hypothalamic nesfatin-1 stimulates sympathetic...
TRANSCRIPT

Mamoru Tanida,1 Hitoshi Gotoh,2 Naoki Yamamoto,3 Mofei Wang,1 Yuhichi Kuda,1
Yasutaka Kurata,1 Masatomo Mori,4 and Toshishige Shibamoto1
Hypothalamic Nesfatin-1 StimulatesSympathetic Nerve Activity viaHypothalamic ERK SignalingDiabetes 2015;64:3725–3736 | DOI: 10.2337/db15-0282
Nesfatin-1 acts on the hypothalamus and regulates theautonomic nervous system. However, the hypothalamicmechanisms of nesfatin-1 on the autonomic nervoussystem are not well understood. In this study, we found thatintracerebroventricular (ICV) administration of nesfatin-1increased the extracellular signal–regulated kinase (ERK)activity in rats. Furthermore, the activity of sympatheticnerves, in the kidneys, liver, and white adipose tissue(WAT), and blood pressure was stimulated by the ICVinjection of nesfatin-1, and these effects were abolishedowing to pharmacological inhibition of ERK. Renalsympathoexcitatory and hypertensive effects were alsoobserved with nesfatin-1 microinjection into the paraven-tricular hypothalamic nucleus (PVN). Moreover, nesfatin-1increased the number of phospho (p)-ERK1/2–positiveneurons in the PVN and coexpression of the protein inneurons expressing corticotropin-releasing hormone (CRH).Pharmacological blockade of CRH signaling inhibitedrenal sympathetic and hypertensive responses to nesfatin-1.Finally, sympathetic stimulation of WAT and increasedp-ERK1/2 levels in response to nesfatin-1 were preservedin obese animals such as rats that were fed a high-fatdiet and leptin receptor-deficient Zucker fatty rats.These findings indicate that nesfatin-1 regulates the au-tonomic nervous system through ERK signaling in PVN-CRH neurons to maintain cardiovascular function andthat the antiobesity effect of nesfatin-1 is mediated byhypothalamic ERK-dependent sympathoexcitation inobese animals.
Nesfatin-1 is an 82-amino acid neuropeptide produced inthe hypothalamus that acts on the brain to suppress
appetite (1–6), increase energy expenditure (7), and in-duce cardiovascular changes, leading to body weight re-duction (1), blood pressure (BP) elevation (8–10), andincreased insulin sensitivity (11) in animals. Intracerebro-ventricular (ICV) administration of nesfatin-1 reducesfood intake and body weight gain and elevates BP, heartrate (HR) (8), and peripheral glucose uptake (11). Thus,central nesfatin-1 may regulate the function of peripheralorgans through neural activity to maintain homeostasisand regulate a number of physiological processes.
Regarding the neural mechanism of physiological reg-ulation by nesfatin-1, our previous study demonstratedthat sympathetic nervous supply to the kidneys in anes-thetized rats could be stimulated by the ICV injection ofnesfatin-1 (10), suggesting that the sympathetic nervoussystem mediates the action of nesfatin-1. Recently, it hasbeen reported that increased sympathetic stimulation ofwhite adipose tissue (WAT) and the liver resulted in lipol-ysis (12) and glucogenesis (13), respectively, with the in-tracerebral administration of leptin, a feeding regulatorand sympathetic activator. Thus, central nesfatin-1 maymodulate sympathetic nerve outflow to WAT and theliver and regulate lipid and glucose metabolism; however,there are no studies reporting the effect of ICV nesfatin-1on the neural activity of sympathetic nerves to WAT andthe liver.
The hypothalamus performs a crucial role in coordi-nating the autonomic control of abdominal organs bythe sympathetic and parasympathetic nerves. Nesfatin-1expression has been demonstrated in several hypotha-lamic nuclei including the paraventricular nucleus (PVN),supraoptic nucleus, arcuate nucleus (ARC), and lateral
1Department of Physiology II, Kanazawa Medical University, Uchinada, Ishikawa,Japan2Department of Biology, Kyoto Prefectural University of Medicine, Kyoto, Japan3College of Pharmacology, Hokuriku University, Kanazawa, Ishikawa, Japan4Kitakanto Molecular Novel Research Institute for Obesity and Metabolism, MidoriCity, Gunma, Japan
Corresponding author: Mamoru Tanida, [email protected].
Received 27 February 2015 and accepted 20 July 2015.
This article contains Supplementary Data online at http://diabetes.diabetesjournals.org/lookup/suppl/doi:10.2337/db15-0282/-/DC1.
© 2015 by the American Diabetes Association. Readers may use this article aslong as the work is properly cited, the use is educational and not for profit, andthe work is not altered.
Diabetes Volume 64, November 2015 3725
METABOLISM

hypothalamic area (14,15). Moreover, oxytocin neurons inthe PVN play an important role in mediating feeding re-duction induced by nesfatin-1 (2). These reports suggestthat neural transmission in the PVN mediates the hypo-thalamic action of nesfatin-1 on feeding regulation.
Hypothalamic intracellular signaling, including extra-cellular signal–regulated kinase (ERK), phosphoinositol-3kinase (PI3K), and AMPK, plays important roles in an-orexia and sympathetic stimulation following the centraladministration of leptin (12,13,16–18). Although a recep-tor specific to nesfatin-1 has not yet been identified, thedifferent signaling systems mediated by nesfatin-1 havebeen clarified. Nesfatin-1 stimulates the phosphorylationof mitogen-activated protein kinase (MAPK) (19), CREBP(20), and AMPK (11). However, the precise signalingmechanisms underlying nesfatin-1–induced activation ofthe sympathetic nervous tones in the brain remain to bedetermined. Thus, we examined the effect of nesfatin-1on hypothalamic intracellular signaling and, through phar-macological inhibition studies, examined the role of the in-tracellular pathway in mediating the effect of nesfatin-1on sympathetic nerve stimulation of abdominal organs,cardiovascular function, and feeding behavior.
RESEARCH DESIGN AND METHODS
AnimalsMale Wistar rats (weighing 250–270 g) and Zucker fattyrats (weighing 340–385 g) were used in these studies.Animals were housed in a room maintained at 24 6 1°Cand illuminated for 12 h (8:00 A.M. to 8:00 P.M.). Rats hadfree access to food and water, and were allowed to adaptto the environment for at least 1 week before experimen-tation. Dietary obesity was induced in rats via feedingwith a 60% high-fat diet (HFD) (HFD-60; Oriental YeastCo., Ltd., Tokyo, Japan) for 10 weeks. All animal care andhandling procedures were approved by the Animal Re-search Committees of Kanazawa Medical University.
Brain CannulationRats were equipped with ICV, lateral cerebroventricular,and ARC cannulae using a stereotaxic apparatus, asdescribed previously (12,13). The PVN of rats was cannu-lated unilaterally using a 25-gauge guide cannula withcoordinates (215 mm anteroposterior, +0.5 mm medio-lateral, and 27.5 mm dorsoventral with 0°) according tothe atlas of Paxinos and Watson (21). To verify the accu-racy of PVN and ARC injections, the brain was sectioned,and brain slices were counterstained with cresyl violetsolution to visualize the injection site.
Recording of Sympathetic Nerve ActivitySeven to 10 days after recovery from brain cannulation,anesthesia was induced in rats via intraperitoneal injectionsof a urethane (750 mg/kg) and a-chloralose (75 mg/kg)mixture. Autonomic nerve activity measurements were per-formed as described previously (12,13). Renal sympatheticnerve activity (RSNA), WAT sympathetic nerve activity (WAT-SNA) projecting to the adipose tissue of the epididymis,
and liver sympathetic nerve activity (Liv-SNA) in ratswere recorded in separate animals. The BP signal wasalso sampled using PowerLab and was stored on a harddisk for offline analysis calculating mean arterial pres-sure (MAP) and HR.
The baseline measurements of SNAs were made 5–10 minprior to the ICV injection of vehicle (artificial cerebrospinalfluid [aCSF], 10 mL) or nesfatin-1 (200 pmol/10 mL aCSF).The other groups of rats received unilateral nesfatin-1(50 pmol/0.5 mL aCSF) microinjection into the PVN orARC. In experiments using pharmacological inhibitors,animals first received ICV U0126 (7 mg), LY294002 (5 mg),SB203580 (0.5 mg), astressin 2B (30 mg), H4928 (9 nmol), orvehicle (DMSO; 5 mL) followed 15 min later by ICV nesfatin-1or vehicle (aCSF).
