indirect mechanism of lead-induced genotoxicity in cultured mammalian cells
TRANSCRIPT

Mutation Research, 241 (1990) 75-82 75 Elsevier
MUTGEN 01543
Indirect mechanism of lead-induced genotoxicity in cultured mammalian cells
Andrea Hartwig, Regina Schlepegrell and Detmar Beyersmann Biochemistry Group, Department of Biology and Chemistry, University of Bremen, D-2800 Bremen 33 (F.R.G.)
(Received 6 October 1989) (Revision received 4 December 1989)
(Accepted 11 December 1989)
Keywords: Lead; UV; HPRT mutation assay; Sister-chromatid exchange; Comutagenicity; DNA-repair inhibition
Summaff
The data concerning the mutagenic, clastogenic and carcinogenic properties of inorganic lead com- pounds have been conflicting. To investigate whether the genotoxicity of lead is due to indirect effects such as interference with DNA-repair processes, the induction of mutations, sister-chromatid exchanges and strand breaks by lead ions alone as well as in combination with UV light as a standard mutagen were determined. Lead acetate alone does not induce DNA-strand breaks in HeLa cells or mutations at the H P R T locus and sister-chromatid exchanges in V79 Chinese hamster cells. However, at all endpoints tested, lead ions interfere with the processing of UV-induced D N A damage. They inhibit the closing of DNA-st rand breaks after UV irradiation and enhance the number of UV-induced mutations and sister-chromatid exchanges, indicating an inhibition of D N A repair. These data point out the necessity to consider such indirect effects when assessing the genotoxicity of metal compounds. As possible mecha- nisms of repair inhibition we suggest either the interaction with repair enzymes such as polymerase or ligase or else the interaction with calcium-regulated processes, for example with calmodulin.
For many centuries, the adverse health effects to humans of lead and its compounds have been recognized. Even though the environmental pollu-
Abbreviations: BrdU, 5-bromodeoxyuridine; CFA, colony-forming ability; DMSO, dimethyl sulfoxide; EDTA, ethylene diaminotetraacetic acid; HPRT, hypoxanthine- guanine-phosphoribosyltransferase; Hoechst 33258, bisbenzi- mide trihydrochloride; MEM, minimal essential medium; SCE, sister-chromatid exchange; Triton X-100, octyl phenoxy poly- ethoxy ethanol.
Correspondence: Dr. Andrea Hartwig, Biochemistry Group, Department of Biology and Chemistry, University of Bremen, D-2800 Bremen 33 (F.R.G.).
tion will become less mainly due to the use of unleaded gasoline, lead is still present in many workplaces and also in the environment because of its persistence. Although the toxic effects of lead compounds have been studied for many years, inconsistent results have been published about their mutagenic, clastogenic and carcinogenic properties. Inorganic ' lead compounds are classi- fied as possibly carcinogenic to humans by the International Agency for Research on Cancer (IARC, 1987). The evidence for carcinogenicity to humans is still inadequate. While Dingwall-For- dyce and Lane (1963) did not find an excess of overall cancer deaths in workers of lead-related
0165-1218/90/$03.50 © 1990 Elsevier Science Publishers B.V. (Biomedical Division)

76
industries, Cooper and Gaffey (1975) reported an elevated number of deaths from all malignant neoplasms in smelter workers but not in battery plant workers; reevaluated by the IARC, these data were found to be not significant (IARC, 1980). However, inorganic lead compounds have been shown to cause renal tumors in experimental animals after different routes of delivery as well as other tumors (reviewed by IARC, 1980). Addition- ally, lead acetate, lead subacetate and lead oxide enhanced the number of tumors induced by some organic compounds, including 2-(ethylnitrosa- mine)ethanol (Shirai et al., 1984) and N-(4'-flu- oro-4-biphenyl)-acetamide (Tanner and Lipsky, 1984). Much work has been done determining the clastogenicity of lead compounds by investigating chromosomal aberrations in lymphocytes of lead- exposed workers. However, the results remain contradictory, which might in part be due to dif- ferences in culture conditions (Gebhardt and Rossman, 1989). Furthermore, one question that still needs clarification is whether inorganic lead compounds exert clastogenic effects themselves or whether they enhance chromosomal aberrations induced by compounds occurring simultaneously a n d / o r arising during cell culturing (Deknudt and Lronard, 1975; Nordenson and Beckman, 1982; Forni et al., 1980). In a previous report, both soluble and insoluble lead compounds were shown to be mutagenic at the HPRT locus after long-term exposure of V79 cells; additionally, the number of cell transformations in Syrian hamster embryo cells was enhanced after treatment with lead acetate. But again the mode of action remained unclear, since no direct DNA damage such as strand breaks or DNA-protein crosslinks was de- tected (Zelikoff et al., 1988). On the other hand, lead ions were shown to decrease the fidelity of DNA synthesis (Sirover and Loeb, 1976) and to inhibit the DNA polymerase fl in vitro (Popenoe and Schmaeler, 1979). Therefore, we investigated whether the genotoxic potential might be due rather to indirect effects, such as interference with DNA-repair processes. We tested lead ions alone as well as in combination with UV as a standard mutagen. For the determination of mutations at the HPRT locus as well as sister-chromatid ex- changes, the well-established V79 Chinese hamster system was employed. For the detection of strand
breaks during UV repair a human cell line (HeLa) was used, because these cells have been shown to exert a much higher incision rate, which is due to their higher overall repair capacity towards UV- induced photoproducts (Johnson et al., 1984).
