inducible nitric oxide synthase in retinal ischemia-reperfusion injury
TRANSCRIPT

Exp. Eye Res. (1996) 63, 501–509
Inducible Nitric Oxide Synthase in Retinal Ischemia-reperfusion
Injury
MASANORI HANGAI, NAGAHISA YOSHIMURA*a, KANO HIROI, MICHIKO MANDAI
YOSHIHITO HONDA
Department of Ophthalmology and Visual Science, Kyoto University Graduate School of Medicine,
Kyoto 606, and a Department of Ophthalmology, Shinshu University School of Medicine,
Matsumoto 390, Japan
(Received Cleveland 12 September 1995 and accepted in revised form 19 January 1996)
The purpose of this study was to determine whether inducible nitric oxide synthase (iNOS) was implicatedin the pathogenesis of retinal ischemia-reperfusion injury. Semi-quantitative reverse transcription-polymerase chain reaction showed that the level of iNOS mRNA expression was markedly increased inrat retina following transient ischemia, with peak expression at 12 hr after reperfusion (15±7-foldincrease over pre-ischemic levels). In situ hybridization showed that iNOS mRNA was expressed byresident retinal cells, most likely glial cells in the innermost retina, and also by the neutrophils that hadinfiltrated the retina. Intraperitoneal administration of NG-(1-iminoethyl)--ornithine (-NIO), aninhibitor of iNOS, significantly increased the rate of b-wave recovery compared to that of control animals.The values (mean³...) were 55±0³4±4% versus 40±1³5±1% (P!0±05) at 1 day and 68±6³6±6%versus 45±8³3±5% (P!0±05) at 3 days. This study shows that iNOS mRNA is highly expressed by non-neuronal cells of the inner retina during reperfusion following transient retinal ischemia. It also showsthat -NIO treatment provides some protection against ischemia-reperfusion injury. We suggest thatnitric oxide produced by iNOS may mediate retinal ischemia-reperfusion injury.
# 1996 Academic Press LimitedKey words : inducible nitric oxide synthase; ischemia-reperfusion; retina; neutrophils ; retinal glial
cells ; NG-(1-iminoethyl)--ornithine ; reverse transcription-polymerase chain reaction; in situ hybrid-ization.
1. Introduction
Nitric oxide (NO) is an important messenger when
present at low physiological concentrations. However,
at high concentrations NO mediates tissue damage
due to its cytotoxicity (Moncada, Palmer and Higgs,
1991). NO is synthesized by three isoforms of NO
synthase (NOS). Neuronal and endothelial isoforms,
termed constitutive NOS (cNOS), share the same en-
zymatic character ; they are constitutively expressed,
are regulated by intracellular calcium, and release
NO for a short period. In contrast, a third isoform of
NOS (termed inducible NOS, iNOS) is induced at the
transcriptional level by macrophages, neutrophils and
a number of other cells in response to inflammatory
stimuli such as lipopolysaccharide and cytokines.
Once expressed, iNOS tonically synthesizes high levels
of NO, independent of intracellular calcium. Recently,
there is increasing evidence to suggest that iNOS is
implicated in the pathogenic process of various
diseases, including cerebral ischemic injury (Kilbourn
et al., 1990; Kroncke et al., 1991; Corbett et al.,
1992; Mulligan, Moncada and Ward, 1992; Iadecola
et al. 1995a, b).
* For correspondence at : Department of Ophthalmology, ShinshuUniversity School of Medicine, Matsumoto 390, Japan.
Inflammatory reactions occur in the retina fol-
lowing transient ischemia (Szabo et al., 1991a, b), and
we previously showed that mRNA for interleukin-1, a
proinflammatory cytokine that can induce iNOS, was
upregulated in rat retina following transient ischemia
(Hangai et al., 1995). It is thus possible that iNOS is
induced in the retina following transient ischemia,
and that it subsequently participates in the patho-
genesis of retinal ischemia-reperfusion injury. The first
objective of this study was to determine whether iNOS
is induced in retina following transient ischemia; the
second objective was to determine whether such
putative iNOS mediates retinal ischemia-reperfusion
injury. To answer these questions, mRNA expression
for iNOS in rat retina was examined following
transient ischemia by semi-quantitative reverse trans-
cription-polymerase chain reaction (RT-PCR) and in
situ hybridization histochemistry. The effects of NG-(1-
iminoethyl)--ornithine (-NIO), which is known to
inhibit iNOS (McCall et al., 1991) were also examined,
on the changes in b-wave of the electroretinogram.
2. Materials and Methods
Animals
All animal studies were conducted in compliance
with the ARVO Statement for the Use of Animals in
0014-4835}96}11050109 $25.00}0 # 1996 Academic Press Limited

502 M. HANGAI ET AL.
Ophthalmic and Vision Research. Seventy-seven adult
male Sprague-Dawley rats weighing 250 to 300 g
were used for polymerase chain reaction (PCR)
experiments, and six were used for in situ hybridization
experiments ; 17 rats weighing 250 g were used for
the electroretinogram (ERG) experiments.
Animal Surgery
Rats were anesthetized by intramuscular injections
of ketamine-HCl (70 mg kg−") and xylazine
(10 mg kg−"). The pupils were dilated with the
instillation of 1% atropine sulfate and 0±5% phenyl-
ephrine. After a lateral conjunctival peritomy and
disinsertion of the lateral rectus muscle, the optic
nerve of the right eye was exposed by blunt dissection.
