hyaluronic acid-based scaffold for central neural...
TRANSCRIPT

Interface Focus (2012) 2, 278–291
on August 31, 2018http://rsfs.royalsocietypublishing.org/Downloaded from
doi:10.1098/rsfs.2012.0016Published online 21 March 2012
REVIEW
*Author for c
One contribuChina’.
Received 6 JaAccepted 20 F
Hyaluronic acid-based scaffold forcentral neural tissue engineering
Xiumei Wang*, Jin He, Ying Wang and Fu-Zhai Cui
Institute for Regenerative Medicine and Biomimetic Materials, State Key Laboratory of NewCeramics and Fine Processing, Department of Materials Science and Engineering,
Tsinghua University, Beijing 100084, People’s Republic of China
Central nervous system (CNS) regeneration with central neuronal connections and restorationof synaptic connections has been a long-standing worldwide problem and, to date, no effectiveclinical therapies are widely accepted for CNS injuries. The limited regenerative capacity of theCNS results from the growth-inhibitory environment that impedes the regrowth of axons. Centralneural tissue engineering has attracted extensive attention from multi-disciplinary scientists inrecent years, and many studies have been carried out to develop cell- and regeneration-activating biomaterial scaffolds that create an artificial micro-environment suitable for axonalregeneration. Among all the biomaterials, hyaluronic acid (HA) is a promising candidate for cen-tral neural tissue engineering because of its unique physico-chemical and biological properties.This review attempts to outline current biomaterials-based strategies for CNS regenerationfrom a tissue engineering point of view and discusses the main progresses in research of HA-basedscaffolds for central neural tissue engineering in detail.
Keywords: central neural tissue engineering; hyaluronic acid; central nervoussystem; regeneration; scaffold
1. INTRODUCTION
Damage to the central nervous system (CNS) includingthe brain and the spinal cord, which is mostly causedby trauma, tumour or disease, usually results in severeneurological impairments with irretrievable nervoussystem functional devastations or limitations such asparalysis, limited mobility and sensory loss. In China, itis roughly estimated that more than 1 500 000 peoplelive with spinal cord injury (SCI), and there are approxi-mately 10 000 new cases every year. Therefore, functionalrecovery of injured CNS has been extremely significant inclinical cases for improving the qualities of patients’ lives.
Unlike peripheral nervous system (PNS) injury,completely functional restoration in damaged CNS isnot possible in most clinical cases. Consequently, CNSregeneration with central neuronal connections andrestoration of synaptic connections has been a long-standing worldwide problem, which has attractedextensive attention from multi-disciplinary scientists.The CNS has limited capacity for regeneration mainlybecause of the growth-inhibitory environment thatimpedes the regrowth of axons. Owing to injuries, aseries of complex cellular and biochemical reactionsare triggered within the CNS and they are usually
orrespondence ([email protected]).
tion of 11 to a Theme Issue ‘Biomaterials research in
nuary 2012ebruary 2012 278
divided into two phases: primary injury and secondaryinjury. Primary injury is an acute phase of injury that iscaused by contusion, laceration, compression or severerotation of the tissue, transection of axons and localblood vessel damage, and finally it leads to cell necrosisat the injury epicentre [1,2]. Except for direct cell deathand haemorrhage, there is no more damage to the CNScaused by the primary injury [3]. However, the adventof secondary injury slowly deteriorates the condition,resulting in complete and permanent trauma to cellsand axons, and functional disabilities in the CNS. Sec-ondary injury is the chronic phase injury, which lastsseveral months or years. After injury, owing to death ofneural cells, breakdown of blood–brain barrier andinflux of inflammatory cells, reactive gliosis is initiatedand glial scars are formed [4]. Although, at the beginning,glial scars seclude the injury site from the healthy tissueand play an important role in restoration of the blood–brain barrier, they finally develop into a tenacious andrubbery membrane acting as a physical barrier thatblocks the outgrowth, penetration and reconnection ofaxons [5]. Besides this physical barrier, in the glial scar,there are several kinds of molecules that act as a bio-chemical barrier, including Nogo, myelin-associatedglycoprotein (Mag), oligodendrocyte myelin glycoprotein(Omgp) and chondroitin sulphate proteoglycan (CSPG),all of which have been demonstrated to be potentinhibitors of neurite outgrowth [6–13].
This journal is q 2012 The Royal Society

Review. HA-based scaffold for CNS regeneration X. Wang et al. 279
on August 31, 2018http://rsfs.royalsocietypublishing.org/Downloaded from
Based on the knowledge of physiology of the CNSand pathology of CNS injuries, the main factors thatdeprive the CNS of the ability to regenerate aremulti-faceted and can be summarized as: inability ofadult neurons to proliferate; absence of neurotrophicfactors; formation of glial scars; existence of inhibitorymolecules. To date, no effective clinical therapies arewidely accepted for CNS injuries, although some arecommonly applied, including methylprednisolone treat-ment and cell therapy, both of which fail to achievecomplete recovery because they overcome only somebut not all of the inhibitory factors. Hence, a multi-faceted approach is needed to design and produce asystem that integrates several factors to conquer allthe inhibitory factors. Tissue engineering uses scaffolds,living cells and regulators to develop ideal biologicalsubstitutes for restoring or regenerating damaged bodytissues. Consequently, by integrating the use of biomater-ials, cells and regulators, such tissue engineering strategysheds new light on CNS regeneration. Since it is bioma-terials (scaffolds) that play a critical role in tissueengineering by acting as biodegradable engineered extra-cellular matrix (ECM) for in vitro or in vivo cellattachment, proliferation, migration, differentiation andtissue formation, the key to successful CNS regenerationwith tissue engineering approaches lies in developingmaterials-based strategies to overcome the inhibitoryenvironment. Recently, higher requirements are raisedin bioactivity of materials [14,15]. Instead of merelybeing able to promote cell adhesion, migration and pro-liferation, the truly bioactive materials are expected tobe capable of inducing certain cellular responses and acti-vating certain gene expressions in the patient’s tissue, soas to make use of their self-healing potential. These idealproperties are underlaid by a special design of the bioma-terial system and it is up to the materials scientists toachieve this goal with multi-disciplinary knowledge.
Here in this review, we first summarize currentbiomaterials-based strategies for central neural tissueengineering, and then we focus on recent progress indesigning and fabricating hyaluronic acid (HA)-basedscaffolds.
2. CURRENT BIOMATERIALS-BASEDSTRATEGIES FOR CENTRAL NERVOUSSYSTEM REGENERATION FROM ATISSUE ENGINEERING POINT OF VIEW
As we all know, tissue engineering triad consists ofscaffolds, cells and regulators (biomolecules), all ofwhich are quite critical for tissue repair or regeneration.However, cells and/or biomolecules are thought to benot definitely necessary to be loaded into scaffoldsand cultured for a while in vitro before transplantation.There is no doubt that if a biomaterial scaffold itself hassufficient bioactivities to recruit endogenous cells andgrowth factors to help tissue regeneration, exogenouscells and molecules could be omitted. Because in vitroapplications of cells and/or biomolecules have encoun-tered a number of translational, manipulation, safetyand regulatory problems, currently biomaterials-basedstrategies for in situ tissue engineering have been an
Interface Focus (2012)
attractive area. Biomaterials scientists have been tryingto design and fabricate ideal biomaterial scaffolds,which are capable of delivering chemical, physical andbiological cues to regulate cell attachment, proliferation,migration, differentiation and neotissue formation byacting as biodegradable engineered ECM. Therefore, bio-materials-based strategies have great promise for tissueengineering and regenerative medicine. Biomaterialscaffolds are not only simply ‘tissue-engineered scaffolds’for cell delivery or cell migration, but also ‘cell- andregeneration-activating systems’ for in situ tissue engin-eering. Numerous studies have focused on design andfabrication of biomaterials. Here, we classify current pro-gress according to the three factors in central neuraltissue engineering.
2.1. Scaffolds
For use in central neural tissue engineering, biomater-ials should meet the following criteria. Biomaterialsshould: integrate well with host tissue without inducinginflammatory reaction and glial scar formation; havesimilar physical properties to the brain or the spinalcord; allow infiltration of cells and axons, and transpor-tation of nutrients and metabolites; exhibit a suitablerate of degradation with no inflammation caused bythe degradation products. On the basis of these require-ments, it is found that among all the biomaterials,hydrogels, electrospun nanofibres and self-assemblingpeptides are ideal candidates and have been applied inmany studies.
Hydrogels are three-dimensional networks of hydro-philic polymer held together by chemical or physicalcross-linking. Hydrogels are glassy in the dry state butthey swell in water and form elastic gels, retaining alarge quantity of water in their mesh-like structures.Many kinds of hydrogels have been used and they can beclassified into naturally derived hydrogels, namely HA,chitosan, alginate, agarose, fibrin, and methylcellulose,and synthetic hydrogels, namely poly(2-hydroxyethylmethacrylate), poly[N-(2-hydroxypropyl) methacryla-mide] and polyethylene glycol. Made of polysaccharides,glycosaminoglycans or ECM constituents, naturallyderived hydrogels are inherently bioactive and allow forcell attachment. Besides, under physiological conditions,naturally derived hydrogels are degradable via enzymaticaction, which facilitates infiltration of cells and axons intothe hydrogels [16]. However, batch variation and the riskof disease transmission may limit the application of natu-rally derived hydrogels. For synthetic hydrogels, they arebiologically inert and cells hardly adhere to them, whichmakes it necessary to adoptmodification such as tetheringof natural polymers or adhesive motifs.
Electrospinning is effective in producing fine fibres ofnanoscale diameters that range from several nanometresto 1 mm [17]. Made of nanofibrous meshes, electrospunscaffolds exhibit high surface-to-volume ratio and highporosity, and thus mimic the hierarchical structuresof laminin and collagen of the ECM, which facilitatescell and axon penetration, offers guidance cues to neuriteextension and enhances scaffold–tissue integration. Alot of polymers have been employed to produce electro-spun fibres, including chitosan [18], poly-L-lactic acid

280 Review. HA-based scaffold for CNS regeneration X. Wang et al.
on August 31, 2018http://rsfs.royalsocietypublishing.org/Downloaded from
(PLLA) [19,20], polycaprolactone (PCL) [21], poly-lactic-glycolic acid (PLGA) [22], polyether sulphone[23], poly(L-lactic acid)-co-poly(3-caprolactone) [24] andpolyurethane [25].
Self-assembly is another method for producingnanofibre networks. Via hydrophobic interaction,oligopeptides or amphiphilic peptides assemble intoself-assembling peptide nanofibre scaffolds (SAPNSs),with hydrophobic backbones forming the core, andthe hydrophilic head groups forming the sheath [26].Compared with electrospun fibres, SAPNSs more clo-sely mimic the hierarchical structure of ECM owing tosmaller fibre diameters and high-density presentationof peptide sequences.