Materials and methods
Cell culture Chinese hamster V79 cells were grown as
monolayers in minimal essential medium (MEM), a modified, containing 10% fetal calf serum, 100 units penicillin/ml, 100 /~g s t reptomycin/ml and 0.0013 t~g fungizone (Squibb)/ml. HeLa cells were cultured in MEM supplemented with 5% fetal calf serum and antibiotics as described for V79 cells. All cultures were incubated at 37 °C with 5% CO 2 in air and 100% humidity.
Materials MEMa, MEM, the trypsin, penicillin-strep-
tomycin and Colcemid solutions are products of Gibco, Karlsruhe. 5-Bromodeoxyuridine, bisben- zimidetrihydrochloride (Hoechst 33258) and 2- amino-6-mercaptopurine (6-thioguanine) were ob- tained from Sigma, Munich. Fetal calf serum was purchased from Gibco, Karlsruhe or Biochrom KG, Berlin and assayed for minimum mutation background. Triton X-100 was bought from Pierce, Oud-Beijerland (The Netherlands). All other chemicals, including lead acetate ((CH3COO)2Pb × 2 HzO, Cat. No. 7375) and Giemsa stain, were of p.a. grade and were from Merck, Darmstadt. The culture dishes were supplied by Nunc, Wiesbaden. 6-Thioguanine was acidified with 0.1 N HC1 before diluting with DMSO and bidistilled water (1 :2 v/v) . Lead acetate was dissolved in bidistilled water. All solutions were made up freshly before use. The cells were irradiated with UV as monolayers in petri dishes without cover and medium.
Cytotoxicity Cytotoxicity was determined as colony-forming
ability (CFA). 300 cells/100-mm dish were al- lowed to attach for 5 h and treated with lead acetate for 24 h. After 6 days (V79 cells) or 10 days (HeLa cells) of incubation, colonies were

77
fixed with ethanol (96%), stained with Giemsa (25% in ethanol), counted and calculated as % of control. All values were determined in triplicate and averaged.
Mutation assay The procedure is based mainly on the H P R T
method of Hsie et al. (1979). 5 h after seeding 5 x 105 V79 cel ls /100-mm dish they were treated with lead acetate for 20 h, irradiated with 5 J / m 2 UV and postincubated for 24 h with lead acetate. During the following expression time, the cells were subcultured twice with 3-days intervals. The first subculture took place 48 h after seeding; at this time, the cytotoxicity was determined. First of all, the cells were counted and calculated as % of control to detect growth inhibition. Secondly, the cells were diluted to 200 cel ls /100-mm dish to determine CFA. After 5 -7 days, the colonies were fixed with ethanol (96%), stained in Giemsa solu- tion (25% in ethanol) and again calculated as % of control. For selection, the cells were trypsinized and reseeded at a density of 2 x 105 cel ls /100-mm dish, taking 5 dishes for each condition. The selec- tive medium contained 10 ttg 6-thioguanine/ml. At the same time, the cells were diluted to about 200 cel ls /100-mm dish in triplicate and incubated in growth medium to test for viability. After 7 days, these colonies as well as mutant colonies were fixed and stained. The mutation frequency was calculated as both mutants /106 seeded cells and mutants /106 viable cells. The complete ex- periment shown in Fig. 2 was carried out twice with essentially the same result.