Feeding ExperimentThe body weight and food intake of individually cagedrats were measured before and after treatment. Seven to10 days after ICV surgery, overnight-fasted rats (n = 7animals per group) were assigned to receive an ICV in-jection of vehicle (DMSO; 5 mL) or U0126 (7 mg) followed15 min later by an ICV injection of vehicle (aCSF; 10 mL)or nesfatin-1 (200 pmol) at 8:00 P.M. Body weight and foodintake were measured 4 and 12 h after ICV nesfatin-1 orvehicle administration. In Zucker fatty or HFD rats (n = 5or 6 animals per group), after overnight fasting, food in-take was measured 4 h after ICV injection of vehicle ornesfatin-1.
Immunohistochemistry and In Situ HybridizationAll rats were fasted overnight. Thirty minutes after thelateral cerebroventricular injection of nesfatin-1 (400pmol) or vehicle (aCSF; 10 mL), rats were anesthetizedand perfused intracardially with saline followed by 4%paraformaldehyde in 0.1 mol/L PBS; lateral cerebroven-tricular injection of nesfatin-1 increased RSNA in anes-thetized rats (Supplementary Fig. 1). Brains wereremoved, postfixed at 4°C overnight, and cryoprotectedin 30% sucrose for 2 nights. The brains were sliced, andthen each section was treated with a 0.15% H2O2 solutionand incubated in a 0.1% BSA solution for 1 h. Thereafter,the immunohistochemical responses of phospho (p)-ERKin the PVN and ARC were measured as follows: sectionswere incubated with primary antibody solutions of spe-cific polyclonal rabbit antibody against p-ERK (in 1:200dilution; Cell Signaling Technology) as long as 48 h at 4°C.After washing, sections were incubated at 4°C overnightwith biotinylated anti-rabbit IgG secondary antibody (in1:500 dilution; Sigma-Aldrich). After washing sections,immunoreactivity was visualized using a Vectastain ABCKit (Vector Laboratories) and 3,39-diaminobenzidine (DojindoMolecular Technologies, Inc.) as the chromogen. Images ofthe slices were examined under a microscope, and the num-ber of p-ERK–immunoreactive cells in the PVN or ARC wascounted using ImageJ software.
For double staining of the hypothalamic slices, usingfluorescent immunohistochemistry, brain sections were
3726 Nesfatin-1 Regulates Sympathetic Nervous System Diabetes Volume 64, November 2015

incubated with a primary antibody at the following dilutions:rabbit anti–p-ERK1/2, 1:200 (Cell Signaling Technology);and mouse anti-oxytocin, 1:600 (MAB5296; Chemicon). Then,sections were incubated with Alexa Fluor 488–labeledand Alexa Fluor 543–labeled secondary antibody andDAPI and were observed using a fluorescence microscope(DP70 and DP71; Olympus) or a confocal microscope.
Using dual in situ hybridization and immunohisto-chemistry, we examined the types of neurons in the PVNthat colocalized with the p-ERK1/2–positive cells followingnesfatin-1 injection. Fixed brain sections were incubatedwith proteinase K and acetylated with 0.1 mol/L triethanol-amine containing 0.25% acetic anhydride. Digoxigenin-labeled probes were applied onto sections and incubatedat 65°C overnight. Slides were washed with 13 sodiumchloride–sodium citrate containing 50% formamide twicefollowed by maleic acid buffer containing 0.1% Tween 20.Sections were incubated with sheep anti–digoxigenin-alkaline phosphatase (Roche, Zürich, Switzerland) at 4°Covernight. Signals were detected via incubation with anNBT/BCIP solution (Roche). For immunohistochemicalstaining after in situ hybridization, sections were treatedwith heat by microwaving for 5 min in 10 mmol/L citratebuffer (pH 6.0) and cooled to room temperature beforeincubation with anti–p-ERK1/2 antibody. The signals ofp-ERK were visualized using a peroxidase reaction with3,39-diaminobenzidine as the substrate. The followingcDNAs were PCR amplified, cloned into p3T (MoBiTec,Göttingen, Germany) vector, and used for digoxigenin-labeled probe synthesis: corticotropin-releasing hormone(CRH; NM_031019; nt_203–758), arginine vasopressin(AVP; NM_016992; nt_27–513), and thyrotropin-releasinghormone (TRH; NM_013046; nt_136–903). Sections wereobserved under a microscope.
Western BlottingRats were fasted overnight before the ICV injection ofvehicle or nesfatin-1 (100 or 200 pmol/10 mL). Thirtyminutes after ICV injections, animals were killed by de-capitation, and the mediobasal hypothalamus was quicklyremoved and homogenized on ice. A total protein assayand Western blotting with primary antibodies (p-ERK1/2,p-Akt, p-AMPK, p-p38, p-CREBP, total ERK1/2, total Akt,total AMPK, total p38, and total CREBP) were performedas described in our previous studies (12,13).
Measurement of CRH ContentOne hundred twenty minutes after ICV injections (vehicleor nesfatin-1), animals were killed by decapitation, brainswere quickly removed, and the PVN area was dissected inthe frozen hypothalamic sections and homogenized onice. The CRH level in the PVN was measured with anELISA Kit (YK131; Yanaihara Co., Shizuoka, Japan).
Data AnalysisAll data were expressed as the mean 6 SEM. When com-paring the responses of nerve activity and cardiovascularparameters between groups, ANOVA with the Bonferroni
post hoc test was used. When comparing the Westernblotting and immunohistochemistry data between vehicleand nesfatin-1, the Student t test was used. P , 0.05 wasconsidered statistically significant.
RESULTS
Hypothalamic Nesfatin-1 Increases ERK1/2 Activity inRatsTo determine the signaling pathways crucial for nesfatin-1action in the hypothalamus, we examined the effect of theICV administration of nesfatin-1 on hypothalamic intracel-lular signaling factors in vivo.
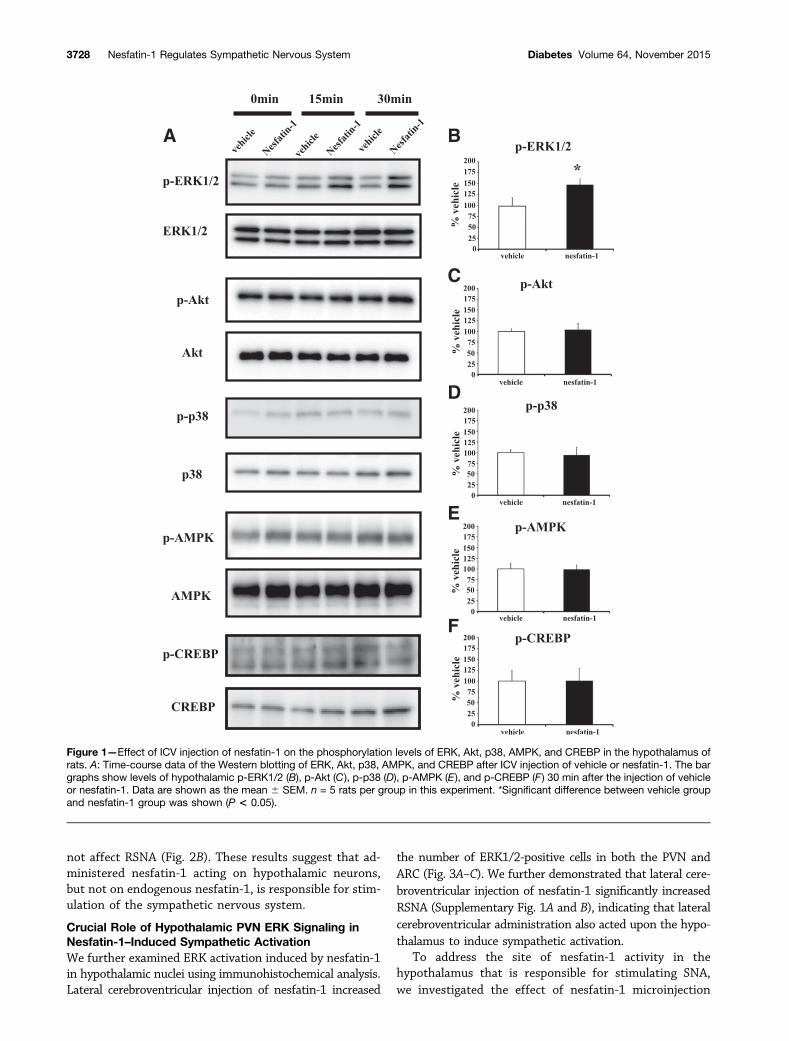
ICV administration of nesfatin-1 increased the levels ofp-ERK1/2 in a dose- and time-dependent manner (Fig. 1Aand Supplementary Fig. 2), with a significant differenceobserved 30 min after injection with nesfatin-1 or vehicle(Fig. 1B). On the contrary, the levels of p38, which is alsoinvolved in MAPK signaling, as well as those of p-Akt,p-AMPK, and p-CREBP were unaltered 30 min after ICVinjection of nesfatin-1 (Fig. 1C–E).
