hsp90 invades the outside
TRANSCRIPT
N E W S A N D V I E W S
NATURE CELL BIOLOGY VOLUME 6 | NUMBER 6 | JUNE 2004 479
If you had to guess which proteins arerequired for the invasive nature of cancercells, your list might include integrins andmatrix metalloproteases (MMPs). It is unlike-ly, however, that it would include a molecularchaperone. Think again! Despite the fact thatthe cytosol and the extracellular space are verydifferent environments, many proteins can bemade to function on both sides of the cellmembrane. However, the question is whetherthey do so in real life. Well-established exam-ples are hard to find, but on page 507 of thisissue1, Jay and colleagues demonstrate thatHsp90 is necessary for extracellular matura-tion of the matrix metalloprotease MMP2. Byassisting MMP2, Hsp90 promotes the migra-tion of cancer cells through the extracellularprotein meshwork.
The story begins with another first: theauthors exploited a previously developed tech-nology for functional proteomics2 to screenfor extracellular proteins that are necessary forcancer cells to move across a basement mem-brane. Fluorophore-assisted light inactivation(FALI) affords the possibility of targeting aparticular surface protein for destructionusing the specificity of an antibody. Excitationof a fluorescein–antibody conjugate with bluelight eventually (after 1 h) leads to the destruc-tion of the antigen within close proximity. Jayand colleagues have now progressed from theproof of principle to a functional screen. Fromlibraries of monoclonal antibodies and phage-expressed single-chain antibodies, they firstidentified 29 antibodies that specifically recog-nise cell-surface proteins of the fibrosarcomamodel cell line HT-1080. These antibodieswere destroyed one at a time with FALI, andthe treated cancer cells were then assayed forinvasiveness. A relatively large proportion ofantibodies mediated inhibition of migrationby FALI, and most of these, perhaps not sur-prisingly, were directed against integrins. Theonly other antigen identified in the screen,recognised by two independently isolated
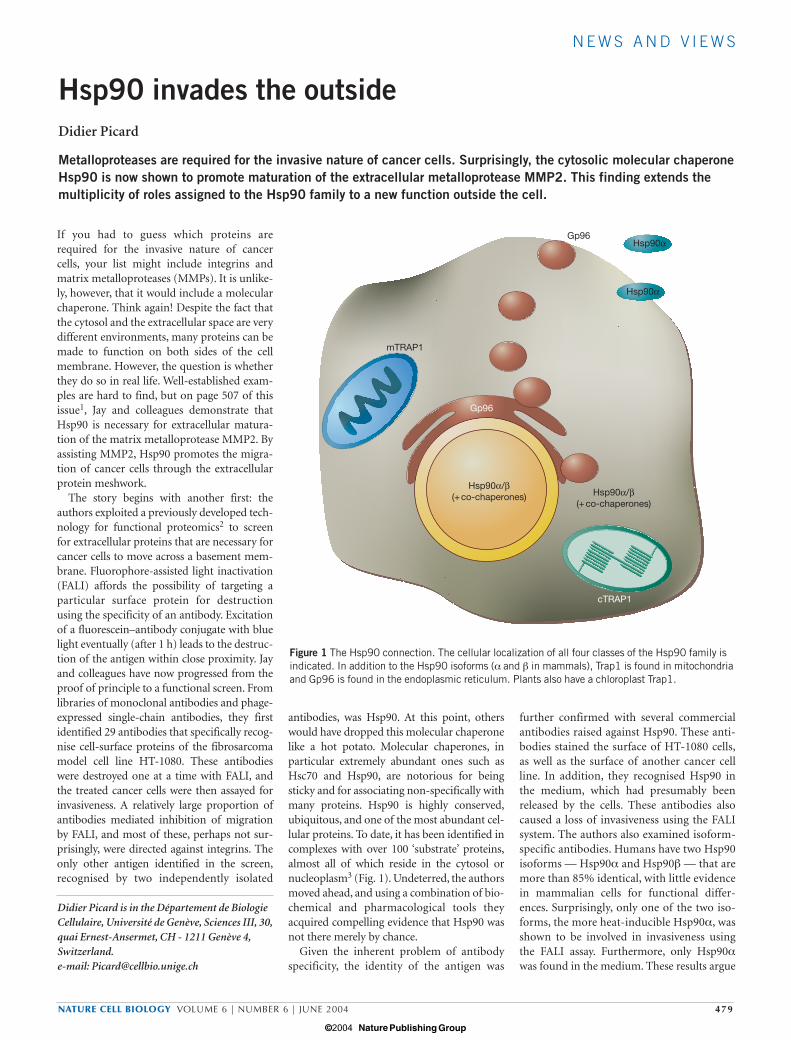
antibodies, was Hsp90. At this point, otherswould have dropped this molecular chaperonelike a hot potato. Molecular chaperones, inparticular extremely abundant ones such asHsc70 and Hsp90, are notorious for beingsticky and for associating non-specifically withmany proteins. Hsp90 is highly conserved,ubiquitous, and one of the most abundant cel-lular proteins. To date, it has been identified incomplexes with over 100 ‘substrate’ proteins,almost all of which reside in the cytosol ornucleoplasm3 (Fig. 1). Undeterred, the authorsmoved ahead, and using a combination of bio-chemical and pharmacological tools theyacquired compelling evidence that Hsp90 wasnot there merely by chance.
