how localized are the distributions of australian scarabs (coleoptera, scarabaeoidea)?
TRANSCRIPT

Diversity and Distributions (1999) 5, 143–149
BIODIVERSITY RESEARCH
How localized are the distributions of Australian scarabs(Coleoptera, Scarabaeoidea)?
P. G. ALLSOPP Bureau of Sugar Experiment Stations, Private Bag 4, Bundaberg DC 4670, Australia, E-mail:[email protected]
Abstract. The degree of localization in the However, the Australian fauna lacks the highproportion of widespread species characteristic of thedistribution of Australian scarabaeine (320 spp.),
aphodiine (160 spp.), geotrupid (166 spp.) and West Palaearctic Scarabaeinae, Aphodiinae andGeotrupidae. This difference reflects the differentdynastine (182 spp.) scarabs is analysed. Localization
differs among these groups, appearing to increase ecological histories of the two regions.marginally from the Dynastinae, to the Aphodiinae,to the Scarabaeinae, to the Geotrupidae, but with Key words. Localized species distributions,
endemic, relictual, Australia, Aphodiinae, Dynastinae,significant differences within groups, e.g. thescarabaeine tribes Onthophagini and Scarabaeini. Geotrupidae, Scarabaeinae.
INTRODUCTION the range of localization of Australian scarabs isanalysed in the same three groups, the Scarabaeinae,
Species diversity is not uniformly distributed across Aphodiinae and Geotrupidae, and for comparison inthe planet’s surface, being characterized by disharmony one of the phytophagous scarab groups, theand impoverishment (Quammen, 1996). Many species Dynastinae.have very limited distributions. These species are oftensaid to be endemic (Horton, 1973; Lumaret & Lobo,
MATERIALS AND METHODS1996), but this term is misleading and misused. Firstly,because it is relative, depending on the spatial scale
The Australian (including Tasmania and nearby off-considered. Secondly, because endemism (originated in
shore islands) and the West Palaearctic (includingthe same place as it is found) is one of two routes by
Britain, Ireland and Mediterranean islands) regionswhich a species’ present distribution can be limited;
have similar land areas, about 7.7×106 andthe other is relictualism (surviving in a given place
12×106 km2, respectively. To allow direct comparisonwhilst disappearing elsewhere) (Quammen, 1996). A
with Lumaret & Lobo’s (1996) analyses, I includedbetter term to describe a species with an observed
Tasmania and other major Australian islands (Lumaretlimited distribution is localized.
& Lobo included Britain, Ireland and MediterraneanAs noted by Lumaret & Lobo (1996), the study of
islands) and defined (Fig. 1) the same six geographicallocalized species is of great use, both for reconstructing
range-size classes as used by them:the biogeographic history of life and areas and for
I species with a distribution in an area less than ormaking decisions about the conservation of areas andequal to 0.1% of the total surface area of Australiaspecies. Lumaret & Lobo (1996) analysed the range of(7.7×103 km2);localization in three groups of scarabaeoid dung beetles
II species with a distribution in an area between 0.1%from the West Palaearctic region and showed that thereand 0.5% of the total surface area of Australiawas significant variation among the groups. This they(3.9×104 km2);attributed to differences among groups in tolerance to
III species with a distribution in an area betweenenvironmental variables and dispersal capacity. Are0.5% and 1% of the total surface area of Australiathese differences the same in a different region with a
different biogeographic history? To help answer this, (7.7×104 km2);
143 1999 Blackwell Science Ltd. http://www.blackwell-science.com/dad

144 P. G. Allsopp
Fig. 1. Sizes of the first five range-size classes defined for Australia.
IV species with a distribution in an area between 1% arrived in Australia in the last 210 years were excluded.and 5% of the total surface area of Australia I used a system of paper chips to estimate range sizes,(3.9×105 km2); similar to that used by Lumaret & Lobo (1996).
