impacts of nighttime warming on the soil nematode community in a winter wheat field of yangtze delta...
TRANSCRIPT

Journal of Integrative Agriculture2014, 13(7): 1477-1485 July 2014RESEARCH ARTICLE
© 2014, CAAS. All rights reserved. Published by Elsevier Ltd.doi: 10.1016/S2095-3119(14)60807-8
Impacts of Nighttime Warming on the Soil Nematode Community in a Winter Wheat Field of Yangtze Delta Plain, China
SONG Zhen-wei1, ZHANG Bin2, TIAN Yun-lu3, DENG Ai-xing1, ZHENG Cheng-yan1, Md Nurul Islam4, Md Abdul Mannaf4 and ZHANG Wei-jian1
1 Institute of Crop Sciences, Chinese Academy of Agricultural Sciences/Key Laboratory of Crop Physiology, Ecology & Production, Ministry of Agriculture, Beijing 100081, P.R.China
2 Institute of Rice Sciences, Guangdong Academy of Agricultural Sciences, Guangzhou 510640, P.R.China3 College of Agriculture, Nanjing Agricultural University, Nanjing 210095, P.R.China4 Bangladesh Agricultural Research Institute, Joydebpur Gazipur-1701, Bangladesh
Abstract
Changes in the soil nematode community induced by global warming may have a considerable influence on agro-ecosystem functioning. However, the impacts of predicted warming on nematode community in farmland (e.g., winter wheat field) have not been well documented. Therefore, a field experiment with free air temperature increase (FATI) was conducted to investigate the responses of the soil nematode community to nighttime warming in a winter wheat field of Yangtze Delta Plain, China, during 2007 to 2009. Nighttime warming (NW) by 1.8°C at 5-cm soil depth had no significant impact on the total nematode abundance compared to un-warmed control (CK). However, NW significantly affected the nematode community structure. Warming favored the bacterivores and fungivores, such as Acrobeles, Monhystera, Rhabditis, and Rhabdontolaimus in bacterivores, and Filenchus in fungivores, while the plant-parasites were hindered, such as Helicotylenchus and Psilenchus. Interestingly, the carnivores/omnivores remained almost unchanged. Hence, the abundances of bacterivores and fungivores were significantly higher under NW than those under CK. Similarly, the abundances of plant-parasites were significantly lower under NW than under CK. Furthermore, Wasilewska index of the nematode community was significantly higher under NW than those under CK, indicating beneficial effect to the plant in the soil. Our results suggest that nighttime warming may improve soil fertility and decrease soil-borne diseases in winter wheat field through affecting the soil nematode community. It is also indicated that nighttime warming may promote the sustainability of the nematode community by altering genera-specific habitat suitability for soil biota.
Key words: climate warming, FATI, soil nematodes, community structure,winter wheat
INTRODUCTION
Global warming has taken place with an increase of mean surface air temperature by 0.74°C for the past 100 years, and the temperature will further increase at
the rate of 1.1-6.4°C by the end of this century (IPCC 2007). Furthermore, the increased level of the daily minimum temperature was higher than that of the daily maximum temperature, and the increasing trend of the former is expected higher than the latter (Easterling et al. 1997; IPCC 2001; Lobell et al. 2011). Nematodes are the key agents of soil processes, such as organic matter
Received 6 September, 2013 Accepted 29 November, 2013SONG Zhen-wei, Tel: +86-10-62128815, E-mail: [email protected]; Correspondence ZHANG Wei-jian, Tel: +86-10-62156856, Fax: +86-62128815, E-mail: [email protected]

1478 SONG Zhen-wei et al.
© 2014, CAAS. All rights reserved. Published by Elsevier Ltd.
decomposition, mineralization, nutrient cycling and soil-borne diseases (Meagher 1977; Bongers and Ferris 1999; Liang et al. 2009). Warming-led alterations of the nematode abundance and its community structure may have a considerable influence on soil ecosystem functioning (Bakonyi et al. 2007).
Temperature change has been evidenced as an important factor determining nematode abundance under many conditions, such as Antarctic (Simmons et al. 2009), subarctic (Ruess et al. 1999), Alpine summit (Hoschitz and Kaufmann 2004), semiarid (Bakonyi et al. 2007; Li et al. 2013), and arid regions (Pen-Mouratov et al. 2004). Many studies have shown that higher temperature might increase nema-tode abundances, however, some studies reported that warming impacts could be different and even negative due to the particular conditions of latitude, soil prop-erties and microhabitat types (Bakonyi et al. 2007). Meanwhile, warming may also affect soil nematode community structure. For example, an increase of soil temperature by 1-2°C could significantly decrease soil nematode species diversity, resulting in great changes in nematode trophic structure and species dominance in subarctic soils (Ruess et al. 1999). Bakonyi et al. (2007) observed that nematode community diversity and multivariate structure were more sensitive to chang-es in soil temperature than soil moisture in a temperate semiarid shrub land. However, Li et al. (2013) found that warming influenced the nematode community di-versity less than N addition in a temperate steppe. As we know, most of existing observations about warming impacts on soil nematodes were conducted in natural soils. Moreover, previous studies related to warming impacts were mostly performed at a plant or plant community scale under controlled conditions, rather than at an ecosystem scale in situ (Okada and Ferris 2001; Aronson and McNulty 2009). The impacts of warming on soil nematode abundance and community structure in agro-ecosystem have not been well docu-mented so far.