In order to enhance neural regeneration, many studieshave been conducted to optimize the design and fabrica-tion of scaffolds. The effect of modulus of elasticity,architectural structures and modification with ECMcomponents has been extensively investigated.
2.1.1. Modulus of elasticity of scaffoldsModulus of elasticity of scaffolds has recently beenshown to be a key factor that influences cell behaviours.Different adhesion and morphologies of astrocytes andneurons in response to changes in modulus wereobserved [27]. Soft gels tended to suppress the adhesionand proliferation of astrocytes, while modulus did notsignificantly alter the actin formation and neuriteextension. In cultures of dissociated embryonic cortices,it was further demonstrated that the control of moduluswas effective in screening the cells that attached ontothe surface. Moreover, modulus was reported to beable to direct stem cells into different lineages.Mesenchymal stem cells (MSCs) were directed into neu-rogenic lineages when they were cultured on soft gelsthat had modulus similar to that of brain tissue [28].Neural stem cells (NSCs) favoured neuronal lineageson soft gels with modulus less than 1 kPa [29,30].Thus, modulus should be paid attention to when scaf-folds are designed and prepared. Hydrogels withmodulus similar to that of the brain or spinal cordtissue are ideal for use in CNS regeneration.
2.1.2. ArchitectureThere have been many studies on repair of the PNS bytubes or nerve guides, with some promising resultsobserved [31,32]. These anisotropic structures offerphysical guidance to the migration of cells and the pen-etration of axons. Thus, in CNS studies, uniaxialstructures are prepared and the effects of guidancecues are investigated.
There are several ways to create uniaxial structureswith hydrogels. Templating, which is straightforwardand reproducible, is highly effective in fabricating scaf-folds with longitudinal channels [33]. After removal ofPCL fibres by acetone, uniaxial channels are formedwhere PCL fibres had been. Thus, the sizes of the chan-nels can be directly controlled by the diameter of thefibres. It is reported that the implantation of templatedagarose scaffolds resulted in linear and organized pen-etration of axons and blood vessels, in distinction tothe random orientation without scaffolds [34]. However,
Interface Focus (2012)
an in vivo study surprisingly showed that channels withdifferent diameters did not differ in their ability to sup-port axon migration [35]. Moreover, the tubularstructure is far from perfect. Wong et al. [36] high-lighted the importance of microstructures. In thisstudy, the effect of scaffolds with different complexstructure was studied, demonstrating that open-pathdesigns better promoted axonal regeneration, whileclosed designs resulted in the encapsulation of fibroustissues and the enlargement of defects. Freeze-dryingis another way to fabricate uniaxial hydrogel scaffolds[37,38]. With a gradient in temperature, scaffolds withhoneycomb structure were created [38]. Along withother factors, linear growth of axons within channelswas observed and the axons successfully bridgedthe lesion.
Many studies have shown that the orientations ofelectrospun nanofibres are vital in controlling cell beha-viours. Aligned electrospun fibres are mainly fabricatedby collecting the polymer stream at a metal groundeddrum or plate that rotates at a suitable speed [19,39–41]. It was reported that the orientation of neuriteextension was highly dependent on the fibres [20].Further analysis measuring the length of neuritesshowed that the aligned fibres increased neurite exten-sion by 20 per cent in length compared with randomcontrols. This effect was more vividly shown by samplescombining both random controls and aligned fibres [42].At the border of aligned and random nanofibres, theneurites from the same dorsal root ganglion (DRG)cells grew without any preference in direction on theside of random fibres, while on the other side where elec-trospun fibres were aligned, they grew along the fibrealignment; apparently, the neurites were much longeron aligned fibres. If pluripotent cells were incorporated,the alignment was shown to influence cell differen-tiation, with more cells being induced towardsneuronal lineages on aligned fibres for mouse embryonicstem cells (ESCs) [21] and MSCs [43].
2.1.3. Modification of engineered extracellular matrixcomponentsThe ECM molecules are important components of thenervous system. Composed of a heterogeneous latticeof proteoglycans and glycoproteins, ECM providesstructural support and anchorage for cells and regulatescell adhesion, migration and proliferation [44]. Amongall the ECM molecules, laminin and fibronectin,together with their peptide motifs, have been recog-nized to facilitate nerve development and offer aneuroprotective function in the nervous system afterinjury [45–47]. Thus, laminin, fibronectin, collagenand their peptide motifs have been used to modify scaf-folds, so as to improve cell adhesion and axon sprouting,and enhance neural regeneration.
Incorporation of whole ECM molecules, either bycoating, blending, covalently immobilizing, or as fillersin tubes, makes scaffolds more biocompatible, especiallyfor synthetic scaffolds that lack bioactive domains tointeract with cells. Modification with laminin, eitherby covalent binding, physical adsorption or blending,was observed to significantly enhance neurite extension

Review. HA-based scaffold for CNS regeneration X. Wang et al. 281
on August 31, 2018http://rsfs.royalsocietypublishing.org/Downloaded from
compared with negative controls [48]. In this study,blending was shown to be more effective than covalentbinding and physical adsorption, which might resultfrom the higher laminin quantity coupled to PLLAnanofibres by blending. The function of collagen in pro-moting spinal cord regeneration was investigated in vivo[49]. One year after implantation, regeneration of axonand functional recovery were achieved in groups withchitosan tubes filled by semifluid type I collagen. Thismodification stimulated regenerative nerve fibres topenetrate the lesion site and grow into the distal end,while this effect was not observed in negative controls.The incorporation of ECM components may influencedifferentiation of pluripotent cells, but it is still debata-ble and extensive studies are needed [50,51].
Peptides of laminin, collagen and fibronectinare alternative ECM molecules in modification of scaf-folds. Arg–Gly–Asp (RGD), Ile–Lys–Val–Ala–Val(IKVAV) and Tyr–Ile–Gly–Srg–Arg (YIGSR) arethe most widely used. Modification of YIGSR betterenhanced neurite extension compared with controlgroups without YIGSR both in vitro and in vivo [52].Similar results were reported in studies on modificationof RGD and IKVAV [53,54]. Schense et al. [55] system-atically investigated the effect of peptide concentrationand combination on neurite extension. Not all peptideshad linear relationships between neurite outgrowth andincorporated peptide concentration. Different co-cross-linked peptides elicited various effects on the neuriteextension, depending on the combination.
2.2. Regulators
Delivery of antibodies, neurotrophic factors or thera-peutic drugs to lesion sites in the CNS can alleviateinflammation, protect spared tissue around the lesionand promote neural regeneration. Direct injections ofthese bioactive molecules have a relatively short half-life[56]; thus, scaffolds are used to achieve sustained releaseof bioactive molecules. Here, we limit our discussion tothe scaffold-based delivery of neurotrophic factors andtherapeutic drugs, while delivery of antibodies will bediscussed in §3.4.
2.2.1. Delivery of neurotrophic factorsSome of the most commonly used growth factors to pro-mote neural regeneration are neurotrophic factors,including nerve growth factor (NGF) [57], neurotro-phin-3 (NT-3) [58], brain-derived neurotrophin factor(BDNF) [59], glial cell line-derived neurotrophic factor(GDNF) [60] and ciliary neurotrophic factor (CNTF)[61]. Different methods have been developed to deliverthese growth factors, namely physical embedding,heparin-binding and covalent-binding.
Physical embedding is a simple method to deliverneurotrophic factors. Factors are blended with hydrogelsolution and they evenly distribute within the hydrogelscaffolds after gelation [62,63]. Delivery of GDNF in thisway resulted in promotion of axonal outgrowth andsuppression of cystic cavitation [62]. To prolong the invivo availability and reduce running away, carriers areintroduced to deliver growth factors. It was shownthat the dosage of NT-3 to induce NSCs to commit
Interface Focus (2012)
neuronal lineages was much lower in the groups employ-ing chitosan carriers, compared with those of directaddition [64].
Sakiyama-Elbert et al. developed a heparin-baseddelivery system for sustained release of growth factors.This system has been successfully applied to deliverb-fibroblast growth factor [65], NGF [66], platelet-derived growth factor [67] and NT-3 [68]. The potentialof this heparin-based delivery system for use in repair-ing CNS injuries was investigated in vivo [58]. Despitesome promising results in the short term, functionalrecovery study between a group with heparin-NT-3and a negative control did not show a significant differ-ence after 12 weeks, indicating that from a clinical pointof view, the time period for growth factor deliveryshould be further prolonged.
Recently, many studies tried to covalently immobilizeneurotrophic factors onto scaffolds. Compared with phys-ical adsorption, a higher concentration of growth factorswas obtained via immobilization [69]. Instead of compro-mising the bioactivity, immobilization in fact increasedthe efficacy of growth factors [57]. It was reported thatimmobilization of vascular endothelial growth factor(VEGF) outperformed in cell adhesion and viabilityeven at a much lower concentration compared withsoluble VEGF [70]. Thus, immobilization of neurotrophicfactors increases the efficiency, reduces the requireddosageand makes it possible to provide a long-term delivery.
2.2.2. Delivery of therapeutic drugsMethylprednisolone, which is capable of improvingneurologic recovery, is widely used for acute SCI. How-ever, high doses of methylprednisolone increase the riskof unwanted side effects. So it is imperative to find aneffective way to deliver methylprednisolone to lesionsites in doses as small as possible. A delivery systemwas developed to provide a sustained release of methyl-prednisolone [71]. Methylprednisolone was encapsulatedin PLGA nanoparticles embedded in agarose hydrogels.Results showed that this system successfully deliveredmethylprednisolone to injured spinal cord tissue with-out compromising its bioactivity. The inflammationand cystic cavitation were significantly reduced owingto the presence of methylprednisolone.
2.3. Cells
Several cell types have been used for CNS injurystudies, including Schwann cells (SCs) [72], hippocam-pal neurons [73], DRGs [74], olfactory ensheathingcells (OECs) [75], NSCs [76,77], neural progenitor cells(NPCs) [67], ESCs [51] and MSCs [78,79].
In many studies, cell–scaffold complexes are fabri-cated and implanted into CNS lesion sites to evaluatetheir abilities in inducing neural regeneration. It wasobserved that implantation of such cell–scaffold com-plexes resulted in suppression of inflammation andastrocytic scarring around the lesion [80–82], reductionof lesion volume [78], infiltration of regenerating axonsinto the implant [80,82,83], ingrowth of new bloodvessels [82,83], formation of synapses [84] and evenfunctional recovery [78,83,85].

