interaction of lectins from soybean and peanut with rhizobia that nodulate soybean, peanut, or both...
TRANSCRIPT

Interaction of lectins from soybean and peanut with rhizobia that nodulate soybean, peanut, or both plants1
STEVEN G. PUEPPKE Departmer~t of Plant Pathology, University of Florida, Gainesuille, FL, U.S.A. 32611
AND
TOM G. FREUND, BRENT C. SCHULZ, A N D HARVEY P. FRIEDMAN Departrnertt of Biology, University of Missouri, 8001 Nat~tral Bridge Road, St. Louis, MO, U.S.A. 63121
Accepted September 17, 1980
PUEPPKE, S. G., T. G. FREUND, B. C. SCHULZ, and H. P. FRIEDMAN. 1980. Interactionof lectins from soybean and peanut with rhizobia that nodulate soybean, peanut, or both plants. Can. J . Microbiol. 26: 1489-1497.
Four of 14 strains of Rhizobi~on japonicum from soybean nodulated peanut (Aracltis hypogaea L. cultivar Jumbo Virginia), and 3 of 8 Rhizobiltm sp. strains from peanut nodulated soybean (Glycine rnax (L.) Merr. cultivar Harosoy 63). Cells of three peanut rhizobia bound fluorescent- and radioisotope-labeled soybean lectin. Two of these strains failed to nodulate soybean, and conversely, two peanut strains that nodulated soybean did not bind to soybean lectin. Both culture medium and age had pronounced effects of the number of peanut rhizobia cells that bound fluorescent-labeled soybean lectin. Harosoy 63 soybean root exudates stimulated the growth of peanut rhizobia, but had no consistent influence on the number of cells that bound soybean lectin. Although extracellular soybean lectin receptors were present in culture fluids from each of the peanut rhizobia whose cells bound the lectin, the titer of receptors was greatest for strain 3G4b5. The affinity constants for the adherence of soybean lectin to Rhizobiurn sp. 3G4b5 cells from cultures of various ages ranged from 4.2 x 106 to 4.9 x 106 M-', and the number of lectin binding sites per cell decreased as cells aged. Cells of the soybean and peanut rhizobia did not bind fluorescent- or radioisotope-labeled peanut lectin. The results indicate that there is no relation- ship between the ability of peanut and soybean rhizobia to nodulate the reciprocal host plant and their ability to bind to the lectin of that plant.
PUEPPKE, S. G., T. G. FREUND, B. C. SCHULZ et H. P. FRIEDMAN. 1980. Interaction of lectins from soybean and peanut with rhizobia that nodulate soybean, peanut, or bothplants. Can. J. Microbiol. 26: 1489-1497.
De 14 souches de Rhizobiurn japonicum extraites de racines de soya, 4 d'entre elles ont cause la nodulation de racines d'arachide (Arachis hypogaea L. cultivar Jumbo Virginia) et, de 8 souches de Rhizobiurn sp. provenant de racines d'arachide, 3 d'entre elles ont cause la nodulation de racines de soya (Glycitle max (L.) Men. cultivar Harasoy 63). Les cellules de trois rhizobiums de plants d'arachide se sont attache des lectines de soya marquees de radio-isotope et de fluores- cence. Deux de ces souches n'ont pas cause de nodulation chez le soya et, inversement, deux souches provenant de racines d'arachide qui ont cause de la nodulation chez le soya ne se sont pas attache de lectines de soya. Le milieu de culture et 1'8ge ont tous deux des effets prononces sur le nombre de cellules de rhizobium qui s'attachent des lectines de soya marquees de fluorescence. Des exsudats de racines de soya Harasoy 63 ont stimule la croissance de rhizobium d'arachide, mais ils n'ont pas influence de f a ~ o n consistante le nombre de cellules qui s'attachent des lectines de soya. Bien que des recepteurs extracellulaires de lectines de soya aient ete presents dans les liquides culturaux de chacun des rhizobiums d'arachides dont les cellules s'attachent les lectines, le titre le plus eleve des recepteurs fut celui de la souche 3G4b5. Les constantes d'affinite pour I'adherence des lectines de soya aux cellules du Rhizobiurn sp. 3G4b5 dans des cultures d'gges varies se situent entre 4,2 x lo6 a 4,9 x lo6 M-' , et le nombrede sites d'attachement de lectines par cellule decroit avec I'avancement en ige des cellules. Les cellules des rhizobiums de soya et d'arachide ne se sont pas attache de lectines d'arachide marquees de fluorescence ou de radio- isotope. Les resultats indiquent qu'il n'y a pas de relation entre la capacite des rhizobiums de soya et d'arachide de causer la nodulation des plantes h6tes reciproques et leur capacite de s'attacher les lectines de ces plantes.
[Traduit par lejournal]
Introduction has not been established. There is, however, a The biochemical basis for the well-known selec- body of data suggesting that plant lectins may have
tivity exhibited by legume-Rhizobium associations some function in the recognition process . -
IFlirida Agricultural Experiment Station Journal Series 1978; Schmidt 1979; and references Paper NO. 241 1. therein). Whereas some correlative studies indicate
0008-4166/80/121489-09$01.00/0 a1980 National Research Council of Canada/Conseil national de recherches du Canada
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MC
GIL
L U
NIV
ER
SIT
Y o
n 11
/20/
14Fo
r pe
rson
al u
se o
nly.

1490 C A N . J . MICROBIOL. VOL. 26, 1980
a role for the 120000 molecular weight soybean lectin (SBL) in symbiont recognition by soybean (Bhuvaneswari and Bauer 1978; Bhuvaneswari et al. 1977; Bohlool and Schmidt 1974), there is gene- tic evidence that SBL does not function in the soy- bean - Rhizobium japonicutn interaction (Pull et al. 1978; Su et al. 1980). Perhaps the most persua- sive evidence favoring SBL involvement is the ob- servation that SBL selectively interacts with rhizobia that nodulate soybean. Thus the lectin binds to over 60% of the symbiotic R. japonicum strains tested, but to only about a quarter of the tested nonsymbiont strains from other species of Rhizobium (Bhuvaneswari et al. 1977; Bohlool and Schmidt 1974; Law and Strijdom 1977). The SBL receptors of symbiont rhizobia are extracellular or capsular polysaccharides and are apparently pres- ent only at certain times during rhizobial growth in culture (Bal et al. 1978; Bhuvaneswari and Bauer 1978; Bhuvaneswari et al. 1977; Calvert et al. 1978; Shantharam et al. 1980).