Winter wheat (Triticum aestivum L.) is one of the most important crops in China, and more than 70% of Chinese winter wheat is sown in the eastern areas (Tian et al. 2012). Yangtze Delta Plain is one of the major regions of Chinese winter wheat cropping. Meanwhile, air tem-perature, especially daily minimum air temperature of
winter wheat growing season has significantly increased over the past decades, and will further increase till 2050 in this area (Chavas et al. 2009; Dong et al. 2011). Thus, to learn about warming impacts on the winter wheat cropping system will greatly facilitate the development of strategies leading to future crop production in China. Many efforts have been made to evaluate the effects of warming on wheat growth and production in this region (Tian et al. 2011, 2012; Zhang et al. 2013), whereas few studies focused on the responses of soil nematode. We, therefore, conducted a field experiment with a facility of free air temperature increase (FATI) in Nanjing, Jiangsu Province, during 2007 to 2009. Our objectives were to investigate the impacts of daily minimum air temperature increase (i.e., nighttime warming) on soil nematode abundance and community structure in a winter wheat field.
RESULTS
Total nematode abundance
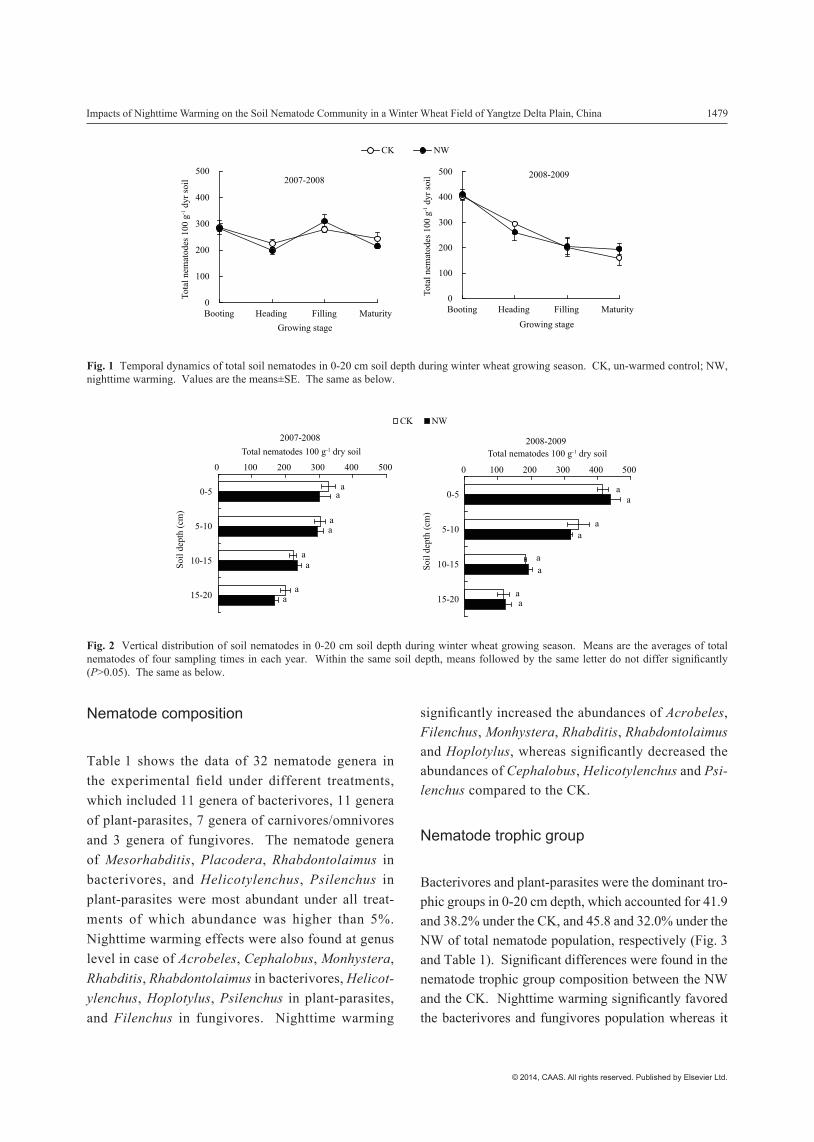
Soil nematode abundance in the 0-20 cm depth showed a decreasing trend with time from wheat booting stage to the maturity stage for the warmed (NW) and un-warmed plots (CK) (Fig. 1). Generally, the nematode abundance reached its highest values at the booting stage except that the highest value under NW occurred at the heading stage in 2008. The average nematode abundance over the sampling time under CK and NW was 257.7 and 251.0 individuals (ind.) per 100 g dry soil in 2008, and 264.6 and 268.3 ind. per 100 g dry soil in 2009, respectively. No significant difference was found in the nematode abundance between the NW and CK plots during the experimental duration.
There were similar vertical distribution patterns of nematode abundance between the treatments, with a decreasing trend along with the increase in the soil depth (Fig. 2). Average nematode abundance in 0-5 cm depth in 2008 and 2009 was 371.8 ind. per 100 g dry soil under the CK and 370.2 ind. per 100 g dry soil under the NW, while the corresponding values in 15-20 cm depth were 158.3 and 145.0 ind. per 100 g dry soil. However, no significant differences occurred in nematode abundance in each soil depth between the treatments.