Sister-chromatid exchange (SCE) test 5 h after seeding 105 V79 cel ls /100-mm dish
they were preincubated with lead acetate where indicated and after this period irradiated with UV. Afterwards, the cells were incubated simulta- neously with 5-bromodeoxyuridine (BrdU, 5 /~g/ml) and lead acetate for 26 h. During this time and during the following 2-h incubation with Col- cemid (0.04 /~g/ml), the dishes were wrapped in aluminum foil to keep the cells in the dark. Mitotic cells were harvested by pipetting, centrifuged at 200 x g for 5 rain, resuspended in 0.56% KC1 and centrifuged again. Then they were fixed twice in freshly prepared methanol /acet ic acid (3:1) for
15 min. Air-dried slides were stained 24 h later by the method of Goto et al. (1978). The slides were incubated for 15 rain in Hoechst 33258 (1 x 10 -5 M) at room temperature, mounted with buffer (0.075 M NaC1 and 0.0075 M sodium citrate), pH 8, and irradiated with UV, 2 x 15 W, 365 nm, at 50 o C for 30 rain from a distance of 7 cm. Finally, they were stained in 3% Giemsa solution buffered with phosphate. The complete experiment was carried out twice with essentially the same results.
Nucleoid sedimentation The procedure is based on the method of Cook
and Brazell (1976) with modifications described by Yew and Johnson (1979). 3-5 × 105 HeLa cells were lysed in the presence of 2 M NaC1, 0.01 M EDTA, pH 8 and 0.5% Triton X-100 and centri- fuged down a 15-30% sucrose gradient containing 2 M NaC1, 0.01 M EDTA and 1 /~g/ml Hoechst 33258 dye at 8000 rpm for 30 rain. The position of the D N A nucleoid band was determined under ultraviolet light and calculated as distance sedi- mented compared to control cells. Each point was averaged from 2 independent treatments and the experiments were repeated at least once.
Results
Cell culture, cytotoxicity and uptake To investigate the mechanisms of lead toxicity
in in vitro test systems, the strong dependence of the action of metal compounds on cell culture conditions has to be considered (Costa and Heck, 1986). Therefore, much care was taken to de- termine appropriate incubation conditions. Com- paring the cytotoxicity measured as CFA after incubation with lead acetate for 24 h, we found that V79 cells are much more sensitive to lead ions than HeLa cells (Table 1). With HeLa cells, it takes about 600/~M to reduce the CFA to 50% of the control. With V79 cells, the same reduction is generated by 5-10 ~tM lead acetate. To investigate whether this effect is due to the impermeabili ty of the cytoplasm membrane of HeLa cells to lead ions, as proposed by Frenkel and Middleton (1987), the uptake of lead acetate into both cell types was measured by atomic absorption spec- trometry. Preliminary results show, however, an even higher uptake in HeLa cells compared to V79
78
TABLE 1
CYTOTOXICITY OF LEAD ACETATE IN HeLa A N D V79 CELLS D E T E R M I N E D BY CFA
lOO
Cell line Lead acetate (#M) CFA (% of control) ~. 80
V79 1 92.0 3 53.1 ~ 60 5 41.5
40 i 10 37.5 25 32.1 "~ 20
HeLa 100 76.1 "~ 0 500 66.3
1000 26.5 2000 14.7
For treatment conditions, see Materials and methods.
cells. Furthermore, almost all of the lead is inter- nalized, since only about 10% can be removed from the outer membrane by EDTA. However, studies of the kinetics of lead uptake indicate that it takes 20-24 h of incubation before considerable amounts are taken up (details on the kinetics of lead uptake in different cell types will be pub- lished elsewhere). Therefore, we chose long prein- cubation times prior to UV irradiation to insure that lead ions are inside the cell at the time of irradiation. Regarding the 2 different cell lines, we applied about equitoxic concentration ranges of lead acetate.
