human fetal auditory stem cells can be expanded in vitro and differentiate into functional auditory...
TRANSCRIPT

TISSUE-SPECIFIC STEM CELLS
Human Fetal Auditory Stem Cells Can Be Expanded In Vitro
and Differentiate Into Functional Auditory Neurons
and Hair Cell-Like Cells
WEI CHEN,a,b STUART L. JOHNSON,b WALTER MARCOTTI,b PETER W. ANDREWS,a,b HARRY D. MOORE,a,b
MARCELO N. RIVOLTAa,b
aCentre for Stem Cell Biology and bDepartment of Biomedical Sciences, University of Sheffield,
Sheffield, United Kingdom
Key Words. Fetal stem cells • Tissue-specific stem cells • Differentiation • Neuron • Hair cells • Ear • Deafness
ABSTRACT
In the quest to develop the tools necessary for a cell-basedtherapy for deafness, a critical step is to identify a suitablestem cell population. Moreover, the lack of a self-renovatingmodel system for the study of cell fate determination in the
human cochlea has impaired our understanding of the mo-lecular events involved in normal human auditory develop-
ment. We describe here the identification and isolation of apopulation of SOX21OCT41 human auditory stem cellsfrom 9-week-old to 11-week-old fetal cochleae (hFASCs).
These cells underwent long-term expansion in vitro andretained their capacity to differentiate into sensory haircells and neurons, whose functional and electrophysiologicalproperties closely resembled their in vivo counterparts dur-
ing development. hFASCs, and the differentiating protocolsdefined here, could be used to study developing human
cochlear neurons and hair cells, as models for drug screeningand toxicity and may facilitate the development of cell-basedtherapies for deafness. STEM CELLS 2009;27:1196–1204
Disclosure of potential conflicts of interest is found at the end of this article.
INTRODUCTION
Deafness is a very common affliction, suffered by more than250 million people world wide and with vast social and eco-nomic implications. Almost 90% of cases are sensorineural innature, involving the loss of hair cells and, to a variabledegree, their associated neurons. The transplantation of stemcells with the potential to produce functional differentiatedcell types might be a promising therapeutic strategy for thetreatment of hearing impairment [1, 2]. However, no actualhuman candidate cell type is yet available to facilitate the de-velopment of this technology. Stem cells have been isolatedfrom adult mouse inner ears [3], and efforts have been madeto induce differentiation of auditory progenitors from mouseembryonic stem cells (ESCs) [4, 5]. Transplantation studieshave been done with undifferentiated and neuralized mouseESCs as well as neural stem cells, but these cell types areless adequate because they are not of otic origin and may notfully resemble the tissue they aim to repair [6–9]. Althoughinner ear stem cells can be isolated from the mouse adult ves-tibular organs [3], attempts to obtain equivalent cells from theadult mouse cochlea have failed, as their population declines
sharply after birth and is almost gone by the third week ofage [10]. An ideal strategy would then be to aim for early,fetal stages. We have previously shown that the human fetalcochlea harbors auditory stem cells [11] and could be a poten-tial source for their isolation, as cells taken before the onsetof terminal differentiation should retain most of their undiffer-entiated features if cultured under appropriate conditions.Aiming to isolate them at early stages of development wouldbring the additional benefit of capturing them at an appropri-ate time, avoiding the potential risk of undifferentiating amore committed cell population from a later stage of develop-ment. Isolated populations of fetal auditory stem cells shouldtherefore be valuable models of human auditory cell develop-ment. With this aim, we have attempted to isolate auditorystem cells from the human fetal cochlea. By using definedculture conditions, we have expanded in vitro cells thatexpress several stem cell/progenitor markers and establishedthem as cell lines. These cell lines can be propagated for pro-longed periods of time, making them amenable to experimen-tation and have the ability to differentiate into sensory neu-rons and hair cell-like cells. Moreover, the differentiatedphenotypes obtained displayed functional properties resem-bling the equivalent developing cell types in vivo.
Author contributions: W.C.: collection and/or assembly of data, data analysis, and interpretation; S.L.J. and W.M.: collection, analysis,and interpretation of electrophysiology data, manuscript writing; P.W.A. and H.D.M.: provision of study material and administrativesupport; M.N.R: conception and design, financial support, collection and/or assembly of data, its analysis and interpretation, manuscriptwriting, final approval of manuscript.
Correspondence: Dr. Marcelo N. Rivolta, Centre for Stem Cell Biology and Department of Biomedical Sciences, University ofSheffield, Alfred Denny Building, Western Bank, Sheffield S10 2TN, UK. Telephone: 44(0)-114-2222385; Fax: (0)-114-2765413;e-mail: [email protected] Received December 15, 2008; accepted for publication March 3, 2009; first published online inSTEM CELLS EXPRESS March 23, 2009. VC AlphaMed Press 1066-5099/2009/$30.00/0 doi: 10.1002/stem.62
STEM CELLS 2009;27:1196–1204 www.StemCells.com

MATERIALS AND METHODS
Establishment of Cultures
Collection of human fetal tissue was done in agreement withthe Polkinhorne guidelines and following ethical approval andinformed consent. Cochleae were removed from 9-week-oldto 11-week-old human fetuses in sterile Hanks solution. Thecartilaginous capsule was removed and the cochlear epithelia(primarily the greater and lesser epithelial ridges, includingKollicker’s organ) was carefully dissected free from surround-ing tissue and explanted as small patches on tissue cultureplastic dishes (In Vitro Fertilization dishes; Corning Life Sci-ences, Acton, MA, http://www.corning.com/lifesciences), in100 ll of high glucose Dulbecco’s modified Eagle’s medium(DMEM; Invitrogen, Carlsbad, CA, http://www.invitrogen.com)-10% FBS. The IVF dish allows the use of a small vol-ume of culture media, while still making the tissue accessiblefor further dissociation. Given that 3–5 hours have normallyelapsed since pregnancy termination until the tissue was avail-able for dissection, explants were incubated overnight at 37�Cas a precautionary measure to increase cell viability and todiminish the stress should further dissociation be appliedimmediately after dissection. Nevertheless, this step couldlikely be omitted if needed to adapt the protocol to GoodManufacturing Practice (GMP) standards. Epithelial patcheswere then lifted from the dishes (using needles and trying toremove only the epithelial tissue and discarding as muchunderlying mesenchyme as possible) and dissociated by incu-bation with 0.125% trypsin in Hanks solution (supplementedwith 5 mM HEPES) at room temperature for 10–15 minutes.Separation was facilitated by gentle mechanical dissociationusing initially, a 1,000-ll pipette tip and then, a 200-ll pip-ette tip. Single cell suspensions obtained from both cochleaefrom a single donor were pooled and plated on gelatin-coateddishes (Nunc, Rochester, NY, http://www.nuncbrand.com;Thermo Fisher) and selected in Otic Stem Cell Full Medium(OSCFM). This medium consists of a 1:1 mixture of high glu-cose DMEM (4.5 g/l D-Glucose; Invitrogen) plus F12 (Invitro-gen) nutrient solution, N2 and B27 supplements, 20 ng/mlrecombinant human basic fibroblast growth factor (bFGF),50 ng/ml recombinant human insulin-like growth factor 1
(IGF-1), and 20 ng/ml recombinant human epidermal growthfactor (EGF). Concentrations of bFGF, EGF, and IGF-1 in thesingle growth factor experiments were the same as used inOSCFM. Growth factors were from R&D Systems Inc. (Min-neapolis, MN, http://www.rndsystems.com). Trypsin was onlyused for the initial dissociation. Subsequent passages weredone using a Hank’s based, enzyme-free cell dissociation so-lution (Chemicon, Temecula, CA, http://www.chemicon.com).For all cell counting data, statistical comparisons of meanswere made using one-way or two-way ANOVA followed byBonferroni’s post-test.
