influence of iron on sulfide inhibition in dark biohydrogen fermentation
TRANSCRIPT

Bioresource Technology 126 (2012) 123–130
Contents lists available at SciVerse ScienceDirect
Bioresource Technology
journal homepage: www.elsevier .com/locate /bior tech
Influence of iron on sulfide inhibition in dark biohydrogen fermentation
Bipro Ranjan Dhar a, Elsayed Elbeshbishy b,⇑, George Nakhla c
a Civil and Environmental Engineering, University of Waterloo, Waterloo, ON, Canada N2L 3G1b Trojan Technologies, London, ON, Canada N5V 4T7c Chemical and Biochemical Engineering, University of Western Ontario, London, ON, Canada N6A 5B9
h i g h l i g h t s
" Sulfide inhibition on dark biohydrogen fermentation has been studied." Low sulfide concentration (25 mg/L) significantly enhanced the hydrogen production." High sulfide concentrations (500 mg/L) completely inhibited hydrogen production." Addition of ferrous iron entirely eliminated the inhibitory effect of sulfide.
a r t i c l e i n f o
Article history:Received 17 July 2012Received in revised form 12 September2012Accepted 13 September 2012Available online 25 September 2012
Keywords:BiohydrogenDark fermentationFerrous ironSulfide inhibition
0960-8524/$ - see front matter � 2012 Elsevier Ltd. Ahttp://dx.doi.org/10.1016/j.biortech.2012.09.043
⇑ Corresponding author. Address: Trojan TechnologON, Canada N5V 4T7. Tel.: +1 519 860 3556.
E-mail addresses: [email protected], eelbetrojan.com (E. Elbeshbishy).
a b s t r a c t
Sulfide impact on biohydrogen production using dark fermentation of glucose at 37 �C was investigated.Dissolved sulfide (S2�) at a low concentration (25 mg/L) increased biohydrogen production by 54% rela-tive to the control (without iron addition). Whereas on initial dissolved S2� concentration of 500 mg/Lsignificantly inhibited the biohydrogen production with total cumulative biohydrogen decreasing by90% compared to the control (without iron addition). At sulfide concentrations of 500 mg S2�/L, additionof Fe2+ at 3–4 times the theoretical requirement to precipitate 100% of the dissolved S2� entirely elimi-nated the inhibitory effect of sulfide.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
To mitigate the worldwide increasing energy demand, scientistsare focusing on the production of energy from various renewablesources. Hydrogen is a promising alternative clean fuel for thefuture that has very high energy content (142 kJ/g) (Sinha andPandey, 2011). Hydrogen can be produced through various pro-cesses such as steam reforming of natural gas, water electrolysis,and biologically through dark fermentation, photofermentation,and photolysis. Due to the increased interest in utilization ofrenewable sources such as waste and wastewater for bio-energyproduction, biological conversion of waste to biohydrogen has re-ceived a considerable attention in recent years (Sinha and Pandey,2011; Wang and Wan, 2009). Biological hydrogen productionthrough dark fermentation is significantly influenced by various
ll rights reserved.
ies, 3020 Gore Road, London,
[email protected], selbeshbishy@
environmental and operating factors such as inoculum, substrate,pH, temperature, availability of macronutrients (N, P, S), andmicronutrients (K, Mg, Ca, Fe, Mn, Co, Cu, Mo, Zn) (Mara andHoran, 2003). If the aforementioned operating conditions, micro-nutrients, and macronutrients are not maintained within a certainrange, they may inhibit the activity of hydrogen producing micro-organisms. Thus, the impacts of the aforementioned parametershave been extensively studied to optimize biohydrogen productionas well as to explore the inhibition levels (Wang and Wan, 2009;Sinha and Pandey, 2011).
Sulfate (SO2�4 ) is a common form of sulfur in many wastes used
as feedstock for anaerobic digestion (AD) such as pulp and paperindustry and food processing industry wastes (Lens et al., 1998).During AD, sulfate can be reduced to sulfide (S2�) by sulfate reduc-ing bacteria (SRB). Additionally, sulfide can be produced throughthe anaerobic degradation of sulfur containing compounds suchas proteins. Sulfide has been identified to be toxic or inhibitoryto the fermentative microorganisms (Hwang et al., 2011; Dezhamet al., 1988). Sulfide can also reduce the bioavailability of essentialmacronutrients (trace metals) through the formation of insoluble

Table 1
124 B.R. Dhar et al. / Bioresource Technology 126 (2012) 123–130
metal sulfides (Mara and Horan, 2003). Sulfide inhibition has beenextensively studied for biological methane production (Chen et al.,2008b; Kalyuzhnyi et al., 1998; Parkin et al., 1990; McCartney andOleszkiewicz, 1993). The reported inhibitory dissolved sulfide (S2�)concentration to methanogens in anaerobic digestion is 100–800 mg/L (Chen et al., 2008a; Kalyuzhnyi et al., 1998; Parkinet al., 1990; McCartney and Oleszkiewicz, 1993). On the otherhand, most of the related research studies on the impact of sulfatereduction by SRB on dark biohydrogen fermentation were con-ducted by using sulfate-rich feedstocks (up to 20000 mg/L SO4
2�)(Lin and Chen, 2006; Chen et al., 2008a,b; Hwang et al., 2011).Since SRBs compete with hydrogen-producing microorganismsfor organic matter, the direct toxicity of sulfide to hydrogen pro-ducing bacteria has not been considered in the aforementionedstudies, and the inhibitory dissolved sulfide concentration rangefor hydrogen producing bacteria is largely unknown.