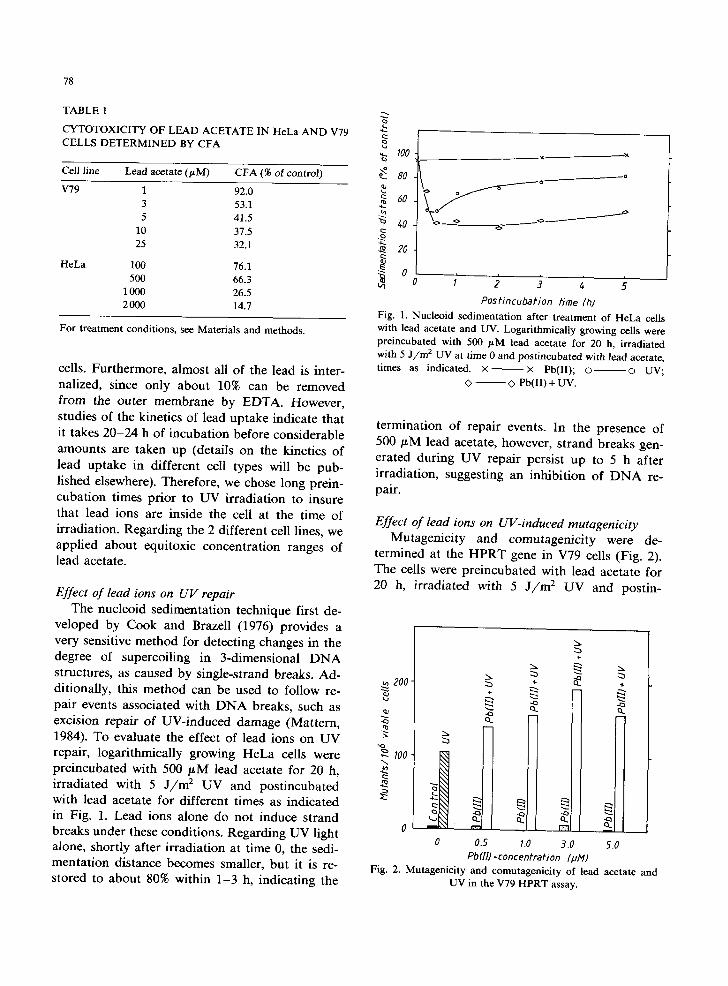
Effect of lead ions on UV repair The nucleoid sedimentation technique first de-
veloped by Cook and Brazell (1976) provides a very sensitive method for detecting changes in the degree of supercoiling in 3-dimensional D N A structures, as caused by single-strand breaks. Ad- ditionally, this method can be used to follow re- pair events associated with D N A breaks, such as excision repair of UV-induced damage (Mattern, 1984). To evaluate the effect of lead ions on UV repair, logarithmically growing HeLa cells were preincubated with 500 /LM lead acetate for 20 h, irradiated with 5 J / / m 2 UV and postincubated with lead acetate for different times as indicated in Fig. 1. Lead ions alone do not induce strand breaks under these conditions. Regarding UV light alone, shortly after irradiation at time 0, the sedi- mentation distance becomes smaller, but it is re- stored to about 80% within 1-3 h, indicating the
~ . ~ . _ . _ . - ~ o ~
o
Postincubation time (h) Fig. 1. Nucleoid sedimentation after t reatment of HeLa cells with lead acetate and UV. Logarithmically growing cells were preincubated with 500 laM lead acetate for 20 h, irradiated with 5 J / m 2 UV at time 0 and postincubated with lead acetate, times as indicated. × ~ x Pb(II); o o UV;
<> ~ Pb(II) + UV.
termination of repair events. In the presence of 500 #M lead acetate, however, strand breaks gen- erated during UV repair persist up to 5 h after irradiation, suggesting an inhibition of D N A re- pair.
Effect of lead ions on UV-induced mutagenicity Mutagenicity and comutagenicity were de-
termined at the H P R T gene in V79 cells (Fig. 2). The cells were preincubated with lead acetate for 20 h, irradiated with 5 J / m 2 UV and postin-
200
~ 100
2~
-x
+
t~
0.5 10 3.0 Pb(H)-concentrafion [pl~)
5.0
2~
4:3
Fig. 2. Mutagenicity and comutagenicity of lead acetate and UV in the V79 HPRT assay.

TABLE 2
CYTOTOXICITY AFTER SINGLE AND COMBINED TREATMENT OF V79 CELLS WITH UV AND LEAD ACETATE
Treatment Cell number CFA (% (% of control) of control)
UV 61.7 82.1
0.5/~M Pb(lI) 98.8 118.1 0.5/LM Pb(II) + UV 66.7 55.8
1.0 ~M Pb(II) 96.3 102.1 1.0 ~M Pb(II)+UV 61.7 66.1
3.0 ~M Pb(II) 74.1 66.9 3.0 ~ M Pb(II) + UV 37.0 47.3
5.0/xM Pb(II) 74.1 90.2 5.0 ,ttM Pb(lI) + UV 35.8 62.6
Cell number and CFA were determined at the time of the first subculture of the HPRT assay, as described in Materials and methods.
