how does ocean acidification impact phytoplankton ... · how does ocean acidification impact...
TRANSCRIPT

1
How does ocean acidification impact phytoplankton productivity and community structure?
Aliza Ray1 and Joe Vallino
2
1Bard College, Annandale-On-Hudson NY 12504
2Marine Biological Laboratory, Ecosystem Center, Woods Hole MA 02543

2
Abstract
Increasing atmospheric CO2 concentration will result in the acidifying and CO2 fertilization of our
oceans. Phytoplankton assemblages will be among the organisms affected by theses changes. We
tested how coastal New England phytoplankton community photosynthetic productivity and
community composition will be changed. This microcosm experiment manipulated the CO2
concentrations to be 370µatm, 925µatm, and 3700µatm over a 3 week incubation period.
Treatments showed reduced pH and changed dissolved inorganic carbon levels relative to each
treatment. Measured parameters included nutrients, Chl a, pCO2, enumeration, and quantification
of phytoplankton taxa using microscopy and flow cytometry. The present study revealed a coastal
plankton community that was slightly affected by ocean acidification. Changes in respiration
across the course of a 24 hour period showed net consumption cycles to be greatest at elevated
CO2 levels. A species shift was seen from control treatments to elevated treatments and over a 5
day period. We have concluded that future research needs to be done to accurately determine how
phytoplankton will be affected by ocean acidification changes.
Key Words:
Ocean acidification, phytoplankton, primary productivity, community composition
Introduction
Elevated atmospheric CO2 levels, primarily due to fossil fuel combustion, has led to
increased CO2 fertilization of oceans (Brewer 2009), also known as ‘ocean acidification’.
Currently the atmospheric CO2 concentration is 390ppm, and is expected to increase to 750 ppm
or higher by the end of the century (Raven et al. 2005). Global elemental cycles are driven by
biological activity. Assessing the impact of ocean acidification on marine microorganisms is
important for understanding how aquatic systems will react. Observing microbe-driven
ecosystem function changes to elevated CO2 has proven challenging (Liu et al. 2010) and (Joint
et al. 2011).
Phytoplankton play an important role as primary producers of aquatic systems. Direct
effects between ocean acidification and photosynthetic ability have been observed to increase
under elevated pCO2 (Rost et al. 2008). The net rate of organic carbon production determines
support for higher trophic levels. Natural phytoplankton assemblages have been shown to
enhance photosynthesis under elevated pCO2 (Egge et al. 2009). This study will focus on coastal
phytoplankton community because they contribute significantly to global primary productivity
(Field et al. 1998).
Changes in community composition can also be caused by elevated pCO2. Global
changes in biodiversity can alter ecosystem services and disturb biogeochemical cycles, such as
control the CO2 taken up by the oceans. Different species play different roles in ecosystem

3
dynamics. For example, the size of phytoplankton determines grazing efficiencies and can alter
the population structure of grazers. Microbial community structure shifts can mean a loss of
biodiversity, compromised of ecosystem robustness, and potentially major consequences for
higher trophic levels. Studies that have observed the effects of increased CO2 inputs on
phytoplankton systems have found mixed results on phytoplankton assemblages (Nielsen et al.
2010). It is likely that different phytoplankton taxa react differently to ocean acidification. The
effects of elevated pCO2 on microzooplankton have shown no consistent effect to ocean
acidification on microbial biodiversity and community composition (Suffrian et al. 2008).
In the following we describe the responses of natural fall coastal phytoplankton groups to
changes in the carbonate chemistry of a microcosm system to determine how future lowered pH
levels and increased CO2 may alter functioning and composition. This study focuses on the
changes to phytoplankton photosynthetic productivity and community assemblages. To examine
the effects we created semi-continuous microcosm culture techniques in which CO2 manipulated
environments were periodically pulsed by pulling sampling and returning medium with nutrients
and filtered seawater. Here, we report on the results of a 3 week experiment demonstrating
effects of elevated CO2 levels of phytoplankton productivity on local North-west Atlantic Ocean
assemblages. We discuss the potential ecological and biogeochemical implications of our
findings.
Methods
A coastal seawater sample was obtained from Woods Hole, MA (41.5264° N, 70.6736°
W) during November. The sample was filtered through 200µm mesh. 1L of sample was allocated
to each microcosm with nutrients. Nutrient concentrations in the microcosms remained at 36µM
KNO3, 52µM NaSiO3, and 2.3µM KH2PO4. The experiment was preformed on triplicates at
925µatm, 3700µatm, and controlled atmospheric levels of 370µatm. Microcosms were incubated
at 20˚C on a 12 hour light cycle and were continuously stirred. CO2 and air input were bubbled
into the sample at 23 mL min-1
. Each day, 10% of the sample was removed and the same volume
of 0.45µm filtered seawater and nutrients were added back in. All air and water samples were
taken +/- 1 hour of growth lights turning on each day. The microcosms sat undisturbed for 4 days
prior to initiation of the pulse chemostat method.

4
10mL of sample was used to measure pH with Accumet pH/conductivity meter (Fisher
Scientific). 10mL was used for fluorescence using a Fluorometer (Turner Designs). 80mL of was
filtered using 25mm GF/F filters for nutrients. Ammonium was measured using a modification of
the phenol-hypochlorite method (Solorzano 1969) analyzed with a Cary UV Visible
Spectrophotometer (Varian). Phosphate was analyzed by a modification of the method of
Murphy and Riley (1962) using UV-VIS Spectrophotometer (Shimadzu). Nitrate was measured
using QuickChem Flow Injection Analyzer (LACHET). Filters were dried, and analyzed for
molar carbon and nitrogen with a PerkinElmer 2400 Series II CHN Elemental Analyzer.
10mL of sample was used to measure dissolved inorganic carbon levels. 20mL of
Ascarite scrubbed CO2-free air was drawn into the syringe, 0.2mL of H2SO4 was added, and
sample was shaken for 1 minute prior to injection of air. pCO2 in the microcosm head space and
CO2 input to the system was also measured. CO2 consumption was calculated by subtracting
pCO2 input from output, then multiplying by flow rate (23mL min-1
). Dissolved inorganic carbon
(DIC) and microcosm air was measured using gas chromatography (GC-8A Shimadzu).
50mL of sample was drawn for microscopy. Autotrophs were fixed in alkaline Lugol’s
solution (10g iodine, 20g potassium iodide, 10g sodium acetate, in 140ml distilled water) for a
concentration of 0.1%, followed by borate-buffered formalin addition of 2.4%, and 3% sodium
thiosulfate for a final concentration of 0.1% (Sherr and Sherr, 1993). Preserved sample were
filtered onto 25mm white 0.8µm membrane filters (Osmonics). Taxa were identified to the
nearest genus or species. Identification was done using differential interference and bright field
light microscopy (Zeiss Axio Imager.M2) at 20X and 40X.
Additional samples (50mL) were fixed to a final concentration of 5% glutaric dialdehyde.
