inhibition with spontaneous reactivation of carboxyl esterases by organophosphorus compounds:...
TRANSCRIPT

Inhibition with Spontaneous Reactivation of Carboxyl Esterases byOrganophosphorus Compounds: Paraoxon as a Model
Jorge Estevez,* Adolfo Garcıa-Perez, Jose Barril, and Eugenio Vilanova
Unidad de Toxicologıa y Seguridad Quımica, Instituto de Bioingenierıa, UniVersidad Miguel Hernandez,Elche (Alicante), Spain
ReceiVed October 7, 2010
In this work kinetic data were obtained for different paraoxon concentrations incubated with chickenserum and the soluble fraction of chicken peripheral nerve. A kinetic model equation was deduced byassuming a multienzymatic system with three different simultaneously occurring molecular phenomena:(1) inhibition; (2) simultaneous spontaneous reactivation; (3) “ongoing” inhibition (inhibition during thesubstrate reaction). A three-dimensional fit of the model was applied to analyze the experimental dataversus the concentration of the inhibitor and the preincubation time in an inhibition experiment. Thebest-fitting model in the soluble fraction of chicken peripheral nerve was compatible with a resistantcomponent (22%) and with two sensitive enzymatic entities (37 and 41%). The corresponding second-order rate constants of inhibition (ki ) 1.8 × 10-3 and 5.1 × 10-3 nM-1 min-1, respectively) and thespontaneous reactivation constants (kr ) 0.428 and 0.011 min-1, respectively) were estimated. The best-fitting model in chicken serum was compatible with a resistant component (5.6%) and with two sensitiveenzymatic entities (22.1 and 72.3%). The corresponding second-order rate constants of inhibition (ki )5.8 × 10-2 and 2.0 × 10-3 nM-1 min-1, respectively) and the spontaneous reactivation constants (kr )0.0044 and 0.0091 min-1, respectively) were estimated. These parameters were similar to those observedin spontaneous reactivation experiments with preinhibited paraoxon samples. The consistency of the resultsof all the experiments is considered an internal validation of the methodology. The results are also consistentwith a significant ongoing inhibition. The proportion of enzymatic components shown in this work bythe inhibition and reactivation of paraoxon is similar to that previously observed in inhibition experimentswith mipafox in both tissues, demonstrating that this kinetic approach provides consistent results in complexenzymatic systems. The high sensitivity (at nanomolar concentrations) of these esterases suggests thatthey may either play a role in toxicity in low-level long-term exposure of organophosphate compoundsor have a protective effect related with the spontaneous reactivation.
1. IntroductionThe acute and delayed neurotoxicity of organophosphorus
compounds (OPs) is mediated by the inhibition of esterases suchas acetylcholinesterase or neuropathy target esterase (NTE) bya covalent organophosphorylating reaction (reaction i in Figure1). It is usually considered irreversible and yields an inactivephosphorylated protein, thus producing a time-progressiveinhibition. This reaction is similar to that of the first catalyticreaction step of the carboxyl-ester substrate, which yields theintermediate acyl-enzyme that is quickly deacylated by hy-drolysis to release the acidic product. In contrast, the phosphoryl-enzyme is not normally reactivated because the dephosphory-lating reaction (reaction r in Figure 1) does not occur, or is veryweak, and so the enzyme is practically inhibited irreversibly.In some cases, the phosphoryl-enzyme can undergo a dealky-lating reaction called “aging” (reaction a in Figure 1), which isnot significant under the experimental conditions of the exampleswith paraoxon provided in this paper because all of the activityis recovered overnight after removal of the inhibitor byultrafiltration. In some cases, the dephosphorylation reactionoccurred at a detectable rate; therefore, spontaneous reactivationshould be considered in the kinetic data analysis.
In vitro experiments involve preincubating the enzymepreparation with an inhibitor concentration (I) during inhibition
times (t) and then incubating with a substrate during theenzyme-substrate reaction time (ts) to measure residual enzymeactivity (E) (Figure 2).
After incubation with the soluble fraction of chicken periph-eral nerve, most soluble phenyl valerate esterase (PVase) activitywas inhibited at a low concentration (nanomolar levels) in assaysin which the substrate was added to a preincubated mixture oftissue with the non-neuropathic organophosphorus compound(OP) paraoxon (O,O′-diethyl p-nitrophenyl phosphate): residualactivity should include the so-called soluble neuropathy targetesterase (S-NTE) which, by definition, is considered resistantto the permanent and progressive (covalent) inhibition byparaoxon. However, paraoxon was proposed to be a potentreversible inhibitor of S-NTE (2), thus creating the need tointroduce profound methodological changes as paraoxon is usedin the standard NTE assay to inhibit only nonrelevant esterases(10). Therefore, the need exists to understand the kineticbehavior of most sensitive soluble PVases in the presence ofparaoxon.
The model equations and approaches for analyzing the kineticbehavior of multienzymatic systems in the presence of aninhibitor, with or without spontaneous reactivation, have beenrecently reviewed (7). If more than one enzyme is present inthe preparation or a partial spontaneous reactivation by dephos-phorylation occurs, a complex kinetic model needs to be applied
* Author to whom correspondence should be addressed (e-mail:[email protected].
Chem. Res. Toxicol. 2011, 24, 135–143 135
10.1021/tx100346c 2011 American Chemical SocietyPublished on Web 12/14/2010

for an accurate calculation of inhibitory potencies and todiscriminate between the different esterases encountered in thepreparation.
A proposed model equation can only be accepted if consistentresults are obtained when different inhibitor concentrations areassayed as follows: the best-fitting model is the same with thesame number of components, the same kinetic constants, andthe same amplitude (proportion of the different components)(7). Nonetheless, when the preparation includes enzymaticcomponents of different sensitivities, the best-fitting modelequation and, therefore, the number of discriminated componentsand their estimated kinetic constants and deduced amplitudevalues may apparently differ for the various inhibitor concentra-tions tested. This has been overcome by using an approach withseveral tiers and by analyzing data with a three-dimensionalmodel equation that has been successfully applied to the analysisof peripheral nerve soluble PVases inhibition by mipafox (N,N′-di-isopropyl phosphorodiamidefluoridate) (6, 7).
The spontaneous reactivation of paraoxon-inhibited esteraseshas been observed in both the chicken peripheral nerve solublefraction and chicken serum (2, 3, 9). The precise analysis ofthe combined spontaneous reactivation and inhibition kineticsof multienzymatic systems has always been hindered not onlyby the systems’ complex mathematical performance but alsoby difficulty in interpretation of the results. Additional complex-ity involves the medium containing a highly sensitive componentthe inhibition of which may continue during the substratereaction (“ongoing inhibition”) (7, 8).
Here a kinetic model has been used, which includes theinhibition process and a spontaneous reactivation in a complexmultienzymatic system in which ongoing inhibition occurs inthe most sensitive component. The model is used to analyzethe chicken peripheral nerve soluble fraction and the serumesterases inhibited by paraoxon.