Certain strains of rhizobia belonging to the soy- bean cross-inoculation group (R. japonicum) can nodulate plants such as cowpea and peanut, whose homologous symbionts are normally classified in the "cowpea miscellany," Rhizobium sp.; the re- ciprocal nodulation of soybean by various strains of Rhizobium sp. has also been observed (Carroll 1934; Leonard 1923; Sears and Carroll 1927; Walker and Brown 1935; Wilson 1944). Unfortu- nately, in recent years scant attention has been paid to such heterologous nodulation, even though studies of this phenomenon may lead to an en- hanced understanding of the basis of legume-Rhizobium interactions. Here we report the results of binding studies using seed lectins from soybean and peanut and rhizobia from the reciprocal host. Our objectives were (i) to deter- mine if strains that nodulate the heterologous host also interact with the heterologous lectin, and (ii) to compare the intereaction of SBL with peanut rhizobia that nodulate soybean and peanut rhizobia that do not nodulate soybean.
Materials and methods Mainterlance of orgunisms
The rhizobial strains used in this study are listed in Tables I and 2. With the exception of Rhizobium sp. 3G4b19 and R. juponicum 61A93, which were grown on yeast extract mannitol medium (Vincent 1970), all strains were maintained on slants of gluconate mannitol medium (Bhuvaneswari et al. 1977). The organisms were stored at 4'C and were routinely transferred every 2 months.
Nodulatiorl The ability of Rhizobium strains to nodulate soybean (Glycine
max (L.) Merr. cultivar Harosoy 63, from Dr..T. Hymowitz, Department of Agronomy, University of Illinois, Urbana, IL)
was assessed as described previously (Su et 01. 1980). Nodula- tion studies with peanut (Arnchis hypogaea L. cultivar Jumbo Virginia, from W. Atlee Burpee, Riverside, CA) were performed in the same way, except that the plants were grown in 15-cm pots of sand. The peanuts were maintained in a growth chamber under a 12-h photoperiod (ca. 25001~ from a fluorescent source) at 28°C day and 25°C night temperatures, and nodulation was evaluated 28 days after inoculation.
Preparntiorl and lubelir~g of lectiris SBL from the soybean cultivar Disoy was purified to
homogeneity by two cycles ofaffinity chromatography using the procedure of Bhuvaneswari et al. (1977). Peanut lectin (PNL) from the peanut cultivar Jumbo Virginia was prepared similarly using the appropriate affinity adsorbent (Pueppke 1979~). The PNL preparation was homogeneous and gave the expected seven isolectin bands upon isoelectric focusing (Pueppke 19790). Both lectins were fluorescent labeled with fluorescein isothiocyanate (FITC, from Sigma Chemical Co., St. Louis, MO) according to Udenfriend (1962). For the FITC-labelled SBL (FITC-SBL), the ratio of absorbance at 493 to 280 nm was 0.25, and the protein concentration, as determined by the method of Lowry et 01. (1951) with bovine serum albumin as the standard, was 320 pg/mL. For the FITC-labeled PNL (FITC- PNL), the absorbance ratio was 0.62, and the protein concen- tration was 605 pg/mL.
SBL (30 mg) was radioisotope labeled with 25 mCi (I mCi = 37 GBq) of sodium [3H]borohydride (509 mCi/mmol, from Amersham Corp., Arlington Heights, IL) according to Lotan, Debray et al. (1975). PNL (12.5 mg) was labeled with 1.OmCi of Na[lZ5I] (ICN Radiochemicals, Irvine, CA) usingthechloramine Tmethod(Selaetal.1971). Bothlectinswererepurifiedby affinity chromatography before use, and thespecific activity ofeach was reduced as necessary by adding unlabeled lectin.
Binding experinients withfluorescent-labeled lectiris Cells were prepared for the initial screening studies by adding
5 x lo8 cells (determined turbidimetrically) from a slant culture to 50mL of gluconate mannitol broth (Bhuvaneswari et 01. 1977) in a 250-mL Erlenmeyer flask. The flasks were incubated at 29 f 1°C on a gyratory shaker operating at 125 rpm. When the cultures reached mid-log phase, they were centrifuged at 7700 x g for IOmin at 4°C. The cells were resuspended in phosphate-buffered saline (PBS: 0.43g KH,PO,, 1.48g Na,- HPO,, 7.2g NaCI/L distilled water, pH 7.2), and the cell con- centrations adjusted turbidimetrically to no more than lo8 cells/mL. Ten microlitres of cells was placed on a clean micro- scope slide and 5 pL of FITC-SBL or FITC-PNL, an amount previously determined to be saturating, was added. After a brief mixing period a cover slip was placed on the slide, sealed with Flo-Texx (Lerner Laboratories, Stamford, CT), and the slide examined at 400x with a Zeiss standard research microscope equipped with incident fluorescent illumination (BG-12 primary filter, FT-460 chromatic beam splitter, LP-478 barrierfilter). As a control for biochemical specificity of lectin adherence, prep- arations in which lectins bound to cells were also made up in the presence of 100 mM D-galactose (Pereira et al. 1974, 1976).
In later experiments the binding of fluorescent-labeled lectins to cells of Rhizobium sp. strains 3G4b4,3G4b5, and 3G4b 16 was examined as a function of culture age. Liquid cultures were initiated as described above using two different growth media. The first was gluconate mannitol broth, and the second was glutamate medium, which contained (per litre of distilled water) 1.1 g sodium glutamate, 2.2g KH,PO,, 0.5g (NH,),SO,, 0. I g MgSO,, 0.8g K,HP04, 5.0g sodium glucpnate, 5.0g mann~tol, 0.5 l g biotin, I mg nicotinic acid, I0 mg thiamine HCI, I mg pyridoxine HCI, and 100mg myoinositol. Over a 7-day period 5 mL of cell suspension was removed daily from a single flask of
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MC
GIL
L U
NIV
ER
SIT
Y o
n 11
/20/
14Fo
r pe
rson
al u
se o
nly.

PUEPPKE ET AL. 1491
each strain in each medium. For each sample of culture, cell number per millilitre was determined turbidimetrically, and the cells were prepared for microscopy by centrifugation and sus- pension in PBS. For each cell preparation a total of l00cells was examined in each of four randomly selected areas of the slide. Cells were counted using phase-contrast optics in conjunction with transmitted visible illumination, and the percentage of cells that fluoresced was determined by periodically replacing the visible light with incident fluorescent illumination. Controls for biochemical specificity were run as described above.