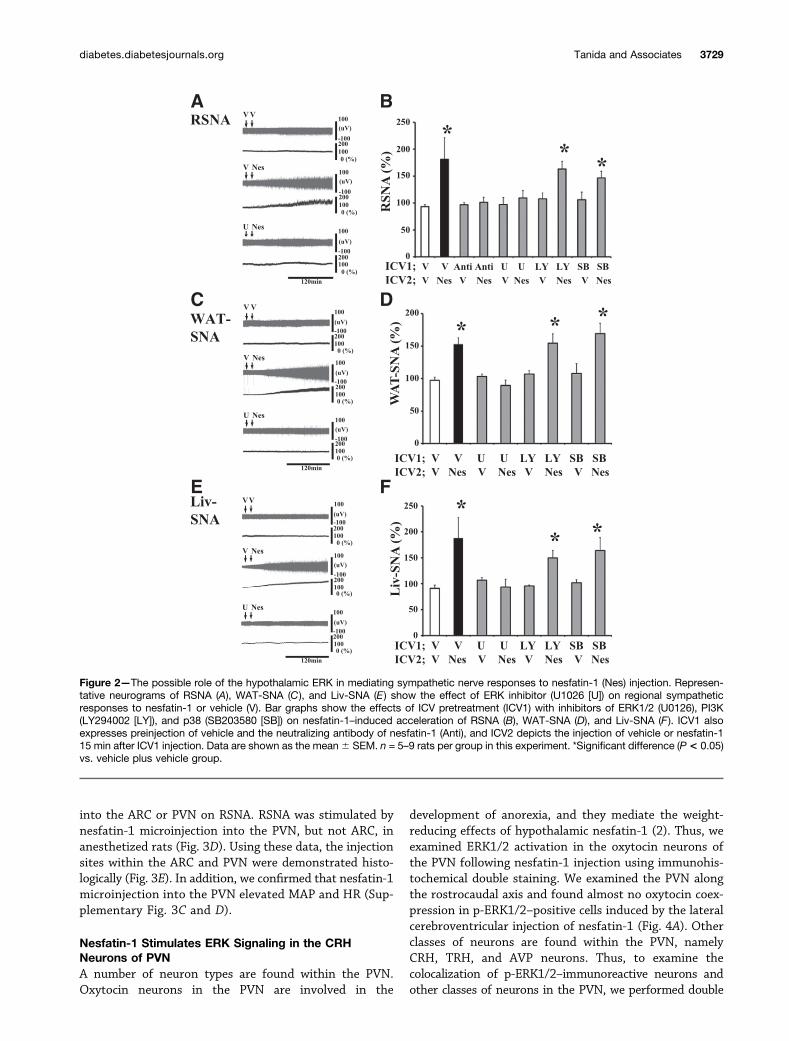
Pharmacological Blockade of ERK AbrogatesSympathetic Nerve Stimulation in Response toNesfatin-1We used a pharmacological approach to examine theeffect of ERK inhibition on sympathetic activation via ICVnesfatin-1 administration. We found that the renal sym-pathetic response to ICV nesfatin-1 administration wasattenuated by ICV preinjection with U0126 (an ERKinhibitor) but not LY20996 (a PI3K inhibitor) orSB203580 (a p38 inhibitor; Fig. 2A and B). On the basisof our previous report (10) demonstrating that the ICVadministration of nesfatin-1 increased MAP and HR, weexamined the role of hypothalamic ERK signaling in cardio-vascular regulation by nesfatin-1. ICV pretreatment withU0126 abrogated the hypertensive and HR-elevating effectof nesfatin-1 (Supplementary Fig. 3A and B). In addition,ICV injection of nesfatin-1 stimulated regional SNA inWAT (Fig. 2C) and liver (Fig. 2E) of anesthetized rats.Increased sympathetic nerve outflows in WAT and theliver in response to nesfatin-1 was blocked by pretreat-ment with U0126, but not by pretreatment withLY20996 or SB203580 (Fig. 2D and F). To reveal theeffect of anesthetics on cardiovascular and sympatheticresponses to nesfatin-1, MAP was measured in consciousrats, and this parameter was elevated 60 min after the ICVinjection of nesfatin-1 (before injection 112 6 4 mmHg,postinjection 1286 5mmHg, change 166 4mmHg, P, 0.05).
Nesfatin-1/NucB2 is expressed in a number of hypo-thalamic nuclei (14,15), and endogenous nesfatin-1 mayaffect sympathetic neurotransmission. To examine thishypothesis, we investigated the effect of neutralizing anti-bodies against nesfatin-1 on ICV nesfatin-1–inducedsympathoexcitation and found that pretreatment withneutralizing antibodies attenuated renal sympathetic ac-tivation by ICV nesfatin-1 (Fig. 2B), whereas neutralizingantibodies administered before the vehicle injection did
diabetes.diabetesjournals.org Tanida and Associates 3727
not affect RSNA (Fig. 2B). These results suggest that ad-ministered nesfatin-1 acting on hypothalamic neurons,but not on endogenous nesfatin-1, is responsible for stim-ulation of the sympathetic nervous system.
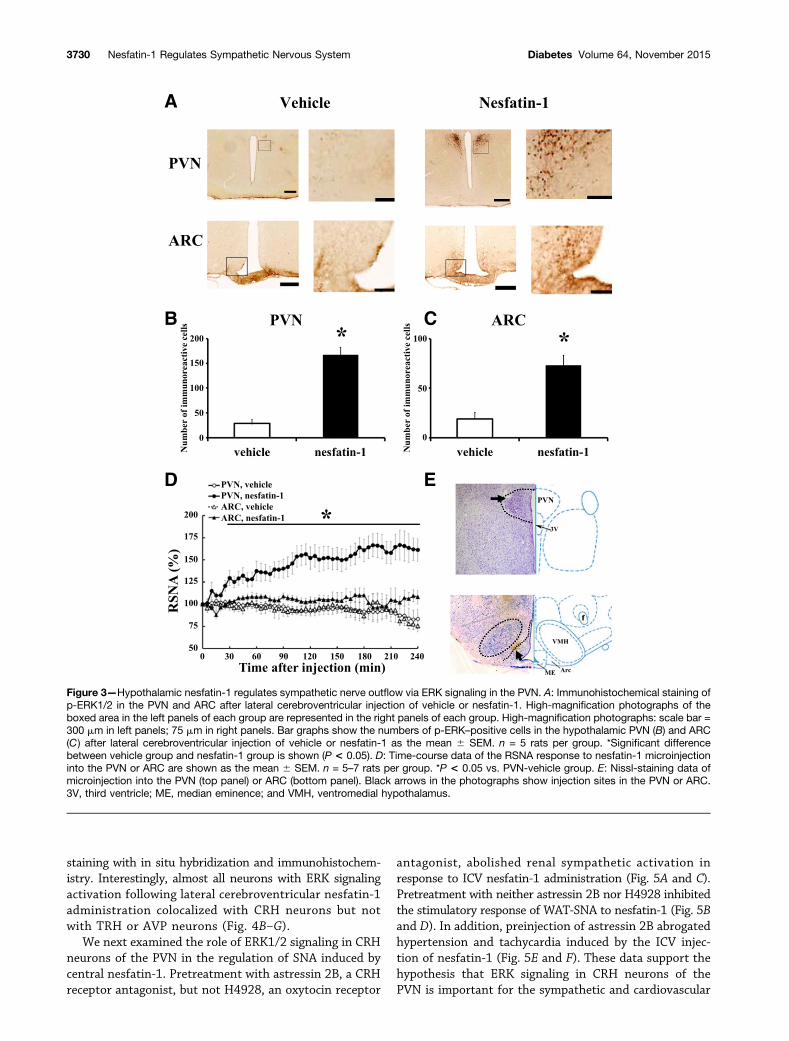
Crucial Role of Hypothalamic PVN ERK Signaling inNesfatin-1–Induced Sympathetic ActivationWe further examined ERK activation induced by nesfatin-1in hypothalamic nuclei using immunohistochemical analysis.Lateral cerebroventricular injection of nesfatin-1 increased
the number of ERK1/2-positive cells in both the PVN andARC (Fig. 3A–C). We further demonstrated that lateral cere-broventricular injection of nesfatin-1 significantly increasedRSNA (Supplementary Fig. 1A and B), indicating that lateralcerebroventricular administration also acted upon the hypo-thalamus to induce sympathetic activation.