This work continues the development of methods for simul-taneously characterizing interactions of organophosphate neu-rotoxins with biological samples that potentially contain nu-merous sites, that is, enzymes, that are susceptible to inhibition.Enumeration of sensitive components and the associated inhibi-
tion and reactivation kinetics are determined, in this case, forparaoxon reactions with chicken serum and the soluble fractionof chicken peripheral nerve by applying the considerations andkinetic models described by Estevez and Vilanova (7). A usefulcommercial software which allows a 3D fit by applying user-defined equations has been used to implement kinetic modelswith the data.
Even though this paper does not deal directly with organo-phophorus-induced delayed polyneuropathy (OPIDP) and doesnot study neuropathy target esterase (NTE), chicken tissues werechosen as the source of biological material and phenyl valerate(PV) as the substrate because there are extensive studies intochicken PVases (brain, spinal cord, peripheral nerve, liver, ...)using paraoxon to eliminate (inhibit) nonrelevant esterases forthe induction of OPIDP because it is neither an inducer of suchneurotoxicity nor an inhibitor of NTE (2, 3, 6, 8).
The model equations are used by means of the 3D fitaccording to the best kinetic model that can explain the differentkinetic phenomena observed, and these are employed to (1)discriminate PVase activity components in the soluble fractionof peripheral nerve searching for permanently inhibited (non-spontaneously reactivated) PVases, which should be discardedas neurotoxicity targets; (2) confirm that the use of complexequations by means of a 3D fit according to the best kineticmodel is both applicable and useful in other enzymatic systemssuch as PVases of chicken serum; and (3) compare the kineticcomponents discriminated within chicken serum PVases withthose in peripheral nerve soluble PVases as the former havebeen suggested to mirror the response to inhibitors of the latter(9).
Materials and Methods
2.1. Chemicals. O,O′-Diethyl p-nitrophenyl phosphate (paraox-on, D9286, total impurities e 10%) was obtained from Sigma-Aldrich S.A. (Madrid, Spain), and phenylvalerate was purchasedfrom Lark Enterprise (Webster, MA). A stock solution of 10 mMparaoxon was prepared in acetone and dissolved in 50 mM Tris-HCl buffer (pH 8.0) containing 1 mM EDTA immediately beforethe kinetic assays; the final acetone concentration was <0.001% in
Figure 1. Reaction of inhibition (i), reactivation (r), and aging (a) of esterases by organophosphorus compounds. This scheme assumes that theformation of a Michaelis-type intermediate complex in the inhibition reaction does not significantly affect the kinetics of the reaction because thelow solubility of the organophosphorus compounds does not allow the concentrations to cause significant saturation (1).
Figure 2. Typical time course in an inhibition experiment. Enzyme preparation is treated with an inhibitor (paraoxon) (first vertical arrow) and ispreincubated during the “inhibition time” (ti) (continuous horizontal line). Then a substrate is added (PV) (second vertical arrow) and incubatedduring the “substrate time” (ts) (dashed horizontal line) to allow the substrate-enzyme reaction to measure the residual active enzyme (mark of theinhibition). Finally, the reaction with the substrate is stopped (third vertical arrow) by adding SDS, and a color reagent is added before absorbanceis measured. The absorbance values are referred to as the absorbance of the enzyme preparation without an inhibitor. The addition of a substrateusually stops inhibition due to dilution and competitive substrate protection.
136 Chem. Res. Toxicol., Vol. 24, No. 1, 2011 EsteVez et al.

volume. A stock solution of substrate phenylvalerate (16.8 mM)was prepared in dried N,N-dimethylformamide and diluted in waterto 0.54 mM immediately before the enzymatic assays. All of theother chemicals were of analytical reagent grades. Sodium dodecylsulfate (SDS)/4-aminoantipyrine (74.5 mM/1.23 mM) was preparedin water and stored in a dark bottle. Potassium hexacyanoferrate(III)(1.21 mM) was prepared in water.
2.2. Preparation of the Chicken Soluble Fraction PeripheralNerve. A soluble fraction of sciatic nerves of chicken legs wasprepared as described by Estevez et al. (6). In short, hen sciaticnerves were obtained from a commercial slaughtering house andkept in cold (0-5 °C) Tris buffer until use (before 1 h). The tissuewas cleaned of fat and blood. The sciatic nerve was cut into 5 mmpieces and homogenized with a Polytron homogenizer (KinematicaGmbH, Germany) using a PTA 10s head at 70% speed (3 times ×30 s) in Tris buffer at a concentration of 200 mg of fresh tissue/mL. Then homogenization particulate and soluble fractions wereseparated by ultracentrifugation at 100000g for 60 min at a 200mg concentration of fresh tissue per milliliter in 50 mM Tris-HClbuffer (pH 8.0) containing 1 mM EDTA. For the inhibition process,this fraction was either concentrated or diluted with buffer (1:20)to obtain the desired concentration.
2.3. Standard Phenylvalerate Esterase Activity (PVase) As-say. Enzyme assays were carried out as previously described (6)following a procedure based on the colorimetric method for theNTE assay, as developed by Johnson (12), using phenylvalerate(PV) as a substrate. One milliliter of tissue preparation and 0.1mL of inhibitor (buffer in controls) at the appropriate concentrationwere preincubated for the corresponding time and then incubatedwith 1 mL of substrate (phenylvalerate ) 0.54 mM) for 10 min.The reaction was stopped by adding 1 mL of a solution of 2%SDS with 1.23 mM aminoantipyrine and 0.5 mL of 1.21 mMpotassium ferrycyanide. After 10 min, the absorbance was read at510 nm. Blanks and spontaneous hydrolysis controls (sampleswithout tissue) were included in the same procedure.
2.4. Inhibition Procedure. The paraoxon inhibition kinetic datawere analyzed in a peripheral nerve soluble fraction at 37 °C. A 1mL volume of peripheral nerve soluble fraction (at an equivalentconcentration of 20 mg mL-1 of original fresh tissue) was incubatedwith 100 µL of paraoxon to reach the indicated paraoxonconcentrations at 37 °C for the indicated inhibition time (see Figure3). Then, the substrate was added and incubated for 10 min tomeasure residual enzymatic activity, following the proceduredescribed by Johnson (12) as modified in ref 6. The residualconcentration of paraoxon in the presence of PV was 0.52 of theinitial concentration. The reaction was stopped, and the results wereexpressed as percent of activity (E/E0 × 100) over the controlwithout an inhibitor and were plotted versus time (t) for each dataset for the respective inhibitor concentration.