(a) (b)
Figure 1. The structure of HA hydrogels observed using scanning electron microscopy at (a) lower and (b) higher magnification.Reproduced with permission from Hou et al. [94], q 2006 Elsevier. Scale bars, (a) 50mm; (b) 20mm.
282 Review. HA-based scaffold for CNS regeneration X. Wang et al.
on August 31, 2018http://rsfs.royalsocietypublishing.org/Downloaded from
To further enhance regeneration, cell co-culture systemis introduced. Co-culture of endothelial cells and NPCsled to enhanced formation of tubular structures [86].After implantation into lesions in spinal cord, the co-culture implant was found to be highly effective inpromoting angiogenesis, with a twofold increase in func-tional vessels over the implant with endothelial cellsalone [87]. Since OECs and SCs are important cells thatensheath axons, co-culture of SCs with NSCs/NPCsmay further enhance axonal formation and infiltrationcompared with encapsulation of either cells alone.
3. RESEARCH OF HYALURONIC ACID-BASED SCAFFOLDS FOR CENTRALNERVOUS SYSTEM REGENERATION
As discussed earlier, different kinds of biomaterials havebeen investigated for their potential in promoting axonalregeneration. Among them, HA is one of the best candi-dates for the following reasons: as a major componentof soft connective tissue, HA is widely found in mostorgans and tissues [88,89], especially in the CNS [90];owing to its high biocompatibility, HA plays a beneficialrole in wound healing [91,92]; in recent studies, it hasbeen shown that implantation of HA scaffolds reducesglial scar formation [93,94]. To further enhance axonalregeneration with HA scaffolds, a series of strategies areimplemented, and promising results are observed[95,96]. Here, we briefly discuss recent progress on theresearches of HA-based scaffolds for CNS regeneration.
3.1. Pure hyaluronic acid hydrogels as scaffoldsfor central nervous system regeneration
HA hydrogels have an interconnected porous structure(figure 1) [94] that allows transportation of nutritionand penetration of cells, nerve fibres and blood vessels.Thus, HA hydrogels are used as implants to enhanceneural regeneration. HA was demonstrated to be effec-tive in reducing scar formation and enhancing neuralregeneration in both the PNS [97] and CNS [98]. Anin vivo study showed that treatment with HA hydrogelssignificantly inhibited glial scarring, with much smallergliosis thickness and fewer glial fibrillary acidic protein(GFAP)-positive cells around the scarring area [98].However, according to a recent study, only HA hydro-gels with high molecular weight had such an effect to
Interface Focus (2012)
inhibit astrocytic activation, macrophage/microgliainfiltration and CSPG deposition [99]. It was hypoth-esized that this effect resulted from interaction betweenHA and certain receptors via different states of aggrega-tion [100]. However, further experiments are essential toconfirm the results because in the original study, a controlgroup treated with low molecular weight HA was absent,where the influence of low molecular weight cannotdirectly be shown. Besides molecular weight, the modulusof HA hydrogels was reported to be capable of influencingdifferentiation of NPCs [101]. The majority of NPCs cul-tured in hydrogels with similar modulus to that ofneonatal brain tissue differentiated into neurons withextended long processes, while those cultured in hydro-gels with similar modulus to that of adult brain tissuemostly differentiated into astrocytes.
3.2. Hyaluronic acid hydrogels blended with othermaterials as scaffolds for CNS regeneration
A dominant disadvantage of HA is that cells do notadhere to its surface. Hence, HA hydrogels were blendedwith other materials to promote cell adhesion and thusenhance neural regeneration. Spector and his colleaguereported that blending HA with collagen enabledthe fabrication of scaffolds with suitable mechanicalproperties for CNS regeneration [102]. In vitro exper-iments showed that the NSCs cultured in HA–collagen scaffolds favoured neuronal differentiation,which was in accordance with the results reported byForsberg-Nilsson and co-workers [103].
3.3. Modification of hyaluronic acid hydrogels
Another way to promote cell adhesion is modifying HAhydrogels with ECM components, namely laminin, RGD,IKVAV, poly-D-lysine (PDL) and poly-L-lysine (PLL).
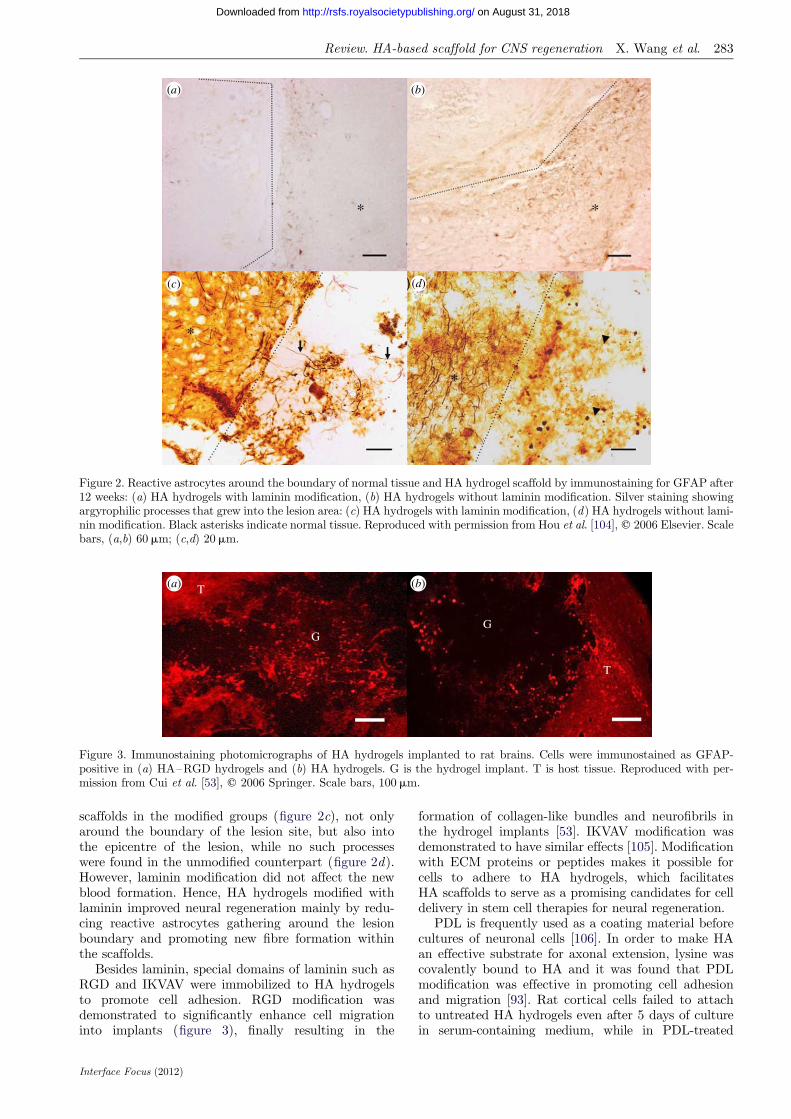
Our group reported that the modification of HA withlaminin better improved tissue reconstruction than thatof negative controls [104]. Figure 2a,b, shows the reac-tive astrocytes around the boundary of the hosttissues and the HA hydrogel scaffold by immunostain-ing after 12 weeks. Fewer GFAP-positive astrocyteswere found in the groups with laminin modification.Statistical analysis showed that after 12 weeks, modifi-cation of laminin reduced one-third of astrocytesaround the boundary. Silver staining revealed the pres-ence of argyrophilic processes that grew into the HA
*
*
*
*
(a) (b)
(c) (d)
Figure 2. Reactive astrocytes around the boundary of normal tissue and HA hydrogel scaffold by immunostaining for GFAP after12 weeks: (a) HA hydrogels with laminin modification, (b) HA hydrogels without laminin modification. Silver staining showingargyrophilic processes that grew into the lesion area: (c) HA hydrogels with laminin modification, (d) HA hydrogels without lami-nin modification. Black asterisks indicate normal tissue. Reproduced with permission from Hou et al. [104], q 2006 Elsevier. Scalebars, (a,b) 60mm; (c,d) 20mm.
(a) T
GG
T
(b)
Figure 3. Immunostaining photomicrographs of HA hydrogels implanted to rat brains. Cells were immunostained as GFAP-positive in (a) HA–RGD hydrogels and (b) HA hydrogels. G is the hydrogel implant. T is host tissue. Reproduced with per-mission from Cui et al. [53], q 2006 Springer. Scale bars, 100 mm.
Review. HA-based scaffold for CNS regeneration X. Wang et al. 283
on August 31, 2018http://rsfs.royalsocietypublishing.org/Downloaded from
scaffolds in the modified groups (figure 2c), not onlyaround the boundary of the lesion site, but also intothe epicentre of the lesion, while no such processeswere found in the unmodified counterpart (figure 2d).However, laminin modification did not affect the newblood formation. Hence, HA hydrogels modified withlaminin improved neural regeneration mainly by redu-cing reactive astrocytes gathering around the lesionboundary and promoting new fibre formation withinthe scaffolds.
Besides laminin, special domains of laminin such asRGD and IKVAV were immobilized to HA hydrogelsto promote cell adhesion. RGD modification wasdemonstrated to significantly enhance cell migrationinto implants (figure 3), finally resulting in the
Interface Focus (2012)
formation of collagen-like bundles and neurofibrils inthe hydrogel implants [53]. IKVAV modification wasdemonstrated to have similar effects [105]. Modificationwith ECM proteins or peptides makes it possible forcells to adhere to HA hydrogels, which facilitatesHA scaffolds to serve as a promising candidates for celldelivery in stem cell therapies for neural regeneration.
PDL is frequently used as a coating material beforecultures of neuronal cells [106]. In order to make HAan effective substrate for axonal extension, lysine wascovalently bound to HA and it was found that PDLmodification was effective in promoting cell adhesionand migration [93]. Rat cortical cells failed to attachto untreated HA hydrogels even after 5 days of culturein serum-containing medium, while in PDL-treated

(a) (b)
5 µm 1 µm
Figure 4. Scanning electron microscopy images of neural cells that adhered to HA–PDL hydrogel. Boxed section in (a) is enlargedin (b) to show the connection (white arrow) between neurons [93].
284 Review. HA-based scaffold for CNS regeneration X. Wang et al.
on August 31, 2018http://rsfs.royalsocietypublishing.org/Downloaded from
groups cells adhered to HA hydrogels and formedinterconnected networks, as shown in figure 4. In a trau-matic brain injury rat model, groups implanted withPDL-treated HA hydrogels showed improved regener-ation of blood vessels and reconstruction of new ECM.These results further demonstrated that PDL-modification was effective in making HA hydrogelsmore biocompatible and promoting angiogenesis.
