immunochemical studies on the insulin-degrading...
TRANSCRIPT

Immunochemical Studies on theInsulin-degrading Enzyme fromPig and Rat Skeletal MuscleKOICHI YOKONO, YOSHIMICHI IMAMURA, KOZUI SHII, NOBUHIKO MIZUNO, HIDEYO SAKAI, AND SHIGEAKI BABA
SUMMARYInsulin-degrading enzyme (IDE), which proteolyticallydegraded insulin with a high degree of specificity, waspurified from pig skeletal muscle by ammonium sul-fate precipitation, chromatography on Bio-Gel P-200and DEAE-cellulose, and finally rechromatography onSephadex G-200 (rechromatography fraction). The en-zyme was also purified by affinity chromatography (af-finity fraction). Both fractions migrated as a singlecomponent at the same position on polyacrylamide-gel disc electrophoresis. Antiserum against pig mus-cle IDE was obtained by immunization of rabbits usingthe rechromatography fraction. By means of anti-serum, it was shown that pig muscle IDE (affinity frac-tion), rat muscle cytosol-, and membrane-IDE gave aprecipitin band of identity in Ouchterlony double-im-munodiffusion systems. Quantitative immunoprecipitindata demonstrated that the antiserum inhibited theactivities of the above three IDEs compared with nor-mal rabbit serum. These data suggest that the insulin-degrading enzyme from porcine muscle and that fromrat muscle have similar immunologic properties.
The antiserum described here should be a usefultool for the examination of subcellular distribution andthe quantitative analysis of insulin-degrading enzyme.It may also be helpful in determining the physiologicSignificance Of IDE. DIABETES 29:856-859, October 1980.
Many reports are currently available concerningthe insulin-degrading enzyme that proteolytic-ally degrades insulin with a high degree ofspecificity. The insulinase reported by Mirsky
was heat-labile,1 restricting the procedures available for en-zymatic purification. However, insulin protease was puri-fied from rat skeletal muscle using a procedure that in-
From The Second Department of Internal Medicine, Kobe University Schoolof Medicine, Kobe, Japan.Address reprint requests to Prof. Shigeaki Baba, The Second Department ofInternal Medicine, Kobe University School of Medicine, Ikuta-ku, Kobe,Japan.Received for publication 31 July 1980.
eluded affinity chromatography.2'3 We reported thatinsulin-degrading enzyme (IDE) could be highly purifiedfrom pig skeletal muscle by the isoelectrofocusing tech-nique. Porcine IDE prepared in this way appeared to beidentical to rat insulin protease in most of its biochemicalproperties.4 These enzymes have been extensively studiedin terms of enzymatic characteristics1-4-5 and biologic signif-icance.6"8 On the other hand, a number of questions remainunanswered, such as whether these enzymes exist in bloodand how they might interact with insulin receptors in the pro-cess of insulin degradation. Immunochemical studies usingantiserum against these enzymes should be a useful tool forthe examination of these problems. However, there is no re-port concerning the immunochemical study of these en-zymes since it has been difficult to obtain enough pure en-zyme protein to form an antiserum.
In the present study, IDE purified from pig skeletal muscleby Sephadex G-200 rechromatography was utilized to prod-uce IDE antibodies in rabbits. The immunologic purity of theantiserum was demonstrated in the Ouchterlony double-im-munodiffusion system, and quantitative antigen-antibody ti-trations of pig and rat muscle IDEs with the antiserum werecarried out.