2.5. Reactivation Kinetics after Removal of Paraoxon byUltrafiltration. For the inhibition and removal procedure, a 1.5mL volume containing the soluble fraction corresponding to 200mg of fresh tissue/mL was incubated with 1 µM paraoxon for 30min at 37 °C in a total volume of 1.65 mL. Controls were incubatedwith only buffer, and the sample was incubated with paraoxon.Both the noninhibited control and the preinhibited samples werediluted to 15 mL with cold buffer to be then subjected to two washesthrough Millipore ultrafree-15 biomax 50 K (15 mL) filters bycentrifuging at 1500-2000g and 4 °C. Centrifugation continueduntil the sample volume reduced to 0.15 mL. Then concentrateswere diluted to 15 mL with cold buffer and filtered as before. Atthe end of the process, buffer was added to obtain the sampleconcentration used in the PVase assay (soluble fraction from 20mg of fresh tissue/mL). The lost PVase activity in the process wasaround 10-20%. The residual paraoxon concentration was negli-gible (from the above-indicated volumes, estimated to be about 0.01nM) and was, therefore, unable to significantly inhibit the remainingenzyme activity. This residual concentration was estimated fromtwo concentration steps at 0.15 mL and three dilution steps at 15mL. At the end of the first step (one dilution at 15 mL, oneconcentration at 0.15 mL, and then one dilution at 15 mL), the
residual concentration was estimated to be 1.1 nM and to be 0.01nM at the end of second step (one concentration at 0.15 mL andthen one dilution at 15 mL).
For reactivation, after dilution, the ultrafiltered samples wereincubated at 37 °C, and 1 mL aliquots were taken at 0, 30, 60,125, and 180 min to allow for reactivation. An overnight time wastaken to allow for reactivation in an additional experiment underthe same experimental conditions. PVase activity was assayed asdescribed above. The results are expressed as percent of activity(E/E0 × 100) over the control without the inhibitor.
2.6. Computerized Estimation of the Kinetic Parameters.Model equations were fitted to the experimental inhibition kineticdata by a nonlinear computerized method based on the least-squaresprinciple using Sigma Plot software, versions 6 and 8. Equationswere applied and fits were fulfilled according to the software user’sguide.
2.7. Model Equation for Inhibition and the ReactivationProcess. The model equations were applied according to theinstructions included in Estevez and Vilanova (7).
In short, the formation of a reversible noncovalent Michaelis-like intermediate is not considered in this paper as the low solubilityof the inhibitor does not allow the concentrations to cause significantsaturation(1).ThereactionoftheinhibitionwithouttheMichaelis-Mentencomplex may be expressed as follows:
The model equation deduced to describe the inhibition andreactivation mechanisms for a system with several enzymaticcomponents is described in Estevez and Vilanova (7). In short, theexpected kinetic equation is as follows:
In a biological preparation, esterase activity may contain severalsensitive enzymatic fractions (E1, E2, E3, etc.), as well as a resistant
Figure 3. Kinetics of the time-progressive inhibition of phenylvalerateesterase activity (PVase) from the soluble fraction of the peripheralnerve at different paraoxon concentrations. PVase activity was assayedas described under Materials and Methods using a preparation contain-ing the soluble fraction of 17 mg of fresh tissue/mL that waspreincubated with 0, 3, 10, 50, or 200 nM paraoxon (upper to lowerplots) for the indicated time. Then the enzymatic activity was assayedwith phenylvalerate for 10 min. Percentages refer to the activity of thesamples preincubated with 0 nM paraoxon at each time. Each curvewas fitted with exponential model equations by selecting the best-fittingequation according to the F test. The inset is an enlargement of thefigure and shows inhibition at 0 min of inhibition time (ongoinginhibition).
E + PX98ki
E - P + X (I)
E ) E0 e-ki·PX·t (1a)
E ) E0 e-ki·I·t (PX ≡ I) (1b)
Paraoxon Model of Inhibition Chem. Res. Toxicol., Vol. 24, No. 1, 2011 137

component (ER). For example, given a situation of two sensitiveenzymatic fractions (exponential components) and a resistantcomponent, the model equation should be expressed as
where k1 and k2 are the second-order rate constants; E10 and E20
are the amplitude or proportion of the enzymatic sensitive com-ponents at the initial time; a1 and a2 are the parameters involvingthe rate constant and the inhibitor concentration (a1 ) k1I and a2
) k2I); and ER is the resistant component.In a simple classical irreversible inhibition mechanism by
organophosphorylation (reaction I, an exponential decay (eq 1a) isexpected with lines of E versus t lowering to the same residuallevel. The rate of decay would depend on the inhibitor concentration(PX or I), but the long-term tendency should be at the same residuallevel. This tendency is expected to be zero if all of the activity issensitive or to be at a residual level if a fraction of the activity isresistant to the inhibitor.
However, when a spontaneous reactivation simultaneously occurs,the reaction to be considered is the following:
For a single enzyme component that reactivates, the deduced modelequations are (7)
variation of EP as a function of t: dEP/dt ) a ·E - kr ·EP
The solution of this system of equations when t ) 0, E(t ) 0) )100, and EP(t ) 0) ) 0, as the initial conditions, is as follows:
where a is the parameter that includes the inhibitor concentrationand the inhibition rate constant: a ) kiI.
This model has been extended to consider not only a multien-zymatic system with a significant spontaneous reactivation of theinhibited enzyme but also the possible interference, which shouldbe the most sensitive component undergoing additional ongoinginhibition during the substrate reactions according to Estevez andVilanova (7). These additional considerations are provided in detailunder Results.
3. Results
3.1. Paraoxon Inhibition Curves in the Peripheral NerveSoluble Fraction. Inhibition of peripheral nerve soluble PVaseactivity with paraoxon shows a time-progressive inhibition,which is coherent with covalent irreversible inactivation. Figure3 presents the curves corresponding to the best fit according tothe F test of the individual curve for each paraoxon concentra-tion used. For a long inhibition time, curves do not converge atthe same residual activity point, but show a parallel linestendency at a level depending on the inhibition concentrationbeing used.
Although each curve apparently shows a good, graphical fitwith the experimental data, no consistent values for the kineticconstants, numbers, and the proportion of the components havebeen obtained. Therefore, the data cannot be interpreted withexponential decay models as they only consider the phospho-rylation reaction. The introduction of two exponential compo-
nents and/or a resistant component did not improve theconsistency of the results and their properties. The number ofsensitive components depends on the inhibition concentrationand not on the remaining constant. The first-order constantsobtained in the inhibition curves are not linearly dependent onthe paraoxon concentration, and the values of the second-orderkinetic constants are not consistent.
One possible interpretation of these observations is theexistence of a spontaneous simultaneous reactivation (7), whichwas confirmed in the experiment described in the next section(3.2).
Extrapolating the data in Figure 3 to the preincubation zerotime did not converge to 100% (see the inset zoomed graph).This indicates that ongoing inhibition during substrate incubationunder the assayed conditions is apparently significant, at leastfor the highest inhibitor concentrations (50 and 200 nM).Therefore, ongoing inhibition during substrate incubation needsto be considered in the model, at least the most sensitivecomponents (7, 8). This effect is not highly significant as it isonly approximately 10% of the highest inhibitor concentration;however, it could hinder the data analysis if not considered.Accordingly, a data analysis was done by introducing thespontaneous reactivation effect and ongoing inhibition, whichare presented in section 3.3.