V species with a distribution in an area between 5% Kolmogorov–Smirnov two-sample tests (Steel &and 10% of the total surface area of Australia Torrie, 1980) were used to assess the hypothesis of(7.7×105 km2); independence among geographical range-size classes
VI species with a distribution in an area greater than and each of the Australian groups (including a10% of the total surface area of Australia. comparison between the Onthophagini and
Scarabaeini). The test was also used to assess theLumaret & Lobo (1996) considered a species localizedhypothesis of independence for each of theif its distribution fell into classes I–IV.Scarabaeinae, Aphodiinae and Geotrupidae of theI took the ranges of the species from: AphodiinaeAustralian and West Palaearctic regions. The(Aphodiidae of Lumaret & Lobo, 1996) – Stebnicka &Kolmogorov–Smirnov test is preferable to the chi-Howden (1994, 1995, 1996, 1997) and Storey & Howdensquare test because it exploits information in the(1996); Dynastinae – Allsopp (1990, 1993), Allsopp &ordering of the categories as well as means andCarne (1986), Carne (1957, 1976, 1978, 1980, 1981,variances (Analytical Software, 1996). However, I used1985a,b), Carne & Allsopp (1987) and Endrodi (1977,the chi-square test to assess the distributions for a1978); Scarabaeinae (Scarabaeidae of Lumaret & Lobo,theoretical evenness of species among range-size classes1996) – Hill (1993), Matthews (1972, 1974, 1976),for each of the major Australian groups; in this caseMatthews & Stebnicka (1986), Storey (1977, 1984, 1986,the ordering of the classes is immaterial and the chi-1991) and Storey & Weir (1988, 1990); Geotrupidae –square test is less conservative than theCarne (1965), Howden (1974, 1975, 1979, 1985a, 1992),
Krikken (1976) and Nikolajev (1990). Species that have Kolgomorov–Smirnov test.
1999 Blackwell Science Ltd, Diversity and Distributions, 5, 143–149
Localized distributions of Australian scarabs 145
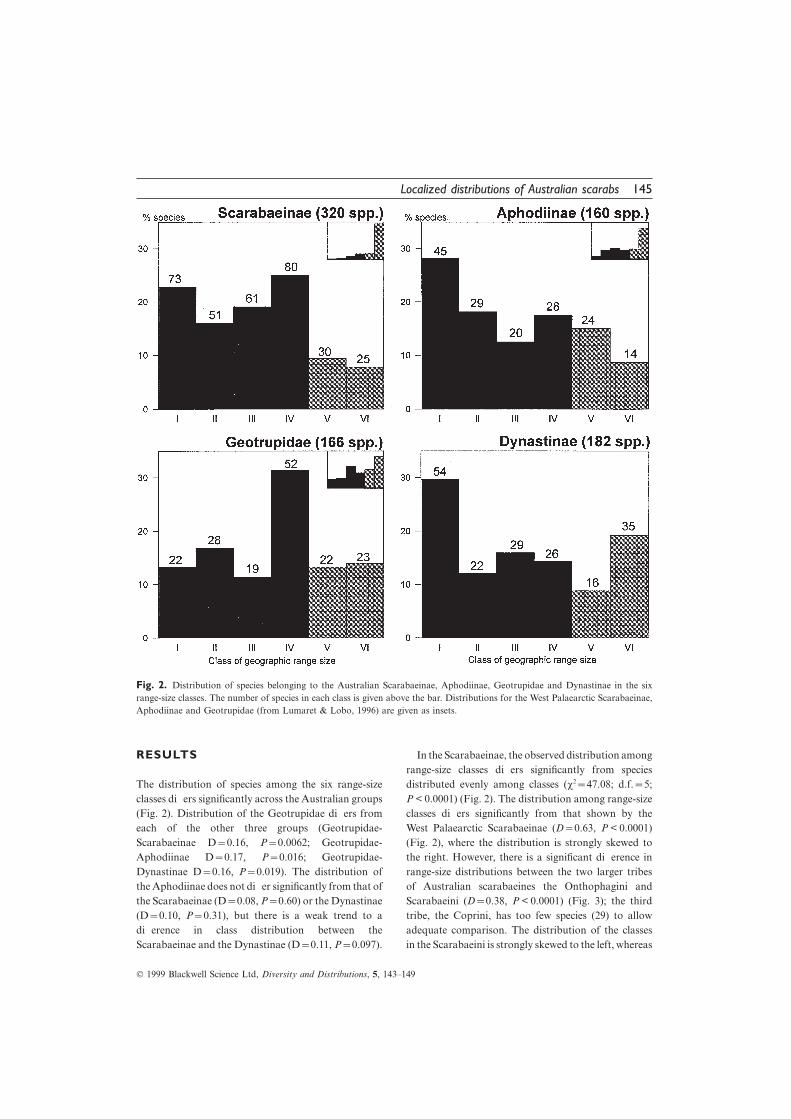
Fig. 2. Distribution of species belonging to the Australian Scarabaeinae, Aphodiinae, Geotrupidae and Dynastinae in the sixrange-size classes. The number of species in each class is given above the bar. Distributions for the West Palaearctic Scarabaeinae,Aphodiinae and Geotrupidae (from Lumaret & Lobo, 1996) are given as insets.