RT-PCR
Total RNA was extracted using Trizol (Invitrogen). cDNAsynthesis was carried out by oligo (dT) primed reverse tran-scription using M-MLV RT (Stratagene, La Jolla, CA, http://www.stratagene.com). Primers and conditions used aredetailed in Table 1.
Immunolabeling
Cells were fixed in cold 4% paraformaldehide in PBS for10 minutes, permeabilized and blocked in 0.1% Triton X-100and 5% normal goat serum in PBS for 20 minutes at RT, andthen incubated with the primary antibody in the same buffer.For cytokeratin staining, cells were fixed in a 1:1 mixture ofice-cold acetone–methanol. Antibodies used against SOX2,NESTIN, p27Kip1, and ATOH1 were from Abcam (Cam-bridge, U.K., http://www.abcam.com); OCT4 and glial fibril-lary acidic protein (GFAP) from Santa Cruz BiotechnologyInc. (Santa Cruz, CA, http://www.scbt.com); NF200 and Pan-cytokeratin from Sigma (St. Louis, MO, http://www.sigmaaldrich.com); b-tubulin III (TUJ1) from Covance (Princeton,NJ, http://www.covance.com); and BRN3c from Abnova(Taipei City, Taiwan, http://www.abnova.com). Alexa Fluor568-conjugated phalloidin was from Molecular Probes Inc.(Eugene, OR, http://probes.invitrogen.com). The MYOSINVIIA antibody was a kind gift from C. Petit, Pasteur Institute.Specific labeling was visualized with either a secondary anti-mouse, anti-goat, or anti-rabbit antibody conjugated to FITCor Rhodamine Red (Jackson ImmunoResearch Labs, WestGrove, PA, http://www.jacksonimmuno.com). Controls were
Table 1. Oligonucleotide primers for RT-PCR
Gene name Forward Reverse
Positiona,
amplicon length
Annealing
T�
GATA3 50-GTACAGCTCCGGACTCTTCCC-30 50-CTGCTCTCCTGGCTGCAGACA-30 803, 260 bp 55�CSOX2 50-ATGCACCGCTACGACCGTGA-30 50-CTTTTGCACCCCTCCCATTT-30 1016, 437 bp 55�CABCG2 50-GTTTATCCGTGGTGTGTCTGG-30 50-CTGAGCTATAGAGGCCTGGG-30 1036, 652 bp 52�CPOU5F1(Oct4) 50-CTTGCTGCAGAAGTGGGTGGA
GGAA-3050-CTGCAGTGTGGGTTTCGGGCA-30 660, 169 bp 55�C
NANOG 50-TGCCTCACACGGAGACTGTC-30 50-TGCTATTCTTCGGCCAGTTG-30 350, 354 bp 55�CZFP42 (Rex1) 50-TGAAAGCCCACATCCTAACG-30 50-CAAGCTATCCTCCTGCTTTGG-30 1283, 554 bp 55�CNEUROG1 50-CCGACGACACCAAGCTCA-30 50-GGAATGAAACAGGGCGTT-30 627, 337 bp 52�CPOU4F1 (Brn3A) 50-GGCCCACCTCAAGATCCCGG-30 50-AGTTTCTCGGCGATGGCGGC-30 1113, 297 bp 57�CPVALB 50-GATGACAGACTTGCTGAACGC-30 50-CTTAGCTTTCAGCCACCAGAG-30 56, 329 bp 52�CATOH1(Hath1) 50-CTCAGCCCCAGCTTCTGC-30 50-AAACAACGACCATCGCAGAG-30 1190, 359 bp 53�CPOU4F3 (Brn3C) 50-TGCAAGAACCCAAATTCTCC-30 50-GAGCTCTGGCTTGCTGTTCT-30 47, 758 bp 50�CMYO7A 50-CACATCTTTGCCATTGCTGAC-30 50-AGAAGAGAACCTCACAGGCAT-30 669, 649 bp 52�CCACNAD1D (Cav1.3) 50-CGAACGGAGGCATCACCAACTT-30 50-ACGGCGGCCCTACATCTTCTG-30 1137, 601 bp 65�CGJB6 (Cx30) 50-GGGCCCTCCAGCTGATCTTCG
TCTCC-3050-TTACTCTCCTTTAGGGCATGATTGGGGTGATTTTTT-30
783, 490 bp 60�C
HPRT 50- AATTATGGACAGGACTGAACGTC-30 50-CGTGGGGTCCTTTTCACCAGCAAG-30
290, 387 bp 52�C
aPosition is the location of the 50 base of the forward primer in the annotated sequence.
Chen, Johnson, Marcotti et al. 1197
www.StemCells.com

performed by replacing the primary antibody with unspecificmouse or rabbit IgGs.
Differentiation Experiments
To define conditions that would support neuronal differentia-tion, cells were split using 0.125% trypsin in Hanks solution.Following dissociation, trypsin was inactivated by using equalvolumes of 0.5 mg/ml Soybean Trypsin Inhibitor (Invitrogen).This cell dissociation treatment appears to be critical in induc-ing neuronal differentiation. Cells were plated on gelatin-coated dishes at a density of 3–4,000 cells per centimetersquared and cultured in DFNB medium (high glucose DMEMplus F12 nutrient solution, N2 and B27) alone or supple-mented with either recombinant human bFGF (20 ng/ml),IGF-1 (50 ng/ml), Shh–C24II (500 ng/ml), bFGF plus IGF-1,or bFGF plus Shh-C24II. The C24II mutation of human Shhenhances its potency. Bipolar cells were counted on a dailybasis. Once the initial conditions were selected (bFGF plusShh-C24II), cultures were supplemented at the third day with10 ng/ml of brain-derived neurotrophic factor (BDNF) andneurotrophin 3 (NT3) and Shh-C24II removed at the fifth day.For hair cell differentiation, cells were disaggregated using anenzyme-free cell dissociation solution (Chemicon), plated at adensity of 4,000 cells per centimeter squared and cultured inDFNB medium supplemented with 10�6 M All-trans RetinoicAcid (Sigma) and 20 ng/ml EGF.