3.2. Spontaneous Reactivation of Paraoxon-Inhibited Es-terases in the Peripheral Nerve Soluble Fraction. Figure 4shows the spontaneous reactivation in the peripheral nervesoluble fraction preinhibited with paraoxon once the inhibitorhad been removed by ultrafiltration. The activity shows a time-progressive reactivation up to a level of about 60% at around180 min. Therefore, this confirms that a spontaneous reactivationtakes place, which should be considered in the kinetic model.An additional reactivation experiment reveals reactivation upto a level of about 100% once the inhibitor had been removedby ultrafiltration and was allowed to reactivate overnight. Thisfull spontaneous reactivation suggests that the aging reactionmay be negligible under these experimental conditions. There-
E ) E10 e-k1·I·t + E20 e-k2·I·t + ER (2a)
E ) E10 e-a1·t + E20 e-a2·t + ER (2b)
E + PX98ki
-XE - P98
krE + P (II)
variation of E as a function of t: dE/dt ) -a ·E + kr ·EP
E ) [(kr ·E0)/(a + kr)] + [(a ·E0)/(a + kr)] e-(a+kr)·t
(3)
Figure 4. Kinetics of the spontaneous reactivation of phenylvalerateesterase in a soluble fraction of the peripheral nerve after inhibition byparaoxon. Three independent preparations were inhibited by preincu-bation with 1 µM paraoxon for 30 min, which were then cooled anddiluted. Then the inhibitor was removed by two washing ultrafiltrationand dilution cycles to be then incubated at 37 °C for 150 min forreactivation. During the reactivation period, activity was monitored bymeasuring the PVase activity in aliquot samples taken at 0, 30, 60,125, and 180 min. Around 100% recovery of activity was observed inan additional experiment under the same conditions, where activity wasmonitored by measuring only the PVase activity in the aliquot samplestaken overnight.
138 Chem. Res. Toxicol., Vol. 24, No. 1, 2011 EsteVez et al.

fore, aging was not considered in the model equations represent-ing the kinetic behavior of this system.
3.3. Kinetic Model Considering Ongoing Inhibition andthe Simultaneous Spontaneous Reactivation Process in thePeripheral Nerve Soluble Fraction. Study of the InhibitionExperiment with Simultaneous Reactivation. The profile ofthe inhibition curve in Figure 3 and the evidence of reactivation(Figure 4) both suggest that a complex kinetic model shouldbe used to fit the data. This model should include both inhibitionand the reactivation process for several enzymatic components,as well as the ongoing inhibition effect.
It is reasonable to assume that ongoing inhibition is significantonly for the most sensitive enzymatic component. Therefore,an additional exponential factor may be applied to the mostsensitive component to correct the effect (7, 8). This factor ise-ka′·I and should be added in the most sensitive component (E1)equation (see eq 4). Parameter ka′ represents the apparent first-order kinetic constant of the observed inhibition withoutpreincubation caused by inhibition during the substrate reactiontime applied to measure activity.
The model that considers inhibition with simultaneousspontaneous reactivation (see Materials and Methods) was usedfor one, two, or three enzymatic components by consideringthe factor for the ongoing inhibition effect in the most sensitivecomponent. The best-fitting model (according to the F test)consisted of three enzymatic entities and was inhibited andspontaneously reactivated. According to Estevez and Vilanova(7), this mathematic model is
where k1, k2, and k3 are the inhibition constants; kr1, kr2, and kr3
are the reactivation constants; and E10, E20, and E30 are theproportion (amplitude) of enzymatic components E1, E2, E3,respectively, where enzymatic components E1, E2, and E3 wereinhibited and spontaneously reactivated.
For the most sensitive enzymatic component, the exponentialfactor (e- ka′·I) was included to correct the ongoing inhibitionduring the substrate reaction, where ka′ is the exponentialconstant of the ongoing inhibition (7, 8). This exponential factoraffects the most sensitive enzymatic component.
For the purpose of obtaining a coherent solution in theinteractive computing estimation, some restrictions were applied:(1) all of the parameters (rate constants and amplitudes) shouldhave positive values (>0); (2) component 1 is the most sensitive,so k1 > k2 and k2 > k3. (3) The following complementaryrestriction was also applied: E10 + E20 + E30 ) 100%. A 3Dfitting (percent of phenylvalerate esterase activity versus t andI) was done with the data described in Figure 3. The results areprovided in Table 1 (line A), and the deduced 3D surface isplotted in Figure 5. The system allows more than one solution,although this depends on the initial value in the interactivecomputing estimation. The results were accepted only if thereactivation constants were of the same order of magnitude asin the reactivation experiment, which is shown later. Theconsistency of the results improved if a correction factor forthe ongoing inhibition during the substrate reaction was includedin the most sensitive component.
The I50(30 min) values for each component (Ei) were obtainedby approximation and by applying the equation
Table 1. Kinetic Constants (ki) and the Proportions of Components Obtained from the Different Inhibition and/or ReactivationExperiments with Paraoxon on Soluble Fractions of Peripheral Nervea
expt E1 (%)k1
(nM-1 min-1)k1r
(min-1)I50
30
(nM) E2 (%)k2
(nM-1 min-1)k2r
(min-1)I50
30
(nM) E3 (%)k3
(nM-1 min-1)k3r
(min-1)I50
30
(nM)
Ab (37) 1.798 0.428 0.24 (41) 0.0051 0.0110 6 (22) 3.5 × 10-5 0.0060 740
B ne (1 + 33)c 0.0092 (35)ne (1 + 33)c 0.0092 (36)ne (1 + 31)c 0.0116 (36)
C (1 + 46)d 1.468 0.387 0.26 (17 + 26)c 0.0027 0.0190 12 (10)
a The I50 values were calculated from the kinetic constants for each component. (A) Experiment for time-progressive inhibition with five differentinhibitor concentrations; (B) reactivation after ultrafiltration; (C) reactivation experiment after diluting. The R2 coefficients were 0.9928 (expt A); 0.9740,0.9724, and 0.9847 (expt B); and 0.9956 (expt C). b The apparent first-order ongoing inhibition constant (ka′) ) 0.0015 nM-1. c These values correspondto E20 + EI20 ) E2 (the free initial enzyme plus the initial inhibited enzyme) of component 2 (ne ) not estimated). d This value corresponds to E10 +EI10 ) E1 (the free initial enzyme plus the initial inhibited enzyme) of component 1.
E ) [e-ka′·I] · {[(kr1 ·E10)/(k1 · I + kr1)] +
[(k1 · I ·E10)/(k1 · I + kr1)] e-(k1·I+kr1)·t} +[(kr2 ·E20)/(k2 · I + kr2)] +
[(k2 · I ·E20)/(k2 · I + kr2)] e-(k2·I+kr2)·t +[(kr3 ·E30)/(k3 · I + kr3)] +
[(k3 · I ·E30)/(kr3 · I + kr3)] e-(k3·I+kr3)·t
(4)
Figure 5. Representation of the inhibition kinetics of soluble peripheralnerve PVases by paraoxon. The inhibitory surface obtained by fittingthe 3D model equation to the data corresponding to paraxon inhibitiondone in a soluble fraction of the peripheral nerve is shown in Figure 3.The surface reflects the result of the best model according to the Ftest. It corresponds to a model with three enzymatic components thatare simultaneously inhibited and reactivated. Details are provided underMaterials and Methods.