A third set of experiments was designed to determine the effect on lectin binding to cells of adding soybean root exudates to culture media. Root exudates were obtained from hy- droponically grown Harosoy 63 soybeans according to Bhuvaneswari and Bauer (1978), and neutral sugar was deter- mined by the phenol - sulfuric acid method (Kabat 1961). The exudates were filter sterilized using Uni-Pore polycarbonate membranes (0.4-pm pore size, from Amicon Corp., Lexington, MA) and stored at 4°C. The final media were prepared according to Bhuvaneswari and Bauer (1978) by dispensing 1.8mL of exudate into each of a series of 12 cm x 2cm screw-capped tubes and then adding 0.2 mL of 10 times concentrated, filter-sterilized gluconate mannitol medium to each tube. A parallel series of control tubes received 1.8 mL of distilled water and 0.2 mL of the concentrated synthetic medium. For each strain tested, three exudate tubes and three control tubes received 40pL of distilled water containing 2 x los cells. The cultures were maintained at 25 k 1°C without agitation, and one control and one exudate tube of each strain were removed at various inter- vals and prepared for examination as described above. In the first experiment the strains tested were Rhizobi~rtn sp. 3G4b4, 3G4b5,3G4b10,3G4b16,3G4b19, and R. japonicutn 505W, and cells were harvested on days 2. 4, and 6. In the second experi- ment the strains were the same except that 3G4b9a replaced 3G4b19, and cells were harvested on days 3.5, and 7.
Soluble lectin receptors in cult~rre-fllrids In order to examine culture fluids for SBL and PNL receptors,
a quantitative assay that relied on the ability of soluble lectin receptors to competitively inhibit lectin-mediated hemaggluti- nation was developed. The culture fluids for assay were super- natant solutions remaining after removal of cells for the above fluorescent binding assays. The solutions had been frozen im- mediately after centrifugation and were stored at - 16°C until needed.
Prior to each experiment, a 25-pL portion of a stock SBL solution (I mg/mL in PBS) was twofold serially diluted with equal volumes of PBS in a Cooke microtiter hemagglutination plate. A 3% (vlv) suspension of trypsinized erythrocytes in PBS (Pueppke 19796) was added to each well, and after 3 h at room temperature, the hemagglutination titer was recorded. A portion of the stock lectin solution was then diluted soas to contain four times the minimum quantity causing hemagglutination. This lectin concentration was designed to be just sufficient to cause hemagglutination in the subsequent hemagglutination inhibition assays.
For the hemagglutination inhibition assays, 25 pL of each culture fluid was twofold serially diluted as above with equal volumes of PBS. Appropriately diluted test lectin solution (25pL) was then added to each well, followed by 25 pL of erythrocyte suspension. D-Galactose (100mM) served as a positive control in each experiment. After 3 h at room tempera- ture, hemagglutination inhibition titer, defined as the reciprocal of the greatest inhibitor dilution completely preventing hemagglutination, was determined for each culture fluid. The corresponding assays using PNL were identical to those for SBL, except that the erythrocytes were desialyzed (Pueppke
1979a) rather than trypsinized, and that the positive control inhibitor was 50 mM lactose rather than D-galactose.
Binding experittlerlts with radioisotope-labeled lectins Bacteria for these experiments were grown in liquid cultures
ofgluconate mannitol medium scaled up to 500 mL/L flask. Cells were harvested by centrifugation at 7700 x g , washed in PBS, and recentrifuged. The cells were then resuspended in PBS and adjusted turbidimetrically to either 5 x lo8 or lo9 cells/mL. Binding was carried out in 15-mL polycarbonate centrifuge tubes that had been treated with Prosil-28 (PCR Research Chemicals, Gainesville, FL). Replicate tubes received 1 mL of cell suspension followed by 0.15 mL of ['HISBL containing from 10 to 80pg of protein (28000 to 166000 counts per minute (cpm)). To control for nonspecific lectin adherence to the tubes, treatments containing 10 or 80pg of ['HISBL were run with I mL of PBS replacing the cell suspension. To control for nonspecific binding of the lectin to the cells, treatments con- taining 10 or 80 pg of ['HISBL were incubated in the presence of I0 mM N-acetyl-D-galactosamine (Aldrich Chemical Co., Mil- waukee, WI). After incubation of the tubes at 4°C for 1 h, the cells were removed by centrifugation at 7700 x g, and 0.5-mL aliquots of the supernatant solutions were counted using Beckman Ready-Solv EP scintillation fluid and a Beckman LS- 150 liquid scintillation spectrometer. Numbers of binding sites per cell and affinity constants were determined from data points plotted according to the method of Steck and Wallach(1965).
Experiments designed to compare the capacities of cells to bind ['HISBL and [lZSI]PNL were also conducted. [lZSI]PNL was supplied at 165 pgltube (60000 to 70000 cpm) and [3H]SBL was supplied at 120 pgltube (22000 cpm). The tubes contained 5 x IOs cells prepared as described above, and controls for bind- ing specificity were done in the presence of l00mM lactose (PNL) or 10mM N-acetyl-D-galactosamine (SBL).
Results
Nodulation and lectin binding A total of 14 R . japonicunz strains from soybean
and 8 Rhizobium sp. strains from peanut were evaluated for nodulation of the reciprocal heterologous host, peanut or soybean, respectively (Tables 1 and 2). Four of the soybean strains pro- duced nodules on peanut, and in each case spheri- cal bacteroids (Staphorst and Strijdom 1972) were present in crushed nodule preparations. Three of the peanut strains also nodulated soybean, and again the nodules contained bacteroids. We have reported that an isolate of 3G4b4 from another source nodulated Harosoy 63 soybean in an earlier experiment (Su et al. 1980). The isolate used in this study, however, did not nodulate soybean, an ob- servation in accord with previous data for the same isolate (Bhuvaneswari and Bauer 1978; Brethauer and Paxton 1977).
The adherence of the heterologous lectins to log phase cells of the heterologous strains is also given in Tables 1 and 2. FITC-SBL bound in a hapten- specific manner to three of the peanut strains. The capacity of peanut strains to bind FITC-SBL did not coincide with the ability of the strains to be symbiotic with soybean, and indeed strain 3G4b16
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MC
GIL
L U
NIV
ER
SIT
Y o
n 11
/20/
14Fo
r pe
rson
al u
se o
nly.