RESULTS In the Scarabaeinae, the observed distribution amongrange-size classes differs significantly from speciesdistributed evenly among classes (v2=47.08; d.f.=5;The distribution of species among the six range-size
classes differs significantly across the Australian groups P < 0.0001) (Fig. 2). The distribution among range-sizeclasses differs significantly from that shown by the(Fig. 2). Distribution of the Geotrupidae differs from
each of the other three groups (Geotrupidae- West Palaearctic Scarabaeinae (D=0.63, P < 0.0001)(Fig. 2), where the distribution is strongly skewed toScarabaeinae D=0.16, P=0.0062; Geotrupidae-
Aphodiinae D=0.17, P=0.016; Geotrupidae- the right. However, there is a significant difference inrange-size distributions between the two larger tribesDynastinae D=0.16, P=0.019). The distribution of
the Aphodiinae does not differ significantly from that of of Australian scarabaeines the Onthophagini andScarabaeini (D=0.38, P < 0.0001) (Fig. 3); the thirdthe Scarabaeinae (D=0.08, P=0.60) or the Dynastinae
(D=0.10, P=0.31), but there is a weak trend to a tribe, the Coprini, has too few species (29) to allowadequate comparison. The distribution of the classesdifference in class distribution between the
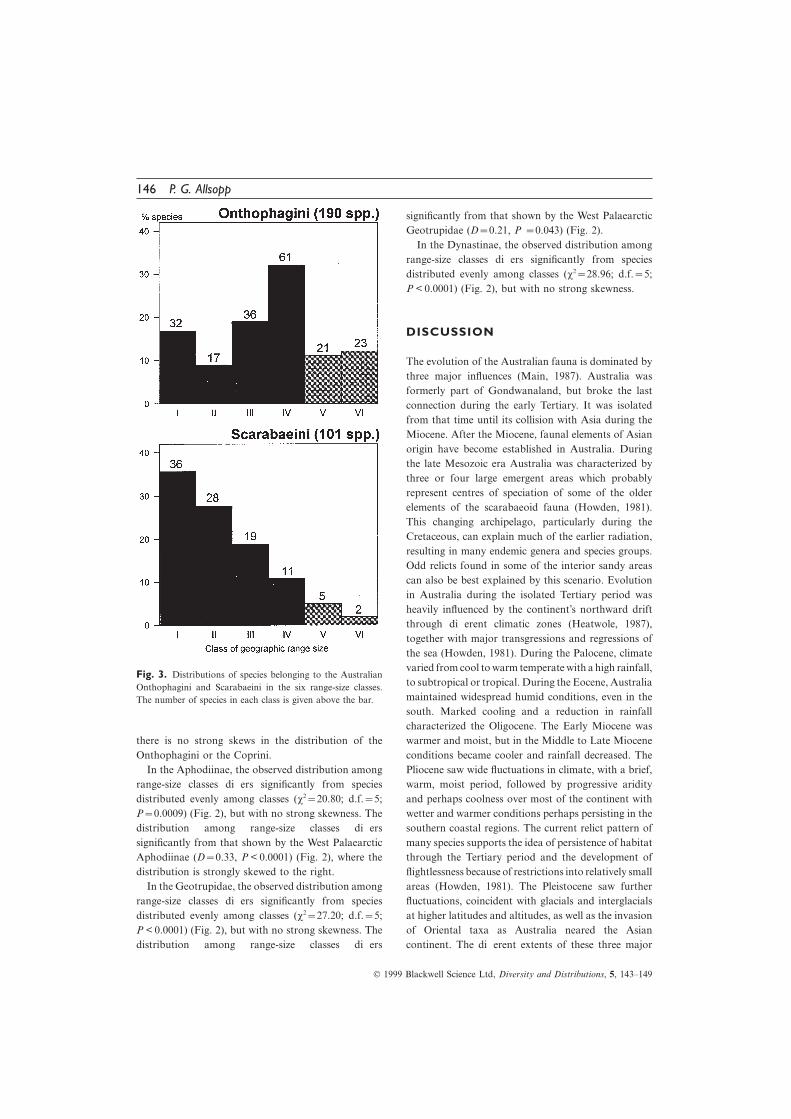
Scarabaeinae and the Dynastinae (D=0.11, P=0.097). in the Scarabaeini is strongly skewed to the left, whereas
1999 Blackwell Science Ltd, Diversity and Distributions, 5, 143–149
146 P. G. Allsopp
significantly from that shown by the West PalaearcticGeotrupidae (D=0.21, P =0.043) (Fig. 2).
In the Dynastinae, the observed distribution amongrange-size classes differs significantly from speciesdistributed evenly among classes (v2=28.96; d.f.=5;P < 0.0001) (Fig. 2), but with no strong skewness.
DISCUSSION
The evolution of the Australian fauna is dominated bythree major influences (Main, 1987). Australia wasformerly part of Gondwanaland, but broke the lastconnection during the early Tertiary. It was isolatedfrom that time until its collision with Asia during theMiocene. After the Miocene, faunal elements of Asianorigin have become established in Australia. Duringthe late Mesozoic era Australia was characterized bythree or four large emergent areas which probablyrepresent centres of speciation of some of the olderelements of the scarabaeoid fauna (Howden, 1981).This changing archipelago, particularly during theCretaceous, can explain much of the earlier radiation,resulting in many endemic genera and species groups.Odd relicts found in some of the interior sandy areascan also be best explained by this scenario. Evolutionin Australia during the isolated Tertiary period washeavily influenced by the continent’s northward driftthrough different climatic zones (Heatwole, 1987),together with major transgressions and regressions ofthe sea (Howden, 1981). During the Palocene, climatevaried from cool to warm temperate with a high rainfall,