Fermentative hydrogen producing microorganisms requiretrace concentrations of various metal ions for their metabolism(Wang and Wan, 2009). Among the various metal ions, Fe2+ isthe most widely researched in dark biohydrogen fermentation, asiron is an important component of hydrogenase and other enzymesrequired for biohydrogen production (Nicolet et al., 2002). Iron isalso a very effective heavy metal for sulfide control in anaerobicdigestion through precipitation (Nielsen et al., 2008; Lens et al.,1998). Additionally, iron-sulfur clusters are essential cofactors ofproteins (Sinha and Pandey, 2011). However, precipitation ofsulfide by iron as ferrous sulfide (FeS) may diminish the impactof adding iron by limiting its bioavailability to the fermentativemicroorganisms (Lens et al., 1998). The impact of iron on darkfermentation has been studied by many researchers within a widerange of iron concentration from 0 to 14,000 mg/L (Wang and Wan,2009; Sinha and Pandey, 2011). Hwang et al. (2009) have observedincreased hydrogen production for iron concentrations raging from580 to 2900 mg/L at high sulfate concentrations (1000–5000mg/L). However, the impact of iron in high sulfidogenic dark fer-mentation systems apparently has not been demonstrated to date.
The main purpose of this work was to explore the dissolved S2�
inhibition levels for hydrogen producing bacteria as well as toinvestigate the feasibility of ferrous iron addition for sulfide inhibi-tion control in dark biohydrogen fermentation. The inhibitoryeffect of sulfide was examined in terms of biohydrogen productionin batch tests. Furthermore, different doses of Fe2+ (as ferrouschloride) were added based on the theoretical requirement ofFe2+ to precipitate dissolved sulfide (S2�) to investigate whetherthe addition of ferrous iron is effective to neutralize the sulfideinhibition of biohydrogen production.
Summary of experimental setup.
Set S2� (mg/L) Fe2+ (mg/L)
1 – –2 25 –3 50 –4 100 –5 250 –6 500 –7 – 258 – 509 – 10010 – 25011 – 50012 50 87.513 50 17514 50 35015 100 17516 100 35017 100 70018 500 87519 500 175020 500 3500
2. Methods
2.1. Seed and substrate
Anaerobic digester sludge (ADS) was collected from a conven-tional anaerobic digester at the St. Mary’s wastewater treatmentplant (St. Mary’s, Ontario, Canada) and used as seed sludge afterultrasonication, using a lab scale ultrasound device (ModelVCX-750, 750 W, 20 kHz, Sonic and Materials, Connecticut, USA)at a specific energy input of 20 kJ/g TS with temperature controlin order to hinder methane-producing bacteria as described inElbeshbishy et al. (2010). The feed containing glucose at a concen-tration of 8.5 g/L, was supplemented by 5 mL/L of a nutrient stocksolution with the following composition per litre of stock: 1000 gNaHCO3, 280 g NH4Cl, 250 g of K2HPO4, 10 g of CaCl2.2H2O, 0.05 gof H3BO3, 0.05 g of ZnCl2, 0.03 g of CuCl2, 0.5 g of MnCl2.4H2O,0.05 g of (NH4)6Mo7O24, 0.05 g of AlCl3, 0.05 g of CoCl2.6H2O, and0.05 g of NiCl2.
2.2. Batch test
To assess the effect of different S2� and Fe2+ concentrations onbiohydrogen production, batch experiments were conducted in aseries of serum bottles (liquid volume of 200 ml). The volumes ofsubstrate (glucose) and seed (13.5 g VSS/L) based on food to micro-organism (F/M) ratio of 2 mg of CODsubstrate/mg of VSSseed (Nasret al., 2010), were 150 mL and 50 mL, respectively. Differentconcentrations of S2� and Fe2+ were added to the substrate asNa2S�9H2O (EMD Chemicals Inc., Germany), and FeCl2�4H2O (EMDChemicals Inc., Germany), respectively. To evaluate the impact ofFe2+ on sulfide inhibition, FeCl2�4H2O was added at 1, 2, and 4times the stoichiometric requirement of 1.75 mg Fe2+/mg of S2�.Control bottles were also prepared using the substrate containingonly glucose without any Na2S�9H2O and FeCl2�4H2O addition.The concentrations of S2� and Fe2+ in the substrate for the differentsamples are summarized in Table 1. The initial pHs of all bottleswere adjusted to 6.8 using 1 N NaOH and 1 N HCl before startingthe experiment. After purging with oxygen-free nitrogen, the ser-um bottles were sealed with rubber septa and agitated in theshaker (MaxQ 4000, incubator and refrigerated shaker, ThermoScientific, Fremont, CA) at 180 rpm, at the mesophilic temperatureof 37 ± 1 �C. The produced biogas volume and hydrogen content ofthe biogas were monitored continuously until biogas productionstopped. All experiments were conducted in duplicates. Differencesbetween duplicate measurements were less than 10% for allparameters, and hence average values are reported here.
2.3. Analytical methods
Samples were analyzed for total suspended solids (TSS), andvolatile suspended solids (VSS) according to the Standard Methods(APHA, 1998). Soluble parameters were analyzed after filtering thesamples through 0.45 lm membrane filters (Sterile membrane fil-ter, Cat. No. 7141104, Whatman Limited, England). HACH vialswere used to measure chemical oxygen demand (COD), and sulfide(S2�) concentrations. Dissolved iron and dissolved sulfur concen-trations were analyzed by Inductively Coupled Plasma OpticalEmission Spectrophotometer (ICP-OES) (Varian, Inc., VistaPro Ax-ial). The concentrations of volatile fatty acids (VFAs) were analyzedusing a gas chromatograph (Model Varian 8500, Varian Inc., Toron-to, Canada) with a flame ionization detector (FID) equipped with afused silica column (30 m � 0.32 mm). Helium was used as the

B.R. Dhar et al. / Bioresource Technology 126 (2012) 123–130 125
carrier gas at a flow rate of 5 mL/min. The temperatures of the col-umn and detector were 110 and 250 �C, respectively. Glucose wasanalyzed by the phenol-sulfuric acid method (Dubois et al., 1956).
Biogas volume was measured by releasing the gas pressure inthe serum vials using appropriately sized glass syringes (Perfek-tum; Popper & Sons Inc., NY, USA) in the 5–100 mL range to equil-ibrate with the ambient pressure (Owen et al., 1979). Hydrogenand methane content of the biogas were analyzed using SRI 310CGas Chromatograph (Model 310, SRI Instruments, Torrance, CA)equipped with a molecular sieve column (Molesieve 5A, mesh80/100, 182.88 � 0.3175 cm) and a thermal conductivity detector(TCD). The temperatures of the column and the TCD detector were90 and 105 �C, respectively. Argon was used as carrier gas at a flowrate of 30 mL/min.