% activity (Ei) ) [(kr ·100)/(ki · I + kr)] +
[(ki·I·100)/(ki·I + kr)] · e-(ki·I+kr)·30 (5)
Paraoxon Model of Inhibition Chem. Res. Toxicol., Vol. 24, No. 1, 2011 139

for the previously estimated kinetic constants and by thencarrying out successive iterations with different I values in anelectronic spreadsheet to obtain the I value for the percent ofactivity equal to 50 ( 0.1%.
Spontaneous Reactivation Constants Obtained from Dilu-tion Experiments. The data reported by Barril et al. (3) in aninhibition experiment with peripheral nerve soluble PVaseactivity with paraoxon, followed by spontaneous reactivationafter dilution, were used to estimate the reactivation andinhibition kinetic constants. In this experiment, samples wereinhibited with 10 nM paraoxon to be then diluted to around 0.5nM paraoxon. The recovery level was similar to the activityexpected if inhibition with 0.5 nM paraoxon was applied in theinhibition experiment shown in Figure 3.
The reactions to be considered are as follows:
The differential equations associated with these reactions are(7)
variation of EP as a function of t: dEP/dt ) a · E - kr ·EP
When the initial conditions were t ) 0 min, E(t ) 0) ) E0
and EP(t ) 0) ) EP0, the solution of this system of equationsis as follows:
Mathematical models were applied and corresponded to one,two, or more inhibited enzymes, which spontaneously reacti-vated. The initial concentrations of the active enzymes and theresidual concentration of the inhibitor were considered signifi-cant. The corresponding model described by Estevez andVilanova (7) was applied as follows:
where A ) E1P0 + E10 ) total concentration of E1 and B )E2P0 + E20 ) total concentration of E2. The followingrestrictions were applied: A + B + R ) 100% and k1 > 1. Therestriction k1 > 1 nM-1 min-1 forces the estimate of the second-order rate kinetic constant of the most sensitive component.
This mathematical model corresponds to two sensitivecomponents capable of spontaneously reactivating, and aresistant component (R). This R component is probably theequivalent to the low sensitive component E3 detected in theinhibition experiment. The system estimated it as a resistantcomponent because the inhibitor concentration was lower to thatneeded to cause significant inhibition. The resulting kineticparameters and I50
30 values are shown in Table 1, line B.
Study of the Reactivation Progress Experiment afterRemoval of the Inhibitor by Ultrafiltration. The enzyme wasinhibited and the inhibitor was thoroughly removed after severalultrafiltration steps (see Materials and Methods). The inhibitorconcentration was practically I ) 0, then a ) 0. The diagramshowing this process is as follows:
and the resulting kinetic equation for this reaction (7) is
where ET ) EP0 + E0 ) total enzyme concentration ) 100%.The mathematical models corresponding to the one, two, or
more inhibited enzymes that had spontaneously reactivated wereused. The best fit according to the F test was
where kr is the reactivation constant, E0 is the proportion(amplitude) of the initial active enzymatic component, EP0 isthe proportion (amplitude) of the initial inhibited enzymaticcomponent, and R is the enzymatic fraction resistant toinhibition.
The following constraint was applied: E0 + EP0 + R ) 100%.The deduced kinetic parameter values are shown in Table 1,
line C, and the curve is plotted in Figure 4.The inhibited nonultrafiltered controls showed 17% PVase
activity.Consistency between the Kinetic Behavior in Inhibition
Experiments and in Reactivation Experiments. The consis-tency of the estimated parameters was checked by comparingthe results of the inhibition experiments with those obtainedwith the reactivation experiments. The kinetic model for thesesituations was the same, but the starting conditions differed.Therefore, different mathematical equations were applied. Thecomparison made is presented in Table 1, lines A-C.
3.4. Kinetic Inhibition Model with a Simultaneous Spon-taneous Reactivation Process in Chicken Serum. Analysisof the Data from the Inhibition Experiment ConsideringSimultaneous Reactivation. The data reported in Garcıa-Perezet al. (9), that is, in the inhibition experiment of chicken serumsoluble PVase activity with paraoxon, were used to estimatethe kinetic constants. Spontaneous reactivation and ongoinginhibition were considered.
The profile of the curves in the time-progressive inhibition(parallel lines for a long time) suggests that spontaneousreactivation occurred simultaneously.
Extrapolating the data in this experiment to the preincubationzero time did not converge to 100%. This indicates that theongoing inhibition during the substrate incubation is apparentlysignificant under the assayed conditions. Therefore, the ongoinginhibition during substrate incubation needs to be considered.
The kinetic inhibition model with simultaneous spontaneousreactivation was applied to the inhibition data by consideringmodel equations with one, two, or three exponential componentswith or without a constant component and by also taking intoaccount the factor for the ongoing inhibition effect in the mostsensitive component. The F test was applied. The best-fittingmodel consisted of three enzymatic entities, two of which wereinhibited and spontaneously reactivated, whereas one waspermanently inhibited. This mathematical model is
E - P98kr
E + P
E + PX98ki
E - P + X
variation of E as a function of t: dE/dt ) -a · E + kr ·EP
E ) (EP0 + E0) ·kr/(k · I + kr) +
[E0 - (EP0 + E0) ·kr/(k · I + kr)] · e-(k·I+kr)·t
E ) A · k1r/(k1 · I + k1r) +
[E10 - A · k1r/(k1 · I + k1r)] · e-(k1·I+k1r)·t +
B · k2r/(k2I + k2r) +
[E20 - B · k2r/(k2I + k2r)] · e-(k2·I+k2r)·t + R
(6)
EP98kr
E + P (III)
E ) ET - EP0 · e-kr·t (7)
E ) (E0 + EP0) - EP0 · e-kr·t + R (8)