Electrophysiology
The whole-cell patch clamp technique was used to measuremembrane currents using an Optopatch (Cairn Research Ltd,U.K., http://www.cairnweb.com) or Multiclamp 700B (Molec-ular Devices Corp., Union City, CA, http://www.molecularde-vices.com) amplifiers. Recordings were performed at roomtemperature (20–25�C) from human fetal auditory stem cells(hFASCs) between 1 and 17 days (D1–D17), in which theinduction of differentiation corresponds to D0. Although re-cording at room temperatures in mammalian cells can slowthe current kinetics, most of the voltage-clamp measurementsperformed from acutely isolated mouse or rat cochleae [12–14] and used as a reference for our findings, were alsoobtained under the same conditions. The extracellular solutioncontained (mM): 135 NaCl, 5.8 KCl, 1.3 CaCl2, 0.9 MgCl2,0.7 NaH2PO4, 5.6 D-glucose, 10 Hepes-NaOH, 2 sodium pyru-vate. Amino acids and vitamins for Eagle’s minimal essentialmedium (MEM) were added from concentrates (Fisher Scien-tific International, Hampton, NH, http://www.fisherscientific.com). The pH was adjusted to 7.5. Stem cells were viewedusing an upright microscope equipped with Nomarski DICoptics (Leica, Germany, http://www.leica-camera. com/home/)and continuously superfused with extracellular solution. Patchelectrodes were pulled from soda glass capillaries (HarvardApparatus Ltd, U.K.) and electrodes had resistances in theextracellular solution of 2–3 MX. The shank of the electrodewas coated with surf wax to minimize the fast electrodecapacitative transients. The pipette solution contained (mM):131 KCl, 3 MgCl2, 1 EGTA-KOH, 5 Na2ATP, 5 Hepes-KOH,10 sodium phosphocreatine (pH 7.3).
Voltage protocol application and data acquisition wereperformed using pClamp software and a Digidata 1320A (Mo-lecular Devices) or Signal software and a Power 1401 acquisi-tion interface (CED, U.K.). Recordings were filtered at 2.5 or10 kHz (8-pole Bessel), sampled at 5 or 50 kHz and storedon computer for off-line analysis using Clampfit, Signal, andOrigin (OriginLab, USA) software. All recordings were cor-rected off-line for linear leakage, calculated between �84 mVand �74 mV (undifferentiated cells: 17.3 � 2.3 nS, n ¼ 27;hair cell-like cells: 1.7 � 0.4 nS, n ¼ 20, p < .05; neuronal-
like cells: 1.5 � 0.2 nS, n ¼ 30), and residual capacitativetransients, unless otherwise stated. Membrane potentials undervoltage clamp were corrected for the voltage drop across theresidual series resistance (Rs) at steady-state current level andfor a liquid junction potential, measured between pipette andbath solutions, of �4 mV. Membrane capacitance (Cm) was54 � 3 pF (n ¼ 77, range 8–105 pF) and series resistanceafter electronic compensation of up to 60% (Rs) was 6.5 �0.3 MX (range 1.9–13.8 MX, n ¼ 77), resulting in voltage-clamp time constants of 343 � 22 ls (n ¼ 77).
Membrane currents were elicited by applying voltagesteps (10 mV nominal values) from the holding potential of�84 mV (for IK, ICa, and INa) or �64 mV (for IK1). All actualtest potentials reached are indicated next to the traces. Cur-rent-voltage (I–V) curves in Figures 3G and 4E were fittedusing the following equation:
I ¼ gmax V � Vrevð Þ= 1þ exp Vhalf � Vð Þ=S½ �gf (1)
where I is the current, V is the membrane potential of thevoltage step, Vrev is the reversal potential extrapolated fromthe fit, gmax is the maximum chord conductance, Vhalf is thepotential of half-maximal activation, and S describes the volt-age sensitivity of activation.
Statistical comparisons of means were made using eitherthe unpaired student’s two-tailed t-test for two data sets or forcomparisons of multiple data sets, using analysis of variance(one-way ANOVA followed by the Tukey post-test). For allstatistical tests, p < .05 was used as the criterion for statisticalsignificance. Mean values are quoted � s.e.m. in text and figures.
RESULTS
Isolation of hFASCs
Our initial aim was to establish the culture conditions thatwill support the expansion of stem cells from the fetal coch-leae and allow their isolation. For this, whole cochleae from9-week-old to 11-week-old human fetuses (stage of ear devel-opment equivalent to E11-13.5 in the mouse) were removedfrom the temporal bones and the primordial organ of Cortimicrodiseccted using fine needles. Efforts were made to dis-sect out the lateral wall of the duct including the inmaturestria vascularis. Small patches of epithelia, containing primar-ily the greater and lesser epithelial ridges (including Kollick-er’s organ), were transferred to a culture dish and allowed torecover for 12–18 hours as an explant. The epithelial patcheswere then lifted, trying to remove as much underlying mesen-chyme as possible, dissociated into a single cell suspensionand plated on gelatin-coated dishes in the presence of eitherbFGF, EGF, IGF, or a combination of the three in a serum-free medium. Different combinations of these factors havebeen used to isolate neural stem cells from the central nervoussystem (CNS) [15] as well as stem cells from mouse ears [3].After 72 hours, bFGF was found to support cell growth betterthan the other factors, with cells reaching a density of 98.0 �13.1 cells per millimeter squared against 49.8 � 8.0 cells permillimeter squared (IGF, p < .01) and 42.4 � 7.8 (EGF, p <.001). A partially additive effect with the three factors com-bined was observed a week after the first (399.4 � 7.5 vs.290.6 � 15.5 for bFGF, p < .001) or second passage (390.0� 14.1 vs. 163.2 � 14.4 for bFGF alone, p < .001). Cells inIGF-1 only survived for a week, dying after the first passage.After 2 weeks, a small population of cells survived in EGFalone (Fig. 1A). Several independent (different donor sample)