2.4. Kinetic analysis
The modified Gompertz equation has been used to describe thecumulative hydrogen production observed for various samples inthe batch study (Zwietering et al., 1990):
H ¼ P � exp � expRm
pðk� tÞ þ 1
� �� �ð1Þ
where, H is the cumulative hydrogen production (ml), P is themaximum hydrogen production (ml), Rm is the maximum hydrogenproduction rate (ml/hr), k is the lag phase time (hr), and e = exp(1) = 2.718. The average cumulative hydrogen production of thetwo duplicates was used for the kinetic analysis, and differences be-tween duplicate measurements were less than 10%. The cumulativehydrogen production data were fitted with Gompertz equationusing the Newton–Raphson method for non-linear numericalestimation.
3. Results and discussion
3.1. Biohydrogen production
3.1.1. Impact of various sulfide concentrationsFig. 1(a) illustrates the cumulative temporal biohydrogen pro-
duction for the control and the batches containing various initialS2� concentration ranging from 25 to 500 mg/L. The highest ulti-mate biohydrogen production was observed for the batch contain-ing 25 mg/L S2�, representing a 54% increase over the control. Withincreasing initial S2� concentration above 50 mg/L, biohydrogenproduction gradually decreased. However, the ultimate biohydro-gen production was substantially higher than the control for initialS2� concentration up to 100 mg/L. Initial dissolved sulfide concen-trations of 250 and 500 mg/L have been found to significantly inhi-bit the hydrogen producing bacteria, with ultimate biohydrogenproductions 40% and 90%, respectively, lower than the control.Thus, it is evident that high initial dissolved S2� concentrationsmay completely inhibit the activity of hydrogen producing bacteriain dark biohydrogen fermentation. Several inhibition mechanismsfor sulfide have been reported in literature. At very high concentra-tions (100–800 mg/L), sulfide has been found to diffuse into themicrobial cell, and interfere with the bioactivity of anaerobicmicroorganisms (Speece, 1983; Chen et al., 2008b). Sulfide mayalso inhibit the microbial activity by denaturing the cell proteinsthrough the formation of sulfide and disulfide cross-links betweenpolypeptide chains, interfering with the activities of variousenzymes and coenzymes, chemically reacting with cytochromes,thus impacting the assimilatory metabolism of sulfur (Mara andHoran, 2003; Chen et al., 2008b; Speece, 1983). However, it wasalso interesting to observe that the highest ultimate biohydrogenproduction (270 mL), which was 54% higher than the control,
occurred at the lowest initial S2� concentration of 25 mg/L. Thus,the results suggest that addition of an optimum amount of sulfuras dissolved S2� can appreciably facilitate the microbial metabo-lism of the hydrogen producing bacteria as well as enhance theultimate biohydrogen production. Sulfur is an important macronu-trient for fermentative microorganisms, and some bacteria storegranules of sulfur as their energy reserves (Mara and Horan,2003). It has been also found that sulfide is an obligatory nutrientfor methanogens with an optimum sulfide concentration in therange of 1–25 mg/L (Speece, 1983). To date, no information onthe optimum sulfide requirement is available in literature forhydrogen producing microorganisms in dark fermentation. How-ever, Maeda et al. (2000) has found that sulfide could be utilizedas hydrogen donor, and observed 3 times enhanced hydrogen pro-duction in photo-fermentation of seawater in the presence of0.2 mM sodium sulfide (Na2S).
3.1.2. Impact of various ferrous iron concentrationsThe influence of different initial Fe2+ concentrations ranging
from 25 to 500 mg/L on cumulative biohydrogen production inbatch tests (no sulfide was added in these batches) is depicted inFig. 1(b). As apparent from Fig. 1(b), ultimate biohydrogen produc-tion for all batches containing Fe2+ were 11–23% higher than thecontrol. Addition of Fe2+ in the substrate has been found toenhance the activity of hydrogen producing hydrogenase enzyme,and hydrogenase enzyme oxidizes reduced ferredoxin to producemolecular hydrogen (Wang and Wan, 2009). Therefore, theincreased biohydrogen production observed might be due to theenhanced hydrogenase activity. However, in this study, the in-crease in hydrogen production did not show any consistent trendwith increasing initial Fe2+ concentrations from 25 to 500 mg/L.The reported impact of Fe2+ concentrations on dark biohydrogenfermentation as well the optimum Fe2+ concentration in literaturestudies varies widely. The optimum Fe2+ concentration reported inliterature for enhanced biohydrogen production is 10–1600 mg/L(Zhang et al., 2005; Liu and Shen, 2004). Zhang et al. (2005) havefound 1600 mg/L Fe2+ as optimum in batch dark biohydrogen fer-mentation of from sucrose using mixed culture, while Liu and Shen(2004) have found 10 mg/L Fe2+ as optimum for biohydrogenproduction from starch. Both studies have been done at the sameinitial pH and temperature. The differences in fermentation operat-ing conditions (e.g. inoculum, substrate etc.) in these studies mightrationalize these widely disparate optimum Fe2+ concentrations.Few studies also reported no impact of iron addition on hydrogenproduction (Lee et al., 2001). Thus, the literature studies as well asthe observed trend in this study confirmed that the optimum Fe2+
dosage for dark biohydrogen fermentation is strictly linked to thefermentation operating conditions.