Fig. 3. Distributions of species belonging to the Australianto subtropical or tropical. During the Eocene, AustraliaOnthophagini and Scarabaeini in the six range-size classes.maintained widespread humid conditions, even in theThe number of species in each class is given above the bar.south. Marked cooling and a reduction in rainfallcharacterized the Oligocene. The Early Miocene waswarmer and moist, but in the Middle to Late Miocenethere is no strong skews in the distribution of the
Onthophagini or the Coprini. conditions became cooler and rainfall decreased. ThePliocene saw wide fluctuations in climate, with a brief,In the Aphodiinae, the observed distribution among
range-size classes differs significantly from species warm, moist period, followed by progressive aridityand perhaps coolness over most of the continent withdistributed evenly among classes (v2=20.80; d.f.=5;
P=0.0009) (Fig. 2), but with no strong skewness. The wetter and warmer conditions perhaps persisting in thesouthern coastal regions. The current relict pattern ofdistribution among range-size classes differs
significantly from that shown by the West Palaearctic many species supports the idea of persistence of habitatthrough the Tertiary period and the development ofAphodiinae (D=0.33, P < 0.0001) (Fig. 2), where the
distribution is strongly skewed to the right. flightlessness because of restrictions into relatively smallareas (Howden, 1981). The Pleistocene saw furtherIn the Geotrupidae, the observed distribution among
range-size classes differs significantly from species fluctuations, coincident with glacials and interglacialsat higher latitudes and altitudes, as well as the invasiondistributed evenly among classes (v2=27.20; d.f.=5;
P < 0.0001) (Fig. 2), but with no strong skewness. The of Oriental taxa as Australia neared the Asiancontinent. The different extents of these three majordistribution among range-size classes differs
1999 Blackwell Science Ltd, Diversity and Distributions, 5, 143–149

Localized distributions of Australian scarabs 147
influences are reflected in the different occurrence of plants nor to animal dung, soil type and rainfall appearto be the most limiting factors (Howden, 1981). Manylocalized species amongst the groups of scarabs that I
analysed. The degree of localization appears to increase of the species from the Eyrean subregion appear to berelicts, surviving over long periods as the continentmarginally from the Dynastinae, to the Aphodiinae,
