interaction between n and c in soil has consequences for global carbon cycling
TRANSCRIPT

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions,research libraries, and research funders in the common goal of maximizing access to critical research.
Interaction between N and C in Soil has Consequences for Global CarbonCyclingAuthor(s): Siegfried FleischerSource: Journal of Resources and Ecology, 3(1):16-19. 2012.Published By: Institute of Geographic Sciences and Natural Resources Research, Chinese Academy ofSciencesDOI: http://dx.doi.org/10.5814/j.issn.1674-764x.2012.01.003URL: http://www.bioone.org/doi/full/10.5814/j.issn.1674-764x.2012.01.003
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological,and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and bookspublished by nonprofit societies, associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance ofBioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercialinquiries or rights and permissions requests should be directed to the individual publisher as copyright holder.

J. Resour. Ecol. 2012 3 (1) 016-019
DOI:10.5814/j.issn.1674-764x.2012.01.003
www.jorae.cn
March, 2012 Journal of Resources and Ecology Vol.3 No.1
Article
Received: 2011-11-25 Accepted: 2011-11-30* The author: Siegfried FLEISCHER. Email: [email protected]
1 IntroductionThe general view until now has been that CO2 emissions from soils (or from soil samples at the laboratory) can be used as a proxy for respiratory activity (Janssens et al. 2010). But gross heterotrophic respiration (GHR) within the soil, and net (NHR) which is CO2 emitted from the soil, were different as a result of within-soil CO2 reduction. Inhibition of N-cycling enabled measurement in the laboratory of a within-soil CO2-sink as the difference between GHR and NHR (Fleischer and Bouse 2008). The outcome of the initial rough estimate deserved further investigation, and resulted in the present broadened study. It is necessary to include within-soil CO2 sequestration to correctly understand the C-cycle.
The eddy covariance technique for flux measurements has largely contributed to increased knowledge of CO2-exchange between the atmosphere and terrestrial ecosystems. If this is the sole technique, however, land areas are ”black boxes” not disclosing within-ecosystem
processes such as the within-soil process reported here. From the presentation of a global database on forest ecosystems (Lyussaert et al. 2007) it appeared that autotrophic, heterotrophic and ecosystem respirations have few direct observations and should be measured independently. They also suggested chamber measurements to increase knowledge on CO2-emissions from different types of land areas, but even with this method included, information on the underlying soil processes resulting in NHR is limited.
It has long been known that N-supply decreases the efflux of CO2 from soils, an effect often viewed upon as respiration impairment in some way (Janssens et al. 2010). Billing and Ziegler (2008) already followed this interpretation, but, importantly, they showed that elevated CO2 increased this process. An increase in NH4
+ does favour the chemoautotrophic process nitrification, but so does CO2 which is the sole carbon source for this process. This was our background when identifying the within-soil CO2-sink which could be prevented by specific inhibition
Interaction between N and C in Soil has Consequences for Global Carbon Cycling
Siegfried FLEISCHER
Rydberg Laboratory, Halmstad University, P.O. Box 823, S-301 18 Halmstad, Sweden
Abstract: Energy-yielding processes in the N-cycle form important links with the global C-cycle.
One example is demonstrated with the supply of nitrogen to soils, initially resulting in lowered CO2
emissions. This well known effect has mostly been interpreted as hampered or delayed soil respiration.
When added in surplus, however, nitrogen supply does not stabilize the minimum emissions initially
obtained, but gradually results in increased CO2 emissions. Specific inhibition of the CO2 consuming
process nitrification in soils, with surplus ammonium supply or with acetylene, mostly results in
additional CO2 emissions. The difference between this disclosed gross heterotrophic respiration
(GHR) and the net CO2 emission (NHR) is the result of a within-soil CO2-sink. Soil respiration solely
determined as CO2 emitted as NHR (the common situation) therefore may lead to misinterpretations
of the function of the soil system, especially in areas with high N-deposition. As a consequence, the
interpreted ’acclimation’ of the soil respiration response in a warmer world should be reconsidered.
The concept of respiration inhibition by nitrogen supply may also be questioned. Disregard of these
processes, including the indicated N-driven within-soil CO2-sink, may prevent adequate measures
counteracting climate change.