To address the site of nesfatin-1 activity in thehypothalamus that is responsible for stimulating SNA,we investigated the effect of nesfatin-1 microinjection
Figure 1—Effect of ICV injection of nesfatin-1 on the phosphorylation levels of ERK, Akt, p38, AMPK, and CREBP in the hypothalamus ofrats. A: Time-course data of the Western blotting of ERK, Akt, p38, AMPK, and CREBP after ICV injection of vehicle or nesfatin-1. The bargraphs show levels of hypothalamic p-ERK1/2 (B), p-Akt (C), p-p38 (D), p-AMPK (E), and p-CREBP (F) 30 min after the injection of vehicleor nesfatin-1. Data are shown as the mean 6 SEM. n = 5 rats per group in this experiment. *Significant difference between vehicle groupand nesfatin-1 group was shown (P < 0.05).
3728 Nesfatin-1 Regulates Sympathetic Nervous System Diabetes Volume 64, November 2015
into the ARC or PVN on RSNA. RSNA was stimulated bynesfatin-1 microinjection into the PVN, but not ARC, inanesthetized rats (Fig. 3D). Using these data, the injectionsites within the ARC and PVN were demonstrated histo-logically (Fig. 3E). In addition, we confirmed that nesfatin-1microinjection into the PVN elevated MAP and HR (Sup-plementary Fig. 3C and D).
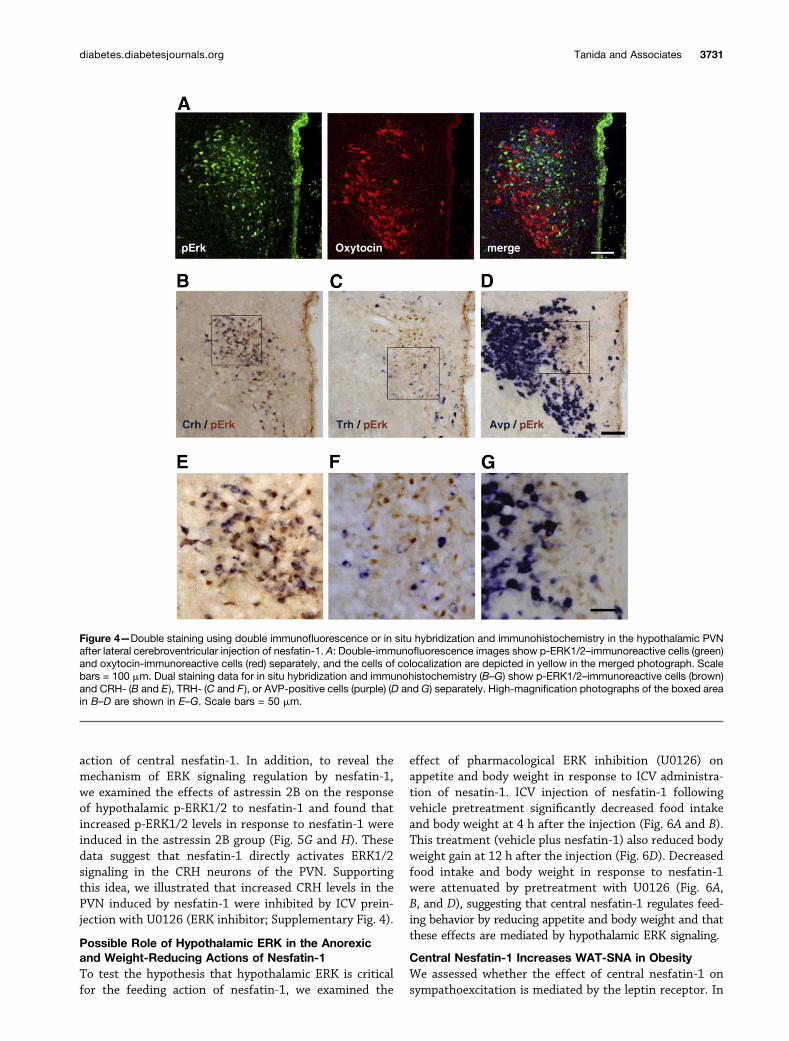
Nesfatin-1 Stimulates ERK Signaling in the CRHNeurons of PVNA number of neuron types are found within the PVN.Oxytocin neurons in the PVN are involved in the
development of anorexia, and they mediate the weight-reducing effects of hypothalamic nesfatin-1 (2). Thus, weexamined ERK1/2 activation in the oxytocin neurons ofthe PVN following nesfatin-1 injection using immunohis-tochemical double staining. We examined the PVN alongthe rostrocaudal axis and found almost no oxytocin coex-pression in p-ERK1/2–positive cells induced by the lateralcerebroventricular injection of nesfatin-1 (Fig. 4A). Otherclasses of neurons are found within the PVN, namelyCRH, TRH, and AVP neurons. Thus, to examine thecolocalization of p-ERK1/2–immunoreactive neurons andother classes of neurons in the PVN, we performed double
Figure 2—The possible role of the hypothalamic ERK in mediating sympathetic nerve responses to nesfatin-1 (Nes) injection. Represen-tative neurograms of RSNA (A), WAT-SNA (C), and Liv-SNA (E) show the effect of ERK inhibitor (U1026 [U]) on regional sympatheticresponses to nesfatin-1 or vehicle (V). Bar graphs show the effects of ICV pretreatment (ICV1) with inhibitors of ERK1/2 (U0126), PI3K(LY294002 [LY]), and p38 (SB203580 [SB]) on nesfatin-1–induced acceleration of RSNA (B), WAT-SNA (D), and Liv-SNA (F ). ICV1 alsoexpresses preinjection of vehicle and the neutralizing antibody of nesfatin-1 (Anti), and ICV2 depicts the injection of vehicle or nesfatin-115 min after ICV1 injection. Data are shown as the mean6 SEM. n = 5–9 rats per group in this experiment. *Significant difference (P< 0.05)vs. vehicle plus vehicle group.
diabetes.diabetesjournals.org Tanida and Associates 3729
staining with in situ hybridization and immunohistochem-istry. Interestingly, almost all neurons with ERK signalingactivation following lateral cerebroventricular nesfatin-1administration colocalized with CRH neurons but notwith TRH or AVP neurons (Fig. 4B–G).