cuba ted with lead aceta te for 24 h, concent ra t ions as indica ted . N o mutagen ic i ty is de tec ted by lead alone, bu t lead ions enhance the UV- induced mutagenic i ty . I t has to be emphas ized that the comutagen ic effect is ob ta ined at very low con- cent ra t ions , be tween 0.5 and 5.0 ~ M Pb(II) . Higher concen t ra t ions up to 100 # M lead aceta te fail to exert comutagen ic i ty (da t a not shown). There is also a marked decrease in cell n u m b e r at the t ime of the first subcul ture at concent ra t ions as low as
TABLE 3
EFFECT OF LEAD IONS ON UV-INDUCED SCEs IN V79 CELLS
Treatment SCEs/cell + SD
Control 8.87 _+ 4.35 UV 23.40 + 8.10
1/zM Pb(II) 8.88 + 4.56 1 btM Pb(lI) + UV 36.68 + 15.28
5/~M Pb(II) 8.72+ 3.03 5/LM Pb(II) + UV 34.20 + 16.38
10 ~M Pb(II) 10.44+ 4.04 10 #M Pb(II) + UV 34.40 _+ 12.35
p < 0.01 a
p < 0.01 a
NS p < 0.01 a
Logarithmically growing cells were incubated as described in Materials and methods. 25 mitotic cells were counted for each value. a Determined by t-test. NS, not significant (t-test).
79
3 /~M Pb(II ) plus 5 J / m z U V (Table 2), which might be due to ei ther g rowth inh ib i t ion a n d / o r the dea th of a cons iderab le pa r t of the cells. The remain ing cells, however, exhibi t a compara t i ve ly high c loning efficiency.
Effect of lead ions on UV-induced SCEs A p p l y i n g bas ica l ly the same cond i t ions as in
the mu ta t i on assay, s i s t e r -chromat id exchanges were de t e rmined in V79 cells. UV light increases the number of spon taneous SCEs f rom 8.9 to 23.4. Lead ions a lone cause only a smal l increase in SCEs at the highest concen t ra t ion app l i ed (10 /zM), which is not s ignif icant as de t e rmined by t-test. However , at all concen t ra t ions tested, l ead s ignif icant ly enhances the n u m b e r of U V - i n d u c e d SCEs, y ie ld ing p r o n o u n c e d har lequ in ch romo- somes (Table 3).
Discussion
Culture conditions, cytotoxicity and uptake The b ioava i lab i l i ty of heavy meta ls is s t rongly
med ia t ed by phys ico-chemica l factors l ike the pH, the presence or absence of che la t ing agents, cer- ta in anions and cat ions, as reviewed by Babich et al. (1985). Us ing cells in cul ture, the up take a n d / o r cy to toxic i ty might be inf luenced by med ia com- pos i t ion and also by the k ind and a m o u n t of serum used (Kussel et al., 1978). In the case of lead ions, it seems i m p o r t a n t to conf i rm the up- take of the lead c o m p o u n d tes ted under the same condi t ions as used for geno tox ic i ty tests, espe- cial ly in cons idera t ion of the incons i s ten t results ob ta ined in shor t - te rm tests r epor t ed in the l i tera- ture. However , the dif ferences in cy to tox ic i ty ob- served with H e L a and V79 cells are not caused by a lower up take of lead aceta te b y H e L a cells. Since lead ions are ab le to b ind to m a n y di f ferent cell componen t s (such as prote ins , etc.), their in- t racel lu lar d i s t r ibu t ion might be different . S imilar observa t ions were m a d e by F ischer (1985); she found Chinese hams te r B14F28 cells to be much more sensit ive to CdC12 than H e L a cells, while the meta l was accumula ted even more in the second cell line.
DNA-strand breaks Lead ions a lone do not induce s t r and b reaks
under the cond i t ions app l i ed for the nuc leo id sedi-

80
mentation assay in HeLa cells. This is in agree- ment with a previous report, where lead acetate failed to induce single-strand breaks, D N A - D N A and DNA-protein crosslinks in V79 cells de- termined by the alkaline elution technique (Zeli- koff et al., 1988). The kinetics of the appearance and disappearance of strand breaks after UV irradiation resemble very closely those observed by Mattern (1984) in cultured mammalian cells and Holmberg (1989) in human lymphocytes in the nucleoid sedimentation and alkaline unwind- ing assay, respectively. Possibly, the breaks are generated during the repair of the (6-4)-photo- product, which was shown to be repaired to 80% during the first 6 h after irradiation, whereas the loss of antibody-binding sites to the second and most frequent photoproduct, the pyrimidine di- mer, occurs much more slowly in Chinese hamster ovary cells (Mitchell et al., 1988). In the presence of lead ions, the completion of these early repair events seems to be inhibited. Since incisions still take place, either the polymerase or the ligase step of excision repair may be affected. In contrast, no inhibition of pyrimidine dimer removal by lead ions in HeLa cells was found by Snyder et al. (1989), as determined by chromatography. How- ever, this discrepancy might be due to differences in incubation conditions, e.g., the lack of long preincubation times in the latter study.