Retinal ischemia and reperfusion were produced in 41
rats according to the technique described previously,
with slight modification (Stefansson et al., 1988). In
brief, a 6–0 nylon suture was passed behind the optic
nerve and tightened until blood flow ceased in all of
the retinal vessels. Complete nonperfusion was con-
firmed through an operating microscope. Reperfusion
was induced by releasing the suture, and was
confirmed by observation through the operating
microscope. Eyes that failed to reperfuse within 5 min
were excluded from the experiment. Sham-operated
control rats underwent similar surgery but without
tightening of the suture. Rats were sacrificed at 0,
0±25, 0±5, 1, 3, 6, 12, 24, 48, 96, and 168 hr after
reperfusion (n¯3–5, at each time).
Retinal ischemia was also produced in nine rats by
elevating the intraocular pressure. The anterior
chamber was cannulated with a 27 gauge infusion
needle connected to a saline container, which was
elevated to produce an intraocular pressure of
140 mmHg for 2 hr. Optic nerve injury was produced
in 9 rats by ligating the optic nerve intraorbitally for
30 sec. It was also produced in nine rats by crushing
the optic nerve intraorbitally between fine forceps for
30 sec at 0±5 mm from the eye. The latter procedure
produced transient retinal ischemia for 2–5 min. Optic
nerve transection was performed intraorbitally in nine
rats at 0±5 mm from the eye, but care was taken not
to produce retinal ischemia. The rats that suffered
pressure-induced ischemia or the optic nerve damages
were killed at 3, 12 and 24 hr (n¯3 at each time)
after reperfusion.
Reverse Transcription-Polymerase Chain Reaction (RT-
PCR)
For cDNA synthesis total RNA was isolated by the
acid guanidinium thiocyanate-phenol chloroform
(AGPC) extraction method (Chomczynski and Sacchi,
1987). The first strand cDNA was synthesized as
described previously (Hangai et al., 1995). Briefly,
before cDNA synthesis the total RNA was treated by
RNase-free DNase (Promega, Madison, WI, U.S.A.).
The first strand cDNA was synthesized from 5 µg
RNA, and then amplified by a step cycle (95°C, 30 sec;
55°C, 30 sec; 72°C, 60 sec) for 25 or 35 cycles (β-
actin, 25 cycles ; iNOS, 35 cycles) (Saiki et al., 1988).
The primers used in this experiment were CGCTACA-
CTTCCAACGCAAC (sense) and AGGAAGTAGGTGAG-
GGCTTG (antisense) for iNOS (Wood et al., 1993), and
AGCTGAGAGGGAAATCGTGC (sense) and ACCAGA-
CAGCACTGTGTTGG (antisense) for β-actin (Nudel et
al., 1983).
To examine ligation-induced retinal ischemia, the
same RNA preparations were used as in the previous
study (Hangai et al., 1995). The RNAs were freshly
isolated to examine the other animal models used in
this study.
Sequencing of the PCR Products
PCR products were separated by 2% agarose gel
electrophoresis and bands of the expected size (407 bp
for iNOS and 295 bp for β-actin) were extracted. The
extracted DNA was subcloned into the pBluescript II
vector (Stratagene, La Jolla, CA, U.S.A.). Before
subcloning, the vector was treated with Eco RV and
the T-vectors were made by Taq polymerase and
deoxythymidine triphosphate (Marchuk et al., 1991).
Nucleotide sequencing of the subcloned DNA was
carried out by the dideoxynucleotide chain termin-
ation method using a Sequenase ver. 2.0 DNA
sequencing kit (United States Biochemicals, Cleveland,
OH, U.S.A.) (Sanger, Nicklen and Coulson et al.,
1977). Double strand template DNAs were denatured
by alkaline treatment and the sequencing reaction
was initiated by adding T3 and}or T7 primer.
Semi-quantitation of Levels of iNOS mRNA by RT-PCR
Semi-quantitative RT-PCR was carried out on the
cDNA from the rats that suffered ligation-induced
retinal ischemia. This procedure was performed as
previously described (Hangai et al., 1995). Briefly,
2 µCi of radiolabeled dCTP was added to the PCR
mixture. The PCR products were then electrophoresed
in a 2% agarose gel and stained with ethidium
bromide. The bands were then excised and the
radioactivity incorporated into the DNA was measured
by Cerenkov scintillation counting. A standard curve
was drawn from the radioactivity measured in serial
dilutions of the template cDNA; this standard curve
was used to calculate the relative quantity of the
expressed genes in each cDNA. First, the cDNA
concentration was normalized by the relative quantity
of β-actin gene expression determined by PCR with the
specific primers. Similar experiments were carried out
using specific primers for iNOS in order to quantify
their relative expression over the time course of
reperfusion after ischemia. The mean of the results
from three sham-operated control rats was used as the

INDUCIBLE NOS IN RETINAL ISCHEMIA 503
control level, and the level of each gene expression
was calculated as a ratio to this control.
Preparation of cRNA Probe
iNOS cDNA was generated by PCR amplification
using specific primers : nucleotides 2207–3418
(1212 bp) of rat iNOS (Wood et al., 1993). This
fragment was subcloned and sequenced to ascertain
that it was indeed derived from the target genes. cRNA
probe labeled with α-$&S-UTP ("1000 Ci mmol−" ;
Amersham Japan, Tokyo, Japan) was generated by
transcription of linearized plasmid DNAs by using the
Riboprobe Gemini II Core System and T3 RNA
polymerase (Promega). Partial alkaline hydrolysis
was carried out at 60°C to yield approximately
100–150 bp. The radiolabeled probe was diluted with
hybridization buffer to approximately 2¬10( cpm
ml−". Specific activity of the probe was approximately
1±1¬10* cpm mg−".