Ultimate hydrogen production observed for different initial Fe2+
doses (25–500 mg/L) with no supplemental sulfur addition wasnotably lower than the hydrogen productions at initial dissolvedS2� concentrations of 25 and 50 mg/L (194–216 mL versus 255–270 mL). Lee et al. (2009) have found that iron and sulfur both playa very important role in enhancing hydrogenase activity throughthe hydrogenase biosynthesis and releasing hydrogen containedin cells, and suggested that the addition of FeSO4 is more beneficialfor dark biohydrogen production than the addition of FeCl3. Allhydrogenases are composed of iron in the form of Fe–S clusters(Cammack et al., 1986). The Fe-hydrogenases act as terminal elec-tron acceptors, and catalyze the reduction of protons to producehydrogen. Therefore, microorganisms can conserve metabolicenergy in iron-sulfur protein clusters by iron and sulfur supple-mentation (Lee et al., 2009). Thus, the hydrogen productivity indark fermentation considerably depends on iron and sulfur ratherthan only iron. Our results, suggest that very low sulfur-containing

Fig. 1. (a) Effect of different sulfide concentrations, (b) effect of different ferrous iron concentrations, and (c) combined effect of sulfide and iron on cumulative hydrogenproduction with time.
126 B.R. Dhar et al. / Bioresource Technology 126 (2012) 123–130
feedstocks should be supplemented with an optimum amount ofsulfur.
3.1.3. Combined effect of iron and sulfideFig. 1(c) shows the impact of different Fe2+ concentrations on
biohydrogen production from the batches containing dissolvedS2� concentrations of 50, 100, and 500 mg/L. The added Fe2+ doseswere based on the 1, 2, and 4 times of the theoretical requirementfor 100% S2� precipitation. Without any Fe2+ addition, the totalcumulative hydrogen production for the substrates containing 50,100, and 500 mg/L S2� were 255, 206, and 18 ml, respectively. Inbatch tests, addition of Fe2+ ranging from 87.5 to 350 mg/L to thesubstrate containing 50 mg/L of dissolved S2� have shown signifi-cantly lower hydrogen productions (197–209 mL H2) than the sub-strate containing 50 mg/L of dissolved S2� without any Fe2+
addition (255 mL). Therefore, the results of hydrogen productionand dissolved iron in final samples (discussed later) suggest thatthe addition of Fe2+ to the substrate containing low initialdissolved S2� concentration (50 mg/L) may diminish the positiveimpact of sulfide supplementation on dark fermentation by precip-itation. However, the addition of Fe2+ to the substrate with initialdissolved sulfide concentration of 100 mg/L S2� have shown almostthe same total cumulative biohydrogen as the batches without anyFe2+ addition. At the high initial dissolved sulfide concentration of500 mg/L S2�, the addition of Fe2+ at the stoichiometric require-ment for complete (100%) S2� precipitation (875 mg/L Fe2+ for500 mg/L S2�) has substantially compensated for the inhibitoryeffect of the 500 mg/L S2� concentration, and increased the
cumulative biogas production from 18 to 153 ml; doubling theFe2+ dose has increased the hydrogen production to 169 mL, stillslightly lower than that the control (175 mL). The addition ofFe2+ at 4 times the stoichiometric dose (3500 mg/L Fe2+ for500 mg/L S2�) has totally offset the inhibitory effect of sulfide ondark fermentation, with biohydrogen production 11% higher thanthe control (194 mL versus 175 mL). The elimination of the inhib-itory effect of the high initial sulfide concentration (500 mg/L) isdue to the precipitation of sulfide as ferrous sulfide. Thus, it isevident that the addition of ferrous iron can be used as a sulfideinhibition control strategy for dark biohydrogen fermentation.The results of this study also suggested that Fe2+ at 3–4 timesthe stoichiometric dose may be required to completely compen-sate for the inhibitory/toxicity effect of sulfide on hydrogen pro-ducing bacteria.
3.2. Hydrogen yield and COD balance
At the end of batch fermentation tests, glucose, and COD con-centrations were analyzed. Glucose was completely consumed inall bottles with 98–100% conversion efficiency of glucose apartfrom set#6, the batch containing 500 mg/L S2� (73% conversionefficiency). Fig. 2(a) shows the hydrogen yields calculated basedon the glucose consumed. For most of the samples, hydrogen yieldswere observed within the range of hydrogen yields from glucosereported in the literature (1–2.2 mol H2/mol glucoseconsumed)(Wang and Wan, 2009). The maximum hydrogen yield (1.7 molH2/mol glucoseconsumed) was observed for the batch containing
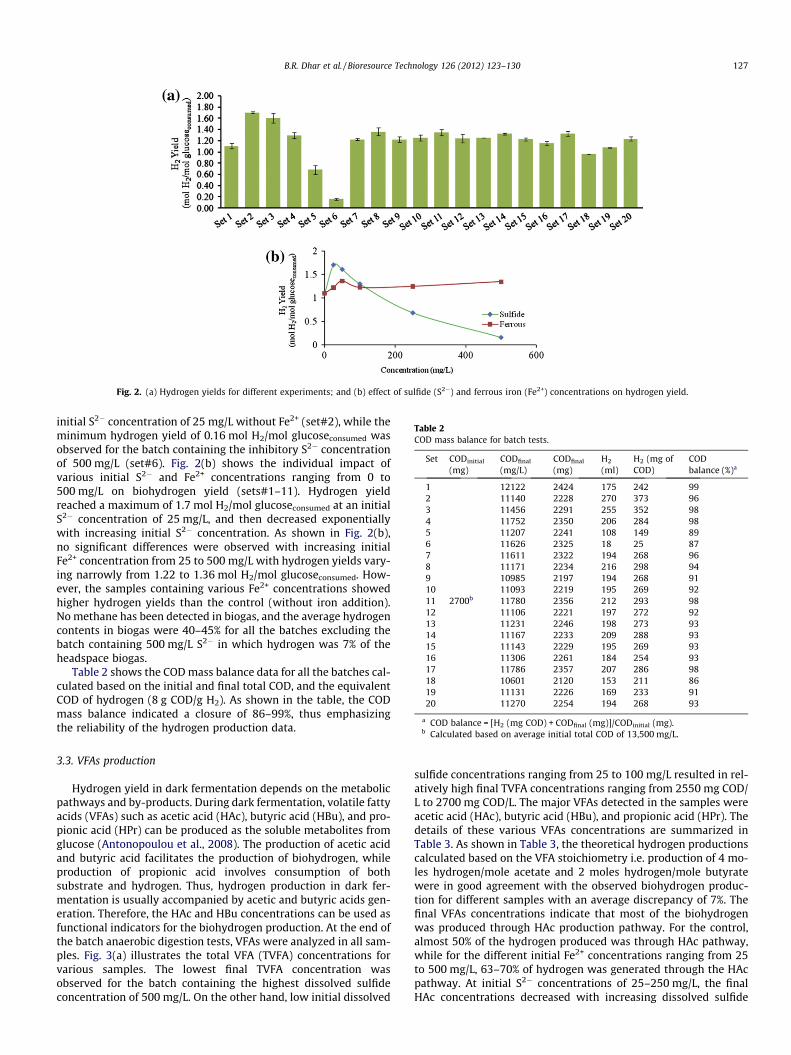
Fig. 2. (a) Hydrogen yields for different experiments; and (b) effect of sulfide (S2�) and ferrous iron (Fe2+) concentrations on hydrogen yield.