Key words: soil respiration; carbon sink; CO2 emissions; temperature response; climate change; N-supply
Siegfried FLEISCHER: Interaction between N and C in Soil has Consequences for Global Carbon Cycling 17
of nitrification (Fleischer and Bouse, 2008). A strong CO2-sink did not generally seem to coincide with increased NO3
- in the soil water. A possible interpretation was that repeated N-cycling generated the energy needed for the extensive CO2-reduction in different soils, as such an expression of “gross nitrification”. Terrestrial sinks as a result solely of the extensive photosynthetic CO2 uptake into green plants above the soil surface was questioned.
These initial findings are the basis for the hypothesis that N-supply under current atmospheric N-deposition conditions does not decrease heterotrophic soil respiration – it enforces the within-soil CO2-sink, in this way leading to decreased CO2-emissions. At very high N-supply rates nitrification is gradually inhibited, and CO2-emissions are again increasing (Fleischer and Bouse 2008), thus contradicting the prevailing interpretation that heterotrophic respiration is inhibited by N-supply (Olsson et al. 2005; Janssens et al. 2010). Results presented here support the role of the within-soil CO2-sink.
2 MethodsTwenty-five to forty composite soil samples (φ25 mm) from the 0–5 cm horizon (in a vertical profile 0–5, 5–10 and 10–15 cm) were transported at sampling temperatures and sieved (2 mm) immediately after arrival to the laboratory. Possible remains of fine roots were included in the saprophytic system. Preparation and incubation for 3–7 hours in darkness of each soil sample included series of 12 ml exetainers without (references), and with additions of NH4