Direct DAPI (4',6-diamino-2-phenylindole dihydrochloride) counts were taken from the
glutaraldehyde preserved samples. 1 ml of sample and 50µl of 200µg/ml working solution DAPI
was incubated for 5 minutes then drawn onto 1µm black polycarbonate filters. Phosphate
buffered saline was used to rinse. Samples were viewed under 20X magnification using blue
fluorescence. Samples were quantified:
cells ml-1 =(cells/field of view)´ (area of filter covered by sample)
(field of view area)´ (preseravtion dilution factor)´ (ml filtered)
Flow cytometry was done on live, unpreserved samples using a flow cytometer (BD
FACSCalibur) using CellQuest Pro software. 1mL samples were drawn and filtered using 35µm

5
mesh, and 5µL of 1µm beads were added to the sample. Particles were enumerated based on size,
complexity, and fluorescence.
Results
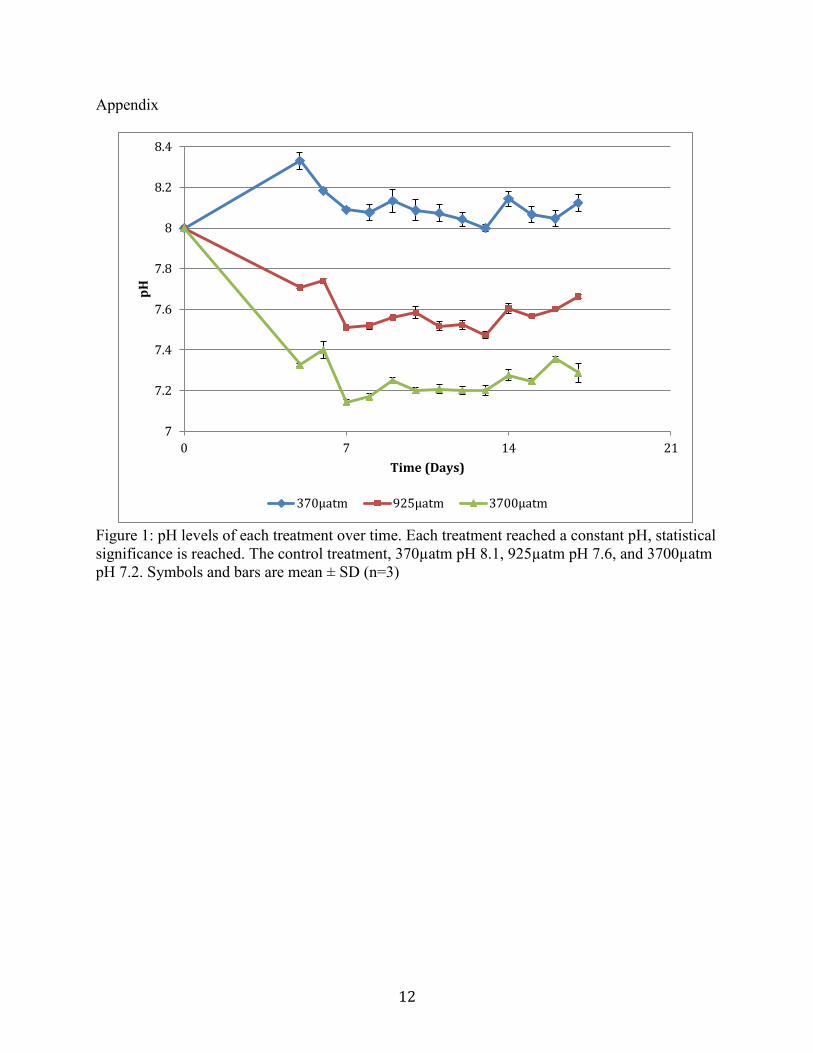
Measured pH during the experimental period remained constant once a steady pH was
reached (Figure 1). There were significant differences in pH between treatments. The 370µatm
(control) maintained a pH of 8.1, 925µatm was 7.6, and 3700µatm was 7.3. Seawater sampled at
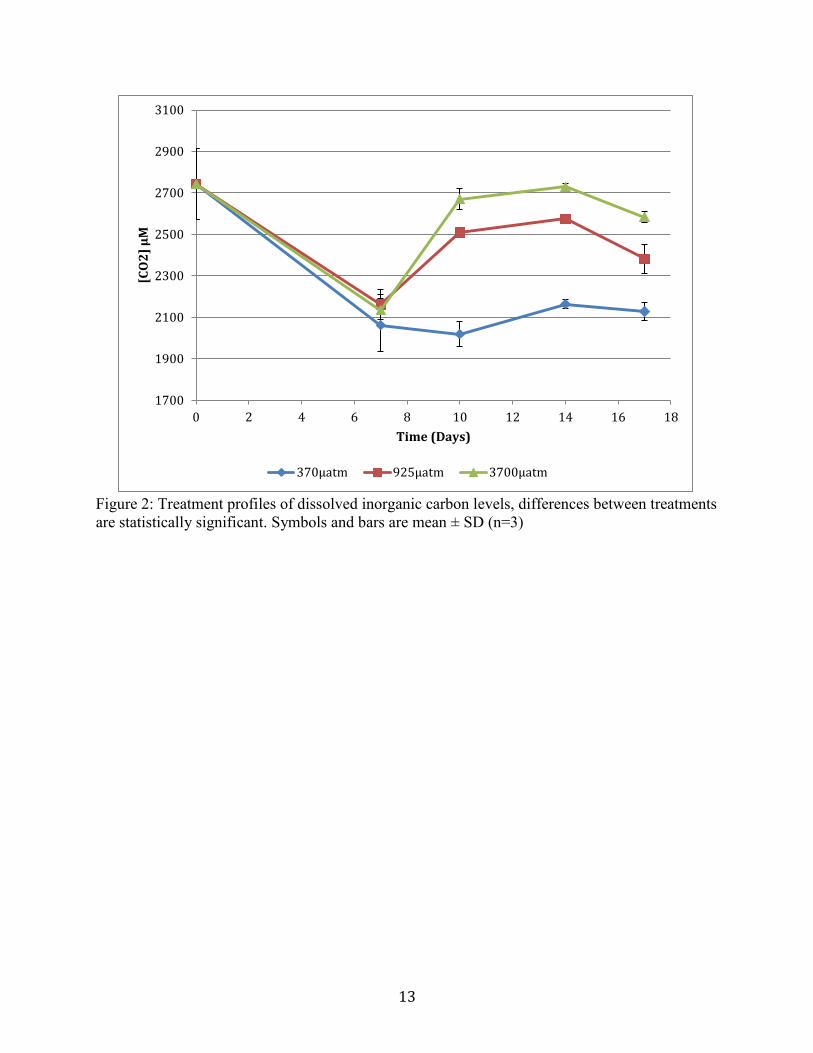
the start of the experiment measured a pH of 8. The initial level of DIC of the sampled water was
2700µM, and for all treatment levels the DIC changes occurred during the experimental period
(Figure 2).