MATERIALS AND METHODSMaterials. 125l-labeled porcine insulin (specific activity:150-200 fiCUfig) was purchased from Dinabott Rl Labora-tories, Tokyo. Pork monocomponent insulin was obtainedfrom Novo Research Institute, Copenhagen. Bovine serumalbumin (BSA) was from Sigma Chemical Co., St. Louis,Missouri. Sephadex G-25, G-200, and CNBr-activated Se-pharose 4B were manufactured by Pharmacia Fine Chemi-cals, Inc., Piscataway, New Jersey. Bio-Gel P-200 wasbought from Bio-Rad Laboratories, Richmond, California,and DEAE-cellulose from Brown Co., Berlin, New Hamp-shire. Freund's complete adjuvant was obtained from DifcoLaboratories, Detroit, Michigan. Other chemicals were of re-agent grade.Enzyme purification. Freshly prepared pig leg muscleswere trimmed of fat and connective tissue, minced, and ho-
856 DIABETES, VOL 29, OCTOBER 1980

KOICHI YOKONO AND ASSOCIATES
mogenized in 0.35 M sucrose in a Waring blender for five15-sec intervals. The homogenate was centrifuged at10,000 x g for 20 min. The supernatant was fractionated byammonium sulfate precipitation, Bio-Gel P-200, and DEAE-cellulose column chromatography as previously de-scribed.4 The enzyme protein of the DEAE-cellulose fractionwas lyophylized and then further purified by Sephadex G-200 column rechromatography or affinity chromatography.For the former, the lyophylizate, dissolved in 25 mM potas-sium phosphate buffer (KPB), pH 7.5, containing 1 mM di-thiothreitol (DTT), was chromatographed on a Sephadex G-200 column (3.2 x 93 cm) equilibrated with the same buffercontaining 1 mM DTT. Fractions comprising the peak en-zyme activity were pooled, salted out with 80% saturation ofammonium sulfate solution, desalted by passage through aSephadex G-25 column, and concentrated with a collodionbag. This preparation was rechromatographed on a Sepha-dex G-200 column, following the exact procedure of the firstchromatography. The final concentrate was lyophilized with1% sucrose and stored at -80°C until used (rechroma-tography fraction). A second batch of the DEAE-cellulosefraction was purified by affinity chromatography accordingto Duckworth et al.2 Cyanogen bromide-activated se-pharose 4B was reacted with insulin in 0.2 M sodium citratebuffer at pH 5. The enzyme was adsorbed to the insulin-Se-pharose column (1.8 x 18 cm), washed with 20 mM acetatebuffer (pH 6.2) containing 1 mM DTT, and eluted with 0.2 MNaCI in the same buffer. The purified enzyme was dialyzedagainst 50 mM KPB (pH 7.5), lyophylized with 1% sucrose,and stored at -80°C until used (affinity fraction).
Insulin-degrading enzymes from the cytosol fraction (cy-tosol-IDE) and the plasma membrane fraction (membrane-IDE) of rat skeletal muscle were prepared as previously de-scribed.8
Immunologic studies. Five New Zealand white female rab-bits (2-2.5 kg body weight) were bled before the immuniza-tion to provide control serum. Each rabbit was injected sub-cutaneously with 0.6 ml of an emulsion prepared from equalvolumes of the rechromatographed fraction of pig muscleIDE (1 mg protein in 0.1 M KPB, pH 7.0) and Freund's com-plete adjuvant. The injections were repeated at 3-wk inter-vals and bleedings were carried out 8 days after each injec-tion. All serum was stored at -20°C until used. Ouchterlonydouble-immunodiffusion studies were performed accordingto the method of Stollar and Levine.9 Diffusion was allowedto proceed at 4°C in a moist chamber for about 48 h. Quanti-tative antigen-antibody titrations were carried out as de-scribed by Varandani.10 Solutions of IDE from pig muscle(affinity fraction) or rat muscle were incubated with anti-serum, normal rabbit serum, or 0.1% BSA in 0.1 : M KPB(pH 7.0) for 1 h at 25°C. After centrifugation aliquots ofsupernatant were assayed for insulin-degrading activity.Analytic procedures. Insulin-degrading activity was esti-mated by determining the quantity of trichloroacetic acid(TCA)-soluble radioactivity produced after incubation with125l-insulin.8 Protein concentrations were measured by theLowry method,11 with BSA used as standard.
RESULTS AND DISCUSSIONAs previously reported,2i4>5 insulin-degrading activity of theammonium sulfate fraction was eluted at the front shoulderof the protein peak on Bio-Gel P-200 filtration; maximum en-
0 10 20 30 40 50 60FRACTION NUMBER
FIGURE 1. Elutlon pattern of the DEAE-cellulose fraction on aSephadex G-200 column (3.2 x 93 cm) eluted with 25 mM phosphatebuffer (pH 7.5) containing 1 mM DTT. The upper panel shows the firstchromatography and the lower panel shows the rechromatography ona Sephadex G-200 column. Shaded areas Indicate the fractions usedfor additional studies.