Mutations at the HPRT locus In our experiments, no mutagenicity was ob-
served with lead acetate alone, which is in accor- dance with results presented by Amacher and Paillet (1980), where lead acetate did not enhance the frequency of spontaneous trifluorothymidine- resistant mutants in L 5 1 7 8 Y / T K +/- cells. In con- trast, our findings do not support the mutagenicity of lead nitrate reported previously by Zelikoff et al. (1988) after a 5-day exposure of V79 cells, which may be due to the shorter incubation period in our present study. The comutagenic effect is very likely a consequence of the inhibition of D N A repair demonstrated in HeLa cells with the nucleoid sedimentation assay. Even though there is still a debate about the relative importance of pyrimidine dimers and (6-4)-photoproducts in- duced by UV light at 254 nm in terms of cyto- toxicity and mutagenicity (Mitchell et al., 1988),
almost all UV-induced mutants have been shown to occur at dipyrimidine sites, which are sites where either photoproduct could have been formed (Vrieling et al., 1989). Therefore, inhibition of excision repair should lead to an enhanced num- ber of mutants. The lack of comutagenicity at higher concentrations of Pb(II) is likely to be a result of cytotoxicity, where potential mutants are killed and are no longer detectable in this test system, whereas the cells that survive the treat- ment resemble those that are damaged less.
Sister-chromatid exchanges The inability of lead ions to induce significant
numbers of SCEs has been reported earlier (Beek and Obe, 1975; Zelikoff et al., 1988). The en- hancement of UV-induced SCEs by lead acetate confirms the results obtained for mutations. Fur- thermore, it strengthens the thesis of the inhibition of D N A repair by lead ions, since there is good concordance between the number of remaining pyrimidine dimers and the number of SCEs (Natarajan, 1982). Enhanced frequencies of SCEs have also been observed in cells from patients with Bloom's syndrome, who are thought to have a D N A ligase I deficiency (Willis and Lindahl, 1987). On the other hand, an inhibition in poly- merase activity should lead to corresponding ef- fects, since the number of UV-induced SCEs was enhanced by the known polymerase inhibitor ara- binofuranosylcytosine (Ishii and Bender, 1980).
Possible mechanisms of repair inhibition by lead The inhibition of D N A repair as one mecha-
nism of genotoxicity is supported by several papers concerning the interaction of lead ions with en- zymes involved in D N A processing and repair. In 1976, Sirover and Loeb showed a decreased fidel- ity of the D N A polymerase from avian myelob- lastosis virus (AMV) in the presence of lead chlo- ride; however, the concentration of 4 mM applied was very high. Frenkel and Middleton (1987) re- ported an inhibition of polymerase a and RNA polymerase II isolated from HeLa cells between 80 and 150 /LM lead acetate. No inhibition of D N A or RNA synthesis, however, was found in intact HeLa cells. Popenoe and Schmaeler (1979) demonstrated an inhibition of isolated human polymerase fl at concentrations as low as 10 /~M

81
lead nitrate. Fur thermore , 250 /~M lead chloride inhibi ted the repair of X-ray- induced D N A damage measured as thymidine uptake in HeLa
cells (Skreb and Habaz in-Novak , 1977). However, besides inhibi t ing repair enzymes di-
rectly, lead ions might also interfere with calcium-regulated processes involved in the regu- la t ion of D N A replicat ion and repair. For exam- ple, specific an t i -ca lmodul in agents have been
shown to inhibi t the repair of UV- induced pyr imidine dimers (Charp and Regan, 1985) and
to potent ia te the lethality of b leomycin (Chafou- leas et al., 1984). The manifo ld interact ions be- tween lead and calcium concern ing uptake, calcium homeostasis and subst i tu t ion of Pb(II) for
Ca(II) have been reviewed by Pounds (1984). The results presented in this paper point out
the necessity of consider ing these indirect effects when assessing the genotoxic potent ia l of lead compounds . No t only the D N A damage itself bu t also the genotoxic effects of other chemical or physical agents may be augmented when repair is
inhibi ted, as i l lustrated by the enhancement of
UV- induced muta t ions and SCEs. Compar ing the data obta ined for lead ions with
those observed for other metal compounds , similar results in m a m m a l i a n cells were reported for Ni(I I ) (Hartwig and Beyersmann, 1989a), Cd(II) and Cr(VI) (Hartwig and Beyersmann, 1989b). In ad-
di t ion, an inh ib i t ion of the repair of D N A damage caused by N-methyl -N-ni t rosourea by sodium
arsenite has been reported by Li and Rossman (1989). Therefore, an interference with D N A re- pair processes has to be considered as an im- por tan t general mechanism of metal genotoxicity.