1198 Isolation of hFASCs
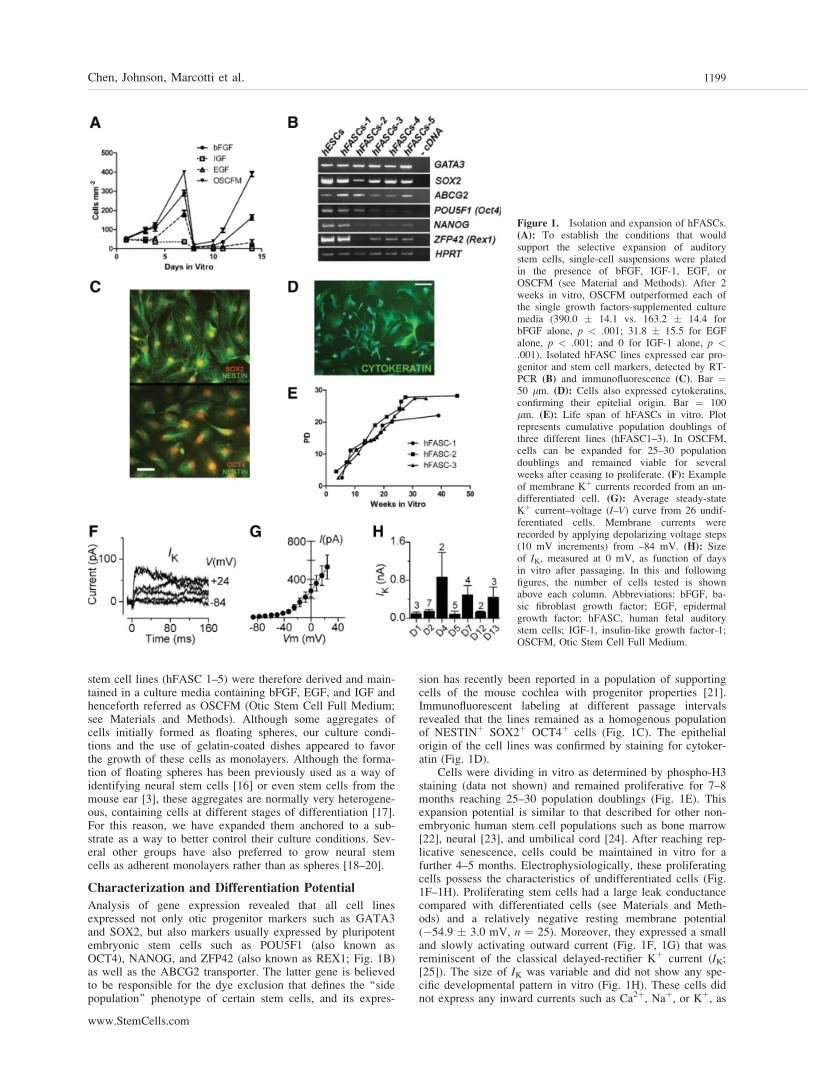
stem cell lines (hFASC 1–5) were therefore derived and main-tained in a culture media containing bFGF, EGF, and IGF andhenceforth referred as OSCFM (Otic Stem Cell Full Medium;see Materials and Methods). Although some aggregates ofcells initially formed as floating spheres, our culture condi-tions and the use of gelatin-coated dishes appeared to favorthe growth of these cells as monolayers. Although the forma-tion of floating spheres has been previously used as a way ofidentifying neural stem cells [16] or even stem cells from themouse ear [3], these aggregates are normally very heterogene-ous, containing cells at different stages of differentiation [17].For this reason, we have expanded them anchored to a sub-strate as a way to better control their culture conditions. Sev-eral other groups have also preferred to grow neural stemcells as adherent monolayers rather than as spheres [18–20].
Characterization and Differentiation Potential
Analysis of gene expression revealed that all cell linesexpressed not only otic progenitor markers such as GATA3and SOX2, but also markers usually expressed by pluripotentembryonic stem cells such as POU5F1 (also known asOCT4), NANOG, and ZFP42 (also known as REX1; Fig. 1B)as well as the ABCG2 transporter. The latter gene is believedto be responsible for the dye exclusion that defines the ‘‘sidepopulation’’ phenotype of certain stem cells, and its expres-
sion has recently been reported in a population of supportingcells of the mouse cochlea with progenitor properties [21].Immunofluorescent labeling at different passage intervalsrevealed that the lines remained as a homogenous populationof NESTINþ SOX2þ OCT4þ cells (Fig. 1C). The epithelialorigin of the cell lines was confirmed by staining for cytoker-atin (Fig. 1D).
Cells were dividing in vitro as determined by phospho-H3staining (data not shown) and remained proliferative for 7–8months reaching 25–30 population doublings (Fig. 1E). Thisexpansion potential is similar to that described for other non-embryonic human stem cell populations such as bone marrow[22], neural [23], and umbilical cord [24]. After reaching rep-licative senescence, cells could be maintained in vitro for afurther 4–5 months. Electrophysiologically, these proliferatingcells possess the characteristics of undifferentiated cells (Fig.1F–1H). Proliferating stem cells had a large leak conductancecompared with differentiated cells (see Materials and Meth-ods) and a relatively negative resting membrane potential(�54.9 � 3.0 mV, n ¼ 25). Moreover, they expressed a smalland slowly activating outward current (Fig. 1F, 1G) that wasreminiscent of the classical delayed-rectifier Kþ current (IK;[25]). The size of IK was variable and did not show any spe-cific developmental pattern in vitro (Fig. 1H). These cells didnot express any inward currents such as Ca2þ, Naþ, or Kþ, as
Figure 1. Isolation and expansion of hFASCs.(A): To establish the conditions that wouldsupport the selective expansion of auditorystem cells, single-cell suspensions were platedin the presence of bFGF, IGF-1, EGF, orOSCFM (see Material and Methods). After 2weeks in vitro, OSCFM outperformed each ofthe single growth factors-supplemented culturemedia (390.0 � 14.1 vs. 163.2 � 14.4 forbFGF alone, p < .001; 31.8 � 15.5 for EGFalone, p < .001; and 0 for IGF-1 alone, p <.001). Isolated hFASC lines expressed ear pro-genitor and stem cell markers, detected by RT-PCR (B) and immunofluorescence (C). Bar ¼50 lm. (D): Cells also expressed cytokeratins,confirming their epitelial origin. Bar ¼ 100lm. (E): Life span of hFASCs in vitro. Plotrepresents cumulative population doublings ofthree different lines (hFASC1–3). In OSCFM,cells can be expanded for 25–30 populationdoublings and remained viable for severalweeks after ceasing to proliferate. (F): Exampleof membrane Kþ currents recorded from an un-differentiated cell. (G): Average steady-stateKþ current–voltage (I–V) curve from 26 undif-ferentiated cells. Membrane currents wererecorded by applying depolarizing voltage steps(10 mV increments) from –84 mV. (H): Sizeof IK, measured at 0 mV, as function of daysin vitro after passaging. In this and followingfigures, the number of cells tested is shownabove each column. Abbreviations: bFGF, ba-sic fibroblast growth factor; EGF, epidermalgrowth factor; hFASC, human fetal auditorystem cells; IGF-1, insulin-like growth factor-1;OSCFM, Otic Stem Cell Full Medium.
Chen, Johnson, Marcotti et al. 1199
www.StemCells.com

previously reported for mouse and human embryonic stemcells [26].