Given the inherent problem of antibodyspecificity, the identity of the antigen was
further confirmed with several commercialantibodies raised against Hsp90. These anti-bodies stained the surface of HT-1080 cells,as well as the surface of another cancer cellline. In addition, they recognised Hsp90 inthe medium, which had presumably beenreleased by the cells. These antibodies alsocaused a loss of invasiveness using the FALIsystem. The authors also examined isoform-specific antibodies. Humans have two Hsp90isoforms — Hsp90α and Hsp90β — that aremore than 85% identical, with little evidencein mammalian cells for functional differ-ences. Surprisingly, only one of the two iso-forms, the more heat-inducible Hsp90α, wasshown to be involved in invasiveness usingthe FALI assay. Furthermore, only Hsp90αwas found in the medium. These results argue
Hsp90 invades the outsideDidier Picard
Metalloproteases are required for the invasive nature of cancer cells. Surprisingly, the cytosolic molecular chaperoneHsp90 is now shown to promote maturation of the extracellular metalloprotease MMP2. This finding extends themultiplicity of roles assigned to the Hsp90 family to a new function outside the cell.
Didier Picard is in the Département de BiologieCellulaire, Université de Genève, Sciences III, 30,quai Ernest-Ansermet, CH - 1211 Genève 4,Switzerland.e-mail: [email protected]
Gp96
Gp96
Hsp90α/β(+ co-chaperones)
Hsp90α/β(+ co-chaperones)
mTRAP1
cTRAP1
Hsp90α
Hsp90α
Figure 1 The Hsp90 connection. The cellular localization of all four classes of the Hsp90 family isindicated. In addition to the Hsp90 isoforms (α and β in mammals), Trap1 is found in mitochondriaand Gp96 is found in the endoplasmic reticulum. Plants also have a chloroplast Trap1.
June N&V final 14/5/04 1:57 PM Page 479
© 2004 Nature Publishing Group

N E W S A N D V I E W S
480 NATURE CELL BIOLOGY VOLUME 6 | NUMBER 6 | JUNE 2004
against simple cell lysis as the source of extra-cellular Hsp90.
The specificity of FALI depends entirely onthe specificity of the antibody; thus, inhibi-tion of Hsp90 with a drug offers a welcomealternative. Geldanamycin (GA) is an exquis-itely specific inhibitor of the Hsp90 family4
that binds to the ATP-binding pocket ofHsp90 and blocks the ATPase cycle. As aresult, most Hsp90 substrates fail to matureproperly and are targeted to the proteasomefor degradation. GA blocks invasiveness justas the antibodies did in the FALI assay.However, as GA is membrane-permeable, thiscould be explained by the loss of intracellularsubstrate proteins of Hsp90. However, animpermeable version of GA attached to beadsworks just as well, strongly supporting theconclusion that extracellular Hsp90 isrequired for invasiveness.
What are the extracellular substrate(s) ofHsp90 that are important for invasiveness?The authors focused on MMP2 because it isknown to be important for invasiveness ofHT-1080 cells. MMPs are secreted as inactivepro-proteins and are activated extracellularlyby proteolysis. Jay and colleagues found byimmunoprecipitation that MMP2 is associat-ed with Hsp90 in the medium — again, onlywith the α isoform — and that GA inhibits itsactivation. Moreover, MMP2 seems to be thesole relevant target of Hsp90α for this partic-ular function, as supplementing cells withactive MMP2 restores invasiveness to cells thathad been inhibited by GA.
This discovery is exciting in two ways: first,from a clinical perspective, because it pro-vides a novel extracellular drug target forwhich promising compounds not only exist,but are currently being tested in clinical trials;second, from a cell biological and biochemi-cal perspective, because it is one of the best-supported cases for an extracellular role of a‘classical’ intracellular molecular chaperone.From both perspectives, however, one of thefirst points on the agenda will be to deter-mine the generality of the finding. Do othertypes of cancer cell lines and genuinetumours secrete Hsp90α, and is Hsp90αrequired for the maturation of other MMPs?If so, membrane-impermeable Hsp90inhibitors with pharmacological properties
entirely different from GA might representpotent anti-metastatic drugs for a large vari-ety of cancers. It should be noted that thisnovel therapeutic approach, which would tar-get extracellular functions of Hsp90, is con-ceptually different from current therapiesusing GA-type compounds; these therapiesare based on the unusually high affinity ofGA for intracellular Hsp90 complexes intumour cells versus Hsp90 in normal cells5.