Table 2COD mass balance for batch tests.
Set CODinitial
(mg)CODfinal
(mg/L)CODfinal
(mg)H2
(ml)H2 (mg ofCOD)
CODbalance (%)a
1 12122 2424 175 242 992 11140 2228 270 373 963 11456 2291 255 352 984 11752 2350 206 284 985 11207 2241 108 149 896 11626 2325 18 25 877 11611 2322 194 268 968 11171 2234 216 298 949 10985 2197 194 268 9110 11093 2219 195 269 9211 2700b 11780 2356 212 293 9812 11106 2221 197 272 9213 11231 2246 198 273 9314 11167 2233 209 288 9315 11143 2229 195 269 9316 11306 2261 184 254 9317 11786 2357 207 286 9818 10601 2120 153 211 8619 11131 2226 169 233 9120 11270 2254 194 268 93
a COD balance = [H2 (mg COD) + CODfinal (mg)]/CODinitial (mg).b Calculated based on average initial total COD of 13,500 mg/L.
B.R. Dhar et al. / Bioresource Technology 126 (2012) 123–130 127
initial S2� concentration of 25 mg/L without Fe2+ (set#2), while theminimum hydrogen yield of 0.16 mol H2/mol glucoseconsumed wasobserved for the batch containing the inhibitory S2� concentrationof 500 mg/L (set#6). Fig. 2(b) shows the individual impact ofvarious initial S2� and Fe2+ concentrations ranging from 0 to500 mg/L on biohydrogen yield (sets#1–11). Hydrogen yieldreached a maximum of 1.7 mol H2/mol glucoseconsumed at an initialS2� concentration of 25 mg/L, and then decreased exponentiallywith increasing initial S2� concentration. As shown in Fig. 2(b),no significant differences were observed with increasing initialFe2+ concentration from 25 to 500 mg/L with hydrogen yields vary-ing narrowly from 1.22 to 1.36 mol H2/mol glucoseconsumed. How-ever, the samples containing various Fe2+ concentrations showedhigher hydrogen yields than the control (without iron addition).No methane has been detected in biogas, and the average hydrogencontents in biogas were 40–45% for all the batches excluding thebatch containing 500 mg/L S2� in which hydrogen was 7% of theheadspace biogas.
Table 2 shows the COD mass balance data for all the batches cal-culated based on the initial and final total COD, and the equivalentCOD of hydrogen (8 g COD/g H2). As shown in the table, the CODmass balance indicated a closure of 86–99%, thus emphasizingthe reliability of the hydrogen production data.
3.3. VFAs production
Hydrogen yield in dark fermentation depends on the metabolicpathways and by-products. During dark fermentation, volatile fattyacids (VFAs) such as acetic acid (HAc), butyric acid (HBu), and pro-pionic acid (HPr) can be produced as the soluble metabolites fromglucose (Antonopoulou et al., 2008). The production of acetic acidand butyric acid facilitates the production of biohydrogen, whileproduction of propionic acid involves consumption of bothsubstrate and hydrogen. Thus, hydrogen production in dark fer-mentation is usually accompanied by acetic and butyric acids gen-eration. Therefore, the HAc and HBu concentrations can be used asfunctional indicators for the biohydrogen production. At the end ofthe batch anaerobic digestion tests, VFAs were analyzed in all sam-ples. Fig. 3(a) illustrates the total VFA (TVFA) concentrations forvarious samples. The lowest final TVFA concentration wasobserved for the batch containing the highest dissolved sulfideconcentration of 500 mg/L. On the other hand, low initial dissolved
sulfide concentrations ranging from 25 to 100 mg/L resulted in rel-atively high final TVFA concentrations ranging from 2550 mg COD/L to 2700 mg COD/L. The major VFAs detected in the samples wereacetic acid (HAc), butyric acid (HBu), and propionic acid (HPr). Thedetails of these various VFAs concentrations are summarized inTable 3. As shown in Table 3, the theoretical hydrogen productionscalculated based on the VFA stoichiometry i.e. production of 4 mo-les hydrogen/mole acetate and 2 moles hydrogen/mole butyratewere in good agreement with the observed biohydrogen produc-tion for different samples with an average discrepancy of 7%. Thefinal VFAs concentrations indicate that most of the biohydrogenwas produced through HAc production pathway. For the control,almost 50% of the hydrogen produced was through HAc pathway,while for the different initial Fe2+ concentrations ranging from 25to 500 mg/L, 63–70% of hydrogen was generated through the HAcpathway. At initial S2� concentrations of 25–250 mg/L, the finalHAc concentrations decreased with increasing dissolved sulfide