Impacts of Nighttime Warming on the Soil Nematode Community in a Winter Wheat Field of Yangtze Delta Plain, China 1479
© 2014, CAAS. All rights reserved. Published by Elsevier Ltd.
Nematode composition
Table 1 shows the data of 32 nematode genera in the experimental field under different treatments, which included 11 genera of bacterivores, 11 genera of plant-parasites, 7 genera of carnivores/omnivores and 3 genera of fungivores. The nematode genera of Mesorhabditis, Placodera, Rhabdontolaimus in bacterivores, and Helicotylenchus, Psilenchus in plant-parasites were most abundant under all treat-ments of which abundance was higher than 5%. Nighttime warming effects were also found at genus level in case of Acrobeles, Cephalobus, Monhystera, Rhabditis, Rhabdontolaimus in bacterivores, Helicot-ylenchus, Hoplotylus, Psilenchus in plant-parasites, and Filenchus in fungivores. Nighttime warming
significantly increased the abundances of Acrobeles, Filenchus, Monhystera, Rhabditis, Rhabdontolaimus and Hoplotylus, whereas significantly decreased the abundances of Cephalobus, Helicotylenchus and Psi-lenchus compared to the CK.
Nematode trophic group
Bacterivores and plant-parasites were the dominant tro-phic groups in 0-20 cm depth, which accounted for 41.9 and 38.2% under the CK, and 45.8 and 32.0% under the NW of total nematode population, respectively (Fig. 3 and Table 1). Significant differences were found in the nematode trophic group composition between the NW and the CK. Nighttime warming significantly favored the bacterivores and fungivores population whereas it
CK NW
0
100
200
300
400
500
Booting Heading Filling Maturity
Tota
l nem
atod
es 1
00 g
-1 d
yr so
il
Growing stage
2007-2008
0
100
200
300
400
500
Booting Heading Filling Maturity
Tota
l nem
atod
es 1
00 g
-1 d
yr so
il
Growing stage
2008-2009
Fig. 1 Temporal dynamics of total soil nematodes in 0-20 cm soil depth during winter wheat growing season. CK, un-warmed control; NW, nighttime warming. Values are the means±SE. The same as below.
a
a
a
a
a
a
a
a
0 100 200 300 400 500
0-5
5-10
10-15
15-20
Total nematodes 100 g-1 dry soil
Soil
dept
h (c
m)
2007-2008
CK NW
a
a
a
a
a
a
a
a
0 100 200 300 400 500
0-5
5-10
10-15
15-20
Total nematodes 100 g-1 dry soil
Soil
dept
h (c
m)
2008-2009
Fig. 2 Vertical distribution of soil nematodes in 0-20 cm soil depth during winter wheat growing season. Means are the averages of total nematodes of four sampling times in each year. Within the same soil depth, means followed by the same letter do not differ significantly (P>0.05). The same as below.

1480 SONG Zhen-wei et al.
© 2014, CAAS. All rights reserved. Published by Elsevier Ltd.
hindered the plant-parasites population as compared with the CK. However, there was no significant difference in the carnivores/omnivores population between the treatments.
Nematode ecological indices
Ecological indices of soil nematode community are shown in Table 2. Nighttime warming significantly increased the Wasilewska index (WI) of nematode com-munity by 35.5% compared to the CK. Though enrich-ment index (EI), Shannon-Wiener index (H´), Pielou’s
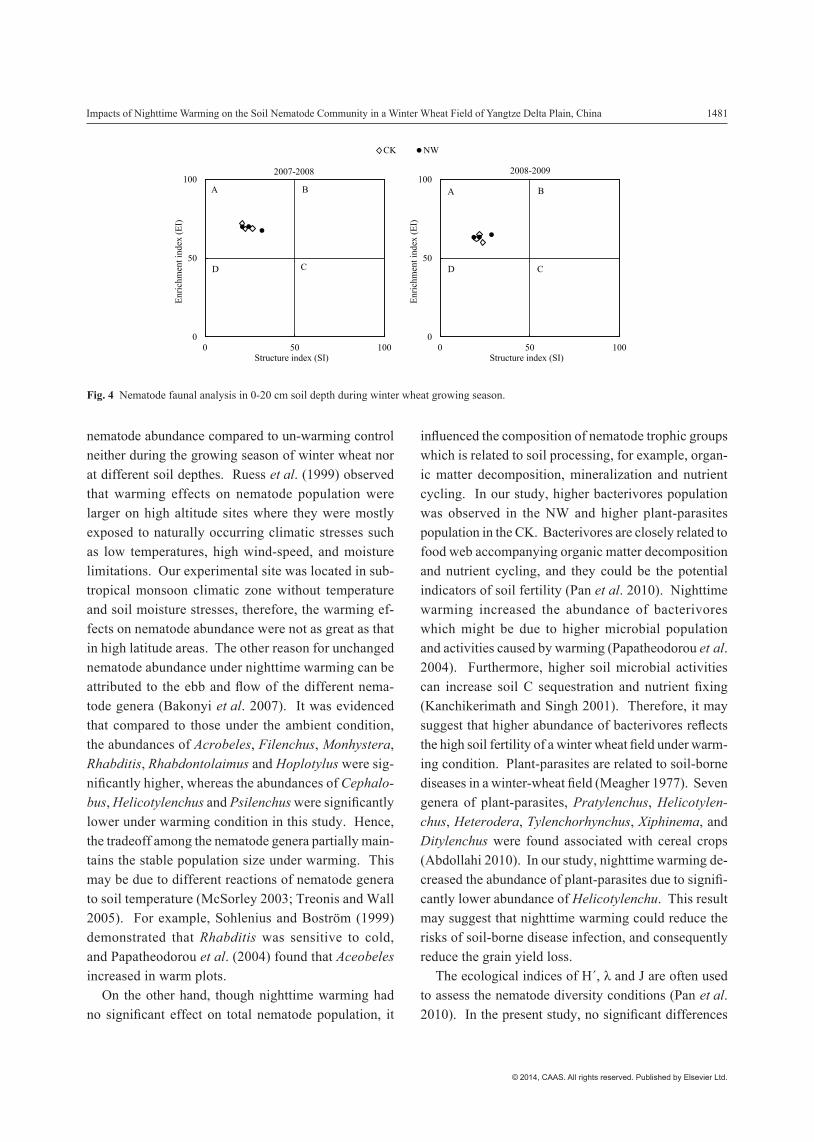
evenness index (J), and structural index (SI) were higher in the NW than that in the CK, no significant difference was found between the treatments. Furthermore, night-time warming decreased the genus dominance index (λ) but not significantly compared to the CK. Nematode faunal analysis indicated that both the CK and NW plots belonged to the A quadrants (Fig. 4), which indicated the soil food webs were disturbed.