In Situ Hybridization
At 12 hr after cessation of 2 hr ischemia, the rats
were deeply anesthetized and perfused with
phosphate-buffered 4% paraformaldehyde at 4°C, after
which the eyes were enucleated and postfixed over-
night in the same fixative. Following fixation, the eyes
were dehydrated in a graded alcohol series and
embedded in paraffin. The sagittal sections of 5 µm
thickness were deparaffinized, rehydrated, treated with
glycine (2 mg ml−"), and then acetylated in freshly
prepared 0±25% acetic anhydride in 100 m tri-
ethanolamine. After dehydration, the slides were pre-
hybridized at 50°C for 1 hr in the following buffer :
4¬SSC, 50% formamide, 5¬Denhardt solution,
10 m ethylenediaminetetraacetic acid (EDTA),
33 µg ml−" polyadenylic acid, 250 µg ml−" yeast
tRNA, 20 m dithiothreitol (DTT), and 500 µg ml−"
heat-denatured salmon sperm DNA. In the case of
hybridization, 10% dextran sulfate was added to the
above described buffer. The slides were hybridized for
18 hr in the hybridization buffer, washed four times in
2¬SSC containing 10 m DTT at 50°C, incubated
with RNase A (50 µg ml−" in 0±5 NaCl, 10 m Tris,
1 m EDTA, pH 8±0) for 30 min at 37°C, and then
washed twice in 50% formamide, 2¬SSC, and 10 m
DTT for 30 min. They were then dehydrated, dried,
and dipped in Kodak NTB3 emulsion (Rochester, NY,
U.S.A.) (diluted 1:1 with water). After 4 weeks
exposure, they were developed and counterstained
with cresylviolet.
L-NIO Treatment of Rats Following Induced Ischemia
-NIO was a generous gift from Dr Thom W. Mittag
(Mount Sinai School of Medicine, New York, NY,
U.S.A.). -NIO (2±5 mg ml−") was prepared in saline.
Ischemia was produced for 1 hr in 17 rats by ligating
the optic nerve. Nine rats received intraperitoneal
administration of -NIO (5 mg kg−") four times, once
every 6 hr, beginning 2 hr after reperfusion (total
dose, 20 mg kg−"). The other rats, which served as
controls, received the same volume of saline adminis-
tered according to the same schedule.
Effects of L-NIO on ERG b-Wave Recovery
Before, and at 1 and 3 days after ischemia, flash
ERGs were recorded from both eyes using a photo-
stimulator lamp placed in front of each eye with
maximum light intensity (the intensity was 3500 Lx
on the surface of the cornea). After dark adaptation for
at least 60 min, a carbon electrode (NEC San-ei,
Tokyo, Japan) was placed on the cornea and stainless
steel needle electrodes (NEC San-ei) placed under the
skin of the nose and the tail, which served as a
reference and ground, respectively. The responses
were amplified with a time constant of 0±3 sec and a
high pass filter of 1000 Hz (Biological Amplifier 1243,
Nihon Kohden, Tokyo, Japan) and were averaged of
four trials (QC-111J, Nihon, Kohden).
Each b-wave amplitude was estimated as percent
recovery compared to the preischemic level, which
was corrected by the ratio of amplitude change in the
nonischemic fellow eye, as shown in the following
equation:
Recovery (%)¯100¬(iA}iApre
)}(niA}niApre
)
where iApre
and niApre
are the preischemic amplitude
of the ischemic eye and the opposite, nonischemic
eye, respectively, and iA and niA are the postischemic
amplitude of the ischemic eye and the nonischemic
fellow eye, respectively. This calculation was per-
formed to eliminate individual and daily variations
(Spivey and Pearlman, 1953; Lawwill, 1972; Stein-
horst et al., 1993).
Light Microscopic Examination
The eyes used in ERG experiments were also
examined by light microscopy. At 7 days after
reperfusion, the rats were killed and the eyes enu-
cleated. Immediately after enucleation the eyes were
cut open, fixed in 1±5% formaldehyde and 1% glutar-
aldehyde in phosphate buffer, dehydrated through
ethanol and xylene, and embedded in paraffin. Sagittal
sections, 5 µm-thick, through the optic nerve were
obtained and stained with hematoxylin and eosin.
