzymol 350:
87–96.
Henriques AO & Moran CP Jr (2000) Structure and assembly of
the bacterial endospore coat. Methods 20: 95–110.
Henriques AO & Moran CP Jr (2007) Structure, assembly and
function of the spore surface layers. Annu Rev Microbiol 61:
555–588.
Henriques AO, Costa TV, Martins LO & Zilhao R (2004) The
functional architecture and assembly of the coat. Bacterial
Spore Formers: Probiotics and Emerging Applications (Ricca RE,
Henriques AO & Cutting SM, eds), pp. 65–86. Horizon
Biosciences, Norfolk.
Kim H, Hahn M, Grabowski P, McPherson DC, Otte MM, Wang
R, Ferguson CC, Eichenberger P & Driks A (2006) The Bacillus
subtilis spore coat protein interaction network. Mol Microbiol
59: 487–502.
Kodama T, Takamatsu H, Asai K, Kobayashi K, Ogasawara N &
Watabe K (1999) The Bacillus subtilis yaaH gene is transcribed
by SigE RNA polymerase during sporulation, and its product
is involved in germination of spores. J Bacteriol 181:
4584–4591.
FEMS Microbiol Lett 299 (2009) 74–85c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
84 D. Mullerova et al.

Krajcikova D, Lukacova M, Mullerova D, Cutting SM & Barak I
(2009) Searching for protein–protein interactions within the
Bacillus subtilis spore coat. J Bacteriol 191: 3212–3219.
Levin PA, Fan N, Ricca E, Driks A, Losick R & Cutting S (1993)
An unusually small gene required for sporulation by Bacillus
subtilis. Mol Microbiol 9: 761–771.
McPherson DC, Kim H, Hahn M, Wang R, Grabowski P,
Eichenberger P & Driks A (2005) Characterization of the
Bacillus subtilis spore morphogenetic coat protein CotO.
J Bacteriol 187: 8278–8290.
Nicholson WL, Munakata N, Horneck G, Melosh HJ & Setlow P
(2000) Resistance of Bacillus endospores to extreme terrestrial
and extraterrestrial environments. Microbiol Mol Biol R 64:
548–572.
Ozin AJ, Henriques AO, Yi H & Moran CP Jr (2000) Morpho-
genetic proteins SpoVID and SafA form a complex during
assembly of the Bacillus subtilis spore coat. J Bacteriol 182:
1828–1833.
Ozin AJ, Samford CS, Henriques AO & Moran CP Jr (2001)
SpoVID guides SafA to the spore coat in Bacillus subtilis.
J Bacteriol 183: 3041–3049.
Piggot PJ & Coote JG (1976) Genetic aspects of bacterial
endospore formation. Bacteriol Rev 40: 908–962.
Pogliano K, Harry E & Losick R (1995) Visualization of the
subcellular location of sporulation proteins in Bacillus subtilis
using immunofluorescence microscopy. Mol Microbiol 18:
459–470.
Price KD & Losick R (1999) A four-dimensional view of assembly
of a morphogenetic protein during sporulation in Bacillus
subtilis. J Bacteriol 181: 781–790.
Ramamurthi KS & Losick R (2008) ATP-driven self-assembly of a
morphogenetic protein in Bacillus subtilis. Mol Cell 31: 406–414.
Roels S, Driks A & Losick R (1992) Characterization of spoIVA, a
sporulation gene involved in coat morphogenesis in Bacillus
subtilis. J Bacteriol 174: 575–585.
Sambrook J, Fritsch EF & Maniatis T (1989) Molecular Cloning: A
Laboratory Manual. Cold Spring Harbor Laboratory Press,
Cold Spring Harbor, NY.
Setlow P (2000) Resistance of bacterial spores. Bacterial
Stress Responses (Storz G & Hengge-Aronis R, eds),
pp. 217–230. American Society for Microbiology,
Washington, DC.
Setlow P (2003) Spore germination. Curr Opin Microbiol 6:
550–556.
Seyler RW Jr, Henriques AO, Ozin AJ & Moran CP Jr (1997)
Assembly and interactions of cotJ-encoded proteins,
constituents of the inner layers of the Bacillus subtilis spore
coat. Mol Microbiol 25: 955–966.
Takamatsu H, Kodama T, Nakayama T & Watabe K (1999)
Characterization of the yrbA gene of Bacillus subtilis, involved
in resistance and germination of spores. J Bacteriol 181:
4986–4994.
Vidal M (1997) The reverse two-hybrid system. The Two-hybrid
System (Bartel P & Fields S, eds), pp. 109–147. Oxford
University Press, New York.
Youngman P, Perkins JB & Losick R (1984) Construction of a
cloning site near one end of Tn917 into which foreign DNA
may be inserted without affecting transposition in Bacillus
subtilis or expression of the transposon-borne erm gene.
Plasmid 12: 1–9.
Zheng L, Donovan WP, Fitz-James PC & Losick R (1988) Gene
encoding a morphogenic protein required in the assembly of
the outer coat of the Bacillus subtilis endospore. Gene Dev 2:
1047–1054.
FEMS Microbiol Lett 299 (2009) 74–85 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
85Interactions of early spore coat proteins