1492 CAN. J . MICROBIOL. VOL. 26, 1980
TABLE 1. Nodulation of Jumbo Virginia peanut and data obtained using fluorescent-labeled lectins. binding of peanut lectin by R. jnporricrrn~ strains from Thus. although none of the strains bound detecta-
soybean ble amounts of PNL, the binding of fluorescent- and radioisotope-labeled SBL to the strains was qual-
Strainn Sourceb Nodules PNL binding itativelv the same. The calculated number of SBL USD A USDA USDA USDA USDA USDA USDA USDA USDA USD A USDA Nitragin Nitragin Nitragin
'1Each stiain nodulated its homologous plant host, Glycirre rrros (L.) Merr. cultivar Harosoy 63.
*USDA, from the United States Department o f Agriculture. Beltsville, M D ; Nitragin, from the Nitragin Co., Milwaukee, W1.
TABLE 2 . Nodulation of Harosoy 63 soybean and binding of soybean lectin by Rlrizobi~rn~ sp. strains
froni peanut
Straina Sourceb Nodules SBL binding
Paxton USDA USDA USDA USDA USDA USDA USDA
.Each strain nodulated its homologous plant host, Aroclris /rj,pogaea L. cultivar Jumbo Virginia.
*Paxton, from Dr. Jack Paxton, Department o f Plant Pa- thology, University of Illinois, Urbana, IL; USDA, from the United States Department o f Agriculture, Beltsville, M D .
was the only SBL-binding, soybean-nodulating strain. In sharp contrast, FITC-PNL did not adhere to any of the soybean strains (Table 1) and failed to bind to the homologous peanut strains (data not shown). The hemagglutinating activity of the FITC-PNL toward desialyzed human type 0 ery- throcytes was the same as that of unlabeled PNL at the same protein concentration, and in positive controls, FITC-PNL caused erythrocytes to fluoresce. Hence, the fluorescent-labeling proce- dure had not destroyed the binding activity of the lectin. Culture fluids from all 22 strains were also screened to determine if soluble, extracellular PNL receptors were produced. None were detected, al- though the control receptor, lactose, was active to a dilution endpoint of 4.28 mg/mL.
Six strains were selected for quantitative testing using theoretically saturating amounts of [lZSI]PNL and [3H]SBL. The results (Table 3) confirmed the
molecdes bound per cell of soybean strain 3Ilb143 was 460000, a value in excellent agreement with estimates obtained previously with this and other soybean strains (Bhuvaneswari et al. 1977; Meyer and Pueppke 1980). Strains 3G4b4 and 3G4b5 from peanut were similar to 3Ilb143, and each bound about 500000 moleculeslcell.
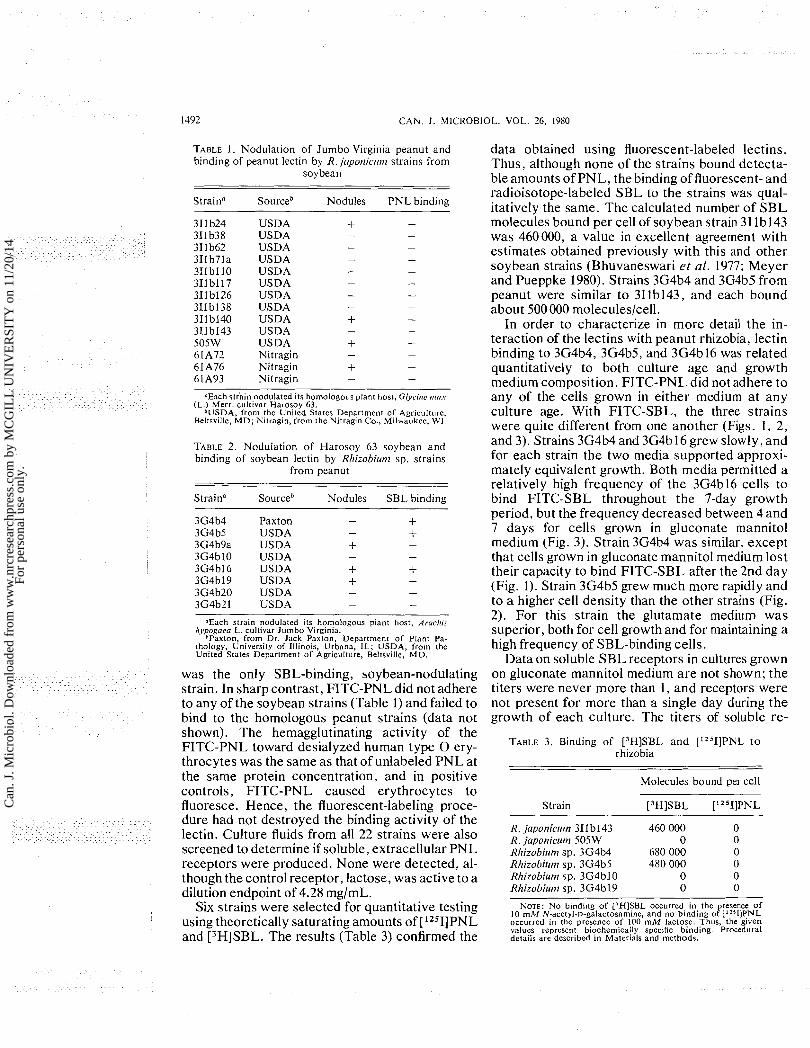
In order to characterize in more detail the in- teraction of the lectins with peanut rhizobia, lectin binding to 3G4b4, 3G4b5, and 3G4b16 was related quantitatively to both culture age and growth medium composition. FITC-PNL did not adhere to any of the cells grown in either medium a t any culture age. With FITC-SBL, the three strains were quite different from one another (Figs. 1, 2, and 3). Strains 3G4b4 and 3G4b 16 grew slowly, and for each strain the two media supported approxi- mately equivalent growth. Both media permitted a relatively high frequency of the 3G4b16 cells to bind FITC-SBL throughout the 7-day growth period, but the frequency decreased between 4 and 7 days for cells grown in gluconate mannitol medium (Fig. 3). Strain 3G4b4 was similar, except that cells grown in gluconate mannitol medium lost their capacity to bind FITC-SBL after the 2nd day (Fig. 1). Strain 3G4b5 grew much more rapidly and to a higher cell density than the other strains (Fig. 2). For this strain the glutamate medium was superior, both for cell growth and for maintaining a high frequency of SBL-binding cells.