+ to increase nitrification, and with acetylene (0.8%–2.0%) to inhibit nitrification, and followed the procedures by Fleischer and Bouse (2008). The study is based on 225 triplets (each with 3 references and 3 with acetylene or N-additions), 8 with five replicates and 8 duplicates. It covered frost free conditions in soils from 2004 to 2009, from central Europe (50°09′ N) to northern Scandinavia (66°03′ N), and two sampling occations in a temperate rainforest in Washington State, USA (47°52′ N). Compared to the previous study (Fleischer and Bouse 2008) the weight of high atmospheric N-deposition areas has been lowered from 25% to 15% and the share of cropland, represented by the Mellby research fields in SW Sweden (56°20′N, 12°59′ E) is 37% which is close to the European situation.
CO2-production in the saprophytic soil system was measured with GS-chromatography on Hay Sep Q in the atmosphere from the sealed exetainers. Calculation of the soil CO2-sink followed (GHR-NHR)/GHR*100 (percent of GHR).
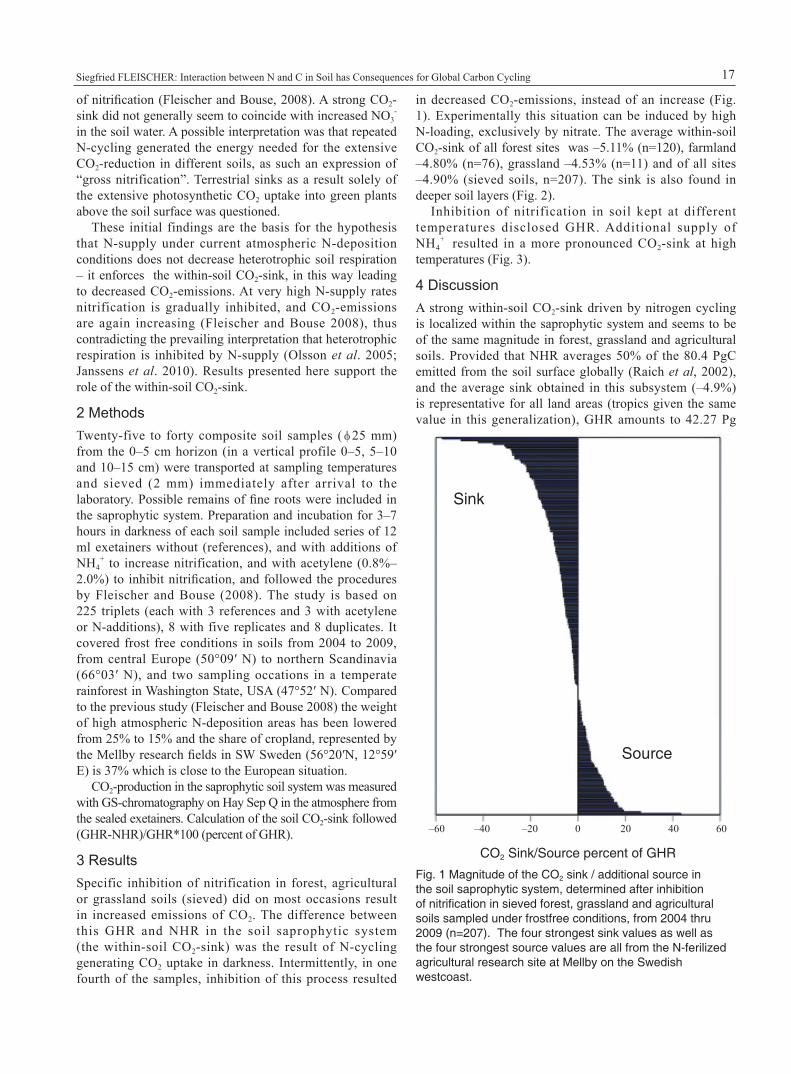
3 ResultsSpecific inhibition of nitrification in forest, agricultural or grassland soils (sieved) did on most occasions result in increased emissions of CO2. The difference between this GHR and NHR in the soil saprophytic system (the within-soil CO2-sink) was the result of N-cycling generating CO2 uptake in darkness. Intermittently, in one fourth of the samples, inhibition of this process resulted
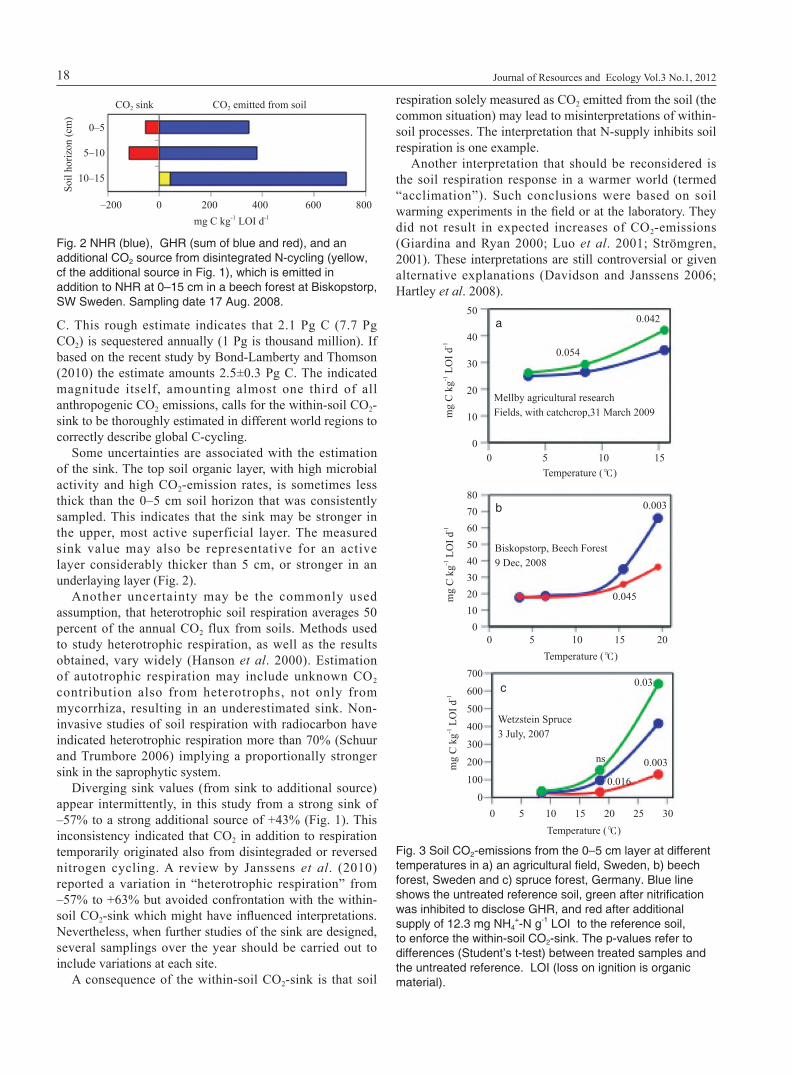
in decreased CO2-emissions, instead of an increase (Fig. 1). Experimentally this situation can be induced by high N-loading, exclusively by nitrate. The average within-soil CO2-sink of all forest sites was –5.11% (n=120), farmland –4.80% (n=76), grassland –4.53% (n=11) and of all sites –4.90% (sieved soils, n=207). The sink is also found in deeper soil layers (Fig. 2).