3.4. Delivery of bioactive agents using hyaluronicacid-based scaffolds
3.4.1. Delivery of Nogo receptor antibodies usinghyaluronic acid-based scaffoldsStudies have shown that the three axonal growth inhibi-tors Nogo, Mag and Omgp require interactions withNogo receptor (NgR) to exert their inhibitory influence[107–109], and blocking of NgR by using polyclonalNogo receptor antibodies (anti-NgRs) successfullyblocked the inhibition of neurite outgrowth by Mag in adose-dependent manner [109]. Hence, sustained deliveryof anti-NgRs in the CNS after injury is effective in over-coming the growth inhibitory environment andpromoting axonal regeneration.
Our group first studied the controlled release ofanti-NgRs via HA hydrogels [110]. The antibodieswere conjugated to the HA backbone by a condensationreaction, with the amount of conjugated antibodiesbeing 135 mg antibody per milligram hydrogel. Therelease was pH-dependent, with nearly 80 per cent ofantibodies being released for up to 400 h in pH 7.4buffer solution. Further in vitro experiments demon-strated that the condensation reaction did notcompromise the bioactivity of anti-NgRs.
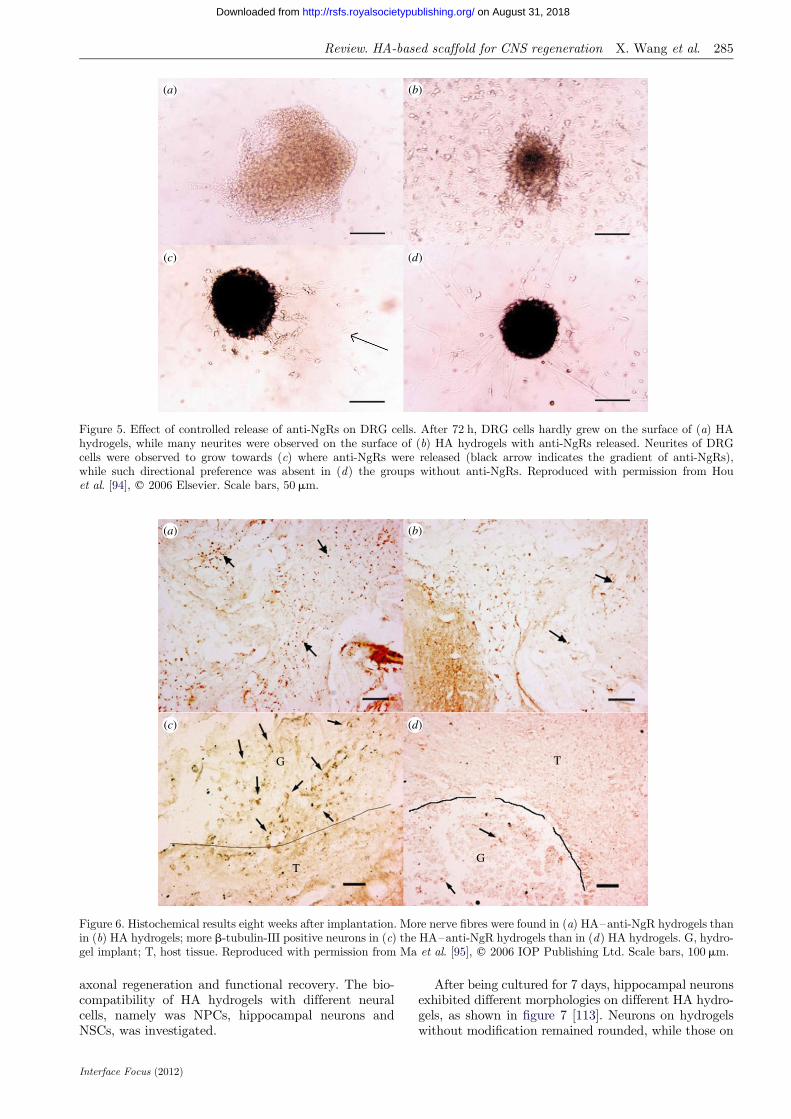
Based on the HA-based delivery system, the effect ofcontrolled release of anti-NgRs via HA hydrogels wasevaluated both in vitro and in vivo. In an in vitrostudy, release of anti-NgRs enhanced adhesion, viabilityand neurite extension of DRG on HA hydrogels [94]. Asshown in figure 5a,b, many more neurites were observedin groups with anti-NgRs than those without. Thiseffect was more vividly shown by releasing anti-NgRsfrom a certain point and thus creating a concentrationgradient of the antibodies. DRG neurites extendedspecifically towards where anti-NgRs were released, asshown in figure 5c.
Interface Focus (2012)
Our group then investigated the effect of controlledrelease of anti-NgRs in a rat stroke model by using HAhydrogels [95]. In behavioural tests, 20 weeks afterimplantation, the groups with HA–anti-NgRs obtainedall pellets in less attempts than the control with HA did,which indicated that the release of anti-NgRs improvedfunctional recovery. The results well matched those ofhistochemical investigations. There are more nerve fibresand b-tubulin-III positive neurons in the HA–anti-NgRhydrogels than in negative controls, as shown in figure6. In an SCI model, similar results were observed, withfew NF-positive fibres discovered in the implants withoutanti-NgRs, while several NF-positive axons were observedentering the HA–anti-NgR–PLL implants [96]. These invivo results demonstrated that controlled delivery of anti-NgRs supported axonal regeneration, which was of greatpotential for clinical applications.
3.4.2. Delivery of neurotrophic factors using hyaluronicacid-based scaffoldsPark et al. [111] reported the effect of controlled release ofBDNF both in vitro and in vivo by using HA hydrogels.BDNF was incorporated into HA hydrogels by electro-static interaction. The release of BDNF significantlychanged the morphology and gene expression of MSCsin culture. In a rat SCI model, HA implants withBDNF better promoted functional recovery as measuredby Basso, Beattie and Bresnahan (BBB) scores. Hence,HA hydrogels had the potential to be ideal carriers ofBDNF. Recently, our group reported the effect of usingPLGA microspheres as carriers for growth factors [112].VEGF and BDNF were incorporated into PLGA micro-spheres by a water-in-oil-in-water emulsion technique,and PLGA microspheres were distributed in HA solutionbefore gelation. The release curves of this HA hydrogel/PLGA microsphere composite began with an initialburst, followed by a stable release phase, with about12 per cent of total loading released after 100 h.
3.5. Delivery of neural cells using hyaluronicacid-based scaffolds
On the basis of studies on the modifications anddelivery of bioactive agents, HA hydrogels are inve-stigated for the purpose of delivering neural cellsto the lesion sites of the CNS, so as to facilitate
(a) (b)
(c) (d)
Figure 5. Effect of controlled release of anti-NgRs on DRG cells. After 72 h, DRG cells hardly grew on the surface of (a) HAhydrogels, while many neurites were observed on the surface of (b) HA hydrogels with anti-NgRs released. Neurites of DRGcells were observed to grow towards (c) where anti-NgRs were released (black arrow indicates the gradient of anti-NgRs),while such directional preference was absent in (d) the groups without anti-NgRs. Reproduced with permission from Houet al. [94], q 2006 Elsevier. Scale bars, 50 mm.
(a)
G
GT
T
(b)
(c) (d)
Figure 6. Histochemical results eight weeks after implantation. More nerve fibres were found in (a) HA–anti-NgR hydrogels thanin (b) HA hydrogels; more b-tubulin-III positive neurons in (c) the HA–anti-NgR hydrogels than in (d) HA hydrogels. G, hydro-gel implant; T, host tissue. Reproduced with permission from Ma et al. [95], q 2006 IOP Publishing Ltd. Scale bars, 100 mm.
Review. HA-based scaffold for CNS regeneration X. Wang et al. 285
on August 31, 2018http://rsfs.royalsocietypublishing.org/Downloaded from
axonal regeneration and functional recovery. The bio-compatibility of HA hydrogels with different neuralcells, namely was NPCs, hippocampal neurons andNSCs, was investigated.
Interface Focus (2012)
After being cultured for 7 days, hippocampal neuronsexhibited different morphologies on different HA hydro-gels, as shown in figure 7 [113]. Neurons on hydrogelswithout modification remained rounded, while those on

(a) (b) (c)
(e) ( f ) (g) (h)
(d)
Figure 7. Morphology of hippocampal neurons cultured on different HA hydrogels for 7 days. (a,e) HA hydrogels without modi-fication. (b, f ) HA–anti-NgR hydrogels. (c,g) HA–PLL hydrogels. (d ,h) HA–PLL–anti-NgR hydrogels [113]. Scale bars,100 mm.
(a) (b)
(c) (d)
Figure 8. Adhesion of NSCs on HA-based hydrogels after 5 days of culture. (a) HA; (b) HA–anti-NgR–PLL; (c) HA–PLL;(d) HA–anti-NgR. Scale bars, (a) 5 mm, (b–d) 10 mm.
286 Review. HA-based scaffold for CNS regeneration X. Wang et al.
on August 31, 2018http://rsfs.royalsocietypublishing.org/Downloaded from
hydrogels modified with PLL or hydrogels cross-linkedwith anti-NgRs exhibited multi-polar morphology. Inaddition, neurons were more evenly distributed on hydro-gels with both PLL and NgR. Interestingly, NPCs wereobserved to behave similarly on these hydrogels [114].