To explore the potential of these cells to differentiate intothe different inner ear lineages, we tested different manipula-tions and culture conditions. On dissociation with trypsin,cells appeared to differentiate readily into neurons acquiring acharacteristic bipolar morphology. To determine whether dif-ferentiation was triggered by the mere dissociation of the cellsor specifically by the action of trypsin, cells were treated inparallel using either 0.125% trypsin or a nonenzymatic disso-ciating solution (see Materials and Methods). Although bothprocesses were equally efficient to produce a suspension ofisolated single cells, a significant difference in the generationof bipolar cells was evident. After trypsin treatment, 82.7% �17.5% cells became bipolar (2.3 � 0.3 pre-treatment, p <.001), whereas only 4.0% � 0.5% (6.3 � 0.4 pretreatment,n.s.) did when dissociated nonenzymatically (Fig. 2A). Non-enzymatic passaging therefore became our method of choicefor maintaining the undifferentiated cells, whereas trypsin wasused to trigger neuronal differentiation. The effect of severalgrowth factors in supporting the neuronal phenotype wastested. One week after trypsination, the combination of Shhtogether with bFGF was the most efficient in supporting neu-ronal differentiation as measured by bipolar morphology(53.4% vs. 15.7% obtained under bFGF, the second most effi-cient treatment, p < .05) and expression of POU4F1(BRN3A; Fig. 2B). On the other hand, IGF-1 appeared to beimportant in preventing differentiation, as shown by the lackof the neuronal marker POU4F1 when cells were alsoexposed to bFGF, justifying its inclusion in the formulation ofOSCFM. We therefore defined the ‘‘neuralizing conditions’’that induced differentiation as splitting by trypsination, incu-bation with 20 ng/ml bFGF plus 500 ng/ml Shh-N(C24II) for3 days and then supplementing the culture media with 10 ng/ml of BDNF and NT3. As Shh has been described to be im-portant for the up-regulation of neurogenin1 and therefore forthe initiation of sensory neurogenesis [27], it was removedfrom the media after 5 days and the cells were maintained inbFGF plus neurotrophins. Neurons obtained in this mannerexpressed NEUROGENIN1, POU4F1, b-TUBULIN III, NEU-ROFILAMENT 200, and PARVALBUMIN (Fig. 2C, 2D). Onthe other hand, when the cells were exposed to retinoic acid(10�6 M) and EGF (20 ng/ml), about 90% retained their epi-thelial morphology, forming small epithelial islands. EGF hasbeen reported to induce the expression of ATOH1 (alsoknown as MATH1/HATH1) in cochlear cultures [28], whereasretinoic acid has been shown to support hair cell differentia-tion in vitro [29]. Under these conditions, �85% of cellsexpressed ATOH1 and POU4F3 (BRN3C). Cells differentiat-ing under ‘‘hair cell conditions’’ (i.e., EGF and RA) alsoexpressed the hair cell marker MYOSIN VIIA (Fig. 2C, 2E).Moreover, some of the MYOSIN VIIA-positive cells dis-played a rearragement of the actin cytoskeleton, with the for-mation of a circumferential ring and an actin-rich area remi-niscent of the cuticular plate (Fig. 2E). These structures havebeen previously described as indicatives of hair cell differen-tiation in vitro using mouse cell lines [29]. PARVALBUMIN,which in the cochlea is expressed not only by neurons butalso by the hair cells, was as well expressed under ‘‘hair cellconditions.’’ There was evidence that at least some supportingcell differentiation was taking place under these conditions.GFAP a traditional marker of astroglial differentiation in theCNS is, in the cochlea, expressed by supporting cells [30]. Itsexpression was detected in some cells (Fig. 2F) together withthe cell cycle inhibitor p27kip1, another supporting cell marker[31]. Moreover, GJB6, encoding CONNEXIN 30 and forming
part of the gap junction between supporting cells in vivo [32],was detected by RT-PCR (Fig. 2C).
Electrophysiological Properties of DifferentiatingNeurons and Hair Cells
The ability of hFASCs to acquire biophysical properties char-acteristic of either neuronal or hair cell-like cells was investi-gated by recording membrane properties following the induc-tion of differentiation. Although cell differentiation wasevident from 2 days after the initiation of the neuronal differ-entiation protocol (D2), a few undifferentiated cells could stillbe recorded at later stages (24 of 77, D2–D17).
Cell differentiation was associated with a significantreduction (p < .0001) in the resting membrane input conduct-ance (see Materials and Methods) and for hair-cell like cells asignificantly more hyperpolarized resting membrane potential(hair cell-like cells: �65.6 � 2.7 mV, n ¼ 20, p < .05; neu-ronal-like cells: �59.9 � 2.3 mV, n ¼ 24). Cell membranecapacitance was not significantly different between undiffer-entiated and differentiated cells. On differentiation, the maincurrent expressed in hair cell-like cells was the inward recti-fier Kþ current (IK1), in which slow decay (Fig. 3A) and volt-age activation range (Fig. 3B) closely resembled that recordedin prehearing mouse cochlear hair cells [33]. Electrophysio-logical differentiation was already present in hair cell-likecells 5 days after the initiation of the RAþEGF treatment,with the size of IK1 increasing significantly with time (p <.001: Fig. 3C). Hair cell-like cells also expressed a small out-ward IK (Fig. 3D), which did not increase significantly withage (Fig. 3E) and is reminiscent of that recorded in undiffer-entiated cells (Fig. 1F), although it was on average muchsmaller. In addition to IK1 and IK, all hair cell-like cells alsoexpressed a sustained inward Ca2þ current (ICa, Fig. 3F). Thelack of ICa inactivation, together with its negative activationrange (positive to �67 mV defined as 5% of gmax: Fig. 3G),indicates the presence of an L-type Ca2þ channel containingthe CaV1.3 subunit [34] as previously found in mammaliancochlear inner hair cells [13]. The maximum size of ICa,measured at around �20 mV, increased significantly (p <.05) with differentiation (Fig. 3H). The expression of theCav1.3 subunit by cells under ‘‘hair cell conditions’’ was con-firmed by RT-PCR (Fig. 2C). The combination of IK1, IK, andICa, together with the biphysical features described earlier,can be interpreted as an electrophysiological signature charac-teristic of developing cochlear hair cells [35].
Neuronal-like cells began to differentiate at around D2by expressing two distinct IK’s and a Naþ current (INa),which was usually seen to precede the much slower outwardKþ current. ICa was not detected in these cells. Typical exam-ples of the two delayed-rectifier Kþ currents and their differ-ent activation voltage-dependencies are shown in Figure 4Aand 4B. The sizes of both IK’s as a function of days afterthe induction of differentiation is shown in Figure 4C. In con-trast to hair cell-like cells, hyperpolarizing voltage steps fromeither �64 mV or �84 mV did not elicit any IK1 (data notshown). Figure 4D shows an example of isolated INa recordedfrom the holding potential of �84 mV. INa activated at mem-brane potentials positive to �45 mV (5% of gmax; Fig. 4E)and reached a maximum size of �398 � 127 pA (n ¼ 15,D2–D7) at �15 mV. The size of INa also increased sig-nificantly (p < .02) over time (Fig. 4F). The expression pro-file of membrane currents in neuronal-like cells closelyresembled that found in primary auditory neurons of the ratcochlea [14].