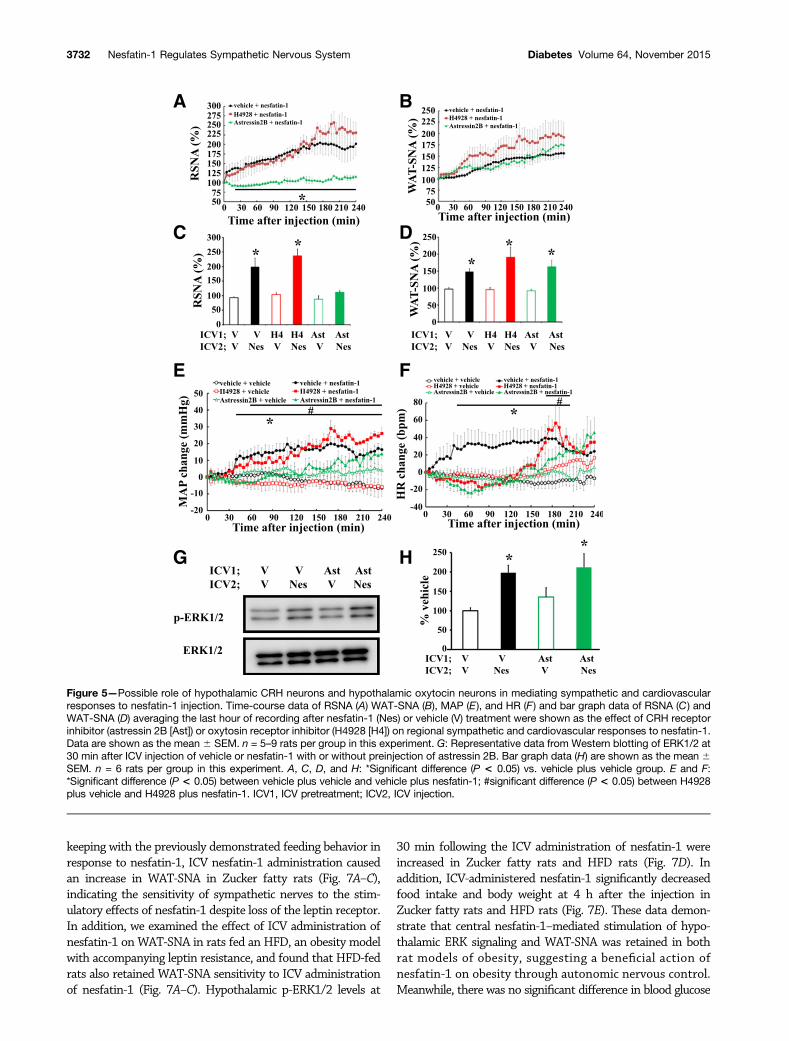
We next examined the role of ERK1/2 signaling in CRHneurons of the PVN in the regulation of SNA induced bycentral nesfatin-1. Pretreatment with astressin 2B, a CRHreceptor antagonist, but not H4928, an oxytocin receptor
antagonist, abolished renal sympathetic activation inresponse to ICV nesfatin-1 administration (Fig. 5A and C).Pretreatment with neither astressin 2B nor H4928 inhibitedthe stimulatory response of WAT-SNA to nesfatin-1 (Fig. 5Band D). In addition, preinjection of astressin 2B abrogatedhypertension and tachycardia induced by the ICV injec-tion of nesfatin-1 (Fig. 5E and F). These data support thehypothesis that ERK signaling in CRH neurons of thePVN is important for the sympathetic and cardiovascular
Figure 3—Hypothalamic nesfatin-1 regulates sympathetic nerve outflow via ERK signaling in the PVN. A: Immunohistochemical staining ofp-ERK1/2 in the PVN and ARC after lateral cerebroventricular injection of vehicle or nesfatin-1. High-magnification photographs of theboxed area in the left panels of each group are represented in the right panels of each group. High-magnification photographs: scale bar =300 mm in left panels; 75 mm in right panels. Bar graphs show the numbers of p-ERK–positive cells in the hypothalamic PVN (B) and ARC(C) after lateral cerebroventricular injection of vehicle or nesfatin-1 as the mean 6 SEM. n = 5 rats per group. *Significant differencebetween vehicle group and nesfatin-1 group is shown (P < 0.05). D: Time-course data of the RSNA response to nesfatin-1 microinjectioninto the PVN or ARC are shown as the mean 6 SEM. n = 5–7 rats per group. *P < 0.05 vs. PVN-vehicle group. E: Nissl-staining data ofmicroinjection into the PVN (top panel) or ARC (bottom panel). Black arrows in the photographs show injection sites in the PVN or ARC.3V, third ventricle; ME, median eminence; and VMH, ventromedial hypothalamus.
3730 Nesfatin-1 Regulates Sympathetic Nervous System Diabetes Volume 64, November 2015
action of central nesfatin-1. In addition, to reveal themechanism of ERK signaling regulation by nesfatin-1,we examined the effects of astressin 2B on the responseof hypothalamic p-ERK1/2 to nesfatin-1 and found thatincreased p-ERK1/2 levels in response to nesfatin-1 wereinduced in the astressin 2B group (Fig. 5G and H). Thesedata suggest that nesfatin-1 directly activates ERK1/2signaling in the CRH neurons of the PVN. Supportingthis idea, we illustrated that increased CRH levels in thePVN induced by nesfatin-1 were inhibited by ICV prein-jection with U0126 (ERK inhibitor; Supplementary Fig. 4).
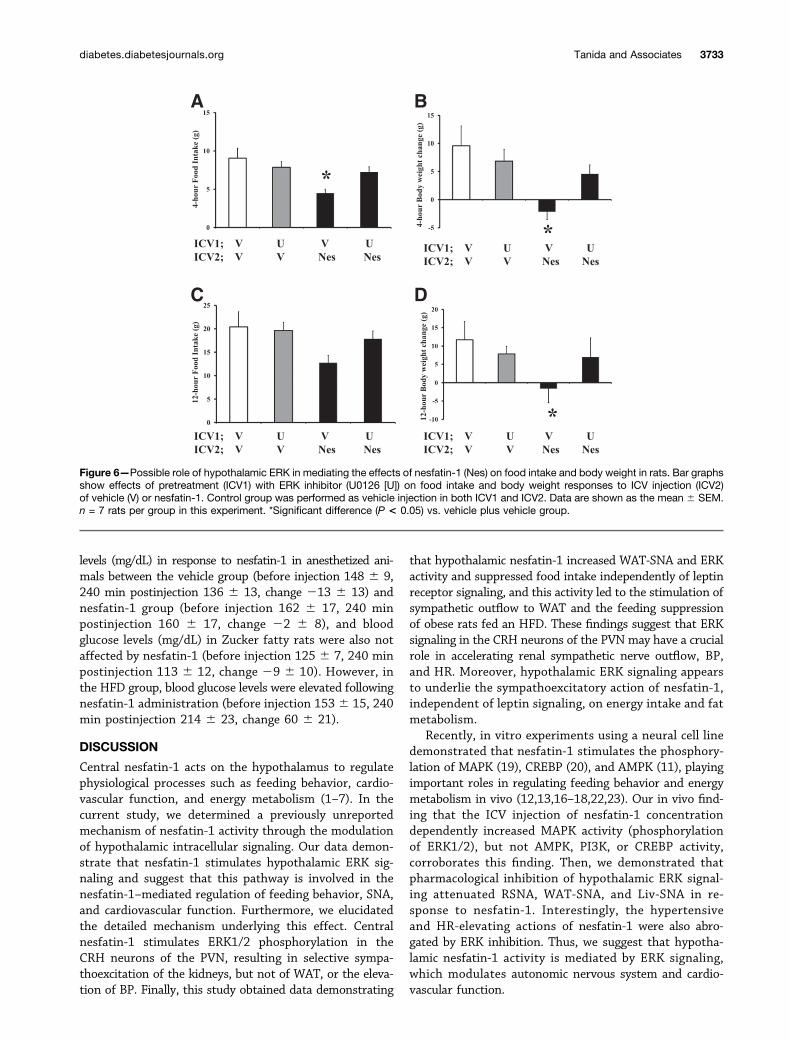
Possible Role of Hypothalamic ERK in the Anorexicand Weight-Reducing Actions of Nesfatin-1To test the hypothesis that hypothalamic ERK is criticalfor the feeding action of nesfatin-1, we examined the
effect of pharmacological ERK inhibition (U0126) onappetite and body weight in response to ICV administra-tion of nesatin-1. ICV injection of nesfatin-1 followingvehicle pretreatment significantly decreased food intakeand body weight at 4 h after the injection (Fig. 6A and B).This treatment (vehicle plus nesfatin-1) also reduced bodyweight gain at 12 h after the injection (Fig. 6D). Decreasedfood intake and body weight in response to nesfatin-1were attenuated by pretreatment with U0126 (Fig. 6A,B, and D), suggesting that central nesfatin-1 regulates feed-ing behavior by reducing appetite and body weight and thatthese effects are mediated by hypothalamic ERK signaling.