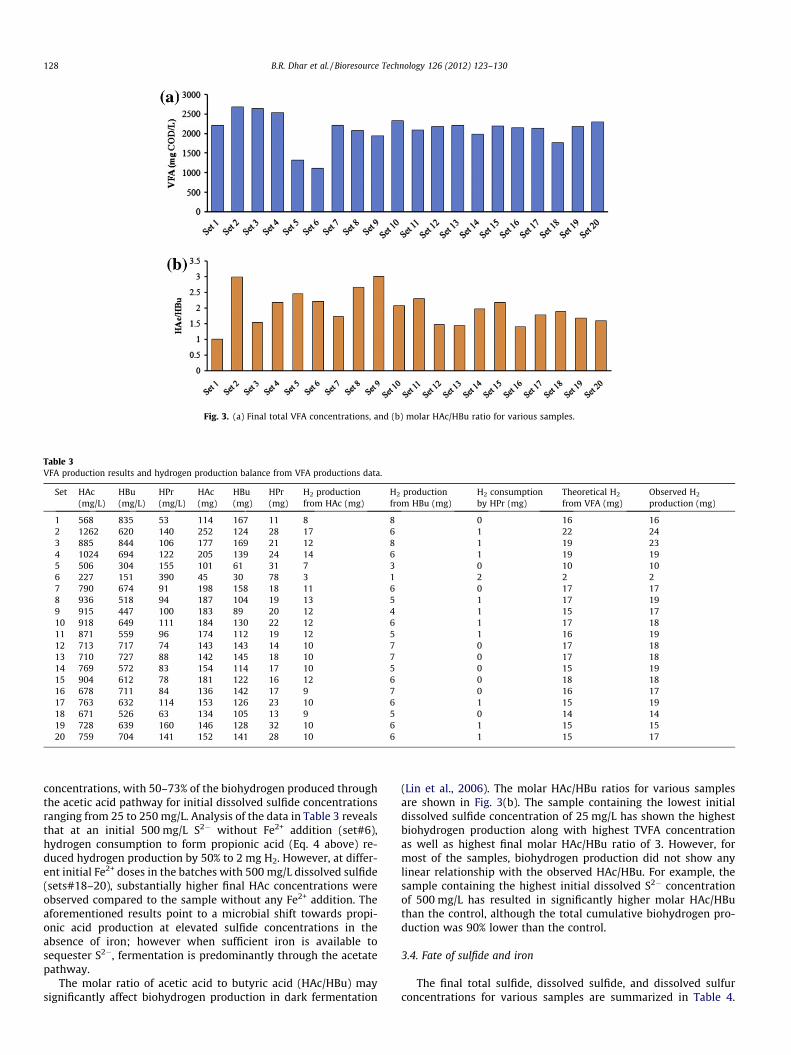
Fig. 3. (a) Final total VFA concentrations, and (b) molar HAc/HBu ratio for various samples.
Table 3VFA production results and hydrogen production balance from VFA productions data.
Set HAc(mg/L)
HBu(mg/L)
HPr(mg/L)
HAc(mg)
HBu(mg)
HPr(mg)
H2 productionfrom HAc (mg)
H2 productionfrom HBu (mg)
H2 consumptionby HPr (mg)
Theoretical H2
from VFA (mg)Observed H2
production (mg)
1 568 835 53 114 167 11 8 8 0 16 162 1262 620 140 252 124 28 17 6 1 22 243 885 844 106 177 169 21 12 8 1 19 234 1024 694 122 205 139 24 14 6 1 19 195 506 304 155 101 61 31 7 3 0 10 106 227 151 390 45 30 78 3 1 2 2 27 790 674 91 198 158 18 11 6 0 17 178 936 518 94 187 104 19 13 5 1 17 199 915 447 100 183 89 20 12 4 1 15 1710 918 649 111 184 130 22 12 6 1 17 1811 871 559 96 174 112 19 12 5 1 16 1912 713 717 74 143 143 14 10 7 0 17 1813 710 727 88 142 145 18 10 7 0 17 1814 769 572 83 154 114 17 10 5 0 15 1915 904 612 78 181 122 16 12 6 0 18 1816 678 711 84 136 142 17 9 7 0 16 1717 763 632 114 153 126 23 10 6 1 15 1918 671 526 63 134 105 13 9 5 0 14 1419 728 639 160 146 128 32 10 6 1 15 1520 759 704 141 152 141 28 10 6 1 15 17
128 B.R. Dhar et al. / Bioresource Technology 126 (2012) 123–130
concentrations, with 50–73% of the biohydrogen produced throughthe acetic acid pathway for initial dissolved sulfide concentrationsranging from 25 to 250 mg/L. Analysis of the data in Table 3 revealsthat at an initial 500 mg/L S2� without Fe2+ addition (set#6),hydrogen consumption to form propionic acid (Eq. 4 above) re-duced hydrogen production by 50% to 2 mg H2. However, at differ-ent initial Fe2+ doses in the batches with 500 mg/L dissolved sulfide(sets#18–20), substantially higher final HAc concentrations wereobserved compared to the sample without any Fe2+ addition. Theaforementioned results point to a microbial shift towards propi-onic acid production at elevated sulfide concentrations in theabsence of iron; however when sufficient iron is available tosequester S2�, fermentation is predominantly through the acetatepathway.
The molar ratio of acetic acid to butyric acid (HAc/HBu) maysignificantly affect biohydrogen production in dark fermentation
(Lin et al., 2006). The molar HAc/HBu ratios for various samplesare shown in Fig. 3(b). The sample containing the lowest initialdissolved sulfide concentration of 25 mg/L has shown the highestbiohydrogen production along with highest TVFA concentrationas well as highest final molar HAc/HBu ratio of 3. However, formost of the samples, biohydrogen production did not show anylinear relationship with the observed HAc/HBu. For example, thesample containing the highest initial dissolved S2� concentrationof 500 mg/L has resulted in significantly higher molar HAc/HButhan the control, although the total cumulative biohydrogen pro-duction was 90% lower than the control.
3.4. Fate of sulfide and iron
The final total sulfide, dissolved sulfide, and dissolved sulfurconcentrations for various samples are summarized in Table 4.

Table 4Final dissolved iron, dissolved sulfur, total and dissolved sulfide concentrations.
Set Dissolvediron (mg/L)
Dissolvedsulfur (mg/L)
Totalsulfide (mg/L)
Dissolvedsulfide (mg/L)
1 1 24 5 02 5 22 6 03 9 32 17 04 3 27 37 05 4 54 69 06 6 92 216 07 5 17 6 08 11 18 6 19 18 16 6 010 71 16 5 011 162 13 4 012 11 28 21 013 30 27 12 014 10 25 4 015 8 25 36 116 3 23 20 017 240 25 7 018 6 33 133 319 288 49 76 120 96 5 20 0
Table 5Gompertz kinetic coefficients for different experiments.