Acknowledgement
This work was supported by G r a n t 01 H K 9369 of the Bundesminis ter fiJr Forschung and Tech-
nologie, Bonn.
References
Amacher, D.E., and S.C. Paillet (1980) Induction of trifluoro- thymidine-resistant mutants by metal ions in L5178Y/ TK +/- cells, Mutation Res., 78, 279-288.
Babich, H., M.A. Devanas and H. Stotzky (1985) The media- tion of mutagenicity and clastogenicity of heavy metals by physicochemical factors, Environ. Res., 37, 253-286.
Beek, B., and G. Obe (1975) The human leukocyte test system. VI. The use of sister chromatid exchanges as possible indicators for mutagenic activities, Humangenetik, 29, 127- 134.
Chafouleas, J.G., W.E. Bolton and A.R. Means (1984) Potenti- ation of bleomycin lethality by anticalmodulin drugs: a role for calmodulin in DNA repair, Science, 224, 1346-1348.
Charp, P.A., and J.D. Regan (1985) Inhibition of DNA repair by trifluoperanzine, Biochim. Biophys. Acta, 824, 34-39.
Cook, P.R., and I.A. Brazell (1976) Detection and repair of single-strand breaks in nuclear DNA, Nature (London), 263, 679-682.
Cooper, W.C., and W.R. Galley (1975) Mortality of lead workers, J. Occup. Med., 17, 100-107.
Costa, M., and J.D. Heck (1986) Metal ion carcinogenesis: mechanistic aspects, in: H. Sigel (Ed.), Metal Ions in Bio- logical Systems, Vol. 20, Marcel Dekker, New York, Basel, pp. 259-278.
Deknudt, G., and A. L6onard (1975) Cytogenetic investiga- tions on leukocytes of workers from a cadmium plant, Environ. Physiol. Biochem., 5, 319-327.
Dingwall-Fordyce, I., and R.E. Lane (1963) A follow-up study of lead workers, Br. J. Ind. Med., 20, 313-315.
Fischer, A.B. (1985) Factors influencing cadmium uptake and cytotoxicity in cultured cells, Xenobiotica, 15, 751-757.
Forni, A., A. Sciame, B.A. Bertazzi and L. Alessio (1980) Chromosome and biochemical studies in women occupa- tionally exposed to lead, Arch. Environ. Health, 35, 139- 146.
Frenkel, G.D., and C. Middleton (1987) Effects of lead acetate on DNA and RNA synthesis by intact HeLa cells, isolated nuclei and purified polymerases, Biochem. Pharmacol., 36, 265-268.
Gebhart, E., and T.G. Rossman (1989) Mutagenicity, carcino- genicity and teratogenicity, in: E. Merian (Ed.), Metals and their Compounds in the Environment, VCH Verlagsgesell- schaft, Weinheim, in press.
Goto, K., S. Maeda, Y. Kano and T. Sugiyama (1978) Factors involved in differential Giemsa-staining of sister-chro- matids, Chromosoma, 66, 351-359.
Hartwig, A., and D. Beyersmann (1989a) Enhancement of UV-induced mutagenesis and sister-chromatid exchanges by nickel ions in V79 cells: evidence for inhibition of DNA repair, Mutation Res., 217, 65-73.
Hartwig, A., and D. Beyersmann (1989b) Comutagenicity and inhibition of DNA repair by metal ions in mammalian cells, Biol. Trace Elem. Res., 21, 359-365.
Holmberg, M. (1989) The effect of deoxynucleosides on repair of DNA breaks in UVC-irradiated human lymphocytes, Mutation Res., 218, 33-39.
Hsie, A.W., D.B. Couch, J.P. O'Neill, J.R.S. Sebastian, P.A. Brimer, R. Machanoff, J.C. Riddle, J.C. Li, J.C. Fuscoe, N. Forbes and M.H. Hsie (1979) Utilization of a quantitative mammalian cell mutation system, CHO/HGPRT, in ex: perimental mutagenesis and genetic toxicology, in: B.E. Butterworth (Ed.), Strategies for Short-term Testing for Mutagens/Carcinogens, CRC Press, Palm Beach, FL, pp. 39-54.

82
IARC (1980) Monographs on the Evaluation of the Carcino- genic Risk of Chemicals to Humans, Vol. 23, IARC, Lyon, pp. 1-210.