DISCUSSION
In the present study, the increase of soil temperature at 5 cm depth by 1.8°C didn’t significantly affect the
Table 1 Genus abundances of soil nematodes under different treatments
Trophic groups Genus Guild1)Genus abundance (%)2)
CK NWBacterivores 41.9±1.2 b 45.8±1.51 a
Acrobeles Ba2 0.9±0.27 b 1.8±0.11 aAcrobeloides Ba2 1.6±0.33 a 1.0±0.25 aCephalobus Ba2 2.9±0.26 a 2.0±0.22 b
Mesorhabditis Ba1 5.0±0.78 a 5.2±0.64 aMonhystera Ba2 3.1±0.30 b 4.1±0.21 a
Panagrolaimus Ba1 2.6±0.33 a 2.7±0.33 aPelodera Ba1 0.9±0.34 a 1.1±0.37 aPlacodira Ba2 13.0±1.06 a 13.0±1.01 aRhabditis Ba1 3.4±0.22 b 4.2±0.33 a
Rhabdontolaimus Ba3 8.2±0.44 b 10.4±0.52 aWilsonena Ba2 0.3±0.15 a 0.2±0.04 a
Plant-parasites 38.2±1.35 a 32.0±2.11 bHelicotylenchus H3 20.5±0.55 a 14.8±1.07 b
Heterodera H3 0.8±0.05 a 0.7±0.05 aHoplotylus H3 0.8±0.17 b 1.3±0.18 aLongidorus H5 2.0±0.34 a 2.1±0.19 aMalenchus H2 1.0±0.15 a 1.2±0.14 a
Paratylenchus H2 2.5±0.13 a 2.5±0.13 aPratylenchus H3 0.4±0.13 a 0.6±0.15 aPsilenchus H2 8.2±0.23 a 6.0±0.31 b
Trichodorus H4 0.3±0.11 a 0.3±0.04 aTylenchorhynchus H3 0.9±0.19 a 1.2±0.13 a
Tylenchus H2 1.0±0.24 a 1.4±0.11 aCarnivores/Omnivores 15.4±0.78 a 15.7±0.33 a
Aporcelaimus Om5 0.8±0.13 a 0.7±0.08 aDorylaimus Om4 2.8±0.21 a 2.0±0.15 b
Eduorylaimus Om4 0.6±0.09 b 0.9±0.35 aLabronema Om4 1.2±0.19 a 1.1±0.13 a
Mesodorylaimus Om5 4.0±0.11 a 4.0±0.21 aMononchus Ca4 2.2±0.07 a 2.2±0.07 aPungentus Om4 3.7±0.27 b 4.8±0.70 a
Fungivores 4.5±0.33 b 6.5±0.28 aAphelenchoides Fu2 1.5±0.17 a 1.6±0.12 a
Aphelenchus Fu2 0.8±0.16 a 1.2±0.20 aFilenchus Fu2 2.2±0.38 b 3.7±0.43 a
1) Ba, bacterivores; H, plant-parasites; Ca/Om, carnivores/omnivores; Fu, fungivores. Values are trophic group plus colonizer-persister (c-p) value.
2) CK, un-warmed control; NW, nighttime warming. Values are the means±SE. Means followed by the same letter do not differ significantly (P>0.05). The same as below.
b a
a
b
a
b
a
a
0
40
80
120
160
Ba H Ca/Om Fu
Tota
l nem
atod
es 1
00 g
-1 d
ry so
il
Trophic groups
2007-2008
CK NW
ba
a
b
a
b
a
a
0
40
80
120
160
Ba H Ca/Om Fu
Tota
l nem
atod
es 1
00 g
-1 d
ry so
il
Trophic groups
2008-2009
Fig. 3 Trophic group composition of soil nematodes in 0-20 cm soil depth during winter wheat growing season. Ba, bacterivores; H, plant-parasites; Ca/Om, carnivores/omnivores; Fu, fungivores.