140 Chem. Res. Toxicol., Vol. 24, No. 1, 2011 EsteVez et al.
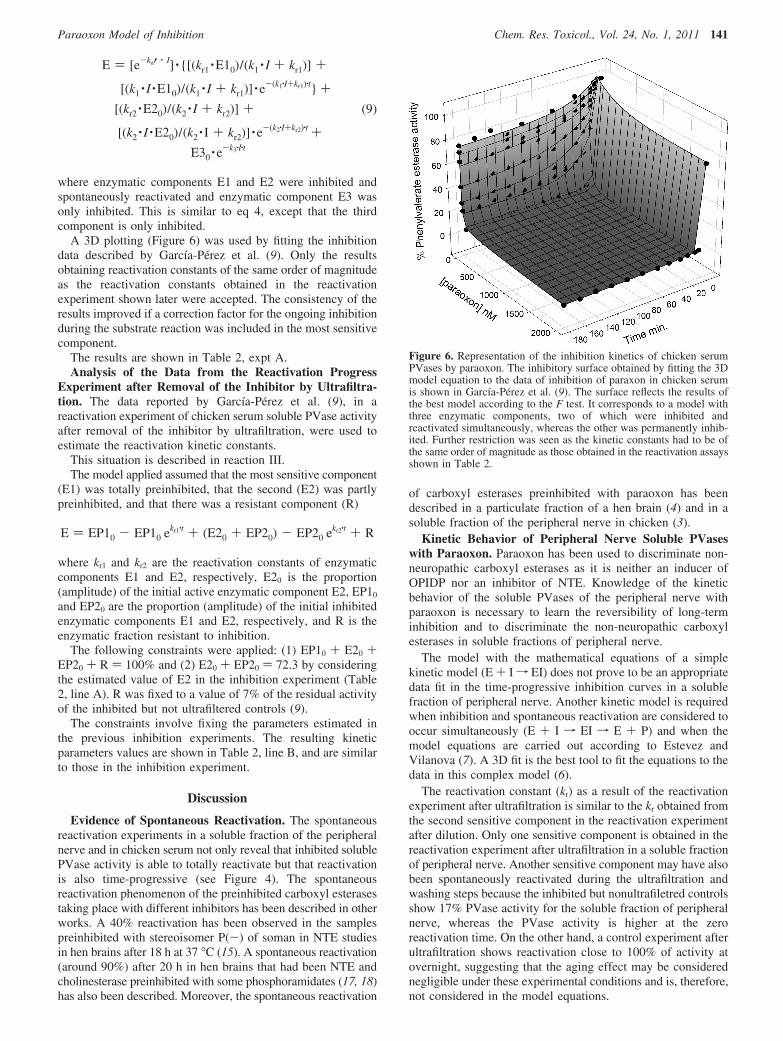
where enzymatic components E1 and E2 were inhibited andspontaneously reactivated and enzymatic component E3 wasonly inhibited. This is similar to eq 4, except that the thirdcomponent is only inhibited.
A 3D plotting (Figure 6) was used by fitting the inhibitiondata described by Garcıa-Perez et al. (9). Only the resultsobtaining reactivation constants of the same order of magnitudeas the reactivation constants obtained in the reactivationexperiment shown later were accepted. The consistency of theresults improved if a correction factor for the ongoing inhibitionduring the substrate reaction was included in the most sensitivecomponent.
The results are shown in Table 2, expt A.Analysis of the Data from the Reactivation Progress
Experiment after Removal of the Inhibitor by Ultrafiltra-tion. The data reported by Garcıa-Perez et al. (9), in areactivation experiment of chicken serum soluble PVase activityafter removal of the inhibitor by ultrafiltration, were used toestimate the reactivation kinetic constants.
This situation is described in reaction III.The model applied assumed that the most sensitive component
(E1) was totally preinhibited, that the second (E2) was partlypreinhibited, and that there was a resistant component (R)
where kr1 and kr2 are the reactivation constants of enzymaticcomponents E1 and E2, respectively, E20 is the proportion(amplitude) of the initial active enzymatic component E2, EP10
and EP20 are the proportion (amplitude) of the initial inhibitedenzymatic components E1 and E2, respectively, and R is theenzymatic fraction resistant to inhibition.
The following constraints were applied: (1) EP10 + E20 +EP20 + R ) 100% and (2) E20 + EP20 ) 72.3 by consideringthe estimated value of E2 in the inhibition experiment (Table2, line A). R was fixed to a value of 7% of the residual activityof the inhibited but not ultrafiltered controls (9).
The constraints involve fixing the parameters estimated inthe previous inhibition experiments. The resulting kineticparameters values are shown in Table 2, line B, and are similarto those in the inhibition experiment.
Discussion
Evidence of Spontaneous Reactivation. The spontaneousreactivation experiments in a soluble fraction of the peripheralnerve and in chicken serum not only reveal that inhibited solublePVase activity is able to totally reactivate but that reactivationis also time-progressive (see Figure 4). The spontaneousreactivation phenomenon of the preinhibited carboxyl esterasestaking place with different inhibitors has been described in otherworks. A 40% reactivation has been observed in the samplespreinhibited with stereoisomer P(-) of soman in NTE studiesin hen brains after 18 h at 37 °C (15). A spontaneous reactivation(around 90%) after 20 h in hen brains that had been NTE andcholinesterase preinhibited with some phosphoramidates (17, 18)has also been described. Moreover, the spontaneous reactivation
of carboxyl esterases preinhibited with paraoxon has beendescribed in a particulate fraction of a hen brain (4) and in asoluble fraction of the peripheral nerve in chicken (3).
Kinetic Behavior of Peripheral Nerve Soluble PVaseswith Paraoxon. Paraoxon has been used to discriminate non-neuropathic carboxyl esterases as it is neither an inducer ofOPIDP nor an inhibitor of NTE. Knowledge of the kineticbehavior of the soluble PVases of the peripheral nerve withparaoxon is necessary to learn the reversibility of long-terminhibition and to discriminate the non-neuropathic carboxylesterases in soluble fractions of peripheral nerve.
The model with the mathematical equations of a simplekinetic model (E + If EI) does not prove to be an appropriatedata fit in the time-progressive inhibition curves in a solublefraction of peripheral nerve. Another kinetic model is requiredwhen inhibition and spontaneous reactivation are considered tooccur simultaneously (E + I f EI f E + P) and when themodel equations are carried out according to Estevez andVilanova (7). A 3D fit is the best tool to fit the equations to thedata in this complex model (6).
The reactivation constant (kr) as a result of the reactivationexperiment after ultrafiltration is similar to the kr obtained fromthe second sensitive component in the reactivation experimentafter dilution. Only one sensitive component is obtained in thereactivation experiment after ultrafiltration in a soluble fractionof peripheral nerve. Another sensitive component may have alsobeen spontaneously reactivated during the ultrafiltration andwashing steps because the inhibited but nonultrafiletred controlsshow 17% PVase activity for the soluble fraction of peripheralnerve, whereas the PVase activity is higher at the zeroreactivation time. On the other hand, a control experiment afterultrafiltration shows reactivation close to 100% of activity atovernight, suggesting that the aging effect may be considerednegligible under these experimental conditions and is, therefore,not considered in the model equations.