3. Results
Specific Amplification of iNOS mRNA by RT-PCR
Reverse transcription-polymerase chain reaction
(RT-PCR) analysis was carried out at each time point
using specific primers for iNOS. Control amplification
with primers for β-actin was also performed at the
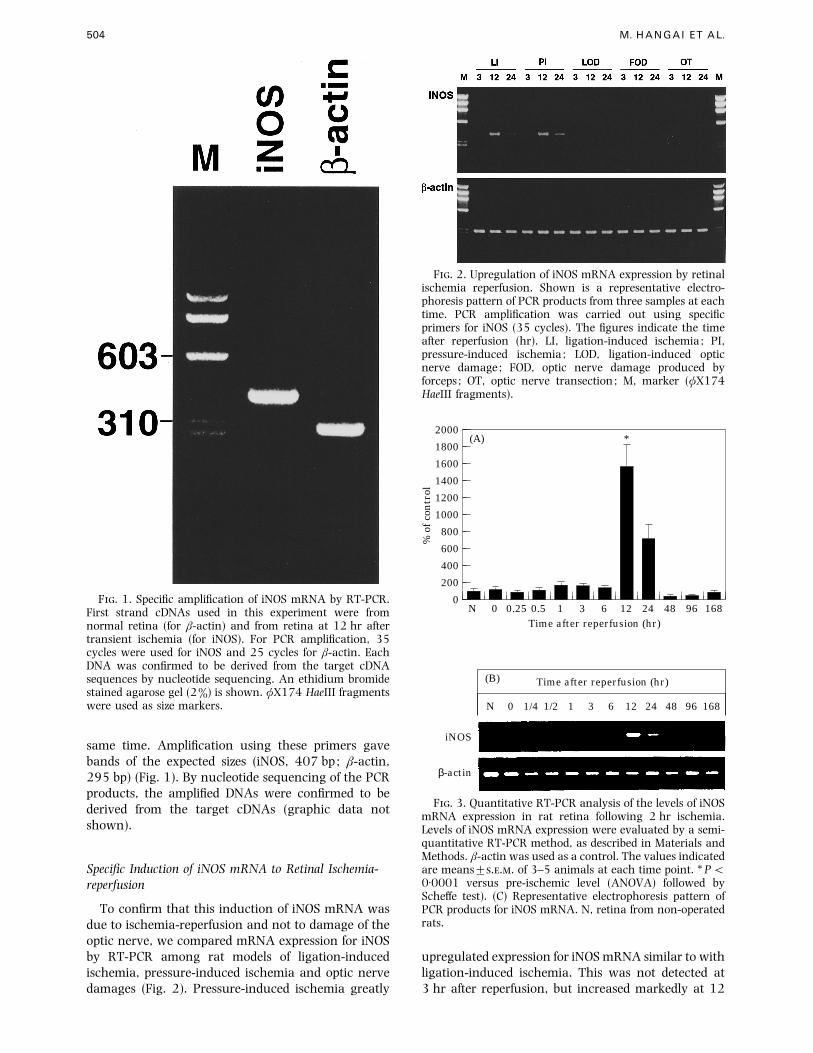
504 M. HANGAI ET AL.
F. 1. Specific amplification of iNOS mRNA by RT-PCR.First strand cDNAs used in this experiment were fromnormal retina (for β-actin) and from retina at 12 hr aftertransient ischemia (for iNOS). For PCR amplification, 35cycles were used for iNOS and 25 cycles for β-actin. EachDNA was confirmed to be derived from the target cDNAsequences by nucleotide sequencing. An ethidium bromidestained agarose gel (2%) is shown. φX174 HaeIII fragmentswere used as size markers.
same time. Amplification using these primers gave
bands of the expected sizes (iNOS, 407 bp; β-actin,
295 bp) (Fig. 1). By nucleotide sequencing of the PCR
products, the amplified DNAs were confirmed to be
derived from the target cDNAs (graphic data not
shown).
Specific Induction of iNOS mRNA to Retinal Ischemia-
reperfusion
To confirm that this induction of iNOS mRNA was
due to ischemia-reperfusion and not to damage of the
optic nerve, we compared mRNA expression for iNOS
by RT-PCR among rat models of ligation-induced
ischemia, pressure-induced ischemia and optic nerve
damages (Fig. 2). Pressure-induced ischemia greatly
F. 2. Upregulation of iNOS mRNA expression by retinalischemia reperfusion. Shown is a representative electro-phoresis pattern of PCR products from three samples at eachtime. PCR amplification was carried out using specificprimers for iNOS (35 cycles). The figures indicate the timeafter reperfusion (hr). LI, ligation-induced ischemia; PI,pressure-induced ischemia; LOD, ligation-induced opticnerve damage; FOD, optic nerve damage produced byforceps ; OT, optic nerve transection; M, marker (φX174HaeIII fragments).
168
2000
0N
Time after reperfusion (hr)
% o
f co
ntr
ol
0.5
1000
1800
1600
1200
1400
800
600
400
200
0 0.25 1 3 6 9612 24 48
*
Time after reperfusion (hr)
168N 1/20 1/4 1 3 6 9612 24 48
β-actin
iNOS
(B)
(A)
F. 3. Quantitative RT-PCR analysis of the levels of iNOSmRNA expression in rat retina following 2 hr ischemia.Levels of iNOS mRNA expression were evaluated by a semi-quantitative RT-PCR method, as described in Materials andMethods. β-actin was used as a control. The values indicatedare means³... of 3–5 animals at each time point. *P!0±0001 versus pre-ischemic level (ANOVA) followed byScheffe test). (C) Representative electrophoresis pattern ofPCR products for iNOS mRNA. N, retina from non-operatedrats.
upregulated expression for iNOS mRNA similar to with
ligation-induced ischemia. This was not detected at
3 hr after reperfusion, but increased markedly at 12
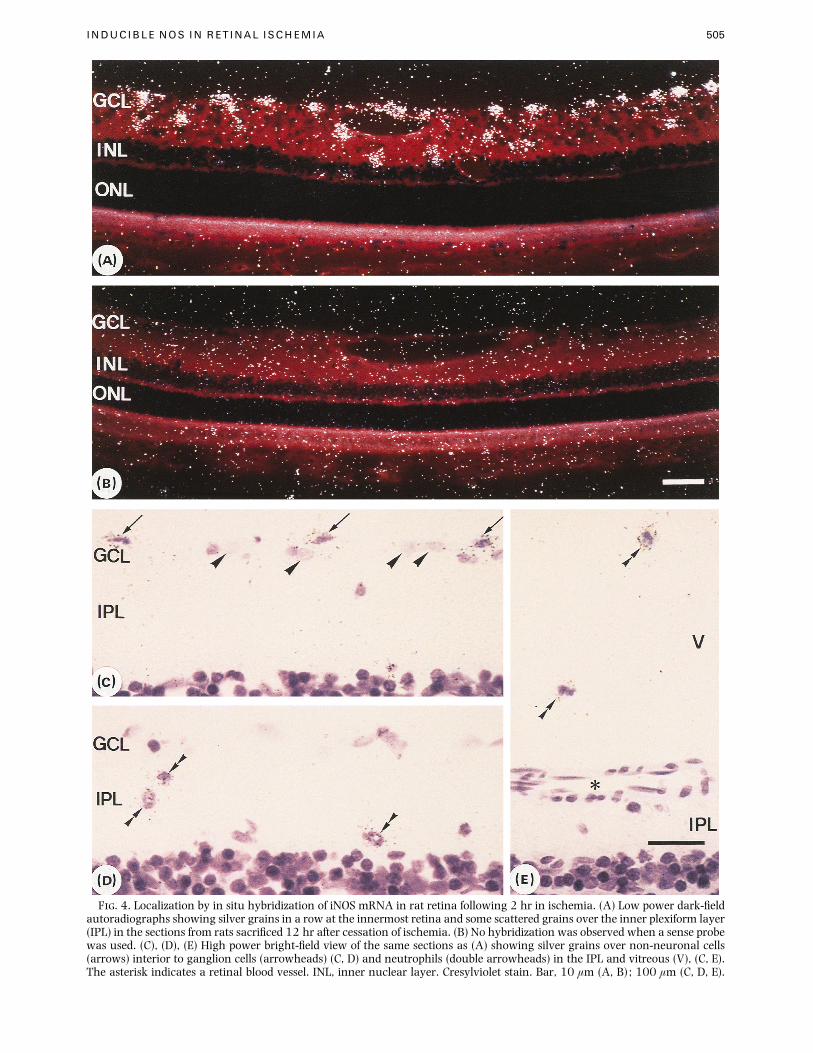
INDUCIBLE NOS IN RETINAL ISCHEMIA 505
F. 4. Localization by in situ hybridization of iNOS mRNA in rat retina following 2 hr in ischemia. (A) Low power dark-fieldautoradiographs showing silver grains in a row at the innermost retina and some scattered grains over the inner plexiform layer(IPL) in the sections from rats sacrificed 12 hr after cessation of ischemia. (B) No hybridization was observed when a sense probewas used. (C), (D), (E) High power bright-field view of the same sections as (A) showing silver grains over non-neuronal cells(arrows) interior to ganglion cells (arrowheads) (C, D) and neutrophils (double arrowheads) in the IPL and vitreous (V), (C, E).The asterisk indicates a retinal blood vessel. INL, inner nuclear layer. Cresylviolet stain. Bar, 10 µm (A, B) ; 100 µm (C, D, E).

506 M. HANGAI ET AL.
T I