IARC (1987) Monographs on the Evaluation of the Carcino- genic Risk of Chemicals to Humans, Suppl. 7, IARC, Lyon, pp. 230-232.
Ishii, Y., and M.A Bender (1980) Effects of inhibitors of DNA synthesis on spontaneous and ultraviolet light-induced sis- ter-chromatid exchanges in Chinese hamster cells, Mutation Res., 79, 19-32.
Johnson, R.T., A.R.S. Collins, G.C. Elliott and S. Squires (1984) Comparative studies using inhibitors, in: A.R.S. Collins, C.S. Downes and R.T. Johnson (Eds.), DNA Re- pair and its Inhibition, IRL Press, Oxford, pp. 255-277.
Kussel, M., S. O'Cheskey and L.E. Gerschenson (1978) Cellu- lar and molecular toxicology of lead. I. Effect of lead on cultured cell proliferation, J. Toxicol. Environ. Health, 4, 503-513.
Li, J.H., and T.G. Rossman (1989) Mechanism of comutagene- sis of sodium arsenite with N-methyl-N-nitrosourea, Biol. Trace Elem. Res., 21, 373-381.
Mattern, M.R. (1984) The relation of three-dimensional DNA structure to D N A repair as studied by nucleoid sedimenta- tion, in: A.R.S. Collins, C.S. Downes and R.T. Johnson (Eds.), DNA Repair and its Inhibition, IRL Press, Oxford, pp. 35-50.
Mitchell, D.L., R.M. Humphrey, G.M. Adair, L.H. Thompson and J.M. Clarkson (1988) Repair of (6-4)-photoproducts correlates with split-dose recovery in UV-irradiated normal and hypersensitive rodent cells, Mutation Res., 193, 53-63.
Natarajan, A.T. (1982) Relationship between sister-chromatid exchanges and chromosomal aberrations, in: D. Miiller, A.T. Natarajan, G. Obe and G. RiShrborn (Eds.), Sister- Chromatid Exchange Test, Georg Thieme Verlag, Stuttgart, New York, pp. 6-10.
Nordenson, I., and L. Beckman (1982) Occupational and en- vironmental risks in and around a smelter in northern Sweden. VII, Hereditas, 96, 175-181.
Popenoe, E.A., and M.A. Schmaeler (1979) Interaction of human polymerase fl with ions of copper, lead, and cadmium, Arch. Biochem. Biophys., 196, 109-120.
Pounds, J.G. (1984) Effect of lead intoxication on calcium homeostasis and calcium-mediated cell function: a review, Neurotoxicology, 5, 295-322.
Shirai, T., M. Ohshima, A. Masuda, S. Tomano and N. Ito (1984) Promotion of 2(ethylnitrosamino)ethanol induced renal carcinogenesis in the rat by nephrotoxic compounds. Positive responses with folic acid, J. Natl. Cancer Inst., 72, 477-480.
Sirover, M.A., and L.A. Loeb (1976) Infidelity of DNA synthe- sis in vitro: screening for potential rnutagens or carcino- gens, Science, 194, 1434-1436.
Skreb, Y., and V. Habazin-Novak (1977) Lead induced modifi- cations of the response to X-rays in human cells in culture, Stud. Biophys., 63, 97-103.
Snyder, R.D., G.F. Davis and P.J. Lachmann (1989) Inhibition by metals of X-ray and ultraviolet-induced DNA repair in human cells, Biol. Trace Elem. Res., 21, 389-398.
Tanner, D.C., and M.M. Lipsky (1984) Effect of lead acetate on N-(4 ' - f luoro-4-b iphenyl )ace tamide- induced renal carcinogenesis in the rat, Carcinogenesis, 5, 1109-1113.
Vrieling, H., M.L. van Rooijen, N.A. Groen, M.Z. Zdzienicka, J.W.I.M. Simons, P.H.M. Lohman and A.A. van Zeeland (1989) DNA strand specificity for UV-induced mutations in mammalian cells, Mol. Cell. Biol., 9, 1277-1283.
Willis, A.E., and T. Lindahl (1987) DNA ligase deficiency in Bloom's syndrome, Nature (London), 325, 355-357.
Yew, F.-H., and R.T. Johnson (1979) Ultraviolet-induced DNA excision repair in human B and T lymphocytes, II. Effects of inhibitors and DNA precursors, Biochim. Biophys. Acta, 562, 240-251.
Zelikoff, J.T., J.H. Li, A. Hartwig, X.W. Wang, M. Costa and T.G. Rossman (1988) Genetic toxicology of lead com- pounds, Carcinogenesis, 9, 1727-1732.