Overall, about 49% of cells showed a neuronal-like phe-notype under neuralizing conditions whereas 56% of cells
1200 Isolation of hFASCs
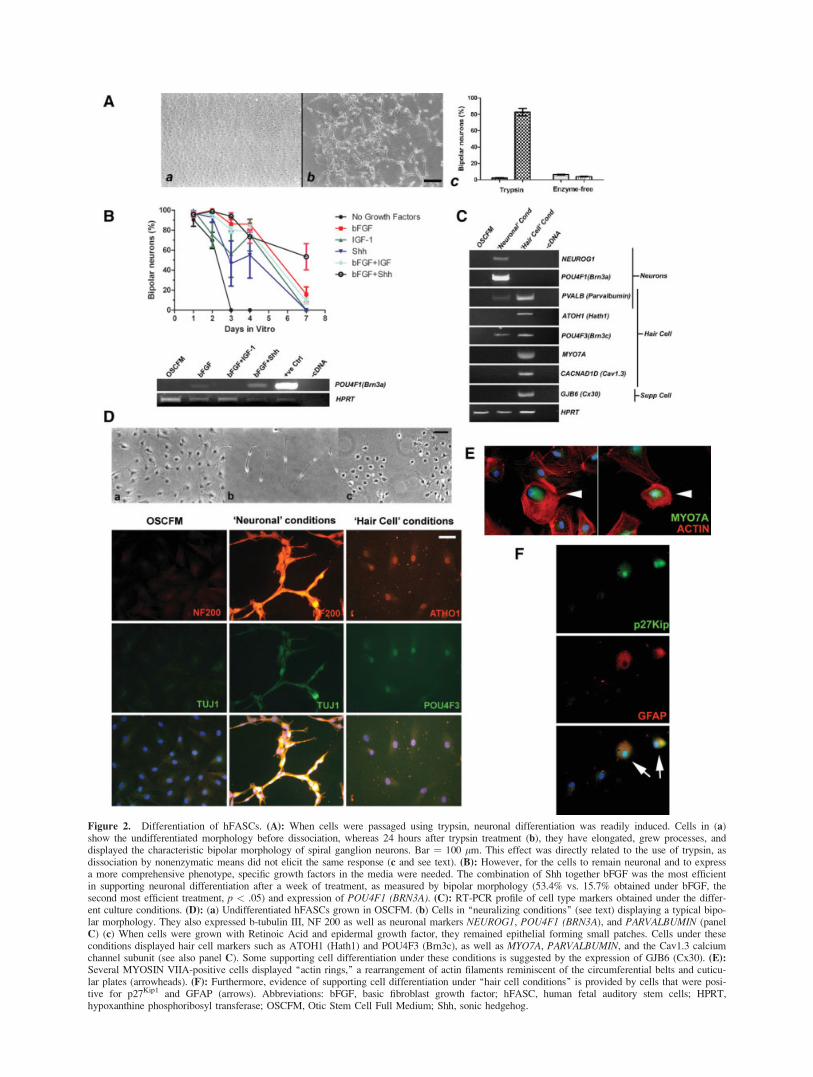
Figure 2. Differentiation of hFASCs. (A): When cells were passaged using trypsin, neuronal differentiation was readily induced. Cells in (a)show the undifferentiated morphology before dissociation, whereas 24 hours after trypsin treatment (b), they have elongated, grew processes, anddisplayed the characteristic bipolar morphology of spiral ganglion neurons. Bar ¼ 100 lm. This effect was directly related to the use of trypsin, asdissociation by nonenzymatic means did not elicit the same response (c and see text). (B): However, for the cells to remain neuronal and to expressa more comprehensive phenotype, specific growth factors in the media were needed. The combination of Shh together bFGF was the most efficientin supporting neuronal differentiation after a week of treatment, as measured by bipolar morphology (53.4% vs. 15.7% obtained under bFGF, thesecond most efficient treatment, p < .05) and expression of POU4F1 (BRN3A). (C): RT-PCR profile of cell type markers obtained under the differ-ent culture conditions. (D): (a) Undifferentiated hFASCs grown in OSCFM. (b) Cells in ‘‘neuralizing conditions’’ (see text) displaying a typical bipo-lar morphology. They also expressed b-tubulin III, NF 200 as well as neuronal markers NEUROG1, POU4F1 (BRN3A), and PARVALBUMIN (panelC) (c) When cells were grown with Retinoic Acid and epidermal growth factor, they remained epithelial forming small patches. Cells under theseconditions displayed hair cell markers such as ATOH1 (Hath1) and POU4F3 (Brn3c), as well as MYO7A, PARVALBUMIN, and the Cav1.3 calciumchannel subunit (see also panel C). Some supporting cell differentiation under these conditions is suggested by the expression of GJB6 (Cx30). (E):Several MYOSIN VIIA-positive cells displayed ‘‘actin rings,’’ a rearrangement of actin filaments reminiscent of the circumferential belts and cuticu-lar plates (arrowheads). (F): Furthermore, evidence of supporting cell differentiation under ‘‘hair cell conditions’’ is provided by cells that were posi-tive for p27Kip1 and GFAP (arrows). Abbreviations: bFGF, basic fibroblast growth factor; hFASC, human fetal auditory stem cells; HPRT,hypoxanthine phosphoribosyl transferase; OSCFM, Otic Stem Cell Full Medium; Shh, sonic hedgehog.
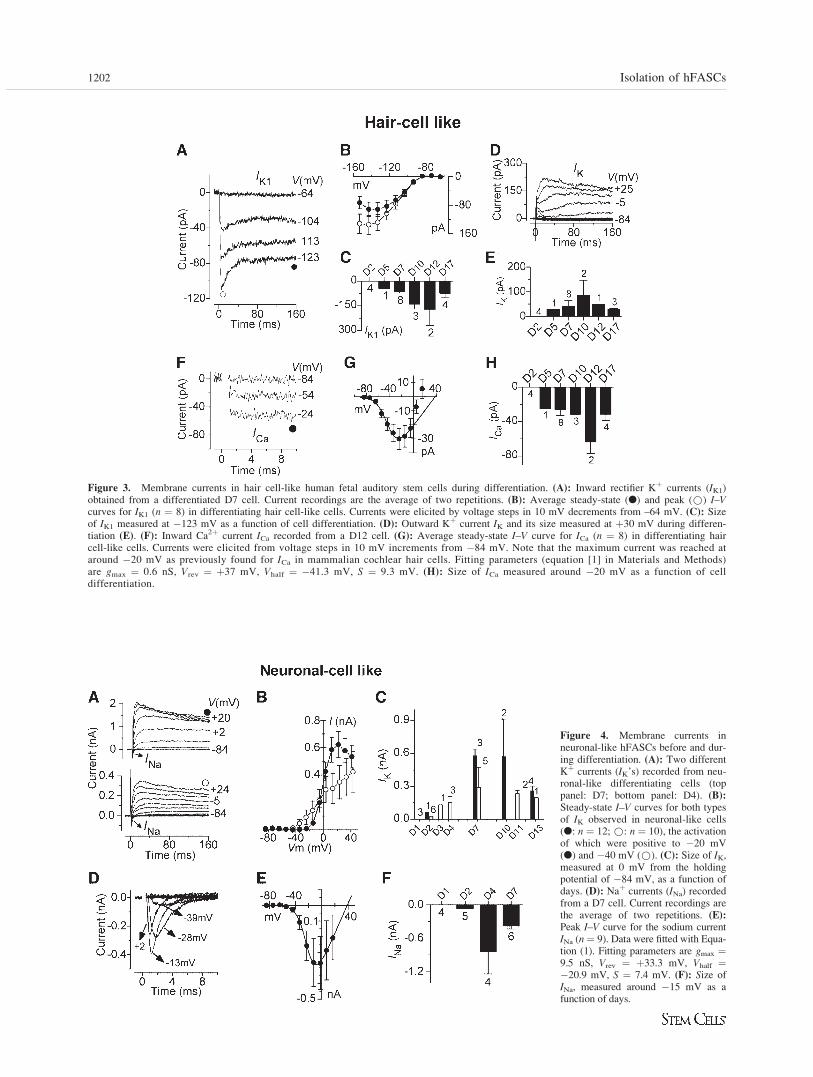
Figure 3. Membrane currents in hair cell-like human fetal auditory stem cells during differentiation. (A): Inward rectifier Kþ currents (IK1)obtained from a differentiated D7 cell. Current recordings are the average of two repetitions. (B): Average steady-state (l) and peak (*) I–Vcurves for IK1 (n ¼ 8) in differentiating hair cell-like cells. Currents were elicited by voltage steps in 10 mV decrements from –64 mV. (C): Sizeof IK1 measured at �123 mV as a function of cell differentiation. (D): Outward Kþ current IK and its size measured at þ30 mV during differen-tiation (E). (F): Inward Ca2þ current ICa recorded from a D12 cell. (G): Average steady-state I–V curve for ICa (n ¼ 8) in differentiating haircell-like cells. Currents were elicited from voltage steps in 10 mV increments from �84 mV. Note that the maximum current was reached ataround �20 mV as previously found for ICa in mammalian cochlear hair cells. Fitting parameters (equation [1] in Materials and Methods)are gmax ¼ 0.6 nS, Vrev ¼ þ37 mV, Vhalf ¼ �41.3 mV, S ¼ 9.3 mV. (H): Size of ICa measured around �20 mV as a function of celldifferentiation.