Data on soluble SBL receptors in cultures grown on gluconate mannitol medium are not shown; the titers were never more than 1, and receptors were not present for more than a single day during the growth of each culture. The titers of soluble re-
TABLE 3. Binding of [3H]SBL and ['251]PNL t o rhizobia
Molecules bound per cell
Strain [3H]SBL [L2511PNL
R. japoniclrrn 311 b143 460 000 0 R. japonicrrnt 505W 0 0 Rhizobirrm sp. 3G4b4 680 000 0 Rllizobilmt sp. 3G4b5 480 000 0 Rhizobirrm sp. 3G4b10 0 0 Rlrizobiut~t sp. 3G4b19 0 0
NOTE: N O binding o f [3H]SBL occurred in the presence o f 10 mM N-acetyl-D-galaclosamine, and no binding o f ['1511PNL occurred in the presence o f 100 m M lactose. Thus, the given values represent biochemica!ly specific binding. Procedural details are described in Materials and methods.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MC
GIL
L U
NIV
ER
SIT
Y o
n 11
/20/
14Fo
r pe
rson
al u
se o
nly.

PUEPPKE ET AL. 1493
E : [ I- I m m . r d I 2 3 4 5 6 7
C U L T U R E A G E (DAYS1 32
FIG. 1. Influence of growth medium composition and culture o I 2 3 4 5 6 7
age on the binding of SBL to cells and soluble lectin receptors of CULTURE A O E ( D A Y S )
~%izob iu t ,~ sp. 3 ~ 4 b 4 . Solid lines represent gluconate mannitol medium, and dashed lines represent glutamate medium. Circles indicate total cells per millilitre, and squares indicate cells bind- ing FITC-SBL per millilitre. The vertical bars represent hemagglutination inhibition titers of soluble lectin receptors from cells grown in glutamate medium. Lectin binding was prevented in all experiments when the binding mixtures were made 100 mM with respect to D-galactose, and for the binding of FITC-SBL to cells, standard deviations were less than f 5 cells1100 cells counted.
ceptors for glutamate-grown 3G4b4 and 3G4b16 were similarly low, but the receptors were present for extended periods of time and seemed to appear coincident with a decrease in the frequency of cells that bound the lectin (Figs. 1 and 3). Strain 3G4b5 produced many more receptors, and a maximum titer of 128 was recorded on day 4 (Fig. 2). By day 5, however, the 3G4b5 receptors had abruptly disap- peared. D-Galactose controls were included with each hemagglutination inhibition assay, and the hapten was active when diluted to from 283 to 565 pg/mL (1.57 to 3.13 mM).
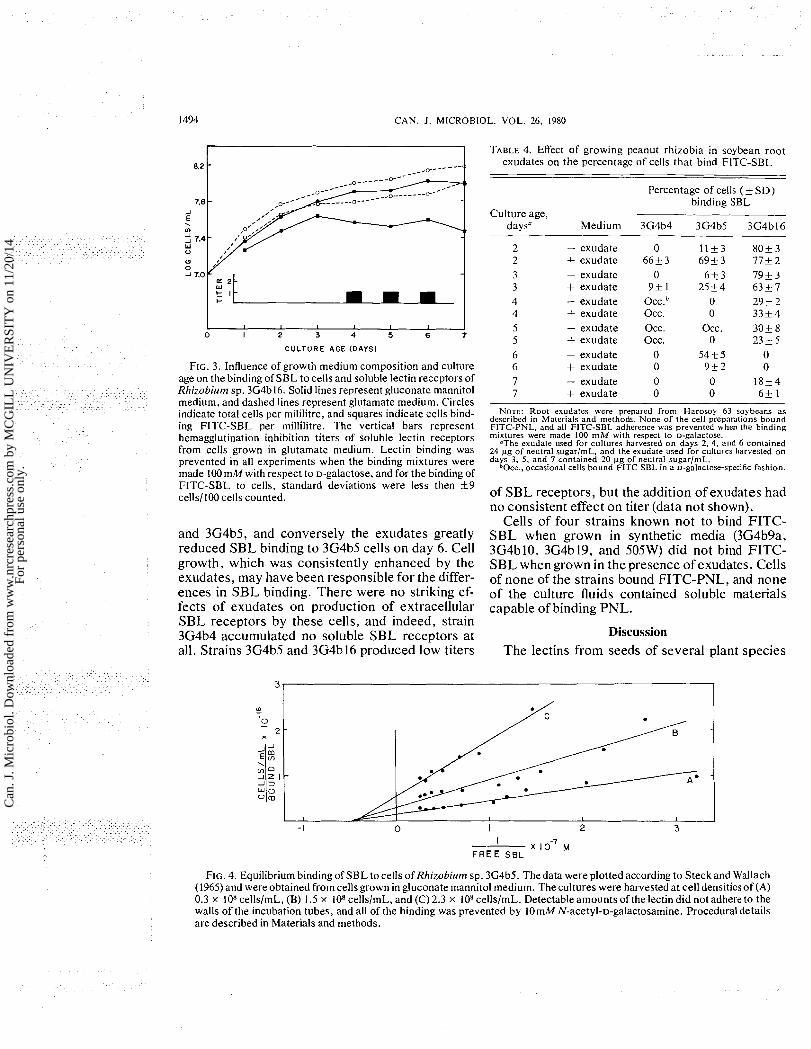
Equilibriuln binding Because it was not possible to estimate accu-
rately variations in the intensity of fluorescence associated with cells that bound FITC-SBL, data on the affinity of SBL to cells of different ages were obtained from equilibrium binding experiments using cells of 3G4b5. The data were calculated by the method of Steck and Wallach (1965), and linear regressions, which had correlation coefficients varying from f0.96 to +0.99, were plotted (Fig. 4). The values of the affinity constant, K, betw'een SBL and cells from cultures having 0.3 x lo8, 1.5 x lo8, or 2.3 x lo8 cells/mL were4.9 x lo6, 4.2 x lo6,
FIG. 2. Influence of growth medium composition and culture age on the binding of SBL to cells and soluble lectin receptors of Rllizobi~it?~ sp. 3G4b5. Solid lines represent gluconate mannitol medium, and dashed lines represent glutamate medium. Circles indicate total cells per millilitre, and squares indicate cells bind- ing FITC-SBL per millilitre. The vertical bars represent hemagglutination inhibition titers of soluble lectin receptors from cells grown in glutamate medium. Lectin binding was prevented in all experiments when the binding mixtures were made 100 mM with respect to D-galactose, and for the binding of FITC-SBL to cells, standard deviations were less than +7 cells1100 cells counted.
and 4.3 x lo6 M - I , respectively. Incontrast withthe rather close agreement of affinity constants, the values for n , the number of binding sites per cell, decreased from 44 1 000 to 258 000 to 128 000/cell in the corresponding cultures. N-Acetyl-D- galactosamine prevented 100% of the binding in each experiment, and thus the adherence of the lectin to the cells was entirely biochemically specific.