to the Scarabaeinae, to the Geotrupidae. became drier (Howden, 1981) – this may account forthe high proportion of species in range-size class IV.Much of the Australian dynastine fauna appears to
have its origin in the pre-Jurrasic to Jurrasic periods The distribution of range-size classes of theAustralian and West Palaearctic faunas differs greatly.and is concentrated in the arid Eyrean subregion
(Carne, 1957; Howden, 1981; Allsopp, 1995). The much Why is this so? In Australia, the significant, but notsevere, fluctuations in climate during the Mesozoic andyounger and much smaller Malayan element is
concentrated in the north and north-east (Torresian Cenozoic eras presumably led to isolation ofpopulations and evolution of new species. In contrast,subregion) and derives from ancestors arriving in the
Cenozoic, presumably after the Miocene. Speciation in the West Palaearctic fauna was heavily influenced bythe severe glacials and interglacials of the Pleistocene,the older element presumably benefited from the
climate fluctuations during the Mesozoic and early which necessitated recolonization of large areas(Hanski, 1991). This is most noticeable in the cool-Cenozoic eras.
The Australian aphodiines were poorly studied until susceptible Scarabaeinae, which is dominated bywidespread, dispersing species; the more cool-tolerantrecently. Howden (1981) speculated that, because they
are generally the smallest Australian scarabs, they Aphodiinae has a much higher proportion of localizedspecies. The Geotrupidae is the least cool-tolerant ofare the more widely dispersed. My analysis does not
support this, and the revisions of the Australian the groups analysed by Lumaret & Lobo (1996) andits degree of localization appears to be heavilyaphodiines by Stebnicka & Howden (1994, 1995, 1996,
1997) recognized a large number of localized species. influenced by the flightless genus Thorectes and thegroup’s basal lineages in the western Mediterranean.Among the indigenous Australian Aphodiinae there
are a number of forms that do not appear to have A further factor that has undoubtedly influenced thedistributions of both faunas is the impact of humans.close relatives elsewhere. Species of both old and recent
elements are concentrated around the south-eastern Within Australia, habitat destruction has beenimportant in eliminating species from the Australianand eastern periphery of the continent (Stebnicka &
Howden, 1995). Many of these appear to have had a fauna and in endangering the survival of others(Heatwole, 1987). There is no evidence that anylong residency in Australia, similar to the Dynastinae.