Inhibition of nitrification in soil kept at different temperatures disclosed GHR. Additional supply of NH4
+ resulted in a more pronounced CO2-sink at high temperatures (Fig. 3).
4 DiscussionA strong within-soil CO2-sink driven by nitrogen cycling is localized within the saprophytic system and seems to be of the same magnitude in forest, grassland and agricultural soils. Provided that NHR averages 50% of the 80.4 PgC emitted from the soil surface globally (Raich et al, 2002), and the average sink obtained in this subsystem (–4.9%) is representative for all land areas (tropics given the same value in this generalization), GHR amounts to 42.27 Pg
Fig. 1 Magnitude of the CO2 sink / additional source in the soil saprophytic system, determined after inhibition of nitrification in sieved forest, grassland and agricultural soils sampled under frostfree conditions, from 2004 thru 2009 (n=207). The four strongest sink values as well as the four strongest source values are all from the N-ferilized agricultural research site at Mellby on the Swedish westcoast.
Sink
Source
–60 –40 –20 0 20 40 60
CO2 Sink/Source percent of GHR
Journal of Resources and Ecology Vol.3 No.1, 201218
C. This rough estimate indicates that 2.1 Pg C (7.7 Pg CO2) is sequestered annually (1 Pg is thousand million). If based on the recent study by Bond-Lamberty and Thomson (2010) the estimate amounts 2.5±0.3 Pg C. The indicated magnitude itself, amounting almost one third of all anthropogenic CO2 emissions, calls for the within-soil CO2-sink to be thoroughly estimated in different world regions to correctly describe global C-cycling.
Some uncertainties are associated with the estimation of the sink. The top soil organic layer, with high microbial activity and high CO2-emission rates, is sometimes less thick than the 0–5 cm soil horizon that was consistently sampled. This indicates that the sink may be stronger in the upper, most active superficial layer. The measured sink value may also be representative for an active layer considerably thicker than 5 cm, or stronger in an underlaying layer (Fig. 2).
Another uncertainty may be the commonly used assumption, that heterotrophic soil respiration averages 50 percent of the annual CO2 flux from soils. Methods used to study heterotrophic respiration, as well as the results obtained, vary widely (Hanson et al. 2000). Estimation of autotrophic respiration may include unknown CO2 contribution also from heterotrophs, not only from mycorrhiza, resulting in an underestimated sink. Non-invasive studies of soil respiration with radiocarbon have indicated heterotrophic respiration more than 70% (Schuur and Trumbore 2006) implying a proportionally stronger sink in the saprophytic system.
Diverging sink values (from sink to additional source) appear intermittently, in this study from a strong sink of –57% to a strong additional source of +43% (Fig. 1). This inconsistency indicated that CO2 in addition to respiration temporarily originated also from disintegraded or reversed nitrogen cycling. A review by Janssens et al. (2010) reported a variation in “heterotrophic respiration” from –57% to +63% but avoided confrontation with the within-soil CO2-sink which might have influenced interpretations. Nevertheless, when further studies of the sink are designed, several samplings over the year should be carried out to include variations at each site.
A consequence of the within-soil CO2-sink is that soil
respiration solely measured as CO2 emitted from the soil (the common situation) may lead to misinterpretations of within-soil processes. The interpretation that N-supply inhibits soil respiration is one example.