Effect of soybean root exudates o n lectin binding Root exudates from soybean and pea (Pisum
satiuuln L.) have been reported to stimulate soy- bean rhizobia to express SBL receptors (Bhuvaneswari and Bauer 1978). A pair of experi- ments was accordingly performed to evaluate the effect of soybean root exudates on the frequency of rhizobial cells that bind FITC-SBL and FITC- PNL. The results with strains known to bind FITC-SBL are in Table 4, and in general the effect of the exudates on the binding of cells to FITC-SBL was minimal. Nevertheless, on day 2 the exudates greatly stimulated SBL binding by strains of 3G4b4
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MC
GIL
L U
NIV
ER
SIT
Y o
n 11
/20/
14Fo
r pe
rson
al u
se o
nly.
1494 CAN. J . MICROBIOL. VOL. 26, 1980
I I I I I 0 I 2 3 4 5 6 7
CULTURE AGE (DAYS)
FIG. 3. Influence of growth medium composition and culture age on the binding of SBL to cells and soluble lectin receptors of Rhizobiutn sp. 3G4b16. Solid lines represent gluconate mannitol medium, and dashed lines represent glutamate medium. Circles indicate total cells per mililitre, and squares indicate cells bind- ing FITC-SBL per millilitre. The vertical bars represent hemagglutination inhibition titers of soluble lectin receptors from cells grown in glutamate medium. Lectin binding was prevented in all experiments when the binding mixtures were made 100 mM with respect to D-galactose, and for the binding of FITC-SBL to cells, standard deviations were less then +9 cells1100 cells counted.
and 3G4b5, and conversely the exudates greatly reduced SBL binding to 3G4b5 cells on day 6. Cell growth, which was consistently enhanced by the exudates, may have been responsible for the differ- ences in SBL binding. There were no striking ef- fects of exudates on production of extracellular SBL receptors by these cells, and indeed, strain 3G4b4 accumulated no soluble SBL receptors at all. Strains 3G4b5 and 3G4b16 produced low titers
TABLE 4. Effect of growing peanut rhizobia in soybean root exudates on the percentage of cells that bind FITC-SBL
Percentage of cells ( f SD) binding SBL
Culture age, daysa Medium 3G4b4 3G4b5 3G4bIG
- exudate + exudate - exudate + exudate - exudate + exudate - exudate + exudate - exudate + exudate - exudate + exudate
0 66k3
0 9+ 1
O C C . ~ Occ. occ. occ.
0 0 0 0
l l t 3 8 0 t 3 6 9 t 3 77+2 6 t 3 7 9 t 3
25+4 6 3 k 7 0 29k 2 0 3 3 t 4
Occ. 3 0 t 8 0 2 3 k 5
54+ 5 0 9 + 2 0 0 18+4 0 6 t 1
NOTE: Root exudates were prepared from Harosoy 63 soybeans as described in Materials and methods. None of the cell preparations bound FITC-PNL, and all FITC-SBL adherence was prevented when the binding mixtures were made 100 m M with respect to D-galactose.
"The exudate used for cultures harvested o n davs 2. 4 . and 6 contained 24 wg of neutral sugarlmL, and the exudate used fbr chtures harvested o n days 3, 5, and 7 contained 20 wg of neutral sugar/mL.
b O ~ ~ . , occasional cells bound FITC-SBL in a D-galactose-specific fashion.
of SBL receptors, but the addition of exudates had no consistent effect on titer (data not shown).
Cells of four strains known not to bind FITC- SBL when grown in synthetic media (3G4b9a, 3G4b10, 3G4b19, and 505W) did not bind FITC- SBL when grown in the presence of exudates. Cells of none of the strains bound FITC-PNL, and none of the culture fluids contained soluble materials capable of binding PNL.
Discussion The lectins from seeds of several plant species
I I o - ~ F R E E SBL
FIG. 4. Equilibrium binding of SBL to cells of Rhizobium sp. 3G4b5. Thedata were plotted according to Steckand Wallach (1965) and were obtained from cells grown in gluconate mannitol medium. The cultures were harvested at cell densitiesof (A) 0.3 x lo8 cellslmL, (B) 1.5 x lo8 cellslmL, and (C) 2.3 x lo8 cellsImL. Detectable amounts of the lectin did not adhere to the walls of the incubation tubes, and all of the binding was prevented by 10mM N-acetyl-D-galactosamine. Procedural details are described in Materials and methods.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MC
GIL
L U
NIV
ER
SIT
Y o
n 11
/20/
14Fo
r pe
rson
al u
se o
nly.

PUEPPKE ET AL. 1495
have been shown to adhere preferentially to homologous rhizobial symbionts (Bhuvaneswari et al. 1977; Bohlool and Schmidt 1974; Dazzo and Hubbell 1975). These and ensuing studies led to formulation of the hypothesis that seed lectins may function in symbiosis, perhaps to specify host range (Bauer 1977; Dazzo and Hubbell 1975; Schmidt 1979). In soybean the initial screening study demonstrated a striking specificity of SBL for homologous R. japonicum strains; although 88% of the tested symbiont strains bound SBL, not a single heterologous nonsymbiont strain interacted with the lectin (Bohlool and Schmidt 1974). More recent experiments have revealed that a number of heterologous nonnodulating strains, including rep- resentatives of R. leguminosarum, R. trifolii, R. meliloti, Rhizobium sp., and R. phaseoli, do in fact bind SBL, but these strains have not been investi- gated in detail (Bhuvaneswari and Bauer 1978; Brethauer and Paxton 1977; Chen and Phillips 1976; Law and Strijdom 1977).