E ) [e-ka′ · I] · {[(kr1 ·E10)/(k1 · I + kr1)] +
[(k1 · I ·E10)/(k1 · I + kr1)] · e-(k1·I+kr1)·t} +
[(kr2 ·E20)/(k2 · I + kr2)] +
[(k2 · I ·E20)/(k2 · I + kr2)] · e-(k2·I+kr2)·t +
E30 · e-k3·I·t
(9)
E ) EP10 - EP10 ekr1·t + (E20 + EP20) - EP20 ekr2·t + R
Figure 6. Representation of the inhibition kinetics of chicken serumPVases by paraoxon. The inhibitory surface obtained by fitting the 3Dmodel equation to the data of inhibition of paraxon in chicken serumis shown in Garcıa-Perez et al. (9). The surface reflects the results ofthe best model according to the F test. It corresponds to a model withthree enzymatic components, two of which were inhibited andreactivated simultaneously, whereas the other was permanently inhib-ited. Further restriction was seen as the kinetic constants had to be ofthe same order of magnitude as those obtained in the reactivation assaysshown in Table 2.
Paraoxon Model of Inhibition Chem. Res. Toxicol., Vol. 24, No. 1, 2011 141

The 3D fit enables all of the data in the same fit to be includedsimultaneously. The amplitudes (proportion) of the obtainedcomponents by 3D fitting in inhibition data are similar to thoseobtained in the inhibition experiments with mipafox (Table 3).The outcomes indicate that the PVases with paraoxon behavein the same way as three different enzymatic entities. Table 1shows how both the inhibition constants (and the correspondingI50
30 values) and the reactivation constants are consistent withthose obtained in the reactivation experiments after eitherdilution or ultrafiltration. These observations may be consideredan internal validation of the applied kinetic model and the 3Dfit.
It is concluded that a soluble fraction of peripheral nervecontains two paraoxon-sensitive components of 37 and 41%(I50) 0.24 and 6 nM, respectively, for 30 min) and that 22% oftotal activity is resistant to the highest tested concentration (200nM). The same number and proportion of components areobserved in the same tissue by inhibition with mipafox (Table3).
However, the relative sensitivity of the time-progressiveinhibition for mipafox and paraoxon differs (Table 3). The firstcomponent is the most sensitive for paraoxon, whereas thesecond component is the most sensitive component for mipafox.In any case, both components are very sensitive enzymes (I50
on the order of nanomolar) if compared with other esterases(i.e., neuropathy target esterases bound to brain membranes),with I50 for 30 min on the order of micromolar (5, 11, 14, 19).
Finally, the consistency of the results obtained in theexperiments done with paraoxon and mipafox in both tissuesmay be considered an internal validation of the strategies tocharacterize kinetic behavior.
Model Equation with Spontaneous Reactivation and a3D Fit Applied to the Data from Chicken Serum PVaseswith Paraoxon. The data from the chicken serum PVases (9)were reviewed to corroborate the applicability of the obtainedequations from the different kinetic mechanisms and the 3Dfit. No kinetic inhibition constants were provided in the citedwork.
A similar consistency of the results, as in peripheral nervesoluble PVases with paraoxon, is obtained. The reactivationconstants (kr1 and kr2), as a result of the reactivation experimentafter ultrafiltration, are similar to those obtained in the inhibitionexperiment. In addition, three components have been obtainedwith proportions similar to those in the inhibition experimentwith mipafox at a fixed time (Table 3) (8).
However, the relative sensitivities of the time-progressiveinhibitions for mipafox and paraoxon differ (Table 3). The firstmost sensitive component (E1, 21-22%) is the most sensitivefor paraoxon, whereas the second most sensitive component (E2,72-75%) is the most sensitive component for mipafox. In anycase, both components are very sensitive enzymes (I50 on theorder of nanomolar). With both inhibitors, a minor resistant (I50
> 1000 nM) component of about 5-6% was also observed. Theconsistency of the results obtained in the experiments withparaoxon and mipafox may be considered an internal validation.
Thus, it is confirmed that using complex equations by meansof a 3D fit in accordance with the best kinetic model isapplicable and useful in other kinetic systems such as the PVasesof chicken serum.
Toxicological Meaning and Applications for MeasuringNTE. To discriminate and measure neuropathy target esterases(NTE), those esterases sensitive to paraoxon (an organophos-phate that does not induce neuropathy) are excluded as targetsof the neurotoxic process (10, 13, 16). This work proves thatthe soluble PVases of the peripheral nerve can be progressivelyinhibited by paraoxon with time, but not permanently for it tobe spontaneously reactivated. The soluble PVase activity is ableto reactivate about 65% of total activity when it is preinhibitedwith 40 µM paraoxon and when residual PVase activity is <5%throughout the procedure to obtain the P tissue (3).
The results of this work confirm that paraoxon does notpermanently inhibit the soluble PVases of the chicken peripheralnerve in the long term and that it is not the most appropriatetool to discriminate NTE and other potential targets of neuro-toxicity in a soluble fraction of the peripheral nerve. In fact,these PVases slowly hydrolyze the paraoxon and are irreversibly
Table 2. Kinetic Constants (ki) and Proportions of Components Obtained from the Different Inhibition and/or ReactivationExperiments with Paraoxon in Chicken Seruma
expt E1 (%)k1
(nM-1 min-1)k1r
(min-1) I5030 (nM) E2 (%)
k2
(nM-1 min-1)k2r
(min-1) I5030 (nM) E3 (%)
k3
(nM-1 min-1) I5030 (nM)
Ab 22.1 0.0584 0.0044 0.43 72.3 0.0020 0.0091 13.7 5.6 4.3 × 10-6 5373B 20.7 0.0021 (19.4 + 52.9)c
(fixed E20 +E2I0 ) 72.3)
0.0112 7 (fixed)
a I50 values were calculated from the kinetic constants for each component. (A) Experiment for time-progressive inhibition with eight differentinhibitors concentrations; (B) reactivation after ultrafiltration with additional restrictions. The R2 coefficients were 0.9869 (expt A) and 0.9983 (expt B).b Apparent first-order ongoing inhibition constant (ka′) ) 0.0025 nM-1. c This value corresponds to E20 + EI20 ) E2 (the free initial enzyme plus theinitial inhibited enzyme) of component 2.
Table 3. Sensitivity of the Different Components Discriminated by Inhibition with Mipafox and Paraoxona
PXN MPX
component sensitivity I50 (nM) % sensitivity I50 (nM) %
peripheral nerve soluble esterases b cI (36.6 -37%) +++ 0.24-0.26 37 ++ 69-71 36.6II (41-47.8%) ++ 6-12 41 +++ 11-12 47.8III (15.6-22%) + 740 22 - - 15.6serum esterases d eI (21-22%) +++ 0.43 22.1 ++ >100 21II (72-75%) ++ 13.7 72.3 +++ 3.6-4 74III (5-5.6%) + >2000 5.6 + .1000 5
a I50 values are indicated (for 30 min) with the proportion of the component given in %. b From the data in this paper. c From the 3D fitting in thepaper of Estevez et al. (6). d The results obtained in this paper from the time-progressive inhibition data in the paper of Garcia-Perez et al. (9). e Fromthe experiment with a fixed inhibition time in the paper of Garcia-Perez et al. (9): +++, very high; ++, high; +, low; -, resistant.