Central Nesfatin-1 Increases WAT-SNA in ObesityWe assessed whether the effect of central nesfatin-1 onsympathoexcitation is mediated by the leptin receptor. In
Figure 4—Double staining using double immunofluorescence or in situ hybridization and immunohistochemistry in the hypothalamic PVNafter lateral cerebroventricular injection of nesfatin-1. A: Double-immunofluorescence images show p-ERK1/2–immunoreactive cells (green)and oxytocin-immunoreactive cells (red) separately, and the cells of colocalization are depicted in yellow in the merged photograph. Scalebars = 100 mm. Dual staining data for in situ hybridization and immunohistochemistry (B–G) show p-ERK1/2–immunoreactive cells (brown)and CRH- (B and E), TRH- (C and F ), or AVP-positive cells (purple) (D and G) separately. High-magnification photographs of the boxed areain B–D are shown in E–G. Scale bars = 50 mm.
diabetes.diabetesjournals.org Tanida and Associates 3731
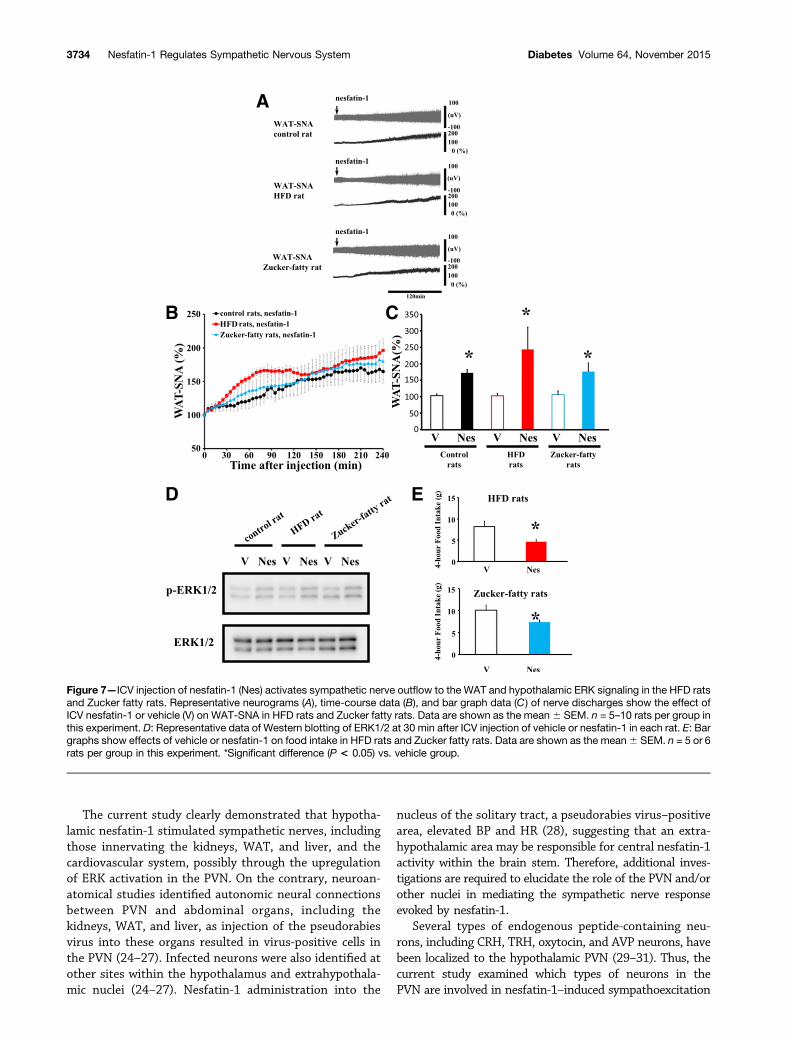
keeping with the previously demonstrated feeding behavior inresponse to nesfatin-1, ICV nesfatin-1 administration causedan increase in WAT-SNA in Zucker fatty rats (Fig. 7A–C),indicating the sensitivity of sympathetic nerves to the stim-ulatory effects of nesfatin-1 despite loss of the leptin receptor.In addition, we examined the effect of ICV administration ofnesfatin-1 on WAT-SNA in rats fed an HFD, an obesity modelwith accompanying leptin resistance, and found that HFD-fedrats also retained WAT-SNA sensitivity to ICV administrationof nesfatin-1 (Fig. 7A–C). Hypothalamic p-ERK1/2 levels at
30 min following the ICV administration of nesfatin-1 wereincreased in Zucker fatty rats and HFD rats (Fig. 7D). Inaddition, ICV-administered nesfatin-1 significantly decreasedfood intake and body weight at 4 h after the injection inZucker fatty rats and HFD rats (Fig. 7E). These data demon-strate that central nesfatin-1–mediated stimulation of hypo-thalamic ERK signaling and WAT-SNA was retained in bothrat models of obesity, suggesting a beneficial action ofnesfatin-1 on obesity through autonomic nervous control.Meanwhile, there was no significant difference in blood glucose
Figure 5—Possible role of hypothalamic CRH neurons and hypothalamic oxytocin neurons in mediating sympathetic and cardiovascularresponses to nesfatin-1 injection. Time-course data of RSNA (A) WAT-SNA (B), MAP (E), and HR (F ) and bar graph data of RSNA (C) andWAT-SNA (D) averaging the last hour of recording after nesfatin-1 (Nes) or vehicle (V) treatment were shown as the effect of CRH receptorinhibitor (astressin 2B [Ast]) or oxytosin receptor inhibitor (H4928 [H4]) on regional sympathetic and cardiovascular responses to nesfatin-1.Data are shown as the mean 6 SEM. n = 5–9 rats per group in this experiment. G: Representative data from Western blotting of ERK1/2 at30 min after ICV injection of vehicle or nesfatin-1 with or without preinjection of astressin 2B. Bar graph data (H) are shown as the mean 6SEM. n = 6 rats per group in this experiment. A, C, D, and H: *Significant difference (P < 0.05) vs. vehicle plus vehicle group. E and F:*Significant difference (P < 0.05) between vehicle plus vehicle and vehicle plus nesfatin-1; #significant difference (P < 0.05) between H4928plus vehicle and H4928 plus nesfatin-1. ICV1, ICV pretreatment; ICV2, ICV injection.
3732 Nesfatin-1 Regulates Sympathetic Nervous System Diabetes Volume 64, November 2015
levels (mg/dL) in response to nesfatin-1 in anesthetized ani-mals between the vehicle group (before injection 148 6 9,240 min postinjection 136 6 13, change 213 6 13) andnesfatin-1 group (before injection 162 6 17, 240 minpostinjection 160 6 17, change 22 6 8), and bloodglucose levels (mg/dL) in Zucker fatty rats were also notaffected by nesfatin-1 (before injection 125 6 7, 240 minpostinjection 113 6 12, change 29 6 10). However, inthe HFD group, blood glucose levels were elevated followingnesfatin-1 administration (before injection 1536 15, 240min postinjection 214 6 23, change 60 6 21).
DISCUSSION
Central nesfatin-1 acts on the hypothalamus to regulatephysiological processes such as feeding behavior, cardio-vascular function, and energy metabolism (1–7). In thecurrent study, we determined a previously unreportedmechanism of nesfatin-1 activity through the modulationof hypothalamic intracellular signaling. Our data demon-strate that nesfatin-1 stimulates hypothalamic ERK sig-naling and suggest that this pathway is involved in thenesfatin-1–mediated regulation of feeding behavior, SNA,and cardiovascular function. Furthermore, we elucidatedthe detailed mechanism underlying this effect. Centralnesfatin-1 stimulates ERK1/2 phosphorylation in theCRH neurons of the PVN, resulting in selective sympa-thoexcitation of the kidneys, but not of WAT, or the eleva-tion of BP. Finally, this study obtained data demonstrating
that hypothalamic nesfatin-1 increased WAT-SNA and ERKactivity and suppressed food intake independently of leptinreceptor signaling, and this activity led to the stimulation ofsympathetic outflow to WAT and the feeding suppressionof obese rats fed an HFD. These findings suggest that ERKsignaling in the CRH neurons of the PVN may have a crucialrole in accelerating renal sympathetic nerve outflow, BP,and HR. Moreover, hypothalamic ERK signaling appearsto underlie the sympathoexcitatory action of nesfatin-1,independent of leptin signaling, on energy intake and fatmetabolism.