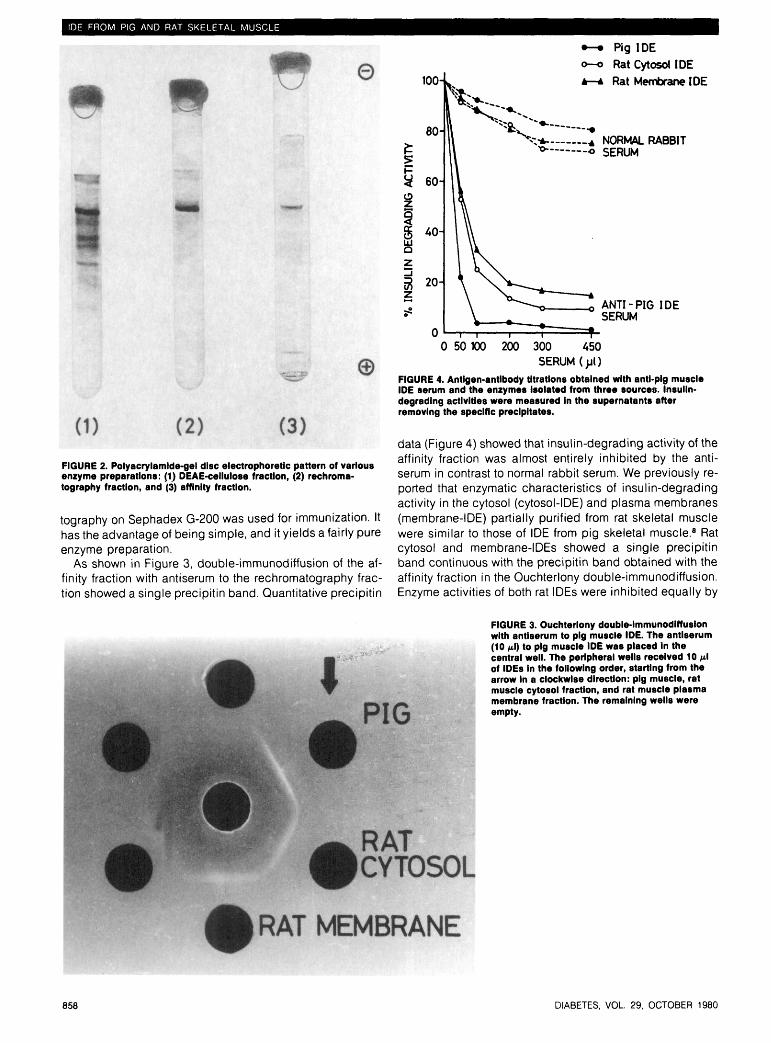
zyme activity appeared in the 0.25 M NaCI fraction onDEAE-cellulose chromatography. Figure 1 shows the elutionpattern of DEAE-cellulose purified enzyme after passageover the Sephadex G-200 column. The upper panel showsthe first chromatography on the Sephadex G-200 column.The highest enzyme activity appeared in the front proteinpeak. The fraction indicated by the shaded area was rechro-matographed on the same column (lower panel). Fractionscomprising the maximum enzyme activity after the secondchromatography, indicated by shaded area, were pooled(rechromatography fraction). With pig muscle homogenateas starting material, an 1100-fold purification was obtained.The DEAE-cellulose fraction was also purified by affinitychromatography and a 3000-fold purification was obtainedby this method (data not shown). This value was of the sameorder reported by Duckworth and Kitabchi in rat skeletalmuscle.2*3 The enzyme purified by DEAE-cellulose chroma-tography showed several bands on polyacrylamide-gel discelectrophoresis. However, the enzyme protein obtainedfrom Sephadex G-200 rechromatography migrated as a sin-gle component at the same position as the fraction from af-finity chromatography (Figure 2). Thus, IDE was highly puri-fied not only by affinity chromatography but also bySephadex G-200 column rechromatography of the DEAE-cellulose fraction. Since the procedure of affinity chroma-tography is more difficult and not suitable for handling largequantities of enzyme protein, the method of rechroma-
DIABETES, VOL. 29, OCTOBER 1980 857
IDE FROM PIG AND RAT SKELETAL MUSCLE
(1)
FIGURE 2. Polyacrylamlde-gel disc electrophoretic pattern of variousenzyme preparations: (1) DEAE-cellulose fraction, (2) rechroma-tography fraction, and (3) affinity fraction.
tography on Sephadex G-200 was used for immunization. Ithas the advantage of being simple, and it yields a fairly pureenzyme preparation.