The interaction of SBL with cells of homologous R. japonicum strains is a complex matter. In a given population the frequency of cells capable of binding the lectin is highly variable, and for the lectin- binding cells the calculated number of binding sites per cell fluctuates with culture age (Bhuvaneswari et al. 1977). Strains may consist of stable (Meyer and Pueppke 1980) or unstable (Shantharam et al. 1980) subpopulations of cells, and soluble SBL re- ceptors, presumably extracellular polysaccharides (Bal et al. 1978; Calvert et al. 1978), may accumu- late in culture fluids (Bhuvaneswari et al. 1977). Moreover, there is evidence that R. japorzic~rm strains incapable of binding the lectin when grown in synthetic media can be induced to do so when grown in media containing host or nonhost root exudates (Bhuvaneswari and Bauer 1978).
We have found that the interaction of SBL with peanut rhizobia, either symbionts or nonsymbionts of soybean, is generally similar to the interaction of the lectin with homologous R. japonicum strains. Thus for Rhizobi~rrn sp. strains 3G4b4 and 3G4b5 (soybean nonsymbionts) and 3G4b 16 (soybean symbiont) the frequency of cells binding SBL is initially great and may decline, sometimes to zero, as cultures age. The affinity constant for the adher- ence of SBL to cells of strain 3G4b5, however, did not change as cultures aged. This implies that con- current with a decrease in the number of receptors per cell, there was no alteration in receptor compo- sition. In contrast, the SBL binding plots obtained with strains of R. japonicum are biphasic and com- plex, suggesting that receptor structures are
heterogeneous or variable (Bhuvaneswari et al. 1977).
Unlike the rather cumbersome procedure used in the past to detect extracellular receptors (Bhuvaneswari et al. 1977), the hemagglutination inhibition assay was both rapid and quantitative. Extracellular receptors invariably appeared when cellular receptors began to decline, indicating that the former may have been detached cellular re- ceptors, as has been suggested previously (Bhuvaneswari et al. 1977). Both the cellular and extracellular receptors of strain 3G4b5 eventually disappeared, and thus the degradation or masking of extracellular receptors is an additional com- plicating factor with this, and perhaps other strains.
Medium composition had a major influence on the capacity of cells from liquid cultures to bind SBL. Soybean root exudates did not permit four strains normally incapable of binding the lectin to do so, although for other strains the exudates af- fected both cell growth rates and the frequencies of cells binding the lectin. These results are not in accord with those of Bhuvaneswari and Bauer (1978), who claimed that similar exudates permit- ted from 1 to 5% of the cells in normally nonbinding populations to bind the lectin. Although our exu- dates were prepared according to earlier methods, we used Harosoy 63 rather than Beeson soybeans, and this may account for the discrepancy. Nevertheless it is clear that host root exudates do not invariably convert cells of soybean symbionts such as 3G4b9a, 3G4b10, and 3G4b19 from the nonbinding to the binding condition.
Taken together, the data on SBL do not lend much support to the hypothesis that SBL functions in symbiosis by preferentially adhering to strains that nodulate soybean. As a group the peanut strains resembled previously studied soybean strains in their interactions with SBL. Moreover, with respect to frequency of'cells that bind SBL, dependence of lectin binding on culture age, and influence of exudates on lectin adherence, the peanut strains capable of nodulating soybean are not distinguishable from the peanut strains that do not nodulate soybean.
Incontrast withsoybean, peanut has notbeen well investigated with respect to the lectin hypothesis. PNL is well characterized (Lotan, Skutelsky et al. 1975) and is composed of six or seven isolectins, depending on cultivar (Pueppke 1979a). The Jumbo Virginia cultivar used here contained seven isolec- tins, and several are present in seedling roots (Pueppke 1979a). Nonetheless, the lectin failed to bind to any of the rhizobia, irrespective of growth
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MC
GIL
L U
NIV
ER
SIT
Y o
n 11
/20/
14Fo
r pe
rson
al u
se o
nly.

1496 C A N . J. MICROBIOL. VOL. 26. 1980
medium composition and culture age. Similarly, we could obtain no evidence for the presence of solu- ble PNL receptors in any cultures.
While this work was in progress, Bhagwat and Thomas (1980) reported that two strains of peanut rhizobia bind PNL, although R. japonicum, R. meliloti, Escherichia coli, and Bacillus subtilis (one strain each) do not. The number of molecules bound per cell of peanut strain TG-3 at lectin sat- uration was about 6000, and the receptors were apparently both lipopolysaccharides and extracel- lular polysaccharides. The R. japonicum strain unfortunately was not identified, and it was as- sumed, but evidently not confirmed, to be a peanut nonsymbiont. Regardless, it is clear that homolog- ous peanut symbionts capable of binding small amounts of PNL do exist, although the biochemical specificity of the lectin adherence, and indeed the isolectin composition of the PNL, has not been demonstrated. The aggregate available data nevertheless indicate that the interaction of rhizobia with PNL is not a necessary event in nodulation of peanut.
Acknowledgements We thank Lee-Chin Su and Ulla Benny for ex-
cellent laboratory assistance. This work was sup- ported by National Science Foundation grant No. DEB 77-24444.
BAL, A. K., S. SHANTHARAM, and S. RATNAM. 1978. Ultra- structure ofRhizobiurn japorzicurn in relation to its attachment to root hairs. J. Bacteriol. 133: 1393-1400.
BAUER, W. D. 1977. Lectins as determinants of specificity in legume-Rhizobiurn symbiosis. In Genetic engineering for nit- rogen fixation. Edited by A. Hollaender. Plenum Press, New ~ o r k . pp. 283-295.
BHAGWAT. A. A.. and J. THOMAS. 1980. Dual binding sites for peanut lectin on rhizobia. J. Gen. Microbiol. 117: 119-125.
BHUVANESWARI, T. V., and W. D. BAUER. 1978. Role oflectins in plant-microorganism interactions. Ill. Influence of rhizo- spherelrhizoplane culture conditions on the soybean lectin- binding properties of rhizobia. Plant Physiol. 62: 71-74.
BHUVANESWARI, T. V., S. G. PUEPPKE, and W. D. BAUER. 1977. Role of lectins in plant-microorganism interactions. I . Binding of soybean lectin to rhizobia. Plant Physiol. 60: 486-49 1.