Recently, in vitro experiments using a neural cell linedemonstrated that nesfatin-1 stimulates the phosphory-lation of MAPK (19), CREBP (20), and AMPK (11), playingimportant roles in regulating feeding behavior and energymetabolism in vivo (12,13,16–18,22,23). Our in vivo find-ing that the ICV injection of nesfatin-1 concentrationdependently increased MAPK activity (phosphorylationof ERK1/2), but not AMPK, PI3K, or CREBP activity,corroborates this finding. Then, we demonstrated thatpharmacological inhibition of hypothalamic ERK signal-ing attenuated RSNA, WAT-SNA, and Liv-SNA in re-sponse to nesfatin-1. Interestingly, the hypertensiveand HR-elevating actions of nesfatin-1 were also abro-gated by ERK inhibition. Thus, we suggest that hypotha-lamic nesfatin-1 activity is mediated by ERK signaling,which modulates autonomic nervous system and cardio-vascular function.
Figure 6—Possible role of hypothalamic ERK in mediating the effects of nesfatin-1 (Nes) on food intake and body weight in rats. Bar graphsshow effects of pretreatment (ICV1) with ERK inhibitor (U0126 [U]) on food intake and body weight responses to ICV injection (ICV2)of vehicle (V) or nesfatin-1. Control group was performed as vehicle injection in both ICV1 and ICV2. Data are shown as the mean 6 SEM.n = 7 rats per group in this experiment. *Significant difference (P < 0.05) vs. vehicle plus vehicle group.
diabetes.diabetesjournals.org Tanida and Associates 3733
The current study clearly demonstrated that hypotha-lamic nesfatin-1 stimulated sympathetic nerves, includingthose innervating the kidneys, WAT, and liver, and thecardiovascular system, possibly through the upregulationof ERK activation in the PVN. On the contrary, neuroan-atomical studies identified autonomic neural connectionsbetween PVN and abdominal organs, including thekidneys, WAT, and liver, as injection of the pseudorabiesvirus into these organs resulted in virus-positive cells inthe PVN (24–27). Infected neurons were also identified atother sites within the hypothalamus and extrahypothala-mic nuclei (24–27). Nesfatin-1 administration into the
nucleus of the solitary tract, a pseudorabies virus–positivearea, elevated BP and HR (28), suggesting that an extra-hypothalamic area may be responsible for central nesfatin-1activity within the brain stem. Therefore, additional inves-tigations are required to elucidate the role of the PVN and/orother nuclei in mediating the sympathetic nerve responseevoked by nesfatin-1.
Several types of endogenous peptide-containing neu-rons, including CRH, TRH, oxytocin, and AVP neurons, havebeen localized to the hypothalamic PVN (29–31). Thus, thecurrent study examined which types of neurons in thePVN are involved in nesfatin-1–induced sympathoexcitation
Figure 7—ICV injection of nesfatin-1 (Nes) activates sympathetic nerve outflow to the WAT and hypothalamic ERK signaling in the HFD ratsand Zucker fatty rats. Representative neurograms (A), time-course data (B), and bar graph data (C) of nerve discharges show the effect ofICV nesfatin-1 or vehicle (V) on WAT-SNA in HFD rats and Zucker fatty rats. Data are shown as the mean6 SEM. n = 5–10 rats per group inthis experiment. D: Representative data of Western blotting of ERK1/2 at 30 min after ICV injection of vehicle or nesfatin-1 in each rat. E: Bargraphs show effects of vehicle or nesfatin-1 on food intake in HFD rats and Zucker fatty rats. Data are shown as the mean6 SEM. n = 5 or 6rats per group in this experiment. *Significant difference (P < 0.05) vs. vehicle group.
3734 Nesfatin-1 Regulates Sympathetic Nervous System Diabetes Volume 64, November 2015

through ERK signaling. The results demonstrated thatERK1/2-positive cells induced by the central administra-tion of nesfatin-1 colocalized with CRH-expressing neu-rons in PVN but not with neurons expressing TRH, oxytocin,or AVP, and supporting these data, ICV nesfatin-1 admin-istration increased CRH levels in PVN. In addition, wedetermined that hypothalamic nesfatin-1 selectively stim-ulated SNA in the kidneys, but not in WAT or the cardio-vascular system, through CRH neurons. It appears thatERK signaling in CRH neurons in the PVN might contrib-ute to the regulation of autonomic and cardiovascularfunctions by central nesfatin-1 as an underlying mech-anism of the hypothalamic action of nesfatin-1, but themechanism by which nesfatin-1 activates ERK signalingin PVN is unknown. Our data indicated that CRH re-ceptor blocking did not affect the increased phosphor-ylation of ERK1/2 induced by ICV administration ofnesfatin-1, suggesting that stimulated ERK signalingin CRH neurons in PVN is induced by a direct actionof nesfatin-1, opposed to secondary action of CRF re-leased from CRH neurons. Similarly, an in vitro study(32) revealed that CRH failed to stimulate ERK. In ad-dition, an increase in CRH levels in PVN in response tonesfatin-1 was attenuated by preinjection of an ERKinhibitor, supporting our aforementioned idea.
Leptin, an appetite suppressor released from WAT, hasbeen demonstrated to act on the hypothalamus througha similar mechanism as nesfatin-1 in stimulating SNA inrats (12,13,16); however, the anorexic effects of hypotha-lamic nesfatin-1 are not mediated by the same mechanismobserved with activation of the leptin receptor (2). In thecurrent study, the effects of nesfatin-1 on WAT-SNA werealso preserved in Zucker fatty rats lacking the leptin re-ceptor. Interestingly, rats fed an HFD, causing obesity andleptin resistance, also had increased WAT-SNA in re-sponse to nesfatin-1. In addition, both HFD and Zuckerfatty rats had intact hypothalamic ERK signaling sensi-tivity and anorexic responses to central nesfatin-1. Theseresults suggest that central nesfatin-1 suppresses appe-tite and activates SNA in rats, independent of leptinsignaling. Thus, nesfatin-1 may have antiobesity activityin obese animals through a neural pathway mediated byhypothalamic ERK signaling, leading to sympatheticstimulation of WAT and a consequent increase in energymetabolism.
The current study had a number of limitations thatshould be addressed. First, our study could not determinewhether nesfatin-1 action on the CRH neurons in PVN isassociated with hypothalamic proopiomelanocortin neu-rons because previous studies of neural circuits mediatingthe hypertensive effect of hypothalamic nesfatin-1 in thehypothalamus indicated that proopiomelanocortin neu-rons in the ARC are the primary neurons activated bynesfatin-1 before signaling to CRH neurons in the PVN assecondary neurons (9). CRH neurons in the PVN are stim-ulated by nesfatin-1 via two distinct mechanisms; none-theless, our data demonstrated that nesfatin-1 injection into
the PVN, but not into the ARC, caused renal sympatheticnerve activation and BP elevation. This suggests that thedirect action of nesfatin-1 on CRH neurons in PVN isimportant in the regulation of SNA and cardiovascularfunction. Second, the physiological relevance of nesfatin-1–induced sympathoexcitation appears to be tissue spe-cific and dependent on the innervation of the organ. Forinstance, in our study, increased neural activity in thekidneys and WAT induced by nesfatin-1 resulted in BPelevation and metabolic acceleration resulting in bodyweight reduction, respectively. On the contrary, previousstudies (13) on the physiological significance of sympa-thetic innervation of the liver demonstrated hepatic au-tonomic control of glucose metabolism, as stimulation ofliver sympathetic nerves resulted in hyperglycemia. How-ever, our data illustrating that ICV nesfatin-1–inducedincreases in Liv-SNA did not affect blood glucose levelsare inconsistent with those of previous studies of hepaticautonomic innervation. Because increased parasympa-thetic stimulation of the liver suppresses glucose produc-tion (13), central nesfatin-1 may also increase hepaticparasympathetic activity, resulting in unchanged bloodglucose levels. Of course, we will need to investigate thishypothesis in the future.
In conclusion, we demonstrated that ERK signaling inCRH neurons of the hypothalamic PVN plays a crucial rolein the regulation of SNA in the kidneys and cardiovascularfunction by central nesfatin-1. In addition, we describedan ERK-mediated effect of nesfatin-1 on the activation ofWAT-SNA in Zucker fatty rats and a diet-induced ratmodel of obesity, suggesting that nesfatin-1, independentof leptin activity, has beneficial effects in improvingobesity through hypothalamic ERK-SNA signaling.