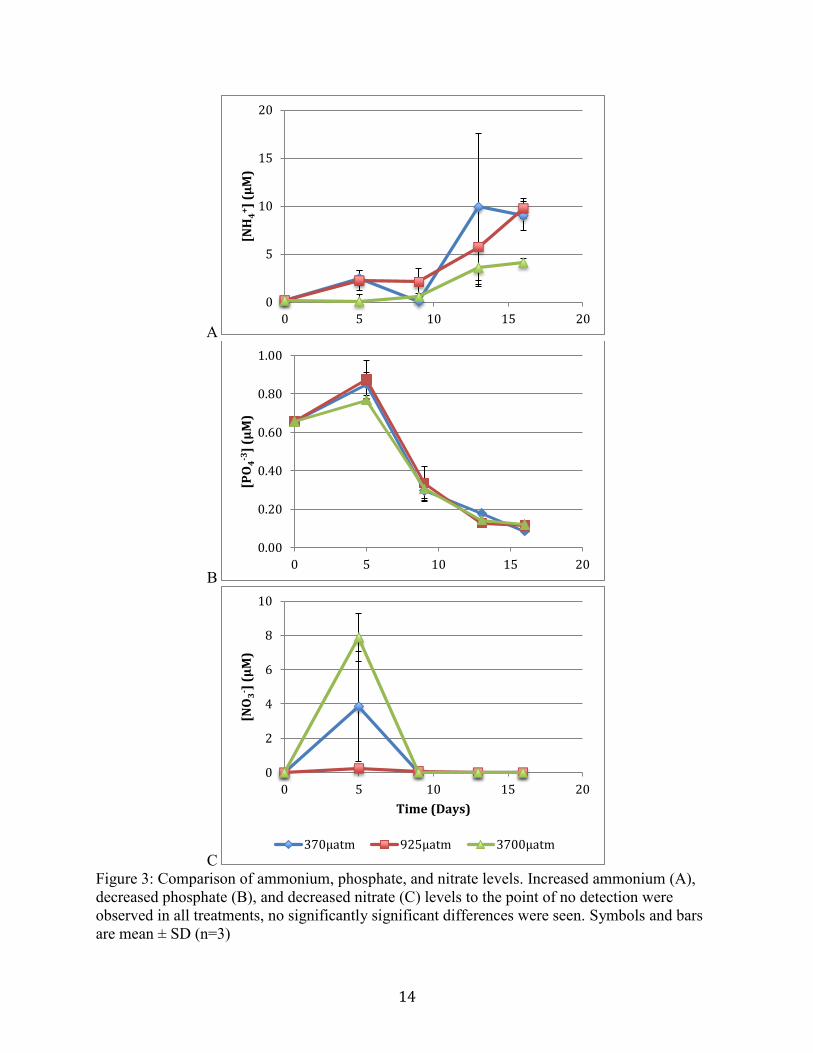
Nutrient profiles phosphate and ammonium were measured throughout the course of the
experiment. Initial concentrations of ammonium were undetectable, phosphate measured
0.66µM, and nitrate was also undetectable in the seawater sampled. Ammonium levels gradually
increased in all treatments over the course of the experiment (Figure 3A). Phosphate levels
gradually decreased in all treatments (Figure 3B). Levels of nitrate after day 5 were below
detection limit (Figure 3C). No statistically significant differences were found between
treatments.
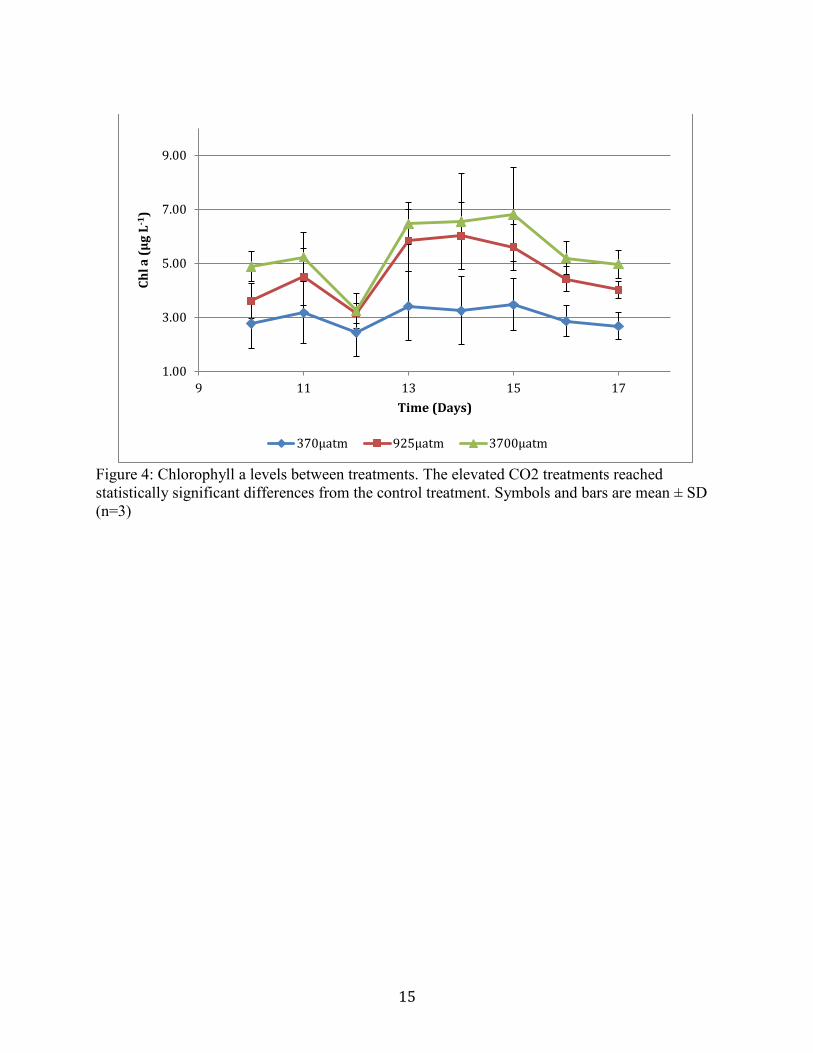
Chlorophyll a changes between treatments reflect changes based on treatment. Final Chl
a measurements reflect significant differences between treatments, with the 3700µatm treatment
having the highest Chl a values, and the ambient CO2 treatment measuring the lowest (Figure 4).