Recovery rate of b-wave amplitude of ischemic eyes
Recovery rate (%)
Time (days) -NIO Control P
1 55±0³4±4 40±1³5±1 !0±053 68±6³6±6 45±8³3±5 !0±05
The values indicated are means³...
and 24 hr. In contrast, iNOS mRNA was not detected
by RT-PCR at any time in retinas following the optic
nerve injuries or optic nerve transection.
Time Course of iNOS mRNA Expression Following
Transient Retinal Ischemia
In order to monitor levels of iNOS mRNA expression,
a semi-quantitative PCR procedure was carried out
(Fig. 3) ; β-actin, which is expressed constitutively,
was used as an internal control. iNOS mRNA
expression showed no obvious changes before 6 hr,
but then rapidly increased after 12 hr (15±7-fold,
P!0±0001 vs. preischemic level, ANOVA followed by
Scheffe test). This upregulation of iNOS mRNA was
transient, as the expression declined to baseline levels
at 48 hr.
Localization of iNOS mRNA
iNOS mRNA was localized using in situ hybridi-
zation with specific riboprobes prepared from the target
GCL
(A)ILM
OPL
INL
IPL
ONL
(B) (C)
F. 5. Light micrographs of posterior rat retina. Thinning of the inner retina was observed equivalently throughout all retinalareas in both saline-treated and -NIO-treated retinas, but no apparent difference was detected between them anywhere. Shownare representative photographs taken in the region 0±5–1 mm from the optic disc. (A) Normal, uninjected retina. ILM, innerlimiting membrane; GCL, ganglion cell layer ; IPL, inner plexiform layer ; INL, inner nuclear layer ; OPL, outer plexiform layer ;ONL, outer nuclear layer. (B) Saline-treated retina 7 days after reperfusion. Note severe retinal ganglion cell (RGC) loss andatrophy of the NFL. (C) NIO-treated retina 7 days after reperfusion. Note that the RGC and NFL appear better preserved.Hematoxylin and eosin stain. Bar, 25 µm.
sequences. No iNOS mRNA expression was detected in
normal retinas, but at 12 hr after reperfusion of the
ischemic rats, iNOS mRNA was expressed predom-
inantly in the innermost retina: that is, the ganglion
cell layer (GCL) or the nerve fiber layer (NFL) [Fig.
4(A)]. In addition, occasional signals were seen in the
inner plexiform layer (IPL). There were no signals in
the inner nuclear layer (INL), the outer nuclear layer
(ONL), or the pigmented epithelium. No hybridization
was observed in parallel experiments when a sense
probe for iNOS mRNA was used [Fig. 4(B)]. High-
power bright field examination showed that at least
two types of cells expressed iNOS mRNA [Fig. 4(C)–(E)].
One type appeared to reside in the nerve fiber layer
(NFL), as they were located in a row interior to the
ganglion cells [Fig. 4(A) and (C)], although the NFL
was thinned at this time by the ischemic insult. These
cells were also densely stained by cresylviolet, and
some of them showed spindle-shaped nuclei [Fig.
4(C)]. Other types of cells had segmented or poly-
morphous nuclei and were scattered in the IPL, the
GCL and the vitreous (Fig. 4(D) and (E)].
Effects of L-NIO on the Rate of ERG b-Wave Recovery
To determine whether iNOS induction mediates
retinal ischemia-reperfusion injury, the effects of -
NIO treatment on the rate of b-wave recovery were
examined (Table I). The rate of the b-wave recovery
(mean³SEM) was 40±1³5±1% and 45±8³3±5% of
the pre-ischemic level at 24 and 72 hr, respectively, in
the control (n¯8). In -NIO treated rats, the b-wave
amplitude was 55±0³4±4% and 68±6³6±6% at 24

INDUCIBLE NOS IN RETINAL ISCHEMIA 507
and 72 hr, respectively (n¯9). This effect of -NIO
was statistically significant (P!0±05; ANOVA fol-
lowed by Scheffe test). Administration of -NIO
according to the protocol did not alter the ERG of the
fellow, nonischemic eyes.