Our recent work focuses on the delivery of NSCs viaHA-based scaffolds. The adhesion of NSCs to HAhydrogels is shown in figure 8. The modifications withPLL and the delivery of anti-NgRs helped neurospheres
Interface Focus (2012)
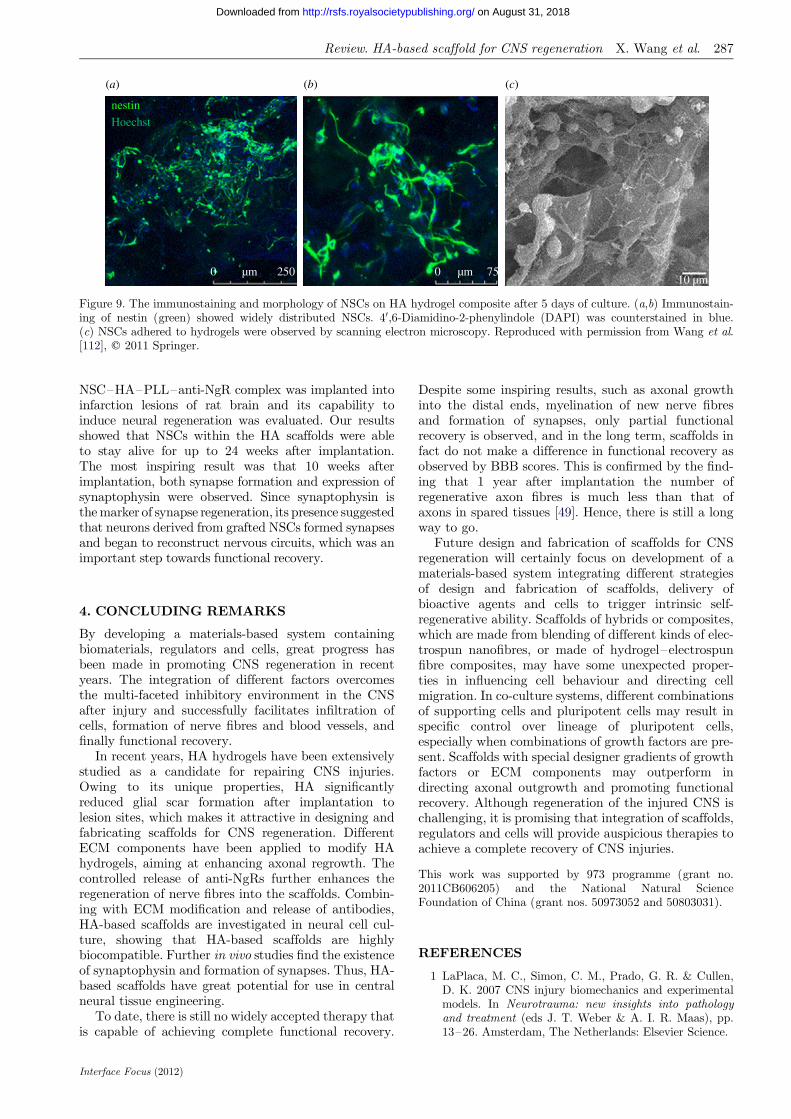
to attach to and spread on HA hydrogels. Incorpora-tion of PLGA microspheres encapsulating VEGF andBDNF further promoted NSC adhesion and pro-liferation [112]. After 5 days of culture, neuritesextended along the wall of the scaffold and formedan extensive network (figure 9). The proliferationassay showed that NSCs on the HA hydrogel withgrowth-factor-containing PLGA microspheres grewfaster than negative controls. In our latest work, the
(a) (b) (c)
nestin
Hoechst
0 250µm 0 75µm10 µm
Figure 9. The immunostaining and morphology of NSCs on HA hydrogel composite after 5 days of culture. (a,b) Immunostain-ing of nestin (green) showed widely distributed NSCs. 40,6-Diamidino-2-phenylindole (DAPI) was counterstained in blue.(c) NSCs adhered to hydrogels were observed by scanning electron microscopy. Reproduced with permission from Wang et al.[112], q 2011 Springer.
Review. HA-based scaffold for CNS regeneration X. Wang et al. 287
on August 31, 2018http://rsfs.royalsocietypublishing.org/Downloaded from
NSC–HA–PLL–anti-NgR complex was implanted intoinfarction lesions of rat brain and its capability toinduce neural regeneration was evaluated. Our resultsshowed that NSCs within the HA scaffolds were ableto stay alive for up to 24 weeks after implantation.The most inspiring result was that 10 weeks afterimplantation, both synapse formation and expression ofsynaptophysin were observed. Since synaptophysin isthe marker of synapse regeneration, its presence suggestedthat neurons derived from grafted NSCs formed synapsesand began to reconstruct nervous circuits, which was animportant step towards functional recovery.
4. CONCLUDING REMARKS
By developing a materials-based system containingbiomaterials, regulators and cells, great progress hasbeen made in promoting CNS regeneration in recentyears. The integration of different factors overcomesthe multi-faceted inhibitory environment in the CNSafter injury and successfully facilitates infiltration ofcells, formation of nerve fibres and blood vessels, andfinally functional recovery.
In recent years, HA hydrogels have been extensivelystudied as a candidate for repairing CNS injuries.Owing to its unique properties, HA significantlyreduced glial scar formation after implantation tolesion sites, which makes it attractive in designing andfabricating scaffolds for CNS regeneration. DifferentECM components have been applied to modify HAhydrogels, aiming at enhancing axonal regrowth. Thecontrolled release of anti-NgRs further enhances theregeneration of nerve fibres into the scaffolds. Combin-ing with ECM modification and release of antibodies,HA-based scaffolds are investigated in neural cell cul-ture, showing that HA-based scaffolds are highlybiocompatible. Further in vivo studies find the existenceof synaptophysin and formation of synapses. Thus, HA-based scaffolds have great potential for use in centralneural tissue engineering.
To date, there is still no widely accepted therapy thatis capable of achieving complete functional recovery.
Interface Focus (2012)
Despite some inspiring results, such as axonal growthinto the distal ends, myelination of new nerve fibresand formation of synapses, only partial functionalrecovery is observed, and in the long term, scaffolds infact do not make a difference in functional recovery asobserved by BBB scores. This is confirmed by the find-ing that 1 year after implantation the number ofregenerative axon fibres is much less than that ofaxons in spared tissues [49]. Hence, there is still a longway to go.
Future design and fabrication of scaffolds for CNSregeneration will certainly focus on development of amaterials-based system integrating different strategiesof design and fabrication of scaffolds, delivery ofbioactive agents and cells to trigger intrinsic self-regenerative ability. Scaffolds of hybrids or composites,which are made from blending of different kinds of elec-trospun nanofibres, or made of hydrogel–electrospunfibre composites, may have some unexpected proper-ties in influencing cell behaviour and directing cellmigration. In co-culture systems, different combinationsof supporting cells and pluripotent cells may result inspecific control over lineage of pluripotent cells,especially when combinations of growth factors are pre-sent. Scaffolds with special designer gradients of growthfactors or ECM components may outperform indirecting axonal outgrowth and promoting functionalrecovery. Although regeneration of the injured CNS ischallenging, it is promising that integration of scaffolds,regulators and cells will provide auspicious therapies toachieve a complete recovery of CNS injuries.
This work was supported by 973 programme (grant no.2011CB606205) and the National Natural ScienceFoundation of China (grant nos. 50973052 and 50803031).
REFERENCES
1 LaPlaca, M. C., Simon, C. M., Prado, G. R. & Cullen,D. K. 2007 CNS injury biomechanics and experimentalmodels. In Neurotrauma: new insights into pathologyand treatment (eds J. T. Weber & A. I. R. Maas), pp.13–26. Amsterdam, The Netherlands: Elsevier Science.

288 Review. HA-based scaffold for CNS regeneration X. Wang et al.
on August 31, 2018http://rsfs.royalsocietypublishing.org/Downloaded from
2 Hulsebosch, C. E. 2002 Recent advances in pathophysiol-ogy and treatment of spinal cord injury. Adv. Physiol.Educ. 26, 238–255. (doi:10.1152/advan.00039.2002)
3 Beattie, M. S., Hermann, G. E., Rogers, R. C. &Bresnahan, J. C. 2002 Cell death in models of spinalcord injury. In Spinal cord trauma: regeneration, neuralrepair and functional recovery (eds L. McKerracher,G. Doucet & S. Rossignol), pp. 37–47. Amsterdam,The Netherlands: Elsevier Science.
4 Yiu, G. & He, Z. 2006 Glial inhibition of CNS axon regen-eration. Nat. Rev. Neurosci. 7, 617–627. (doi:10.1038/nrn1956)
5 Silver, J. & Miller, J. H. 2004 Regeneration beyond theglial scar. Nat. Rev. Neurosci. 5, 146–156. (doi:10.1038/nrn1326)
6 Chen, M. S., Huber, A. B., van der Haar, M. E.,Frank, M., Schnell, L., Spillmann, A. A., Christ, F. &Schwab, M. E. 2000 Nogo-A is a myelin-associated neur-ite outgrowth inhibitor and an antigen for monoclonalantibody IN-1. Nature 403, 434–439. (doi:10.1038/35000219)
7 GrandPre, T., Nakamura, F., Vartanian, T. &Strittmatter, S. M. 2000 Identification of the Nogoinhibitor of axon regeneration as a reticulon protein.Nature 403, 439–444. (doi:10.1038/35000226)
8 Prinjha, R., Moore, S. E., Vinson, M., Blake, S., Morrow,R., Christie, G., Michlovich, D., Simmons, D. L. &Walsh, F. S. 2000 Neurobiology: inhibitor of neurite out-growth in humans. Nature 403, 383–384. (doi:10.1038/35000287)
9 Huber, A. B., Weinmann, O., Brosamle, C., Oertle, T. &Schwab, M. E. 2002 Patterns of Nogo mRNA and proteinexpression in the developing and adult rat and after CNSlesions. J. Neurosci. 22, 3553–3567.
10 Wang, X. X., Chun, S. J., Treloar, H., Vartanian, T.,Greer, C. A. & Strittmatter, S. M. 2002 Localization ofNogo-A and Nogo-66 receptor proteins at sites ofaxon-myelin and synaptic contact. J. Neurosci. 22,5505–5515.
11 McKerracher, L., David, S., Jackson, D. L., Kottis, V.,Dunn, R. J. & Braun, P. E. 1994 Identification ofmyelin-associated glycoprotein as a major myelin-derivedinhibitor of neurite growth. Neuron 13, 805–811. (doi:10.1016/0896-6273(94)90247-X)
12 Davies, S. J. A., Goucher, D. R., Doller, C. & Silver, J.1999 Robust regeneration of adult sensory axons indegenerating white matter of the adult rat spinal cord.J. Neurosci. 19, 5810–5822.