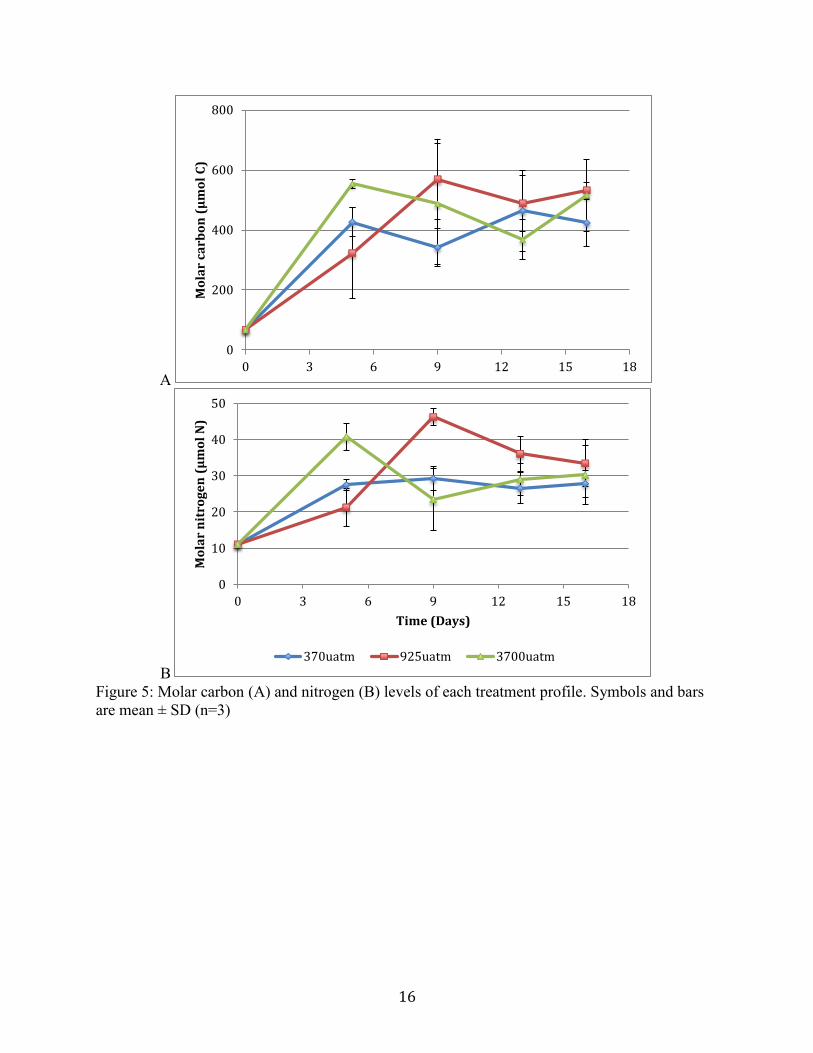
Molar carbon and nitrogen levels were not significant between treatments, all treatments showed
increasing fluctuation of C and N (Figure 5).
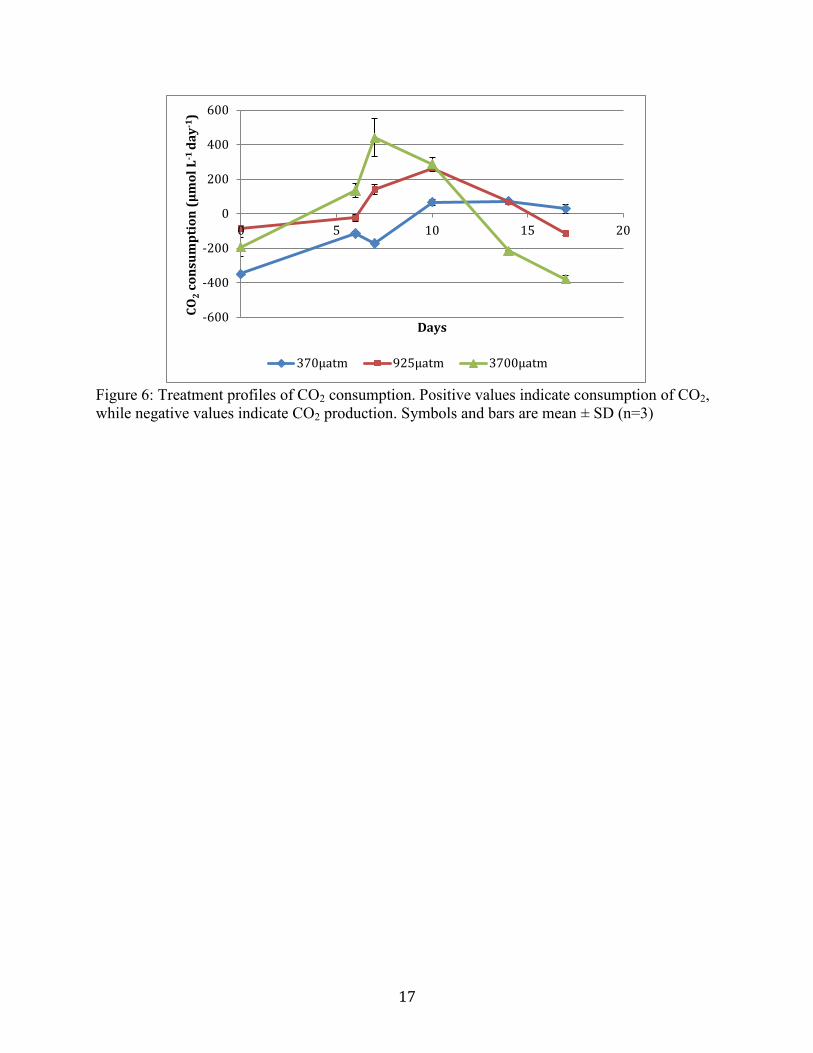
Measured carbon dioxide consumption showed significant difference between treatments.