Figure 4. Membrane currents inneuronal-like hFASCs before and dur-ing differentiation. (A): Two differentKþ currents (IK’s) recorded from neu-ronal-like differentiating cells (toppanel: D7; bottom panel: D4). (B):
Steady-state I–V curves for both typesof IK observed in neuronal-like cells(l: n ¼ 12; *: n ¼ 10), the activationof which were positive to �20 mV(l) and �40 mV (*). (C): Size of IK,measured at 0 mV from the holdingpotential of �84 mV, as a function ofdays. (D): Naþ currents (INa) recordedfrom a D7 cell. Current recordings arethe average of two repetitions. (E):
Peak I–V curve for the sodium currentINa (n¼ 9). Data were fitted with Equa-tion (1). Fitting parameters are gmax ¼9.5 nS, Vrev ¼ þ33.3 mV, Vhalf ¼�20.9 mV, S ¼ 7.4 mV. (F): Size ofINa, measured around �15 mV as afunction of days.
1202 Isolation of hFASCs

displayed hair cell-like features under hair cell conditions.About 6% of cells investigated under hair cell conditionsshowed not only a small IK like that found in undifferentiatedcells (Fig. 1F–1H) but also, in some cases, an additional,small inward Kþ current yielding an electrophysiological pro-file similar to that of supporting cell types [36].
DISCUSSION
The hFASCs described here are the first in vitro renewablestem cell system derived from the human auditory organ andhave the potential for a variety of applications. Their progres-sive differentiation into the two main sensory lineages underwell-defined conditions, including the manifestation of func-tional electrophysiological characteristics of cochlear haircells [35] and auditory neurons [14], defines them as an idealmodel to study early developmental events. A previous studyby Rask-Andersen [37] showed that it is possible to isolate apopulation of neural progenitors from the adult human coch-lea. Those cells had a lineage-restricted capability, differenti-ating only into neurons and were able to multiply in vitro forup to 4 weeks. On the other hand, hFASCs are able to pro-duce hair cell-like cells and neurons and can be expanded andmaintained in vitro for up to a year.
The hair cell-like cells obtained in vitro express molecularmarkers and electrophysiological characteristics of this celltype, although they did not show the typical formation of ahair bundle. However, it is encouraging that the actin cyto-skeleton shows rearrangements indicative of a certain degreeof apical maturation. The gradual expression of ICa and IK1under hair cell conditions indicates that differentiation in vitrois a progressive phenomenon. For hair bundle differentiation,complex polarity cues are likely to be required that could beabsent in these culture conditions. Future experiments willaim to improve the differentiation system to achieve a morecomplete cell phenotype.
It is particularly intriguing the fact that dissociation bytrypsin can trigger neuronal differentiation. Certain inhibitorypathways are controlled by ligands and receptors located inadjacent cells and their signaling can be released by physi-cally separating the cells (i.e., lateral inhibition mediated bydelta/notch interactions). It was therefore unexpected thatonly the dissociation of cells by trypsin had an inductiveeffect, whereas an equally efficient separation (as judged bythe homogeneity of the single-cell suspension) by nonenzy-matic means failed to induce differentiation. This suggests a
direct role of the protease and will be the base for futureresearch.
Although considerable information has been obtainedabout the embryology of the ear using animal models, thelack of a human system has impaired the validation of suchinformation and delayed the translation to the clinic. Thedirect access to human cells from a well-defined developmen-tal stage that also have the ability to differentiate at will,should allow the exploration of features unique to humansthat may not be possible to investigate in a model system. Inthis context, it is interesting to highlight the fact that IGF-1failed to support growth and survival of the stem cells duringisolation. This was surprising given previous reports of itseffects in rat, mouse, and chick [38–40], and emphasizes theimportance of studying the human system. However, IGF-1seemed to be important for maintaining the cells in the undif-ferentiated state.
The versatility of a human model to study ear differentia-tion in vitro could prove very useful in the adaptation of theseconditions to other systems. At present, we are using theknowledge gathered with hFASCs to optimize the controlleddifferentiation of human ESCs into ear cell types (unpublishedobservations).
The search for auditory drugs and high-throughput toxicityassays, which have so far relied on the use of animal cells orunrelated human tissues, could benefit from stem cell use.Furthermore, the protocol employed to isolate and expandhFASCs has been designed using a chemically defined, serumfree medium without animal components. This should facili-tate the translation of the process into GMP standards for thederivation of clinical-grade cells and their potential therapeu-tic application.
ACKNOWLEDGMENTS
This work was supported by The Royal National Institute forDeaf People (M.N.R.), Deafness Research UK (M.N.R. andW.M.), and the Wellcome Trust (M.N.R. and W.M.). W.M. is aRoyal Society University Research Fellow.
DISCLOSURE OF POTENTIAL CONFLICTS
OF INTEREST
The authors indicate no potential conflicts of interest.
REFERENCES
1 Li H, Corrales CE, Edge A et al. Stem cells as therapy for hearingloss. Trends Mol Med 2004;10:309–315.
2 Rivolta MN, Holley MC.Gene arrays, cell lines, stem cells and sen-sory regeneration in mammalian ears. In: Salvi RJP, Popper AN, FayRR, eds. Hair Cell Regeneration, Repair and Protection. New York:Springer, 2008:257–307.
3 Li H, Liu H, Heller S. Pluripotent stem cells from the adult mouseinner ear. Nat Med 2003;9:1293–1299.
4 Li H, Roblin G, Liu H et al. Generation of hair cells by stepwise dif-ferentiation of embryonic stem cells. Proc Natl Acad Sci USA 2003;100:13495–13500.
5 Rivolta MN, Li H, Heller S.Generation of inner ear cell types fromembryonic stem cells. In: Turksen K, ed. Embryonic Stem Cells:Methods and Protocols. 2nd ed. Totowa, NJ: Humana Press, 2005:71–92.