Table 2 Ecological indices of soil nematode community under different treatmentsEcological index CK NWShannon-Wiener index (H´) 1.16±0.02 a 1.19±0.02 aGenus dominance index (λ) 0.35±0.01 a 0.34±0.01 aPielou’s evenness index (J) 0.83±0.01 a 0.86±0.02 aEnrichment index (EI) 66.05±1.17 a 66.55±0.15 aStructural index (SI) 22.57±1.28 a 24.40±3.06 aWasilewska index (WI) 1.21±0.02 b 1.64±0.14 a
Impacts of Nighttime Warming on the Soil Nematode Community in a Winter Wheat Field of Yangtze Delta Plain, China 1481
© 2014, CAAS. All rights reserved. Published by Elsevier Ltd.
nematode abundance compared to un-warming control neither during the growing season of winter wheat nor at different soil depthes. Ruess et al. (1999) observed that warming effects on nematode population were larger on high altitude sites where they were mostly exposed to naturally occurring climatic stresses such as low temperatures, high wind-speed, and moisture limitations. Our experimental site was located in sub-tropical monsoon climatic zone without temperature and soil moisture stresses, therefore, the warming ef-fects on nematode abundance were not as great as that in high latitude areas. The other reason for unchanged nematode abundance under nighttime warming can be attributed to the ebb and flow of the different nema-tode genera (Bakonyi et al. 2007). It was evidenced that compared to those under the ambient condition, the abundances of Acrobeles, Filenchus, Monhystera, Rhabditis, Rhabdontolaimus and Hoplotylus were sig-nificantly higher, whereas the abundances of Cephalo-bus, Helicotylenchus and Psilenchus were significantly lower under warming condition in this study. Hence, the tradeoff among the nematode genera partially main-tains the stable population size under warming. This may be due to different reactions of nematode genera to soil temperature (McSorley 2003; Treonis and Wall 2005). For example, Sohlenius and Boström (1999) demonstrated that Rhabditis was sensitive to cold, and Papatheodorou et al. (2004) found that Aceobeles increased in warm plots.
On the other hand, though nighttime warming had no significant effect on total nematode population, it
influenced the composition of nematode trophic groups which is related to soil processing, for example, organ-ic matter decomposition, mineralization and nutrient cycling. In our study, higher bacterivores population was observed in the NW and higher plant-parasites population in the CK. Bacterivores are closely related to food web accompanying organic matter decomposition and nutrient cycling, and they could be the potential indicators of soil fertility (Pan et al. 2010). Nighttime warming increased the abundance of bacterivores which might be due to higher microbial population and activities caused by warming (Papatheodorou et al. 2004). Furthermore, higher soil microbial activities can increase soil C sequestration and nutrient fixing (Kanchikerimath and Singh 2001). Therefore, it may suggest that higher abundance of bacterivores reflects the high soil fertility of a winter wheat field under warm-ing condition. Plant-parasites are related to soil-borne diseases in a winter-wheat field (Meagher 1977). Seven genera of plant-parasites, Pratylenchus, Helicotylen-chus, Heterodera, Tylenchorhynchus, Xiphinema, and Ditylenchus were found associated with cereal crops (Abdollahi 2010). In our study, nighttime warming de-creased the abundance of plant-parasites due to signifi-cantly lower abundance of Helicotylenchu. This result may suggest that nighttime warming could reduce the risks of soil-borne disease infection, and consequently reduce the grain yield loss.
The ecological indices of H´, λ and J are often used to assess the nematode diversity conditions (Pan et al. 2010). In the present study, no significant differences
0
50
100
0 50 100
Enric
hmen
t ind
ex (E
I)
Structure index (SI)
2007-2008
CK NW
A B
CD
0
50
100
0 50 100
Enric
hmen
t ind
ex (E
I)
Structure index (SI)
2008-2009
A B
CD
Fig. 4 Nematode faunal analysis in 0-20 cm soil depth during winter wheat growing season.

1482 SONG Zhen-wei et al.
© 2014, CAAS. All rights reserved. Published by Elsevier Ltd.
were observed between the treatments considering H´, λ and J, which implies that nighttime warming has no effects on the nematode diversity. This result was consis-tent with previous findings that temperature did not affect several parameters, such as total nematode population and species diversity, but insignificantly affected the nematode abundance of different genera (Bakonyi et al. 2007). Moreover, EI and SI are often used to assess the nematode community structure and indicate the dynam-ics of soil food webs (Ferris et al. 2001; Li et al. 2010). Our study showed that no significant differences in EI and SI were observed between the treatments, indicating that nighttime warming has no significant effects on soil food webs. It was also confirmed by the nematode faunal analysis that both the CK and NW belonged to the A quadrants with disturbed soil food webs (Ferries et al. 2001; Li et al. 2010). Furthermore, WI can demonstrate the proportion of free nematodes and plant-parasites and reflect the health of plants in the soil (Wasilewska 1994). In this study, WI was significantly higher under the NW compared to the CK due to higher abundance of bacterivores and fungivores, which suggests that nighttime warming may benefit to the plant health and soil environmental sustainability (Chen et al. 2009).