13 Vinson, M., Strijbos, P. J. L. M., Rowles, A., Facci, L.,Moore, S. E., Simmons, D. L. & Walsh, F. S. 2001Myelin-associated glycoprotein interacts with gangliosideGT1b: a mechanism for neurite outgrowth inhibition.J. Biol. Chem. 276, 20 280–20 285. (doi:10.1074/jbc.M100345200)
14 Gumera, C., Rauck, B. & Wang, Y. 2011 Materials for cen-tral nervous system regeneration: bioactive cues. J. Mater.Chem. 21, 7033–7051. (doi:10.1039/c0jm04335d)
15 Hench, L. L. & Polak, J. M. 2002 Third-generation bio-medical materials. Science 295, 1014–1017. (doi:10.1126/science.1067404)
16 Namba, R. M., Cole, A. A., Bjugstad, K. B. & Mahoney,M. J. 2009 Development of porous PEG hydrogels thatenable efficient, uniform cell-seeding and permit earlyneural process extension. Acta Biomater. 5, 1884–1897.(doi:10.1016/j.actbio.2009.01.036)
17 Smith, L. A. & Ma, P. X. 2004 Nano-fibrous scaffolds fortissue engineering. Colloids Surf. B 39, 125–131. (doi:10.1016/j.colsurfb.2003.12.004)
Interface Focus (2012)
18 Liu, T., Teng, W. K., Chan, B. P. & Chew, S. Y. 2010Photochemical crosslinked electrospun collagen nanofi-bers: synthesis, characterization and neural stem cellinteractions. J. Biomed. Mater. Res. A 95A, 276–282.(doi:10.1002/jbm.a.32831)
19 Hurtado, A., Cregg, J. M., Wang, H. B., Wendell, D. F.,Oudega, M., Gilbert, R. J. & McDonald, J. W. 2011Robust CNS regeneration after complete spinal cordtransection using aligned poly-L-lactic acid microfibers.Biomaterials 32, 6068–6079. (doi:10.1016/j.biomaterials.2011.05.006)
20 Corey, J. M., Lin, D. Y., Mycek, K. B., Chen, Q., Samuel,S., Feldman, E. L. & Martin, D. C. 2007 Aligned electro-spun nanofibers specify the direction of dorsal rootganglia neurite growth. J. Biomed. Mater. Res. A 83A,636–645. (doi:10.1002/jbm.a.31285)
21 Xie, J., Willerth, S. M., Li, X., Macewan, M. R.,Rader, A., Sakiyama-Elbert, S. E. & Xia, Y. 2009 Thedifferentiation of embryonic stem cells seeded on electro-spun nanofibers into neural lineages. Biomaterials 30,354–362. (doi:10.1016/j.biomaterials.2008.09.046)
22 Lee, J. Y., Bashur, C. A., Goldstein, A. S. & Schmidt,C. E. 2009 Polypyrrole-coated electrospun PLGA nano-fibers for neural tissue applications. Biomaterials 30,4325–4335. (doi:10.1016/j.biomaterials.2009.04.042)
23 Christopherson, G. T., Song, H. & Mao, H. Q. 2009The influence of fiber diameter of electrospun substrateson neural stem cell differentiation and proliferation.Biomaterials 30, 556–564. (doi:10.1016/j.biomaterials.2008.10.004)
24 Jin, G. Z., Kim, M., Shin, U. S. & Kim, H. W. 2011 Neur-ite outgrowth of dorsal root ganglia neurons is enhancedon aligned nanofibrous biopolymer scaffold with carbonnanotube coating. Neurosci. Lett. 501, 10–14. (doi:10.1016/j.neulet.2011.06.023)
25 Carlberg, B., Axell, M. Z., Nannmark, U., Liu, J. &Kuhn, H. G. 2009 Electrospun polyurethane scaffoldsfor proliferation and neuronal differentiation of humanembryonic stem cells. Biomed. Mater. 4, 045004.(doi:10.1088/1748-6041/4/4/045004)
26 Zhao, X., Pan, F., Xu, H., Yaseen, M., Shan, H., Hauser,C. A. E., Zhang, S. & Lu, J. R. 2010 Molecular self-assem-bly and applications of designer peptide amphiphiles.Chem. Soc. Rev. 39, 3480–3498. (doi:10.1039/b915923c)
27 Georges, P. C., Miller, W. J., Meaney, D. F., Sawyer,E. S. & Janmey, P. A. 2006 Matrices with compliancecomparable to that of brain tissue select neuronal overglial growth in mixed cortical cultures. Biophys. J. 90,3012–3018. (doi:10.1529/biophysj.105.073114)
28 Engler, A. J., Sen, S., Sweeney, H. L. & Discher, D. E.2006 Matrix elasticity directs stem cell lineage specifica-tion. Cell 126, 677–689. (doi:10.1016/j.cell.2006.06.044)
29 Leipzig, N. D. & Shoichet, M. S. 2009 The effect of sub-strate stiffness on adult neural stem cell behavior.Biomaterials 30, 6867–6878. (doi:10.1016/j.biomaterials.2009.09.002)
30 Hynes, S. R., Rauch, M. F., Bertram, J. P. & Lavik, E. B.2009 A library of tunable poly(ethylene glycol)/poly(L-lysine) hydrogels to investigate the material cues thatinfluence neural stem cell differentiation. J. Biomed.Mater. Res. A 89A, 499–509. (doi:10.1002/jbm.a.31987)
31 Meek, M. F., Den Dunnen, W. F. A., Schakenraad, J. M. &Robinson, P. H. 1999 Long-term evaluation of functionalnerve recovery after reconstruction with a thin-walled bio-degradable poly(DL-lactide-1-caprolactone) nerve guide,using walking track analysis and electrostimulation tests.Microsurgery 19, 247–253. (doi:10.1002/(SICI)1098-2752(1999)19:5,247::AID-MICR7.3.0.CO;2-E)

Review. HA-based scaffold for CNS regeneration X. Wang et al. 289
on August 31, 2018http://rsfs.royalsocietypublishing.org/Downloaded from
32 Mackinnon, S. E. & Dellon, A. L. 1990 Clinical nervereconstruction with a bioabsorbable polyglycolic acidtube. Plast. Reconstr. Surg. 85, 419–424.
33 Flynn, L., Dalton, P. D. & Shoichet, M. S. 2003 Fibertemplating of poly(2-hydroxyethyl methacrylate) forneural tissue engineering. Biomaterials 24, 4265–4272.(doi:10.1016/s0142-9612(03)00334-x)
34 Stokols, S., Sakamoto, J., Breckon, C., Holt, T., Weiss, J.& Tuszynski, M. H. 2006 Templated agarose scaffoldssupport linear axonal regeneration. Tissue Eng. 12,2777–2787. (doi:10.1089/ten.2006.12.2777)
35 Scott, J. B., Afshari, M., Kotek, R. & Saul, J. M. 2011The promotion of axon extension in vitro using poly-mer-templated fibrin scaffolds. Biomaterials 32, 4830–4839. (doi:10.1016/j.biomaterials.2011.03.037)
36 Wong, D. Y., Leveque, J. C., Brumblay, H., Krebsbach,P. H., Hollister, S. J. & LaMarca, F. 2008 Macro-architectures in spinal cord scaffold implants influenceregeneration. J. Neurotraum. 25, 1027–1037. (doi:10.1089/neu.2007.0473)
37 Stokols, S. & Tuszynski, M. H. 2004 The fabrication andcharacterization of linearly oriented nerve guidance scaf-folds for spinal cord injury. Biomaterials 25, 5839–5846.(doi:10.1016/j.biomaterials.2004.01.041)
38 Stokols, S. & Tuszynski, M. H. 2006 Freeze-dried agarosescaffolds with uniaxial channels stimulate and guide linearaxonal growth following spinal cord injury. Biomaterials27, 443–451. (doi:10.1016/j.biomaterials.2005.06.039)
39 Mukhatyar, V. J., Salmeron-Sanchez, M., Rudra, S.,Mukhopadaya, S., Barker, T. H., Garcia, A. J. &Bellamkonda, R. V. 2011 Role of fibronectin in topogra-phical guidance of neurite extension on electrospunfibers. Biomaterials 32, 3958–3968. (doi:10.1016/j.biomaterials.2011.02.015)
40 Cooper, A., Bhattarai, N. & Zhang, M. Q. 2011 Fabrica-tion and cellular compatibility of aligned chitosan-PCLfibers for nerve tissue regeneration. Carbohydr. Polym.85, 149–156. (doi:10.1016/j.carbpol.2011.02.008)
41 Yang, F., Murugan, R., Wang, S. & Ramakrishna, S.2005 Electrospinning of nano/micro scale poly(L-lacticacid) aligned fibers and their potential in neural tissueengineering. Biomaterials 26, 2603–2610. (doi:10.1016/j.biomaterials.2004.06.051)
42 Xie, J., MacEwan, M. R., Li, X., Sakiyama-Elbert,S. E. & Xia, Y. 2009 Neurite outgrowth on nanofiberscaffolds with different orders, structures, and surfaceproperties. ACS Nano 3, 1151–1159. (doi:10.1021/nn900070z)
43 Cho, Y. I., Choi, J. S., Jeong, S. Y. & Yoo, H. S. 2010Nerve growth factor (NGF)-conjugated electrospunnanostructures with topographical cues for neuronal differ-entiation of mesenchymal stem cells. Acta Biomater. 6,4725–4733. (doi:10.1016/j.actbio.2010.06.019)
44 Platt, C. I., Krekoski, C. A., Ward, R. V., Edwards, D. R.& Gavrilovic, J. 2003 Extracellular matrix and matrixmetalloproteinases in sciatic nerve. J. Neurosci. Res.74, 417–429. (doi:10.1002/jnr.10783)
45 Duan, W. M., Zhao, L. R., Westerman, M., Lovick, D.,Furcht, L. T., McCarthy, J. B. & Low, W. C. 2000Enhancement of nigral graft survival in rat brain withthe systemic administration of synthetic fibronectin pep-tide V. Neuroscience 100, 521–530. (doi:10.1016/S0306-4522(00)00299-2)
46 Zhao, L. R., Spellman, S., Kim, J., Duan, W. M.,McCarthy, J. B. & Low, W. C. 2005 Synthetic fibronectinpeptide exerts neuroprotective effects on transient focalbrain ischemia in rats. Brain Res. 1054, 1–8. (doi:10.1016/j.brainres.2005.04.056)
Interface Focus (2012)
47 King, V. R., Hewazy, D., Alovskaya, A., Phillips, J. B.,Brown, R. A. & Priestley, J. V. 2010 The neuroprotectiveeffects of fibronectin mats and fibronectin peptides fol-lowing spinal cord injury in the rat. Neuroscience 168,523–530. (doi:10.1016/j.neuroscience.2010.03.040)
48 Koh, H. S., Yong, T., Chan, C. K. & Ramakrishna, S.2008 Enhancement of neurite outgrowth using nano-structured scaffolds coupled with laminin. Biomaterials29, 3574–3582. (doi:10.1016/j.biomaterials.2008.05.014)
49 Li, X., Yang, Z., Zhang, A., Wang, T. & Chen, W. 2009Repair of thoracic spinal cord injury by chitosan tubeimplantation in adult rats. Biomaterials 30, 1121–1132. (doi:10.1016/j.biomaterials.2008.10.063)
50 Prabhakaran, M. P., Venugopal, J. R. & Ramakrishna, S.2009 Mesenchymal stem cell differentiation to neuronalcells on electrospun nanofibrous substrates for nervetissue engineering. Biomaterials 30, 4996–5003.(doi:10.1016/j.biomaterials.2009.05.057)
51 Hashemi, S. M., Soudi, S., Shabani, I., Naderi, M. &Soleimani, M. 2011 The promotion of stemness and plur-ipotency following feeder-free culture of embryonic stemcells on collagen-grafted 3-dimensional nanofibrous scaf-fold. Biomaterials 32, 7363–7374. (doi:10.1016/j.biomaterials.2011.06.048)
52 Borkenhagen, M., Clemence, J. F., Sigrist, H. &Aebischer, P. 1998 Three-dimensional extracellularmatrix engineering in the nervous system. J. Biomed.Mater. Res. 40, 392–400. (doi:10.1002/(SICI)1097-4636(19980603)40:3,392::AID-JBM8.3.0.CO;2-C)
53 Cui, F. Z., Tian, W. M., Hou, S. P., Xu, Q. Y. & Lee, I. S.2006 Hyaluronic acid hydrogel immobilized with RGD pep-tides for brain tissue engineering. J. Mater. Sci. Mater.Med. 17, 1393–1401. (doi:10.1007/s10856-006-0615-7)
54 Suzuki, M., Itoh, S., Yamaguchi, I., Takakuda, K.,Kobayashi, H., Shinomiya, K. & Tanaka, J. 2003Tendon chitosan tubes covalently coupled with syn-thesized laminin peptides facilitate nerve regenerationin vivo. J. Neurosci. Res. 72, 646–659. (doi:10.1002/jnr.10589)
55 Schense, J. C., Bloch, J., Aebischer, P. & Hubbell, J. A.2000 Enzymatic incorporation of bioactive peptides intofibrin matrices enhances neurite extension. Nat. Biotech-nol. 18, 415–419. (doi:10.1038/74473)
56 Krewson, C. E., Klarman, M. L. & Saltzman, W. M. 1995Distribution of nerve growth-factor following direct deliv-ery to brain interstitium. Brain Res. 680, 196–206.(doi:10.1016/0006-8993(95)00261-N)
57 Yu, L. M. Y., Wosnick, J. H. & Shoichet, M. S. 2008 Min-iaturized system of neurotrophin patterning for guidedregeneration. J. Neurosci. Methods 171, 253–263.(doi:10.1016/j.jneumeth.2008.03.023)
58 Mo, L. H., Yang, Z. Y., Zhang, A. F. & Li, X. G. 2010The repair of the injured adult rat hippocampus withNT-3-chitosan carriers. Biomaterials 31, 2184–2192.(doi:10.1016/j.biomaterials.2009.11.078)
59 Jain, A., Kim, Y. T., McKeon, R. J. & Bellamkonda,R. V. 2006 In situ gelling hydrogels for conformal repairof spinal cord defects, and local delivery of BDNF afterspinal cord injury. Biomaterials 27, 497–504. (doi:10.1016/j.biomaterials.2005.07.008)
60 Wang, Y. C., Wu, Y. T., Huang, H. Y., Lin, H. I., Lo,L. W., Tzeng, S. F. & Yang, C. S. 2008 Sustained intrasp-inal delivery of neurotrophic factor encapsulated inbiodegradable nanoparticles following contusive spinalcord injury. Biomaterials 29, 4546–4553. (doi:10.1016/j.biomaterials.2008.07.050)
61 Burdick, J. A., Ward, M., Liang, E., Young, M. J. &Langer, R. 2006 Stimulation of neurite outgrowth by