142 Chem. Res. Toxicol., Vol. 24, No. 1, 2011 EsteVez et al.

inhibited by mipafox (an organophosphate that induces neur-opathy) (6). Therefore, it should be excluded from the dif-ferential standard assay with the target tissue because all of thesoluble PVase activity is resistant to a permanent inhibition byparaoxon despite paraoxon transiently inhibiting most solubleesterase.
There have been suggestions that different mipafox concen-trations may be used to discriminate components in the nervesoluble fraction (6).
The results of this work confirm that chicken serum esterasesmay be considered a “mirror” of the soluble esterases ofperipheral nerve for monitoring studies (9) because the kineticbehaviors of the serum PVases in the presence of paraoxon andmipafox are similar to those of the soluble peripheral nervePVases and that this could prove convenient for evaluations inmammals.
In short, the toxicological meaning of the transient inhibitionof soluble esterases by paraoxon is still not fully understood.High sensitivity, accompanied by transitory inhibition withreactivation, suggests potential roles in toxicity and/or detoxi-cation during low-dose long-term exposure to organophosphoruscompounds that warrant further research. The kinetic model thatconsiders spontaneous reactivation with simultaneous inhibition(7) is appropriate to fit the data from inhibition and reactivationexperiments with the paraoxon of PVases in a soluble fractionof peripheral nerve and serum. The 3D fit is a good method toestimate the kinetic parameters in complex kinetic models withone or more sensitive components. The estimated number ofcomponents and their proportion of paraoxon-inhibited PVaseactivity are coherent with those estimated by mipafox inhibitionin both the peripheral nerve and serum. This consistency withdifferent inhibitors in two different biological preparations maybe considered a validation, thus demonstrating that the proposedstrategy and the kinetic models used are efficient tools toestimate kinetic parameters. Paraoxon is not an appropriate toolto discriminate NTE and other potential targets of neurotoxicityin a soluble fraction of the peripheral nerve given the simulta-neous spontaneous reactivation of the paraoxon-inhibited esterases.
Acknowledgment. This work has been partly supported byProject A051/2007/3.14.4 of the Spanish Ministry of Environ-ment and by institutional support from the University MiguelHernandez de Elche.
References
(1) Aldridge, W. N., and Reiner, E. (1972) Enzyme Inhibitors AsSubstrates, North-Holland Publishing, Amsterdam, The Netherlands.
(2) Barril, J., and Vilanova, E. (1997) Reversible inhibition can profoundlymislead studies on progressive inhibition of enzymes: the interactionof paraoxon with soluble neuropathy target esterase. Chem.-Biol.Interact. 108, 19–25.
(3) Barril, J., Estevez, J., Escudero, M. A., Cespedes, M. V., Nıguez, N.,Sogorb, M. A., Monroy, A., and Vilanova, E. (1999) Peripheral nervesoluble esterases are spontaneously reactivated after inhibition byparaoxon: Implication for a new definition of neuropathy targetesterase. Chem.-Biol. Interact. 119-120, 541–550.
(4) Carrington, C. D., and Abou-Donia, M. B. (1985) Paraoxon reversiblyinhibits neurotoxic esterase. Toxicol. Appl. Pharmacol. 79 (1, June15), 175–178.
(5) Escudero, M. A., Barril, J., Tormo, N., and Vilanova, E. (1995)Separation of two forms of neuropathy target esterase in the solublefraction of the hen sciatic nerve. Chem.-Biol. Interact. 97 (3, Aug18), 247–255.
(6) Estevez, J., Garcıa-Perez, A. G., Barril, J., Pellın, M., and Vilanova,E. (2004) The inhibition of the high sensitive peripheral nerve solubleesterases by mipafox: a new mathematical processing for the kineticsof inhibition of esterases by organophosphorus compounds. Toxicol.Lett. 151, 171–181.
(7) Estevez, J., and Vilanova, E. (2009) Model equations for the kineticsof covalent irreversible enzyme inhibition and spontaneous reactiva-tion: esterases and organophosphorus compounds. Crit. ReV. Toxicol.39 (5), 427–448.
(8) Estevez, J., Barril, J., and Vilanova, E. (2010) Inhibition withspontaneous reactivation and the “ongoing inhibition” effect ofesterases by biotinylated organophosphorus compounds: S9B as amodel. Chem.-Biol. Interact. 187, 397–402.
(9) Garcıa-Perez, A. G., Barril, J., Estevez, J., and Vilanova, E. (2003)Properties of phenyl valerate esterase activities from chicken serumare comparable with soluble esterases of peripheral nerves in relationwith organophosphorus compounds inhibition. Toxicol. Lett. 142 (1-2, April 30), 1–10.
(10) Johnson, M. K. (1969) The delayed neurotoxic effect of someorganophosphorus compounds. Identification of the phosphorylationsite as an esterase. Biochem. J. 114, 711–717.
(11) Johnson, M. K. (1975) Structure-activity relationships for substratesand inhibitors of hen brain neurotoxic esterase. Biochem. Pharmacol.24 (7, April 1), 797–805.
(12) Johnson, M. K. (1977) Improved assay of neurotoxic esterase forscreening organophosphates for delayed neurotoxicity potential. Arch.Toxicol. 37, 113–115.
(13) Johnson, M. K. (1982) The target for initiation of delayed neurotoxicityby organophosphorus esters: biochemical studies and toxicologicalapplications. ReV. Biochem. Toxicol. 4, 141–212.
(14) Johnson, M. K. (1982) Initiation of organophosphate-induced delayedneuropathy. NeurobehaV. Toxicol. Teratol. 4 (6, Nov-Dec), 759–765.
(15) Johnson, M. K., Read, D. J., and Benschop, H. P. (1985) Interactionof the four stereoisomers of soman (pinacolyl methylphosphonofluo-ridate) with acetylcholinesterase and neuropathy target esterase of henbrain. Biochem. Pharmacol. 34 (11, June 1), 1945–1951.
(16) Johnson, M. K. (1990) Organophosphates and delayed neuropathy-is NTE alive and well? Toxicol. Appl. Pharmacol. 102 (3, March 1),385–399.
(17) Jokanovic, M., and Johnson, M. K. (1993) Interactions in vitro of someorganophosphoramidates with neuropathy target esterase and acetyl-cholinesterase of hen brain. J. Biochem. Toxicol. 8 (1, March), 19–31.
(18) Vilanova, E., Johnson, M. K., and Vicedo, J. L. (1987) Interaction ofsome unsubstituted phosphoramidate analogs of methamidophos (O,S-dimethyl phosphorothioamidate) with acetylcholinesterase and neur-opathy target esterase of hen brain. Pestic. Biochem. Physiol. 28, 224–238.
(19) Vilanova, E., Barril, J., Carrera, V., and Pellin, M. C. (1990) Solubleand particulate forms of the organophosphorus neuropathy targetesterase in hen sciatic nerve. J. Neurochem. 55 (4), 1258–1265.
TX100346C
Paraoxon Model of Inhibition Chem. Res. Toxicol., Vol. 24, No. 1, 2011 143