Funding. This study was supported by grants (to M.T.) from the Ministry ofEducation, Culture, Sports, Science and Technology of Japan (Grant-in-Aid forYoung Scientists 21689008 and 26870672), the Promoted Research fromKanazawa Medical University (S2014-2), and the Takeda Science Foundation.Duality of Interest. No potential conflicts of interest relevant to this articlewere reported.Author Contributions. M.T. conceived and designed the experiments,performed the experiments, analyzed the data, contributed reagents/materials/analysis tools, and wrote the article. H.G. performed the experiments and wrotethe article. N.Y. performed the experiments. M.W. and Y. Kud. analyzed the data.Y. Kur., M.M., and T.S. contributed reagents/materials/analysis tools. M.T. is theguarantor of this work and, as such, had full access to all the data in the studyand takes responsibility for the integrity of the data and the accuracy of the dataanalysis.
References1. Oh-I S, Shimizu H, Satoh T, et al. Identification of nesfatin-1 as a satietymolecule in the hypothalamus. Nature 2006;443:709–7122. Maejima Y, Sedbazar U, Suyama S, et al. Nesfatin-1-regulated oxytocinergicsignaling in the paraventricular nucleus causes anorexia through a leptin-independent melanocortin pathway. Cell Metab 2009;10:355–3653. Stengel A, Goebel M, Wang L, et al. Central nesfatin-1 reduces dark-phasefood intake and gastric emptying in rats: differential role of corticotropin-releasingfactor2 receptor. Endocrinology 2009;150:4911–4919
diabetes.diabetesjournals.org Tanida and Associates 3735

4. Könczöl K, Pintér O, Ferenczi S, et al. Nesfatin-1 exerts long-term effect onfood intake and body temperature. Int J Obes 2012;36:1514–15215. Gotoh K, Masaki T, Chiba S, et al. Nesfatin-1, corticotropin-releasing hor-mone, thyrotropin-releasing hormone, and neuronal histamine interact in thehypothalamus to regulate feeding behavior. J Neurochem 2013;124:90–996. Stengel A, Mori M, Taché Y. The role of nesfatin-1 in the regulation of foodintake and body weight: recent developments and future endeavors. Obes Rev2013;14:859–8707. Wernecke K, Lamprecht I, Jöhren O, Lehnert H, Schulz C. Nesfatin-1 in-creases energy expenditure and reduces food intake in rats. Obesity (SilverSpring) 2014;22:1662–16688. Yosten GL, Samson WK. The anorexigenic and hypertensive effects ofnesfatin-1 are reversed by pretreatment with an oxytocin receptor antagonist. AmJ Physiol Regul Integr Comp Physiol 2010;298:R1642–R16479. Yosten GL, Samson WK. Neural circuitry underlying the central hypertensiveaction of nesfatin-1: melanocortins, corticotropin-releasing hormone, and oxy-tocin. Am J Physiol Regul Integr Comp Physiol 2014;306:R722–R72710. Tanida M, Mori M. Nesfatin-1 stimulates renal sympathetic nerve activity inrats. Neuroreport 2011;22:309–31211. Yang M, Zhang Z, Wang C, et al. Nesfatin-1 action in the brain increasesinsulin sensitivity through Akt/AMPK/TORC2 pathway in diet-induced insulin re-sistance. Diabetes 2012;61:1959–196812. Tanida M, Yamamoto N, Shibamoto T, Rahmouni K. Involvement of hypo-thalamic AMP-activated protein kinase in leptin-induced sympathetic nerve ac-tivation. PLoS One 2013;8:e5666013. Tanida M, Yamamoto N, Morgan DA, Kurata Y, Shibamoto T, Rahmouni K.Leptin receptor signaling in the hypothalamus regulates hepatic autonomic nerveactivity via phosphatidylinositol 3-kinase and AMP-activated protein kinase. JNeurosci 2015;35:474–48414. Foo KS, Brismar H, Broberger C. Distribution and neuropeptide coexistenceof nucleobindin-2 mRNA/nesfatin-like immunoreactivity in the rat CNS. Neuro-science 2008;156:563–57915. Goebel M, Stengel A, Wang L, Lambrecht NW, Taché Y. Nesfatin-1 im-munoreactivity in rat brain and spinal cord autonomic nuclei. Neurosci Lett 2009;452:241–24616. Rahmouni K, Sigmund CD, Haynes WG, Mark AL. Hypothalamic ERK me-diates the anorectic and thermogenic sympathetic effects of leptin. Diabetes2009;58:536–54217. Harlan SM, Guo DF, Morgan DA, Fernandes-Santos C, Rahmouni K. Hy-pothalamic mTORC1 signaling controls sympathetic nerve activity and arterialpressure and mediates leptin effects. Cell Metab 2013;17:599–60618. Dagon Y, Hur E, Zheng B, Wellenstein K, Cantley LC, Kahn BB. p70S6 kinasephosphorylates AMPK on serine 491 to mediate leptin’s effect on food intake. CellMetab 2012;16:104–112
19. Tagaya Y, Miura A, Okada S, Ohshima K, Mori M. Nucleobindin-2 isa positive modulator of EGF-dependent signals leading to enhancement of cellgrowth and suppression of adipocyte differentiation. Endocrinology 2012;153:3308–331920. Ishida E, Hashimoto K, Shimizu H, et al. Nesfatin-1 induces the phos-phorylation levels of cAMP response element-binding protein for intracellularsignaling in a neural cell line. PLoS One 2012;7:e5091821. Paxinos G, Watson C. The Rat Brain in Stereotaxic Coordinates. 6th ed. SanDiego, CA, Academic Press, 200722. Minokoshi Y, Alquier T, Furukawa N, et al. AMP-kinase regulates food intakeby responding to hormonal and nutrient signals in the hypothalamus. Nature2004;428:569–57423. Xu Y, Hill JW, Fukuda M, et al. PI3K signaling in the ventromedial hypo-thalamic nucleus is required for normal energy homeostasis. Cell Metab 2010;12:88–9524. Schramm LP, Strack AM, Platt KB, Loewy AD. Peripheral and centralpathways regulating the kidney: a study using pseudorabies virus. Brain Res1993;616:251–26225. Huang J, Weiss ML. Characterization of the central cell groups regulatingthe kidney in the rat. Brain Res 1999;845:77–9126. Adler ES, Hollis JH, Clarke IJ, Grattan DR, Oldfield BJ. Neurochemicalcharacterization and sexual dimorphism of projections from the brain to ab-dominal and subcutaneous white adipose tissue in the rat. J Neurosci 2012;32:15913–1592127. Buijs RM, la Fleur SE, Wortel J, et al. The suprachiasmatic nucleus balancessympathetic and parasympathetic output to peripheral organs through separatepreautonomic neurons. J Comp Neurol 2003;464:36–4828. Mimee A, Smith PM, Ferguson AV. Nesfatin-1 influences the excitability ofneurons in the nucleus of the solitary tract and regulates cardiovascular function.Am J Physiol Regul Integr Comp Physiol 2012;302:R1297–R130429. Sawchenko PE, Swanson LW. Immunohistochemical identification of neu-rons in the paraventricular nucleus of the hypothalamus that project to themedulla or to the spinal cord in the rat. J Comp Neurol 1982;205:260–27230. Sawchenko PE. Evidence for differential regulation of corticotropin-releasingfactor and vasopressin immunoreactivities in parvocellular neurosecretory andautonomic-related projections of the paraventricular nucleus. Brain Res 1987;437:253–26331. de Greef WJ, Rondeel JM, van Haasteren GA, Klootwijk W, Visser TJ.Regulation of hypothalamic TRH production and release in the rat. Acta MedAustriaca 1992;19(Suppl. 1):77–7932. Cao J, Cetrulo CL, Theoharides TC. Corticotropin-releasing hormone inducesvascular endothelial growth factor release from human mast cells via the cAMP/protein kinase A/p38 mitogen-activated protein kinase pathway. Mol Pharmacol2006;69:998–1006
3736 Nesfatin-1 Regulates Sympathetic Nervous System Diabetes Volume 64, November 2015