The Scarabaeinae, as a whole, have a large Australian scarab has become extinct, but one canspeculate that large species of Scarabaeinae may haveproportion of localized species. However, differences
among groups, especially the Onthophagini and been associated with the dung of larger native mammalsin the same way that the large Heliocopris spp. areScarabaeini, are even more evident than in, say, the
Dynastinae. The extreme tendency towards localized associated with elephant dung in Africa. These largenative mammals went extinct at the close of thespecies within the Scarabaeini reflect the group’s long
residency in Australia and a high proportion of Pleistocene and any associated large dung beetles mayhave disappeared at the same time. However, otherflightless species, reflecting limitation of habitat area
coupled with environmental stability (Matthews, 1974; species have benefited from changes to habitat,becoming more common (e.g. scarabs associated withHowden, 1981). Concentration of species along the
periphery of the continent (Matthews, 1974; Allsopp, sugarcane, Allsopp & Logan, 1999), if not more widelydistributed. Similar changes have probably occurred1995) probably reflects early expansion and later
extinction in unfavourable habitats. Many of the in the West Palaearctic region.In summary, the occurrence of localized speciesOnthophagini, in contrast, seem to be good dispersers
and their ancestors apparently arrived much more differs among groups of Australian scarabs, reflectingthe continent’s past isolation and recent exchange fromrecently from the north in less than 34 separate
incursions (Matthews, 1972; Howden, 1981). Asia. The occurrence is very different from that in thesame groups of West Palaearctic scarabs, lacking theThe Geotrupidae generally has patterns like the older
elements of the scarabaeine subfamilies. Many species high proportion of widespread species. The comparisonillustrates Howden’s (1985b) contention that bothare cool or cold tolerant and occur in mesic and arid
environments. Because they are not tied to various evolutionary time and ecological attributes are
1999 Blackwell Science Ltd, Diversity and Distributions, 5, 143–149

148 P. G. Allsopp
from north Queensland (Coleoptera: Scarabaeidae).important in determining the taxon cycle/speciesJournal of the Australian Entomological Society, 24,packing concepts.75–76.
Carne, P.B. & Allsopp, P.G. (1987) Novapus macfarlandisp. n. and notes on other species of Novapus Sharp
REFERENCES (Coleoptera: Scarabaeidae: Dynastinae) from Australia.Journal of the Australian Entomological Society, 26,309–312.Allsopp, P.G. (1990) Phyllognathus carnei sp. n. &
Endrodi, S. (1977) Monographie Dynastinae VIII. Tribus:Oryctoderus latitarsis Boisduval (Coleoptera:Phileurini (Coleoptera: Lamellicornia). EntomologischeScarabaeidae: Dynastinae), two additions to theAbhandlungen staatliches Museum fur Tierkunde inAustralian fauna. Journal of the AustralianDresden, 41, 93–134.Entomological Society, 29, 229–232.
Endrodi, S. (1978) Oryctoderinus walfordorum gen. et sp.Allsopp, P.G. (1993) Trissodon howdenorum and T.n. (Coleoptera: Scarabaeidae: Dynastinae). Journal ofbidentatus spp. n. from Western Australia with furtherthe Australian Entomological Society, 17, 159–161.records of other Trissodon spp. (Coleoptera:
Hanski, I. (1991) North temperate dung beetles. DungScarabaeidae: Dynastinae). Journal of the AustralianBeetle Ecology (ed. by I. Hanski & Y. Cambefort), pp.Entomological Society, 32, 201–205.75–96. Princeton University Press, Princeton.Allsopp, P.G.. (1995) Biogeography of the Australian