Light Microscopic Study
To confirm the detrimental effects of iNOS induction,
light microscopy was used to examine the effect of
-NIO treatment on the histology. At 7 days after
reperfusion, thinning of the inner retina was observed
equivalently throughout all retinal areas in both
saline-treated and -NIO-treated rats. This change was
identical in the two groups throughout the entire
retina, although there was a trend that retinal
ganglion cells and the NFL were better preserved in
-NIO-treated retinas than in retinas of saline-treated
rats (Fig. 5). No remarkable changes were seen in the
outer retina of ischemic retinas of either group. No
prominent changes were observed in -NIO-treated
fellow non-ischemic eyes.
4. Discussion
This study in rats shows that iNOS mRNA is highly
expressed by non-neuronal cells of the inner retina
during reperfusion following transient retinal
ischemia. Semi-quantitative RT-PCR showed that
iNOS mRNA was transiently induced, with peak
expression at 12 hr after reperfusion (Fig. 3). This
induction of iNOS mRNA appeared to be specifically
due to ischemia-reperfusion, and was not related to
damage to the optic nerve, as it also occurred in the
eyes in which ischemia-reperfusion was produced by
elevating intraocular pressure and because it was not
detected by RT-PCR in rat retina following optic nerve
injuries or transection (Fig. 2). In situ hybridization
revealed that the iNOS mRNA was localized in the
inner retina: the GCL, NFL, IPL and the vitreous
(Fig. 4).
At least two types of cells within the inner retina
appeared to express the iNOS mRNA (Fig. 4). One
appeared to be resident retinal cells, which were
located in a row interior to the ganglion cells and
whose nuclei appeared to be smaller and more densely
stained by cresylviolet than those of the ganglion cells
[Fig. 4(A) and (C)]. Some of these cells had spindle-
shaped nuclei. From the location and morphology, we
suggest that these cells are astrocytes (Schnitzer,
1988), although microglial cells cannot be ruled out.
iNOS has been shown to be expressed by retinal glial
cells, such as Mueller cells and astrocytes in response
to lipopolysaccharide, cytokines and infection
(Dighiero et al., 1994; Goureau et al., 1994). The
other type of cells were randomly scattered in the IPL,
the GCL and the vitreous cavity [Fig. 4(D) and (E)] and
had segmented or polymorphous nuclei. It was
reported that cells infiltrating the retina during the
first 24 hr are neutrophils (Szabo et al., 1991a, b;
Hangai et al., 1995). Therefore, we suggest that cells
seen in this study are neutrophils recruited into the
ischemic retina. It has been reported that retinal
pigment epithelial cells (RPE) in culture can also
express iNOS in response to cytokines and activated T
cells (Goureau et al., 1993; Liversidge et al., 1994),
but we could not detect iNOS mRNA in RPE in this
study.
Expression of iNOS mRNA was relatively delayed
during reperfusion; it was not detected before 6 hr
after the end of ischemia (Fig. 3). We previously
showed that interleukin-1 (IL-1) mRNA expression
was upregulated during reperfusion in this same
model (Hangai et al., 1995). According to the results
of semi-quantitative RT-PCR in that study, induction
of iNOS mRNA was preceded by that of IL-1 mRNA.
Because IL-1 is one of the cytokines that can induce
iNOS (Moncada, Palmer and Higgs, 1991), it may
mediate iNOS induction in the retina following
transient ischemia. In addition, the in situ hybrid-
ization experiments showed cellular localization of
iNOS mRNA to be similar to that of IL-1β mRNA
(Hangai et al., 1995), although iNOS mRNA was not
detected in endothelial cells (Fig. 4). This temporal and
spatial correlation between IL-1 and iNOS mRNA
expression suggests that IL-1 initiates and promotes
inflammatory}immunologic events such as activation
of glial cells, and recruitment and stimulation of
neutrophils, and, as a result, induces iNOS mRNA in
these cells in an autocrine and}or paracrine manner.
Whether iNOS mediates retinal ischemia-
reperfusion injury was the next issue that we wished
to address. We found that -NIO treatment signi-
ficantly increased the rate of b-wave recovery (Table
I). -NIO is thought to be a potent and irreversible
inhibitor of inducible NOS, because it effectively
inhibits NOS activity in phagocytic cells (McCall et al.,
1991), and also inhibits immune complex injury, a
process in which iNOS is involved (Mulligan, Moncada
and Ward, 1992). The results raise the possibility that
NO generated by iNOS induced in ischemic retinas
mediates retinal ischemia-reperfusion injury. How-
ever, it cannot be denied that constitutive isoforms of
NOS were also inhibited by the -NIO treatment in this
study (Rees et al., 1990; Moore et al., 1994), and that
the inhibition of cNOS was involved in the process of
increased b-wave recovery. Further studies to confirm
this hypothesis are needed.
In contrast to our results, Ostwald et al. found that
pretreatment of -NG-nitro-arginine-methylester (-
NAME) did not alter early recovery of b-wave in cats
(Ostwald et al., 1995). The reason for this discrepancy
is uncertain in this study. One possible explanation for
this is that these differences may result from the
different time schedules of drug administration. They
infused -NAME intravenously for 30 min starting
1 hr before the ischemic insult, whereas we repeatedly
gave -NIO intraperitoneally starting 2 hr after reper-

508 M. HANGAI ET AL.
fusion. -NAME administered 1 hr before ischemia
might not effectively inhibit iNOS activity, because
iNOS mRNA expression exhibited delayed upregula-
tion (12–24 hr) after reperfusion.