Set S2� (mg/L) Fe2+ (mg/L) P (ml) Rm (ml/hr) k (h) R2
1 – – 174 25 8 0.9992 25 – 263 30 12 0.9993 50 – 251 25 9 0.9994 100 – 194 29 11 0.9995 250 – 108 9 20 0.9996 500 – 18.1 0.7 17 0.9997 – 25 195 22 6 0.9998 – 50 214 28 11 0.9999 – 100 194 33 10 0.99910 – 250 192 32 12 0.99911 – 500 209 35 10 0.99912 50 87.5 194 34 9 0.99913 50 175 196 34 8 0.99914 50 350 207 36 8 0.99915 100 175 194 34 8 0.99916 100 350 183 25 6 0.99917 100 700 205 36 8 0.99918 500 875 153 23 6 0.99919 500 1750 167 21 6 0.99920 500 3500 192 33 9 0.999
B.R. Dhar et al. / Bioresource Technology 126 (2012) 123–130 129
Total sulfide is composed of dissolved sulfide and insoluble sulfide(i.e. ferrous sulfide). For all of the samples, the final dissolvedS2� concentrations were almost 0 mg/L irrespective of initialconcentration and iron dose. Although the initial dissolved sulfideconcentrations were different, the samples without Fe2+, the finaltotal sulfide concentrations (in Table 4) decreased significantly,and correlated linearly with the initial dissolved sulfide concentra-tions (R2 = 0.965, figure not shown) with the final total sulfideconcentration less than 42% of the initial dissolved sulfide, thusindicating that the dissolved sulfides might are emitted as hydro-gen sulfide (H2S) in the biogas (Dhar et al., 2011; Peu et al., 2011).
In the batches with both sulfide and iron, the final total sulfideconcentrations decreased linearly with the addition of Fe2+ due tochemical precipitation. In the samples without sulfide, the finaldissolved iron concentrations were significantly below the initialconcentrations. The final soluble iron concentrations increasedlinearly with the initial Fe2+ concentrations (R2 = 0.993, figure notshown). Thus, it is evident that Fe2+ is consumed not only in reac-tions with the dissolved sulfides to form ferrous sulfide (FeS), butalso with phosphate to from ferrous phosphate, and by theanaerobic microorganisms for other biological processes (Singer,1972; Straub et al., 1996).
3.5. Kinetic study
The three Gompertz parameters: maximum hydrogen produc-tion potential, maximum hydrogen production rate, and lag phasetime for all batches were calculated and tabulated in Table 5. Thecoefficient of determination (R2) of 0.999 for all the regressionsindicated a strong correlation between the experimental dataand the fit. The results of the kinetic analysis show that the hydro-gen production rate decreased with increasing initial S2� concen-trations, and higher initial dissolved sulfide concentrations(250–500 mg/L) resulted in significantly extended lag-phase times(17–20 h) along with lower hydrogen production rates comparedto the control due to sulfide inhibition. Hydrogen production ratesincreased with increasing Fe2+ concentrations. However, the lag-phase time slightly increased for the initial Fe2+ concentrationabove 25 mg/L. Although the highest initial S2� concentration of500 mg/L has completely inhibited biohydrogen production withthe lowest biohydrogen production rate of 0.9 ml H2/h, additionof Fe2+ from 875 to 3500 mg/L has increased the biogas productionrates by 23–33%. As well as decreased the lag phase from 17 to 9 hafter Fe2+ for this sample. Thus, the predicted maximum hydrogenproduction rates for different batches estimated by the modifiedGompertz model suggested that the addition of Fe2+ to the highinitial sulfide samples not only helped to compensate for theinhibitory effect of sulfide but also enhanced the biohydrogen pro-duction rate.
4. Conclusions
Biohydrogen production was completely inhibited at a dis-solved sulfide (S2�) concentration of 500 mg/L. However, a S2�
concentration of 25 mg/L significantly enhanced for biohydrogenproduction, thus indicating that maintaining an optimum S2� con-centration is very important for dark biohydrogen fermentation, assulfur limitation may adversely impact the biohdyrogen produc-tion. Addition of Fe2+ to the substrate mitigated sulfide inhibitionof dark fermentation. However, overdosing Fe2+ at the lowdissolved sulfide concentration of 50 mg/L eliminated the positiveimpact of sulfide by reducing the bioavailability of sulfur throughprecipitation.
References
Antonopoulou, G., Gavala, H.N., Skiadas, I.V., Angelopoulos, K., Lyberatos, G., 2008.Biofuels generation from sweet sorghum: fermentative hydrogen productionand anaerobic digestion of the remaining biomass. Bioresour. Technol. 99 (1),110–119.
APHA, 1998. Standard Methods for the Examination of Water and Wastewater.American Public Health Association, Washington DC, USA.
Cammack, R., Fernandez, V.M., Schneider, K., 1986. Activation and active sites ofnickel containing hydrogenases. Biochimie 68, 85–91.
Chen, C.C., Chen, H.P., Wu, J.H., Lin, C.Y., 2008a. Fermentative hydrogen productionat high sulfate concentration. Int. J. Hydrogen Energy 33, 1573–1578.
Chen, Y., Cheng, J.J., Creamer, K.S., 2008b. Inhibition of anaerobic digestion process:a review. Bioresour. Technol. 99, 4044–4064.
Dezham, P., Rosenblum, E., Jenkins, D., 1988. Digester gas H2S control using ironsalts. J. Water Pollut. Control Fed. 60, 514–517.
Dhar, B.R., Youssef, E., Nakhla, G., Ray, M.B., 2011. Pretreatment of waste activatedsludge for volatile sulfur compounds control in anaerobic digestion. Bioresour.Technol. 102 (4), 3776–3782.
Dubois, M., Gilles, K.A., Hamilton, J.K., Rebers, P.A., Smith, F., 1956. Colorimetricmethod for determination of sugars and related substances. Anal. Chem. 28,350–356.