CONCLUSION
In this study, an increase of nighttime temperature at 5 cm soil depth by 1.8°C had no significant effects on nematode abundance neither during the growing season of winter wheat nor at different soil layers. Moreover, nighttime warming significantly increased the abundances of Acrobeles, Monhystera, Rhabditis, Rhabdontolaimus and Hoplotylus in bacterivores, and Filenchus in fungivores, and decreased the abundances of Psilenchus and Helicotylenchus in plant-parasites. No significant warming impacts were found in the abundances of carnivores/omnivores. Hence, nighttime warming favored the bacterivores and fungivores while it hindered the plant-parasites in winter wheat field soil. Our results suggest that nighttime warming may improve the soil fertility, maintain soil environmental sustainability, and reduce soil-borne disease infection risk for winter wheat growth through altering soil nematode community.
MATERIALS AND METHODS
Experimental site description
The experimental site was located at the Experimental Station of Jiangsu Academy of Agricultural Sciences, Nanjing, Jiangsu Province, China (32°02´N, 118°52´E, 11 m a.s.l.). This location belongs to the climatic zone of subtropical monsoon climate. The mean air temperature, the annual precipitation, the sunshine hours and the frost-free days are 16.7°C, 1 050 mm, 1 900 h and 237 d, respectively. The soil at the experimental site is brunisolic silt loam soil (Alfisols in USA-ST) with 0.5% sand, 75.3% silt and 24.2% clay, respectively. The nutrient conditions were as follows: soil contains 8.2 g kg-1 organic C, 2.6 g kg-1 total N, 0.6 g kg-1 total P, 14.0 g kg-1 total K, 166.2 mg kg-1 available P, and 165.0 mg kg-1 available K. The cropping pattern is a double cropping system of annually winter wheat-rice cropping.
Experimental design and management
The field experiment was started in winter 2007 and continued up to summer 2009, including two treatments: nighttime warming (NW) and un-warmed control (CK). The field was laid out in a randomized block design with three replicates. The plot size was 30 m2 (6 m×5 m). The field warming system was constructed according to the design of free air temperature increase (FATI) facility located at the Great Plain Apiaries, USA (Wan et al. 2002). In each warmed plot, a single 180 cm × 20 cm infrared heater (Jiangsu Tiande Special Light Source Co., Ltd., China) was suspended 1.5 m above the ground, which provided with continuous warming from 18:00 to 6:00. In the un-warmed plot, a ‘dummy’ heater of the same shape and size was suspended at the same height to simulate the shading effects of the heater. The distance between the adjacent plots was about 5 m to avoid heating contamination between plots. This FATI facility could provide 2 m×2 m sampling areas with uniform and reliable warming effects. The FAIT facility could significantly increase the mean soil temperature at 5 cm depth by 1.8°C. The same soil water content in 0-20 cm depth was maintained in both the treatments during the entire growth period of wheat (Tian et al. 2010).
The winter wheat cultivar Yangmai 11 was manually sown on 15 November, 2007, and 2 November, 2008 at a density of 225 plants m-2 with a row spacing of 20 cm and were harvested on 2 Jun,e 2008 and 24 May, 2009, respectively. In accordance with local agronomic practices, the fertilizer applications of N, P and K in each plot were 225, 75 and 75 kg ha-1, respectively. The total P and K and 50% N were applied 2 days prior to sowing as basal dressing. The other 10% N was applied as side dressing at early tillering stage at the beginning of March, and 40% at the spike initiation stage at the beginning of April. The

Impacts of Nighttime Warming on the Soil Nematode Community in a Winter Wheat Field of Yangtze Delta Plain, China 1483
© 2014, CAAS. All rights reserved. Published by Elsevier Ltd.
precipitations were 396.9 mm in 2007-2008 growing season and 354.1 mm in 2008-2009 growing season, thus, no irrigation was applied during the entire growth period of winter wheat.
Soil sampling
For temporal nematode population measurements, soil samples were collected at four key growth stages of winter wheat viz., booting stage (28 March, 2008 and 16 March, 2009 ), heading stage (27 April, 2008 and 9 April, 2009), grain filling stage (13 May, 2008 and 4 May, 2009) and maturity stage (30 May, 2008 and 24 May, 2009). Each soil sample was collected from six randomly selected sampling points at 5 cm intervals from 0 to 20 cm depth. Six sub-samples for each soil depth class were blended to get one composite sample for each depth class per plot. Each soil sample was passed through a 6-mm-mesh soil sieve to remove the plant leaves and roots, and large stones. All samples were stored in a refrigerator at 4°C until subsequent analysis.
Nematode extraction and identification
Sucrose solution-elutriation-centrifugation method was used to extract soil nematodes from about 100 g fresh soil (Bulluck et al. 2002). Then the nematodes were heat-killed by immersion in 65°C water for 2 min and preserved in triethanolamine formaldehyde (TAF) solution (Shepherd, 1970). Total nematodes in each sample were counted under an anatomical lens (40×). For each sample, 100 nematodes were randomly selected to identify the genera, trophic groups, and colonizer-persister (c-p) value with the aid of an optical microscope (Yeates et al. 1993; Bongers 1994; Yin 1998; Okada and Harada 2007; Li et al. 2012). In the present study, the nematodes were classified as four trophic groups characterized by feeding habits, bacterivores (Ba), plant-parasites (H), carnivores/omnivores (Ca/Om), and fungivores (Fu). Soil moisture was determined by oven-drying samples at 105°C. The density of total and different trophic-group nematodes were adjusted to the number per 100 g dry soil. The nematode abundance for each genus was the percentage of specific genus in total nematodes.