290 Review. HA-based scaffold for CNS regeneration X. Wang et al.
on August 31, 2018http://rsfs.royalsocietypublishing.org/Downloaded from
neurotrophins delivered from degradable hydrogels.Biomaterials 27, 452–459. (doi:10.1016/j.biomaterials.2005.06.034)
62 Iannotti, C., Li, H. Y., Yan, P., Lu, X. B., Wirthlin, L. &Xu, X. M. 2003 Glial cell line-derived neurotrophicfactor-enriched bridging transplants promote propriosp-inal axonal regeneration and enhance myelination afterspinal cord injury. Exp. Neurol. 183, 379–393. (doi:10.1016/S0014-4886(03)00188-2)
63 Piantino, J., Burdick, J. A., Goldberg, D., Langer, R. &Benowitz, L. I. 2006 An injectable, biodegradable hydrogelfor trophic factor delivery enhances axonal rewiring andimproves performance after spinal cord injury. Exp. Neurol.201, 359–367. (doi:10.1016/j.expneurol.2006.04.020)
64 Li, X., Yang, Z. & Zhang, A. 2009 The effect of neurotro-phin-3/chitosan carriers on the proliferation anddifferentiation of neural stem cells. Biomaterials 30,4978–4985. (doi:10.1016/j.biomaterials.2009.05.047)
65 Sakiyama-Elbert, S. E. & Hubbell, J. A. 2000 Develop-ment of fibrin derivatives for controlled release ofheparin-binding growth factors. J. Control. Release 65,389–402. (doi:10.1016/S0168-3659(99)00221-7)
66 Willerth, S. M., Johnson, P. J., Maxwell, D. J., Parsons,S. R., Doukas, M. E. & Sakiyama-Elbert, S. E. 2007Rationally designed peptides for controlled release ofnerve growth factor from fibrin matrices. J. Biomed.Mater. Res. A 80A, 13–23. (doi:10.1002/jbm.a.30844)
67 Johnson, P. J., Tatara, A., Shiu, A. & Sakiyama-Elbert,S. E. 2010 Controlled release of neurotrophin-3 andplatelet-derived growth factor from fibrin scaffoldscontaining neural progenitor cells enhances survivaland differentiation into neurons in a subacute modelof SCI. Cell Transplant. 19, 89–101. (doi:10.3727/096368909X477273)
68 Taylor, S. J., McDonald, J. W. & Sakiyama-Elbert, S. E.2004 Controlled release of neurotrophin-3 from fibrin gelsfor spinal cord injury. J. Control. Release 98, 281–294.(doi:10.1016/j.jconrel.2004.05.003)
69 Rahman, N., Purpura, K. A., Wylie, R. G., Zandstra,P. W. & Shoichet, M. S. 2010 The use of vascular endo-thelial growth factor functionalized agarose to guidepluripotent stem cell aggregates toward blood progenitorcells. Biomaterials 31, 8262–8270. (doi:10.1016/j.bioma-terials.2010.07.040)
70 Shen, Y. H., Shoichet, M. S. & Radisic, M. 2008 Vascularendothelial growth factor immobilized in collagen scaffoldpromotes penetration and proliferation of endothelialcells. Acta Biomater. 4, 477–489. (doi:10.1016/j.actbio.2007.12.011)
71 Chvatal, S. A., Kim, Y.-T., Bratt-Leal, A. M., Lee, H. &Bellamkonda, R. V. 2008 Spatial distribution and acuteanti-inflammatory effects of methylprednisolone aftersustained local delivery to the contused spinal cord. Bio-materials 29, 1967–1975. (doi:10.1016/j.biomaterials.2008.01.002)
72 Schnell, E.,Klinkhammer,K., Balzer, S., Brook,G.,Klee,D.,Dalton, P. & Mey, J. 2007 Guidance of glial cell migrationand axonal growth on electrospun nanofibers of poly-1-caprolactone and a collagen/poly-1-caprolactone blend.Biomaterials 28, 3012–3025. (doi:10.1016/j.bio materials.2007.03.009)
73 Xu, T., Molnar, P., Gregory, C., Das, M., Boland, T. &Hickman, J. J. 2009 Electrophysiological characterizationof embryonic hippocampal neurons cultured in a 3D col-lagen hydrogel. Biomaterials 30, 4377–4383. (doi:10.1016/j.biomaterials.2009.04.047)
74 Novikova, L. N., Mosahebi, A., Wiberg, M., Terenghi, G.,Kellerth, J. O. & Novikov, L. N. 2006 Alginate hydrogel
Interface Focus (2012)
and matrigel as potential cell carriers for neurotransplan-tation. J. Biomed. Mater. Res. A 77A, 242–252. (doi:10.1002/jbm.a.30603)
75 Shen, Y. X., Qian, Y. Q., Zhang, H. X., Zuo, B. Q., Lu, Z. F.,Fan, Z. H., Zhang, P., Zhang, F. & Zhou, C. L. 2010 Gui-dance of olfactory ensheathing cell growth and migrationon electrospun silk fibroin scaffolds. Cell Transplant. 19,147–157. (doi:10.3727/096368910x492616)
76 Frampton, J. P., Hynd, M. R., Shuler, M. L. & Shain, W.2011 Fabrication and optimization of alginate hydrogelconstructs for use in 3D neural cell culture. Biomed.Mater. 6, 015002. (doi:10.1088/1748-6041/6/1/015002)
77 Freudenberg, U. et al. 2009 A star-PEG-heparin hydrogelplatform to aid cell replacement therapies for neurode-generative diseases. Biomaterials 30, 5049–5060.(doi:10.1016/j.biomaterials.2009.06.002)
78 Lu, D., Mahmood, A., Qu, C., Hong, X., Kaplan, D. &Chopp, M. 2007 Collagen scaffolds populated withhuman marrow stromal cells reduce lesion volume andimprove functional outcome after traumatic braininjury. Neurosurgery 61, 596–602. (doi:10.1227/01.neu.000028004/.85651.4f)
79 Barralet, J. E., Wang, L., Lawson, M., Triffitt, J. T.,Cooper, P. R. & Shelton, R. M. 2005 Comparison ofbone marrow cell growth on 2D and 3D alginate hydro-gels. J. Mater. Sci. Mater. Med. 16, 515–519. (doi:10.1007/s10856-005-0526-z)
80 Hejcl, A. et al. 2010 HPMA–RGD hydrogels seeded withmesenchymal stem cells improve functional outcome inchronic spinal cord injury. Stem Cells Dev. 19, 1535–1546. (doi:10.1089/scd.2009.0378)
81 Kadoya, K. 2009 Combined intrinsic and extrinsic neur-onal mechanisms facilitate bridging axonal regenerationone year after spinal cord injury. Neuron 64, 165–172.(doi:10.1016/j.neuron.2009.09.016)
82 Guo, J., Su, H., Zeng, Y., Liang, Y.-X., Wong, W. M.,Ellis-Behnke, R. G., So, K.-F. & Wu, W. 2007 Reknittingthe injured spinal cord by self-assembling peptide nanofi-ber scaffold. Nanomed. Nanotechnol. Biol. Med. 3, 311–321. (doi:10.1016/j.nano.2007.09.003)
83 Xiong, Y., Qu, C. S., Mahmood, A., Liu, Z. W., Ning,R. Z., Li, Y., Kaplan, D. L., Schallert, T. & Chopp, M.2009 Delayed transplantation of human marrow stromalcell-seeded scaffolds increases transcallosal neural fiberlength, angiogenesis, and hippocampal neuronal survivaland improves functional outcome after traumatic braininjury in rats. Brain Res. 1263, 183–191. (doi:10.1016/j.brainres.2009.01.032)
84 Yu, H. W., Cao, B., Feng, M. Y., Zhou, Q., Sun, X. D.,Wu, S. L., Jin, S. Z., Liu, H. W. & Jin, L. H. 2010 Com-binated transplantation of neural stem cells and collagentype I promote functional recovery after cerebral ischemiain rats. Anat. Rec. 293, 911–917. (doi:10.1002/ar.20941)
85 Mahmood, A., Qu, C. S., Ning, R. Z., Wu, H. T., Gous-sev, A., Xiong, Y., Irtenkauf, S., Li, Y. & Chopp, M.2011 Treatment of TBI with collagen scaffolds andhuman marrow stromal cells increases the expression oftissue plasminogen activator. J. Neurotraum. 28, 1199–1207. (doi:10.1089/neu.2010.1694)
86 Ford, M. C., Bertram, J. P., Hynes, S. R., Michaud, M.,Li, Q., Young, M., Segal, S. S., Madri, J. A. & Lavik,E. B. 2006 A macroporous hydrogel for the coculture ofneural progenitor and endothelial cells to form functionalvascular networks in vivo. Proc. Natl Acad. Sci. USA103, 2512–2517. (doi:10.1073/pnas.0506020102)