Elbeshbishy, E., Hafez, H., Nakhla, G., 2010. Enhancement of biohydrogen producingusing ultrasonication. Int. J. Hydrogen Energy 35, 6184–6193.
Hwang, J.H., Choi, J.A., Oh, Y.K., Abou-Shanab, R.A.I., Song, H., Min, B., Cho, Y., Na, J.G.,Koo, J., Jeon, B.H., 2011. Hydrogen production from sulfate- and ferrous-enriched wastewater. Int. J. Hydrogen Energy 36 (21), 13984–13990.
Hwang, J.H., Choi, J.A., Abou-Shanab, R.A.I., Bhatnagar, A., Min, B., Song, H., Kumar,E., Choi, J., Lee, E.S., Kim, Y.J., Um, S., Lee, D.S., Jeon, B.H., 2009. Effect of pH andsulfate concentration on hydrogen production using anaerobic mixedmicroflora. Int. J. Hydrogen Energy 34, 9702–9710.

130 B.R. Dhar et al. / Bioresource Technology 126 (2012) 123–130
Kalyuzhnyi, S., Fedorovich, V., Lens, P., Hulshoff, P.L., Lettinga, G., 1998.Mathematical modelling as a tool to study population dynamics betweensulfate reducing and methanogenic bacteria. Biodegradation 9,187–199.
Lee, D.Y., Li, Y.Y., Oh, Y.K., Kim, M.S., Noike, T., 2009. Effect of iron concentration oncontinuous H2 production using membrane bioreactor. Int. J. Hydrogen Energy34 (3), 1244–1252.
Lee, Y.J., Miyahara, T., Noike, T., 2001. Effect of iron concentration on hydrogenfermentation. Bioresour. Technol. 80, 227–231.
Lens, P.N.L., Visser, A., Janssen, A.J.H., Hulshoff Pol, L.W., Lettinga, G., 1998.Biotechnological treatment of sulfate-rich wastewaters. Crit. Rev. Environ. Sci.Technol. 28 (1), 41–88.
Lin, C.Y., Chen, H.P., 2006. Sulfate effect on fermentative hydrogen production usinganaerobic mixed microflora. Int. J. Hydrogen Energy 31, 953–960.
Lin, C.Y., Hung, C.H., Chen, C.H., Chung, W.T., Cheng, L.H., 2006. Effects of initialcultivation pH on fermentative hydrogen production from xylose using naturalmixed cultures. Process Biochem. 41, 1383–1390.
Liu, G., Shen, J., 2004. Effects of culture medium and medium conditions onhydrogen production from starch using anaerobic bacteria. J. Biosci. Bioeng. 98,251–256.
Maeda, I., Mizoguchi, T., Miura, Y., Yagi, K., Shioji, N., Miyasaka, H., 2000. Influence ofsulfate-reducing bacteria on outdoor hydrogen production by photosyntheticbacterium with seawater. Curr. Microbiol. 40, 210–213.
Mara, D., Horan, N.J., 2003. Handbook of Water and Wastewater Microbiology.Academic Press, UK.
McCartney, D.M., Oleszkiewicz, J.A., 1993. Competition between methanogens andsulfate reducers: effect of COD:sulphate ratio and acclimation. Water Environ.Res. 65 (5), 655–664.
Nasr, N., Elbeshbishy, E., Hafez, H., Nakhla, G., El Naggar, M.H., 2010. Bio-hydrogenproduction from thin stillage using conventional and acclimatized anaerobicdigester sludge. Int. J. Hydrogen Energy 36, 12761–12769.
Nicolet, Y., Cavazza, C., Fontecilla-Camps, J.C., 2002. Fe-only hydrogenases:structure, future and evolution. J. Inorg. Biochem. 91, 1–8.
Nielsen, A.H., Hvitved-Jacobsen, T., Vollertsen, J., 2008. Effects of pH and ironconcentrations on sulfide precipitation in wastewater collection systems. WaterEnviron. Res. 80 (4), 380–384.
Owen, W.F., Stuckey, D.C., Healy, J.B., Young, L.Y., McCarty, P.L., 1979. Bioassay formonitoring biochemical methane potential and anaerobic toxicity. Water Res.13, 485–492.
Parkin, G.F., Lynch, N.A., Kuo, W., Van Keuren, E.L., Bhattacharya, S.K., 1990.Interaction between sulfate reducers and methanogens fed acetate andpropionate. Res. J. Water Pollut. Control Fed. 62, 780–788.
Peu, P., Sassi, J.F., Girault, R., Picard, S., Saint-Cast, P., Béline, F., Dabert, P., 2011.Sulphur fate and anaerobic biodegradation potential during co-digestion ofseaweed biomass (Ulva sp.) with pig slurry. Bioresour. Technol. 102,10794–10802.
Singer, P.C., 1972. Anaerobic control of phosphate by ferrous iron. J. Water Pollut.Control Fed. 44 (4), 663–669.
Sinha, P., Pandey, A., 2011. An evaluative report and challenges for fermentativebiohydrogen production. Int. J. Hydrogen Energy 36 (13), 7460–7478.
Speece, R.E., 1983. Anaerobic biotechnology for industrial waste treatment. Environ.Sci. Technol. 17, A416–A427.
Straub, K.L., Benz, M., Schink, B., Widdel, F., 1996. Anaerobic, nitrate-dependentmicrobial oxidation of ferrous iron. Appl. Environ. Microbiol. 62 (4),1458–1460.
Wang, J., Wan, W., 2009. Factors influencing fermentative hydrogen production: areview. Int. J. Hydrogen Energy 34, 799–811.
Zhang, Y.F., Liu, G.Z., Shen, J.Q., 2005. Hydrogen production in batch culture ofmixed bacteria with sucrose under different iron concentrations. Int. J.Hydrogen Energy 30, 855–860.
Zwietering, M.H., Jongenburger, I., Rombouts, F.M., Vantriet, K., 1990. Modeling ofthe bacterial-growth curve. Appl. Environ. Microbiol. 56, 1875–1881.