Positive CO2 consumption values and slopes indicate net photosynthetic productivity, while
negative values and slopes indicate net CO2 production. Initial pCO2 values show net production,
while CO2(g) values after a 5 day acclimation period show increased consumption. Consumption
decreased in both elevated CO2 treatments. The control treatment reached a steady CO2
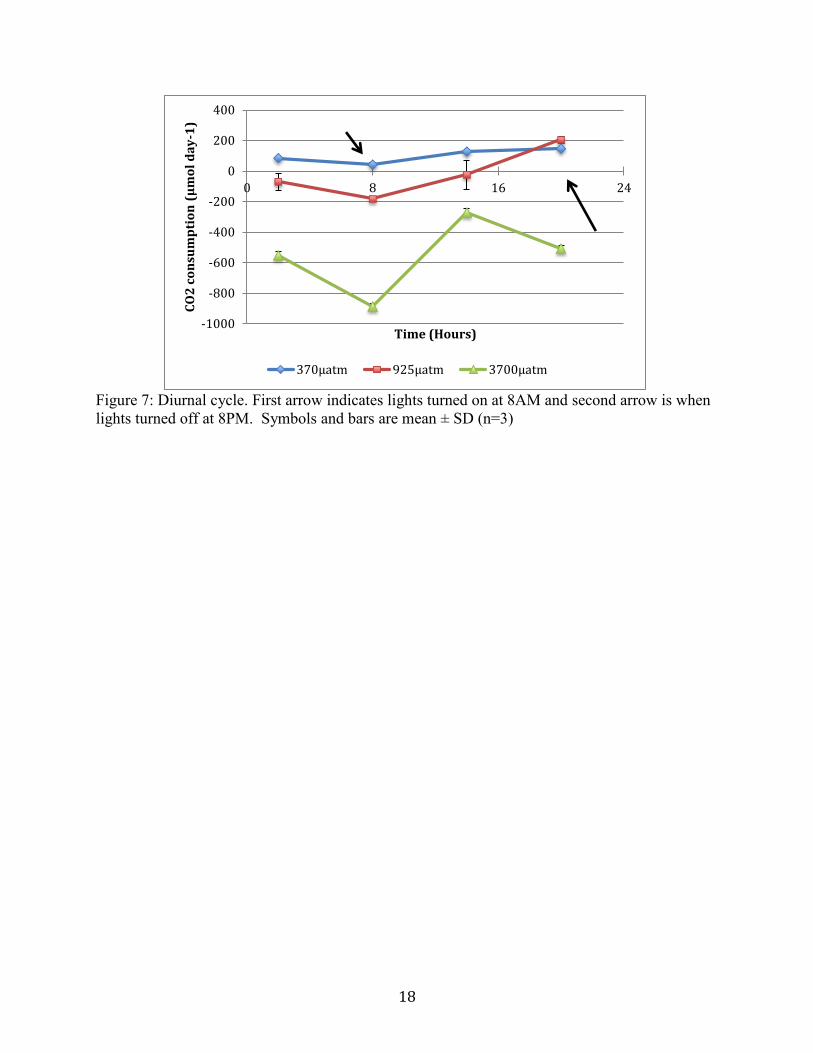
consumption rate (Figure 6). A diurnal pCO2 cycle shows significantly different consumption
values between treatments. Increased consumption is seen in each treatment during the 12 hours
the lights were on, except the highest CO2 treatment saw increased CO2 production from the
peak of midday until lights turn off at night. All treatments have negative consumption during

6
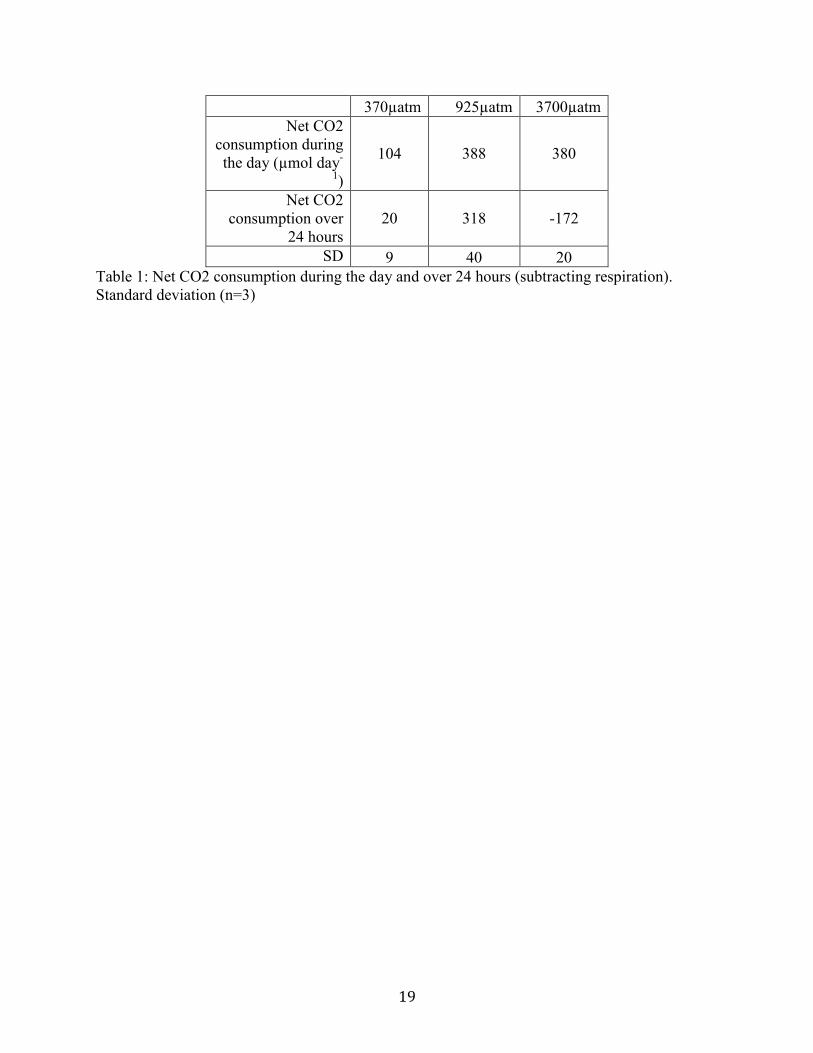
the 12 hours the lights were not on (Figure 7). Net CO2 consumption values due to
photosynthesis (during the period the lights were on) are increased from the 925µatm and
3700µatm treatments (Table 1). Net CO2 consumption during the course of 24 hours is negative
only for the 3700µatm treatment, meaning there is more CO2 produced than was consumed.
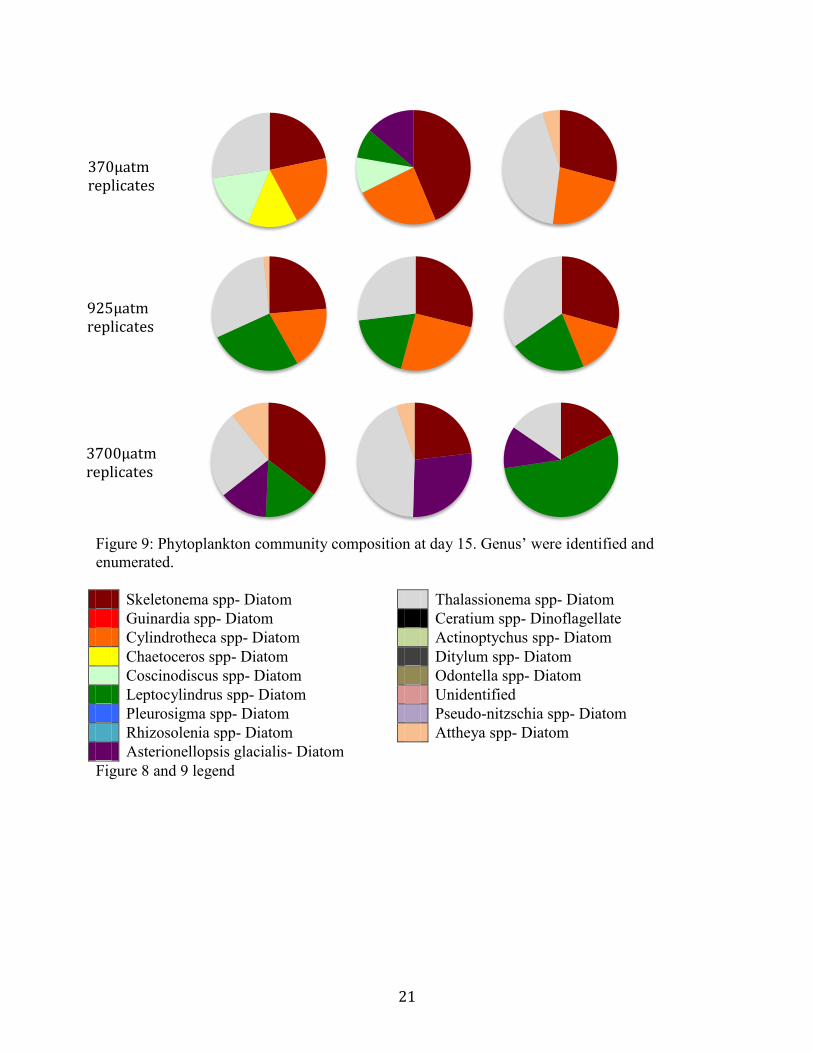
Changes to species composition from the control were seen in both between treatments
and changed over a 5 day period. Samples analyzed on day 10 and 15 showed variance between
treatments, and some variance between replicates. Sixteen diatom genus’ and one dinoflagellate
were identified. On day 10, control and 925µatm treatments were dominated by a diverse
assemblage of diatoms including Skeltonema spp, Guinardia spp, Cylindrotheca spp,
Chaetoceros spp, Coscinodiscus spp, Leptocylindrus spp, Pleurosigma spp, and Rhizolenia spp.