Extracellular functions have already beendescribed for molecular chaperones that arenormally resident within cells. For example,Hsp70 and Gp96 (the Hsp90-family memberfrom the endoplasmic reticulum; Fig. 1) seemto modulate the immune response by present-ing small peptides6. In addition, recent evi-dence has accumulated to indicate that Hsp90itself can be secreted and may have additionalextracellular functions. Vascular smooth mus-cle cells secrete Hsp90α (not Hsp90β)7, andmembranes of other cells contain immobileHsp90 as part of a multimeric lipopolysaccha-ride (LPS) receptor complex8,9. Exogenousaddition of Hsp90 to culture medium hassome cell stimulatory effects. More impor-tantly, antibodies to Hsp90 block LPS sig-nalling8,9 and the activation of prekallikreinon the membrane of endothelial cells10.Intriguingly, both MMP2 and prekallikreinare proteases that are made as pro-proteinsand require proteolytic activation. The matu-ration of prekallikrein by high-molecular-weight kininogen can be promoted by Hsp90in a purified system10.
So how does Hsp90 get to the membraneand/or the extracellar space? The knownHsp90 isoforms do not carry recognisable sig-nal sequences that would target them to thesecretory pathway or to membranes. However,we cannot exclude the possibility that an alter-natively spliced version of Hsp90α contains asecretory signal sequence; Hsp90α was identi-fied on the basis of antibody recognition andpartial mass spectrometric analysis, so the pre-cise amino-terminal sequence is not known.Interestingly, the Ensembl.org entry for thehuman Hsp90α gene points out an alternativetranscript with two extra exons preceding thestandard first exon. Closer inspection revealsthat translation initiation at an ATG within thesecond exon would produce an extended
Hsp90 with a predicted N-terminal signal pep-tide. After its cleavage, the secreted Hsp90would encompass the entire known Hsp90sequence with only a 3K N-terminal exten-sion. It is also conceivable that the ‘canonical’Hsp90α ends up associated with the outside ofcells through an unconventional protein secre-tion pathway. In the latter scenario, however,one would have to explain why Hsp90β doesnot follow the same route as Hsp90α.
A primarily cytosolic protein such as Hsp90can be relatively easily pictured as a solubleprotein in the extracellular space, but how arewe to imagine its membrane association? Theexact topology remains to be determined withmore detailed analyses. The experiments withthe GA-coupled beads suggest that, at the veryleast, the ATPase domain is close to the surfaceand is required for MMP2 maturation,although it is noteworthy that Hsp90 is capa-ble of certain ATP-independent chaperonefunctions11. If Hsp90α is both membrane-associated and soluble, which of the twoforms is the functional one for a given sub-strate, say MMP2?
A major remaining challenge will be tounderstand the molecular function of extra-cellular Hsp90. How does a molecular chaper-one, ‘accustomed’ to the cytosol, work in anoxidative environment with considerablylower ATP concentrations? Perhaps extracellu-lar Hsp90 works less as a chaperone and moreas a scaffolding protein, linking to activatingproteases for example. A lonely extracellularshift indeed, if it is to function without itsusual cohort of co-chaperones3, none of whichhave yet been sighted outside the cell.
1. Eustace, B. K. et al. Nature Cell Biol. 6, 507–510(2004).
2. Beck, S. et al. Proteomics 2, 247–255 (2002).3. Picard, D. Cell. Mol. Life Sci. 59, 1640–1648
(2002).4. Neckers, L. & Ivy, S. P. Curr. Opin. Oncol. 15,
419–424 (2003).5. Kamal, A. et al. Nature 425, 407–410 (2003).6. Tsan, M. F. & Gao, B. Am. J. Physiol. Cell Physiol.
286, C739–C744 (2004).7. Liao, D. F. et al. J. Biol. Chem. 275, 189–196
(2000).8. Triantafilou, K., Triantafilou, M. & Dedrick, R. L.
Nature Immunol. 2, 338–345 (2001).9. Triantafilou, K. et al. J. Cell Sci. 114, 2535–2545
(2001).10. Joseph, K., Tholanikunnel, B. G. & Kaplan, A. P. Proc.
Natl Acad. Sci. USA 99, 896–900 (2002).11. Wiech, H., Buchner, J., Zimmermann, R. & Jakob, U.
Nature 358, 169–170 (1992).
June N&V final 14/5/04 1:57 PM Page 480
© 2004 Nature Publishing Group