The finding of b-wave recovery must be explained
with caution, as it does not necessarily represent the
entire spectrum of ischemic damage in the retina. In
fact, no prominent difference in retinal thickness was
detected by the light microscopic study (Fig. 5). The
results imply, therefore, that -NIO treatment de-
creased injury of the localized retinal tissue associated
specifically with the generation of the b-wave. In
general, heterogeneous actions of NO are thought to
exist depending upon its local redox forms. It has been
suggested that retinal ischemia-reperfusion injury
involves free radical formation (Szabo et al., 1991a, b;
Nayak, Kita and Marmor, 1993; Peachey, Green and
Ripps, 1993) and excitatory amino acid release (Yoon
and Marmor, 1989; Louzada et al., 1992). NO can
react with superoxide anion (O#
−) to generate the
highly cytotoxic free radical, peroxynitrite (dONOO−)
and hydroxyl free radical (dOH−) (Beckman et al.,
1990; Radi et al., 1991). In contrast, oxidation of NO
produces nitrosonium, which can inactivate receptors
for -methyl--aspartate (NMDA), and thus can
prevent NMDA-mediated neurotoxicity (Manzoni et
al., 1992, Lipton et al., 1993). The results do not
exclude the possibility that these heterogeneous effects
of NO are involved in the retinal ischemia-reperfusion
injury.
In conclusion, this study has shown that iNOS
mRNA is highly expressed by non-neuronal cells of the
inner retina during reperfusion following transient
retinal ischemia. It has also shown that -NIO
treatment provides some protection against ischemia-
reperfusion injury. The results suggest that nitric
oxide produced by iNOS induced during reperfusion
mediates retinal ischemia-reperfusion injury.
Acknowledgements
The authors thank Professor Thom W. Mittag (MountSinai School of Medicine, New York, NY, U.S.A.) for -NIOand Ms Hisako Okuda for her help with the histologictechniques.
This work was supported in part by a Grant-in-Aid forScientific Research from the Ministry of Education, Science,Sports and Culture of the Japanese Government.
References
Beckman, J. S., Beckman, T. W., Chen, J., Marshall, P. A.and Freeman, B. A. (1990). Apparent hydroxyl radicalproduction by peroxynitrite : implications for endo-thelial injury from nitric oxide and superoxide. Proc.Natl. Acad. Sci. U.S.A. 87, 1620–4.
Chomczynski, P. and Sacchi, N. (1987). Single-step methodof RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 162,156–9.
Corbett, J. A., Tilton, R. G., Chang, K., Hasan, K. S., Ido, Y.,Wang, J. L., Sweetland, M. A., Lancaster, J., Jr.,Williamson, J. R. and McDaniel, M. L. (1992). Amino-
guanidine, a novel inhibitor of nitric oxide formation,prevents diabetic vascular dysfunction. Diabetes 41,552–6.
Dighiero, P., Reux, I., Hauw, J. J., Fillet, A. M., Courtois, Y.and Goureau, O. (1994). Expression of inducible nitricoxide synthase in cytomegalovirus-infected glial cells ofretinas from AIDS patients. Neurosci. Lett. 166, 31–4.
Goureau, O., Lepoivre, M., Becquet, F. and Courtois, Y.(1993). Differential regulation of inducible nitric oxidesynthase by fibroblast growth factors and transforminggrowth factor beta in bovine retinal pigmented epithelialcells : inverse correlation with cellular proliferation.Proc. Natl. Acad. Sci. U.S.A. 90, 4276–80.
Goureau, O., Hicks, D., Courtois, Y. and De-Kozak, Y.(1994). Induction and regulation of nitric oxidesynthase in retinal Mueller glial cells. J. Neurochem. 63,310–7.
Hangai, M., Yoshimura, N., Yoshida, M., Yabuuchi, K. andHonda, U. (1995). Interleukin-1 gene expression intransient retinal ischemia in the rat. Invest. Ophthalmol.Vis. Sci. 36, 571–8.
Iadecola, C., Xu, X, Zhang, F., el-Fakahany, E. E. and Ross,M. E. (1995a). Marked induction of calcium-indepen-dent nitric oxide synthase activity after focal cerebralischemia. J Cereb. Blood Flow Metab. 15, 52–9.
Iadecola, C., Zhang, F. and Xu, X. (1995b). Inhibition ofinducible nitric oxide synthase ameliorates cerebral.ischemic damage. Am. J. Physiol. 268, 286–92.
Kilbourn, R. G., Jubran, A., Gross, S. S., Griffith, O. W., Levi,R., Adams, L. and Lodato, R. F. (1990). Reversal ofendotoxin-mediated shock by NG-methyl--arginine, aninhibitor of nitric oxide synthesis. Biochem. Biophys. Res.Commun. 172, 1132–8.
Kroncke, K. D., Kolb-Bachofen, V., Berschick, B., Burkart, V.and Kolb, H. (1991). Activated macrophages killpancreatic syngeneic islet cells via arginine-dependentnitric oxide generation. Biochem. Biophys. Res. Commun.175, 752–8.
Lawwill, T. (1972). Practical rabbit electroretinography.Am. J. Ophthalmol. 74, 135–41.
Lipton, S. A., Choi, Y. B., Pan, Z. H., Lei, S. Z., Chen, H. S.,Sucher, N. J., Loscalzo, J., Singel, D. J. and Stamler, J. S.(1993). A redox-based mechanism for the neuro-protective and neurodestructive effects of nitric oxideand related nitroso-compounds. Nature 364, 626–32.
Liversidge, J., Grabowski, P., Ralston, S., Benjamin, N. andForrester, J. V. (1994). Rat retinal pigment epithelialcells express an inducible form of nitric oxide synthaseand produce nitric oxide in response to inflammatorycytokines and activated T cells. Immunology 83, 404–9.