The 3500µatm had additional abundance dominance of Asterionellopsis glacialis and
Thalassionema spp (Figure 8). A succession and a shift in abundances occurred during the
following 5 day period. Leptocylindrus spp and Thalassionema spp showed increased dominance
in all treatments at each CO2 level. Cylindrotheca spp, previously seen on day 10 in all
treatments, was no longer seen in the 3700µatm microcosms. Asterionellopsis glacialis became
abundant in the elevated 3700µatm treatment, and Skeletonema spp remained dominant at all
treatment levels (Figure 9). The dionglagellate Ceratium spp was present in one replicate at
925µatm on day 10.
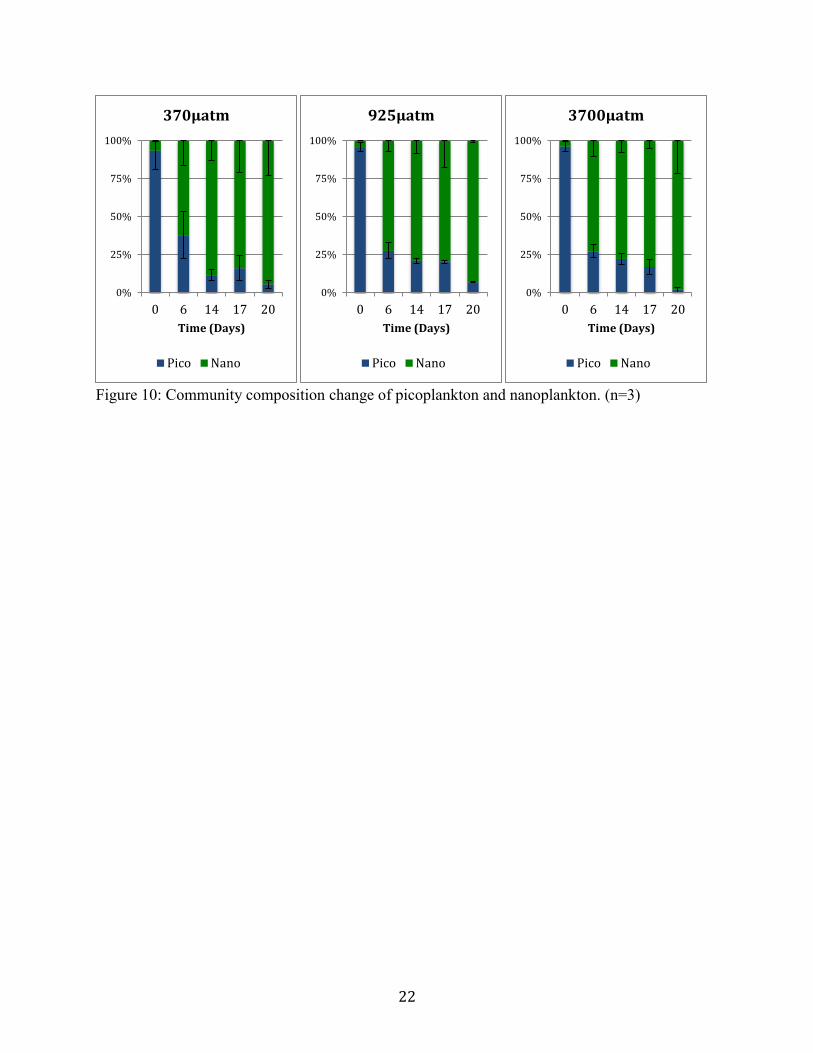
Nanoplankton and picoplankton community structure dynamics show no significant
difference between treatments. All treatments experienced a shift from a picoplankton
dominance to an nanoplankton dominance (Figure 10). In all treatments nanoplankton abundance
increased, while picoplankton abundance decreased (Table 2).
Discussion
The aim of the study was to test how predicated ocean acidification would affect primary
productivity and community composition of coastal phytoplankton assemblages. Microcosms
experienced increased acidity and altered DIC levels respective to each treatment level. This
established that the microcosm environment achieved a state that mimicked carbonate processes
occurring due to ocean acidification.

7
Despite relatively low nitrate levels, there was growth in the microcosms. Daily nitrate
additions of nutrient stock were enough to sustain growth, despite levels being undetectable 24
hours later.
Growth of phytoplankton in each treatment was observed. The initial drop in DIC over
the 5 day assimilation period occurred because of extreme growth in all treatments.
Photosynthetic carbon fixation causes a CO2 and DIC decrease. Phosphate levels decreased
significantly as well in all treatments, indicating growth, due to phosphate uptake. In addition,
Chlorophyll a levels of each treatment level had shown significant differences by the end of the
experiment.
Increased ammonium can mean more grazing on phytoplankton by larger zooplankton,
however, based on microcopy, no grazers were identified in any of the treatments. Nitrate uptake
by phytoplankton has been shown to inhibit by ammonium levels (l'Helguen et al. 2008).
Phytoplankton productivity, measured by the consumption of CO2, in all microcosms
increased during the first half of the experimental period. Decreased productivity in the elevated
CO2 treatments does not mean that consumption was not occurring, only that consumption was
lowered. Further examination of the carbonate chemistry of the systems would allow for
calculations of CO2(aq) in the water. The 24 hour diurnal cycle shows the fluctuations of CO2
consumption throughout the day. CO2 consumption is greatest in the elevated treatments. The
3700µatm treatment experienced net CO2 production during the course of 24 hours. When
photosynthesis was occurring during the lights on cycle, CO2 consumption occurs in all
treatments. CO2 consumption was greatest at the 3700µatm treatment level. Further works need
to be done to determine the CO2(aq) of the microcosms and carbonate chemistry changes to the
systems. Diatom dominated systems have shown 27% to 39% CO2 uptake increases from control
(~370µatm) (Riebesell et al. 2007).
Molar carbon level differences between treatments do not indicate elevated carbon
content at higher CO2 treatment levels. This could be due to carbon content difference based on
diversity of species seen.