BOHLOOL, B. B., and E. L. SCHMIDT. 1974. Lectins: a possible basis for specificity in the Rhizobium - legume root nodule symbiosis. Science, 185: 269-271.
BRETHAUER, T. S., and J. D. PAXTON. 1977. The role of lectin in soybean - Rhizobium japonicum interactions. In Cell wall biochemistry related to specificity in host-plant pathogen interactions. Edited by B. Solheim and J. Raa. Univer- sitetsforlaget, Oslo. pp. 381-384.
BROUGHTON, W. J. 1978. Control of specificity in legume-Rhizobium associations. J . Appl. Bacteriol. 45: 165-194.
CALVERT, H. E., M. LALONDE, T. V. BHUVANESWARI, and W. D. BAUER. 1978. Role of lectins in plant-microorganism
interactions. IV. Ultrastructural localization of soybean lectin binding sites on Rhizobium japonicum. Can. J. Microbiol. 24: 785-793.
CARROLL, W. R. 1934. A study ofRhizobium speciesin relation to nodule formation on the roots of Florida legumes: I. Soil Sci. 37: 117-135.
CHEN, A. T., and D. A. PHILLIPS. 1976. Attachment of Rhizobiurn to legume roots as the basis for specific interac- tions. Physiol. Plant. 38: 83-88.
D ~ z z o , F. B., and D. H. HUBBELL. 1975. Cross-reactive anti- gens and lectin as determinants of symbiotic specificity in the Rl~izobium-clover association. Appl. Microbiol. 30: 1017- 1033.
KABAT, E. A. 196 1. Kabat and Mayer's experimental immuno- chemistry. Charles Thomas, Springfield, IL. p. 504-5 13.
LAW, I. J., and B. W. STRIJDOM. 1977. Some observations on plant lectins and Rllizobium specificity. Soil Biol. Biochem. 9: 79-84.
LEONARD, L. T. 1923. Nodule-production kinship between the soybean and the cowpea. Soil Sci. 15: 277-283.
LOTAN, R. , H. DEBRAY, M. CACAN, R. CACAN, and N. SHA- RON. 1975. Labeling of soybean agglutinin by oxidation with sodium periodate followed by reduction with sodium [3H]borohydride. J. Biol. Chem. 250: 1955-1957.
LOTAN, R., E. SKUTELSKY, D. DANON, and N. SHARON. 1975. The purification. composition, and specificity of the anti-T lectin from peanut (Arachis hypogaea). J. Biol. Chem. 250: 85 18-8523.
LOWRY, 0. H., N. J. ROSEBROUGH, A. L. FARR, and R. J. RANDALL. 1951. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 193: 265-275.
MEYER, M. C., and S. G. PLJEPPKE. 1980. Differentiation of Rhizobium japonicum strain derivatives by antibiotic sen- sitivity patterns, lectin binding, and utilization of biochemi- cals. Can. J. Microbiol. 26: 606-612.
PEREIRA, M. E. A., E. A. KABAT, R. LOTAN, and N. SHARON. 1976. Immunochemical studies on thespecificity ofthe peanut (Arachis hypogaea) agglutinin. Carbohydr. Res. 51: 107-1 18.
PEREIRA, M. E. A,, E. A. KABAT, and N. SHARON. 1974. Im- munochemical studies on the specificity of soybean aggluti- nin. Carbohydr. Res. 37: 89-102.
PLJEPPKE, S. G. 1979a. Distribution of lectins in the Jumbo Virginia and Spanish varieties of the peanut, Arachis llypogaea L. Plant Physiol. 64: 575-580.
19796. Purification and characterization of a lectin from seeds of the winged bean, Psophocarpus tetragonolobus (L.)DC. Biochim. Biophys. Acta, 581: 63-70.
PULL, S. P., S. G. PUEPPKE, T. HYMOWITZ, and J. H. ORF. 1978. Soybean lines lacking the 120,000-dalton soybean lec- tin. Science, 200: 1277-1279.
SCHMIDT, E. L. 1979. Initiation of plant root - microbe interac- tions. Annu. Rev. Microbiol. 33: 355-376.
SEARS, 0. H., and W. R. CARROLL. 1927. Cross inoculation with cowpea and soybean nodule bacteria. Soil Sci. 24: 413-419.
SELA, B., H. LIS, N. S H A R O N , ~ ~ ~ L . SACHS. 1971. Quantitation of N-acetyl-D-galactosamine-like sites on the surface mem- brane of normal and transformed mammalian cells. Biochim. Biophys. Acta, 249: 564-568.
SHANTHARAM, S., J . A. GOW, and A. K. BAL. 1980. Fractiona- tion and characterization of two morphologically distinct types of cells in Rhizobiutn japonicurn broth culture. Can. J. Microbiol. 26: 107-1 14.
STAPHORST, J. L., and B. W. STRIJDOM. 1972. Some observa- tions on the bacteroids in nodules of Arachis spp. and the isolation of rhizobia from these nodules. Phytophylactica, 4: 87-92.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MC
GIL
L U
NIV
ER
SIT
Y o
n 11
/20/
14Fo
r pe
rson
al u
se o
nly.

PUEPPKE ET AL. 1497
STECK, T. L., and D. F. H. WALLACH. 1965. The binding of kidney-bean phytohemagglutinin by Ehrlich ascites car- cinoma. Biochim. Biophys. Acta, 97: 510-522.
Su, L. C., S. G. PUEPPKE, and H. P. FRIEDMAN. 1980. Lectins and the soybean-Rhizobiurn symbiosis. I. Immunological in- vestigations of soybean lines, the seeds of which have been reported to lack the 120,000 dalton soybean lectin. Biochim. Biophys. Acta, 629: 292-304.
UDENFRIEND, S. 1962. Fluorescence assay in biology and medicine. Academic Press, New York. p. 22 1.
VINCENT, J. M. 1970. A manual for the practical study of root- nodule bacteria. Blackwell Scientific Publications, Oxford.
WALKER, R. H., and P. E. BROWN. 1935. The nomenclature of the cowpea group of root-nodule bacteria. Soil Sci. 39: 22 1-225.
WILSON, J . R. 1944. Over five hundred reasons for abandoning the cross-inoculation groups of the legumes. Soil Sci. 58: 61-69.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
MC
GIL
L U
NIV
ER
SIT
Y o
n 11
/20/
14Fo
r pe
rson
al u
se o
nly.