87 Rauch, M. F., Hynes, S. R., Bertram, J., Redmond, A.,Robinson, R., Williams, C., Xu, H., Madri, J. A. &Lavik, E. B. 2009 Engineering angiogenesis following

Review. HA-based scaffold for CNS regeneration X. Wang et al. 291
on August 31, 2018http://rsfs.royalsocietypublishing.org/Downloaded from
spinal cord injury: a coculture of neural progenitor andendothelial cells in a degradable polymer implant leadsto an increase in vessel density and formation ofthe blood–spinal cord barrier. Eur. J. Neurosci. 29,132–145. (doi:10.1111/j.1460-9568.2008.06567.x)
88 Mori, M., Yamaguchi, M., Sumitomo, S. & Takai, Y. 2004Hyaluronan-based biomaterials in tissue engineering. ActaHistochem. Cytochem. 37, 1–5. (doi:10.1267/ahc.37.1)
89 Laurent, T. C., Laurent, U. B. G. & Fraser, J. R. E. 1996The structure and function of hyaluronan: a overview.Immunol. Cell Biol. 74, A1–A7. (doi:10.1038/icb.1996.32)
90 Costa, C., Tortosa, R., Domenech, A., Vidal, E., Pumar-ola, M. & Bassols, A. 2007 Mapping of aggrecan,hyaluronic acid, heparan sulphate proteoglycans andaquaporin 4 in the central nervous system of the mouse.J. Chem. Neuroanatomy 33, 111–123. (doi:10.1016/j.jchemneu.2007.01.006)
91 Manuskiatti, W. & Maibach, H. I. 1996 Hyaluronic acidand skin: wound healing and aging. Int. J. Dermatol.35, 539–544. (doi:10.1111/j.1365-4362.1996.tb03650.x)
92 Chen, W. Y. J. & Abatangelo, G. 1999 Functions of hya-luronan in wound repair. Wound Repair Regen. 7, 79–89.(doi:10.1046/j.1524-475X.1999.00079.x)
93 Tian, W. M., Hou, S. P., Ma, J., Zhang, C. L., Xu, Q. Y.,Lee, I. S., Li, H. D., Spector, M. & Cui, F. Z. 2005 Hya-luronic acid–poly-D-lysine-based three-dimensionalhydrogel for traumatic brain injury. Tissue Eng. 11,513–525. (doi:10.1089/ten.2005.11.513)
94 Hou, S., Tian, W., Xu, Q., Cui, F., Zhang, J., Lu, Q. &Zhao, C. 2006 The enhancement of cell adherence andinducement of neurite outgrowth of dorsal root gangliaco-cultured with hyaluronic acid hydrogels modifiedwith Nogo-66 receptor antagonist in vitro. Neuroscience137, 519–529. (doi:10.1016/j.neuroscience.2005.09.029)
95 Ma, J., Tian, W.-M., Hou, S.-P., Xu, Q.-Y., Spector, M.& Cui, F.-Z. 2007 An experimental test of stroke recoveryby implanting a hyaluronic acid hydrogel carrying aNogo receptor antibody in a rat model. Biomed. Mater.2, 233–240. (doi:10.1088/1748-6041/2/4/005)
96 Wei, Y. T., He, Y., Xu, C. L., Wang, Y., Liu, B. F., Wang,X. M., Sun, X. D., Cui, F. Z. & Xu, Q. Y. 2010 Hyaluronicacid hydrogel modified with nogo-66 receptor antibodyandpoly-(L)-lysine to promote axon regrowth after spinal cordinjury. J. Biomed. Mater. Res. B. 95B, 110–117. (doi:10.1002/jbm.b.31689)
97 Ozgenel, G. Y. 2003 Effects of hyaluronic acid on periph-eral nerve scarring and regeneration in rats. Microsurgery23, 575–581. (doi:10.1002/micr.10209)
98 Lin, C.-M. et al. 2009 Hyaluronic acid inhibits the glialscar formation after brain damage with tissue loss inrats. Surg. Neurol. 72(Suppl. 2), S50–S54. (doi:10.1016/j.wneu.2009.09.004)
99 Khaing, Z. Z., Milman, B. D., Vanscoy, J. E., Seidlits, S.K., Grill, R. J. & Schmidt, C. E. 2011 High molecularweight hyaluronic acid limits astrocyte activation andscar formation after spinal cord injury. J. Neural Eng.8, 046033. (doi:10.1088/1741-2560/8/4/046033)
100 Campo, G. M., Avenoso, A., Campo, S., D’Ascola, A.,Nastasi, G. & Calatroni, A. 2010 Molecular size hyaluro-nan differently modulates toll-like receptor-4 in LPS-induced inflammation in mouse chondrocytes. Biochimie92, 204–215. (doi:10.1016/j.biochi.2009.10.006)
101 Seidlits, S. K., Khaing, Z. Z., Petersen, R. R., Nickels,J. D., Vanscoy, J. E., Shear, J. B. & Schmidt, C. E.2010 The effects of hyaluronic acid hydrogels with
Interface Focus (2012)
tunable mechanical properties on neural progenitor celldifferentiation. Biomaterials 31, 3930–3940. (doi:10.1016/j.biomaterials.2010.01.125)
102 Wang,T.-W.&Spector,M. 2009Developmentof hyaluronicacid-based scaffolds for brain tissue engineering. Acta Bio-mater. 5, 2371–2384. (doi:10.1016/j.actbio.2009.03.033)
103 Brannvall, K., Bergman, K., Wallenquist, U., Svahn, S.,Bowden, T., Hilborn, J. & Forsberg-Nilsson, K. 2007Enhanced neuronal differentiation in a three-dimensionalcollagen-hyaluronan matrix. J. Neurosci. Res. 85,2138–2146. (doi:10.1002/jnr.21358)
104 Hou, S. P., Xu, Q. Y., Tian, W. M., Cui, F. Z., Cai, Q.,Ma, J. & Lee, I. S. 2005 The repair of brain lesion byimplantation of hyaluronic acid hydrogels modified withlaminin. J. Neurosci. Methods 148, 60–70. (doi:10.1016/j.jneumeth.2005.04.016)
105 Wei, Y. T., Tian, W. M., Yu, X., Cui, F. Z., Hou, S. P.,Xu, Q. Y. & Lee, I.-S. 2007 Hyaluronic acid hydrogelswith IKVAV peptides for tissue repair and axonalregeneration in an injured rat brain. Biomed. Mater. 2,S142–S146. (doi:10.1088/1748-6041/2/3/s11)
106 Yavin, E. & Yavin, Z. 1974 Attachment and culture ofdissociated cells from rat embryo cerebral hemisphereson polylysine-coated surface. J. Cell Biol. 62, 540–546.(doi:10.1083/jcb.62.2.540)
107 Fournier, A. E., GrandPre, T. & Strittmatter, S. M. 2001Identification of a receptor mediating Nogo-66 inhibitionof axonal regeneration. Nature 409, 341–346. (doi:10.1038/35053072)
108 Wang, K. C., Koprivica, V., Kim, J. A., Sivasankaran,R., Guo, Y., Neve, R. L. & He, Z. G. 2002 Oligodendro-cyte-myelin glycoprotein is a Nogo receptor ligand thatinhibits neurite outgrowth. Nature 417, 941–944.(doi:10.1038/nature00867)
109 Domeniconi, M. et al. 2002 Myelin-associated glyco-protein interacts with the Nogo66 receptor to inhibitneurite outgrowth. Neuron 35, 283–290. (doi:10.1016/S0896-6273(02)00770-5)
110 Tian, W. M., Zhang, C. L., Hou, S. P., Yu, X., Cui, F. Z.,Xu, Q. Y., Sheng, S. L., Cui, H. & Li, H. D. 2005 Hyaluro-nic acid hydrogel as Nogo-66 receptor antibody deliverysystem for the repairing of injured rat brain: in vitro.J. Control. Release 102, 13–22. (doi:10.1016/j.jconrel.2004.09.025)
111 Park, J., Lim, E., Back, S., Na, H., Park, Y. & Sun, K.2010 Nerve regeneration following spinal cord injuryusing matrix metalloproteinase-sensitive, hyaluronicacid-based biomimetic hydrogel scaffold containingbrain-derived neurotrophic factor. J. Biomed. Mater.Res. A 93A, 1091–1099. (doi:10.1002/jbm.a.32519)
112 Wang, Y., Wei, Y. T., Zu, Z. H., Ju, R. K., Guo, M. Y.,Wang, X. M., Xu, Q. Y. & Cui, F. Z. 2011 Combination ofhyaluronic acid hydrogel scaffold and PLGA microspheresfor supporting survival of neural stem cells. Pharm. Res.28, 1406–1414. (doi:10.1007/s11095-011-0452-3)
113 Wei, Y. T., Sun, X. D., Xia, X., Cui, F. Z., He, Y., Liu,B. F. & Xu, Q. Y. 2009 Hyaluronic acid hydrogel modi-fied with Nogo-66 receptor antibody and poly(L-lysine)enhancement of adherence and survival of primaryhippocampal neurons. J. Bioact. Compat. Polym. 24,205–219. (doi:10.1177/0883911509102266)
114 Pan, L. J., Ren, Y. J., Cui, F. Z. & Xu, Q. Y. 2009 Via-bility and differentiation of neural precursors onhyaluronic acid hydrogel scaffold. J. Neurosci. Res. 87,3207–3220. (doi:10.1002/jnr.22142)