The initial species dominance shift showed differences between treatments, primarily
diatoms. Resilience of Skeletonema spp in 3700µatm treatments was consistent with findings of
previous studies (Nielsen et al. 2012). The abundance of Asterionellopsis glacialis under
elevated CO2 was especially apparent. Individual phytoplankton physiology or nutrient

8
availability have not been closely assessed. Coastal phytoplankton communities have also been
observed to be impervious to CO2 elevated changes (Nielsen et al. 2010). While we acknowledge
the changes to species composition changes we saw, we also suggest that coastal phytoplankton
species could be tolerant of broad level pH fluctuations due to respiratory and photosynthetic
processes (Hansen 2002). Our phytoplankton may have been showing resilience to abrupt CO2
changes, previously seen (Vogt et al. 2008).
Flow cytometry results did not yield significant changes between treatment levels of
nanoplankton (2-20µm) and picoplankton (0.2-2µm) populations. Results were evidence of a
microcosm experiment, in which all treatments experienced a bottle-effect, in which
nanoplankton abundance increased, and picoplankton abundance decreased. Previous studies
have found only slight changes to picoplankton community composition under elevated CO2
levels (Newbold et al. 2012).
In a high CO2 ocean, phytoplankton may over-consume CO2, increasing their C:N ratio
(Toggweiler 1993). Elevated molar carbon level increased in all treatments, and the 925µatm and
3700µatm microcosm did not show significant differences in higher molar C compared to control
treatments. Increased CO2 and increased light have been found to decrease primary production at
light intensities representative of surface layer light levels (Gao et al 2012).
Further research on the effect of increasing CO2 levels on coastal, estuarine, and open-
ocean phytoplankton community assemblages. Our research is an example of one coastal
scenario. Both individual species effects and community productivity and resilience impacts
need to be further examined. Multiple environmental factors, such as nutrient level changes, need
to be examined in isolation of other variables, because of phytoplankton sensitivity to light and
nutrient levels. The ability of certain species to adapt to sudden pH changes is largely unknown.
It is important to study the impacts of ocean acidification to better understand its implications.
Acknowledgements
This exploration would not have been possible without the generosity of everyone at the
Marine Biological Lab. Joe Vallino for all his help, tremendous guidance, and carbonate
chemistry lessons. Hugh Ducklow, Matthew Erickson, Hap Garritt, and Ken Foreman for sharing
their lab and equipment. Jim McIlvain for endless Zeiss microcopy tutorials. Rich McHorney,
Alice Carter, and Carrie Harris for their endless patience.

9
Literature Cited
Brewer, P.G. 2009. A changing ocean seen with clarity. Proceedings of the National Academy of
Sciences USA, 106: 12213-14.
Field C.B., Behrenfeld, M.J., Randerson, J.T., Falkowski, P. 1998. Primary production of the
biosphere: integrating terrestrial and oceanic components. Science 281: 237-40.
Gao, K., Xu, J., Gao, G., Li, Y., Hutchins, D.A., Huang, B., Wang, L., Zheng, Y., Jin, P., Cai, X.,
Hader, D.P., Li, W., Xu, K., Liu, N., Ribesell, U. 2012. Rising CO2 and increased light
exposure synergistically reduce marine primary productivity. Nature Climate Change 2:
519-523
Hansen P.J. (2002) Effect of high pH on the growth and survival of marine phytoplankton:
implications for species succession. Aquatic Microbial Ecology 28: 79-88
Joing, I., Doney, S.C., Karl, D.M. 2011. Will ocean acidification affect marine microbes? The
ISME Journal, 5: 1-7.
Kemp, P.F., Cole, J.J., Sherr, B.F., Sherr, E.B., 1993. “Handbook of Methods in Aquatic
Microbial Ecology”. Lewis Publishers, Ann Arbor- Lugol’s protocol
L'Helguen, S., Maguer, J.F., Caradec, J. 2008. Inhibition kinetics of nitrate uptake by ammonium
in size-fractionated oceanic phytoplankton communities: implications for new production
and f-ratio estimates. Journal Plankton Research. 2008 30: 1179-1188
Lau, J., Weinbauer, M.G., Maier, C., Dai, M., Gattuso, J.P. 2010. Effect of ocean acidification on
microbial diversity and on microbe-driven biogeochemistry and ecosystem functioning.
Aquatic Microbial Ecology, 61: 291-305.
Murphy, J., Riley, J.P, 1962. A modified single solution method for the determination of
phosphate in natural waters. Analytica Chemica Acta 27: 31-6
Newbold, L.K., Oliver, A.E., Booth, T., Tiwari, B., DeSantis, T., Maguire, M., Andersen, G.,
van der Gast, C.J., Whiteley, A.S. 2012. The response of marine picoplankton to ocean
acidification. Environmental Microbiology 14(9): 2293-307
Nielsen, L.T., Jakobsen, H.H., Hansen, P.J. 2010. High resilience of two coastal plankton
communities to twenty-first century seawater acidification: evidence from microcosm
studies. Marine Biology Research: 6(6): 542-55

10
Nielsen, L.T., Hallegraeff, G.M., Wright, S.W., Hansen, P.J. 2012. Effects of experimental
seawater acidification on an estuarine plankton community. Aquatic Microbial Ecology
65: 271-85
Raven, J., Caldeira, K., Elderfield, H., Hoeg-Guldberg, O. 2005. Ocean acidification due to
increasing atmospheric carbon dioxide. Policy Document 12/05, The Royal Society,
London, available at: www.royalsoc.ac.uk
Riebesell, U., Schulz, K.G., Bellerby, R.G.J., Botros, M., Fritsche, P., Meyerhofer, M., Neill, C.,
Nondal, G., Oschlies, A., Wohlers, J., Zollner, E. 2007. Enhanced biological carbon
consumption in a high CO2 ocean. Nature, 450: 545-8.
Rost, B., Zondervan, I., Wolf-Gladrow, D. 2008. Sensitivity of phytoplankton to future changes
in ocean carbonate chemistry: current knowledge, contradictions and research directions.
Mar Ecological Progress Series, 373: 227-37.
Suffrian, K., Simonelli, P., Nejstgaard, J.C., Putzeys, S., Carotenuto, Y., Antia, A.N. 2008.
Microzooplankton grazing and phytoplankton growth in marine mesocosms with
increased CO2 levels. Biogeosciences Discussions 5: 411-433.
Solórzano, L. 1969. Determination of Ammonia in Natural Waters by the Phenol Hypochlorite
Method. Limnology and Oceanography 14.5: 799-801
Toggweiler, J.R. 1993. Carbon overconsumption. Nature 363: 210-11
Vogt, M., Steinke, M., Turner, S., Paulino, A., Meyerhofer, M., Riebesell, U., LeQuere, C. and
Liss, P. 2008. Dynamics of dimethylsulphoniopropionate and dimethylsulphide under
different CO2 concentrations during a mesocosm experiment. Biogeosciences 5: 407-419.