Nematode community structure was characterized by Shannon-Wiener index (H´) (Shannon and Weaver 1949), Pielou’s evenness index (J) (Kennedy and Smith 1995), genus dominance index (λ) (Bongers 1990), enrichment index (EI), structure index (SI) (Ferris et al. 1997), and Wasilewska index (WI) (Yeates 2003).
Statistical analysis
The analysis of variances for soil nematode population
density, abundance and community structure were performed independently using the procedure of SPSS 13.0 (SPSS Inc, Chicago, IL, USA). Least significant differences (LSD) at P<0.05 were used to detect significant differences among means (Steel and Torrie 1980).
AcknowledgementsThis research was supported by the National Basic Research Program of China (2010CB951501), the Key Technologies R&D Program of China during the 12th Five-Year Plan period (2011BAD16B14), the National Natural Science Foundation of China (30771278) and the Innovation Program of Chinese Academy of Agricultural Sciences, China.
ReferencesAbdollahi M. 2010. Plant-parasitic nematodes associated with
wheat in Kohgiluyeh and Boyer-Ahmad Province, Iran. Journal of Agricultural Research, 48, 131-136.
Aronson E L, McNulty S G. 2009. Appropriate experimental ecosystem warming methods by ecosystem, objective, and practicality. Agricultural and Forest Meteorology, 149, 1791-1799.
Bakonyi G, Nagy P, Kovacs-Lang E, Kovacs E, Barabas S, Repasia V, Seres A. 2007. Soil nematode community structure as affected by temperature and moisture in a temperate semiarid shrubland. Applied Soil Ecology, 37, 31-41.
Bongers T. 1990. The maturity index: An ecological measure of environmental disturbance based on nematode species composition. Oecologia, 83, 14-19.
Bongers T. 1994. De Nematoden van Nederland Pirola. Schoorl, Netherlands. (in Dutch)
Bongers T, Ferris H. 1999. Nematode community structure as a bioindicator in environmental monitoring. Trends in Ecology and Evolution, 14, 224-228.
Bulluck L R, Barker K R, Ristaino J B. 2002. Influences of organic and synthetic soil fertility amendments on nematode trophic groups and community dynamics under tomatoes. Applied Soil Ecology, 21, 233-250.
Chavas D R, Izaurralde R C, Thomson A M, Gao X. 2009. Long-term climate change impacts on agricultural productivity in eastern China. Agriculture and Forest Meteorology, 149, 1118-1128.
Chen G, Qin J, Shi D, Zhang Y, Ji W. 2009. Diversity of soil nematodes in areas polluted with heavy metals and polycyclic aromatic hydrocarbons (PAHs) in Lanzhou, China. Environmental Management, 44,163-172.
Dong W, Chen J, Zhang B, Tian Y, Zhang W. 2011. Responses of biomass growth and grain yield of midseason rice to the anticipated warming with FATI facility in East China. Field Crop Research, 123, 259-265.
Easterling D R, Peterson T C, Karl T R. 1997. Maximum and minimum temperature trends for the globe. Science, 277, 364-367.

1484 SONG Zhen-wei et al.
© 2014, CAAS. All rights reserved. Published by Elsevier Ltd.
Ferris H, Bongers T, de Goede R G M. 2001. A framework for soil food web diagnostics: Extension of the nematode faunal analysis concept. Applied Soil Ecology, 18, 13-29.
Ferris H, Venette R C, Lau S S. 1997. Population energetics of bacterial-feeding nematodes: Carbon and nitrogen budgets. Soil Biology & Biochemistry, 29, 1183-1194.
Hoschitz M, Kaufmann R. 2004. Soil nematode communities of Alpine summits-site differentiation and microclimatic influences. Pedobiologia, 48, 313-320.
IPCC. 2001. Climate change 2001: The scientific basis. In: Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, United Kingdom.
IPCC. 2007. Climate change 2007: The physical science basis. In: Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, United Kingdom.
Kanchikerimath M, Singh D. 2001. Soil organic matter and biological properties after 26 years of maize-wheat-cowpea cropping as affected by manure and fertilization in a Cambisol in semiarid region of India. Agriculture, Ecosystems and Environment, 86, 155-162.
Kennedy A C, Smith K L. 1995. Soil microbial diversity and the sustainability of agricultural soils. Plant and Soil, 170, 75-86.
Li Q, Bai H, Liang W, Xia J, Wan S, van der Putten W H. 2013. Nitrogen addition and warming independently influence the belowground micro-food web in a temperate steppe. PLOS ONE, 8, e60441.
Li Q, Bao X, Lu C, Zhang X, Zhu J, Jiang Y, Liang W. 2012. Soil microbial food web responses to free-air ozone enrichment can depend on the ozone-tolerance of wheat cultivars. Soil Biology & Biochemistry, 47, 27-35.
Li Q, Jiang Y, Liang W, Lou Y, Zhang E, Liang C. 2010. Long-term effect of fertility management on the soil nematode community in vegetable production under greenhouse conditions. Applied Soil Ecology, 46, 111-118.