Another interpretation that should be reconsidered is the soil respiration response in a warmer world (termed “acclimation”). Such conclusions were based on soil warming experiments in the field or at the laboratory. They did not result in expected increases of CO2-emissions (Giardina and Ryan 2000; Luo et al. 2001; Strömgren, 2001). These interpretations are still controversial or given alternative explanations (Davidson and Janssens 2006; Hartley et al. 2008).
Fig. 2 NHR (blue), GHR (sum of blue and red), and an additional CO2 source from disintegrated N-cycling (yellow, cf the additional source in Fig. 1), which is emitted in addition to NHR at 0–15 cm in a beech forest at Biskopstorp, SW Sweden. Sampling date 17 Aug. 2008.
Fig. 3 Soil CO2-emissions from the 0–5 cm layer at different temperatures in a) an agricultural field, Sweden, b) beech forest, Sweden and c) spruce forest, Germany. Blue line shows the untreated reference soil, green after nitrification was inhibited to disclose GHR, and red after additional supply of 12.3 mg NH4
+-N g-1 LOI to the reference soil, to enforce the within-soil CO2-sink. The p-values refer to differences (Student’s t-test) between treated samples and the untreated reference. LOI (loss on ignition is organic material).
80706050403020100
0 5 10 15 20
b
Temperature (℃)
0.003
0.045
Biskopstorp, Beech Forest9 Dec, 2008
mg
C k
g-1 L
OI d
-1
50
40
30
20
10
00 5 10 15
a 0.042
0.054
Temperature (℃)
mg
C k
g-1 L
OI d
-1
Mellby agricultural researchFields, with catchcrop,31 March 2009
700
600
500
400
300
200
100
00 5 10 15 20 25 30
c
Temperature (℃)
0.003
0.016
ns
0.03
mg
C k
g-1 L
OI d
-1
Wetzstein Spruce3 July, 2007
–200 0 200 400 600 800
0–5
5–10
10–15
Soil
horiz
on (c
m)
mg C kg-1 LOI d-1
CO2 sink CO2 emitted from soil

Siegfried FLEISCHER: Interaction between N and C in Soil has Consequences for Global Carbon Cycling 19
At in situ conditions (no sieved soils) Lamersdorf and Borken (2004) found that soil respiration had increased significantly in a “clean rain plot” after seven years of reduced N-supply, in relation to a plot with remaining high N-deposition. The effect was most pronounced at higher temperatures and they suggested that more energy was invested into fine root growth to maintain nitrogen uptake from the low N-deposition soil. Mo et al. (2008) showed reduced CO2 emission from a tropical forest soil (unsieved) with high N supply, and this effect was in the same way most pronounced at high temperatures.
The present experimental study with the saprophytic system (no roots present) suggests that there is a CO2 increase at higher temperature, but the increase is partially hidden by the within-soil CO2-sink, and this counteracting process may have led to biological acclimation as an explanation of the response. This effect may occur concurrently with a gradual decrease in fine root respiration, a long term effect of high N-deposition, of which Lamersdorf and Borken (2004) described the reversal process. Both effects may vary between areas with varying atmospheric N-deposition, and both may have contributed to the interpretation termed “acclimation” which might be questioned.
One explanat ion for the remaining knowledge gaps may be that N and C cycles are too often treated separately. Transformation processes within the N-cycle, such as nitrification, dissimilatory nitrate reduction and denitrification, are not ends in themselves. They should be treated as the important links with the C-cycle that they are. The within-soil CO2-sink is daily influenced by human activities such as land use changes or nitrogen deposition. Directed influence on these processes may open for the most significant CO2 removal from cycling discussed so far. Understanding soil processes leading to CO2 exchange between soil and atmosphere, including both GHR and NHR, is an initial need.
Acknowledgements
I thank Harald Grip and Ulf Johansson, Swedish University of Agricultural Sciences, for discussions and review which have
improved the quality of the paper. The study was supported by the Brita and Sven Rahmn foundation and the Bertebo foundation.
ReferencesBillings S A, S E Ziegler. 2008. Altered patterns of soil carbon substrate
usage and heterotrophic respiration in a pine forest with elevated CO2 and N fertilization. Global Change Biology, 14: 1025-1036.