11
Figures and Tables
Figure 1. pH levels of each treatment over time. Each treatment reached a constant pH, statistical
significance is reached. The control treatment, 370µatm pH 8.1, 925µatm pH 7.6, and
3700µatm pH 7.2. Symbols and bars are mean ± SD (n=3)
Figure 2. Treatment profiles of dissolved inorganic carbon levels, differences between treatments
are statistically significant. Symbols and bars are mean ± SD (n=3)
Figure 3. Comparison of ammonium, phosphate, and nitrate levels. Increased ammonium (A),
decreased phosphate (B), and decreased nitrate (C) levels to the point of no detection
were observed in all treatments, no significantly significant differences were seen.
Symbols and bars are mean ± SD (n=3)
Figure 4. Chlorophyll a levels between treatments. The elevated CO2 treatments reached
statistically significant differences from the control treatment. Symbols and bars are mean
± SD (n=3)
Figure 5. Molar carbon (A) and nitrogen (B) levels of each treatment profile. Symbols and bars
are mean ± SD (n=3)
Figure 6. Treatment profiles of CO2 consumption. Positive values indicate consumption of CO2,
while negative values indicate CO2 production. Symbols and bars are mean ± SD (n=3)
Figure 7. Diurnal cycle. First arrow indicates lights turned on at 8AM and second arrow is when
lights turned off at 8PM. Symbols and bars are mean ± SD (n=3)
Table 1. Net CO2 consumption during the day and over 24 hours (subtracting respiration).
Standard deviation (n=3)
Figure 8. Phytoplankton community composition at day 10. Genus’s were identified and
enumerated.
Figure 9. Phytoplankton community composition at day 15. Genus’ were identified and
enumerated.
Figure 8 and 9 legend.
Figure 10. Community composition change of picoplankton and nanoplankton. (n=3)
Table 2. Changes in abundance of the nanoplankton and picoplankton population in each
treatment. (n=3)
12
Appendix
Figure 1: pH levels of each treatment over time. Each treatment reached a constant pH, statistical
significance is reached. The control treatment, 370µatm pH 8.1, 925µatm pH 7.6, and 3700µatm
pH 7.2. Symbols and bars are mean ± SD (n=3)
7
7.2
7.4
7.6
7.8
8
8.2
8.4
0 7 14 21
pH
Time (Days)
370µatm 925µatm 3700µatm
13
Figure 2: Treatment profiles of dissolved inorganic carbon levels, differences between treatments
are statistically significant. Symbols and bars are mean ± SD (n=3)
1700
1900
2100
2300
2500
2700
2900
3100
0 2 4 6 8 10 12 14 16 18
[CO
2]
µM
Time (Days)
370µatm 925µatm 3700µatm
14
A
B
C
Figure 3: Comparison of ammonium, phosphate, and nitrate levels. Increased ammonium (A),
decreased phosphate (B), and decreased nitrate (C) levels to the point of no detection were
observed in all treatments, no significantly significant differences were seen. Symbols and bars
are mean ± SD (n=3)
0
5
10
15
20
0 5 10 15 20
[NH
4+]
(µM
)
0.00
0.20
0.40
0.60
0.80
1.00
0 5 10 15 20
[PO
4-3
] (µ
M)
0
2
4
6
8
10
0 5 10 15 20
[NO
3- ]
(µ
M)
Time (Days)
370µatm 925µatm 3700µatm
15
Figure 4: Chlorophyll a levels between treatments. The elevated CO2 treatments reached
statistically significant differences from the control treatment. Symbols and bars are mean ± SD
(n=3)
1.00
3.00
5.00
7.00
9.00
9 11 13 15 17
Ch
l a
(µ
g L
-1)
Time (Days)
370µatm 925µatm 3700µatm
16
A
B
Figure 5: Molar carbon (A) and nitrogen (B) levels of each treatment profile. Symbols and bars
are mean ± SD (n=3)
0
200
400
600
800
0 3 6 9 12 15 18
Mo
lar
carb
on
(µ
mo
l C
)
0
10
20
30
40
50
0 3 6 9 12 15 18
Mo
lar
nit
rog
en
(µ
mo
l N
)
Time (Days)
370uatm 925uatm 3700uatm
17
Figure 6: Treatment profiles of CO2 consumption. Positive values indicate consumption of CO2,
while negative values indicate CO2 production. Symbols and bars are mean ± SD (n=3)
-600
-400
-200
0
200
400
600
0 5 10 15 20
CO
2 c
on
sum
pti
on
(µ
mo
l L
-1 d
ay
-1)
Days
370µatm 925µatm 3700µatm
18
Figure 7: Diurnal cycle. First arrow indicates lights turned on at 8AM and second arrow is when
lights turned off at 8PM. Symbols and bars are mean ± SD (n=3)
-1000
-800
-600
-400
-200
0
200
400
0 8 16 24 C
O2
co
nsu
mp
tio
n (
µm
ol
da
y-1
)
Time (Hours)
370µatm 925µatm 3700µatm
19
370µatm 925µatm 3700µatm
Net CO2
consumption during
the day (µmol day-
1)
104 388 380
Net CO2
consumption over
24 hours
20 318 -172
SD 9 40 20
Table 1: Net CO2 consumption during the day and over 24 hours (subtracting respiration).
Standard deviation (n=3)

20
Figure 8: Phytoplankton community composition at day 10. Genus’s were identified and
enumerated.
370µatm replicates
925µatm replicates
3700µatm replicates
21
Figure 9: Phytoplankton community composition at day 15. Genus’ were identified and
enumerated.
Skeletonema spp- Diatom Thalassionema spp- Diatom
Guinardia spp- Diatom Ceratium spp- Dinoflagellate
Cylindrotheca spp- Diatom Actinoptychus spp- Diatom
Chaetoceros spp- Diatom Ditylum spp- Diatom
Coscinodiscus spp- Diatom Odontella spp- Diatom
Leptocylindrus spp- Diatom Unidentified
Pleurosigma spp- Diatom Pseudo-nitzschia spp- Diatom
Rhizosolenia spp- Diatom Attheya spp- Diatom
Asterionellopsis glacialis- Diatom
Figure 8 and 9 legend
370µatm replicates
925µatm replicates
3700µatm replicates
22
Figure 10: Community composition change of picoplankton and nanoplankton. (n=3)
0%
25%
50%
75%
100%
0 6 14 17 20
Time (Days)
370µatm
Pico Nano
0%
25%
50%
75%
100%
0 6 14 17 20
Time (Days)
925µatm
Pico Nano
0%
25%
50%
75%
100%
0 6 14 17 20
Time (Days)
3700µatm
Pico Nano

23
Nanoplankton
Change in Abundance SD
320µatm 5E+04 4E+03
925µatm 8E+04 1E+03
3700µatm 8E+04 4E+03
Picoplankton
Change in Abundance SD
320µatm -1E+04 1E+03
925µatm -1E+04 4E+02
3700µatm -1E+04 7E+02
Table 2: Changes in abundance of the nanoplankton and picoplankton population in each
treatment. (n=3)