Heatwole, H. (1987) Major components and distributionsDynastinae, Rutelinae, Scarabaeinae, Melolonthini,of the terrestrial fauna. Fauna of Australia, Vol. 1A,Scitalini and Geotrupidae (Coleoptera: Scarabaeoidea).General Articles (ed. by G.R. Dyne & D.W. Walton),Journal of Biogeography 22, 31–48.pp. 101–135. Australian Government Publishing Service,Allsopp, P.G. & Carne, P.B. (1986) Teinogenys mooniensisCanberra.sp. n. & new locality records for other species of
Hill, C.J. (1993) The species composition and seasonalityTeinogenys Sharp (Coleoptera: Scarabaeidae:of an assemblage of tropical Australian dung beetlesDynastinae). Journal of the Australian Entomological(Coleoptera: Scarabaeidae: Scarabaeinae). AustralianSociety, 25, 89–93.Entomologist, 20, 121–126.Allsopp, P.G. & Logan, D.P. (1999) Seasonal flight activity
Horton, D.R. (1973) Endemism and zoogeography.of scarab beetles (Coleoptera: Scarabaeidae) associatedSystemic Zoology, 22, 84–86.with sugarcane in southern Queensland. Australian
Howden, H.F. (1974) A revision of the Australian genusJournal of Entomology, 38, 219–226.Stenaspidius Westwood (Coleoptera, Scarabaeidae,Analytical Software (1996). Statistix for Windows. User’sGeotrupinae). Records of the South Australian Museum,Manual. Analytical Software, Tallahassee.17, 11–21.Carne, P.B. (1957) A Systematic Revision of the Australian
Howden, H.F. (1975) A sixth species of StenaspidiusDynastinae (Coleoptera: Scarabaeidae). CSIRO,Westwood with a note on adult behaviour (Coleoptera:Melbourne.Scarabaeidae: Geotrupinae). Journal of the AustralianCarne, P.B. (1965) A revision of the genus ElephastomusEntomological Society, 14, 439–441.Macleay (Coleoptera: Geotrupidae). Journal of the
Howden, H.F. (1979) A revision of the Australian genusEntomological Society of Queensland, 4, 3–13.Blackburnium Boucomont (Coleoptera: Scarabaeidae;Carne, P.B. (1976) Cheiroplatys volsellus sp. n. and notesGeotrupinae). Australian Journal of Zoology,on other related species (Coleoptera: Scarabaeidae:Supplementary Series, 72, 1–88.Dynastinae). Journal of the Australian Entomological
Howden, H.F. (1981) Zoogeography of some AustralianSociety, 15, 85–88.Coleoptera as exemplified by the Scarabaeoidea.Carne, P.B. (1978) Dasygnathus blattocomes sp. n.Ecological Biogeography of Australia (ed. by A. Keast),(Coleoptera: Scarabaeidae: Dynastinae). Journal of thepp. 1009–1035. Junk, The Hague.Australian Entomological Society, 17, 91–93.
Howden, H.F. (1985a) A revision of the Australian beetleCarne, P.B. (1980) Pseudoryctes storeyi sp. n. and newgenera Bolbaleaus Howden & Cooper, Blackbolbusrecords of other species of Pseudoryctes SharpHowden & Cooper, and Bolborhachium Boucomont(Coleoptera: Scarabaeidae: Dynastinae). Journal of the(Scarabaeidae: Geotrupinae). Australian Journal ofAustralian Entomological Society, 19, 255–258.Zoology, Supplementary Series, 111, 1–179.Carne, P.B. (1981) Cryptoryctes minchami sp. n. and notes
Howden, H.F. (1985b) Expansion and contraction cycles,on other Cryptoryctes spp. (Coleoptera: Scarabaeidae:endemism and area: the taxon cycle brought full circle.Dynastinae). Journal of the Australian EntomologicalTaxonomy, Phylogeny and Zoogeography of Beetles andSociety 20, 249–252.Ants (ed. by G.E. Ball), pp. 473–487. Junk, Dordrecht.Carne, P.B. (1985a) Neonastes uptoni sp. n. from the
Howden, H.F. (1992) A revision of the Australian beetleNorthern Territory, with notes on the holotype andgenera Eucanthus Westwood, Bolbobaineus Howden &distribution of N. glabricollis (Macleay) (Coleoptera:Cooper, Australobolbus Howden & Cooper andScarabaeidae: Dynastinae). Journal of the AustralianGilletinus Boucomont (Scarabaeidae: Geotrupinae).Entomological Society, 24, 69–72.
Carne, P.B. (1985b) A new genus and species of Dynastinae Invertebrate Taxonomy, 6, 605–717.