Liang W, Lou Y, Li Q, Zhong S, Zhang X, Wang J. 2009. Nematode faunal response to long-term application of nitrogen fertilizer and organic manure in Northeast China. Soil Biology & Biochemistry, 41, 883-890.
Lobell D B, Schlenker W, Costa-Roberts J. 2011. Climate trends and global crop production since 1980. Science, 333, 616-620.
McSorley R. 2003. Adaptations of nematodes to environmental extremes. Florida Entomologist, 86, 138-142.
Meagher J W. 1977. World dissemination of the cereal-cyst nematode (Heterodera avenae) and its potential as a pathogen of wheat. Journal of Nematology, 9, 9-15.
Okada H, Ferris H. 2001. Effect of temperature on growth and nitrogen mineralization of fungi and fungal-feeding nematodes. Plant and Soil, 234, 253-262.
Okada H, Harada H. 2007. Effects of tillage and fertilizer on nematode communities in a Japanese soybean field. Applied Soil Ecology, 35, 582-598.
Pan F, McLaughlin N B, Yu Q, Xue A G, Xu Y, Han X, Li C, Zhao D. 2010. Responses of soil nematode community structure to different long-term fertilizer strategies in the soybean phase of a soybean-wheat-corn rotation. European Journal of Soil Biology, 46, 105-111.
Papatheodorou E M, Argyropoulou M D, Stamou G P. 2004. The effects of large- and small-scale differences in soil temperature and moisture on bacterial functional diversity and the community of bacterivorous nematodes. Applied Soil Ecology, 25, 37-49.
Pen-Mouratov S, He X, Steinberger Y. 2004. Spatial distribution and trophic diversity of nematode populations under Acacia raddiana along a temperature gradient in the Negev Desert ecosystem. Journal of Arid Environment, 56, 339-355.
Ruess L, Michelsen A, Schmidt I K, Jonasson S. 1999. Simulated climate change affecting microorganisms, nematode density and biodiversity in subarctic soils. Plant and Soil, 212, 63-73.
Shannon C E, Weaver W. 1949. The Mathematical Theory of Communication. University of Illinois Press, Urbana, IL, USA.
Shepherd A M. 1970. Preparation of nematodes for electron microscopy. In: Southey J F, ed., Laboratory Methods for Work with Plant and Soil Nematodes. HMSO, London, UK. pp. 88-95.
Simmons B L, Wall D H, Adams B J, Ayres E, Barrett J E, Virginia R A. 2009. Long-term experimental warming reduces soil nematode populations in the McMurdo Dry Valleys, Antarctica. Soil Biology & Biochemistry, 41, 2052-2060.
Sohlenius B, Boström S. 1999. Effects of climate change on soil factors and metazoan microfauna (nematodes, tardigrades and rotifers) in a Swedish tundra soil - a soil transplantation experiment. Applied Soil Ecology, 12, 113-128.
Steel R G D, Torrie J H. 1980. Principles and Procedures of Statistics, A Biometrical Approach. 2nd ed. McGraw-Hill, New York, USA.
Tian Y, Chen J, Chen C, Deng A, Song Z, Zheng C, Hoogmoed W, Zhang W. 2012. Warming impacts on winter wheat phenophase and grain yield under field conditions in Yangtze Delta Plain, China. Field Crop Research, 134, 193-199.
Tian Y, Chen J, Deng A, Zheng J, Zhang W. 2011. Effects of asymmetric warming on the growth characteristics and yield components of winter wheat under free air temperature increased. Chinese Journal of Applied Ecology, 22, 681-686. (in Chinese)
Tian Y, Zheng J, Zhang B, Chen J, Dong W, Yang F, Zhang W. 2010. Design of free air temperature increasing (FATI) system for upland with three diurnal warming scenarios and their effects. Scientia Agricultura Sinica, 43, 3724-3731. (in Chinese)
Treonis A M, Wall D H. 2005. Soil nematodes and desiccation survival in the extreme arid environment of the Antarctic

Impacts of Nighttime Warming on the Soil Nematode Community in a Winter Wheat Field of Yangtze Delta Plain, China 1485
© 2014, CAAS. All rights reserved. Published by Elsevier Ltd.
dry valleys. Integrative and Comparative Biology, 45, 741-750.
Wan S, Luo Y, Wallace L L. 2002. Change in microclimate induced by experimental warming and clipping in tallgrass prairie. Global Change Biology, 8, 754-768.
Wasilewska L. 1994. The effect of age of meadows on succession and diversity in soil nematode communities. Pedobiologia, 38, 1-11.
Yeates G W, Bongers T, de Goede R G M, Freckman D W, Georoieva S S. 1993. Feeding habits in soil nematode families and genera - an outlines for soil ecologists. Journal
of Nematology, 25, 315-331.Yeates G W. 2003. Nematodes as soil indicators: Functional
and biodiversity aspects. Biology and Fertility of Soils, 37, 199-210.
Yin W Y. 1998. Pictorial Keys to Soil Animals of China. Science Press, Beijing, China. (in Chinese)
Zhang M, Chen J, Guo J, Tian Y, Yang S, Zhang L, Yang B, Zhang W. 2013. Effects of nighttime warming on winter wheat root growth and soil nutrient availability. Chinese Journal of Applied Ecology, 24, 445-450. (in Chinese)
(Managing editor SUN Lu-juan)