As shown in Figure 3, double-immunodiffusion of the af-finity fraction with antiserum to the rechromatography frac-tion showed a single precipitin band. Quantitative precipitin
100
•—• Pig IDE
o—o Rat Cytosol IDE
Rat Membrane IDE
+ NORMAL RABBITo SERUM
ANTI-PIG IDESERUM
0 50 100 200 300 450SERUM
FIGURE 4. Antigen-antibody titratlons obtained with anti-pig muscleIDE serum and the enzymes isolated from three sources. Insulin-degrading activities were measured In the supernatants afterremoving the specific precipitates.
data (Figure 4) showed that insulin-degrading activity of theaffinity fraction was almost entirely inhibited by the anti-serum in contrast to normal rabbit serum. We previously re-ported that enzymatic characteristics of insulin-degradingactivity in the cytosol (cytosol-IDE) and plasma membranes(membrane-IDE) partially purified from rat skeletal musclewere similar to those of IDE from pig skeletal muscle.8 Ratcytosol and membrane-IDEs showed a single precipitinband continuous with the precipitin band obtained with theaffinity fraction in the Ouchterlony double-immunodiffusion.Enzyme activities of both rat IDEs were inhibited equally by
FIGURE 3. Ouchterlony double-immunodiffusionwith antiserum to pig muscle IDE. The antiserum(10 fi\) to pig muscle IDE was placed in thecentral well. The peripheral wells received 10 MIof IDEs in the following order, starting from thearrow In a clockwise direction: pig muscle, ratmuscle cytosol fraction, and rat muscle plasmamembrane fraction. The remaining wells wereempty.
858 DIABETES, VOL. 29, OCTOBER 1980

KOICHI YOKONO AND ASSOCIATES
the antiserum but not as efficiently as was the pig IDE. Thereason for this difference is not clear. However, these datado suggest that pig muscle insulin protease is both bio-chemically and immunologically similar to the rat enzyme.
The procedure described in this paper should allow wide-spread use of antibodies to insulin-degrading enzyme.Hopefully it will help to dissect the physiologic role playedby IDE in normal and pathologic states.
ACKNOWLEDGMENTSWe acknowledge the valuable advice and the excellenttechnical assistance of Dr. Shigenori Emi, Toyobo Co.
REFERENCES1 Mirsky, I. A.: Insulinase, insulinase-inhibitors, and diabetes mellitus.
Recent Prog. Horm. Res. 73:429-72, 1957.2 Duckworth, W. C, Heinemann, M. A., and Kitabchi, A. E.: Purification
of insulin-specific protease by affinity chromatography. Proc. Natl. Acad. Sci.USA 69:3698-3702, 1972.
3 Duckworth, W. C, and Kitabchi, A. E.: Insulin and glucagon degrada-tion by the same enzyme. Diabetes 23:536-43, 1974.
4 Baba, S., Sakai, K, Imamura, Y., Yokono, K., and Emi, S.: Metabolismof insulin and proinsulin. In Proinsulin, insulin, C-peptide. Baba, S., Ed. Am-sterdam, Excerpta Medica, 1979, pp. 270-282.
5 Burghen, G. A., Kitabchi, A. E., and Brush, J. S.: Characterization of arat liver protease with specificity for insulin. Endocrinology 91:633-42, 1972.
8 Morgan, C. R., Spah, J., Frazier, V., and Fleitz, S.: Insulin, a possibleinducer of the biosynthesis of rat liver insulinase. Proc. Soc. Exp. Biol. Med.728:795-97, 1968.
7 Kitabchi, A. E., Stentz, F. B., Cole, C, and Duckworth, W. C: Acceler-ated insulin degradation: an alternate mechanism for insulin resistance. Dia-betes Care 2:414-17, 1979.
8 Yokono, K., Imamura, Y., Sakai, H, and Baba, S.: Insulin-degradingactivity of plasma membranes from rat skeletal muscle. Its isolation, charac-terization, and biologic significance. Diabetes 28:810-817, 1979.
9 Stollar, D., and Levine, L: In Methods in Enzymology, Vol. VI. Co-lowick, S. P., and Kaplan, N. 0., Eds. New York, Academic Press, 1963, pp.848-54.
10 Varandani, P. T.: Insulin degradation. I. Purification and properties ofglutathione-insulin transhydrogenase of rat liver. Biochim. Biophys. Acta286:126-35, 1972.
11 Lowry, H. 0., Rosebrough, N. J., Farr, A. L, and Randall, R. J.: Pro-tein measurement with the Folin phenol reagent. J. Biol. Chem. 53:265-75,1951.
DIABETES, VOL 29, OCTOBER 1980 859