1999 Blackwell Science Ltd, Diversity and Distributions, 5, 143–149

Localized distributions of Australian scarabs 149
Krikken, J. (1976) Elephastomus carnei, a new species from Stebnicka, Z. & Howden, H.F. (1996) Australian generaQueensland (Coleoptera: Geotrupidae). Entomologische and species in the tribes Odontolochini, Psammodiini,Berichten, 36, 101–103. Rhyparini, Stereomerini and part of the Eupariini
Lumaret, J.-P. & Lobo, J.M. (1996) Geographic (Coleoptera: Scarabaeoidea: Aphodiinae). Invertebratedistribution of endemic dung beetles (Coleoptera, Taxonomy, 10, 97–170.Scarabaeoidea) in the Western Palaearctic. Biodiversity Stebnicka, Z. & Howden, H.F. (1997) Revision of theLetters, 3, 192–199. Australian species of Ataenius Harold (Coleoptera:
Main, A.R. (1987) Evolution and radiation of the Scarabaeoidea: Aphodiinae: Eupariini). Invertbrateterrestrial fauna. Fauna of Australia, Vol. 1A, General Taxonomy, 11, 735–821.Articles (ed. by G.R. Dyne & D.W. Walton), pp. 136–155. Steel, R.G.D. & Torrie, J.H. (1980) Principles andAustralian Government Publishing Service, Canberra. Procedures of Statistics. A Biometrical Approach. 2nd
Matthews, E.G. (1972) A revision of the scarabaeinae dung edn. McGraw-Hill Kogakusha, Tokyo.beetles of Australia I. Tribe Onthophagini. Australian Storey, R.I. (1977) Six new species of Onthophagus LatreilleJournal of Zoology, Supplementary Series, 9, 1–330. (Coleoptera: Scarabaeidae) from Australia. Journal of
Matthews, E.G. (1974) A revision of the scarabaeinae the Australian Entomological Society, 16, 313–320.dung beetles of Australia II. Tribe Scarabaeini. Storey, R.I. (1984) A new species of ApentocanthonAustralian Journal of Zoology, Supplementary Series, 24, Matthews from north Queensland (Coleoptera:1–211. Scarabaeidae: Scarabaeinae). Memoirs of the Queensland
Matthews, E.G. (1976) A revision of the scarabaeinae Museum, 21, 387–390.dung beetles of Australia III. Tribe Coprini. Australian Storey, R.I. (1986) A new flightless species of AulacoprisJournal of Zoology, Supplementary Series, 38, 1–52. White from north Queensland (Coleoptera:
Matthews, E.G. & Stebnicka, Z. (1986) A review ofScarabaeidae: Scarabaeinae). Memoirs of the Queensland
Demarziella Balthasar, with a transfer from AphodiinaeMuseum, 22, 197–203.
to Scarabaeinae (Coleoptera: Scarabaeidae). AustralianStorey, R.I. (1991) New species and new records ofJournal of Zoology, 34, 449–461.
Tesserodon Hope (Coleoptera: Scarabaeidae) fromNikolajev, G.V. (1990) New species of the genusnorthern Australia. Memoirs of the Queensland Museum,Elephastomus Macleay (Coleoptera, Scarabaeidae,30, 577–588.Geotrupinae) from Australia [in Russian]. Vestnik
Storey, R.I. & Howden, H.F. (1996) Revision ofZoologii, 6, 67–69.Australoxenella Howden & Storey in AustraliaQuammen, D. (1996) The Song of the Dodo. Island(Coleoptera: Scarabaeidae: Aphodiinae). Memoirs of theBiogeography in an Age of Extinction. Pimlico, London.Queensland Museum, 39, 365–380.Stebnicka, Z. & Howden, H.F. (1994) A revision of the
Storey, R.I. & Weir, T.A. (1988) New localities andAustralian genus Podotenus A. Schmidt (Coleoptera:biological notes for the genus Onthophagus LatreilleScarabaeoidea: Aphodiini). Invertebrate Taxonomy, 8,(Coleoptera: Scarabaeidae) in Australia. Australian17–62.Entomological Magazine, 15, 17–24.Stebnicka, Z. & Howden, H.F. (1995) Revision of
Storey, R.I. & Weir, T.A. (1990) New species ofAustralian genera in the tribes Aphodiini, Aegialiini andOnthophagus Latreille (Coleoptera: Scarabaeidae) fromProctophanini (Coleoptera: Scarabaeidae: Aphodiinae).
Invertbrate Taxonomy, 9, 709–766. Australia. Invertebrate Taxonomy, 3, 783–815.
1999 Blackwell Science Ltd, Diversity and Distributions, 5, 143–149