Bond-Lambert B, A Thomson. 2010. Temperature-associated increases in the global soil respiration record. Nature, 464: 579-582.
Davidson E A, I A Janssens. 2006. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature, 440: 165-173.
Fleischer S, I Bouse. 2008. Nitrogen cycling drives a strong within-soil CO2-sink. Tellus, 60B: 782-786.
Giardina C P, M G Ryan. 2000. Evidence that decomposition rates of organic carbon in mineral soil do not vary with temperature. Nature, 404: 858-861.
Hanson P J, N T Edwards, C T Garten, J A Andrews. 2000. Separating root and soil microbial contributions to soil respiration: A review of methods and observations. Biogeochemistry, 48: 115-146.
Hartley I P, D W Hopkins, M S Garnett, M Sommerkorn, P A Wookey. 2008. Soil microbial respiration in artic soil does not acclimate to temperature. Ecology Letters, 11: 1092-1100.
Janssens I A, W Dieleman, S Luyssaert, J-A Subke, M Reichstein, et al. 2010. Reduction of soil respiration in response to nitrogen deposition. Nature Geoscience, 3: 315-322.
Lamersdorf N P, W Borken. 2004. Clean rain promotes fine root growth and soil respiration in a Norway spruce forest. Global Change Biology, 10: 1351-1362.
Luo Y, S Wan, D Hui, L L Wallace. 2001. Acclimatization of soil respiration to warming in a tall grass prairie. Nature, 413: 622-624).
Luyssaert S, I Inglima, M Jung, A D Richardson, M Reichstein, et al. 2007. CO2 balance of boreal, temperate, and tropical forests derived from a global database. Global change biology, 13: 2509-2537
Mo J M, W Zhang, W Zhu, P Gundersen, Y Fang, et al. 2008. Nitrogen addition reduces soil respiration in a mature tropical forest in southern China. Global Change Biology, 14: 403-412.
Olsson P, S Linder, R Giesler, P Högberg. 2005. Fertilization of boreal forest reduces both autotrophic and heterotrophic soil respiration. Global Change Biology,11: 1745-1753.
Raich J W, C S Potter, D Bhagawati. 2002. Interannual variability in global soil respiration. 1980-1994. Global Change Biology, 8: 800-812.
Schuur E A G, S E Trumbore. 2006. Heterotrophic contribution to soil respiration from boreal black spruce forest: using isotopes to partition sources to soil carbon dioxide flux. Global Change Biology, 12:165-176.
Strömgren M. 2001. Soil-Surface CO2 Flux and Growth in a Boreal Norway Spruce Stand. Effects of Soil Warming and Nutrition. Acta Universitatis Agriculturae Sueciae, Silvestria, 220: 1-44.
土壤中氮碳间相互作用对全球碳循环的影响
Siegfried FLEISCHER
瑞典哈尔姆斯塔德大学 Rydberg实验室,瑞典,哈尔姆斯塔德 823信箱
摘 要:氮循环中的能量产生过程构成与全球碳循环间的重要关联。例如,氮投入土壤,一开始导致CO2排放降低。这一众所周知的效应被理解为土壤呼吸作用受到阻碍或延缓。然而,当把冗余考虑在内便知,氮投入并不使最初得到的CO2最低排放量保持不变,而是逐渐导致排放增加。土壤中硝化作用对CO2消耗过程的特异性抑制,加上铵冗余投入或乙炔作用,往往导致额外的CO2排放量。这种总自养性呼吸(GHR)与CO2净排放(NHR)之间的差就是土壤内CO2汇。土壤呼吸作用单纯由NHR产生的CO2排放量(通常情况)来决定会导致对土壤系统的曲解,特别是在氮沉降量高的地区。因此,必须重新考虑温暖区域土壤呼吸作用的“适应环境”问题。可能也需要质疑氮投入带来的“呼吸抑制”概念。无视这些过程,包括上述氮驱动下的土壤内CO2汇过程,也许会有碍于采取足够的措施来对抗气候变化。
关键词:土壤呼吸;碳汇;CO2排放;温度反应;气候变化;氮投入