Louzada Jr, P., Dias, J. J., Santos, W. F., Lachat, J. J.,Bradford, H. F. and Coutinho-Netto, J. (1992).Glutamate release in experimental ischaemia of theretina: An approach using microdialysis. J. Neurochem.59, 358–63.
Manzoni, O., Prezeau, L., Marin, P., Deshager, S., Bockaert,J. and Fagni, L. (1992). Nitric oxide-induced blockade ofNMDA receptors. Neuron 8, 653–62.
Marchuk, D., Drumm, M., Saulino, A. and Collins, F. S.(1991). Construction of T-vectors, a rapid and generalsystem for direct cloning of unmodified PCR products.Nucleic Acid Res. 19, 1154.
McCall, T. B., Feelisch, M., Palmer, R. M. and Moncada, S.(1991). Identification of -iminoethyl--ornithine as anirreversible inhibitor of nitric oxide synthase in phago-cytic cells. Br. J. Pharmacol. 102, 234–8.
Moncada, S., Palmer, R. M. and Higgs, E. A. (1991). Nitricoxide : physiology, pathophysiology, and pharm-acology. Pharmacol. Rev. 43, 109–42.
Moore, W. M., Webber, R. K., Jerome, G. M., Tjoeng, F. S.,

INDUCIBLE NOS IN RETINAL ISCHEMIA 509
Misko, T. P. and Currie, M. G. (1994). -N6-(1-iminoethyl)lysine : a selective inhibitor of induciblenitric oxide synthase. J. Med. Chem. 37, 3886–8.
Mulligan, M. S., Moncada, S. and Ward, P. A. (1992).Protective effects of inhibitors of nitric oxide synthase inimmune complex-induced vasculitis. Br. J. Pharmacol.107, 1159–62.
Nayak, M. S., Kita, M. and Marmor, M. F. (1993). Protectionof rabbit retina from ischemic injury by superoxidedismutase and catalase. Invest. Ophthalmol. Vis. Sci. 34,2018–22.
Nudel, U., Zakut, R., Shani, M., Neuman, S., Levy, Z. andYaffe, D. (1983). The nucleotide sequence of the ratcytoplasmic beta-actin gene. Nucleic Acid Res. 11,1759–71.
Ostwald, P., Goldstein, I. M., Pachnanda, A. and Roth, S.(1995). Effects of nitric oxide synthase inhibition onblood flow after retinal ischemia in cats. Invest.Ophthalmol. Vis. Sci. 36, 2396–403.
Peachey, N. S., Green, D. J. and Ripps, H. (1993). Ocularischemia and the effects of allopurinol on functionalrecovery in the retina of the arterially perfused cat eye.Invest. Ophthalmol. Vis. Sci. 34, 58–65.
Radi, R., Beckman, J. S., Bush, K. M. and Freeman, B. A.(1991). Peroxynitrite oxidation of sulfhydryls. Thecytotoxic potential of superoxide and nitric oxide. J.Biol. Chem. 266, 4244–50.
Rees, D. D., Palmer, R. M., Schulz, R., Hodson, H. F. andMoncada, S. (1990). Characterization of three inhibitorsof endothelial nitric oxide synthase in vitro and in vivo.Br. J. Pharmacol. 101, 746–52.
Saiki, R. K., Gelfand, D. H., Stoffel, S., Scharf, S. J., Higuchi,R., Horn, G. T., Mullis, K. B. and Erlich, H. A. (1988).Primer-directed enzymatic amplification of DNA with athermostable DNA polymerase. Science 239, 487–91.
Sanger, F., Nicklen, S. and Coulson, A. R. (1977). DNAsequencing with chain-terminating inhibitors. Proc.Natl. Acad. Sci., U.S.A. 74, 5463–7.
Schnitzer, J. (1988). Astrocytes in the guinea pig, horse, andmonkey retina: their occurrence coincides with thepresence of blood vessels. Glia 1, 74–89.
Spivey, B. E. and Pearlman, J. T. (1953). Day-to-dayvariations in the ERG of humans and rabbits. Am. J.Ophthalmol. 55, 1013–20.
Stefansson, E., Wilson, C. A., Schoen, T. and Kuwabara, T.(1988). Experimental ischemia induces cell mitosis inthe adult rat retina. Invest. Ophthalmol. Vis. Sci. 29,1050–5.
Steinhorst, U. H., Chen, E. P., Hatchell, D. L., Samsa, G. P.,Saloupis, P. T., Westendorf, J. and Machemer, R.(1993). Aclacinomycin A in the treatment of ex-perimental proliferative vitreoretinopathy. Efficacy andtoxicity in the rabbit eye. Invest. Ophthalmol. Vis. Sci. 34,1753–60.
Szabo, M. E., Droy-Lefaix, M. T., Doly, M. and Braquet, P.(1991a). Free radical-mediated effects in reperfusioninjury: a histologic study with superoxide dismutaseand EGB 761 in rat retina. Ophthalmic Res. 23, 225–34.
Szabo, M. E., Droy-Lefaix, M. T., Doly, M., Carre, C. andBraquet, P. (1991b). Ischemia and reperfusion-inducedhistologic changes in the rat retina. Demonstration of afree radical-mediated mechanism. Invest. Ophthalmol.Vis. Sci. 32, 1471–8.
Wood, E. R., Berger, H., Jr., Sherman, P. A. and Lapetina,E. G. (1993). Hepatocytes and macrophages express anidentical cytokine inducible nitric oxide synthase gene.Biochem. Biophys. Res. Commun. 191, 767–74.
Yoon, Y. H. and Marmor, M. F. (1989). Dextromethorphanprotects retina against ischemic injury in vivo. Arch.Ophthalmol. 107, 409–11.