6 Coleman B, Hardman J, Coco A et al. Fate of embryonic stem cellstransplanted into the deafened mammalian cochlea. Cell Transplant2006;15:369–380.
7 Hildebrand MS, Dalh HM, Hardman J et al. Survival of partially dif-ferentiated mouse embryonic stem cells in the scala media of theguinea pig cochlea. JARO 2005;6:341–354.
8 Hu Z, Wei D, Johansson CB et al. Survival and neural differentiationof adult neural stem cells transplanted into the mature inner ear. ExpCell Res 2005;302:40–47.
9 Sakamoto T, Nakagawa T, Endo T et al. Fates of mouse embryonicstem cells transplanted into the inner ears of adult mice and embry-onic chickens. Acta Otolaryngol Suppl 2004;551:48–52.
10 Oshima K, Grimm CM, Corrales CE et al. Differential distribution ofstem cells in the auditory and vestibular organs of the inner ear.J Assoc Res Otolaryngol 2006;8:18–31.
11 Chen W, Cacciabue-Rivolta DI, Moore HD et al. The human fetalcochlea can be a source for auditory progenitors/stem cells isolation.Hear Res 2007;233:23–29.
Chen, Johnson, Marcotti et al. 1203
www.StemCells.com

12 Marcotti W, Kros CJ. Developmental expression of the potassium cur-rent IK,n contributes to maturation of mouse outer hair cells. J Physiol1999;520(Part 3):653–660.
13 Marcotti W, Johnson SL, Rusch A et al. Sodium and calcium currentsshape action potentials in immature mouse inner hair cells. J Physiol2003;552:743–761.
14 Jagger DJ, Robertson D, Housley GD. A technique for slicing the ratcochlea around the onset of hearing. J Neurosci Methods 2000;104:77–86.
15 Ostenfeld T, Svendsen CN. Requirement for neurogenesis to proceedthrough the division of neuronal progenitors following differentiationof epidermal growth factor and fibroblast growth factor-2-responsivehuman neural stem cells. Stem Cells 2004;22:798–811.
16 Reynolds BA, Rietze RL. Neural stem cells and neurospheres—Re-evaluating the relationship. Nat Methods 2005;2:333–336.
17 Suslov ON, Kukekov VG, Ignatova TN et al. Neural stem cell hetero-geneity demonstrated by molecular phenotyping of clonal neuro-spheres. Proc Natl Acad Sci USA 2002;99:14506–14511.
18 Cattaneo E, McKay R. Proliferation and differentiation of neuronal stemcells regulated by nerve growth factor. Nature 1990;347:762–765.
19 Johe KK, Hazel TG, Muller T et al. Single factors direct the differen-tiation of stem cells from the fetal and adult central nervous system.Genes Dev 1996;10:3129–3140.
20 Pollard SM, Conti L, Sun Y et al. Adherent neural stem (NS) cells fromfetal and adult forebrain. Cereb Cortex 2006;16(Suppl 1):i112–i120.
21 Savary E, Hugnot JP, Chassigneux Y et al. Distinct population of haircell progenitors can be isolated from the postnatal mouse cochleausing side population analysis. Stem Cells 2007;25:332–339.
22 Simonsen JL, Rosada C, Serakinci N et al. Telomerase expressionextends the proliferative life-span and maintains the osteogenic poten-tial of human bone marrow stromal cells. Nat Biotechnol 2002;20:592–596.
23 Wright LS, Prowse KR, Wallace K et al. Human progenitor cells iso-lated from the developing cortex undergo decreased neurogenesis andeventual senescence following expansion in vitro. Exp Cell Res 2006;312:2107–2120.
24 Terai M, Uyama T, Sugiki T et al. Immortalization of human fetalcells: The life span of umbilical cord blood-derived cells can be pro-longed without manipulating p16INK4a/RB braking pathway. MolBiol Cell 2005;16:1491–1499.
25 Hille B.Ion Channels of Excitable Membranes.Sunderland, MA: Sina-uer Associates,2001.
26 Wang K, Xue T, Tsang SY et al. Electrophysiological properties ofpluripotent human and mouse embryonic stem cells. Stem Cells 2005;23:1526–1534.
27 Ota M, Ito K. Induction of neurogenin-1 expression by sonic hedge-hog: Its role in development of trigeminal sensory neurons. Dev Dyn2003;227:544–551.
28 Doetzlhofer A, White PM, Johnson JE et al. In vitro growth and dif-ferentiation of mammalian sensory hair cell progenitors: a requirementfor EGF and periotic mesenchyme. Dev Biol 2004;272:432–447.
29 Cacciabue-Rivolta D, Helyer R, Davies D et al. A model for mamma-lian cochlear hair cell differentiation in vitro: Effects of retinoic acidon cytoskeletal proteins and potassium conductances. Eur J Neurosci2007;25:957–973.
30 Rio C, Dikkes P, Liberman MC et al. Glial fibrillary acidic proteinexpression and promoter activity in the inner ear of developing andadult mice. J Comp Neurol 2002;442:156–162.
31 White PM, Doetzlhofer A, Lee YS et al. Mammalian cochlear sup-porting cells can divide and trans-differentiate into hair cells. Nature2006;441:984–987.
32 Zhao HB, Yu N. Distinct and gradient distributions of connexin26 andconnexin30 in the cochlear sensory epithelium of guinea pigs. J CompNeurol 2006;499:506–518.
33 Marcotti W, Geleoc GS, Lennan GW et al. Transient expression of aninwardly rectifying potassium conductance in developing inner andouter hair cells along the mouse cochlea. Pflugers Arch 1999;439:113–122.
34 Platzer J, Engel J, Schrott-Fischer A et al. Congenital deafness andsinoatrial node dysfunction in mice lacking class DL-type Ca2þ chan-nels. Cell 2000;102:89–97.
35 Housley GD, Marcotti W, Navaratnam D et al. Hair cells—Beyondthe transducer. J Membr Biol 2006;209:89–118.
36 Lawlor P, Marcotti W, Rivolta MN et al. Differentiation of mamma-lian vestibular hair cells from conditionally immortal, postnatal sup-porting cells. J Neurosci 1999;19:9445–9458.
37 Rask-Andersen H, Bostrom M, Gerdin B et al. Regeneration of humanauditory nerve. In vitro/in video demonstration of neural progenitorcells in adult human and guinea pig spiral ganglion. Hear Res 2005;203:180–191.
38 Zheng JL, Helbig C, Gao WQ. Induction of cell proliferation by fibro-blast and insulin-like growth factors in pure rat inner ear epithelialcell cultures. J Neurosci 1997;17:216–226.
39 Camarero G, Avendano C, Fernandez-Moreno C et al. Delayed innerear maturation and neuronal loss in postnatal Igf-1-deficient mice.J Neurosci 2001;21:7630–7641.
40 Camarero G, Leon Y, Gorospe I et al. Insulin-like growth factor 1 isrequired for survival of transit-amplifying neuroblasts and differentia-tion of otic neurons. Dev Biol 2003;262:242–253.
1204 Isolation of hFASCs